Верхняя половина т-РНК более древняя.
Не смотря на то, что молекула тРНК относительно небольшая, всё же она является весьма сложно устроенной. Целый ряд фактов даёт основания предполагать, что современная форма тРНК возникла не стразу, и разные её части возникли в разное время.
Майцелс и Вайнер предположили, чтот-РНК-подобная структура древних РНКовых организмов состояла из коаксиальных T-петли и акцепторного стебля [Maizels N., Weiner A.M. 1994]. Это предположение основано на двух группах известных фактов.
Во-первых, верхняя половинасовременных т-РНК является независимым структурным доменом, который узнается РНКазой P (один из ключевых молекулярных реликтов, предположительно существовавших ещё в мире РНК, о ней подробнее далее в статье), фактором элонгации Tu, АРСазами и даже рибосомными РНК. Важность этой части т-РНК почти во всех макромолекулярных взаимодействиях дает основание считать, что она более древняя, чем нижняя половины молекулы.
Во-вторых, способность клеток различать т-РНК разных видов зависит, преимущественно отспецифических нуклеотидов верхней половины молекулы т-РНК, включая «дискриминаторное основание» (это комплементарная пара оснований, находящаяся в полжении 2-72 (о нумерации см. Аминоацил-тРНК-синтетазы - два класса ферментов) и являющаяся ключевой для распознавания АРСазами). Авторы оставляют открытым вопрос о возникновении нижней половины тРНК (рис.1).
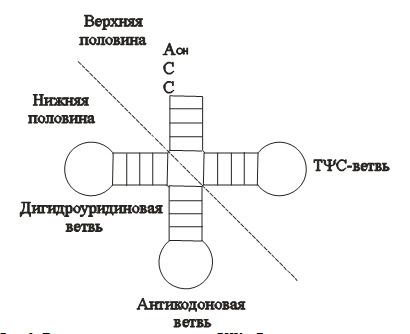
Рис.1.
Две половины современных т-РНК. Верхняя половина структурно и функционально независима и может быть более древней, чем нижняя половина.
Верхнияя половина геометрически обособлена от нижней при формировании третичной структуры тРНК - L-формы (рис. 2).
АРСазы обязательно взаимодействуют с верхней половиной, тогда как взаимодействие с нижней половиной происходит далеко не всегда. В рамках рассмотренной модели считается, что домен АРСазы, взаимодействующий с нижней половиной, возник позднее.
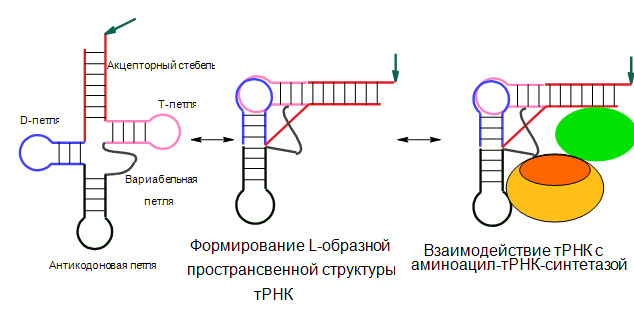
Рис. 2. Схема образования пространственной L-образной стуктуры тРНК состоящей из верхней и нижней и взаимодействия её с АРСазой.
Стрелкой обозначено место присоединенияаминокислоты при аминоацилировании тРНК синтетазой. Красным цветом выделенакцепторный домен тРНК, синим – антикодоновый домен. Овалами обозначены доменыАРСазы: зелёный – каталитический домен, содержащий домен связывания иаминоацилирования акцепторной области тРНК, жёлтым и оранжевым – вариабельныйдомен АРСазы. В зависимости от размера этого домена, АРСаза распознаёт вариабельным доменом антикодоновую область (домен обозначен жёлтым цветом), илине распознаёт (домен обозначен оранжевым цветом).
Гипотеза протокодона.
В.Мёллер и Г.Янссен в начале 90-х годов прошлого века основываясь на статистическом сравнении последовательностей антикодона и акцепторного стебля обнаружили для бактериальных тРНК высокую частоту встречаемости (от 20 до 72%) кодоновой последовательности в акцепторном стебле в позиции 3-5 акцепторного стебля аланиновой (кодоновая последовательность GCC), глициновой (GGC), аспарагиновой (GAC) и валиновой (GTC) тРНК [Moller W., Janssen G.M.C. 1990, 1992]. Эти кодоновые последовательности в акцепторном стебле были названы ПРОТОКОДОНОМ (см. рис.3):
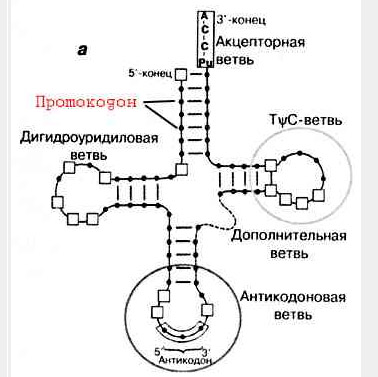
Рис. 3. Расположение протокодона на тРНК.
