Вместо вступления (разминка).
Как это принято писать в начале научных статей, несмотря на очевидный прогресс в области понимания механизмов происхождения жизни, в данном вопросе по-прежнему существует много белых пятен. В данной статье я попытаюсь прояснить некоторые из них, ну а уж как у автора это получится, судить читателям. Для начала стоит отметить, что несколько упрощая, можно выделить два базовых подхода к решению проблемы понимания процесса происхождения жизни. В одном из них учёные сначала сами разбивают весь этот сложный процесс на ряд более простых (с их точки зрения, естественно) этапов, а потом пытаются воспроизвести эти этапы в, так сказать, "идеальных лабораторных условиях". По другому этот подход можно сформулировать как попытку вначале, исходя из некоторой внутренней логики, убрать из каждого предполагаемого этапа всё лишнее, мешающее добиться предполагаемого результата, а уже потом, когда результат на каждом отдельном шаге будет успешно достигнут, начать попытки соединения всех этих этапов в единую непрерывную цепь от исходной смеси на входе до искомого продукта на выходе. В общем, такой своеобразный "научный дизайн".  Альтернативный, так сказать, "синергетический" подход основан на принципе исходного воспроизводства процесса в условиях, максимально приближенных к боевым реальным (хотя, по правде говоря, каковы были эти "реальные условия" у учёных до сих пор тоже нет единодушия). То есть, даже если некий элемент "бульона" кажется лишним, или даже мешающим, но, судя по всему, он был широко представлен в той среде, в которой зарождалась жизнь, не убирать его оттуда, что бы "помочь" процессу зарождения жизни, а оставить в качестве её компонента, авось пригодится. Любопытно отметить, что именно второй подход относительно недавно помог совершить прорыв в вопросе абиогенного синтеза пиримидиновых нуклеотидов. Вплоть до последнего времени химики пытались вначале по отдельности синтезировать из простейшей органики азотистые основания и рибозу, а уже потом соединить их между собой и с фосфатом для получения соответствующих нуклеиновых оснований, "строительных кирпичиков", из которых построена РНК. Но долгое время ничего путного не получалось, если что-то и синтезировалось, то с очень низким выходом. И вот, английские химики решили, что раз по отдельности "крокодил не ловится, не растёт кокос", попробовать "бросить в бульон" сразу горсть простых органических молекул и фосфата, пусть эта затея и казалось на первый взгляд совершенно бесперспективной. И это сработало! Оказалось, что "бульону", содержащему смесь довольно простых органических веществ, нужно лишь дать немного настояться, а потом слегка подогреть и посветить на него ультрафиолетом, образно выражаясь, до полной готовности, и, вуаля, в нём в массовом порядке обнаруживаются сразу два из четырёх "основных" для жизни нуклеотидов - пиримидиновые нуклеотиды цитидин и уридин. Более того, другие "неканонические" нуклеотиды после ультрафиолетовой "стерилизации" разрушаются. Итак, пара компонентов, которые могли в ощутимой концентрации присутствовать в исходной среде, у нас уже есть. Что там ещё могло быть интересного с точки зрения перспективности для происхождения жизни? По этому вопросу у химиков царит практически полное единодушие, конечно аминокислоты! Ещё со времён классических опытов Миллера, впоследствии многократно повторённых в различных вариантах, в восстановительной среде, содержащей простейшие молекулы типа водорода, аммиака, угарного газа, воды и метана, при условии подвода энергии в той или иной форме неизменно синтезировалось около десятка различных аминокислот, причём, четыре наиболее простых их них - валин, аланин, аспарагиновая кислота (аспарат) и глицин синтезировались в наибольших количествах. Так что, смело добавляем в наш условный бульон указанные аминокислоты. Теперь посмотрим, какие ещё упрощения принимают большинство биохимиков, когда они пытаются моделировать ранние этапы жизни, в частности, так называемый "Мир РНК"? Одно из самых существенных из них заключается в том, что априори предполагается, что нуклеотиды при полимеризации в цепочки всегда соединяются одним и тем же способом, 5-й атом углерода рибозы через ортофосфорную кислоту соединяется с 3-м атомом углерода рибозы следующего нуклеотида. Но таких "идеальных" цепочек РНК ни в одном из опытов без помощи взятых "напрокат" у реальных клеток сложных молекулярных машинок (полимераз) никому из химиков синтезировать никогда не удавалось. В реальности цепочка всегда содержит смесь соединённых через 2-й и 3-й атомы углерода рибозы нуклеотидов. Давайте вместо того, что бы закрывать на эту неприятную проблему глаза, с самого начала предположим, что изначально после полимеризации у полинуклеотида связи образуются по сути случайным образом. Может быть, опять "тот кто нам мешает, тот нам и поможет?".
Альтернативный, так сказать, "синергетический" подход основан на принципе исходного воспроизводства процесса в условиях, максимально приближенных к боевым реальным (хотя, по правде говоря, каковы были эти "реальные условия" у учёных до сих пор тоже нет единодушия). То есть, даже если некий элемент "бульона" кажется лишним, или даже мешающим, но, судя по всему, он был широко представлен в той среде, в которой зарождалась жизнь, не убирать его оттуда, что бы "помочь" процессу зарождения жизни, а оставить в качестве её компонента, авось пригодится. Любопытно отметить, что именно второй подход относительно недавно помог совершить прорыв в вопросе абиогенного синтеза пиримидиновых нуклеотидов. Вплоть до последнего времени химики пытались вначале по отдельности синтезировать из простейшей органики азотистые основания и рибозу, а уже потом соединить их между собой и с фосфатом для получения соответствующих нуклеиновых оснований, "строительных кирпичиков", из которых построена РНК. Но долгое время ничего путного не получалось, если что-то и синтезировалось, то с очень низким выходом. И вот, английские химики решили, что раз по отдельности "крокодил не ловится, не растёт кокос", попробовать "бросить в бульон" сразу горсть простых органических молекул и фосфата, пусть эта затея и казалось на первый взгляд совершенно бесперспективной. И это сработало! Оказалось, что "бульону", содержащему смесь довольно простых органических веществ, нужно лишь дать немного настояться, а потом слегка подогреть и посветить на него ультрафиолетом, образно выражаясь, до полной готовности, и, вуаля, в нём в массовом порядке обнаруживаются сразу два из четырёх "основных" для жизни нуклеотидов - пиримидиновые нуклеотиды цитидин и уридин. Более того, другие "неканонические" нуклеотиды после ультрафиолетовой "стерилизации" разрушаются. Итак, пара компонентов, которые могли в ощутимой концентрации присутствовать в исходной среде, у нас уже есть. Что там ещё могло быть интересного с точки зрения перспективности для происхождения жизни? По этому вопросу у химиков царит практически полное единодушие, конечно аминокислоты! Ещё со времён классических опытов Миллера, впоследствии многократно повторённых в различных вариантах, в восстановительной среде, содержащей простейшие молекулы типа водорода, аммиака, угарного газа, воды и метана, при условии подвода энергии в той или иной форме неизменно синтезировалось около десятка различных аминокислот, причём, четыре наиболее простых их них - валин, аланин, аспарагиновая кислота (аспарат) и глицин синтезировались в наибольших количествах. Так что, смело добавляем в наш условный бульон указанные аминокислоты. Теперь посмотрим, какие ещё упрощения принимают большинство биохимиков, когда они пытаются моделировать ранние этапы жизни, в частности, так называемый "Мир РНК"? Одно из самых существенных из них заключается в том, что априори предполагается, что нуклеотиды при полимеризации в цепочки всегда соединяются одним и тем же способом, 5-й атом углерода рибозы через ортофосфорную кислоту соединяется с 3-м атомом углерода рибозы следующего нуклеотида. Но таких "идеальных" цепочек РНК ни в одном из опытов без помощи взятых "напрокат" у реальных клеток сложных молекулярных машинок (полимераз) никому из химиков синтезировать никогда не удавалось. В реальности цепочка всегда содержит смесь соединённых через 2-й и 3-й атомы углерода рибозы нуклеотидов. Давайте вместо того, что бы закрывать на эту неприятную проблему глаза, с самого начала предположим, что изначально после полимеризации у полинуклеотида связи образуются по сути случайным образом. Может быть, опять "тот кто нам мешает, тот нам и поможет?".  Буквально в прошлом году вышла статья об очень важном эксперименте, проделанном командой, состоящей из американских и английских учёных. Они взяли один из самых известных рибозимов, обладающих каталитической активностью, так называемый hammerhead ribozyme, способный вырезать определённый фрагмент собственной полинуклеотидной цепочки, и изменили у случайно выбранных 25% нуклеотидов стандартную связь через 3-й атом углерода на нестандартную связь через 2-й атом углерода, в некоторой степени как бы имитируя тем самым первые синтезированные абиогенно РНК олигонуклеотиды. При этом выяснилось, что во-первых, модифицированный подобным образом рибозим сохранил свою каталитическую активность, а во-вторых, его температура плавления (то есть, та температура, при которой спаренные комплиментарные цепочки начинают расходиться, делая тем самым возможным механизм так называемой неферментативной репликации РНК) ощутимо понизилась. Всё это делает гораздо более реалистичным сценарий при котором первые синтезированные абиогенно РНК-цепочки, состоящие из случайной смеси связанных между собой как через 3-й, так и через 2-й атомы углерода нуклеотидов при понижении температуры (например, ночью) катализировали некоторые предбиологические химические реакции, а при её повышении (например, днём) водородные связи, формирующие их вторичную структуру (и связывающие синтезированные на предыдущем этапе взаимно комплиментарные цепочки) разрывались, обеспечивая тем самым условия для размножения рибозимов. Как видим, в проблеме происхождения жизни многое не случайно, так что, по-видимому, учёным ещё многому придётся научиться у природы, во многих вопросах она проявляет себя гораздо более продвинутым дизайнером, чем "доценты с кандидатами".
Буквально в прошлом году вышла статья об очень важном эксперименте, проделанном командой, состоящей из американских и английских учёных. Они взяли один из самых известных рибозимов, обладающих каталитической активностью, так называемый hammerhead ribozyme, способный вырезать определённый фрагмент собственной полинуклеотидной цепочки, и изменили у случайно выбранных 25% нуклеотидов стандартную связь через 3-й атом углерода на нестандартную связь через 2-й атом углерода, в некоторой степени как бы имитируя тем самым первые синтезированные абиогенно РНК олигонуклеотиды. При этом выяснилось, что во-первых, модифицированный подобным образом рибозим сохранил свою каталитическую активность, а во-вторых, его температура плавления (то есть, та температура, при которой спаренные комплиментарные цепочки начинают расходиться, делая тем самым возможным механизм так называемой неферментативной репликации РНК) ощутимо понизилась. Всё это делает гораздо более реалистичным сценарий при котором первые синтезированные абиогенно РНК-цепочки, состоящие из случайной смеси связанных между собой как через 3-й, так и через 2-й атомы углерода нуклеотидов при понижении температуры (например, ночью) катализировали некоторые предбиологические химические реакции, а при её повышении (например, днём) водородные связи, формирующие их вторичную структуру (и связывающие синтезированные на предыдущем этапе взаимно комплиментарные цепочки) разрывались, обеспечивая тем самым условия для размножения рибозимов. Как видим, в проблеме происхождения жизни многое не случайно, так что, по-видимому, учёным ещё многому придётся научиться у природы, во многих вопросах она проявляет себя гораздо более продвинутым дизайнером, чем "доценты с кандидатами".
Почему вообще стало возможным зарождение жизни и её последующая эволюция? (по большей части философия, желающие могут пропустить этот раздел без ущерба для понимания последующего текста).
Теперь давайте немного отвлечёмся от биохимии и чуть-чуть коснёмся проблем комбинаторики. Дело в том, что количество вариантов, которые должна была перебрать природа, прежде чем выпало "счастливое число", давшее ей входной билет на сцену дарвиновской эволюции, уже давно является предметом яростных споров между креационистами всевозможного рода и последователями уникальности земной жизни во Вселенной (в данном случае они сидят в одной лодке) с одной стороны, и сторонниками широкой распространённости в ней живых систем с другой стороны. Итак, предположим, что мы хотим оценить вероятность реализации некой уникальной последовательности событий. Если количество исходов для конкретного события равно M, а длина цепочки равна N, то в случае независимости между собой указанных необходимых событий итоговая вероятность может быть вычислена по формуле Pfinal= M-N. Как эту сухую формулу пощупать руками? Предположим, что мы оцениваем вероятность сборки конкретной пептидной цепочки длиной в 10 аминокислотных остатков (это близко к минимальной длине крошечных белков, которые всё ещё способны проявлять некоторую каталитическую акивность). Подставляя в формулу M= 20 (количество реально используемых всеми земными организмами универсальных аминокислот) получим Pfinal= 20-10≈ 10-13. Число действительно не сильно воодушевляет, конечно, не "ужос-ужос", но всё таки... Но не всё на самом деле так плохо, во многих случаях активность фермента может сохраняться и при замещении одних аминокислот другими. Например, как выяснилось в процессе исследований, в активных центрах наиболее древних пептидов встречаются консервативные мотивы, для которых важно не столько то, какая конкретно аминокислота находится в данной позиции, сколько такое её химическое свойство, как полярность (она же гидрофобность). Таким образом, для таких случаев имеем Pfinal= 2-10≈ 10-3. Что называется, почувствуйте разницу, тысяча это уже вполне, так сказать, нестрашное, бытовое число. Примерно такая же ситуация с вариантами встречаемости уникальных цепочек полинуклеотидов. Если исходить из того, что для кодирования сразу же стали использоваться все 4 "теперешних" нуклеотида, то цепочка длиной в 10 оснований может быть закодирована 410≈ 106 способами. Может, выглядит и не так страшно, как для аминокислот, но нужно учитывать, что при увеличении длины цепочки данное число будет быстро увеличиваться. Однако и здесь, как и в случае с первыми пептидными цепочками, всё можно в конечном итоге свести к бинарному случаю. Действительно, каталитическая активность рибозима в значительной степени определяется его вторичной структурой, которая, в свою очередь, во многом зависит от того, как чередуются в ней "стебли" и "петли", то есть, склеенные между собой и разъединённые (свободно "висящие") фрагменты. При "классической" схеме соединяются между собой пары гуанин - цитозин и урацил - аденин, а все остальные сочетания пар не образуют (в реальности всё, конечно, несколько сложнее, но нам пока важно понять генеральную концепцию). Таким образом, пусть и несколько загрубляя, можно сказать, что и здесь, в принципе, всё можно глобально свести к бинарному случаю - пары нуклеотидов либо "склеиваются", либо нет. Мораль сей басни, думается, ясна, если изначально жизнь в своих "вычислениях" использовала преимущественно двоичную систему исчисления, структуры с аналогичным количеством элементов нижнего уровня могли быть достигнуты методом случайного перебора за на порядки меньшее время эволюции, чем следует из оценок некоторых креационистов.
