Системная нейрофизиология (nan)
Имитационное научение (подражание) у животных
РедактироватьСущественную роль в формировании поведения высших животных играют явления подражания, которые в основном, хотя и не все, относятся к сфере научения. Мы здесь не будем рассматривать формы подражания, которые относятся к инстинктивному поведению. Это широко распространенная среди животных взаимная стимуляция - аллеломиметическое поведение, при котором выполнение видотипичных действий одних животных является побуждающим фактором для других. Последние в результате начинают выполнять такие же действия (одновременные отдых, собирание пищи и т. д.). Следовательно, в данном случае имеет место взаимопоощрение видотипичной деятельности.
Научение же путем подражания ("имитационное научение") заключается в индивидуальном формировании новых форм поведения, но путем одного лишь непосредственного восприятия действий других животных. Таким образом, мы имеем здесь дело с научением на основе общения. Имитационное научение, как и всякое научение вообще, можно подразделить на облигатное и факультативное. При облигатном имитационном научении результат научения вполне укладывается в рамки видового стереотипа. Особенно это относится к молодым животным, которые путем подражания научаются выполнять некоторые жизненно необходимые действия обычного поведенческого "репертуара" своего вида. Так, у молоди стайных рыб защитная реакция на появление хищника (бегство) формируется в результате подражания поведению других рыб при одном лишь виде поедания хищником членов стаи. Л. А. Орбелн считал такое имитационное поведение "главным охранителем вида", ибо "громадное преимущество заключается в том, что "зрители", присутствующие при акте повреждения члена их же стада или их сообщества, вырабатывают рефлекторные защитные акты и таким образом могут в будущем избежать опасности". Облигатное имитационное научение является также важным элементом реакции следования и распознавания молодыми млекопитающими пищевых объектов. Путем облигатного имитационного научения молодые животные накапливают опыт в гнездостроении у птиц (В. П. Промптов) и шимпанзе (Дж. ван Лазик-Гу-долл) и т.д.
Факультативное имитационное научение в простейших формах представлено в имитации невидотипичных движений на основе облигатного (аллеломиметического) стимулирования. Сюда относятся, например, случаи имитирования обезьянами действий человека, особенно при их содержании в домашней обстановке. Производимые ими при этом действия с предметами быта или инструментами, конечно, выходят за рамки видового поведения. Поскольку здесь имеет место научение новым приемам манипулирования, в данном случае можно говорить о невидотипичном имитационном манипулировании.
Высшим проявлением факультативного имитационного научения следует явно считать решение задач путем подражания (или хотя бы облегчение решения). При таком "имитационном решении задач" у животного-"зрителя" вырабатывается определенный навык в результате одного лишь созерцания действий другой особи, направленных на решение соответствующей задачи. Способность к этому установлена у разных млекопитающих: человекообразных и низших обезьян, собак, кошек, мышей. У обезьян имитационное решение задач играет, очевидно, особенно большую роль. Советский исследователь поведения животных А. Д. Слоним, например, считает, что формирование условных рефлексов происходит в •обезьяньем стаде преимущественно на основе подражания. В пользу этого говорят и полевые наблюдения, выполненные рядом исследователей в последние годы.
Правда, на основе имитационного научения у "зрителей", очевидно, не могут формироваться инструментальные навыки. Об этом свидетельствуют, в частности, опыты американского исследователя Б. Б. Бека, в которых павианы-зрители" наблюдали за употреблением орудия при решении задачи у сородича. "Зрители" не оказались способными к имитационному решению столь сложной задачи, но впоследствии они намного чаще и интенсивнее, чем до опытов, манипулировали этим орудием. Это показывает активирующую роль аллеломиметического поведения и невидотипичного имитационного манипулирования при выработке сложных навыков в условиях общения.
Соотношения отдельных категорий и форм подражания у животных можно проиллюстрировать нижеследующей схемой.
