Системная нейрофизиология (nan)
Карта роста аксонов
РедактироватьДетерминация процессов, посредством которых RGCs образуют топографические связи, крайне важна для выявления роли graded guidance молекул в становлении карты и создания моделей этого процесса. Тщательный количественный анализ показал, что это единственный механизм образования карты и в настоящее время начала вырисовываться относительная важность направления роста аксонов, их ветвления и роль «управляющих» молекул в контроле создания топографических карт
Вес уверенности:
Основная функция нервной системы - интерпретация средовых сигналов, осуществляемая через различные органы чувств, и передача информации в мозг. Для решения этой задачи, поступающая информация должна быть организована соответствующим и эффективным образом. Наиболее эффективная организация достигается с помощью так называемых топографических карт, присутствующих во всем мозге (Kaas,1997). Топографическая карта – это проекция из одной группы нейронов в другую, когда «получающая» группа клеток отражает связи соседних проецирующих клеток. В нервной системе высших позвоночных топографические карты включают сенсорные карты тела, tonotopic карты слуховых стимулов и карты визуального поля. Более того, топографические карты персистируют по схеме от первого порядка к более высокому порядку связей.
| THE ACCESSORY OLFACTORY SYSTEM The vomeronasal organ (VNO), located in the ventral nasal cavity, projects to the accessory olfactory bulb (AOB), found posterior-dorsal to the main olfactory bulb using zonal topography. Neurons in apical VNO project to anterior AOB, and neurons in basal VNO project to posterior AOB (Figure 5). VNO neurons expressing a given pheromone receptor form multiple glomeruli within eachAOBhalf (Belluscio et al. 1999, Rodriguez et al. 1999). The distributions and known activities of several families of guidance molecules are described in Figure 5. As in the visual system, distinct combinations of attractive and repellent molecular activities guide VNO axons to their topographically appropriate zones (Knoll & Drescher 2002). The possibility that neuropilin-2 shapes the gradient of secreted semaphorins is an important concept to be considered in all systems. Furthermore, the apparent use of the GPI-linked ephrin-As as guidance receptors (or part of a receptor complex) has important consequences for models of topographic mapping. The accessory olfactory system, owing to its expression of several molecular guidance families in relatively simple patterns and its zonal topography, is an excellent model system to examine the combinatorial interactions between, and within, families of guidance molecules. |
Развитие топографических карт было изучено в нескольких проекционных системах позвоночных – таламокортикальной (Dufour et al. 2003, Vanderhaeghen & Polleux 2004), гиппокампсептальной (Gao et al. 1996, Yue et al. 2002), ольфакторно/вомероназальной (Sidebar 1), системе двигательных аксонов в мышцы (Feng et al. 2000, Nguyen et al.2002), и в сетчатке. Последняя система, сформированная аксонами ганглиозных клеток сетчатки (retinal ganglion cells - RGCs) к их наиболее известным мишеням среднего мозга – оптическому тектуму (optic tectum - OT) у рыб, амфибий и цыплят или к верхнему колликулусу (superior colliculus - SC) у млекопитающих, стала основной моделью для изучения развития топографических карт и градиентов сигнальных («управляющих») молекул, контролирующих их формирование. Авторы акцентировали свое внимание на зрительной системе и на механизмах картирования в мишенях с тем, чтобы детализировать развитие нейральной карты у позвоночных и осветить молекулярные механизмы контроля этого процесса. Роль ростовых конусов и запутанность множества взаимодействующих сигнальных путей рассмотрены в работах (обзоры Huber et al. 2003, van Horck et al. 2004). Наибольшее внимание уделяется млекопитающим и куриному эмбриону, у которых аксональные механизмы развития карты нуждаются в уникальных действиях топографических молекул-гидов (guidance molecules) в специфической временной последовательности. Наряду с этим приводятся примеры механизмов и молекул, участвующих в развитии карт низших позвоночных – лягушек и рыб. Репрезентация сетчатки на OT и SC может быть довольно простой для картирования двух ортогонально ориентированных осей – темпорально-назальной (temporal-nasal - TN) оси сетчатки вдоль передне-задней оси (anterior-posterior axis -AP) OT/SC, и дорсо-вентральной (dorsal-ventral - DV) оси сетчатки вдоль lateral-medial (LM) оси OT/SC (соответственно ventral-dorsal OT оси у немлекопитающих позвоночных). Критерием топографических молекул-гидов в ретинотектальной проекции является их градуированная и ограниченная экспрессия в сетчатке или в OT/SC, а также тот факт, что RGC аксоны из различных частей сетчатки по разному реагируют на эти молекулы и оказывают эффект на RGS картирование in vivo.
