Первые намёки на некое подобие принципа наименьшего действия можно усмотреть ещё в трудах Аристотеля, утверждавшего, что "природа ничего не делает напрасно и во всех своих проявлениях избирает кратчайший или легчайший путь". Несколько позже Птолемей несколько конкретизировал изречение известного древнегреческого философа, показав, что законы отражения света таковы, что из всех возможных путей световой луч выбирает кратчайший. Далее, во времена средневековья, философов больше интересовали религиозные дебаты, но по мере развития в Европе промышленности постепенно возродился и интерес к абстрактным наукам. В 17-м веке автор ставшей впоследствии знаменитой теоремы, получившей его имя, Пьер Ферма, подметил, что если принять в качестве критерия оптимизации не длину пройденного пути, а время, потраченное на его происхождение, то принцип наименьшего "усилия" можно обобщить и на случай распространения света в неоднородной среде (при этом принцип "наименьшего пути" оказывается частным случаем принципа "наименьшего времени", справедливым только для однородных сред). Вскоре Лейбниц ввёл понятие действия, определив его как "произведение количества материи, расстояний, на которые они передвигаются, и скорости", либо, в современных терминах, произведения импульса на пройденный путь, а Мопертюи, в свою очередь, воспользовался им, что бы провозгласить, что "путь, которого придерживается свет, является путем, для которого количество действия будет наименьшим". Данный принцип оказался на удивление универсальным, и, впоследствии, нашёл свои подтверждения практически во всех областях физики от классической механики до квантовой теории поля. В настоящее время есть много различных его формулировок, но мы в данной статье будем опираться на следующую - если известны начальное состояние системы A и её конечное состояние B, то траектория перехода системы из состояния A в состояние B будет такова, что бы минимизировать значение интеграла по времени от величины свободной энергии системы вдоль всей траектории. Рассмотрим находящуюся вдали от состояния термодинамического равновесия замкнутую систему и применим к ней указанный принцип. Исход эволюции такой системы известен, в соответствии со вторым началом термодинамики она в конце-концов неизбежно придёт к состоянию с максимальной энтропией и нулевой свободной энергией. Таким образом, величина изменения свободной энергии системы не зависит от той траектории, по которой она придёт в конечную точку своей эволюции. В соответствии с принципом наименьшего действия это означает, что, как и в случае с распространением света, система будет пытаться выбрать такую траекторию, которая бы привела к состоянию с максимальной энтропией настолько быстро, насколько это возможно (в скобках отметим, что тут есть одна тонкость, формально время достижения системой состояния с максимальной энтропией равно бесконечности, но её можно обойти, например, учитывая естественные флуктуации термодинамических параметров системы, либо выбирая конечную точку траектории системы в некоей эписилон окрестности от состояния с максимальной энтропией). При желании, можно даже рассматривать данный вывод как "усиленный" второй закон термодинамики - замкнутая система не только стремится к состоянию с максимальной энтропией, но и пытается достичь его максимально быстро. Данный вывод говорит о том, что если у системы будет возможность ускорить эволюцию к своему конечному состоянию, то она должна ею непременно воспользоваться. В этом плане сразу бросается в глаза то обстоятельство, что живые системы часто используют те же цепочки химических реакций, которые и так идут в неживой природе, но они приспособились "эксплуатировать" потоки энергии, сопровождающие указанные процессы, присваивая часть этой энергии себе. Можно даже сказать, что жизнь "паразитирует" на законах термодинамики, ускоряя эволюцию системы, и получая взамен некоторое количество свободной энергии, используя её, ровно как и информацию, для ещё большего ускорения собственной эволюции.