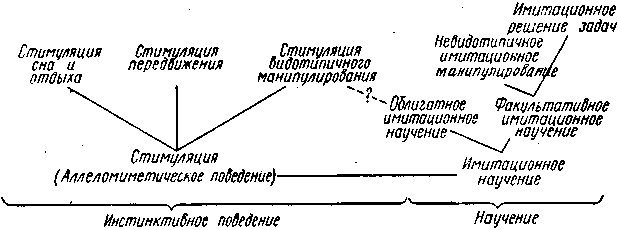
Эта схема неполная в том отношении, что не отражает явления подражания в области сигнализации и коммуникации (например, звукоподражание у птиц). Однако здесь нет принципиальных отличий от отмеченных общих закономерностей и характеристик, и поэтому данная классификация в соответствующих своих разделах вполне применима и для этой области. Так, например, к стимуляции видотипичного поведения относится и стимуляция видотипичной акустической сигнализации ("хоровое пение" у лягушек или птиц), невидотипичному имитационному манипулированию соответствует подражание чужим звукам и песням у птиц, а к облигатному имитационному научению относится, в частности, освоение птенцами видотипичных звуков путем подражания пению взрослых особей. Еще в 1773, г. английский ученый Д. Баррингтон воспитывал молодых коноплянок совместно с птенцами жаворонков. В результате коноплянки перенимали пение жаворонков. С тех пор аналогичные опыты ставились на птицах многократно.
Что же касается обезьян, то в опытах, поставленных Н. А. Тих на молодых гамадрилах, намечались, правда нечетко, некоторые намеки на возможность выработки даже таких навыков, которые можно было бы квалифицировать как имитационное решение задач на основе звукоподражания (в этих опытах обезьяны получали пищевое подкрепление за подражание звукам, которым в их присутствии обучали других обезьян).
Исходя из приведенной классификации можно определить два принципиально различных подхода к изучению подражания у животных.
Аллеломиметическое поведение изучается путем раздельного обучения изолированных друг от друга животных, которые затем сводятся вместе. Советский зоопсихолог Б. И. Хотин, например, обучал одних животных (рыб, птиц, млекопитающих) реагировать положительно на определенный сигнал, а других - отрицательно. После того как животные были сведены вместе и подвергнуты воздействию этого раздражителя, выяснилось, что преобладает: результаты обычного (неимитационного) научения или взаимная стимуляция. В результате можно судить о силе аллеломиметической реакции, т. е. инстинктивного подражания.
При изучении же имитационного научения животные с самого начала общаются между собой. Одна особь (или несколько особей) выполняет при этом роль "актера", т. е. обучается экспериментатором за соответствующее вознаграждение на виду у других особей - "зрителей". Если в результате последние без собственных упражнений и подкрепления вследствие одних лишь наблюдений за действием "актера" также научаются решать поставленную.ему задачу, то можно говорить о факультативном имитационном научении и даже об имитационном решении задач. Так, например, в экспериментах с обезьянами все участвующие в опыте животные устремляются к месту его проведения и внимательно наблюдают, как вожак добывает и получает подкрепление. В итоге соответствующие навыки формируются у всех "зрителей", что проявляется в успешном решении ими той же задачи в отсутствие вожака.
На этом примере видно, как проявляется еще одна особенность, характерная для проявлений подражания в естественных условиях. Дело в том, что наряду с взаимным стимулированием к совместному выполнению определенных (инстинктивных) действий в сообществах животных действует и фактор противоположного рода - подавление действий членов сообщества "господствующими" особями. Так, в приведенном примере обезьяны боялись вплотную подойти к экспериментальной установке, а тем более брать пищу из кормушки. Вместе с тем у обезьян существуют особые "примиряющие" сигналы, оповещающие доминирующую особь о готовности "зрителей" "только смотреть", чем обеспечивается возможность осуществления аллеломиметического поведения и имитационного научения. Отсюда явствует, что явления подражания сложным образом переплетаются с внутригрупповыми отношениями животных.
Подводя итог обзору инстинктивного поведения и научения у животных, вернемся еще раз к описанной раньше общей схеме поведенческого акта. Эта схема, разумеется, не отражает всего многообразия вариантов поведенческих актов в их конкретных проявлениях.
Но во всех этих вариантах остается в силе основная закономерность: каждое действие животного начинается с внутреннего стимула, который в виде потребности активирует животное, кладет начало его поиску раздражителей. Это начало, как и общее направление этого поиска, всегда генетически фиксировано, равно как и конец - заключительные движения животного в завершающей фазе. Как пройти этот путь от начала до конца, между этими "жестко запрограммированными" основными элементами поведения, зависит от лабильных компонентов поведения животного, от его умения быстро и правильно ориентироваться во всех конкретных ситуациях, в которые оно попадает. Как мы видели, особое значение это имеет в начале пути.