Поехали дальше. За счёт чего жизнь сохраняется на нашей планете вот уже миллиарды лет? Ответ (естественно, несколько упрощённый) - за счёт автокаталитических реакций, непрерывно воспроизводящих составляющие её материальные элементы, и эффективного отбора, позволяющего элиминировать (редкие) ошибки копирования и адаптироваться к изменениям в окружающей среде. Но как ни крути, а самосборка прямо из "органического бульона" систем репликации ДНК-РНК и транскрипции-трансляции белков, даже если изначально они были намного проще, всё равно представляется достаточно фантастичной (если, конечно, не учитывать модель мультиверса и инфляционной Вселенной, подразумевающую одновременное существование огромного (возможно, даже бесконечного) количества альтернативных Вселенных, принципиально невидимых для нас, см. книгу Е. Кунина "The logic of chance"). Тем не менее, на мой взгляд, всё же существует вполне реалистичный путь от "бульона" до первых самореплицирующихся систем. Задумаемся, если не учитывать непрерывно самокопирующиеся системы, что ещё более-менее долго сохраняется в нашем мире? Очевидно, либо что-то "крепкое", находящееся в высокоэнтропийном состоянии с минимальной свободной энергией (для разрушения такого объекта приходится использовать достаточно большую внешнюю энергию), либо нечто, постоянно синтезирующееся, запасы которого непрерывно пополняются. Например, из всех химических элементов в нашей галактике наиболее широко представлены по своей массовой доле (в порядке убывания) водород, гелий, кислород, углерод, неон и железо. Водород и гелий присутствуют там ещё со времён так называемого Большого Взрыва, когда они синтезировались в массовом порядке. Кислород и углерод постоянно производятся внутри звёзд в результате термоядерных реакций. Неон тоже активно синтезируется в ядрах массивных звёзд на поздних стадиях их эволюции. А вот железо (точнее, его изотоп 56Fe) является элементом с самой низкой массой в расчёте на один нуклон, что в соответствии с известной формулой E= mc2 говорит о том, что у него самая большая удельная доля массы составляющих его ядро нуклонов конвертирована во внутриядерную энергию. Неудивительно, что этот "крепкий орешек" столь широко распространён во Вселенной. Если те же универсальные принципы и следующие из них результаты применить к эволюции живой природы, то очевидный вывод заключается в том, что представление о равновероятности встречаемости элементов нижнего уровня при оценке количества различных вариантов объектов верхнего уровня, которые могут быть из них собраны, в корне ошибочно. Например, разные химические элементы таблицы Менделеева представлены во Вселенной крайне неравномерно. Этот факт, а так же физико-химические законы, ограничивающие возможные сочетания атомов в молекулах, в свою очередь, диктуют очень неравномерное распределение различных веществ во Вселенной. Каких-то молекул, например, молекулярного водорода или монооксида углерода в ней много, а огромное количество формально возможных сочетаний атомов практически отсутствует вообще. Таким образом, исходная посылка, из которой часто неявно исходят при всевозможных оценках вероятности самозарождения жизни, а именно, тезис о равновероятности встречаемости элементов нижнего уровня, из которых затем строятся элементы следующего уровня иерархии, очевидно неверна. По факту на каждом уровне существует весьма ограниченное количество "выживших" в жёсткой конкурентной борьбе элементов, из которых, в свою очередь, конструируется ограниченное число перспективных в плане выживаемости элементов следующего уровня и так далее, до самого верха условной пирамиды. Таким образом, благодаря всё тому же естественному отбору элементов, ужасного комбинаторного взрыва, которым так часто кошмарят своих противников сторонники интеллектуального дизайна, по сути не происходит. В этом свете вполне естественной представляется и возможность постепенного усложнения органических систем даже на том этапе биологической эволюции, на котором ещё не заработала схема мутации при репликации плюс отбор. До определённого предела отбор может действовать в направлении облегчения процесса усложнения предбиологических систем и в одиночку. Так, вполне определённые короткие пептидные цепочки, состоящие изначально из очень ограниченного количества аминокислот, могли отбираться по признакам устойчивости к деградации в исходной среде и способности к катализу синтеза некоторых других пептидных цепочек (включая самих себя), а так же (что особенно важно) небольших цепочек РНК. В свою очередь, и вполне конкретные небольшие РНК, состоящие всего из пары десятков нуклеотидов, могли отбираться по точно таким же принципам. На следующем уровне иерархии, отобранные на предыдущем этапе короткие пептидные и полинуклеотидные цепочки, в свою очередь, могли собираться во взаимодействующие между собой структуры (включая гибридные образования, состоящие из коротких РНК "прошитых" короткими пептидными цепочками) и т.д. По сути процесс можно наглядно сравнить с постепенным появлением разговорной речи, когда вначале из всего звукового ряда формируются устойчивые образования нижнего уровня - фонемы, потом на основе сформированного ограниченного множества фонем формируется (опять же ограниченный) набор слов, которые, в свою очередь, формируют набор осмысленных предложений и т.п.
Так с чего же всё начиналось?
Всё изложенное в предыдущем разделе, это пока лишь достаточно абстрактные теоретические рассуждения, как же конкретно они могли реализоваться в процессе зарождения жизни? Начнём с очевидного предположения, что в самом низу эволюционной пирамиды должны лежать наиболее просто синтезируемые органические молекулы. С другой стороны, в силу принципа эволюционного консерватизма, предполагающего сохранение эволюционно более древних элементов нижнего уровня в составе эволюционно более "продвинутых" элементов высших уровней, можно ожидать, что химический состав базовых химических элементов жизни был выбран ещё на самых ранних этапах её эволюции. Судя по составу липидов, белков, коферментов, а так же нуклеиновых кислот, таковыми были углерод, кислород, азот, водород, фосфор, а так же (в существенно меньших количествах) сера, железо, никель и ряд других металлов. Белки состоят из легко синтезируемых в восстановительных условиях аминокислот, в составе которых встречаются все вышеперечисленные элементы кроме фосфора и металлов. Липиды, представляющие из себя достаточно простые цепочки атомов, состоящие из углерода и водорода с содержащим кислород "хвостом" на одном конце тоже легко синтезируются абиогенно. Коферменты (активные центры) как правило представляют собой либо фрагменты природных минералов (грейгит и т.д.), либо что-то вроде небольших нуклеотидно-аминокислотно-липидных комплексов. Сложнее с нуклеиновыми кислотами, каким образом они были синтезированы абиогенно до сих пор нет особой ясности. Попробуем поискать ответ в строении современных клеток. Как было недавно показано, в клетках организмов, принадлежащих всем трём доменам современной жизни на Земле (эубактерии, археи и эукариоты), присутствует одна и та же органелла, судя по всему имевшаяся уже у их последнего общего предка (ЛУКА) - асидокальциcома. Кислая среда, заполняющая внутренность данной органеллы, богата катионами кальция и полифосфатами. Любопытно, что схожий состав имеет минерал апатит, до 90% вещества которого составляют фосфаты и кальций, а так же считающийся его прекурсором брушит с химической формулой CaHPO4*2H2O. Ещё более любопытно, что так называемые "нанобактерии", которые до того успешно имитируют метаболизм живых клеток, что почти 20 лет по поводу природы этих объектов среди учёных шли бурные споры, в итоге оказались продуктом сложного взаимодействия всё тех же кристаллов апатита и различных протеинов, в частности, производимого печенью белка фетуин-А. Не есть ли это некий намёк на то, что именно взаимодействие полифосфатов с синтезированными абиогенно короткими полипептидными цепочками и могло породить всё более усложняющийся цикл метаболических реакций, который на каком-то этапе приобрёл свойство автокаталитического самовоспроизводства?
Так что же такое полифосфаты? Это цепочки соединённых посредством атомов кислорода остатков молекул ортофосфорной кислоты различной (от двух до сотен единиц) длины. Если обозначить одну молекулу остатка ортофосфорной кислоты как P, то первичную структуру полифосфатов можно условно представить следующим образом: PPPPPPP.... Как будет описано ниже, в присутствии апатитов из простого органического вещества формальдегида (CH2O) при определённых (довольно мягких) условиях с хорошим выходом синтезируется рибоза. Это уже интересно, так как она (наровне с ортофосфоной кислотой) входит в состав нуклеотидов, кирпичиков, из которых состоят нуклеиновые кислоты. Исходя из указанного факта можно заключить, что в богатой кальцием и фосфатами среде при наличии простых органических веществ вполне вероятно присутствие ощутимого количества молекул рибозы. Соответсвенно, вполне естественно предположить, что какая-то часть из них будет встраиваться в цепочки полифосфатов, "разбавляя" их рибозой. Если обозначить молекулу рибозы как R, то получающиеся при этом молекулы можно представить примерно вот таким образом: PPPPPRPPPPRPPP..... Остались ли в современных клетках какие-нибудь "воспоминания" об этих самых ранних этапах эволюции? Пожалуй, на этот вопрос можно дать осторожный утвердительный ответ. Сейчас "с нуля" все нуклеотиды в любых клетках собираются на основе молекулы под замысловатым названием 5-фосфорибозил 1-пирофосфат (сокращённо PRPP). Как, видимо, уже догадались некоторые читатели, общепринятая сокращённая форма этой молекулы в точности соответсвует предложенным выше обозначениям (хотя стоит отметить, что дифосфат в ней прикреплён к рибозе не стандартным для полинуклеотидных молекул образом). Если пойти ещё дальше, и ввести обозначения для азотистых оснований в виде надстрочных символов (A, U, С, G для аденина, урацила, гуанина и цитозина соответсвенно), то легко заметить, что и на этом уровне по видимому видны следы цепочек полифосфатов, на базе которых могли строиться органические молекулы на начальных этапах эволюции. Скажем, основная энергетическая молекула клетки - АТФ при таком способе обозначений предстанет перед нами в виде PPPRA, а одна из основных сигнальных молекул бактериальных клеток (гуанозин 5-дифосфат 3-дифосфат) будет выглядеть так: PPRGPP. Другая важная сигнальная молекула клетки - циклический аденозинмонофосфат (изомер АМФ) в такой записи выглядит как PRA. Если к этому добавить ещё и тот факт, что сам по себе ион кальция Ca2+ тоже является важной сигнальной молекулой, а интенсивный синтез полифосфатов по сути является стандартным ответом клетки на многие стрессовые условия, сам собой напрашивается вывод, что упомянутая выше асидокальцисома является прямо-таки складом сигнальных молекул и их заготовок. Но вернёмся к нашему "алфавиту молекул жизни". Обозначив в нём буквой N дополнительное неканоническое азотистое основание, тем не менее, очень распространённое в живой природе - никотинамид, мы легко разглядим "полифосфатный скелет" и в ещё двух незаменимых для любой клетки кофакторах - НАД и НАДФ, которые в этом случае можно будет записать, соответсвенно, как RNPPRA и RNPPRAP.
