Происхождение жизни
Происхождение жизни
http://www.macroevolution.narod.ru/paleobac.htm
Александр Марков
В природе проявляется такая высокая степень разума, что вся значимость, которую люди придают своему мышлению, кажется по сравнению с ней абсолютно ничтожной.
Эйнштейн
Маленькая колба с бульоном порождает большие проблемы
Наши далекие предки и не подозревали, что вопрос о происхождении живого из неживого может стать серьезной проблемой для их потомков. Для них все вокруг было живым, одухотворенным: солнце и воздух, горы и реки, облака и море. Мудрецы Античности и Средневековья тоже не видели непроходимой грани между живым и неживым. Вслед за Аристотелем все ученые вплоть до 17 века полагали самозарождение жизни самым обычным явлением. В гниющих отходах зарождаются черви и мухи, в старом тряпье – мыши, на подводных камнях и днищах кораблей - моллюски. Могучая "животворная сила" пронизывает мироздание; она-то и заставляет косную материю порождать жизнь. Это учение – витализм – не противоречило и библейской версии космогенеза.
"И сказал Бог: да произрастит земля зелень…"
"И сказал Бог: да произведет вода пресмыкающихся…"
Бог дал стихиям творческие силы. И с тех пор – чему же удивляться – вода производила, а земля произращивала…
Первым усомнился итальянец Франческо Реди (1626 – 1698), заявивший, что, по его мнению, всякий живой организм происходит только от другого живого организма. Доказать "принцип Реди" удалось лишь два века спустя, в 1860 г, Луи Пастеру. В серии изящных опытов с хитро изогнутыми колбами он показал, что "зарождение" микроорганизмов в стерильном бульоне происходит только в том случае, если их зародыши могут попасть в бульон из воздуха или иным путем. Если преградить путь "зародышам" (оставив при этом доступ воздуху), никакого самозарождения не происходит.
Неотъемлемое свойство науки – самой себе создавать проблемы. С витализмом было покончено, но все дивиденды достались не науке, а церкви. Живое не может самозарождаться? Превосходно! Но откуда тогда вообще появилась жизнь? Одно из двух: либо она существовала изначально (эту точку зрения позже отстаивал В.И.Вернадский), либо сотворена богом. Второе, конечно, вероятнее: представить себе "изначальность" жизни (как и "вечность" Вселенной) довольно трудно. А вот теория божьего творения проста и понятна каждому.
Нелегкая задача встала перед учеными. Доказав невозможность самозарождения, они должны были теперь долго и мучительно доказывать его возможность. Пусть не здесь и сейчас, а очень давно, и не за час-другой, а за миллионы лет.
Поначалу дело казалось безнадежным, ведь химики еще не умели получать органические вещества из неорганических. Грань между живой и неживой материей казалась непреодолимой. Правда, еще 1828 г. Ф.Вёлер синтезировал мочевину, но это никого не убедило: подумаешь, мочевина! Не органика, а так, "отход жизнедеятельности". Однако в 1854 г. П.Э.М.Бертло сумел искусственно получить липиды, а в 1864 г. А.М.Бутлеров открыл реакцию синтеза углеводов из формальдегида. Впоследствии химики научились получать и многие другие органические вещества из неорганических.
Стало ясно, что между живой и косной материей на химическом уровне нет непреодолимой грани. Следовательно, хотя прямое самозарождение живых существ невозможно, жизнь могла появиться постепенно в результате долгой "химической эволюции". С тех пор (и до сего дня) усилия ученых направлены на поиски доказательств и развитие этой гипотезы. Что касается идеи об изначальности жизни, то она сейчас практически не имеет сторонников, поскольку на первых этапах развития Вселенной синтез даже самых простых органических соединений был невозможен.
Рождение Вселенной: первые подступы к жизни
В последние годы стало модным рассуждать о развитии Вселенной как о едином направленном процессе, в ходе которого самопроизвольно и неизбежно возникают все более сложные структуры. Возникло особое междисциплинарное направление – Big History, или Универсальная История (любопытно, что основоположник этого направления – профессор Дэвид Кристиан из университета Сан-Диего (США) – изначально специализировался на истории России). При взгляде на историю мироздания "с высоты птичьего полета" создается впечатление, что каждый новый шаг в эволюции Вселенной логически вытекал из предыдущего и, в свою очередь, предопределял следующий. Возникновение жизни предстает уже не случайностью, а закономерным итогом развития. Вселенная словно была изначально спроектирована так, чтобы в ней появилась жизнь, и проект был чрезвычайно точен: малейшее изменение базовых физических констант сделало бы жизнь невозможной. От религиозного толкования этого обстоятельства придумана защита – "антропный принцип". Дескать, если бы константы были иные, то в такой Вселенной некому было бы и рассуждать о мудрости ее устройства. Кто знает, может быть, существует много разных Вселенных, и только в нашей все так удачно сложилось, так что удивляться нечему.
Физики утверждают, что Вселенная появилась в результате Большого Взрыва из некого бесконечно малого объекта. В первые мгновения не было даже атомов и молекул. Потом появились элементарные частицы, из них образовались атомы водорода; скопления атомов превратились в звезды первого поколения. В них происходила реакция ядерного синтеза, в ходе которой водород превращался в гелий. Самые крупные звезды после истощения запасов водорода взрывались. При этом давление и температура в недрах звезды достигали колоссальных величин. Это создавало необходимые условия для синтеза тяжелых элементов. Все элементы тяжелее гелия, в том числе необходимые для жизни углерод, кислород, азот, фосфор, сера и другие, могли образоваться только во время таких взрывов. Звезды первого поколения стали фабрикой по производству атомов, необходимых для будущей жизни.
Взрыв звезды не только создавал тяжелые элементы, но и рассеивал их в пространстве. Из новых скоплений атомов образовались звезды второго поколения, в том числе и наше Солнце. Облака рассеяных частиц, не вошедших в состав центральной звезды, вращались вокруг нее и постепенно разделялись на отдельные сгустки – будущие планеты. Именно на этом этапе и мог начаться синтез первых органических молекул.
Возможность органического синтеза в протопланетном облаке предполагалась давно, но подтверждена была лишь недавно, во многом благодаря работам академика В.Н.Пармона и его коллег из Новосибирского Института Катализа. При помощи сложных расчетов и компьютерного моделирования было показано, что в газово-пылевых протопланетных облаках имеются необходимые условия для синтеза разнообразной органики из водорода, азота, угарного газа, цианистого водорода и других простых молекул, обычных в космосе. Непременным условием является присутствие твердых частиц-катализаторов, содержащих железо, никель и кремний.
Молодая Земля могла иметь в своем составе большое количество органики уже с самого начала своего существования. Абиогенный синтез органики продолжался уже на Земле.
Преджизнь
Вместе с Землей возник и так называемый "геохимический круговорот". Одни вещества поступали из сдавленных, разогревшихся недр Земли, формируя первичную атмосферу и океаны. Другие приходили из космоса в виде валящихся с неба остатков протопланетного облака, метеоритов и комет. В атмосфере, на поверхности суши и в водоемах все эти вещества смешивались, вступая друг с другом в химические реакции, и превращались в новые соединения, которые, в свою очередь, тоже вступали в реакции друг с другом.
Между химическими реакциями возникала своеобразная конкуренция – борьба за одни и те же субстраты (исходные вещества – "пищу" для реакций). В такой борьбе всегда побеждает та реакция, которая идет быстрее. Начинается "естественный отбор" среди химических процессов. Медленные реакции постепенно затухают и прекращаются, вытесняемые более быстрыми.
Важнейшую роль в этом соревновании играли катализаторы – вещества, ускоряющие те или иные химические превращения. Огромное преимущество должны были получать реакции, катализируемые своими собственными продуктами. Такова, например, знаменитая реакция Бутлерова, в ходе которой из формальдегида образуются сахара, которые сами и являются катализаторами этой реакции.
Следующий этап – формирование автокаталитических циклов, в ходе которых происходит не только синтез катализаторов, но и частичное возобновление расходуемых субстратов. От сложного и эффективного автокаталитического цикла уже недалеко и до настоящей жизни, ведь жизнь в основе своей – это самоподдерживающийся, автокаталитический процесс.
Проблема возникает с "затравкой": та же реакция Бутлерова не начнется, если в среде изначально не будет хотя бы небольшого количества углевода-катализатора. Несколько упрощает проблему гипотеза, предложенная А.Д.Пановым из института Ядерной Физики. Известно, что небесные тела могут обмениваться веществом: при столкновении планеты с крупным астероидом из ее поверхности выбиваются фрагменты породы, которые могут улететь в космос и попасть на другие планеты. По расчетам Панова, благодаря такому "метеоритному обмену" возникшее в ходе химической эволюции на одной из планет полезное новшество (например, эффективный катализатор) может в течение обозримого времени попасть в другие звездные системы, а за несколько сотен миллионов лет ареал распространения новшества может охватить всю галактику – разумеется, при условии, что запас данного вещества будет возобновляться в цепочке спровоцированных им химических превращений. Гипотеза Панова расширяет масштаб химической "кухни", в которой подготавливались ингредиенты будущей жизни, от планетарного до галактического.
Главная молекула жизни
Общепринятого определения жизни не существует. Одни ученые полагают, что жизнь – скорее процесс, чем структура, и определяют ее, например, как процесс сохранения неравновесного состояния органической системы извлечением энергии из среды.Такому определению могут соответствовать и системы, не имеющие четких пространственных границ – автокаталитические циклы, "живые растворы". Другие подчеркивают обязательную дискретность живых объектов и считают, что понятие "жизнь" неотделимо от понятия "организм".
Нам известна только одна жизнь – земная, и мы не знаем, какие из ее свойств являются обязательными для любой жизни вообще. Рискнем, однако, два таких свойства все-таки назвать. Это, во-первых, наличие наследственной информации, во-вторых – активное осуществление функций, направленных на самоподдержание, рост и размножение, а также на получение энергии, необходимой для выполнения всей этой работы.
Все живое на Земле справляется с этими задачами при помощи трех классов сложных органических соединений: ДНК, РНК и белков. ДНК взяла на себя первую задачу – хранение наследственной информации. Белки отвечают за вторую: они выполняют все виды активных "работ". Разделение труда у них очень строгое. Белки не хранят наследственную информацию, ДНК не совершает активной работы.