У высших животных факультативное научение, сопряженное с формированием операций, является основным средством для продвижения к завершающей фазе. Но поскольку операция направлена на преодоление преграды, т. е. условий, в которых дан объект завершающего поведения, то здесь особенно важна более полноценная по возможности ориентировка животного во времени и пространстве. Ясно, что эта ориентировка будет тем более совершенной, чем выше уровень психической деятельности животного, его познавательные способности. Наибольший эффект обеспечивают в этом отношении высшие психические функции, особенно интеллектуальные способности, придающие всему поведению животного наибольшую гибкость и адаптивность. К этому вопросу мы еще вернемся при рассмотрении эволюции психики.
Явления подражания играют существенную роль в формировании поведения животных. Подражание может иметь инстинктивную природу, или относиться к сфере научения. Для инстинктивного подражания характерно то, что выполнение видотипичных действий одним животным является побуждающим фактором для выполнения аналогичных действий другими животными данного вида.
Иными словами в этом случае имеет место взаимная стимуляция или взаимопоощрение видотипичной деятельности. Такая форма поведения носит название аллеломиметическое поведение. Примерами такого поведения являются стимуляция сна, отдыха, передвижения или видотипичного манипулирования, когда поведение одной или нескольких особей вызывает аналогичное поведение всего сообщества. Аллеломиметическое поведение изучается путем раздельного обучения изолированных друг от друга животных, которые затем сводятся вместе, в результате можно судить о силе аллеломиметической реакции, то есть инстинктивного подражания.
Научение, которое осуществляется за счет подражания, называется – имитационным научением. При имитационном научении формирование новых индивидуальных форм поведения, происходит на основе общения, то есть путём одного лишь непосредственного восприятия действий других животных. Имитационное научение, как и всякое научение вообще делится на облигатное и факультативное.
Облигатное имитационное научение наиболее типично для молодых животных, когда они осваивают путём подражания характерные для своего вида поведенческие реакции, результат такого научения обычно не выходит за рамки видового стереотипа. В результате имитационного научения животное научается выполнять некоторые жизненно необходимые действия, например, распознавать сородичей, хищников, добычу и так далее. Так, например, у молодых рыб защитная реакция на появление хищника вырабатывается в результате подражания поведению других рыб при виде поедания хищником членов стаи. К облигатному имитационному научению относится и освоение птенцами видотипичных звуков путём подражания пению взрослых особей. Однако с таким же успехом эти птенцы смогут освоить и пение других видов, если будут слышать его с раннего детства. Результатом облигатного имитационного научения является и накопление опыта в гнездостроении.
Факультативное имитационное научение в простейших формах представлено в имитации невидотипичных действий на основе облигатного (аллеломиметического) стимулирования. Здесь можно выделить имитационное манипулирование. В качестве примера данной формы научения можно привести освоение обезьяной действий человека с предметами быта или инструментами. Эти навыки хотя и вырабатываются в результате имитации, но не являются характерными для данного вида. Высшим проявлением факультативного имитационного научения считается имитационное решение задач. При этом у животного – "зрителя" вырабатывается определенный навык за счет одного лишь созерцания действий другой особи решающей определенную задачу. Эта форма научения установлена у разных млекопитающих: мышей, кошек, собак и др., но особенно большую роль она играет у обезьян, у которых формирование сложных условных рефлексов происходит преимущественно на основе подражания вожаку или другим более опытным особям. В заключении следует отметить, что явления подражания сложным образом переплетаются с внутригрупповыми отношениями животных.
Имитационное поведение дельфинов
Одним из наиболее интересных и биологически важных видов группового поведения является подражание, или имитация. Имитационное поведение широко распространено в животном мире и играет существенную роль в приспособлении отдельных особей и вида в целом к меняющимся условиям среды. У позвоночных животных подражание является мощным цементирующим фактором в стадной жизни животных и имеет большое биологическое значение, поскольку служит основой накопления индивидуального опыта. Л.А. Орбели (1949) указывал на большое эволюционное значение имитационного поведения, считая его «главным охранителем вида».