Итак, есть определённые основания полагать, что первые простейшие органические молекулы были построены на различных сочетаниях таких структур как аминокислоты, азотистые основания, рибоза, ортофосфат, липиды. И действительно, именно они, в основном, входят в состав таких широко распространённых в клетке молекулярных "машинок", как коферменты. Про основной переносчик энергии в клетке, молекулу АТФ, состоящую из аденина, рибозы, и трёх остатков ортофосфорной кислоты, а так же основной кофермент окислительно-восстановительных реакций в клетке (дегидрогеназу) НАД(Ф) (аденин, никотинамид, две рибозы, два или три остатка ортофосфорной кислоты) мы выше уже писали. Важнейший кофермент SAM (S-аденозилметионин), являющийся переносчиком метильной группы для более сорока метаболических реакций в клетке, состоит из аденина, рибозы и аминокислоты метионина. Несколько более сложно, но на примерно похожих "идеях" основаны и многие другие важнейшие для жизни коферменты - витамин B12, коэнзим А, FAD. Всех их объединяет одно обстоятельство - они состоят из двух основных частей: аденозина (то есть, связки аденина и рибозы) и соединённого с ним (через остатки ортофосфорной кислоты или напрямую) собственно катализирующего необходимые реакции молекулярного блока. При этом важно отметить, что, насколько мы можем судить, аденозин сам по себе никакого участия в происходящих реакциях не принимает. Зачем же он нужен? Обычно принято считать, что он является своего рода маркером, необходимым для узнавания этих молекул другими органическими молекулами, взаимодействующими с ними. Данный сорт объяснения соответствует так называемой концепции "замороженной случайности". Несомненно, многие важные события в эволюции являются результатом фиксации случайной флуктуации, навечно "вмороженной" в механизмы жизнеобеспечения клетки. Но не мог ли уже на ранних этапах эволюции играть существенную роль и отбор? Представляется, что, по крайней мере, в случае с аденозином именно так и было. Дело в том, что аденин способен активно поглощать энергию в ультрафиолетовой части спектра. Эта энергия может быть оперативно использована для осуществления важных реакций биосинтеза. Например, как было показано ещё в середине прошлого века, упомянутая выше биомолекула НАД эффективно поглощает ультрафиолет своей адениновой частью, а полученная энергия переносится на её никотинамидную половину, где и переизлучается в результате флюоресценции. Представим, что в момент поглощения света аденином второй нуклеотид НАД был химически присоединён к какой-либо другой органической молекуле. В такой ситуации вполне можно ожидать, что вместо переизлучения энергия поглощённого кванта света могла быть использована для осуществления какой-либо полезной эндотермической реакции. Можно вспомнить и про проводимые в 60-х годах прошлого века опыты Поннамперума, в которых при облучении ультрафиолетом смеси АДФ и метил-матафосфата происходил синтез основной энергетической биомолекулы - АТФ. Известный отечественный специалист по биоэнергетике В.П.Скулачёв в своё время предложил гипотетический механизм данной реакции при котором присоединение третей фосфатной группы к АДФ происходит через промежуточный этап фосфорилирования аминогруппы аденина. Следует особо отметить, что факт первоначального использования аденина для улавливания энергии ультрафиолетового света отнюдь не отменяет его использования впоследствии в переносящих энергию молекулах как маркера для их распознавания. Скорее, ровно наоборот, он объясняет, что уже "подсевшим" на аденин метаболическим циклам было проще сохранить его присутствие (пусть и в виде некого анахронизма) в сложившейся цепи энергозависимых реакций, чем перестраивать их с нуля.
Кстати говоря, тут есть ещё одна проблема. Исходя из той универсальной роли, которую играет молекула АТФ для всех без исключения клеточных форм жизни, можно предположить, что в той среде, в которой зарождалась жизнь, аденин должен был быть представлен достаточно широко. Но его выход в опытах по лабораторной имитации абиогенного синтеза органики обычно весьма незначителен. Например, в экспериментах того же Поннамперума был зафиксирован выход аденина лишь на уровне 0.01% от соответствующего количества содержащегося в исходном растворе метана. А ведь для сборки полноценных молекул АТФ аденин нужно ещё объединить с рибозой (получив аденозин), а потом фосфорицировать. Кстати, по поводу абиогеного синтеза рибозы среди химиков тоже долгое время царил достаточно сильный скептицизм. Рибоза является типичной молекулой класса моносахаров, широкий спектр которых, как показал Бутлеров ещё во второй половине IXX века, легко синтезируются при комнатной температуре в слабощелочной водной среде из такой простой молекулы, как формальдегид в присутствии ионов кальция или магния. Более того, реакция является автокаталитической, то есть, чем больше концентрация синтезированных сахаров, тем быстрее она идёт. Некоторое число "затравочных" сахаров могло быть получено простым облучением водного раствора формальдегида ультрафиолетовым светом (похоже, этот источник энергии является критически важным для многих ключевых событий на ранних стадиях эволюции жизни). В частности, одним из основных продуктов обсуждаемой реакции, синтезируемых с неплохим выходом, является глюкоза, этот важнейший, а вполне возможно, что и вообще первый источник энергии нарождающихся живых систем. Казалось бы, ура! Но не всё оказалось так просто. Среди достаточно широкого спектра сахаров, синтезируемых в данной реакции, рибоза практически отсутствовала. А ведь именно её, в большинстве своём, и чаяли получить химики. И опять учёным неожиданно помог отказ от попыток собрать какие-то элементарные кирпичики жизни в результате ряда последовательных, логически продуманных шагов, когда вначале из продуктов A и B при определённых условиях синтезируется C, потом полученный продукт заставляют вступать в реакцию с D и так далее вплодь до полной победы коммунизма получения финального продукта. Видимо, по крайней мере в области органической химии, пора отказываться от линейного мышления, предпологающего лишь последовательные бинарные взаимодействия химических веществ. Жизнь явление синергетическое, во многих случаях надо, образно говоря, просто кинуть в горшочек сразу несколько веществ, содержащих химические элементы, которые мы ожидаем обнаружить в конечном продукте, и природа сама замутит клубок химических реакций, ведущий к желаемому результату. И действительно, если опыт удался несколько миллиардов лет назад, что мешает его повторению? Дополнительной иллюстрацией этого принципа стал результат эксперимента, когда вместо ионов кальция решили использовать в качестве катализатора уже упоминаемый выше достаточно широко встречающийся в природе минерал апатит, содержащий, в том числе, и фосфор, которого, судя по составу молекул РНК и ДНК, было в "колыбели жизни" достаточно много. Добавление апатита неожиданным образом в корне изменило состав веществ на выходе реакции. В ней, как по волшебству, стала синтезироваться почти исключительно рибоза! Кстати говоря, то, что рибоза и глюкоза в процессе происхождения жизни играли важнейшую роль, видимо, неслучайно. По сути, эти молекулы - двоюродные сёстры, отличаясь лишь размером "кольца", у рибозы в нём 5 атомов, а у глюкозы - 6 (см. Рис. 1). Попутно можно отметить, что модные сейчас фуллерены тоже состоят из образующих кольца 5-ти или 6-ти угольников, но в отличии от сахаров, в их составе нет кислорода.
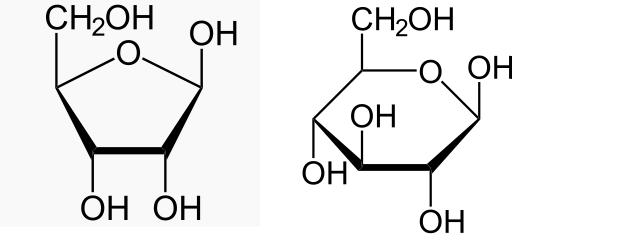
Рис. 1. Две базовые органические молекулы - рибоза (слева) и глюкоза (справа).
Тот факт, что у рибозы и глюкозы есть много общего, подтверждается ещё и тем обстоятельством, что в процессе базовой реакции гликолиза, при которой за счёт расщепления глюкозы выделяется необходимая организму энергия, является её фосфорилирование, когда к "концевому" атому кислорода в левой верхней её части, присоединяется ортофосфорная кислота, причём, в точно такой же позиции, в которой она присоединена к рибозе в молекулах РНК, ДНК, АТФ и т.д. Возможно, этого говорит о том, что на самых ранних этапах эволюции жизни функции этих молекул ещё не были чётко разнесены, и они даже могли участвовать в схожих метаболических циклах, обслуживаемых одними и теми же рибозимами, а глюкоза даже могла иногда замещать рибозу в цепочках органических молекул, которые можно условно назвать "пра-РНК"! Возможность абиогенного синтеза глюкозы подтверждается так же её обнаружением в составе вещества метеорита Мерчисон. Заодно можно упомянуть и ещё об одной химической реакции в результате которой синтезируются углеводороды, составляющие основу таких важнейших биологических молекул, как жирные кислоты. Речь о так называемом процессе Фишера-Тропша. Суть данной реакции - образование цепочек углеводородных молекул из монооксида углерода и молекулярного водорода, причём в качестве побочного продукта реакции полимеризации выделяется вода. Данная реакция лучше всего идёт при достаточно высоких (более 200 градусов Цельсия) температурах в присутствии катализаторов типа железа, никеля или кобальта, но и в упомянутых выше опытах Миллера углеводороды составляли существенную долю от всех синтезированных органических молекул.
Но вернёмся к нашим баранам, то бишь, к вопросам абиогенного синтеза нуклеотидов. Как было изложено выше, по крайней мере нуклеотиды с цитидином и уридином, несколько наиболее простых аминокислот, глюкоза (как эффективный источник энергии) и рибоза вполне могли быть синтезированы абиогенно в достаточно больших количествах при вполне реалистичных условиях. Для полного счастья нам нужно понять пути абиогенного синтеза пуриновых нуклеозидов, комплиментарных пиримидинам (цитидину и уриднину), особенно аденозина, так как факт его присутствия в большей части коферментов "голосует" за очень раннее появление данной молекулы на сцене эволюции. Пурины состоят из рибозы и, соответственно, аденина или гуанина. В принципе, оба эти нуклеиновые основания (как критически важные элементы аденозина и гуанозина) можно синтезировать в условиях умеренно восстановительной среды, примерно соответствующей по составу вышеупомянутым опытам Миллера, если в неё добавить ещё одну простую органическую молекулу - синильную кислоту (HCN). Вначале в результате её реакции с аммиаком получается цианид аммония, а потом, в результате полимеризации последнего, синтезируется некоторое количество молекул аденина и примерно на порядок меньше гуанина, см., например, здесь. Впрочем, такой ощутимый перевес в выходе аденина по сравнению с гуанином в данной реакции неудивителен, так как аденин это единственное нуклеиновое основание, которое по сути является продуктом конденсации 5 простых трёхатомных молекул (5(HCN) => аденин). Может быть ещё более интересными являются недавные опыты американских учёных показавших, что при облучении ультрафиолетом концентрированного раствора формамида в воде при температуре около 100C с хорошим выходом синтезируются как оба "классических" пуриновых основания (аденин и гуанин), так и являющийся в биологических системах их прекурсором гипоксантин. Сам формамид это достаточно простая молекула, которая может быть синтезирована, например, из таких классических для опытов по абиогенному синтезу органики веществ как монооксид углерода и аммиак. При этом, оптимальные условия для осуществления данной реакции примерно соответствуют условиям протекания реакции Фишера-Тропша (см. выше), а в качестве катализатора выступает метилат натрия.
Далее, ещё в 60-70-х годах прошлого века Поннамперума в своих опытах показал, что при облучении ультрафиолетом разбавленных водных растворов аденина, рибозы и фосфорной кислоты при температуре около 40C синтезируется, хоть и с небольшим выходом, аденозин. Как видим, и здесь казалось бы, нужный лишь на последующих этапах фосфор оказывается критически важным. Последний этап - собственно фосфорилирование нуклеозидов и получение важнейших молекул энергетического обмена клетки - АДФ и АТФ может быть осуществлён, например, если продолжать "варить" синтезированные нуклеозиды в растворе фосфорной кислоты, причём если в неё добавить в качестве катализатора этилметафосфат, то выход реакции существенно повышается. Возможен и альтернативный путь синтеза мононуклеотидов - нагревание увлажнённой смеси нуклеозидов и фосфатов натрия или кальция. Он интересен, в частности, тем, что столь эффективно повышающий в реакции Бутлерова выход из всех сахаров именно рибозы апатит является как раз типичным неорганическим фосфатом, тоже содержащим много кальция. И всё же, выход пуриновых оснований в большинстве случаев гораздо более скромный, чем в открытой англичанами реакции синтеза нуклеотидов с пиримидиновыми основаниями. Почему же в итоге из множества возможных вариантов природой были выбраны именно аденин и гуанин? Представляется, что ключевыми здесь оказались два фактора. Во-первых, как уже было отмечено ранее, аденин может служить своеобразной антенной, улавливающей энергию ультрафиолетовой части спектра. Благодаря данному обстоятельству он мог быть удобным источником энергии для предбиологических химических реакций, а так же способствовать запасению свободной энергии про запас в стимулируемых ультрафиолетом реакциях типа АДФ + АМФ => АТФ. Во-вторых, благодаря тому, что аденин комплиментарен к уридину, а гуанин к цитидину, находясь в составе РНК-цепочек они могли "налипать" соответственно на адениновые и уридиновые нуклеотиды той же или другой нуклеотидной цепочки. Данная реакция идёт с поглощением энергии, а это значит, что наличие данных пуриновых оснований в составе РНК будет способствовать её стабилизации в водном растворе и соответствующему увеличению среднего времени её жизни, что критически важно, особенно для ранних этапов эволюции. Оба эти обстоятельства могут работать в направлении своеобразной селекции именно адениновых и гуаниновых нуклеотидов и способствовать постепенному повышению их роли в процессах, происходящих в реакционной среде. И всё же, по-видимому пурины надёжно "прописались" в РНК далеко не сразу, вначале они использовались скорее "на подхвате", но об этом ниже.