Мы собираемся рассмотреть это на конкретном примере, но для начала следует сказать буквально пару слов о двух в значительной степени конкурирующих между собой подходах к вопросу исследования проблемы происхождения жизни. Их можно условно кратко сформулировать как "вначале репликация" и "вначале метаболизм". Сторонники первой предполагают, что жизнь зародилась в результате случайно самособравшегося "первичного репликатора" (скорее всего, РНК-рибозима), который обладал способностью воспроизводить (иногда с ошибками) свою собственную последовательность. Другим вариантом данной гипотезы является предположение, что мог существовать некий замкнутый цикл самовоспроизводящихся репликаторов. Альтернативой данной группе гипотез является подход "вначале метаболизм" предполагающий, что всё началось с некого существовавшего ещё до появления жизни устойчивого химического цикла, на вход которого поступали простые неорганические вещества, а на выходе синтезировалась простейшая органика. Постепенное усложнение данного цикла, включение в него всё более изощрённых способов запасения энергии на промежуточных этапах и её использования для повышения его эффективности и послужило той движущей силой, которая дала старт предбиологической, а потом и биологической эволюции. При таком подходе саморепликация молекул РНК является пусть и важным, но всё же всего лишь одним из многих этапов постепенного совершенствования исходного цикла (или системы циклов). В контексте вышеизложенного появляется соблазн объяснить появление жизни именно как следствие описанного ранее "усиленного" второго закона. Действительно, в духе подхода "вначале метаболизм", и стремления природы максимально ускорить момент наступления термодинамического равновесия вполне логичным выглядит предположение, что любой химический цикл, ведущий к увеличению общей энтропии системы, должен использовать все не противоречащие законам природы средства для того, что бы самоускориться. Но ведь именно этим большинство ферментов и занимаются! Они на многие порядки ускоряют "нужные" химические реакции, например, за счёт увеличения вероятности встречи двух реагентов, которым для встречи и вступления в соответствующую реакцию в неживой природе потребовались бы многие годы, а то и миллионы лет. Недаром ведь слово "оживляться" применяют и к неживой природе в случае, когда в ней резко активизируются какие-то процессы.
Теперь попробуем прикинуть, какой из существующих ныне базовых метаболических процессов, характерных для бактерий, мог вырасти из естественного природного химического цикла? В этом вопросе я согласен с Биллом Мартином, который вот уже много лет "продвигает" на место первичного для жизни метаболического цикла так называемый восстановительный путь Вуда-Люнгдала (использующих данный путь прокариот называют так же ацетогенами). Суть цепочки реакций, составляющих данный путь, заключается в преобразовании молекулярного водорода и углекислого газа в ацетат (СH3COOH), то есть, на бытовом языке, уксус. Есть достаточно много признаков, косвенно свидетельствующих о его древности. Например, в процессе его функционирования во внешнюю среду выталкивается ион натрия, который впоследствии возвращается обратно уже с одновременным синтезом основной энергетической молекулы жизни - АТФ. Как показывают исследования, натриевая энергетика, по-видимому, очень древняя, в частности, она предшествовала протонной. Кроме того, в процессе данного метаболического пути одновременно как синтезируется органика, так и запасается энергия, что, в принципе, для живых систем большая редкость. Далее, в этой реакции вообще не используются столь вездесущие для других метаболических цепочек переносчики электронов через липидную мембрану - убихиноны, что, возможно, говорит о её архаичности. Дополнительным аргументом может являться так же очевидное сходство пути Вуда-Люнгдала с другой, по всей видимости, тоже очень древней метаболической цепочкой - метаногенезом. В частности, обе цепочки принимают на входе одни и те же вещества - CO2 и H2, обе в итоге синтезируют простейшую органику, обе в процессе работы запасают энергию в виде натриевого потенциала. Более того, многие метаногены способны и к ацетогенезу, причём в этом случае первые реакции в обоих путях совпадают, и лишь на одном из "метаболических перекрёстков" ближе концу цепочки данные пути расходятся. Но даже и на этом этапе используемые ими на последних стадиях метаболической цепочки коферменты имеют много общего. В частности, катализирующий предпоследнюю стадию пути Вуда-Люнгдала коэнзим A в общих чертах своей структуры похож на катализирующий последние шаги метаногенного цикла кофермент B (см. рис. 1).
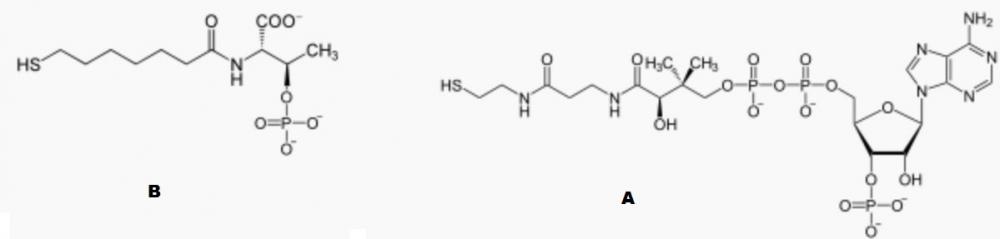
Рис. 1. А - непременный участник основного метаболического пути ацетогенов кофермент A (CoA). В - один из основных катализаторов метаболической цепочки метаногенеза кофермент B.