Исследователями выделяются две основные категории подражания у животных: стимулирование видотипичных действий и научение путем подражания или истинное имитационное поведение (Andrew, 1962; Фабри, 1974). Взаимное стимулирование видотипичных, врожденных действий широко распространено в животном мире как элемент группового поведения. В этологической литературе взаимное стимулирование животных обозначается как аллеломиметрическое поведение. Аллеломиметрическое поведение проявляется в наиболее простой форме — рефлексе следования. Оно обнаруживается у стайных рыб, птиц, копытных, хищных млекопитающих, приматов. Одновременный отдых, собирание пищи, бегство или комфортное поведение животных также являются примерами аллеломиметрического поведения. Результатом этих действий является согласованность, унификация и синхронизация поведения членов стада, что обеспечивает безотлагательное бегство всех животных стада в опасных ситуациях.
Подлинное подражание, или «имитационное научение» (Штодин, 1947; Кряжев, 1955), отличается от аллеломиметрического поведения тем, что животное обучается перенимать те формы поведения, которые наблюдает у другой особи своего или другого вида. Имитационное научение проявляется в виде подражания тем формам поведения, которые не входят в репертуар врожденной видовой деятельности наблюдателя. Случаи истинной имитации известны для птиц, когда они подражают песне другого вида или имитируют человеческий голос. Примеры обучения путем наблюдения известны у рыб (Козаровицкий, 1961), у крыс (del Russo, 1975), кошек (John et al., 1968), но наивысшими способностями в этом отношении обладают обезьяны (Воронин, 1947; Кряжев, 1955; Фабри, 1965; Riopelle, Hill, 1973; и др.).
Формирование и проявление различных видов подражательного поведения в сообществах животных находятся в непосредственной связи с иерархической организацией взаимоотношений членов данной группы. Влияние доминирующих особей в большой степени отражается на проявлениях подражания в сообществах у обезьян (Menzel, 1971; Воронин, 1947; Фабри, 1965). Так, опыты Е. Мензеля (Menzel, 1971) с шимпанзе показали, что группа обезьян, как правило, следует за лидером, которому известно, где находится тайник с лакомствами, и обычно игнорирует попытки низкорангового животного увлечь за собой сородичей к известному ему источнику пищи.
Таким образом, подражание широко распространено в животном мире и играет существенную роль в групповых отношениях разных видов. Вместе с тем наивысшего развития имитационное поведение достигает у приматов и китообразных. Стадная жизнь большинства видов зубатых китообразных связана с развитием у них разных форм подражательного поведения. Многие примеры проявления стадного инстинкта или реакции следования наблюдаются у китообразных в природе.
Одно из самых загадочных явлений в жизни китообразных — групповое самовыбрасывание животных на берег — рассматривается (Norris, 1967; Томилин, 1969) как проявление реакции следования за гибнущим по той или иной причине членом своего стада. В отношении причин самовыбрасывания китообразных высказываются различные предположения: ошибки в работе эхолокационного аппарата, плохо срабатывающего в условиях мелководья и мутной воды (Dudok van Heel, 1962), неизвестные массовые смертельные заболевания, психические расстройства (Geraci, 1978; и др.). Однако ни одна из этих гипотез не получила окончательного подтверждения. Подробное описание наблюдений за выбрасыванием стада гринд у берегов Флориды приводится в работе В. Феринга, Р. Велса (Fehring, Welb, 1976). Группа дельфинов, состоящая из 49 животных, на протяжении четырех дней несколько раз оказывалась на отмели. Операция по спасению китов крайне затруднялась тем, что отбуксированные в море животные вновь выбрасывались на берег. Все стадо удалось вернуть только после того, как далеко в море были отбуксированы два крупных животных, видимо, лидеров стада. После этого остальные дельфины прекратили попытки, выбрасываться на берег и стадо объединилось. Однако когда одного из молодых дельфинов спасенного стада решили взять для океанариума и за веревку, привязанную к его хвосту, начали подтаскивать к берегу, вновь все стадо последовало за ним. Отдельные животные стали выбрасываться на берег, разбиваясь о каменные гряды. Даже после того как молодую гринду оставили, некоторое время дельфины продолжали выбрасываться на берег. Но и в этот раз удалось спасти оставшихся в живых гринд, отбуксировав их вместе с лидером далеко в море. Спустя 4 дня часть этого стада (13 животных) были найдены обсохшими в 275 км от последнего места их обсыхания. Как сообщают авторы, море в эти дни было спокойным и вода прозрачной, что, видимо, исключает ошибки в работе эхолокационного аппарата. Среди выбросившихся животных не наблюдалось паники — они довольно спокойно двигались к берегу и не производили впечатления больных. Таким образом, во время спасательных операций реакция следования проявилась у гринд дважды, когда все стадо ушло в море за лидирующими животными и когда оно последовало за буксируемым к берегу молодым животным.