Подводя промежуточный итог, можно констатировать, что в условиях умеренно восстановительной среды, в которой присутствуют достаточно простые молекулы, состоящие из наиболее распространённых во Вселенной химических элементов (фосфор в этом плане, пожалуй, является единственным исключением) при условии подвода энергии с ощутимым выходом может синтезироваться большинство критически важных для зарождения жизни органических молекул, включая несколько (до десятка) аминокислот, глюкоза, рибоза, некоторые нуклеотиды. Так же при благоприятных условиях возможно накопление в системе свободной энергии, как в виде АТФ и глюкозы, так и в виде полифосфатов (см. ниже). Таким образом, "сырьё" для дальнейшей эволюции вроде бы имеется, но что бы из указанных компонентов стали возникать и самоподдерживаться более сложные структуры типа белков и полинуклеотидов, нужны автокаталитические реакции, в результате которых самовоспроизводство указанных структур было бы поставлено на поток. Как это могло происходить на практике? Начнём с белков. Как было недавно показано, в условиях среды с повышенной концентрацией поваренной соли, даже из набора аминокислот, ограниченного лишь теми их разновидностями, которые могут быть синтезированы абиогенно в опытах, подобных опытам Миллера, с добавлением к нему лишь 2-3 дополнительных аминокислот, могут быть сформированы пептидные цепочки, способные самостоятельно сворачиваться в трёхмерные структуры, имитирующие функционирование "полноценных" белков, собранных из полного спектра в 20 аминокислот. Кстати говоря, повышенная солёность среды тоже, что называется, в тему. С одной стороны, как было показано группой, возглавляемой Е. Куниным, натриевые АТФ-синтетазы, по-видимому, предшествовали протонным, а с другой стороны, древнейшие с точки зрения автора бактерии тоже обитают в воде с очень высокой концентрацией соли. Так же следует отметить, что многие белки могут выполнять свои функции, пусть и с меньшей эффективностью, и в случае, когда они обрезаны до короткого огрызка длиной всего в несколько аминокислот. Впрочем, как известно, всё новое, это хорошо забытое старое. Лет 40-50 назад, когда умы учёных, занимающихся органической химией, ещё были почти целиком заняты поисками подтверждения теории Опарина о возникновении жизни из коацерватных капель, сформированных из прото-белков, ставилось довольно много экспериментов по абиогенному синтезу цепочек своего рода абиогенных протеинов из базовой смеси аминокислот. По видимому, наиболее известны в этом плане опыты американца Фокса и его сподвижников, когда при нагревании до температуры порядка 200 градусов Цельсия сухой смеси аминокислот в ней образовывались так называемые протеиноиды - спонтанно сформировавшиеся цепочки со случайной последовательностью аминокислот, которые, тем не менее, проявляли свойства реальных биологических ферментов. В результате многочисленных экспериментов был сделан вывод о том, что получающаяся смесь протеиноидов может катализировать (хоть и с меньшей эффективностью) большинство реакций, реально идущих в клетке. Характерно, что при нагревании ортофосфорной кислоты до температур порядка 300C тоже осуществляется полимеризация её молекул с образованием полифосфатов. Возможно, нагревание до столь высоких температур не самый благоприятный для абиогенеза сценарий, ибо та же РНК при этом очень быстро деградирует, но возникновение полинуклеотидных цепочек и прото-протеинов могло быть разнесено во времени и/или пространстве. Кроме того, как следует из некоторых опытов, спонтанная полимеризация нуклеотидов в водном растворе всё того же формамида вполне возможна и при гораздо более щадящих температурах в районе (70-90)С. В присутствии монооксида углерода и сероводорода возможен и катализируемый сульфидом железа и никеля синтез из аминокислот коротких цепочек протеноидов. Однако, в связи с тем, что никто не мог предложить достаточно убедительной гипотезы о том, как синтезированные подобным образом протеиноиды могут передавать информацию по наследству, а так же появлением гипотезы РНК-мира, вроде бы отвечающей на этот трудный вопрос, интерес к подобным опытам постепенно сошёл на нет. Представляется, что настало время вновь о них вспомнить, так сказать, на новом витке истории, тем более, что как постепенно выясняется, гипотеза РНК мира тоже имеет свои скелеты в шкафу. Самые главные из них - до сих пор не удалось синтезировать рибозим, способный копировать (полимеризировать) произвольную цепочку полинуклеотидов, с длиной, достаточной для его собственного воспроизводства, а ведь без этого наследственная передача информации невозможна! Кроме того, в последние годы накапливается всё больше доказательств, что эволюция структур типа тРНК и рибосомы не предшествовала эволюции белков, а, скорее, шла параллельно с ней!
Итак, представим, что в результате абиогенных процессов у нас образовался пул протеиноидов, проявляющих ту или иную каталитическую активность. Действительно ли у такой системы принципиально невозможно появление хотя бы зачаточной памяти и возможности передачи накопленных признаков по наследству? Что бы ответить на этот вопрос, нужно для начала отметить, что, как выяснилось в последнее время, те трёхмерные структуры, в которые сворачиваются белки, в процессе эволюции сохраняются гораздо лучше, чем их аминокислотные последовательности (одна и та же структура может быть образована огромным числом различных последовательностей). Соответствующие исследования, проведённые на эту тему, показывают, что все современные укладки белков, по-видимому, произошли из изначального множества размером от 70 до 150 базовых "суперсемейств" укладок, которые впоследствии постепенно диверсифицировались. Не могли ли данные суперсемейства укладок белков выступать на ранних этапах эволюции в качестве своеобразных элементов памяти и передачи наследственной информации? Как уже было сказано выше, в природе синтез достаточно сложных веществ всегда идёт параллельно их неизбежной деградации, так что, сохраняться могут лишь те структуры, которые способны катализировать производство собственных информационных копий (пусть даже и не точных, а приблизительных) из исходного сырья. Одним из неожиданных открытий последних десятилетий оказался как раз факт наличия подобного процесса воспроизводства даже в современной биосфере! Речь, как некоторые читатели, возможно, уже догадались, идёт о прионах. По сути, прионы, это белки, способные изменять третичную структуру других белков, заставляя их принимать такую же трёхмерную укладку, которая характерна для них самих. Эту способность можно считать простейшей реализацией процесса копирования собственной структуры (размножения). Далее, до сих пор во всех клетках экспрессируются так называемые белки теплового шока. Это очень древнее семейство белков, само название которых происходит из того факта, что данный тип белков "заставляет" только что синтезированную цепочку аминокислот сворачиваться именно так, как нужно, минимизируя вероятность их неправильной свёртки. Сама же вероятность этого неприятного для клетки события резко возрастает при повышении температуры, увеличивающей уровень тепловых шумов, которые часто приводят к ситуации, когда воображаемый диктор, комментирующий процесс свёртки очередного белка, вынужден будет сказать "что-то пошло не так...", поэтому экспрессия данных белков при повышении температуры окружающей среды, как правило, увеличивается. На начальных этапах эволюции, когда аминокислотные последовательности протеинов, по-видимому, воспроизводились гораздо менее точно, чем сейчас, данный тип белков должен был играть на эволюционной сцене очень важную роль. Таким образом, на сегодняшний день мы имеем как минимум две любопытные с эволюционной точки зрения группы белков, одна из которых перестраивает третичную структуру некоторых других белков, так сказать "под себя", а другая способна активно влиять на то, какую конформацию в итоге примет только что синтезированная (или получившаяся в результате расплавления своей третичной структуры) цепь аминокислот. По сути, даже если ограничиться только этими двумя группами, перед нами предстают довольно интересные перспективы эволюции подобной системы, элементы которой могут оказывать влияние на функционирование и взаимные превращения друг друга. Адаптация системы в ответ на изменение условий внешней среды (температуры, кислотности и т.д.), образование устойчивых цепочек взаимного катализа (как некого праобраза сложных метаболических цепочек современной клетки), кооперации различных групп протеиноидов для совместного "выживания" (разумеется, в структурно-информационном смысле) и "войны" между указанными группами за ресурсы - вот далеко не полный перечень просматривающихся возможностей. Конечно, "память" (как способность выбирать одну конкретную из нескольких достаточно глубоких потенциальных ям примерно одинаковой глубины на рельефе возможных значений свободной энергии свёрнутой макромолекулы) такой системы весьма ненадёжна и очень мала по объёму, но, тем не менее, она уже обладает рядом важных свойств, присущих памяти, основанной на кодировании информации с помощью РНК/ДНК.
Что ещё, кроме укладки, определяет каталитические свойства белков? Последовательность аминокислот в их, так называемом активном центре, то есть окружающих пространство, внутри которого собственно и происходит катализируемая данным белком реакция. Данная последовательность как бы координирует взаимное расположение реагирующих компонентов, располагая их друг относительно друга таким образом, что вероятность реакции сразу увеличивается на порядки, а то и десятки порядков величины! Довольно часто в активном центре присутствует так же ион какого-нибудь металла (белки с такими активными центрами даже получили специальное название - металлопротеины) или даже что-то вроде небольшого фрагмента кристаллической решётки, как бы "захваченной" протеином (подробнее про это см. статью LUCA Минеральные корни современных ферметов). Так же иногда в активном центре находится относительно простая органическая молекула - коэнзим. Указанные как бы внешние для молекулы белка объекты удерживаются в нужной позиции и с нужной ориентацией аминокислотными остатками, расположенными в районе активного центра. Тем не менее, строго однозначная последовательность аминокислот для этого не нужна. Иногда, как уже было упомянуто выше, важна последовательность чередования гидрофобных и гидрофильных кислот, иногда - наличие в определённых позициях лишь 3-4 ключевых аминокислотных остатков, а какие аминокислотные остатки занимают остальные позиции несущественно. Итак, ключевыми для каталитических свойств того или иного белка во многих случаях являются тип его укладки и наличие в его активном центре на нескольких ключевых позициях определённых аминокислотных остатков. Предположим, что исходно было 100 возможных базовых типов укладки белков, а активность их каталитических центров обеспечивалась в среднем 4-мя ключевыми аминокислотами на определённых позициях. Пусть в нашем условном "органическом бульоне" так же исходно присутствовало 10 синтезированных абиогенно аминокислот. В этом случае, как легко посчитать, мы получим оценку числа принципиально возможных типов синтезированных абиогенно белков в 100*104= 106. Итого, вместо будоражащих воображение гуглообразных чисел с десятками, а то и сотнями нулей, которые так любят в виде своеобразных чёток перебирать сторонники интеллектуального дизайна, перед нами предстаёт всего лишь миллион вариантов. По меркам органической химии это совершенно "детское" число. Чего же после этого удивляться, что среди протеиноидов со случайной последовательностью аминокислотных остатков заведомо находятся молекулы, способные катализировать практическую любую реакцию, важную для функционирования метаболических цепочек клетки!
Но пора вспомнить и о том, что, как и для исправного функционирования любого механизма, кроме его правильной конструкции (пусть и случайно собранной из валяющихся под ногами деталей методом проб и ошибок) и исходного сырья, нужна ещё и энергия! Ведь даже для инициализации работы двигателя внутреннего сгорания, само предназначение которого заключается в добыче свободной энергии, нужен исходный затравочный источник в виде электроэнергии, запасённой в аккумуляторе. Представляется, что на самых ранних стадиях эволюции именно описанные выше полифосфаты при гидролизе которых выделяется энергия, могли играть роль её первичного поставщика. Это тем более вероятно, что у некоторых бактерий (например, пропионовых) как для активации процесса расщепления глюкозы (гликолиза), так и для запасения выделяющей при этом энергии, вместо АТФ используется полифосфат (подробнее см. здесь). Таким образом, выходит, что при наличии всего лишь одного небольшого "затравочного" полифосфата может быть запущена автокаталитическая реакция наращивания длины полифосфатной цепочки! Так же можно отметить, что наравне с протонными и натриевыми помпами (то есть, протеиновыми комплексами, предназначенными для прокачки протонов и катионов натрия через мембрану), использующими энергию АТФ, существуют и протонные и натриевые помпы, использующие в качестве источника свободной энергии гидролиз пирофосфата (PPi). Более того, насосы именно такого типа (наравне с обычными АТФ-помпами) встроены, в частности, и в мембрану асидокальцисомы, обеспечивая внутри неё достаточно кислую среду за счёт неустанной закачки протонов из цитоплазмы внутрь органеллы. Наконец, транспортировка молекул в мембранных каналах определённого типа (состоящих, между прочим, из полимеров бетаоксимасляной кислоты и полифосфатов, соединённых всё теми же ионами кальция!), через которые внутрь клетки могут проникать ионы металлов, цепочки нуклеиновых кислот, и некоторые другие необходимые ей вещества, тоже осуществляется за счёт гидролиза полифосфатов. Несколько позже роль основного хранителя и переносчика свободной энергии могла перейти к упомянутым выше чуть более сложным молекулам PRPP. Реальность подобного сценария косвенно подтверждается тем фактом, что в современных клетках синтез нуклеотидов осуществляется на базе PRPP, причём присоединение азотистого основания к фосфорибозилу (PR) осуществляется за счёт энергии гидролиза фосфатного "хвоста" (PP), а протеин, катализирующий данную реакцию, удерживает при этом молекулу PRPP за второй её фосфатный "хвост" (P), находящийся с другой стороны. Интересно, что активный центр белка, являющийся "карманом" для удержания PRPP за фосфатную группу, состоит всего из 5 аминокислот, и представляет из себя так называемое "гнездо" , короткую группу аминокислот, окружающих анион (то есть, небольшую отрицательно заряженную молекулу) и удерживающих его за счёт взаимодействия с ним аминокислотных групп NH (см. рис. 2).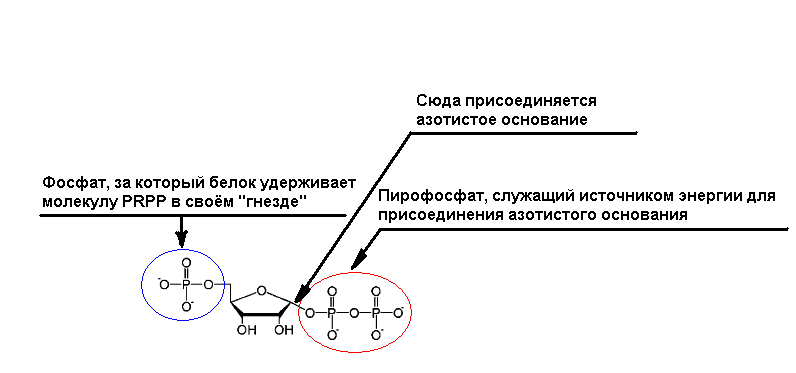
Рис. 2. Схема, поясняющая принцип синтеза нуклеотидов на базе молекулы PRPP.