Молекулы третьего класса веществ – РНК – служат посредниками между ДНК и белками, обеспечивая считывание наследственной информации. При помощи РНК осуществляется синтез белков в соответствии с записанными в молекуле ДНК "инструкциями". Некоторые из функций, выполняемых РНК, очень похожи на функции белков (активная работа по прочтению генетического кода и синтезу белка), другие напоминают функции ДНК (хранение и передача информации). И все это РНК делает не в одиночку, а при активном содействии со стороны белков. На первый взгляд РНК кажется "третьей лишней". Нетрудно представить себе организм, в котором РНК вовсе нет, а все ее функции поделили между собой ДНК и белки. Правда, таких организмов в природе не существует.
Какая из трех молекул появилась первой? Одни ученые говорили: конечно, белки, ведь они выполняют всю работу в живой клетке, без них жизнь невозможна. Им возражали: белки не могут хранить наследственную информацию, а без этого жизнь и подавно невозможна! Значит, первой была ДНК!
Ситуация казалась неразрешимой: ДНК ни на что не годна без белков, белки – без ДНК. Получалось, что они должны были появиться вместе, одновременно, а это трудно себе представить. Про "лишнюю" РНК в этих спорах почти забыли. Ведь она, как тогда думали, не может без посторонней помощи ни хранить информацию, ни выполнять работу.
Потом, правда, выяснилось, что у многих вирусов наследственная информация хранится в виде молекул РНК, а не ДНК. Но это посчитали курьезом, исключением. Переворот произошел в 80-х годах 20 века, когда были открыты рибозимы – молекулы РНК с каталитическими свойствами. Рибозимы – это РНК, выполняющие активную работу, то есть то, что должны делать белки. Среди рибозимов были найдены и катализаторы репликации (копирования, размножения) молекул РНК – своих собственных или чужих.
В итоге РНК из "почти лишней" стала "почти главной". Оказалось, что она, и только она, может выполнять сразу обе главные жизненные задачи – и хранение информации, и активную работу. Стало ясно, что возможен полноценный живой организм, не имеющий ни белков, ни ДНК, в котором все функции выполняются только молекулами РНК. Конечно, ДНК лучше справляется с задачей хранения информации, а белки – с "работой", но это уже мелочи. РНК-организмы могли приобрести белки и ДНК позже, а поначалу обходиться без них.
Так появилась теория РНК-мира, согласно которой первые живые организмы были РНК-организмами без белков и ДНК. А первым прообразом будущего РНК-организма мог стать автокаталитический цикл, образованный самовоспроизводящимися молекулами РНК – теми самыми рибозимами, которые способны катализировать синтез собственных копий.
Теория РНК-мира, вначале чисто умозрительная, очень быстро "обрастает" экспериментальными данными. Химики научились получать рибозимы чуть ли не с любыми желаемыми характеристиками (для этого синтезируют огромное количество разных РНК со случайной последовательностью нуклеотидов, а затем просто отбирают из них молекулы с нужными свойствами). Получены рибозимы, катализирующие синтез нуклеотидов, присоединение аминокислот к РНК и другие биохимические процессы. Стирая грань между живым и неживым, уже растут на искусственных средах в лабораториях возмутительнейшие объекты – колонии размножающихся молекул РНК, способные к тому же синтезировать белки. Весомый вклад в эти исследования вносят ученые из Института химической биологии и фундаментальной медицины (г. Новосибирск) и Института белка (г. Пущино) под руководством академиков В.В.Власова и А.С.Спирина.
РНК – полимер, состоящий из множества похожих "кирпичиков" – рибонуклеотидов, каждый из которых, в свою очередь, собран из трех частей. Первая из них - фосфорная кислота (фосфат) - неорганическое вещество, которого довольно много в земной коре и океанах. Вторая – азотистое основание. В состав РНК входит четыре азотистых основания: аденин, урацил, гуанин и цитозин; соответственно, существует четыре вида рибонуклеотидов. Азотистые основания могли синтезироваться из неорганических молекул (таких, как CO, HCN и NH3) еще в протопланетном облаке. Их находят и в метеоритах. Третья - сахар рибоза – образуется в ходе автокаталитической реакции Бутлерова.
Как РНК решала энергетический вопрос
Чтобы два рибонуклеотида соединились вместе, к одному из них должен быть присоединен дополнительный фосфат (или сразу два). Получившаяся молекула - рибонуклеотид с лишним фосфатом - содержит в себе большое количество энергии. Эта энергия, при наличии подходящих катализаторов, может быть использована для выполнения разных полезных "работ". В том числе для соединения двух рибонуклеотидов в одну молекулу – маленькую РНК. Рибонуклеотиды с дополнительными фосфатами первоначально использовались, скорее всего, только как "строительные кирпичики" при синтезе РНК. Кирпичики, надо сказать, очень удобные – ведь они включают в себя не только строительный материал, но еще и энергию, необходимую для выполнения строительных работ! Впоследствии они стали использоваться для тысяч других важных дел – везде, где для выполнения какой-то работы требуется энергия. Все живое и по сей день пользуется фосфорилированными рибонуклеотидами как универсальными поставщиками энергии при выполнении энергоемких задач. Самая известная из этих "энергетических" молекул – АТФ (аденозинтрифосфат). Это обычный рибонуклеотид, к которому присоединены два дополнительных фосфата. АТФ – одновременно и источник энергии для множества энергоемких реакций, и один из кирпичиков для синтеза РНК. Так земная преджизнь нашла универсальное решение сразу двух задач: запасания энергии в удобной форме и синтеза РНК – главных молекул жизни. Между прочим, другие ключевые "энергетические" молекулы живой клетки – НАД, НАДФ и ФАД – представляют собой пары сцепленных рибонуклеотидов (одного стандартного и одного "неклассического"). Это еще одно наследие РНК-мира.
Первые организмы
Все живые организмы дискретны в пространстве и имеют наружную оболочку. Трудно представить себе живое существо в виде туманного облачка или раствора. Однако поначалу преджизнь существовала именно в виде растворов. Чтобы не раствориться окончательно, не рассеяться в водах древних водоемов, "живые растворы" должны были ютиться в крошечных полостях, которые часто встречаются в минералах. Это тем более удобно, что некоторые минералы (например, пирит) являются неплохими катализаторами для многих биохимических реакций. Кроме того, поверхность минералов могла служить своеобразной матрицей, основой, к которой прикреплялись молекулы РНК. Упорядоченная структура кристаллов помогала упорядочить и структуру этих молекул, придать им нужную пространственную конфигурацию.
Но рано или поздно преджизнь должна была обзавестись собственными оболочками – перейти от доорганизменного уровня к организменному. Идеальным материалом для таких оболочек являются липиды, молекулы которых способны образовывать на поверхности воды тончайшие пленки. Если взболтать такую воду, в ее толще образуется множество мелких пузырьков – водяных капелек, покрытых двуслойной липидной оболочкой (мембраной). Эти капельки проявляют интересные свойства, которые делают их похожими на живые клетки. Например, они способны осуществлять обмен веществ. Липидные мембраны обладают избирательной проницаемостью: одни молекулы сквозь них проходят, другие нет. Благодаря этому одни вещества втягиваются в каплю, другие выводятся, третьи – накапливаются внутри. Правда, для того, чтобы это происходило постоянно, одних мембран недостаточно. Нужно еще, чтобы внутри капли одни вещества превращались в другие, а для этого там должны находиться катализаторы – белки или РНК.
Изучением свойств водно-липидных капель (коацерватов) занимался академик А.И.Опарин. Он считал, что коацерваты были одним из этапов на пути возникновения жизни. Опарин обнаружил, что при определенных условиях коацерваты могут расти и даже "размножаться" делением.
Первые коацерваты могли образоваться самопроизвольно из липидов, синтезированных абиогенным путем. Впоследствии они могли вступить в симбиоз (взаимовыгодное сожительство) с "живыми растворами" – колониями самовоспроизводящихся молекул РНК, среди которых были и рибозимы, катализирующие синтез липидов. Подобное сообщество уже можно назвать организмом. У всех живых существ до сих пор в синтезе липидов важнейшую роль играет кофермент А, представляющий собой не что иное, как модифицированный рибонуклеотид. Это – еще одно напоминание об РНК-мире.
Камнем преткновения для теории РНК-мира в течении некоторого времени была неспособность молекул РНК эффективно взаимодействовать с липидными мембранами. Недавно, однако, было показано, что комплексы из нескольких разных молекул РНК и ионов кальция способны не только прикрепляться к мембранам, но и регулировать их проницаемость.
РНК обзаводится помощниками
В дальнейшем РНК-организмы приобрели два важных усовершенствования. Во-первых, они научились катализировать синтез аминокислотных полимеров – сначала коротких пептидов, а затем и длинных белков. Эти вещества стали для РНК-организмов универсальными помощниками, справляющимися с большинством биологических "работ" гораздо лучше, чем рибозимы. Симбиоз РНК и пептидов, вероятно, начал складываться задолго до появления настоящей клетки и генетического кода. Напоминают об этом этапе недавно открытые комплексы из небольших молекул РНК и пептидов, выполняющие множество регуляторных функций, а также строение некоторых важных молекул, таких как кофермент А – рибонуклеотид с прикрепленным к нему пептидоподобным "хвостом".
Поначалу РНК-катализ белкового синтеза, скорее всего, не был строго специфичным: последовательности аминокислот из раза в раз воспроизводились не точно, а лишь приблизительно. Поскольку точность в данном случае резко повышала стабильность живой системы, естественный отбор способствовал выработке все более специфичных каталитических систем. Дело кончилось возникновением универсальной системы специфичного синтеза любого требуемого пептида. Это и был генетический код вкупе с комплексом рибозимов, необходимых для его прочтения.