Попробуем проанализировать этот факт. Какие мы увидим закономерности?
Нетрудно увидеть, что набор этих протокодонов представлен двумя взаимно комплементарными парами: GGC/GCC и GAC/GTC. Этот факт сам по себе может быть неслучайным. Этот факт находится в хорошем соответствии с моделью кодирования тРНК с комплементарными антикодонами на разных комплементарных цепях (см. Древние тРНК с комплементарными антикодонами кодировались комплементарными РНК-парами).
Далее, антикодоновая последовательность присутствует с заметной частотой именно для тех аминокислот, которые с наибольшей частотой обнаруживаются в экспериментах по моделированию абиогенного синтеза в подвергающихся электрическим разрядам газовых смесях из аммиака, воды, водорода, а также метана или углекислого газа [Miller S.L. 1986]. Именно эти четыре аминокислоты являются наиболее часто встречаемыми в углеродистых метеоритах [Kvenvolden K.A. et. al. 1970]. Однако не все избыточные аминокислоты, полученные в опытах, подобным опытам Миллера и в углеродистых метеоритах включены в ГК: в код не включены такие распространённые аминокислоты, как гамма-аминомасляная кислота, норвалин, норлейцин. Это можно объяснить тем, что возможно для формирования раннего пептидного синтеза важна была не только избыточность аминокислот сама по себе, но стереохимическое соответствие аминокислот нуклеотидным конфигурациям, а также такой важный параметр, как способность этих аминокислот обеспечивать функциональность пептидных молекул.
Предложенная модель допускает, что синтез белков мог быть обусловлен прямым. взаимодействием аминокислот с кодон-антикодоновой пары в соответствии со стереохимическими правилами. Один из возможных вариантов – аминоацилирование таких миниспиралей было обусловлено в том числе и прямым взаимодействием с кодон-антикодоновй парой. Сам матричный синтез пептидов при этом мог отсутствовать, однако могли образовываться случайные пептиды с неслучайной композицией в результате какого-то предкового варианта транспептидации (синтез белка происходит только за счёт переноса аминокислоты с одной аминоацилированной тРНК на другую, это и есть транспептидация) между двумя тРНК или на одной тРНК в соответсвии с гипотезой Д.Мораса (см. Аминоацил-тРНК-синтетазы - два класса ферментов).
Основываясь на полученных данных, В.Мёллер и Г.Янссен предположили, что древние тРНК первоначально представляли собой миниспирали (рис. 4 (a)), содержащие кодон-антикодоновую пару в акцепторном участке, вблизи позиции аминоацилирования. После репликации (рис. 4 (b)) таких коротких миниспиралей мог (возможно автокаталитически) выщепляться фрагмент, содержащий кодоновый участок одной из цепей (рис. 4 (c)), Появление же антикодоновой петли по их мнению было вызвано каталитическим (или автокаталитическим) выщеплением фрагмента РНК, содержащего кодон, и последующим лигированием концов большего фрагмента РНК, которое привело к образованию антикодоновой петли (рис. 4 (e)).
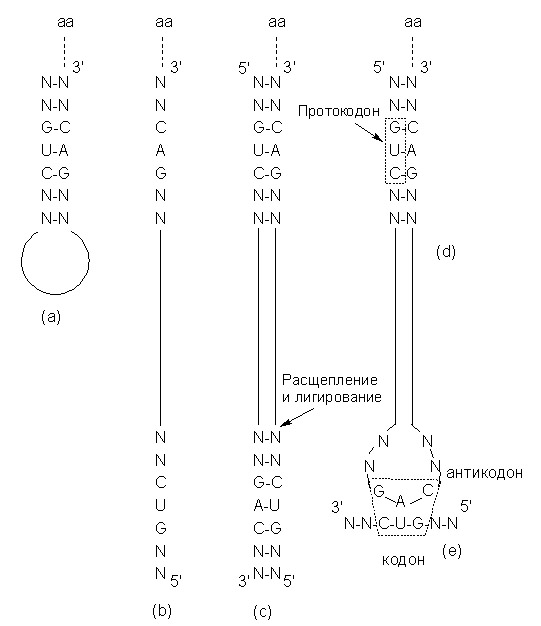
Рис. 4. Модель превращения древней короткой тРНК в тРНК современного размера.