Так как данная группа встречается у всех используемых в биологии аминокислот кроме пролина, можно сказать, что почти любая аминокислотная последовательность способна "пристыковаться" к фосфату, что, возможно, и явилось тем ключевым моментом, который определил выбор природы в пользу амино, а не каких-либо других органических кислот (кето и т.д.). Ведь именно взаимодействие аминокислотных цепочек и нуклеотидов лежит в самом основании всего сложного комплекса реакций, который мы именуем жизнью! В дальнейшем роль "штатного" поставщика энергии видимо перешла к молекулам АТФ. Одной из главных причин этого мог стать тот факт, что, как уже отмечалось выше, АТФ может синтезироваться из АДФ и фосфатов напрямую под действием ультрафиолетового излучения. Кроме того, при наличии соответствующих ферментов, АТФ может синтезироваться из АДФ и при расщеплении глюкозы (которая, напомним, тоже, скорее всего в достаточно больших объёмах синтезировалась в реакции Бутлерова). Таким образом, уже на ранних этапах эволюции жизни могла быть обеспечена возможность непрерывной энергетической "подпитки" предбиологических автокаталитических процессов как днём, так и ночью.
С учётом вышеизложенного, а так же наличия в некоторых коэнзимах как нуклеотидов, так и "пришитых" к ним аминокислот, или даже некого подобия коротких пептидных цепочек, представляется целесообразным рассмотреть вариант совместной параллельной эволюции структур, составленных из нуклеотидов и аминокислот. И на этом пути нас ждёт ещё много интересных открытий. Например, как было в своё время показано, особенности распределения аминокислот по кодонам генетического кода и исследование метаболических путей синтеза аминокислот в современных клетках позволяют выдвинуть достаточно обоснованную гипотезу о том, что изначально аминокислоты могли синтезироваться из кето-кислот (это близкие родственники аминокислот, у которых аминогруппа заменена кислородом) на динуклеотидах, причём каждой синтезируемой аминокислоте соответствовал свой динуклеотид, что и привело в итоге к зарождению первичного кода, вырожденного по третьему основанию. Подробнее об этом см., опять же, в соответсвующей статье LUCA. Действительно, исходное количество молекул аминокислот в среде, естественно, было ограниченным. Что бы не зависеть от "прихотей" природы, меняющихся в зависимости от изменения внешних условий, для зарождающейся биоты очень важно было взять под контроль процесс синтеза столь необходимых ей аминокислот, служащих строительным материалом для первых белков. И, похоже, именно с помощью нуклеотидов биоте удалось успешно решить эту задачу! Но, позвольте, могут спросить некоторые читатели, а откуда, собственно, взялись сами кето-кислоты, они ведь сами по себе тоже являются не такими уж простыми молекулами? Как выяснилось относительно недавно, при подходящих условиях кето-кислоты непрерывно синтезируются в так называемом восстановительном цикле трикарбоновых кислот, который, подобно циклу реакции Бутлерова является автокаталитическим. Другими словами, в процессе функционирования этого цикла концентрация молекул трикарбоновых кислот непрерывно увеличивается. При этом дополнительные кето-кислоты синтезируются за счёт энергии, поставляемой АТФ, из широко распространённого во Вселенной сырья - водорода и углекислого газа. Например, за один оборот цикла из каждой "затравочной" молекулы лимонной кислоты (цитрата) синтезируется в конечном итоге сразу две соответсвующие молекулы. Таким образом, куда ни посмотри, везде у самого основания дерева (можно образно сказать, под его корнями) биологической эволюции мы натыкаемся на автокаталитические циклы. В принципе, именно так и должно быть, ведь и сама жизнь, по существу, это, в конечном итоге, очень изощрённый процесс автокаталитической репликации.
Итак, даже синтезированные в случайном порядке цепочки аминокислот могут катализировать многие важные биологические реакции. Но дело, несомненно, пойдёт гораздо веселее, если не ждать милостей от природы, а целенаправленно влиять на процесс синтеза прото-протеинов, для начала размещая хотя бы несколько ключевых аминокислот в активном центре молекулы на тех позициях, в которых скорость катализируемой реакции ускоряется на порядки, что резко увеличит активность катализируемых процессов. Возможен ли в принципе такой промежуточный вариант синтеза белков на условной шкале, один конец которой оккупируют процессы сборки небольших белков с полностью случайными аминокислотными последовательностями, а другой - стопроцентно контролируемые рибосомой белки, содержащие сотни аминокислотных остатков? Оказывается, такой вариант есть, и, более того, он активно используется даже современными бактериями и некоторыми грибами! Речь идёт о таких пептидах, которые синтезируются вообще без участия рибосомы. Несмотря на всю разнородность данной группы аминокислотных молекулярных комплексов, у них есть некие общие черты, позволяющие предположить их очень древнее происхождение. Во-первых, все они достаточно небольшие по размерам, и содержат обычно от двух до дюжины аминокислот (например, один из самых известных нерибосомных пептидов циклоспорин состоит из 11-ти аминокислотных остатков). Во-вторых, в них очень часто присутствуют неканонические аминокислоты, отсутствующие в стандартном генетическом коде, либо аминокислоты с нестандартной (правой) хириальностью. В третьих, данные полипептидные комплексы, в отличии от белков, синтезируемых на рибосоме, часто имеют не линейную, а циклическую (как тот же циклоспорин) или разветвлённую (как цианофицин) структуру. При этом, что любопытно, во многих реакциях, имеющих важнейшее значение для жизнедеятельности клетки, например, синтезе муреиновой клеточной стенки бактерий, присоединении определённых аминокислот к липидной мамбране, непосредственном соединении соседних аминокислот в нерибосомном пептиде классической аминной связью и т.д., участвуют активированные аминокислоты, присоединённые к очень похожим на тРНК молекулам ровно таким же образом, как они присоединяются к "классическим" тРНК перед доставкой их к рибосоме. Мало того, сами реакции, в которых участвуют эти аминокислоты, обычно катализируются протеинами, имеющими большую степень сходства с "классическими" аминоцил-тРНК-синтетазами (далее АРСазами)! Подробнее про всё это "неканоническое" безобразие см., например, здесь. К ещё более интересным выводам пришли авторы другой вышедшей совсем недавно статьи. В частности, в соответствии с результатами проведённых ими исследований наиболее древней частью всех АРСаз (и других родственных им молекул) является тот участок протеина, который отвечает за присоединение аминокислоты к самой синтетазе и её так называемое активирование (для облегчения последующего присоединения аминокислоты к тРНК) с помощью ATФ. Впоследствии АРСазы постепенно "научились" специализироваться на вполне определённых аминокислотах (спектр которых шаг за шагом расширялся) и последующем присоединении их к соответствующим молекулам тРНК. При этом, важным участком для распознавания наиболее древних тРНК является участок в верхней её половине, так называемый акцепторный стебель. Судя по всему, изначально молекулы тРНК (точнее, их предки) были существенно проще, чем сейчас, и состояли лишь из одной шпильки длиной порядка дюжины нуклеотидов. Впоследствии структура молекул тРНК усложнилась, в частности, в их нижней части появилась антикодонная петля, которая нынче используется аминоцил-тРНК синтетазами как основной элемент их распознавания, а так же абсолютно необходима иРНК и рибосоме для правильной трансляции генетического кода. Один из основных выводов указанной статьи заключается в том, что дифференцация тРНК и АРСаз шла параллельно и поэтапно, причём начался этот процесс ещё до появления рибосомы на сцене эволюции. Из анализа свойств наиболее древних аминокислот, используемых АРСазами, так же вытекает, что они были преимущественно гидрофобными, что позволяет предположить их локализацию внутри мембраны, а не в цитоплазме. Кроме того, в данной статье ещё раз подтверждена родственная связь белков, катализирующих (нерибосомный) синтез ди-пептидов и АРСаз. Кстати, о дипептидах. Как рассказано вот в этой статье LUCA все АРСазы чётко делятся на два класса. Ферменты, принадлежащие к разным классам, "обвивают" тРНК по разному и прикрепляют аминокислоту к разным концам рибозы. Что ещё более интересно, как показали исследования, одну и ту же тРНК могут одновременно "обвить" (и, соответственно, прикрепить "свою" аминокислоту на "свою" гидроксильную группу рибозы), ферменты, принадлежащие к разным классам, при этом не мешая друг другу. Данный факт заставляет задуматься о возможности синтеза дипептидов на одной и той же про-тРНК у которой в концевом аденине аминокислоты присоединены одновременно сразу к двум (2-ой и 3-ей) ОH группам рибозы. Действительно, такое расположение аминокислот на соседних группах рибозы могло бы облегчить образование между ними пептидной связи примерно по такому же сценарию, как это сейчас происходит внутри рибосомы с той лишь разницей, что в реакции задействованы не 2 тРНК как при рибосомном синтезе, а одна и та же, выполняющая одновременно роль как донора там и акцептора присоединяемой аминокислоты. Если эта гипотеза верна, то можно предположить, что "отзвуки" тех далёких времён могут до сих пор частично сохраниться в аминокислотных последовательностях наиболее древних белков, ведь при вышеописанном механизме в составе синтезированного дипептида всегда должны присутствовать аминокислоты, соответствующие разным классам АРСаз, но никак не одному и тому же. Конечно, если указанные дипептиды впоследствии сшивались между собой с помощью каких-то других механизмов, стопроцентного чередования в белках аминокислот, присоединяемых АРСазами I и II классов не было даже изначально, тем не менее, статистически пары аминокислот, имеющих АРСазы разных классов должны были встречаться чаще, чем пары аминокислот, присоединяемые к тРНК АРСазами одного и того же класса. Итак, можно ли попробовать поискать следы возможного древнего синтеза дипептидов в последовательностях ДНК современных организмов? Воспользуемся для этого результатами статистического исследования встречаемости дипептидов из более, чем двух тысячах аминокислотных последовательностей доменов белков, считающихся наиболее древними. Указанные результаты приведены в таблице S3 среди дополнительных материалов к осуждаемой статье про эволюцию АРСаз. Взяв данные из этой таблицы, а так же с учётом известной принадлежности АРСаз различных аминокислот к первому и второму классам (см., например, упомянутую выше статью LUCA) мы легко можем подсчитать, что для указанных белков вероятность того, что взятые в них наугад две соседние аминокислоты принадлежат к "зоне ответственности" АРСаз разных классов равна 56.9%, то есть, несмотря на прошедшие со времён господства механизмов нерибосомного синтеза белков миллиарды лет, корреляция, судя по всему, всё ещё не выветрилась до конца!
На основании всего этого, а так же результатов своих предыдущих исследований, авторы обсуждаемой статьи выдвинули гипотезу о том, что у истоков биологической эволюции мог лежать не мир РНК, а, условно говоря, "мир протеиноидов и кофакторов". В соответствии с этой идеей исходно синтезируемые в среде абиогенно короткие (длиной не более нескольких десятков аминокислотных остатков) пептидные цепочки со случайными последовательностями аминокислот исходно взаимодействовали с небольшими органическими молекулами (предтечами современных кофакторов). Данные кофакторы (типа современных CoA, NADP и т.д.) как правило, включали один или два нуклеотида (я бы лично добавил к предбиологическим кофакторам так же ионы некоторых металлов и растворённые в среде фрагменты минералов, обладающих сильной каталитической активностью, например, железно-серные кластеры). Из-за наличия в подобной среде многочисленных положительных и отрицательных обратных связей постепенно началась эволюция пептидных цепочек и отбор тех из них, которые проявляли наиболее сильные автокаталитические свойства. Так как они, в свою очередь, очень сильно зависят от местоположения на некоторых позициях специфических аминокислот, на определённом этапе эволюции постепенно начали появляться прото-протеины, способные в статистическом плане закреплять некоторые простые аминокислотные мотивы, благоприятные для надёжной фиксации коферментов в конкретной ориентации, обеспечивающей наиболее эффективный ход катализируемых ими реакций. Благодаря всё более эффективной работе прото-протеинов, в частности, появившейся у них способности удерживать некоторые ключевые коэнзимы за фосфатный "хвост" и узнавать некоторые динуклеотидные последовательности, начиная с какого-то момента начали так же целенаправленно синтезироваться и более длинные цепочки полинуклеотидов, некоторые из которых, в частности, и стали предками будущих молекул тРНК и рРНК. Поэтапное совершенствование всей этой всё более усложняющейся системы автокаталитических циклов привело в итоге к постепенному разделению метаболических цепочек рибосомного и нерибосомного синтеза. Описанный сценарий на взгляд автора выглядит вполне правдоподобно и позволяет объяснить некоторые трудные для классической модели РНК мира странности эволюции, например, почему несмотря на то, что даже простые рибозимы in vitro (то есть, в пробирке) часто выполняют операции узнавания конкретной тРНК гораздо лучше протеинов, они для этой цели in vivo (то есть, в реальных живых системах) всё равно не используются.
От себя могу добавить, что в рамках рассматриваемого сценария можно провести следующую аналогию с развитием технологий в процессе эволюции земной цивилизации. До внедрения мануфактур, и уж, тем более, конвейеров, все технологические операции выполнялись мастерами вручную, из-за чего общая производительность труда была низкой, а экземпляры одной и той же продукции могли заметно отличаться друг от друга в зависимости от доступных мастеру материалов, особенностей его индивидуального стиля работы и т.д. По мере усиления тренда на разделение труда, автоматизацию и унификацию, товары, производимые оставшимися ремесленниками были в значительной степени вытеснены из основного товарооборота, но, тем не менее, они не исчезли полностью, а заняли определённые узкие рыночные ниши, например, в области продажи туристам своей сделанной вручную продукции в качестве сувениров. Примерно так же и после "изобретения" универсального рибосомного синтеза белков цепочки внерибосомного синтеза небольших пептидов не исчезли вовсе, а переместились в те области, в которых ограничения, накладываемые "конвейерным способам производства" оказываются непреодолимыми. Например, при синтезе антибиотков часто необходимо на определённые позиции в итоговом мини-протеине включить "правые" аминокислоты, но таких белков на "рибосомном заводе" не делают, ибо нестандарт! Тут то и приходят на помощь "дедовские" способы сборки пептидных цепочек.
