Идея о том, что тРНК с комплементарными антикодонами могли первоначально кодироваться комплементарными цепями одной двуспиральной РНК была выдвинута относительно давно - ещё в 70-х годах прошлого века Эйгеном и Шустером. Они предположили, что древние тРНК могли появиться наиболее коротким путём – синтезом комплементарных нитей РНК с комплементарными антикодонами [Эйген М., Шустер П. 1982; Eigen M., Schuster P. 1979].
Зачем нужно было объединять РНК в пары? Дело в том, что если мы рассмотрим современные вирусные РНК-геномы, то увидим плачевную картину. ОНИ ОЧЕНЬ МАЛЕНЬКИЕ. РНК-вирусы НЕ МОГУТ БЫТЬ БОЛЬШИМИ, они не могут кодировать много генов. Дело в том, что РНК так устрроена, что в отличие от ДНК, она не может столь же точно копироваться.
Если бы РНК-вирусы были такими же большими, что и ДНК-вирусы, то всё потомство у них было бы со смертельными мутациями. Поэтому все современные РНК-вирусы всячески хитрят и извращаются для увеличения количества своих генов. Одна из хитростей - РНК- вирусы кодируют гены не на одной, а на обоих комплементарных цепях.
Эти авторы сравнили последовательности имеющиеся в наличии тРНК с комплементарными антикодонами и не обнаружили статистически достоверной комплементарности последовательностей и заключили, что гипотеза комплементарности таких тРНК является непроверяемой, вследствие накопившегося мутационного шума (радномизации) в современных тРНК. Этот анализ проводился во второй половине 70-х годов прошлого века, когда отсеквенированных последовательностей тРНК было очень мало.
Прошло очень много лет по меркам быстро развивающейся молекулярной биологии (революцию, которую обыватель и не заметил). Новое экспериментальное тестирование гипотезы комплементарности последовательностей тРНК с комплементарными антикодонами было продолжено С.Н. Родиным с соавторами в начале 90-х годов прошлого века [Rodin S. et. al.1993], когда количество отсеквенированных последовательностей тРНК приблизилось к 900.
Для того, чтобы вычленить из частично рандомизированной последовательности информацию для сравнения тРНК с комплементарными антикодонами авторы учитывали следующие ньюансы.
1. Нужно сравнивать не все последовательности тРНК, а только достаточно медленно изменяющиеся, но не константрые. Быстроизменяющиеся последовательности не подойдут - у них уже стёрлась информация о самом древнем предке, жившем больше 3 млрд. лет назад. Данное сравнение позволило исключить из рассмотрения гипервариабельные позиции, к которым, в частности относится последовательность акцепторного стебля [Eigen M. et. al. 1988, 1989; Родин С. Н. 1991]. Аминоацилируемый стебель, однако может служить как внутренний контроль радномизации последовательностей. Поэтому в рассмотрение также включалась полследовательность акцепторного домена за искючением терминальной полностью консервативной последовательности RCCA (находится на 3'-конце, к которому присоединяется аминокислота). Напомню, что R - это пурин, то есть А или G.
2. Чтобы анализируемые последовательности соответствовали более древним, они начали сравнивать не сами последовательности, а результаты их статистического усреднения для каждой отдельной аминокислоты.
То есть брали выборку тРНК из разных организмов, переносящих одну и ту же аминокислоту, и усредняли.
Молекулы РНК асимметричны. То есть у них есть голова - это 5'-конец (посмотрите, кому интересно в википедии) и хвост - 3'-конец. В двойной цепи ДНК или РНК 5'-конец всегда находится напротив 3'-конца. Авторы стали сравнивать тРНК с комплементарными антикодонами, располагая их параллельно или антипараллельно.
Так вот, авторы сравнили усреднённые последовательности, располагая их друг напротив друга параллельно (голова к голове) и антипараллельно (голова к хвосту, как в двойной цепи).
И обнаружили интереснейшую вещь. Оказалось, что тРНК с комплементарными антикодонами, также имеют комплементарыне антикодоновые ветви и Т-ветви. Это сходство обнаруживается только при антипараллельном стравнении, но никак не при параллельном сравнении (см. рис. 1).