Для синтеза белков все живые организмы по сей день пользуются специальными молекулярными "машинками" – рибосомами, основу которых составляют молекулы РНК. Правда, белки тоже входят в состав рибосом. И белки непростые – маленькие, очень древние, крайне консервативные (в геномах древнейших живых организмов – бактерий и архей - гены рибосомных белков по не вполне ясным причинам обычно располагаются рядом, все вместе, в почти одинаковом порядке, образуя так называемый "рибосомный супероперон". Это резко контрастирует с поведением других бактериальных генов, которые могут гулять по хромосоме, как им вздумается). Однако удалось показать, что рибосомные РНК могут синтезировать белок и сами, без помощников – медленно, с трудом, но все-таки могут.
Кроме рибосом, для синтеза белка необходимы особые молекулы–посредники, которые "читают" инструкции, записанные в информационной молекуле РНК, и в соответствии с этими инструкциями присоединяют к синтезируемой молекуле белка нужные аминокислоты. В роли этих посредников выступают опять-таки молекулы РНК (транспортные РНК, или тРНК). Как видим, "главная молекула жизни" так и не доверила синтез белков самим белкам и продолжает, как встарь, выполнять его почти самостоятельно.
Вторым крупным усовершенствованием РНК-организмов было приобретение ДНК. Молекулы ДНК более устойчивы, чем РНК, и потому являются более надежными хранителями наследственной информации. Платой за стабильность стала неспособность молекул ДНК сворачиваться в глобулы и выполнять какие-либо активные действия. Изначально ДНК, скорее всего, была чем-то вроде покоящейся фазы в жизненном цикле самовоспроизводящихся колоний РНК, и лишь много позднее она стала основным носителем наследственной информации.
ДНК и РНК – хранители наследственной информации
ДНК, как и РНК, формируются из кирпичиков – нуклеотидов, только чуть-чуть других. Чтобы из РНК-кирпичика (рибонуклеотида) сделать ДНК-кирпичик (дезоксирибонуклеотид) достаточно одной простой реакции. Молекулы ДНК (как и РНК) способны к самокопированию, правда, для этого нужны катализаторы – белки или рибозимы. Наследственная информация, хранящаяся в ДНК в виде последовательности нуклеотидов, может "переписываться" на РНК (так создаются информационные РНК) и обратно. Точность копирования обеспечивается в значительной мере автоматически, благодаря особому свойству нуклеотидов: против каждого нуклеотида исходной молекулы (матрицы) в синтезируемой копии (реплике) может встать только один строго определенный нуклеотид из четырех возможных (например, напротив гуанина – только цитозин). Когда на этой реплике синтезируется новая реплика, она окажется точной копией исходной молекулы.
В начале было сообщество
Многие биологи полагают, что все разнообразие жизни на нашей планете происходит от единственного исходного вида – "универсального предка". Другие, в том числе крупнейший микробиолог академик Г.А.Заварзин, несогласны с этим. Устойчивое существование биосферы возможно только при условии относительной замкнутости биогеохимических циклов. В противном случае живые существа очень быстро израсходуют все ресурсы или отравят себя продуктами собственной жизнедеятельности.
Замкнутость циклов может быть обеспечена только сообществом из нескольких разных видов микроорганизмов, разделивших между собой биогеохимические функции (примером такого сообщества являются циано-бактериальные маты). Заварзин считает, что организм, способный в одиночку замкнуть круговорот, столь же невозможен, как и вечный двигатель.
Для этапа химической преджизни это еще более очевидно. Никакая отдельно взятая органическая молекула не сможет устойчиво самовоспроизводиться и поддерживать гомеостаз в окружающей среде. На это способны только комплексы из довольно большого числа разных молекул, поделивших между собой функции.
Скорее всего, общим предком всего живого был не один вид, а полиморфное сообщество, в котором происходил активный обмен наследственным материалом между организмами. Разнообразие, симбиоз, разделение функций, информационный обмен – изначальные свойства земной жизни.
Следы древней жизни
Время появления жизни на Земле точно не известно. Ясно одно: если наша планета когда-то и была безжизненной, то не очень долго. Земля сформировалась 4,5 – 4,6 млрд лет назад, но от первых 700-800 млн лет ее существования в земной коре практически не осталось следов. Ископаемые организмы встречаются в основном в осадочных породах, но самые древние из известных осадочных пород (формация Ишуа в Гренландии) имеют возраст около 3,8 млрд лет. И в них уже есть следы жизни. Правда, не совсем понятно, какой – РНК-жизни или уже современной, ДНК-белковой. Эти следы - чисто химические, связанные с изотопным составом углерода. Чуть позже (3,5 млрд лет назад) начинают встречаться остатки целых живых организмов – бактерий. Таким образом, жизнь появилась на Земле не позже, чем 3,8 млрд лет назад. Эпоху РНК-мира некоторые специалисты помещают где-то между 4,3 и 3,8 млрд лет назад.
Обсуждение этой статьи и проблемы происхождения жизни в целом см. здесь.
Бактерии в метеоритах
Елена Наймарк
Двенадцатикилограммовый метеорит Оргэй (Orgueil) упал в мае 1864 г. близ одноименной французской деревни, расколовшись на 20 кусочков. Вещество метеорита можно было спокойно разрезать ножом, поточить как карандаш и рисовать: это был обычный углистый метеорит с очень высоким (почти как в грифеле карандаша) содержанием углерода. Обычный, да не совсем, потому что именно с этим метеоритом связаны наиболее яростные споры – "есть ли жизнь на Марсе – нет ли жизни на Марсе"…
Химический анализ показал, что метеорит состоит из горючего углеродистого материала, включающего соли аммония, магнетит и кремниевую кислоту. В целом минеральный состав метеорита напоминал глины и вулканический пепел, но в земном пепле гораздо меньше магния и больше кремния, алюминия и кальция. Содержание углерода было таким высоким, что химики заподозрили биологическое происхождение метеоритного материала. В абиогенных земных породах такого количества углерода не бывает. Если метеорит образовался при участии микроорганизмов, то и земная жизнь вполне могла быть порождением космических сил.
В то время, в 1864 г., споры о происхождении жизни были в самом разгаре. Несколькими годами раньше Луи Пастер доказал невозможность самозарождения живых существ. Четкая логика опытов и авторитет Пастера (который сам был убежденным креационистом) убедили многих ученых в божественном – и, конечно, земном, а не космическом – происхождении жизни.
Поэтому когда Пастеру принесли для изучения кусочек метеорита Оргэй, он был заранее уверен в отрицательном результате. Тем не менее он тщательно подготовил опыты: стерильными инструментами выкрошил из середины обломка метеоритный материал, посадил на питательную среду и попробовал вырастить космических микробов. Естественно, ничего не выросло. Заключение, сделанное великим ученым, гласило, что ничего живого в метеорите Оргэй нет, так что вся ответственность за жизнь на Земле лежит на Всевышнем.
Однако идея космического происхождения жизни не погибла. В бестселлере Н.К. Фламмариона «Популярная астрономия», вышедшем в 1864 году, был даже специальный раздел, посвященный внеземной жизни, а несколько лет спустя У. Томпсон сформулировал теорию панспермии ("космических семян" жизни).
Спустя столетие изучение метеорита Оргэй возобновилось. К этому времени в руках землян оказалось немало и других метеоритов с высоким содержанием углерода. В 1961 г. американские ученые объявили о новых результатах анализа старого знакомца Оргэй и других углистых метеоритов (Ивуна, Тонк, Алаис). В метеоритном веществе обнаружилось множество разнообразных органических соединений: алифатические и ароматические углеводороды, аминокислоты, жирные кислоты и липиды, азотистые основания обоих основных типов (пуриновые и пиримидиновые), порфирины и другие.
Многие из этих веществ заметно отличались от земных. Так, аминокислоты были преимущественно низкомолекулярными, некоторые из них не имели земных аналогов. Состав порфиринов тоже свидетельствовал о неземном происхождении метеоритной органики.
Но еще важнее было заключение, что набор метеоритных органических молекул вполне мог складывать некие живые существа. Остатки этих существ были также представлены учеными. Найденные в метеоритах Оргэй и Ивуна образования по форме и размерам напоминали земные микроорганизмы. Это были трубочки с пережимами, похожие на цепочки клеток, грибовидные тела, сферы с выростами, обрывки оболочек…
Ученые все же не решились объявить эти структуры клетками, дав им осторожное название «организованные элементы». В 60-е годы 20 века в научных журналах то и дело появлялись сообщения о находках новых «организованных элементов». Некоторые специалисты занялись даже их систематикой. А журнал «Наука и Жизнь» в 1964 году опубликовал большую статью о жизни на Марсе, в которой всерьез обсуждался вопрос о климатических природных поясах Марса, выявленных с помощью сезонных спектральных наблюдений, и о видах водорослей, которые населяют эти природные зоны. Иными словами, после обнаружения "организованных элементов" и химических свидетельств внеземного происхождения метеоритной органики смелая мысль уносила ученых далеко за пределы строгой науки. Наука же требовала методической точности и разнообразия гипотез.
Скептиков, в числе которых был и Нобелевский лауреат Х. Урей, не убедили приведенные доводы. Они продолжали утверждать, что метеоритная органика – не более чем загрязнения, набранные космическим объектом на пути от верхних слоев атмосферы до научной лаборатории. И вправду, обнаружилось, что оптический состав аминокислот напоминает земной – преобладают левозакрученные молекулы, а большая часть органики приурочена к поверхностным слоям метеоритов.
"Организованные элементы" тоже не произвели большого впечатления на скептически настроенных ученых. Обнаружилось, что неорганические кристаллы могут вполне успешно маскироваться под микроорганизмы, будь то кокки, бациллы или более сложные формы. В итоге в 70-е годы панспермия полностью вышла из моды, а термин "астробиология" стал почти ругательным.
Однако уже в следующем десятилетии, в конце 80-х, изучение углистых метеоритов возобновилось. К этому времени появился не только новый материал для исследования, но и мощные электронные микроскопы нового поколения, совмещенные с химическим анализаторами. Это позволило найти огромное количество разнообразных "организованных элементов". Активное участие в исследованиях принимают российские ученые под руководством члена-корреспондента РАН, палеонтолога А.Ю.Розанова.