В заключение данного раздела можно так же отметить, что наиболее архаичные белки, скажем, суперсемейство протеинов, характеризующихся наличием укладки Россмана (что позволяет им связывать нуклеотиды и обеспечивать тем самым своеобразный мостик между ДНК/РНК и белками), как правило характеризуются и достаточно простой структурой, заключающейся в чередованиии стандартных элементов типа альфа-спиралей и бета-листов. При этом наличие строго определённых аминокислот на конкретных позициях критически важно всего лишь для нескольких аминокислотных остатков, расположенных вблизи активного центра. Можно предположить, что именно такого рода белки, позволяющие "подносить снаряды" (в виде АТФ и полифосфатов), необходимые для активации многих важных для биологии химических реакций (включая и те, которые помогали синтезироваться им самим), и формировались в ходе самых ранних предбиологических метаболических циклов. Их простая структура являлась залогом того, что они могли автокаталитически воспроизводиться ещё до появления рибосомы.
Но как всё-таки появились первые РНК?
Итак, некоторые, хотя пока ещё и довольно смутные, но всё же проступающие всё более чётко контуры возможного механизма появления рибосомного синтеза протеинов на переднем фронте современной науки постепенно начинают просматриваться. Это весьма приятный для большинства учёных факт, ибо отсутствие реалистичных сценариев этого процесса приводило бы к необходимости использования достаточно экзотических решений типа предложенной одним из наиболее известных молекулярных биологов современности Евгением Куниным гипотезы случайной самосборки прото-рибосомы и прото РНК-репликазы в результате очень маловероятного процесса, повторение (и уж, тем более, воспроизведение в лаборатории) которого в видимой нам части Вселенной совершенно исключено. Подобные гипотезы, которые очень затруднительно, если вообще принципиально возможно, проверить экспериментально, обычно вызывают у большинства учёных чувство глубокого неудовлетворения.
Выше автор пытался убедить своих читателей, что есть надежда на то, что тайна происхождения "мира протеинов" постепенно приподнимает свою завесу. Но можно ли сказать то же самое про тайну происхождения мира РНК? Это вторая из проблем, одним из возможных решений которой Евгений Кунин считает случайную самосборку макромолекулы-репликазы. Возможно, эта проблема даже важнее проблемы происхождения синтеза белков на рибосоме, ибо связана с самой базовой чертой всех живых систем - их способностью к саморепликации на основе схем дискретного кодирования, а так же изменчивости на основе мутаций с последующим отбором наиболее приспособленных. Итак, каково же состояние современной теории происхождения механизма репликации полинуклеотидных цепочек? Для начала можно отметить, что полимеризация смеси нуклеотидов в РНК-цепочку при определённых условиях идёт сравнительно легко. Например, при активации нуклеотидов имидазолом или метилированным аденином на поверхности достаточно распространённого минерала монтмориллонита (разновидность глины) могут синтезироваться РНК-цепочки длиной до 50 нуклеотидов, что уже сравнимо с длиной простейших биологических РНК - тРНК, или, по крайней мере, их вероятных более простых предшественников. Другой путь активации нуклеотидов - присоединение остатка ортофосфорной кислоты к рибозе сразу по двум атомам углерода (2-му и 3-му) с образованием так называемого циклофосфата. Хотя при этом полимеризация идёт и менее активно, зато этот путь представляется более реалистичным, тем более, что, в описанных ранее опытах по абиогенному синтезу нуклеотидов, содержащих циотзин и урацил, они сразу синтезируются в активированной форме (с циклофосфатом), что избавляет от необходимости дополнительного выдумывания путей синтеза имидазола и активирования им ощутимой доли нуклеотидов. Кроме того, в первых небольших "прото-РНК" азотистые основания могли "насаживаться" на рибозу, которая, в свою очередь, могла "разбавлять" синтезированные абиогенно короткие цепочки полифосфатов (см. раздел "Так с чего же всё начиналось?"). В качестве косвенного подтверждения такой возможности можно упомянуть тот факт, что до сих пор при синтезе клеткой новых пиримидиновых нуклеотидов цитозин и урацил прикрепляются напрямую к рибозе, с обеих сторон соединённой с фосфатными группами (см. выше).
Следующий этап - синтез комплиментарных полинуклеотидных цепочек на абиогенно образовавшихся РНК-матрицах. В последние 10-15 лет в данной области очень активно велись исследования по получению каталитической РНК-цепочки (рибозима), способной синтезировать собственную копию. Думается, в случае успеха авторы решающего эксперимента могут смело рассчитывать на Нобелевку, ведь данная проблема по всеобщему признанию в настоящее время является центральной для теории РНК-мира. Тем не менее, несмотря на большое количество предпринятых попыток, результаты пока весьма скромны. Наилучшие из полученных к настоящему времени РНК-репликаз имеют длину около 200 но, и при этом могут воспроизвести лишь фрагмент произвольной РНК-цепочки примерно на порядок меньший собственной длины (подробнее см., например, здесь). Даже если в итоге для РНК длиной в несколько сот но и удастся достичь самовоспроизводимости, самосборка такой длинной РНК-цепочки представляется крайне маловероятной, тем более, что пока не заработал механизм репликации, эволюция в дарвиновском смысле невозможна. Кроме того, эволюция практически никогда не отказывается от уже принятых решений, а, как правило, впоследствии их лишь улучшает. По этой причине, если бы на ранних этапах эволюции репликация РНК действительно осуществлялась бы рибозимами, скорее всего какие-то варианты или хотя бы отголоски этой эпохи можно было бы обнаружить и у некоторых современных организмов. Однако, в отличии, например, от рибосомного синтеза белков, никаких содержащих РНК элементов в репликазах земных организмов не обнаружено. Есть и ещё одна трудность - все "выведенные" учёными в пробирке РНК-репликазы могут достраивать комплиментарную цепочку лишь в том случае, если уже имеется в наличии её "затравка", так называемый праймер. Строить комплиментарную цепочку "с нуля" рибозимы на сегодняшний день, насколько известно автору, не умеют.
С учётом вышеизложенного представляется вполне логичным подумать и об альтернативных сценариях возникновения репликации РНК. Достаточно большое количество исследований на эту тему было осуществлено ещё в 80-е годы прошлого века (существенная часть из них была впоследствии свёрнута, так как начиная со второй половины 90-х годов более перспективной и амбициозной стала казаться задача получения универсальной РНК-репликазы). В частности, было показано, что на цепочке РНК, содержащей в качестве нуклеинового основания цитозин, при наличии в растворе всех четырёх нуклеиновых оснований, активированных близким родственником имидазола - 2-метилимидазолом, избирательно синтезируется комплиментарная ей цепочка с основанием в виде гуанина, причём точность синтеза комплиментарной копии превышала 99%. Тем не менее, если в молекуле олигонуклеотида, служащего исходной матрицей, существенный процент составляли нуклеотиды с другими основаниями (гуанином, аденином или уридином), то синтез комплиментарной копии молекулы практически прекращался из-за эффекта "слипания" различных фрагментов молекулы, подавляющего неферментативную полимеризацию. Любопытно, что в последнее время интерес к неферментативной репликации полинуклеотидных цепочек, похоже, начал возрождаться. Например, как было продемонстрировано в недавних опытах, нуклеотиды, активированные всё тем же имидазолом, могут успешно проникать через примитивные липидные мембраны. В итоге учёным удалось продемонстрировать, что если "скармливать" созданным ими неким прототипам примитивных клеток активированные нуклеотиды монофосфата гуанозина, то они уверенно проникают сквозь её мембрану и успешно достраивают 15 недостающих позиций против соответствующих цитидинов в молекуле ДНК. И всё же, как уже отмечено выше, пока не будет предложен механизм, решающий проблему неконтролируемого слипания различных участков прото-РНК, о существенном прогрессе в вопросе возникновения механизма саморепликации РНК говорить не приходится. Кроме того, на пути более-менее реалистичной теории возникновения механизма саморепликации полинуклеотидов есть ещё ряд проблем. Во-первых, в живых организмах нуклеотиды в РНК связаны между собой строго определённым образом, всегда через 3-й и 5-й атомы рибозы. А в экспериментах по неферментативному синтезу обычно получается цепочка, в которой часть нуклеотидов связана с соседями не через 3-й, а через 2-й атом углерода. Во-вторых, после успешного синтеза комплиментарной копии для следующего этапа репликации исходную и синтезированную на ней цепочки необходимо разделить (так сказать, отделить слепок от формы). Как ни удивительно, недавно выяснилось, что первая из указанных выше проблем мало того, что не является критической для каталитической активности рибозимов, но попутно и помогает решить вторую проблему, то есть, опять мы наблюдаем ситуацию, когда эффект, на первый взгляд отдаляющий нас от цели, на самом деле помогает к ней приблизиться, хотя и несколько иным путём (подробнее об этом см. раздел "Вместо вступления" выше). Вернёмся теперь к проблеме склеивания между собой фрагментов достаточно длинных РНК-цепочек, делающей невозможной сборку комплиментарных копий РНК (достаточно эффективно синтезируются лишь G-олигомеры на C-матрицах). Можно ли как то обойти эту трудность? Один из вариантов решения данной проблемы заключается в поиске комплиментарного цитозину азотистого основания, которое могло бы на начальных этапах эволюции заменять выступающий в настоящее время в данном качестве гуанин. Удивительно, что такое основание не только есть, но, более того, оно активно используется и в современных клетках! Это входящий в состав НАД никотинамид. Кроме того, что он способен образовывать комплиментарные пары как с цитозином, но и с уридином, у него есть ещё целый букет свойств, делающих его вероятным кандидатом в качестве заменителя гуанина в "младенчестве" эволюции. Во-первых, у него нет входящего в состав всех пуриновых оснований имидазола. Это очень важно, так как именно посредством дополнительных связей через атом азота, входящего в имидазоловое кольцо, обычно и завязываются на РНК "узлы" (см. рис. 3), препятствующие формированию канонических двухцепочечных структур, что является необходимым условием успешности процесса их неферментативного копирования.

Рис. 3. Типичный "узел", который может образоваться на одноцепочечной молекуле РНК, богатой гуанином. В центре схематически обозначен ион атома магния. Взято отсюда.
Во-вторых, в отличии от гуанина, никотинамид может быть легко синтезирован из описанной выше предполагаемой смеси простых молекул (полифосфаты, рибоза, аминокислоты и т.д.), видимо, положившей начало предбиологической эволюции. Например, никотин мононуклеотид (содержащий никотинамид) неферментативно синтезируется из аспарагиновой кислоты, рибозы и вездесущего 5-фосфорибозил 1-пирофосфата (PRPP) являющегося, как мы помним, так же прекурсором и для синтеза "классических" мононуклеотидов. В третьих, при синтезе НАД, так же как и при синтезе пиримидиновых оснований, к PRPP присоединяется уже практически собранное азотистое основание, в то время как при синтезе пуриновых нуклеотидов азотистое основание поэтапно синтезируется на молекуле PRPP в результате достаточно сложного многоэтапного процесса. Данный факт может говорить о большей древности цепочки синтеза НАД и пиримидинов по сравнению пуринами. Наконец, в-четвёртых, у бактерий есть фермент, позволяющий полимеризировать молекулы никотинамид мононуклеотида в цепочку, в точности повторяющую структуру РНК, но с никатинамидом вместо "канонических" азотистых оснований (см. рис. 4.)! Приведённые выше факты, ровно как и следующий ниже рисунок, взяты из интересной работы Нади Рафаэлли "Nicotinamide Coenzyme Synthesis: A Case of Ribonucleotide Emergence or a Byproduct of the RNA World?".

Рис. 4. Синтезируемый бактериями полинуклеотид, включающий молекулы никотинамида, способные к комплиментарному спариванию с РНК, содержащей цитозин и аденин.
Но и этим способности "чудодейственного" кофактора НАД не ограничиваются. Оказывается, иногда клетка использует данную молекулу в качестве "затравки" при репликации цепочки нуклеиновых кислот, присоединяя её к первому нуклеотиду копируемой РНК. Таким образом, кроме всего прочего, наличие НАД на ранних этапах эволюции может по крайней мере частично решить отмеченную выше проблему "праймера", столь необходимого как для ферментативной, так и неферментативной репликации РНК. Ну и, наконец, вполне естественным выглядит предположение, что вдобавок ко всему прочему, миллиарды лет назад, на заре зарождения жизни, НАД, как и сегодня, мог служить эффективным агентом, обслуживающим первые окислительно-восстановительные реакции в органической среде. По видимому, на первых порах молекулы, подобно НАД бывшие "универсалами-многостаночниками", превалировали, а их диверсификация, более узкая специализация и специфическая настройка их функций произошла позже.