Более сложный вид подражания — обучение путем наблюдений, или истинную имитацию, исследовали у многих видов зубатых китообразных в неволе: у атлантических афалин (McBride, Kritzler, 1951; Caldwell et al., 1965), у индийских афалин (Тауler, Saayman, 1973), у гринды (Kritzler, 1952), у малой косатки (Brown et al., 1966), у амазонского дельфина (Pilleri et al., 1980).
Наиболее обстоятельное изучение различных форм моторного имитационного поведения у афалин проведено С. Тайлером, Г. Сайманом (Tayler, Saayman, 1973). Две взрослые самки афалины содержались в одном бассейне со взрослым самцом морского котика (Arctocephalus pusillus); в этом же бассейне находилось несколько пингвинов, черепахи и скаты.
В первые месяцы их совместной жизни обе самки избегали котика, но постепенно степень их контактов стала возрастать, в результате чего в поведении афалин появились многие элементы поведения, присущие котику во время сна, груминга и активного плаванья. Так, дельфины стали плавать, подгребая плавниками как ластами, а хвостовой плавник в это время был совершенно неподвижен.
Как известно, дельфины те занимаются грумингом. Однако, наблюдая эту форму поведения у котика, они переняли многие позы и действия, связанные с грумингом. Котики с помощью передних и задних ласт разглаживают и очищают свой мех, при этом они принимают разные позы: вращаются вокруг собственной оси у поверхности воды или принимают вертикальную позу выставив из воды только задние ласты. В это время к поверхности поднимаются пузырьки воздуха, выдыхаемого котиком. Дельфины, подражая котику, с помощью грудных плавников научились потирать живот, лежа на спине у поверхности воды, принимать вертикальную позу, выставив хвостовой плавник и выпуская при этом пузырьки воздуха из дыхала. Выныривая на поверхность, они широко раскрывали рот, подражая зевоте котиков, хотя известно, что зевота не свойственна дельфинам.
В первое время своего пребывания в бассейне самки афалин плавали даже во время ночного отдыха. Котики же спали неподвижно у поверхности воды, лежа на одном боку, при этом задние ласты выставлены из воды; они имели дугообразную форму. Иногда они лежали на поверхности на спине, скрестив ласты на брюхе. Спустя некоторое время дельфины переняли позы котика, которые он принимал во время сна. Стали замечать, что время от времени афалины неподвижно лежали у поверхности на боку или на спине, прижав плавники к брюху и выставив хвостовой плавник из воды. Хотя эти позы были неудобны афалинам, они часто их принимали, находясь рядом со спящим котиком. Следует отметить, что котик не перенимал поз и движений дельфинов.
Дельфины разных видов, содержащихся в одном бассейне, часто перенимают друг у друга некоторые формы поведения. Так, Д. Браун и др. (Brown et al., 1966) описывают имитацию прыжков вертящегося дельфина (Stenella lingirostris) афалинами, которые содержались в том же бассейне. Гринда пыталась балансировать на носу надувную игрушку, подобно тому как это делали афалины. Но поскольку у гринд отсутствует вытянутый рострум, она манипулировала игрушкой с помощью головы.