Действительно, как несложно заметить, с одного конца каждого кофермента мы видим атом серы, а с другого конца некую полярную "головку" включающую, в частности, фосфат. Оба конца соединены достаточно длинной "нитью" из цепочкой молекул, в которой преобладают углеводородные участки, но встречаются и другие "мотивы", в частности, CH3-CO-NH. Данный факт заставляет нас предположить, что обе молекулы могли возникнуть в результате отбора во вполне определённых (и сходных) условиях внешней среды. Что же это могли быть за условия? Для ответа на этот вопрос обратим внимание, что оба кофермента по структуре отдалённо напоминают фосфолипиды клеточной мембраны. Те тоже состоят из полярной "головы", погружённой в воду, и длинного углеводородного "хвоста", обращённого внутрь мембраны. Не могли ли и рассматриваемые коферменты в своей "прошлой жизни" располагаться на границе сред сходным образом? Предположим, что у нас есть слой воды, над которым расположен слой углеводородов, назовём его условно нефтью. Известно, что сера хорошо растворима в нефти, и обычно присутствует в ней. В таких условиях она может легко прилипать к углеводородным "хвостам" органических молекул, образуя некие аналоги тиолов, часто выступающих в органической химии в качестве катализаторов реакций, например, соединяющих молекулы в линейные цепочки. Не могли ли некоторые коферменты, ныне используемые белками в своих активных центрах, ранее располагаться на границе воды и нефти, причём их гидрофильная часть (например, остатки фосфатов) была погружена в воду, а гидрофобная, в свою очередь, в слой нефти? Такое расположение молекул, катализирующих первые органические реакции, могло давать массу преимуществ. Во-первых, все реакции происходят на границе сред, так что, поиск потенциальными участниками реакций "партнёра" по сути перемещается из трёхмерного в двумерное пространство, а это огромный выигрыш по времени потенциальной эволюции системы! Во-вторых, в определённой степени нормализуется и ориентация молекул в пространстве (более полярная часть молекулы расположена в воде, менее полярная - в нефти), а это тоже существенно уменьшает размерность пространства перебора. В-третьих, если в районе гидрофильной части молекулы находятся потенциальные участники следующего этапа необходимой реакции, то "подцепленная на крючок" с помощью атома серы молекула оказывается чисто механически связанной с гидрофильной частью кофермента, что практически гарантирует возможность их близкого сближения за относительно короткий промежуток времени. Более того, регулируя длину "удочки", ведущей к "крючку" в виде серы, можно подбирать оптимальные условия для протекания в липидном слое реакций между молекулами, сидящими на хвостах различных коферментов. Таким образом, конструкция с "плавающими" в воде, подобно лодкам, коэнзимами, высовывающими при этом в липидный слой свои "удочки" в виде оканчивающихся атомом серы хвостов, сулит массу преимуществ. Рассмотрим, например, упрощённый вариант упоминавшегося выше метаболического пути Вуда-Люнгдала в котором углекислый газ замещён угарным. Если из пород, расположенных ниже уровня воды, в результате дегазации недр планеты или серпантинизации поднимаются вверх молекулы водорода, то они должны накапливаться в вышерасположенном слое нефти, так как газы очень хорошо в ней растворяются. В свою очередь, монооксид углерода может попадать в слой нефти из лежащего выше слоя атмосферы. Если в нефти содержится так же растворённая в ней сера, то образование в таких условиях метантоиола СH3SH, с последующим присоединением к нему CO и превращением (при условии присутствия кристаллов сульфидов железа и никеля), таким образом, в тиоэфир CH3-CO-SCH3, с учётом ставших уже почти классическими опытов Вэхтерсхойзера, да и современных исследований, представляется вполне вероятным, если не сказать неизбежным. Аналогичные реакции могут происходить и в том случае, если сера находится на "хвосте" рассмотренных выше прото-коферментов. В таких условиях протекание энергетически выгодной реакции синтеза ацетата выглядит вполне логичным и тоже подтверждается экспериментально.
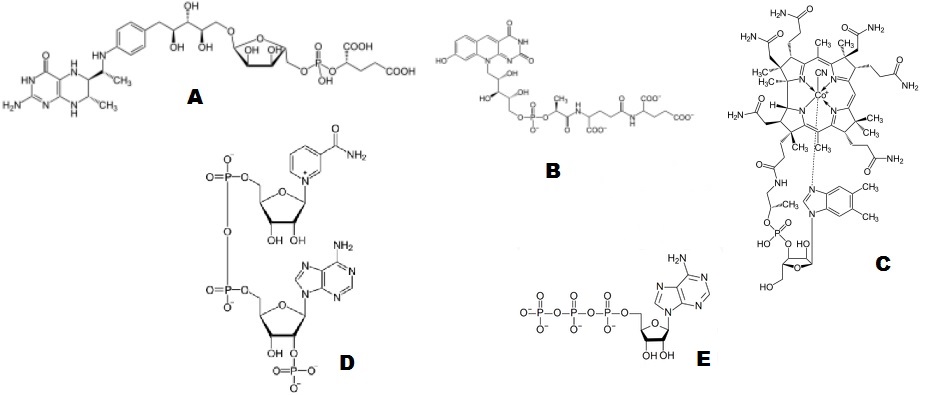
Рис. 2. A - один из основных коферментов метаногенов тетрагидрометаноптерин. B - другая важнейшая молекула, катализирующая реакции основного метабилического пути метаногенов, кофермент F-420. С - кофермент B12, катализирующий передачу метильной группы у ацетогенов. D - один из основных переносчиков протонов и электронов в окислительно-восстановительных реациях кофермент NADP. E - основная универсальная энергетическая молекула любой живой клетки АТФ.