Итак, с учётом вышеизложенного, возможность осуществления на ранних стадиях эволюции неферментативной репликации РНК представляется вполне реальной. Тем не менее, не будем забывать, что данный процесс идёт весьма медленно, и с достаточно большим количеством ошибок. Для того, что бы не заснуть на этом этапе, эволюции нужно было рано или поздно перейти к чему-то более эффективному, то есть, без специализированных молекул-репликаз в конце концов было не обойтись. Другой вопрос, были ли эти молекулы РНК-цепочками (рибозимами) или пептидными цепочками (энзимами)? Второй вариант представляется автору более вероятным. Во-первых, как уже было упомянуто выше, ни одной РНК-репликазы в живой природе до сих пор не обнаружено, видимо, их попросту нет. В силу тенденции эволюции строить все свои последующие этапы на базе предыдущих, данный факт говорит не в пользу рибозимной гипотезы. С другой стороны, анализ существующих сегодня белковых репликаз позволяет сделать вывод, что в них критически важными для правильного функционирования активного центра фермента оказываются три находящихся там на определённых позициях остатка аспарагиновой кислоты (заодно вспоминаем, что та же аспарагиновая кислота нужна и для абиогенного синтеза НАД, а так же обоих пиримидиновых нуклеиновых оснований!), удерживающие ион магния в необходимой для катализа процесса репликации позиции. Невольно напрашивается мысль, что и небольшая по размерам пептидная цепочка, обеспечивающая аналогичное расположение остатков аспарата, тоже может (в союзе с ионом магния естественно) катализировать репликацию, пусть и с меньшей эффективностью. Так это или нет - предмет для дальнейших исследований.
Вместо заключения первой части (наброски некоторых идей относительно возможных механизмов запуска предбиогических этапов эволюции).
В данном разделе мы попытаемся связать изложенные выше фрагментированные лоскутки в некое подобие одеяла, сшивая их по возможности не белыми нитками. Основу той среды, в которой зарождалась жизнь, по видимому составляли достаточно простые вещества, состоящие из наиболее широко распространённых во Вселенной элементов, таких как H2, CO, CO2, H2O, CH4, NH3, CNH, СH2O, H2S. При условии подвода к этой среде энергии, например, в виде ультрафиолетового света или путём её нагревания, в ней синтезировались более сложные молекулы - углеводороды, жирные кислоты, кетокислоты, аминокислоты, причём при определённых условиях могла осуществляться их полимеризация. Несколько сложнее с фосфатами которые, скорее всего, тоже играли важнейшую роль уже на самых ранних этапах эволюции. Тем не менее, и здесь в последнее время достигнуты определённые успехи. Например, в проведённом недавно группой учёных из Великобритании исследовании было показано, что вещество богатых фосфором метеоритов при попадании в воду с повышенной кислотностью растворяется в ней с образованием фосфитов, которые после выпаривания воды полимеризуются в полифосфиты и, в первую очередь, в содержащие два атома фосфора пирофосфиты. Пирофосфиты уже либо могли использоваться в качестве самого первого источника энергии для ранних предбиологических органических реакций, либо путём замещения водорода на кислород быть трансформированы в пирофосфаты (авторы указанной статьи сообщают, что это было ими показано в другой серии экспериментов, результаты которых пока ещё не опубликованы). Далее, вполне естественно предположить, что в нашей гипотетической "колыбели жизни" могли присутствовать так же минералы, содержащие наиболее распространённые во Вселенной металлы, такие как кальций, магний, натрий, железо, никель. Как было показано выше, в условиях восстановительной внешней среды из формальдегида в присутствии кальция или магния с хорошим выходом синтезируется глюкоза, являющаяся с одной стороны ключевой молекулой в основной метаболической цепочке фиксации углекислого газа живыми организмами - цикле Кальвина, а с другой стороны одной из основных форм запасения клеткой свободной энергии для долговременных целей. Мало того, один из вариантов цепочки неполного окисления глюкозы приводит к образованию бетаоксимасляной кислоты, которая после полимеризации (при помощи катионов всё того же кальция) способна образовывать с полифосфатами двойные спирали, являющиеся по сути каналами, по которым через мембрану внутрь клетки могут проникать не только катионы металлов, но так же и фрагменты цепочек нуклеиновых кислот. При этом, гидрофобные участки бетаоксимасляной кислоты контактируют с имеющей к ним химическое сродство липидной мембраной, а её полярные, содержащие кислород группы, ориентированы внутрь канала и через катионы кальция связаны с несущими отрицательный заряд атомами кислорода в составе полифосфатов (см. рис. 5). Такая структура, с одной стороны, вполне естественна, так как минимизирует внутреннюю энергию комплекса, а с другой стороны имеет некое очень отдалённое структурное сходство с двойной спиралью ДНК. Не могли ли и первые короткие полимерные РНК-цепочки или их предшественники быть примерно таким же образом "прописаны" внутри первичных мембран, так сказать, "на постоянной основе"? По крайней мере, это, по-видимому, позволяло бы им сохраняться намного дольше, чем в окружении не очень дружественной для них водной среды. В любом случае, исходя из простоты устройства данных каналов логично предположить, что они были одним из первых механизмов регулируемого транспорта различных молекул сквозь примитивную мембрану.
Рис. 5. Строение простейшего мембранного канала, состоящего из полифосфатов, полимерной цепочки бетаоксимасляной кислоты и катионов кальция (символически обозначены частично закрашенными шариками). Ножницы символизируют возможное отщепление (гидролиз) концевой фосфатной группы, за счёт чего высвобождается энергия, используемая для прокачки через канал необходимых клетке химических веществ. Взято с небольшими изменениями отсюда.
Таким образом, складывается любопытная ситуация. Для синтеза одной группы важных биологических молекул (белки, жиры и т.д.) нужны восстановительные условия внешней среды, для синтеза других, обычно ассоциирующихся с концепцией РНК-мира (полифосфаты, рибоза, пиримидиновые нуклеотиды) предпочтительна нейтральная или даже умеренно подкисленная среда (дополнительные аргументы в пользу полезности повышенной кислотности внешней среды для РНК-мира см. так же здесь). При этом, большинство жизненно важных для клетки процессов сконцентрировано либо внутри, либо вблизи мембран, отделяющих среды с различной кислотностью. Не могла ли и сама жизнь зародиться на поверхности некого подобия тонкой нефтяной плёнки, разделяющей среды, различающиеся по своим химическим свойствам, в частности, кислотности? Действительно, во-первых, разность химических потенциалов в этих средах могла обеспечивать энергию, необходимую для реализации предбиологических химических реакций. Во-вторых, с одной стороны мембраны могли существовать условия, благоприятные для появления условного "мира протеинов", а условия с другой её стороны могли благоприятствовать появлению "мира полифосфатов, рибонуклеиновых кофакторов и РНК". В-третьих, расположение первых комплексов органических молекул непосредственно внутри самой мембраны и на обеих её сторонах позволяло регулировать обмен молекулами, производимыми обоими указанными выше "мирами", де факто беря под контроль химические реакции, протекающие на границе двух сред. В это связи уместно напомнить "кибернетическое" определение жизни, данное в своё время выдающимся российским математиком А.А.Ляпуновым, который предложил считать, что жизнь это "высокоустойчивое состояние вещества, использующее для выработки сохраняющих реакций информацию, кодируемую состояниями отдельных молекул". Регулируемый обмен достаточно сложными молекулами, синтезируемыми по обе стороны мембраны, мог приводить к появлению "перспективных монстров", например, в виде коферментов, имеющих как нуклеотидные, так и напоминающие ультракороткие пептиды элементы. Изначально в "кислой" половине, по-видимому, плавали лишь небольшие цепочки полифосфатов с относительно редкими вкраплениями в них рибозы и других сахаров и ещё более редкими вкраплениями пиримидиновых нуклеотидов. За счёт гидродиза таких цепочек (и, в первую очередь, простейшей из них - пирофосфата) и высвобождалась вначале энергия, необходимая для взятия под контроль первых метаболических реакций. Да и вплодь до настоящего времени этот источник энергии с успехом используется клеткой, вот, например, свежая статья про открытие молекулярных помп, способных транспортировать через мембрану протоны и катионы натрия за счёт энергии гидролиза пирофосфатов. По мере усложнения цепочек химических реакций могли начать использоваться и более сложные молекулы, включающие рибозу, типа уже упомянутой PRPP, и динуклеотиды типа НАД(Ф). Кстати, как никотинамид, имеющий всего один атом кислорода, так уж, тем более, и аденин, в котором кислород вовсе отсутствует, вряд ли могли эффективно синтезироваться в кислой среде. По этой причине гораздо более естественным выглядел бы их синтез по другую сторону мембраны в "щелочном мире". Кроме того, короткие липиды и пептиды, синтезируемые в "щелочном мире" оказались очень эффективными в качестве катализаторов, способных активно "работать" с богатыми энергией короткими цепочками полифосфатов, "разбавленных" рибозой, к которой могли дополнительно опционно крепиться пиримидиновые основания, никотинамид и аденин (см, например, выше про полипептидные цепочки, способные удерживать при себе полифосфаты и их производные за их фосфатный "хвост"). Постепенно в результате действия различных автокаталитическиих циклов появился и начал всё более совершенствоваться взаимовыгодный симбиоз, при котором в результате естественного отбора в среде начали накапливаться преимущественно те молекулярные комплексы, которые прямо или косвенно катализировали собственный синтез. Но механизмов молекулярной памяти ни в виде иРНК, ни, тем более, ДНК в то время ещё не существовало. При этом вряд ли можно относиться серьёзно к тем версиям теории РНК-мира, в которых предполагается сборка готовых функциональных блоков РНК непосредственно из активированных нуклеотидов. Исходя из предполагаемого состава "прокисшей" части первичного бульона гораздо более реалистичной выглядит исходная конденсация небольших цепочек полифосфатов с вкраплением в них рибозы и/или нуклеозидов цитидина и уридина. Указанные полифосфаты могли образовываться, например, на наночастицах кальцита или монтмориллонита. Как было показано, оба минерала обладают способностью селективно абсорбировать из раствора на разных участках кристаллической решётки органические молекулы, обладающие вполне определённой хириальностью. Кроме того, монтмориллонит способен катализировать синтез не только полинуклеотидных и полифосфатных цепочек (см., выше), но и коротких пептидов. Если ко всему вышеперечисленному добавить ещё и свойство глин выступать в качестве своеобразной губки, способной впитывать в себя большое количество жидкости (например, воды или нефти), тем самым обеспечивая растворённым в ней химическим веществам наилучшие возможности для разнообразных химических реакций, катализируемых молекулами, составляющими их кристаллическую решётку (натрий, кальций, магний и т.д.) а так же способность существенно стабилизировать РНК-цепочки не ухудшая при этом их каталитических свойств, то становится понятно, почему глинистые материалы выглядят одними из самых перспективных кандидатов на роль "колыбели жизни". Часто высказывается мнение, что поверхность минералов является идеальным местом для предбиологической химической эволюции, так как существенно облегчает как вероятность встречи реагирующих веществ (как бы перенося перебор вариантов из трёхмерного пространства на двумерную плоскость), так и катализ самой реакции. В этом плане глины обладают обоими вышеперечисленными преимуществами (так как реакции могут идти на плоском слое кристаллической решётки), но при этом сохраняя реакционноспособность во всём объёме, ибо указанными слоями заполнен весь объём минерала. Более того, как было показано, структуры типа монтмориллонита способствуют самосборке из жирных кислот пузырьков (везикул), по своей структуре родственных двухслойным липидным мембранам, а по свойствам чем-то отдалённо напоминающим коарцетивные капли в теории Опарина. В частности, они способны расти в объёме и "размножаться" делением. При этом, мельчайшие частицы глины вместе с абсорбированными на них цепочками нуклеотидов (и полифосфатов?) могут попадать внутрь везикул, и впоследствии, по крайней мере, теоретически, реплицироваться внутри них. Естественно, что при описанном сценарии упомянутая выше проблема проникновения активированных нуклеотидов сквозь липидную мембрану протоклеток попросту не стоит. Таким образом, можно констатировать, что монтмориллонит и сходные с ним глины представляют из себя не только почти идеальный химический реактор для синтеза всевозможной органики, но и способствуют появлению структур, которые могли служить прототипами будущих клеток.