В метеоритах Мурчисон и Ефремовка найден целый комплекс удивительных образований: различные нити с пережимами, сферы, ленты, ажурные плетения. Микробиологи легко подбирают аналогичные им по форме земные бактерии, современные и ископаемые. Все эти образования микронных размеров, как и большинство бактерий. Образования нанометровых размерностей, не имеющие известных земных аналогов, обнаружены в метеорите Мигей. Это четко ограниченные войлокоподобные слои нитей, в которых запутаны отдельные глобулы, соединенные иногда в цепочки. Сейчас стали известны и земные нанобактерии, но наши знания об их разнообразии еще слишком фрагментарны, так что об их сравнении с метеоритными говорить рано.
В современных лабораториях легко отличить земные загрязнения от метиоритного материала. Химические микроанализаторы мгновенно предоставляют отчет об элементном составе подозрительного объекта. Если состав отличается от матрикса метеорита, это загрязнение. Если нет – приходится признать в подозреваемом подлинного пришельца из космоса. Гораздо труднее отличить фоссилизированные остатки микроорганизмов от схожих образований неорганического происхождения. Приходится признать, что наши знания о микрокристаллах и тем более нанокристаллах пока недостаточны. Поэтому вопрос о бактериях в метеоритах остается открытым. Есть, однако, немало оптимистов (как среди российских палеонтологов, так и среди американских сотрудников NASA), которые уже сейчас убеждены в биологической природе "организованных элементов" и даже пытаются по этим остаткам реконструировать внеземные микробные сообщества. По их мнению, эти сообщества были подобны земным циано-бактериальным матам, а развиваться они могли на различных небесных телах: Марсе, гипотетическом Фаэтоне и даже в ядрах комет.
|
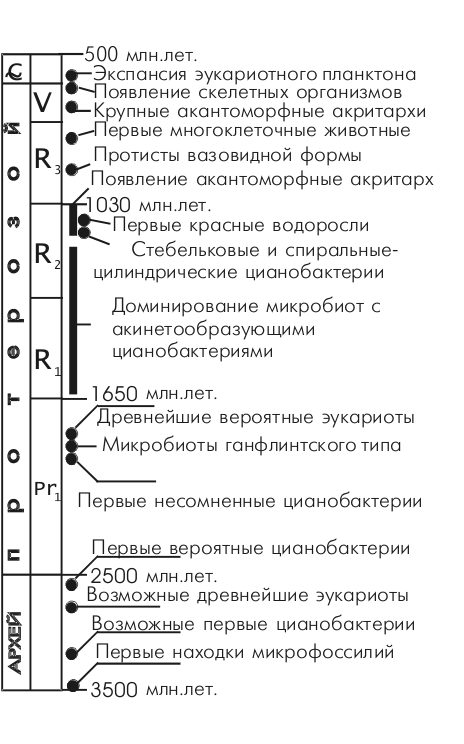
Пожалуй, самый удивительный факт, открытый палеонтологами, состоит в том, что следы жизни присутствуют уже в самых древних породах земной коры. Похоже, жизнь появилась почти одновременно с Землей, или, по крайней мере, наша планета оставалсь безжизненной очень недолго (см. схему периодизации истории биосферы по А.Ю.Розанову, 2002). Уже 3,5 млрд. лет назад, возможно, существовали фотосинтезирующие цианобактерии (сине-зеленые водоросли), а ведь это весьма сложные организмы. С точки зрения сложности организации, пропасть между неживым "первичным бульоном", каким бы он ни был, и прокариотическими организмами (бактериями) не меньше, а скорее больше, чем между бактериями и млекопитающими. Ранние, самые важные, этапы становления жизни: возникновение генетического кода, систем репликации, транскрипции и синтеза белка, липидных мембран – все эти великие ароморфозы не оставили никакого следа в палеонтологической летописи и могут реконструироваться только теоретически. Разброс противоречивых мнений, гипотез пока очень велик. Общее впечатление такое, что исследователи блуждают во мраке (см. подброрку статей в конце этой страницы). Единственная мысль, которая в последнее время более-менее прижилась, состоит в том, что самые первые живые организмы могли быть построены только из РНК (в отличие от современных, построенных из ДНК, РНК и белков). Дело в том, что только РНК способна выполнять и информационные, и структурные, и каталитические функции. Наследием этого древнего этапа "РНК-жизни" остался тот удивительный факт, что у всех живых организмов в системе синтеза белка РНК участвует не только как носитель информации, но выполняет также структурную и транспортную функции (рибосомальная и транспортная РНК). Это действительно ОЧЕНЬ странно, ведь теоретически с этой работой вполне могли бы справиться белки, которые и выполняют в клетке всю подобную работу (когда нужно транспортировать или синтезировать какие-либо молекулы); белки прекрасно "умеют" и распознавать определенные последовательности нуклеотидов, прикрепляться к ним (это т.н. факторы транскрипции). В целом, несмотря на обилие гипотез, следует признать, что удовлетворительной теории возникновения жизни на сегодняшний день не существует. Хорошо уже то, что возникновение и развитие жизни благодаря открытиям И.Пригожина перестало "противоречить" законам физики (а ведь был период, когда казалось, что только Бог может противостоять Второму Началу Термодинамики)…
В.Н.Снытников. Астрокатализ как стартовый этап геобиологических процессов. Жизнь создает планеты?
Г.А.Заварзин. Алкалофильное микробное сообщество как аналог наземной биоты протерозоя.
М.М.Астафьева. Архей Карелии и бактериальная палеонтология.
|
Весьма познавательная дискуссия автора сайта с креационистами по вопросу о возможности самопроизвольного зарождения жизни (там много новейших данных, обзор типичных возражений против теории абиогенеза и ответы на эти возражения) Появление жизни. Из кн. П.Т. де Шардена "Феномен человека" Происхождение жизни. Обзор из нового учебника К.Ю.Еськова. О происхождении жизни и древнейших этапах ее эволюции. Главы из популярной книги А.Ю.Журавлева. О происхождении жизни из кн. В.А.Красилова "Метаэкология" А.С.Спирин. Биосинтез белков, мир РНК и происхождение жизни А.С.Спирин. Рибонуклеиновые кислоты как центральное звено живой материи. В.В.Власов, А.В.Власов. Жизнь начиналась с РНК. Загадка начала жизни (из книги Ю.В.Чайковского). Популярный, современный обзор. Геохимический и биоценотический подход. Мои выписки и заметки на полях: здесь. А.Д.Кернс-Смит. Первые организмы. (кристаллы глины могли служить "матрицей" для возникновения первых организмов?) Мой доклад об эволюционном прогрессе, прочитанный на философском семинаре 18.11.2003 Обзор древнейших этапов эволюции жизни. Б.С.Соколов, М.А.Фелонкин, 1988. Г.А.Заварзин. Развитие микробных сообществ в истории Земли Г.А.Заварзин. Эволюция микробных сообществ (доклад) Г. А. Заварзин. Становление системы биогеохимических циклов. 2003. В.Н.Сергеев. Окремненные микрофоссилии докембрия. В.Н.Сергеев. Цианобактериальные сообщества на ранних этапах эволюции биосферы. А.Ф.Вейс. Органостенные микрофоссилии докембрия - важнейший компонент древней биоты А.Ю. Розанов. Ископаемые бактерии, седиментогенез и ранние стадии эволюции биосферы (2003 г. Палеонтологический журнал. В печати). A.Yu.Rozanov. Bacterial Paleontology (в статье - уникальные фотографии ископаемых бактерий) |
{kind=link}

современные строматолиты - слоистые минеральные образования,
созданные сообществами микроорганизмов
-
В нашей галактике должны быть планеты пригодные для жизни. (Антон Образцов)
Популярные статьи по происхождению жизни из журнала "Наука из первых рук", 2004.
Что мы знаем и чего не знаем об эволюции (Н. Добрецов) .pdf
Жизнь создает планеты? (В. Снытников, В. Пармон) .pdf
Естественный отбор среди молекул (В. Пармон) .pdf
Рожденные из кристаллов? (Н. Юшкин) .pdf
Ловчие сети эволюции (Н. Колчанов) .pdf
Сложность как мерило эволюционного прогресса (В. Бердников) .pdf
Микробы держат небо (Г. Заварзин) pdf
Похвальное слово катастрофам (А.Каныгин) pdf
Хронология древнейших этапов эволюции жизни по палеонтологическим данным
(из работы В.Н.Сергеева)
Дополнительные материалы:
-
Из классики: А. И. Опарин. ЖИЗНЬ, ЕЕ ПРИРОДА, ПРОИСХОЖДЕНИЕ И РАЗВИТИЕ. 1968 г.
-
Химические основы возникновения жизни (популярная стайтья на сайте www.nature.ru)
-
Последние новости о рибозимах и гипотезе "РНК-мира": http://www.scientific.ru/journal/news/1202/n231202.html http://www.nature.com/nsu/021216/021216-9.html
-
А.В.Керимбеков. Механизм выделения L - формы аминокислот на ранней стадии молекулярной эволюции. Предложено простое объяснение старой "загадки" - почему все белки построены только из L-аминокислот.
-
С.Б.Пашутин. Биогенез: мотивы и феномены возникновения жизни (материал сайта www.nature.ru)
-
Обсуждение идей Р.Шелдрейка об эволюции материи и происхождении жизни в связи с развитием информационных и "морфических" полей, с эволюцией в "мире идей". Натурфилософская, идеалистическая концепция. Но занятно.
-
Сайт Шелдрейка (англ.)
Эволюция прокариот по молекулярным данным
Весьма интересная статья на эту тему вышла в 2004 г.: Battistuzzi F.U., Feijao A., Hedges S.B. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land // BMC Evol Biol. 2004; 4: 44. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=533871
Авторы реконструировали эволюцию прокариот на основе последовательностей 32 белков, общих для 72 видов прокариот, относящихся ко всем основным группам. Причем выбраны были такие белки, для которых вероятность горизонтального переноса минимальна (в основном связанные с "хранением и обработкой информации"). Полученные результаты подтвердили "правильность" выделения большинства высших таксонов прокариот. Но есть и новшества, самое важное из которых - выделение новой "хорошо обоснованной" группы, включающей цианобактерий, актинобактерий и Deinococcus. Авторы назвали эту группу Terrabacteria и связали ее появление с ранней колонизацией суши.