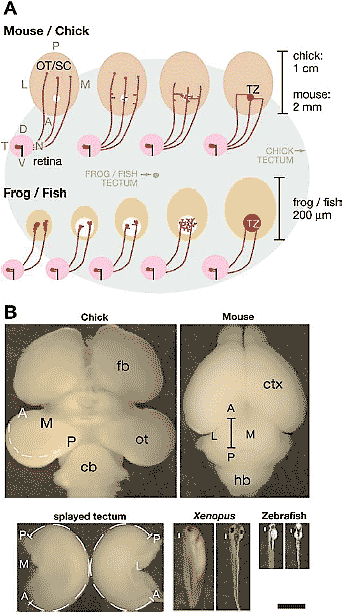 Figure 1 Development of the retinotopic projection and relative scale of the tectum in primary model species. (A, Top) In mouse and chick, RGC axons enter the OT/SC and initially extend well posterior to the location of their future termination zone (TZ) (circle). Interstitial branches form along the axon shaft in a distribution biased for the AP location of the TZ and subsequently exhibit bidirectional growth along the LM axis toward their correct TZ. Upon reaching their TZ, branches elaborate complex arbors and the initial axon overshoot is eliminated. All arbors are formed by interstitial branches. (A, Bottom) In frog (Xenopus laevis) and zebrafish, the tectum and retina expand throughout the development of the retinal projection. During retinotopic map development the tectum is much smaller in relation to a typical growth cone in frog and fish than in chick and mouse. RGC axons extend into the tectum and elaborate many small branches from the base of the growth cone. Arbors elaborate from these backbranches and the thinned growth cone. The TZ becomes dense and refines as the tectum enlarges. (A, Background ) The two ovals in the background represent the relative sizes of the chick tectum (large oval ) and frog or zebrafish tectum (small oval ). (B ) The photographs are at the same scale. The chick OT rotates during development such that the posterior pole (P) is near the midline. The OT is cut along the AP axis at the LM midline (dashed line) and splayed. The distance from the anterior to posterior pole along the cut edge is 1 cm (dashed lines in the splayed tectum). The mouse SC is about 2 mmalong the AP pole at the LM midline (bar). For frog and zebrafish the entire animal is shown in lateral and dorsal views. The white bar on the left of each panel represents the approximate AP position and size of the tectum. The tecta for these organisms are approximately 200 µm along the AP axis. cb, cerebellum; ctx, cortex; fb, forebrain; hb, hindbrain. Scale bar = 2 mm. Adapted from McLaughlin et al. 2003a.
Figure 1 Development of the retinotopic projection and relative scale of the tectum in primary model species. (A, Top) In mouse and chick, RGC axons enter the OT/SC and initially extend well posterior to the location of their future termination zone (TZ) (circle). Interstitial branches form along the axon shaft in a distribution biased for the AP location of the TZ and subsequently exhibit bidirectional growth along the LM axis toward their correct TZ. Upon reaching their TZ, branches elaborate complex arbors and the initial axon overshoot is eliminated. All arbors are formed by interstitial branches. (A, Bottom) In frog (Xenopus laevis) and zebrafish, the tectum and retina expand throughout the development of the retinal projection. During retinotopic map development the tectum is much smaller in relation to a typical growth cone in frog and fish than in chick and mouse. RGC axons extend into the tectum and elaborate many small branches from the base of the growth cone. Arbors elaborate from these backbranches and the thinned growth cone. The TZ becomes dense and refines as the tectum enlarges. (A, Background ) The two ovals in the background represent the relative sizes of the chick tectum (large oval ) and frog or zebrafish tectum (small oval ). (B ) The photographs are at the same scale. The chick OT rotates during development such that the posterior pole (P) is near the midline. The OT is cut along the AP axis at the LM midline (dashed line) and splayed. The distance from the anterior to posterior pole along the cut edge is 1 cm (dashed lines in the splayed tectum). The mouse SC is about 2 mmalong the AP pole at the LM midline (bar). For frog and zebrafish the entire animal is shown in lateral and dorsal views. The white bar on the left of each panel represents the approximate AP position and size of the tectum. The tecta for these organisms are approximately 200 µm along the AP axis. cb, cerebellum; ctx, cortex; fb, forebrain; hb, hindbrain. Scale bar = 2 mm. Adapted from McLaughlin et al. 2003a.