Итак, берём, наконец, быка за рога и переходим к описанию собственно гипотезы. В недавно вышедшей статье наших соотечественников предложено отказаться от традиционной гипотезы зарождения жизни в водах мирового океана, и перенести "колыбель эволюции" в небольшие водоёмы, образующиеся на суше путём конденсации выходящих из под земли вулканических газов. Один из основных аргументов - состав химических элементов, из которых состоят вулканические газы, гораздо больше похож на состав химических элементов цитоплазмы, чем состав элементов морской воды. Примем эту гипотезу за отправную точку в наших рассуждениях и посмотрим, что из этого выйдет. В составе вулканических газов обычно присутствуют в ощутимых количествах в том числе такие вещества как угарный газ (CO), сероводород (H2S) и водород (H2). При температурах выше 200C и давлении, превышающем атмосферное, в условиях восстановительной атмосферы и наличии в породах, через которые проходят вулканические газы, железа, никеля и/или кобальта, складываются благоприятные условия для синтеза углеводородов по схеме реакции Фишера-Тропша (см. выше). Кроме того, факт наличия большого количества углеводородов в метеоритах типа углистых хондритов говорит о том, что на ранних стадиях эволюции битумнообразные вещества могли доставляться с метеоритами из космоса. Но вместе с газами из жерла вулкана часто выходит и лава. Каков её химический состав? Измерения, выполненные недавно геофизиками при извержении ставшего знаменитым исландского вулкана с труднопроизносимым названием Эйяфьятлайокудль показали, что около половины процента лавы составлял оксид фосфора P2O5. Это весьма ощутимая концентрация. Учитывая, что в составе вулканических газов существенную часть составляют пары воды и, кроме того, вода так же выделяется при реакции Фишера-Тропша, можно предположить, что существенная часть оксида фосфора в результате реакции с водой будет превращаться в ортофосфорную кислоту H3PO4. Кстати говоря, данная реакция идёт с существенным выделением тепла, так что, она будет одновременно способствовать как подсушиванию породы, так и повышению её температуры, что создаёт весьма благоприятные условия для синтеза многих органических молекул, в частности нуклеотидов (см. выше). В воде ортофосфорная кислота частично диссоциирует вплоть до аниона PO43-. Отрицательно заряженные анионы естественным образом будут группироваться с растворёнными в воде катионами металлов, в частности, широко представленного в выходящем на поверхность магматическом веществе кальция (Ca2+). С другой стороны, синтезированные в процессе Фишера-Тропша цепочки углеводородов и жирных кислот так же естественным образом будут формировать везикулы - крошечные пузырьки, состоящие из липидообразных молекул, причём, в качестве каталитических центров для образования подобных структур могут выступать как раз скопления фосфатов и катионов металлов, ведь именно в районах их локальной концентрации среда поляризована наиболее сильно, следовательно, с точки зрения минимизации свободной энергии, их то и нужно в первую очередь "прикрыть" сильно поляризованными "головками", из которых состоит поверхностный слой гидрофобной внутри мембраны везикул. Кроме того, в связи с повышенным скоплением ионов в данной области можно ожидать локального повышения в этих районах кислотности среды. Уважаемые читатели, вам ничего не напоминает получающаяся структура, представляющая из себя скопление фосфатов вместе с катионами металлов, в частности, кальция, окружённое липидной мембраной? Ну конечно же, как говорится, встречайте асидокальцисому, единственную клеточную органеллу, присутствующую во всех трёх надцарствах жизни! Таким образом, представляется, что в рассматриваемом сценарии вполне могла сложиться ситуация, когда внутри среды с нейтральными или умеренно щелочными свойствами могли находиться окружённые простейшими липидными мембранами вкрапления с повышенной кислотностью, содержащие достаточно концентрированный ортофосфат. Ортофосфат, как и некоторые другие небольшие полярные молекулы, мог проникать через примитивную липидную мембрану так называемым методом флип-флоп, когда они переносятся с одной её стороны на другую при спонтанном перевороте на 180 градусов самих составляющих мембрану липидных молекул (для большей ясности см. второй сверху рисунок вот в этой статье А.Маркова на Элементах). Кстати говоря, данный метод и до сих пор частенько применяется бактериями, когда им нужно перенести какую то молекулу через фосфолипидную мембрану. При нагревании ортофосфорной кислоты до 150 градусов Цельсия она превращается в пирофосфорную кислоту H4P2O7 (что можно рассматривать как первый шаг к её полимеризации до полифосфатов), а гидроортофосфат кальция CaHPO4 начинает превращаться в прирофосфат кальция Ca2P2O7 уже при температурах порядка 100 градусов Цельсия. Но, пожалуй, ещё более интересен тот факт, что некоторые бактерии в процессе гликолиза синтезируют не АТФ, а полифосфаты. А ведь глюкоза в ощутимых количествах должна была присутствовать в условном "органическом бульоне", и усвоение запасённой в ней энергии издавна считалось одним из первых возможных источников энергии для реакций синтеза органических молекул на начальных стадиях эволюции! Так же полифосфаты могли накапливаться и внутри самой прото-асидокальцисомы, тем более, что в условиях характерной для неё среды с повышенной кислотностью фосфоэфирная связь наиболее стабильна (см. здесь). Их дальнейший транспорт на наружную сторону мембраны мог осуществляться через примитивные небелковые мембранные каналы похожие на те, что изображены на Рис. 5. При этом связка полифосфата с полимером бетаоксимасляной кислоты могла служить в качестве своеобразного бесконечного штопора, один конец которого (с внутренней стороны мембраны) непрерывно удлиняется за счёт присоединения ортофосфорной кислоты, а другой (с внешней её стороны) используется в качестве источника энергии (за счёт гидролиза фосфоэфирной связи) для первых предбиологических химических реакций и активного транспорта небольших полярных молекул через мембрану. Но, что бы окончательно "встать на ноги", первым метаболическим циклам нужно было нащупать такой способ запасения свободной энергии, который бы базировался на химических реакциях, сырьём для которых служат доступные, так сказать, "в промышленных масштабах" вещества. Для поиска такого метаболического пути полезно посмотреть, какой из ныне используемых способов запасения энергии является наиболее древним. Ряд признаков, на которых мы не будем здесь более подробно останавливаться, указывает на то, что это может быть окислительно-восстановительная реакция между углекислым газом и водородом. В частности, для многих грам-положительных бактерий и архей данная реакция является основным способом запасения энергии. При этом, начальные стадии пути, связанные с формированием метильной группы CH3 у эубактерий и архей различаются, но финальная стадия, связанная с формированием молекулярного комплекса ацетил-коэнзим А, катализируется одним и тем же ферментом. Данный факт позволяет предположить, что именно эта часть метаболической цепочки является наиболее древней. В свою очередь, рассматриваемый реакционный комплекс состоит из двух кластеров, в одном из которых (CODH) осуществляется восстановление углекислого газа до монооксида углерода (CO), а во втором (ACS) полученный в результате данной реакции угарный газ вместе с метильной группой формирует ацетил, прикреплённый к одному из самых распространённых в живой природе витаминов - коэнзиму А. Но угарный газ, в принципе, достаточно часто встречается и в неживой природе, более того, это (между прочим, вместе с другом компонентом данной реакции - водородом) одна из самых распространённых молекул во Вселенной! В частности, монооксид углерода составляет ощутимую долю в газах, выделяющихся при извержениях вулканов. У ацетогенов ацетильная группа в итоге отсоединяется от коэнзима А (с запасением свободной энергии в виде АТФ), в результате чего получается ацетат CH3CO2, или, говоря по простому, уксус. Данная реакция является уникальной в том смысле, что в дополнение к запасению энергии из неорганики синтезируется органическое вещество в форме ацетата. Обычно в результате цикла органических реакций происходит либо то, либо другое. В данном же случае, по меткому выражению одного из исследователей, это примерно как "если бы Вы имели обед, за поглощение которого Вам бы ещё и приплачивали". Кстати говоря, некоторые бактерии, живущие в среде, богатой CO, не имеют первой части комплекса CODH/ACS, обходясь лишь его второй частью. Не существует ли в неживой природе некоторых аналогов реакции, осуществляемой ацетил-коэнзим А синтетазой, которые могли бы дать первоначальный толчёк эволюции данного метаболического пути? Перефразируя специалиста по революционным переворотам, можно сказать "Есть такая реакция!". Она была открыта ещё в прошлом веке, и суть её заключается в том, что из угарного газа и сероводорода (H2S) в присутствии в качестве катализаторов часто встречающихся в природе кристаллов, содержащих железо, никель и серу, и растворённого в воде небольшого количества селена при температуре около 120C синтезируется ацетат. Все "действующие лица" данной реакции, за исключением селена, достаточно широко распространены в природе и должны были присутствовать в районе действующих вулканов. Что же касается селена, то с учётом того, что содержащая его аминокислота (селеноцестеин) входит в состав белков многих организмов, можно предположить, что и он был относительно широко представлен в среде, положившей начало биологической эволюции. Можно ли использовать данную реакцию для запасения свободной энергии в полифосфатах? И здесь, видимо, можно дать положительный ответ. В качестве прекурсора при синтезе коэнзима А используется цистеамин, являющийся продуктом распада одной из стандартных аминокислот - цистеина. В 1981-м году был проведён следующий эксперимент - цистеамин, активированный с обоих его сторон ("серной" и "азотной") ацетилом (что можно, в принципе, рассматривать как некий упрощённый аналог молекулы ацетил-коэнзим А), смешанный с имидазолом и ортофосфатом был нагрет до 50C. В результате, через несколько дней в пробирке было обнаружено наличие пирофосфата (то есть, по существу дифосфата) и трифосфата, которые можно рассматривать как упрощённые аналоги соответственно АДФ и АТФ. Конечно, описанные выше факты ещё не доказывают, что именно по таким тропкам и бродила в начала своего пути эволюция, тем не менее, как мне кажется, можно констатировать, что здесь если уже и не "горячо", то, как минимум, "тепло".
На основании вышеизложенных рассуждений можно предположить существование в среде, которая положила начало эволюции жизни, полифосфатов, устойчивых к ультрафиолетовому излучению пиримидиновых нуклеотидов (монофосфат цитидин, монофосфат уридин) и некоторых других сравнительно простых органических молекул. При этом, взятие под контроль поведения обладающих существенными запасами свободной энергии полифосфатов, молекул типа PRPP, и обладающих определённой каталитической активностью коротких нуклеотидных цепочек, способных формировать простые вторичные и третичные структуры, было одним из основных факторов, способных придать какую-то направленность стихийным химическим процессам и их циклам. В решении этой задачи неоценимую помощь могли оказать короткие цепочки аминокислот - протеиноиды. Благодаря способности протеиноидов удерживать вышеперечисленные молекулы за их фосфатный "хвост", аминокислоты оказались селективно отобраны из всех многочисленных синтезируемых абиогенно органических молекул как обеспечивающие своеобразный механизм управления цепочками полифосфатов и их производными. При этом, функциональность первых протеиноидов по видимому изначально слабо зависела как от конкретной последовательности составляющих их аминокислот, так и от специфики удерживаемого ими в "гнезде" аниона. В скобках заметим, что этим, в частности, может объясняться и тот факт, что второй древнейшей метаболической цепочкой у бактерий считается сульфаторедукция. Сульфаты с химической формулой SO42- структурно похожи на фосфаты и, как правило, могут удерживаться теми же протеиноидными "гнёздами", что и они. Как бы там ни было, в итоге мог сложиться своеобразный симбиоз - протеиноиды направляли в желательном направлении синтез первичных цепочек РНК, а те, в свою очередь, помогали синтезироваться постепенно расширяющемуся набору всё более специализированных протеиноидов (см. выше про нерибосомный синтез протеинов). Реальность возможности существования сравнительно сложных форм жизни ещё до возникновения рибосомного синтеза белков косвенно подтверждается и тем фактом, что в базовый состав мембраны, отделяющей клетку от внешнего мира, "классические" протеины практически не входят, а при синтезе липидов, из которых они по большей части состоят, ключевую роль играет кофактор коэнзим А. Кстати говоря, в состав мембраны и клеточной стенки грам-положительных бактерий входят тейхоевые кислоты, по структуре иногда отдалённо напоминающие РНК у которых аналогом рибозы является глюкоза, а на месте нуклеинового основания присоединён остаток аминокислоты (см. Рис. 6) так что не исключено, что в начале и РНК могла попросту служить строительным материалом для какой-нибудь биологической структуры, и лишь позже начала использоваться в качестве катализатора некоторых химических реакций и синтеза первых "классических" протеинов. При этом изначально она могла включать лишь нуклеотиды с цитозином (под действием ультрафиолета частично превращающегося в уридин), а позже и с никотинамидом.
Рис 6. Структура одного элемента полимера глицеринтейхоевой кислоты клеточной стенки актинобактерий семейства Micrococcacrae (взято из Википедии).
Добавление никотинамида позволило РНК формировать вторичные и третичные структуры (так как никотинамид способен комплиментарно спариваться с пиримидинами, но не с самим собой) и начать целенаправленный синтез аминокислот на динуклеотидах. Правда, здесь, вроде бы, есть одна заковырка. Исходная версия гипотезы о синтезе аминокислот на динуклеотидах базировалась на предположении о "классической" четвёрке нуклеотидов. Нет ли тут противоречия? Как ни странно, по видимому, нет! Дело в том, что ключевым свойством гуанина, позволяющим ему активно участвовать в аминировании альфа-кетокислот, прикреплённых по второй позиции к рибозе, является возможность атаки его экзоциклического амина на карбонильную группу кетокислоты (см. рисунок Рис. 6 в соответствующей статье LUCA). Но ведь и никотинамид тоже имеет соответствующий экзоциклический амин примерно в том же месте своего азотистого основания (см. Рис. 4 выше)! Таким образом, отнюдь не исключено, что никотинамид мог первоначально замещать гуанин не только в качестве комплиментарного партнёра цитозина, но и в качестве катализатра при аминировании альфа-кетокислот. Добавление в химический репертуар РНК нуклеотидов с гуанином и аденином по-видимому произошло позже, именно поэтому генетический код проявляет ярко выраженную зависимость полярности кодируемых аминокислот и факта прикрепления их ко 2-му или 3-му атому углерода рибозы при аминацилировании тРНК от наличия в центре триплета цитидина и уридина, но не аденина и гуанина. Последние изначально, по-видимому, использовались только для доставки свободной энергии к реагентам химических реакций (см., выше про гипотезу Скулачёва о механизме синтеза АТФ из АДФ под действием ультрафиолета).
История человеческой цивилизации насчитывает не менее 10 тысяч лет, но лишь примерно начиная от середины этого промежутка оно стало обладать письменностью. Таким образом, многие тысячи лет культурные традиции, знания и навыки успешно передавались лишь с помощью устной речи. Появление письменности позволило, с одной стороны, зафиксировать уже накопленную информацию на более надёжном носителе, а с другой стороны, дало мощный импульс дальнейшему ускоренному развитию цивилизации. Представляется, что аналогичным образом и у биологической эволюции был достаточно длительный период, предшествовавший современной записи информации в геноме. В этот период времени применялись другие, более простые способы хранения и передачи информации, которые, тем не менее, уже позволяли поддерживать клеточные структуры на достаточно высоком уровне сложности, в частности, синтезировать полимерные молекулы и формировать из них липидную мембрану и полифосфаты, целенаправленно производить необходимые для метаболических циклов аминокислоты, воспроизводить синтезируемые нерибосомно короткие протеиноиды и кофакторы, запасать получаемую из окислительно-восстановительных реакций и облучения среды ультрафиолетовым светом свободную энергию. По-видимому, мы ещё только начинаем приоткрывать завесу над одним из самых интересных этапов эволюции жизни, её дорибосомным периодом, и на этом пути нас ещё ждёт масса интересных и неожиданных открытий.