Датировки событий (моментов расхождений эволюционных линий - клад) делались при помощи метода "локальных часов" (local clock method). Результаты получились следующие.
| Событие | Датировка (млрд. лет назад) | Прим. |
| Появление жизни. | < 4,1 | Поскольку анализ шел по белкам, имеется в виду, естественно, ДНК-РНК-белковая жизнь (а не РНК-организмы, которые должны были появиться еще раньше). |
| Появление метаногенеза (CO2 + H2 = CH4 + H2O + энергия). | 3,8 - 4,1 | Метаногены, действительно, могли быть первыми. Такие существа способны жить хоть на Юпитере, будь там подходящая температура. |
| Расхождение крупнейших эволюционных линий архей | 3,1-4,1 | |
| Появление бескислородного фотосинтеза | <3,2 | По мнению авторов, архейские строматолиты были построены бактериальными сообществами (матами), основу которых составляли анаэробные фотосинтетики. |
| Появление анаэробной метанотрофии | >3,1 | По мнению авторов, это соответствует изменениям изотопного состава углерода в породах возрастом 2,8 - 2,6 млрд. лет |
| Заселение суши | 2,8-3,1 | Авторы предполагают, что актино- и цианобактерии могли быть первопоселенцапми суши, и сам кислородный фотосинтез возник как адаптация к сухопутной жизни. |
| Появление цианобактерий | 2,6 | Авторы осознают, что это противоречит общепринятым взглядам о более раннем появлении ЦБ! Что поделаешь, так получается. |
| Появление аэробной метанотрофии | 2,5-2,8 | Может быть, кислород все-таки уже был в атмосфере к концу архея, несмотря на отсутствие цианобактерий? |
| Расхождение крупнейших эволюционных линий бактерий | 2,5 - 3,5 | Стали развиваться позже, чем археи. Все-таки археи - древнейшие из всех. |
"Террабактерии" (предполагаемые первопоселенцы суши, = актинобактерии + цианобактерии) выработали адаптации, направленные на защиту от высыхания и сильного солнечного излучения. Из этого авторы делают вывод, что их общий предок жил на суше. Если так, то появление кислородного фотосинтеза тоже можно рассматривать как приспособление к сухопутной жизни.
Надо сказать, что цианобактерии и актинобактерии и сейчас отлично живут на суше. Чрезвычайно интересно, что на выходах карбонатных пород развиваются особые водорослево-бактериальные сообщества, где большую роль играют актинолишайники - симбиотические организмы, состоящие из актинобактерий (мицелиальных прокариот, их раньше считали грибами и называли актиномицетами) и одноклеточных водорослей. Может быть, подобные актинолишайники (с цианобактериями в качестве водорослевого компонента) составляли основу сухопутной "растительности" в протерозое.
См. статью о лишайниках вообще и об актинолишайниках в частности:
Раннее появление жизни. : результат, вполне адекватный имеющимся представлениям.
Раннее появление метаногенеза: Очень вероятно. Мне всегда казалось, что организмы, которым для жизни не нужно ничего, кроме углекислого газа и водорода, должны быть очень древними.
Анаэробные метанотрофы. Бескислородное окисление метана осуществляют некоторые археи, живущие в ассоциациях с бактериями-сульфатредукторами. По-видимому, эти два процесса сопряжены (метан окисляется - сульфат восстанавливается). Есть, однако, археи, которые окисляют метан и без кислорода, и без помощи сульфат-редукторов; как они это делают - не известно, но у некоторых архей (Archaeoglobus fulgidus) присутствуют вместе гены обоих метаболических путей: окисления метана и сульфатредукции. Возможно, именно такие организмы появились в числе первых, >3,1 млрд лет назад.
Аэробные метанотрофы. Сюда относятся некоторые альфа- и гамма-протеобактерии. Им нужен кислород, следовательно, они, по идее, должны были появиться позже цианобактерий.
Анаэробные фотосинтетики. Как видно из текста, дата их появления (ок. 3,2 млрд. лет) основана на довольно зыбких основаниях. Вполне возможно, что это произошло существенно раньше.
Заселение прокариотами суши. Это один из ключевых моментов в статье. "Эволюция фототрофии была скорее всего связана с эволюцией других свойств, необходимых для выживания в стрессовых условиях. Важнейшим средством защиты от ультрафиолета являются пигменты - каротиноиды. Способностью к их синтезу обладают все фототрофные бактерии, а также группы, тесно связанные с сухопутными местообитаниями (такие как цианобактерии, актинобактерии и Deinococcus-Thermus. Пигментация, возможно, была фундаментальным шагом в колонизации "поверхностнгых" местообитаний. Помимо одинаковых светозащитных веществ, эти три группы (ц.-б., актинобактерии и дейнококкус) объединяет также повышенная устойчивость к высыханию (переносят обезвоживание). Это позволяет предположить, что их общий предок обитал на суше. Предложено общее название для этой группы: Terrabacteria. Заселение суши произошло ПОЗЖЕ расхождения этой группы и Firmicutes (3.05 (2.70–3.49) млрд. лет назад) и РАНЬШЕ отделения Actinobacteria от Cyanobacteria + Deinococcus (2.78 (2.49–3.20) млрд. лет назад). Эти "молекулярные" датировки соответствуют геологическим (2.6–2.7 млрд. лет) датировкам древнейших следов колонизации суши прокариотами."
Позднее происхождение цианобактерий. Этот вывод авторов противоречит общепринятым представлениям и особенно неприятен палеонтологам, которые интерпретируют некоторые древнейшие микрофоссилии именно как цианобактерии (см. выше на схеме из работы Сергеева: "Первые возможные цианобактерии", "Первые вероятные цианобактерии", "Первые несомненные цианобактерии"...). К тому же имеются архейские строматолиты, а главными строматолитостроители принято считать цианобактерий. Лично мне тоже "не подходит" такая датировка появления ц.-б. (2,6 млрд. лет назад), поскольку становится совершенно непонятно, что делают эвкариотические биомаркеры - стеролы - в отложениях возрастом 2,7 млрд. лет (см. ниже реферат статьи Brocks et al., 1999). Ведь эвкариоты по идее не могли появиться до тех пор, пока кто-то не начнет активно выделять кислород. Да что эвкариоты! Сам по себе синтез стеролов - кто бы их не синтезировал в те давние времена - требует свободного кислорода! Стало быть, если утверждается, что цианобактерии появились лишь 2,6 млрд. лет назад, то необходимо объяснить, кто (или что) производило кислород до этого момента.
Авторы указывают, что не только у них, но и у других исследователей по филогенетическим деревьям и молекулярным часам неизменно "получается", что цианобактерии появились "позже, чем надо". Это какая-то удивительная загадка.
Характерная цитата, показывающая, что все-таки абсолютно доверять предлагаемым авторами датировкам не следует: "On the other hand an inconsistency with the fossil record is represented by the divergence of unicellular (Thermosynechococcus elongatus) and heterocyst-forming (Nostoc sp.) cyanobacteria. Our time estimate for this divergence is 0.70–1.41 Ga while microfossils of both groups have been identified in Mesoproterozoic (1.5–1.3 Ga) and Paleoproterozoic (2.12–2.02 Ga) rocks. However the identification of these latter fossils has been debated... The reason for the discrepancy between the molecular and fossil times remains unclear but a possible misinterpretation of the fossil record cannot be dismissed."
Самые интересные рисунки из статьи:
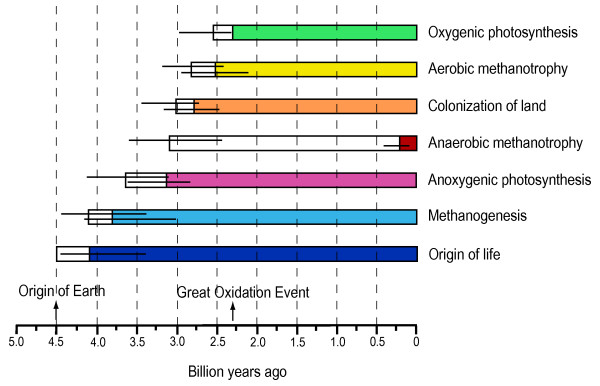
A
time line of metabolic innovations and events on Earth. The minimum time for
oxygenic photosynthesis is constrained by the Great Oxidation Event (2.3 Ga)
whereas the maximum time for the origin of life is constrained by the origin of
Earth (4.5 Ga). Horizontal lines indicate credibility intervals, white boxes
indicate minimum and maximum time constraints on the origin of a metabolism or
event, and colored boxes indicate the presence of the metabolism or
event.
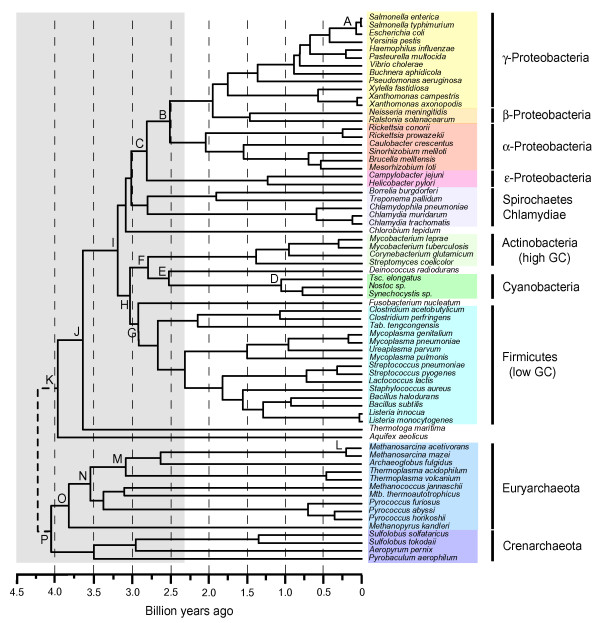
A
timescale of prokaryote evolution. Letters indicate nodes discussed in the text.
The last common ancestor was arbitrarily placed at 4.25 Ga in the tree, although
this placement was not part of the analyses. The grey rectangle shows the time
prior to the initial rise in oxygen (presumably anaerobic conditions). Mtb:
Methanothermobacter, Tab: Thermoanaerobacter, Tsc:
Thermosynechococcus.