Благодаря успехам молекулярной биологии и расшифровке генома, можно предположить, что большинство «игроков» топографического «управления» известно, но их роли и взаимодействие пока не изучены. Странным является то, что в настоящее время единственными молекулами, удовлетворяющими критерию молекул топографического «управления», являются рецепторы и лиганды семейства Eph/ephrin, причем не только в ретинотектальной, но и в других системах, где было изучено развитие топографической карты. Некоторые молекулы, например, RGM (repulsive guidance molecule) и semaphorins, также в некоторой степени удовлетворяют описанным выше критериям. Разнообразие и сложность паттерна экспрессии Ephs и ephrins в пространстве и времени и особенности их функций обеспечивают широкие возможности в отношении их сигнальных свойств и, следовательно, в отношении их активности в качестве молекул-гидов. Регуляция mRNA, локализация белка и внутриклеточная интеграция ориентиров-гидов (ориентиров управления), так же как и влияние на рост субстрата освещены в обзорах (Bassell & Kelic 2004, Huber et al. 2003, van Horck et al. 2004).
Многие молекулы-гиды, включая Ephs и ephrins, имеют множественную, нередко противоположную активность, и выбор доминирующей активности в определенном контексте является критическим для развития топографической карты. Сигнальные пути, соединяющие guidance рецепторы с цитоскелетом и с молекулами cell-attachment (например, integrins), определяют функциональную активность (Davy & Robbins 2000, Gallo & Letourneau 2004, Nakamoto et al. 2004).
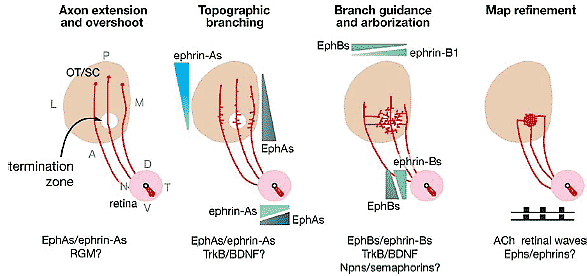
Figure 2 Mechanisms and molecules controlling retinotopic mapping in chicks and rodents. The names and/or distributions of molecules known, or potentially able, to control the dominant mechanisms at each stage are listed. The gradients represent the consensus distribution for a combination of related molecules (i.e., ephrin-A’s), which are not listed individually owing to distinctions in the individual members expressed and the precise distributions between species. Molecules other than those listed are likely to participate.
Успехи в изучении развития ретинотопической (retinotopic) карты доказали участие guidance molecules в картировании DV оси сетчатки вдоль LM оси OT/SC. Кроме того, идентифицирована роль молекул, участвующих в картировании TN оси сетчатки вдоль AP оси OT/SC. Благодаря компьютерным моделям и генетическим исследованиям ретинотопическая карта была значительно усовершенствована в последние годы.
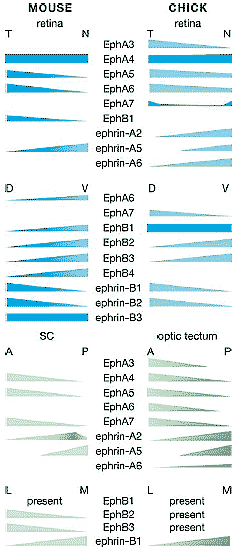
Figure 3 Expression of Ephs and ephrins in the retinocollicular/retinotectal systems of mouse and chick. The table represents our view of the consensus expression patterns for individual Ephs and ephrins in the retina (and likely RGCs) and OT/SC (in positions likely to affect mapping) during the primary molecular-dependent events in topographic map formation (about E15-P7 in mouse and E6-E14 in chick). The list may not be complete, and exclusion from this chart does not necessarily signify absence of expression in vivo. The sizes and shapes of the gradients are generalized, and relative expression levels should not be inferred (for an example of relative expression levels of Eph receptors see Reber et al. 2004).We have included only Eph family members with published expression domains determined by in situ hybridization with antisense riboprobes or specific antibodies in preparations where the listed pattern is evident at an appropriate age. A, anterior; D, dorsal; L, lateral; M, medial; N, nasal; P, posterior; present, receptor is expressed but the pattern is unclear; T, temporal; V, ventral.