Некоторые виды деятельности дельфины, главным образом афалины, могут перенимать у человека. Так, С. Тайлер, Г. Сайман (Tayler, Saayman, 1973) описывают некоторые виды орудийной деятельности афалин, которую они наблюдали у людей. Во время уборки бассейна афалины проявляли большой интерес к этой процедуре. Прибор для уборки бассейна состоял из полого скребка со шлангом, куда втягивался мусор, который собирали со дна бассейна с помощью скребка. Однажды аппарат был оставлен на ночь в бассейне. Им завладела одна самка и долго с ним манипулировала, таская во рту по бассейну. Затем, подражая действиям человека, она, придерживая шланг грудными плавниками, взяла в рот совок и стала водить им по дну бассейна. Когда аппарат убрали, обе самки стали использовать для «уборки» бассейна куски кафельной плитки (6x8 см), зажимая их зубами. Действуя сообща, они научились с помощью кафельных плиток собирать в кучу куски морских водорослей (Enteromorpha), которые они употребляли в пищу. Эти примеры использования орудий как в форме игры (чистка бассейна), так и для сбора пищевых объектов свидетельствуют о том, что они могут не только копировать определенные виды деятельности человека, но и модифицировать их сообразно своим потребностям.
Имитация человеческой деятельности свойственна не только афалинам, но и речным дельфинам. В гл. 2 мы приводили примеры того, как амазонские дельфины, подражая человеку, «чистили» стенки бассейна и окна с помощью небольших щеточек, которые они держали во рту (Pilleri et al., 1980).
Было проведено (Adler, Adler, 1978) экспериментальное исследование способности афалин к обучению путем наблюдений на основе использования инструментальной условнорефлекторной методики. Опыты были проведены на трех афалинах, которые находились в круглом бассейне, разделенном на четыре отсека крупноячеистой сеткой. Дельфинам предложили четыре задачи: втягивание свисающего конца веревки в бассейн; дифференцирование двух квадратов с горизонтальной и вертикальной исчерченностью; дифференцировку двух квадратов красного и зеленого цвета; различение цели по ее местоположению (на левой или правой стороне). Все цели предъявлялись животным над водой.
С одним из дельфинов — «демонстратором» проводили эксперимент, а два других находились в это время в соседних отсеках и могли наблюдать все, что происходит в экспериментальном отсеке. После закрепления навыка у «демонстратора» одного из «наблюдателей» переводили в экспериментальный отсек и проводили с ним процедуру обучения. В опытах те же дельфины были и в роли «демонстраторов» и в роли «наблюдателей». В последней задаче «наблюдателя» помещали в экспериментальную камеру спустя четыре дня после проведения опытов с «демонстратором».
Оценка результатов опытов показывает, что дельфины могут обучаться сложным задачам дифференцировать цели путем наблюдений даже с отсрочкой на несколько дней. У дельфинов-наблюдателей успешное решение задачи происходило уже с первых предъявлений условных раздражителей. В то же время для обучения дельфина-демонстратора требовалось большее число тренировок. У «наблюдателей» по сравнению с «демонстраторами» заметно сокращался латентный период ответов (от 64 до 25 с).
Авторы полагают, что наблюдения за действиями «демонстратора» уже сами по себе являются для дельфинов положительным подкреплением всей ситуации и снижают у них пассивно-оборонительную реакцию перед незнакомыми предметами, которая в сильной степени развита у этих животных. Л. Г. Воронин (1977) также считает, что подражательный рефлекс способствует приучению обезьян к экспериментальной обстановке, сокращая начальный период формирования условнорефлекторной деятельности.
Приведенные примеры демонстрируют высокоразвитую способность дельфинов к воспроизведению сложных моторных действий, но вместе с тем у них в равной степени развита и «вокальная» имитация.
Исследования показали, что дельфины способны имитировать широкий диапазон самых разнообразных звуков — от булькающих звуков поднимающихся пузырьков воздуха из аквалангов до отдельных слов и простых фраз, произносимых человеком (Lilly, Miller, 1962; Caldwell, Caldwell, 1972a; Beach, Papper, 1973; Tayler, Saayman, 1973; Тарчевская и др., 1978; Richard et al., 1981).