В общем и целом можно отметить одну характерную особенность, присущую большинству наиболее архаичных коферментов - в большинстве случаев они содержат в своём составе как минимум один остаток фосфата (см. Рис. 2). Это, по-видимому, говорит о том, что его было много и в той среде, в которой зарождалась жизнь. Как правило, фосфат входит в состав коферментов вместе с рибозой. К рибозе, в свою очередь, достаточно часто крепится нуклеиновое основние, причём, не обязательно "каноническое", то есть, входящее в состав РНК и ДНК. Как же подобные структуры могли естественным образом возникнуть в неживой природе? Как уже отмечалось выше, это могло стать естественным следствием проявления универсального принципа наименьшего действия. Действительно, если не считать химически инертного гелия, угарный газ и водород являются самыми распространёнными веществами во Вселенной. При этом, химическая реакция между ними с образованием более сложных органических молекул (ацетата, метана и т.д.), идёт с выделением энергии, то есть, она термодинамически выгодна. Соответсвенно, в соответсвии с принципом наименьшего действия из пула непрерывно синтезирующихся в органическом бульоне (нефти) органических молекул будут постепенно отбираться те, которые в той или иной степени катализируют её протекание. При этом, в некоторых случаях для катализа нужна свободная энергия, с помощью которой могут инициироваться те промежуточные химические реакции, которые сами идут с поглащением энергии, но в итоге "подталкивают" систему к более эффективному протеканию основной термодиномически выгодной реакции. Временные "затраты" на точечное подталкивание процессов в системе в нужную сторону в этом случае в лихвой окупаются конечной термодиномичкской "выгодой". Ситуацию в известном смысле можно сравнить с ведением бизнеса, когда для того, что бы получить хорошую прибыль, часто приходится вначале брать кредит, локально увеличивая расходы, которые окупятся лишь после завершения всего бизнес-цикла. Роль подобного кредита в нашем случае может играть высокоэнергичная эфирная связь между фосфатом и тиоэфиром.
Итак, вернёмся к нашей модельной системе. Как мы помним, она состоит из слоя воды, над которым располагается слой нефти, снизу поступает водород, а сверху, из атмосферы, в неё поступает успешно растворяющийся в нефти угарный газ. Так как в нефти, при условии наличия внешнего источника энергии, (например, ультрафиолетового излучения), неизбежно будет синтезироваться относительно сложная органика, включая метантиол, то образование в этом случае ацетата в результате реакции водорода и угарного газа по формуле 2CO+2H2 => CH3COOH в теоретическом плане представляется неизбежным и подтверждается экспериментально (см. выше и в предыдущих статьях про опыты Вэхтерсхойзера). То есть, мы имеем как бы химический реактор, на вход которого поступает CO и Н2, а на выходе получается уксусная кислота. Как мы помним, в лежащем под нефтью слое воды, скорее всего, было достаточно много растворённых в ней молекул ортофосфата PO4, а образование ацетата может идти через промежуточную стадию синтеза тиоэфира CH3-CO-SCH3. Как резонно предполагают некоторые исследователи, в данном случае метантиол может выступать в роли примитивного аналога кофермента CoA, и поныне работающего в таком качестве у ацетогенов. При этом, на следующем шаге данного метаболического пути, у современных организмов CoA вытесняется ортофосфатом, образуя молекулу CH3-CO-PO4. Аналогичным образом, и в нашем "реакторе" матантиол, входящий в состав тиоэфира "подплывшего" к границе воды и нефти, мог вытесняться ортофосфатом. Кстати говоря, замещение CoA оротофосфатом активизируется в присутствии калия и аммония и подавляется в присутствии натрия. Этот факт позволяет предположить, что выталкивание натрия за пределы клетки (ровно как и импорт калия) изначально появилось как средство активизации процесса фосфорилирования ацетила, и лишь впоследствии созданный градиент концентрации натрия начал использоваться для запасения энергии в форме АТФ за счёт работы натриевой помпы. На последнем шаге реакции у ацетогенов остаток ортофосфата присоединяется к АДФ (заряжая её, тем самым, энергией путём превращения в АТФ), а ацетат частично выводится во внешнюю среду в качестве конечного продукта жизнедеятельности. Продолжая аналогию, в примитивном ацетогенном цикле