Обозначения: аа - аминокислота, N - любой возможный нуклеотид.
Каталитическое (в частности, автокаталитическое) выщепление фрагмента тРНК выглядит вполне правдоподобным в свете известных фактов по выщеплению интронных областей из РНК-транскриптов генов тРНК. Каталитическое расщепление транскриптов тРНК самой РНК в современных организмах – довольно распространённое явление. РНК являетеся ключевым компонентом в рибонуклеазе P, катализирующей вырезание из РНК-транскрипта интронных областей. Особенно примечательно, что ряд дрожжевых тРНК содержат короткие интроны, локализованные на расстоянии 1-2 оснований от 3’-конца антикодона созревшей тРНК. Особенно впечатляет, что этот интрон в современных организмах может нести кодон для аминокислоты, переносимой соответствующей тРНК. Сайт выщепления этой интронной последовательности лежит вблизи 3’, а не 5’-конца антикодона, то есть расположение выщепляемого фрагмента пре-тРНК сходно с расположением выщепляемого фрагмента в предлагаемой модели. Этот факт как раз находится в полном соответствии с тем фактом, что протокодоновая последовательность обнаруживается вблизи 5’, а не 3’-конца тРНК.
В пользу предложенной гипотезы удвоения предковой шпильки говорит также тот факт, что антикодоновая последовательность в современных тРНК расположена почти посередине.
Предковый вариант транспептидации мог осуществляться при взаимодействии шпилек на аминоацилированных миниспиралях, который как раз и приводил к синтезу пула случайных последовательностей пептидов с неслучайным аминокислотным составом, и, возможно, с преимущественным синтезом каких-то вариантов последовательностей.
Кодируемый пептидный синтез возник лишь после удвоения миниспирали и выщепления кодонового участка из двуцепочечной с последующим соединением образовавшихся концов и формированием антикодоновой петли.
Причиной, по которой сохранилась эта протокодоновая последовательность в современных тРНК – это то, что данная последовательность содержит элементы узнавания для АРСаз.
Предложенная модель также нахоидтся в хорошем соответствии с тем фактом, что данных четырёх аминокилот достаточно для образования функционального разнообразия образуемых ими пептидов [Ikehara K. et al. 2002] (данный вопрос в рамках модели первоначального GNC-кода будет рассмотрен отдельно).
Резюме.
1. Современные аланиновые, глициновые, валиновые и глутаматные тРНК бактерий несут в акцепторном стебле в положении 3-5 кодоновую область, кодирующую в точности ту аминокислоту, которой они аминоацилируются. 4 кодоновые последовательности представляют собой две комплементарные пары: GGC/GCC и GAC/GTC.
2. Антикодоновый стебель современных тРНК образовался в результате удвовения относительно короткой предковой молекулы, несущей кодон-антикодоновую пару в акцепторном стебле, по крайней мере для этих четырёх аминокислот.
3. Сами предковые миниспирали, способны были аминоацилироваться теми тРНК, которые преимущественно образовывались абиогенным образом.
4. Не все аминокислоты, образованные абиогенно в существенных количествах, были использованы для формирования генетического кода. Это было обусловлено тем, что наряду с избыточностью первых аминокислот, важным также было стереохимическое соответствие между аминокислотами и нуклеотидными композициями.
Словарь:
Аминоацилирование - присоединение аминокислоты к акцепторному стеблю тРНК. Без аминоацилирования синтез белка невозможен.
Лигирование - сшивка двух нуклеотидных полимеров в один (стык в стык).
Транспептидация - перенос на растущую синтезируемую белковую цепь новой аминокислоты.
Литература.
1. Maizels N., Weiner A.M. Phylogeny from function: Evidence from the molecular fossil record that tRNA originated in replication, not translation. // Proc.Nat.Acad.Sci.USA. 1994. P.6729-6734.
2. Moller W., Janssen G.M.C. Statistical evidence for remnants of the Primordial code in the acceptor stem of prokaryotic transfer RNA. // J. Mol. Evol. 1992. P.471-477.
3. Moller W., Janssen G.M.C. Transfer RNA for primordial amino acid contain remants of a primitive code at position 3 to 5. // Biochimie. 1990. V.72. P.361-368.
4. Miller S.L. Current status of the prebiotic synthesis of small molecules. // Chem. Scr. 1986. V.26B. P.361-368.
5. Ikehara K., Omori Y., Arai R., Hirose A. A novel theory on theorigin of the genetic code: A GNC-SNS hypothesis // J.Molec. Evol. 2002. V.54. P.530-538