Изучение имитации тональных сигналов с частотной модуляцией у афалин было проведено (Тарчевская и др., 1978) на двух взрослых афалинах. Эталонные сигналы с частотной модуляцией от 5 до 10 кГц, близкие к естественным свистам дельфинов, транслировались под водой. Оценка сходства эталонных сигналов с сигналами дельфинов осуществлялась на экране осциллографа. Было выяснено, что дельфины имитировали эталонные сигналы свистами, превышающими последние по частоте в 1,2 — 1,8 раз. Иногда дельфины могли повторять эталонный сигнал два-три раза в пределах одного имитационного. Наблюдалось как непосредственное повторение только что проигранного сигнала, так и постепенное преобразование собственных свистов в имитационные.
Недавно проведенные исследования способности афалин к имитации искусственных свистовых сигналов (Richard et al., 1981) показали, что они легко справляются с этой задачей. Самец афалина, имевший до опытов определенный набор естественных свистовых сигналов, которые часто и стереотипно повторялись, был обучен имитировать различные вариации звуковых «моделей», излучаемых генератором. Все предложенные искусственные звуки дельфин научился точно воспроизводить, используя свои «вокальные» возможности. Эти звуковые сигналы отличались от его естественных свистов. При проведении опытов авторы отметили большие возможности дельфина к обобщению, что позволило ему воспроизводить новые сигналы после 1 — 3 предъявлений. Скорость обучения воспроизведению искусственных звуковых сигналов свидетельствует, по мнению авторов, о высокоразвитой способности дельфинов оперировать большим объемом слуховой информации, а способность к «вокальной» имитации играет существенную роль при установлении коммуникативных связей в их природных сообществах.
Широко известные эксперименты Дж. Лилли (Lilly, 1962), целью которых было изучение способов установления контактов между человеком и дельфинами, показали, что дельфины обладают чрезвычайно развитыми «вокальными» подражательными способностями. Опыты Дж. Лилли подтвердили, что афалина может в какой-то степени подражать человеческому голосу и после обучения способна воспроизводить слоги, отдельные слова, простые фразы, восклицания, смех. В своих экспериментах в качестве подкрепления он применял электрическое раздражение подкорковых структур мозга, так называемых «центров удовольствия».
Сравнение виброграмм с записью голосов дельфинов и человека подтвердило звукоподражание афалины. Человек произносил слова в пределах колебаний от 200 до 3 000 Гц, а дельфин воспроизводил их в частотах от 1 000 до 8 000 Гц. Если магнитофонную ленту с голосом дельфина, имитировавшим слова человека, пропускать с замедленной в 2 — 4 раза скоростью, то воспроизводится известное сходство с голосом человека. Дальнейшие эксперименты Дж. Лилли по обучению дельфинов подражанию человеческой речи, как известно, не подтвердили ожиданий автора о возможности установления сознательных контактов между дельфином и человеком. Тем не менее эти опыты продемонстрировали большие возможности дельфинов в сфере имитации «гомоноидных» звуков, что само по себе можно рассматривать как большую удачу исследователя.
Интересные исследования взаимоотношений дельфинов в группе при выработке у них пищедобывательных условных рефлексов и некоторых форм имитационного поведения проведены Л. Г. Ворониным и др. (1971, 1974). Группа состояла из трех черноморских афалин: двух самцов и одной самки. Один самец занимал доминирующее положение в группе. Было проведено несколько серий экспериментов, где выяснилось распределение между дельфинами манипуляторов, при воздействии на которые они сразу получали рыбу. В первой серии опытов дельфины могли нажимать на три рычага, во второй серии им оставляли только один рычаг, а в третьей — получение рыбы и манипуляторы были пространственно разделены, т. е. нажав на рычаг, дельфин должен был плыть за рыбой в другой конец бассейна.
В результате проведенных экспериментов было выяснено, что при наличии трех рычагов внимание дельфинов привлекал в основном тот, который выбирал доминантный самец. Второй самец предпочитал подбирать рыбу, которая оставалась от работы доминанта. Насытившись, доминант охотно работал для второго самца, усердно нажимая на рычаг, и отдавал ему всю рыбу. Самка сначала без всякого предпочтения нажимала на разные рычаги, но затем чаще стала пристраиваться к самцам и работала обычно в паре с доминантой.