Надо заметить, что для
"калибровки" своих датировок авторы все-таки использовали палеонтологические
данные. Правда, я, к сожалению, не смог из текста до конца понять, как именно
они их использовали. Но они "пробовали" разные варианты палеонтологических
датировок, и вроде бы их результаты не очень зависят от выбранных
палеонтологических маркеров.
Подборка рефератов по темам: "Палеонтологические данные о древнейших следах жизни на Земле. Бактериальная палеонтология Архея и Протерозоя"; "Происхождение жизни"
J Paleontol 1999 Sep;73(5):744-64
Acritarchs and microfossils from the Mesoproterozoic
Bangemall Group, northwestern Australia.
Buick
R, Knoll AH.
School of Geosciences, University of Sydney, Australia.
buick@es.su.oz.au
Three microfossil assemblages occur in the
Mesoproterozoic Bangemall Group (1625-1000 Ma) of northwestern Australia, each
occupying a different environmental and taphonomic setting. In peritidal
environments, benthic prokaryotic filaments and spheroids of matting habit and
small size were permineralized by early diagenetic silicification of
stromatolitic carbonates. In shallow subtidal environments, benthic filaments of
large size and nonmatting habit and planktonic sphaeromorph acritarchs with thin
walls and moderate dimensions were compressed in mildly kerogenous shale. In
deeper subtidal environments, planktonic megasphaeromorph acritarchs with thick
walls were initially entombed in concretionary nodules in highly kerogenous
shale and then permineralized by silica during later diagenesis. Taxonomic
diversity and numerical abundance evidently decrease offshore. The three
assemblages have typical Mesoproterozoic aspects: peritidal benthic habitats
were dominated by
Siphonophycus-Sphaerophycus-Eosynechococcus-Myxococcoides-Palaeopleurocapsa,
shallow subtidal settings were occupied by
Siphonophycus-Leiosphaeridia-Pterosphermopsimorpha-Satka, and offshore plankton
consisted solely of very large chuarid acritarchs. Because of its taphonomic
restriction to mid-intertidal stromatolites, the peritidal assemblage can be
equated in microenvironment with a similar assemblage in the Neoproterozoic
Draken Conglomerate, suggesting that ecological stasis at the community level
can last for intervals up to 900 million years. In the deeper subtidal
assemblage, the common chuarid has an unusual mode of preservation, in three
dimensions in early diagenetic concretions, revealing that it possesses a thick
multilamellate wall. Because of this distinctive ultrastructure, the new genus
Crassicorium is erected for these fossils, which are among the oldest
indubitable eukaryotes. Very large (34-55 micrometers in diameter) filaments
from shallow subtidal habitats are assigned to the emended species Siphonophycus
punctatum.
PMID: 11543499 [PubMed - indexed for MEDLINE]
Nature 2002 Mar 7;416(6876):76-81
Comment in: Nature. 2002 Mar 7;416(6876):28.
Questioning the evidence for Earth's oldest
fossils.
Brasier MD, Green OR, Jephcoat AP,
Kleppe AK, Van Kranendonk MJ, Lindsay JF, Steele A, Grassineau NV.
Earth
Sciences Department, University of Oxford, Parks Road, Oxford OX1 3PR, UK.
martinb@earth.ox.ac.uk
Structures resembling remarkably preserved
bacterial and cyanobacterial microfossils from about 3,465-million-year-old Apex
cherts of the Warrawoona Group in Western Australia currently provide the oldest
morphological evidence for life on Earth and have been taken to support an early
beginning for oxygen-producing photosynthesis. Eleven species of filamentous
prokaryote, distinguished by shape and geometry, have been put forward as
meeting the criteria required of authentic Archaean microfossils, and contrast
with other microfossils dismissed as either unreliable or unreproducible. These
structures are nearly a billion years older than putative cyanobacterial
biomarkers, genomic arguments for cyanobacteria, an oxygenic atmosphere and any
comparably diverse suite of microfossils. Here we report new research on the
type and re-collected material, involving mapping, optical and electron
microscopy, digital image analysis, micro-Raman spectroscopy and other
geochemical techniques. We reinterpret the purported microfossil-like structure
as secondary artefacts formed from amorphous graphite within multiple
generations of metalliferous hydrothermal vein chert and volcanic glass.
Although there is no support for primary biological morphology, a
Fischer--Tropsch-type synthesis of carbon compounds and carbon isotopic
fractionation is inferred for one of the oldest known hydrothermal systems on
Earth.
PMID: 11882895 [PubMed - indexed for MEDLINE]
Nature 1999 Aug 5;400(6744):554-7
2-Methylhopanoids as biomarkers for
cyanobacterial oxygenic photosynthesis.
Summons RE, Jahnke LL, Hope JM, Logan GA.
Australian Geological Survey
Organisation, Canberra, ACT. Roger.Summons@agso.gov.au
Oxygenic
photosynthesis is widely accepted as the most important bioenergetic process
happening in Earth's surface environment. It is thought to have evolved within
the cyanobacterial lineage, but it has been difficult to determine when it
began. Evidence based on the occurrence and appearance of stromatolites and
microfossils indicates that phototrophy occurred as long ago as 3,465 Myr
although no definite physiological inferences can be made from these objects.
Carbon isotopes and other geological phenomena provide clues but are also
equivocal. Biomarkers are potentially useful because the three domains of extant
life-Bacteria, Archaea and Eukarya-have signature membrane lipids with
recalcitrant carbon skeletons. These lipids turn into hydrocarbons in sediments
and can be found wherever the record is sufficiently well preserved. Here we
show that 2-methyl-bacteriohopanepolyols occur in a high proportion of cultured
cyanobacteria and cyanobacterial mats. Their 2-methylhopane hydrocarbon
derivatives are abundant in organic-rich sediments as old as 2,500 Myr. These
biomarkers may help constrain the age of the oldest cyanobacteria and the advent
of oxygenic photosynthesis. They could also be used to quantify the ecological
importance of cyanobacteria through geological time.
PMID: 10448856
[PubMed - indexed for MEDLINE]
Биомаркеры, по которым можно определить присутствие Архей, Бактерий, Эукариот
Science 1999 Aug 13;285(5430):1033-6
Comment in: Science. 1999 Aug 13;285(5430):1025-7
Archean molecular
fossils and the early rise of eukaryotes.
Brocks JJ, Logan GA, Buick R, Summons RE.
School of
Geosciences, University of Sydney, Sydney, NSW 2006, Australia.
jochen.brocks@agso.gov.au
Molecular fossils of biological lipids are
preserved in 2700-million-year-old shales from the Pilbara Craton, Australia.
Sequential extraction of adjacent samples shows that these hydrocarbon
biomarkers are indigenous and syngenetic to the Archean shales, greatly
extending the known geological range of such molecules. The presence of abundant
2alpha-methylhopanes, which are characteristic of cyanobacteria, indicates that
oxygenic photosynthesis evolved well before the atmosphere became oxidizing. The
presence of steranes, particularly cholestane and its 28- to 30-carbon analogs,
provides persuasive evidence for the existence of eukaryotes 500 million to 1
billion years before the extant fossil record indicates that the lineage
arose.
PMID: 10446042 [PubMed - indexed for MEDLINE]
появление эукариот 2,7 млрд. лет назад
Nature. 2001 Feb 22;409(6823):1083-91.
The habitat and nature of early life.
Nisbet EG, Sleep NH.
Department of Geology, University of
London, Egham, UK.
Earth is over 4,500 million years old. Massive
bombardment of the planet took place for the first 500-700 million years, and
the largest impacts would have been capable of sterilizing the planet. Probably
until 4,000 million years ago or later, occasional impacts might have heated the
ocean over 100 degrees C. Life on Earth dates from before about 3,800 million
years ago, and is likely to have gone through one or more hot-ocean
'bottlenecks'. Only hyperthermophiles (organisms optimally living in water at
80-110 degrees C) would have survived. It is possible that early life
diversified near hydrothermal vents, but hypotheses that life first occupied
other pre-bottleneck habitats are tenable (including transfer from Mars on
ejecta from impacts there). Early hyperthermophile life, probably near
hydrothermal systems, may have been non-photosynthetic, and many housekeeping
proteins and biochemical processes may have an original hydrothermal heritage.
The development of anoxygenic and then oxygenic photosynthesis would have
allowed life to escape the hydrothermal setting. By about 3,500 million years
ago, most of the principal biochemical pathways that sustain the modern
biosphere had evolved, and were global in scope.
Палеонтологические свидетельства существования наземных микробов 2,6-2,7 млрд. лет назад
Nature. 2000
Nov 30;408(6812):574-8.
Geochemical
evidence for terrestrial ecosystems 2.6 billion years
ago.
Watanabe Y, Martini JE, Ohmoto H.
Astrobiology Research
Center and Department of Geosciences, The Pennsylvania State University,
University Park 16802, USA. yumiko@essc.psu.edu
Microorganisms have
flourished in the oceans since at least 3.8 billion years (3.8 Gyr) ago, but it
is not at present clear when they first colonized the land. Organic matter in
some Au/U-rich conglomerates and ancient soils of 2.3-2.7 Gyr age has been
suggested as remnants of terrestrial organisms. Some 2.7-Gyr-old stromatolites
have also been suggested as structures created by terrestrial organisms.
However, it has been disputed whether this organic matter is indigenous or
exogenic, and whether these stromatolites formed in marine or fresh water.
Consequently, the oldest undisputed remnants of terrestrial organisms are
currently the 1.2-Gyr-old microfossils from Arizona, USA. Unusually carbonaceous
ancient soils--palaeosols--have been found in the Mpumalanga Province (Eastern
Transvaal) of South Africa. Here we report the occurrences, elemental ratios (C,
H, N, P) and isotopic compositions of this organic matter and its host rocks.
These data show that the organic matter very probably represents remnants of
microbial mats that developed on the soil surface between 2.6 and 2.7 Gyr ago.
This places the development of terrestrial biomass more than 1.4 billion years
earlier than previously reported.