Toward Discovering Graded Topographic Guidance Molecules
Механизмы, контролирующие развитие топографических карт, изучаются уже несколько десятилетий. Однако только в последнее время стала возможной идентификация молекулярного контроля этого процесса. Хемоаффинная (chemoaffinity гипотеза, предложенная Roger Sperry почти полвека назад, предсказала молекулярные механизмы развития топографических карт (1963). Sperry предположил, что молекулярные метки (tags) на проецирующихся аксонах и их клетках-мишенях детерминируют специфичность аксональных связей в нейральной карте. Более того, он предположил, что такие молекулярные метки могут распределяться по градиенту, который метит соответствующие участки в сенсорных структурах и в их мишенях. И хотя гипотеза Sperry базировалась на изучении регенерации ретинотектальной проекции у тритонов и лягушек, она определила направление исследований в области изучения развития топографических карт. Основной принцип гипотезы был довольно прост (Benson et al. 2001), однако последующие математические модели усовершенствовали гипотезу - добавились контрградиенты аттрактантов ( Gierer 1983) и градуированные репелленты (Gierer,1987), проведены более точные расчеты, особенно последовательных фаз сложного поведения RGC аксонов во время развития карты в OT/SC высших позвоночных (Yates et al. 2004).
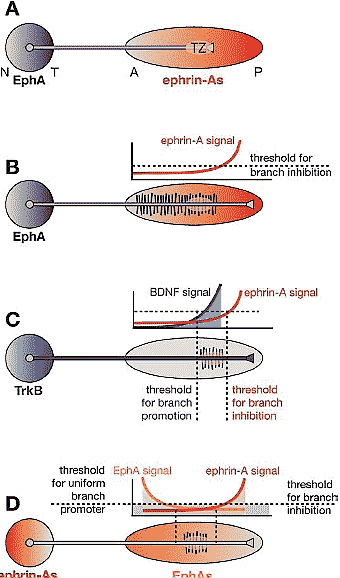
Figure 4 Requirement for two molecular activities to control branch distribution along the AP axis. (A) A gradient of repulsive guidance molecules, such as ephrin-As, is in principle sufficient to guide RGC axonal growth cones topographically to their appropriate TZ. This is the mapping mechanism in lower vertebrates. (B) However, a single repulsive gradient cannot result in the topographic branching observed in chicks and rodents. If the inhibitory signal allows for branching at the TZ, it must also allow for branching anterior to the TZ at the same, or higher, levels. This is not observed in vivo. (C) Gradients of molecules with branch-promoting activities could act in concert with inhibitory activities to result in branching restricted to the topographically appropriate domain. TrkB, in a similar distribution to EphAs in the retina (and/or if TrkB were graded along each RGC axon), and brain-derived neurotrophic factor (BDNF), in the OT/SC, have the appropriate activities to act with EphAs/ephrin-As in mapping. (D) Gradients of branch-inhibiting molecules in gradients opposite the EphA/ephrin-A gradients depicted in (A) could also result in topographic branching. Dual inhibitory gradients would require either a branching mechanism intrinsic to RGC axons or a branch-promoting molecule (blue shading) distributed to allow branching at the low point of the combined inhibitory gradients. The distributions of ephrin-As in the retina and EphAs in the OT/SC fulfill these requirements
В соответствии с хемоаффинной гипотезой каждая точка в OT/SC должна иметь уникальный молекулярный адрес, детерминированный градуированным распределением топографических молекул-гидов вдоль двух тектальных осей и таким же образом каждая RGS должна иметь уникальный профиль рецепторов для этих молекул, что приводило бы к зависимой от расположения и дифференциальной реакции на них RGC аксонов. В последующие десятилетия была изучена специфика проекций RGC аксонов к тектальным клеткам с использованием анализа проекций аксонов у нормальных животных и после экспериментальных манипуляций – сначала в регенерирующей ретинотектальной системе рыб и амфибий и позже, по мере развития высокоразрешающих методов исследования (мечение аксонов), - в развивающихся ретинотектальных/ретиноколликулярных проекциях. Эти исследования