остаток ортофосфата мог присоединяться к другой молекуле ортофосфата образуя, тем самым, пирофосфат. Кстати, некоторые бактерии до сих пор частично используют пирофосфат в качестве заменителя АТФ. Возможно, это некий атавизм, оставшийся ешё со времени первых шагов в становлении жизни. Наличие в водном растворе пирофосфата даёт шансы как бы на "второе дыхание" в эволюции системы, ибо позволяет синтезировать не только те более-менее сложные органические молекулы, реакции формирования которых термодинамически выгодны, но и молекулы, требующие для своего синтеза подвода свободной энергии. Это существенно расширяет спектр органических молекул, которые могут появляться в условном "бульоне", что, в свою очередь, открывает новые горизонты возможностей для эволюции системы. Действительно, пусть с помощью энергии пирофосфатов изредка случайно синтезируются про-коферменты, лучше, по сравнению с другими, катализирующие образование ацетата. К чему это в конечном итоге приведёт? Очевидно, к тому, что в нашей системе будет увеличиваться не только концентрация уксусной кислоты, но и пирофосфата. В свою очередь, больше пирофосфата - больше свободной энергии, а, значит, и больше вероятность синтеза самих этих про-коферментов. Налицо петля положительной обратной связи и все предпосылки для возникновения автокаталитических процессов! Если теперь несколько про-коферментов, занятых в полном цикле ацетогенеза, тем или иным способом (например, путём их соединения между собой при помощи небольших протеноидов или полифосфатов) объединятся в единые функциональные комплексы, то де-факто получится некий примитивный прототип химической системы, способной к эволюции, и обладающий многими признаками живых систем. Действительно, роль генов в этом случае будут играть сами про-коферменты, входящие в комплекс, роль мутаций берут на себя термодинамические флуктуации, регулярно их модифицирующие, а сами комплексы конкурируют между собой за максимизацию КПД превращения водорода и угарного газа в ацетат и устойчивость их автокаталитических циклов. Данная конкуренция способствует дальнейшей эволюции, допуская появление всё более изощрённых механизмов повышения скорости роста энтропии. Можно сказать, что такая система будет находиться на грани между живой и неживой природой, так как, с одной стороны, функционирует в полном соответствии с законами физики и химии, но, с другой стороны, уже обладает многими чертами, которые принято связывать с понятием жизнь.
В заключение, по традиции, ещё раз вкратце набросаем возможный сценарий первых шагов природы, которые могли привести к появлению характерных для живых систем процессов, что, возможно, даже более важно для понимания, чем сценарии появления базовых органических молекул, используемых всеми живыми организмами (чему были, в основном, посвящены первые две статьи данного цикла). Итак, в системе, состоящей из слоя воды и слоя нефти при выполнении ряда условий (наличие в ней небольшого количества железа, никеля, серы, фосфатов и т.д.) при поступлении в неё из внешней среды двух наиболее распространённых во Вселенной веществ, состоящих из более, чем одного атома - водорода и монооксида углерода, создаются благоприятные условия для её эволюции в направлении увеличения КПД реакции синтеза простейшей органической молекулы - уксусной кислоты (ацетата). Данная эволюция является следствием фундаментального физического закона - принципа наименьшего действия, предписывающего системе способствовать наиболее быстрому, из всех возможных, пути увеличения её энтропии. В данном случае высокая скорость "стока" энтропии обеспечивается путём возникновения достаточно сложных комплексов органических молекул, способных использовать энергию, выделяющуюся на промежуточных стадиях реакции, для её ускорения путём поддержания и дальнейшего усложнения собственной структуры благодаря отбору наиболее эффективных автокаталитических процессов. Данное состояние можно рассматривать как один из самых ранних этапов на пути эволюции жизни, предшествующий, в частности, гипотетическому РНК-миру и возникновению организма, являющегося общим предком всего живого на Земле - LUCA. Впоследствии данный химический цикл мог дать начало двум наиболее архаичным метаболическим путям - ацетогенезу и метаногенезу. Но, как говорится, более подробно об этом мы узнаем в следующей серии статье.