При ситуации, когда оставляли только один рычаг, все животные поочередно на него нажимали, но чаще других доминантный самец. Несколько раз доминант отгонял второго самца от манипулятора и преследовал его с агрессивными намерениями. В том случае, когда работа с манипулятором вознаграждалась в другом месте, чаще проявлялось деспотическое поведение доминирующего самца. Он отгонял не только второго самца, но и самку; установилась очередь в работе: сначала к манипулятору подходил только доминант, затем к добыванию пищи присоединялся второй самец, потом к работе приступала самка.
Из приведенной работы следует, что конкурентные отношения между доминантой и подчиненным дельфином возникали не по поводу получения рыбы, а из-за доступа к рычагу, т. е. способу добывания пищи. Таким образом, активный вид деятельности, приводящий к добыванию пищи, является, по-видимому, прерогативой доминирующего животного в сообществе дельфинов. Рефлекс подражания у дельфинов данной группы проявился в том, что наибольшее внимание животных привлекает тот манипулятор, который выбирал доминирующий самец. Два «свободных» рычага мало интересовали дельфинов, хотя с их помощью они также могли добывать пищу.
Обсуждая результаты проведенных экспериментов, авторы отмечают миролюбие и отсутствие жестких конкурентных отношений дельфинов в условиях группового эксперимента. Редко возникающие агрессивные стычки очень быстро сменялись миролюбивыми взаимоотношениями. Способность доминирующего самца нажимать на рычаг после собственного насыщения и таким образом «кормить» другого дельфина рассматривается авторами как проявление сотрудничества в сообществе дельфинов.
На основании приведенных в этом разделе данных можно заключить, что у дельфинов в сильной степени развиты различные формы подражательной деятельности, которые проявляются в виде реакции следования за лидерами, синхронного плавания, а также в более сложных формах научения путем подражания. Дельфины имитируют движения, позы и звуки не только у особей своего вида, но и у животных, далеко отстоящих от них в таксономическом отношении. Некоторые формы орудийной деятельности, которые они могут перенимать у человека, можно рассматривать одним из показателей их развитого интеллекта. Среди млекопитающих по развитию имитационных способностей дельфины находятся на том же уровне, что и человекообразные обезьяны.
Несомненно, развитая способность к сложным формам подражания играет весьма существенную роль в социальной жизни дельфинов. Этот вопрос вызывает большой интерес у исследователей. Л. Герман (Herman, 1980) полагает, что способность дельфинов к «вокальной» и моторной имитации является той основой, на которой развиваются различные формы координированной групповой активности: синхронное плавание, коллективная охота, защитное поведение. Интересную мысль по поводу функциональной роли имитационного поведения дельфинов в их общественной жизни высказывают С. Тайлер, Г. Сайман (Tayler, Saayman, 1973). Авторы предполагают, что имитация половых форм поведения в период ухаживания может играть существенную роль при выборе полового партнера. Синхронность выполнения ритуальных поз ухаживания и координация движений во время совместного плавания, по мнению авторов, может служить стимулом для селективного спаривания и для упрочения социальных связей в сообществе. Другими словами, в каждом сообществе могут существовать собственные «традиции» половых взаимоотношений, которые сохраняются и усиливаются в результате подражания друг другу.
Развитая способность к «вокальной» имитации у дельфинов, по мнению Е. Вильсона (Wilson, 1975), может быть причиной большого сходства опознавательных сигналов у животных, принадлежащих к одному сообществу, что дает им возможность быстро узнавать своих сородичей.
Известно, что у афалин обнаружены (Caldwell, Caldwell, 1972а) устойчивые индивидуальные «свисты-автографы», представляющие собой определенные контуры частотных сигналов. Они служат опознавательными сигналами для каждого члена группы. Вместе с тем можно предположить, что физические характеристики этих «свистов-автографов» у членов одного сообщества в результате подражания могут приобрести общие черты, или свойства, отличающие их от подобных индивидуальных сигналов другого сообщества. По этим сигналам дельфины могут узнавать своих соплеменников и отличать особей, принадлежащих к другим сообществам.
Приведенные выше предположения исследователей о роли имитационных рефлексов в социальной жизни дельфинов весьма интересны, но требуют экспериментальных подтверждений.
Источник:
Последнее редактирование: 07.03.2024
Чтобы оставить комментарии нужно авторизоваться.