[Proterozoic history and present state of cyanobacteria]
Mikrobiologiia 2002 Nov-Dec;71(6):725-40 Related Articles, Links
[Article in Russian]
Sergeev VN, Gerasimenko LM, Zavarzin GA.
Geological Institute, Russian Academy of Sciences, Pyzhevskii per. 7, Moscow, 109017 Russia.
The paper delves into the main regularities of the distribution of fossil microorganisms in Precambrian rocks, beginning from the Archean Eon about 3.5 billion years ago and ending in the Cambrian Period about 0.5 billion years ago. The paper analyzes facial peculiarities in the lateral differentiation of microfossils in Proterozoic basins and the main stages of temporal changes in fossil cyanobacterial communities, which are based on the irreversible succession of physicochemical conditions on the Earth and the evolution of eukaryotic microorganisms and their incorporation into prokaryotic ecosystems. To gain insight into Proterozoic fossil records, modern stratified cyanobacterial mats built up from layers of prokaryotes are considered. The analysis of phosphatization, carbonatization, and silification processes in modern algal-bacterial communities suggests that analogous processes took place in Proterozoic microbiotas. A comparison of modern and Precambrian living forms confirms the inference that cyanobacterial communities are very conservative and have changed insignificantly both morphologically and physiologically during the past two billion years.
PMID: 12526193
Paleobiology of the Mesoproterozoic Billyakh Group, Anabar Uplift, northern Siberia.
J Paleontol 1995 Jan;69(1 Pt 2):1-37
Sergeev VN, Knoll AH, Grotzinger JP.
Geological Institute, Russian Academy of Sciences, Moscow.
Silicified peritidal carbonates of the Mesoproterozoic Kotuikan and Yusmastakh Formations, Anabar Uplift, northeastern Siberia, contain exceptionally well-preserved microfossils… complex forms that typify Neoproterozoic assemblages are absent. The combination in Billyakh assemblages of exceptional preservation and low eukaryotic diversity supports the hypothesis that nucleated organisms diversified markedly near the Mesoproterozoic-Neoproterozoic boundary. The assemblages also demonstrate the antiquity of cyanobacteria capable of cell differentiation and suggest the importance of both changing peritidal substrates and evolving eukaryotes in determining stratigraphic patterns of Proterozoic prokaryotes…
PMID: 11539491
Annu Rev Earth Planet Sci 1999;27:313-58
Stromatolites in
Precambrian carbonates: evolutionary mileposts or environmental
dipsticks?
Grotzinger JP, Knoll
AH.
Department of Earth, Atmospheric and Planetary Sciences,
Massachusetts Institute of Technology, Cambridge 02139, USA.
grotz@mit.edu
Stromatolites are attached, lithified sedimentary growth
structures, accretionary away from a point or limited surface of initiation.
Though the accretion process is commonly regarded to result from the sediment
trapping or precipitation-inducing activities of microbial mats, little evidence
of this process is preserved in most Precambrian stromatolites. The successful
study and interpretation of stromatolites requires a process-based approach,
oriented toward deconvolving the replacement textures of ancient stromatolites.
The effects of diagenetic recrystallization first must be accounted for,
followed by analysis of lamination textures and deduction of possible accretion
mechanisms. Accretion hypotheses can be tested using numerical simulations based
on modem stromatolite growth processes. Application of this approach has shown
that stromatolites were originally formed largely through in situ precipitation
of laminae during Archean and older Proterozoic times, but that younger
Proterozoic stromatolites grew largely through the accretion of carbonate
sediments, most likely through the physical process of microbial trapping and
binding. This trend most likely reflects long-term evolution of the earth's
environment rather than microbial communities.
PMID: 11543060 [PubMed - indexed for MEDLINE]
Подвергается сомнению то, что изменения строматолитов отражают изменения биоты: возможно, только физических факторов.
Precambrian Res 1995 Nov;75(1-2):65-90
Microfossils from the
Neoarchean Campbell Group, Griqualand West Sequence of the Transvaal Supergroup,
and their paleoenvironmental and evolutionary implications.
Altermann W, Schopf JW.
IGPP Center for the Study of
Evolution and the Origin of Life, University of California, Los Angeles 90095,
USA.
The oldest filament- and colonial coccoid-containing microbial
fossil assemblage now known is described here from drill core samples of
stromatolitic cherty limestones of the Neoarchean, approximately 2600-Ma-old
Campbell Group (Ghaap Plateau Dolomite, Lime Acres Member) obtained at Lime
Acres, northern Cape Province, South Africa. The assemblage is biologically
diverse, including entophysalidacean (Eoentophysalis sp.), probable
chroococcacean (unnamed colonial coccoids), and oscillatoriacean cyanobacteria
(Eomycetopsis cf. filiformis, and Siphonophycus transvaalensis), as well as
filamentous fossil bacteria (Archaeotrichion sp.); filamentous possible
microfossils (unnamed hematitic filaments) also occur. The Campbell Group
microorganisms contributed to the formation of stratiform and domical to
columnar stromatolitic reefs in shallow subtidal to intertidal environments of
the Transvaal intracratonic sea. Although only moderately to poorly preserved,
they provide new evidence regarding the paleoenvironmental setting of the
Campbell Group sediments, extend the known time-range of entophysalidacean
cyanobacteria by more than 400 million years, substantiate the antiquity and
role in stromatolite formation of Archean oscillatoriacean cyanobacteria, and
document the exceedingly slow (hypobradytelic) evolutionary rate characteristic
of this early evolving prokaryotic lineage.
PMID: 11542814 [PubMed - indexed
for MEDLINE]
Precambrian Res 1992;54:271-93
Microfossils and possible microfossils from the Early
Archean Onverwacht Group, Barberton Mountain Land, South Africa.
Walsh MM.
Department of Geology and Geophysics,
Louisiana State University, Baton Rouge 70803, USA.
There is widespread
textural evidence for microbial activity in the cherts of the Early Archean
Onverwacht Group. Layers with fine carbonaceous laminations resembling fossil
microbial mats are abundant in the cherty metasediments of the predominantly
basaltic Hooggenoeg and Kromberg Formations. In rare cases, filamentous
microfossils are associated with the laminae. The morphologies of the fossils,
as well as the texture of the encompassing laminae suggest an affinity to modern
mat-dwelling cyanobacteria or bacteria. A variety of spheroidal and ellipsoidal
structures present in cherts of the Hooggenoeg and Kromberg Formations resemble
modern coccoidal bacteria and bacterial structures, including spores. The
development of spores may have enabled early microorganisms to survive the
relatively harsh surficial conditions, including the effects of very large
meteorite impacts on the young Earth.
PMID: 11540926 [PubMed - indexed for
MEDLINE]
Precambrian Res 1988;38:257-79
Distribution and
diagenesis of microfossils from the lower Proterozoic Duck Creek Dolomite,
Western Australia.
Knoll AH,
Strother PK, Rossi S.
Botanical Museum, Harvard University, Cambridge, MA 02138,
USA.
Two distinct generations of microfossils occur in silicified
carbonates from a previously undescribed locality of the Lower Proterozoic Duck
Creek Dolomite, Western Australia. The earlier generation occurs in discrete
organic-rich clasts and clots characterized by microquartz anhedra; it contains
a variety of filamentous and coccoidal fossils in varying states of
preservation. Second generation microfossils consist almost exclusively of
well-preserved Gunflintia minuta filaments that drape clasts or appear to float
in clear chalcedony. These filaments appear to represent an ecologically
distinct assemblage that colonized a substrate containing the partially degraded
remains of the first generation community. The two assemblages differ
significantly in taxonomic frequency distribution from previously described Duck
Creek florules. Taken together, Duck Creek microfossils exhibit a range of
assemblage variability comparable to that found in other Lower Proterozoic iron
formations and ferruginous carbonates. With increasing severity of post-mortem
alteration, Duck Creek microfossils appear to converge morphologically on
assemblages of simple microstructures described from early Archean cherts. Two
new species are described: Oscillatoriopsis majuscula and O. cuboides; the
former is among the largest septate filamentous fossils described from any
Proterozoic formation.
PMID: 11540084 [PubMed - indexed for MEDLINE]
Science 1993 Apr 30;260:640-6
Microfossils of the
Early Archean Apex chert: new evidence of the antiquity of life.
Schopf JW.
Department of Earth and Space Sciences,
University of California, Los Angeles 90024, USA.
Eleven taxa (including
eight heretofore undescribed species) of cellularly preserved filamentous
microbes, among the oldest fossils known, have been discovered in a bedded chert
unit of the Early Archean Apex Basalt of northwestern Western Australia. This
prokaryotic assemblage establishes that trichomic cyanobacterium-like
microorganisms were extant and morphologically diverse at least as early as
approximately 3465 million years ago and suggests that oxygen-producing
photoautotrophy may have already evolved by this early stage in biotic
history.
PMID: 11539831 [PubMed - indexed for MEDLINE]
Photosynth Res 1992;33:75-89
The oldest records of photosynthesis.
Awramik SM.
Department of Geological Sciences, University of California, Santa Barbara 93106, USA.
There is diverse, yet controversial fossil evidence for the existence
of photosynthesis 3500 million years ago. Among the most persuasive evidence is
the stromatolites described from low grade metasedimentary rocks in Western
Australia and South Africa. Based on the understanding of the paleobiology of
stromatolites and using pertinent fossil and Recent analogs, these Early Archean
stromatolites suggest that phototrophs evolved by 3500 million years ago. The
evidence allows further interpretation that cyanobacteria were involved. Besides
stromatolites, microbial and chemical fossils are also known from the same rock
units. Some microfossils morphologically resemble cyanobacteria and thus
complement the adduced cyanobacterial involvement in stromatolite construction.
If cyanobacteria had evolved by 3500 million years ago, this would indicate that
nearly all prokaryotic phyla had already evolved and that prokaryotes
diversified rapidly on the early Earth.
PMID: 11538389 [PubMed - indexed for
MEDLINE]
Фотосинтез появился 3,5 млрд лет назад.
Цитологическая, биохимическая литература по проблеме возникновения жизни. РНК-мир.
1. On the origins of cells: a hypothesis
for the evolutionary transitions from abiotic geochemistry to chemoautotrophic
prokaryotes, and from prokaryotes to nucleated cells.