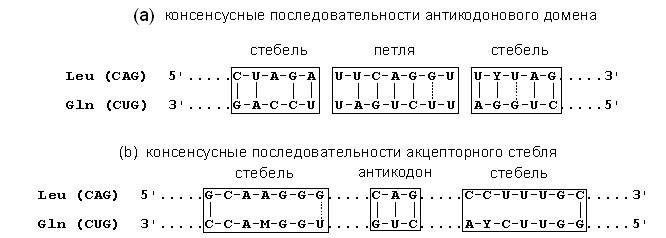
Рис.1. Сравнение консенсусных последовательностей в антипараллельной ориентации лейциновой (CAG) и глутаминовой (CUG) тРНК в акцепторном стебле (a) и антикодоновом домене (b). Сплошные линии показывают каноническое комплементарное спаривание, а пунктирные – неканоническую пару G-U [Rodin S. et. al. 1990]. Сверху - установленная комплементарность для антикодоновой ветви. Снизу - отрицательный контроль - отсутствие комплементарности. Сверху на рисунке приведено сравнение антикодоновых ветвей для двух типов тРНК с комплементарными антикодонами (эта последовательность очень древняя - ей миллиарды лет!). Внизу в качестве отрицательного контроля приведено сравнение акцепторных стеблей этих тРНК. Вертикальные чёрточки как раз и показывают комплементарность. Видно, что вверху последовательности комплементарны, а внизу (отрицательный контроль) - нет.
Этот факт означает - древние тРНК кодировались парами.
Примечательно, что если пары комплементарны, то после расхождения нитей от двойной спирали, полученные тРНК должны иметь одинаковую вторичную структуру. Данная модель схематически изображена на рис. 2.
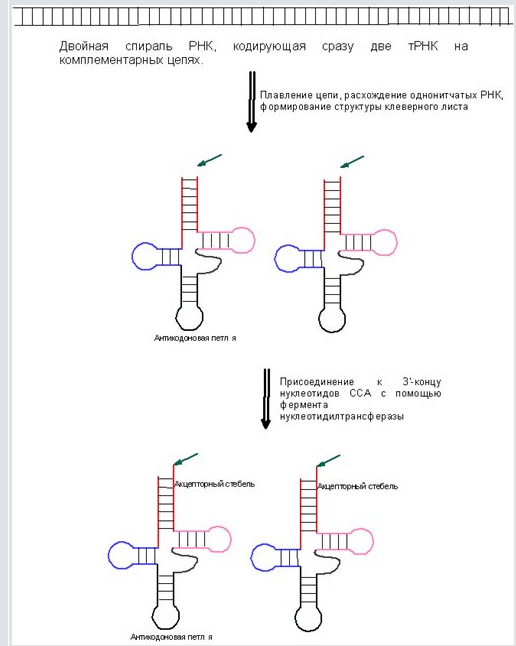
Вначале двуцепочечная РНК представляет собой просто ген, то есть кодирующую последовательность. На первом этапе эти нити расходятся - получаются две нити одноцепочечной РНК, которые пространственно сворачиваются с образованием структуры клеверного листа.
На втором этапе к 3'-хвосту молекулы присоединяются три дополнительных нуклеотида CCA (важно понять, что они не кодируются на последовательности тРНК, а наращиваются позднее, как и в значительной части современных организмов, эта тройка нуклеотидов обязательно присутствует во всех молекулах тРНК без исключения), которые совершенно необходимы для аминоацилирования (присоединения к хвосту аминокислоты).
Таким образом, получаем: одна последовательность - два продукта.
Резюме.
1. В РНК-мире точность воспроизведения была низкая. Поэтому остро стояла проблема компактности генома. Поэтому многие гены могли кодироваться на комплементарных нитях.
2. Древние тРНК кодировались одной и той же двойной спиралью. Послед расхождения отдельный цепей возинкали из одной молекулы две тРНК с одинаковой вторичной структурой.
Примечательно, что к этому же выводу мы придём на основе других данных чуть позднее.
Литература.
1. Rodin S N Ohno S Four primordial modes of tRNA-synthetase recognition,determined by the (G,C) operational code // Proc.Nat.Acad.Sci.USA . - 94,10 . - 1997 . - 5183-5188 .
2. Rodin S., Ohno S., Rodin A. Transfer RNA with complementary anticodon: Could they reflect early evolution of discriminativ genetic code adaptor? // Proc. Natl. Acad. Sci. USA, 1993. V.90. P.4723–4727.
3. Rodin S., Rodin A. Evolution by gene duplication: from the origin of the genetic code to the human genome. In Biosphere origin and evolution. Springer, 2008. P.257-276.
4. Rodin S., Rodin A., Ohno S. The presence of codon-anticodon pairs in the acceptor stem of tRNAs. Proc. Natl. Acad. Sci. USA, 1996. V.93. P.4537–3542.