Philos Trans R Soc Lond B Biol Sci 2003 Jan
29;358(1429):59-85
Martin W, Russell MJ.
Institut fur Botanik
III, Heinrich-Heine Universitaet Dusseldorf, Universitatsstrasse 1, 40225
Dusseldorf, Germany.
All life is organized as cells. Physical
compartmentation from the environment and self-organization of self-contained
redox reactions are the most conserved attributes of living things, hence
inorganic matter with such attributes would be life's most likely forebear. We
propose that life evolved in structured iron monosulphide precipitates in a
seepage site hydrothermal mound at a redox, pH and temperature gradient between
sulphide-rich hydrothermal fluid and iron(II)-containing waters of the Hadean
ocean floor. The naturally arising, three-dimensional compartmentation observed
within fossilized seepage-site metal sulphide precipitates indicates that these
inorganic compartments were the precursors of cell walls and membranes found in
free-living prokaryotes. The known capability of FeS and NiS to catalyse the
synthesis of the acetyl-methylsulphide from carbon monoxide and methylsulphide,
constituents of hydrothermal fluid, indicates that pre-biotic syntheses occurred
at the inner surfaces of these metal-sulphide-walled compartments, which
furthermore restrained reacted products from diffusion into the ocean, providing
sufficient concentrations of reactants to forge the transition from geochemistry
to biochemistry. The chemistry of what is known as the RNA-world could have
taken place within these naturally forming, catalyticwalled compartments to give
rise to replicating systems. Sufficient concentrations of precursors to support
replication would have been synthesized in situ geochemically and
biogeochemically, with FeS (and NiS) centres playing the central catalytic role.
The universal ancestor we infer was not a free-living cell, but rather was
confined to the naturally chemiosmotic, FeS compartments within which the
synthesis of its constituents occurred. The first free-living cells are
suggested to have been eubacterial and archaebacterial chemoautotrophs that
emerged more than 3.8 Gyr ago from their inorganic confines. We propose that the
emergence of these prokaryotic lineages from inorganic confines occurred
independently, facilitated by the independent origins of membrane-lipid
biosynthesis: isoprenoid ether membranes in the archaebacterial and fatty acid
ester membranes in the eubacterial lineage. The eukaryotes, all of which are
ancestrally heterotrophs and possess eubacterial lipids, are suggested to have
arisen ca. 2 Gyr ago through symbiosis involving an autotrophic archaebacterial
host and a heterotrophic eubacterial symbiont, the common ancestor of
mitochondria and hydrogenosomes. The attributes shared by all prokaryotes are
viewed as inheritances from their confined universal ancestor. The attributes
that distinguish eubacteria and archaebacteria, yet are uniform within the
groups, are viewed as relics of their phase of differentiation after divergence
from the non-free-living universal ancestor and before the origin of the
free-living chemoautotrophic lifestyle. The attributes shared by eukaryotes with
eubacteria and archaebacteria, respectively, are viewed as inheritances via
symbiosis. The attributes unique to eukaryotes are viewed as inventions specific
to their lineage. The origin of the eukaryotic endomembrane system and nuclear
membrane are suggested to be the fortuitous result of the expression of genes
for eubacterial membrane lipid synthesis by an archaebacterial genetic apparatus
in a compartment that was not fully prepared to accommodate such compounds,
resulting in vesicles of eubacterial lipids that accumulated in the cytosol
around their site of synthesis. Under these premises, the most ancient divide in
the living world is that between eubacteria and archaebacteria, yet the steepest
evolutionary grade is that between prokaryotes and eukaryotes.
PMID: 12594918
[PubMed - in process]
Симбиоз автотрофной архебактерии с гетеротрофными эубактериями (митохондриями). Происхождение ядра: синтез "эубактериальных" мембран в рез-те экспрессии эубактериальных генов архебактериальным генетическим аппаратом, в среде (цитоплазме), которая не была к таким веществам подготовлена; пузырьки липидных мембран скапливались вокруг места их синтеза – получилось ядро. У архебактерий другие мембраны (isoprenoid ether membranes) чем у эубактерий (fatty acid ester membranes)
Появление жизни в микрополостях сульфида железа. Эти полости были вместо клеток. Поверхность минерала – катализатор. Когда появились мембраны, "это" смогло выйти наружу и жить самостоятельно. Мембраны появились независимо в двух линиях: будущих Археях и Бактериях. Это первое великое разделение жизни. А самое большое эволюционное преобразование (града) – появление эукариот.
Популярная заметка об этой теории (рус.)
2. The origin of DNA genomes and DNA replication proteins.
Curr Opin Microbiol 2002 Oct;5(5):525-32 Related Articles, Links
Forterre P.
Institut de Genetique et Microbiologie, Batiment 409, CNRS UMR 8621, Universite Paris-Sud XI, F-91405 Orsay CEDEX, France. forterre@igmors.u-psud.fr
In recent years, it has became clear that most proteins involved in cellular DNA precursor synthesis or DNA replication have been 'invented' more than once, indicating that the transition from RNA to DNA genomes was more complex than previously thought. Several authors have suggested that DNA viruses, which often encode their own version of these proteins, played an important role in this process. The nature of the genome of the last universal cellular ancestor (LUCA) -- that is, RNA or DNA, prokaryotic-like or eukaryotic-like -- remains in dispute. A hyperthermophilic LUCA would have suggested a circular, double-stranded DNA genome; however, recent data favor a mesophilic or moderately thermophilic LUCA.
PMID: 12354562
3. Omnipotent RNA.
FEBS Lett 2002 Oct 23;530(1-3):4-8 Related Articles, Links
Spirin AS.
Institute of Protein Research, Russian Academy of Sciences, 142290 Pushchino, Moscow Region, Russia. spirin@vega.protres.ru
The capability of polyribonucleotide chains to form unique, compactly folded structures is considered the basis for diverse non-genetic functions of RNA, including the function of recognition of various ligands and the catalytic function. Together with well-known genetic functions of RNA - coding and complementary replication - this has led to the concept of the functional omnipotence of RNA and the hypothesis that an ancient RNA world supposedly preceded the contemporary DNA-RNA-protein life. It is proposed that the Woese universal precursor in the ancient RNA world could be a cell-free community of mixed RNA colonies growing and multiplying on solid surfaces.
PMID: 12387857
4. Small nucleolar RNAs: versatile
trans-acting molecules of ancient evolutionary origin.
Terns MP, Terns RM.
Gene Expr 2002;10(1-2):17-39
Department of Biochemistry and Molecular Biology, University of Georgia,
Athens 30602, USA. mterns@bmb.uga.edu
The small nucleolar RNAs (snoRNAs)
are an abundant class of trans-acting RNAs that function in ribosome biogenesis
in the eukaryotic nucleolus. Elegant work has revealed that most known snoRNAs
guide modification of pre-ribosomal RNA (pre-rRNA) by base pairing near target
sites. Other snoRNAs are involved in cleavage of pre-rRNA by mechanisms that
have not yet been detailed. Moreover, our appreciation of the cellular roles of
the snoRNAs is expanding with new evidence that snoRNAs also target modification
of small nuclear RNAs and messenger RNAs. Many snoRNAs are produced by
unorthodox modes of biogenesis including salvage from introns of pre-mRNAs. The
recent discovery that homologs of snoRNAs as well as associated proteins exist
in the domain Archaea indicates that the RNA-guided RNA modification system is
of ancient evolutionary origin. In addition, it has become clear that the RNA
component of vertebrate telomerase (an enzyme implicated in cancer and cellular
senescence) is related to snoRNAs. During its evolution, vertebrate telomerase
RNA appears to have co-opted a snoRNA domain that is essential for the function
of telomerase RNA in vivo. The unique properties of snoRNAs are now being
harnessed for basic research and therapeutic applications.
PMID:
11868985 [PubMed - indexed for
MEDLINE]
Существуют особые коротенькие РНК, которые участвуют в мдоификации рРНК, т.е., короче, в формировании рибосом. Причем они есть у Эукариот и Археа (указывает на некую связь между ними).
5. Archaeal translation initiation
revisited: the initiation factor 2 and eukaryotic initiation factor 2B
alpha-beta-delta subunit families.
Kyrpides NC, Woese CR.
Proc Natl Acad Sci U S A 1998 Mar
31;95(7):3726-30
Department of Microbiology, University of Illinois at
Urbana-Champaign, B103 Chemical and Life Sciences, MC 110, 407 S. Goodwin,
Urbana, IL 61801, USA. nikos@darwin.life.uiuc.edu
As the amount of
available sequence data increases, it becomes apparent that our understanding of
translation initiation is far from comprehensive and that prior conclusions
concerning the origin of the process are wrong. Contrary to earlier conclusions,
key elements of translation initiation originated at the Universal Ancestor
stage, for homologous counterparts exist in all three primary taxa. Herein, we
explore the evolutionary relationships among the components of bacterial
initiation factor 2 (IF-2) and eukaryotic IF-2 (eIF-2)/eIF-2B, i.e., the
initiation factors involved in introducing the initiator tRNA into the
translation mechanism and performing the first step in the peptide chain
elongation cycle. All Archaea appear to posses a fully functional eIF-2
molecule, but they lack the associated GTP recycling function, eIF-2B (a
five-subunit molecule). Yet, the Archaea do posses members of the gene family
defined by the (related) eIF-2B subunits alpha, beta, and delta, although these
are not specifically related to any of the three eukaryotic subunits. Additional
members of this family also occur in some (but by no means all) Bacteria and
even in some eukaryotes. The functional significance of the other members of
this family is unclear and requires experimental resolution. Similarly, the
occurrence of bacterial IF-2-like molecules in all Archaea and in some
eukaryotes further complicates the picture of translation initiation. Overall,
these data lend further support to the suggestion that the rudiments of
translation initiation were present at the Universal Ancestor
stage.
PMID: 9520434 [PubMed - indexed for MEDLINE]
Важно: факты в пользу того, что трансляция (т.е. синтез белка) присутствовала уже у "последнего общего предка", т.е. жизнь разошлась на линии Археа, Бактериа и Эукариа уже на такой стадии, когда были белки, а не только РНК