подтвердили основной догмат хемоаффинной гипотезы о том, что в установлении топографических проекций участвуют RGC аксоны, реагирующие на позиционную информацию в OT/SC. Изучение молекул, имеющих свойства, которые могли бы играть роль в картировании, выполнялось множеством лабораторий. Несколько поверхностно-клеточных молекул, таких как TRAP (McLoon 1991) и TOPAP (Savitt et al. 1995), с градуированным или ограниченным паттернами в сетчатке и/или в OT/SC были идентифицированы еще в середине 1990-х годов, но их функциональная роль оставалась неизвестной (обзор Roskies et al. 1995). Первое описание graded (ранжированных, градуированных) молекул, имеющих свойства топографических молекул-гидов, появилось только 10 лет назад в связи с клонированием двух родственных генов - ephrin-A2 (ранее Eph ligand family-1, или ELF-1) (Cheng et al. 1995, Cheng & Flanagan 1994) и ephrin-A5, описанный Bonhoeffer, Drescher, и соавт. (изначально названый repulsive axon guidance signal или RAGS) (Drescher et al. 1995). Оба они являются лигандами receptor tyrosine kinase EphA3 (ранее MEK4), экспрессируемые в градуированном паттерне вблизи RGCs (Cheng et al. 1995). Позже было показано, что ephrin-A2 и ephrin-A5 соответствуют функциональным критериям для graded топографических управляющих молекул, т.е. они имели дифференциальную реакцию на них RGC аксонов (ephrin-A2: Nakamoto et al. 1996; ephrin-A5: Drescher et al. 1995, Monschau et al. 1997), оказывали влияние на ретинотопическое картирование in vivo, и были необходимы для корректного развития карты (ephrin-A2: Nakamoto et al. 1996, Feldheim et al. 2000; ephrin-A5: Frisen et al. 1998).
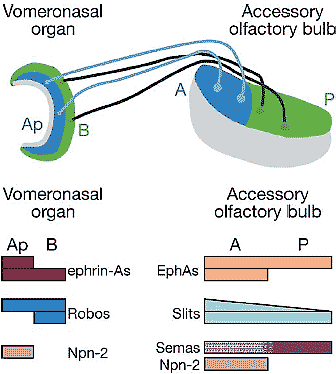
Figure 5 Representation of the accessory olfactory system in cross section. Apical (Ap, blue) vomeronasal (VNO) neurons (light blue) extend axons into the anterior (A, blue) accessory olfactory bulb (AOB) and form glomeruli. Basal (B, green) VNO neurons (dark green) extend axons into posterior (P, green) AOB and form glomeruli. This zonal topography is controlled by the guidance molecules charted below the illustration in representative form. Neuropilin-2 (Npn-2) in the AOB is present primarily in the anterior external plexiform layer of the AOB (gray) and may act to sequester secreted semaphorins (Semas), thus converting the uniform expression of secreted semaphorins in the AOB into a functional distribution of semaphorin protein (purple stippled area) highest in posterior AOB by reducing the availability of ligand in anterior AOB. Apical VNO neurons are guided to anterior AOB by attractive ephrin-A/EphA interactions and repellent Npn-2/sema interactions, whereas basal VNO neurons are guided to posterior AOB by repellent Robo/Slit interactions (Cloutier et al. 2002, 2004; Knoll et al. 2001, 2003;Walz et al. 2002). Other activities may play a role as well, and the described activities likely act in concert with pheromone receptors and other guidance cues (such as MHC class molecules; Loconto et al. 2003) to control the formation of glomeruli (Belluscio et al. 1999, Rodriguez et al. 1999). The gray area in the VNO represents the lumen.
До обнаружения ephrins большинство доказательств о существовании управляющих топографических молекул следовало из работ группы Bonhoeffer, проводимых in vitro –membrane stripe оценка и изучение последствий коллапса ростовых конусов. Используя оценку membrane stripe, авторы показали, что темпоральные аксоны RGС куриного эмбриона, имеющие выбор между ростом на альтернативных lanes передних и задних тектальных мембран, росли преимущественно на своих топографически подходящих anterior мембранах, тогда как назальные RGС аксоны не проявляли никаких преимуществ.
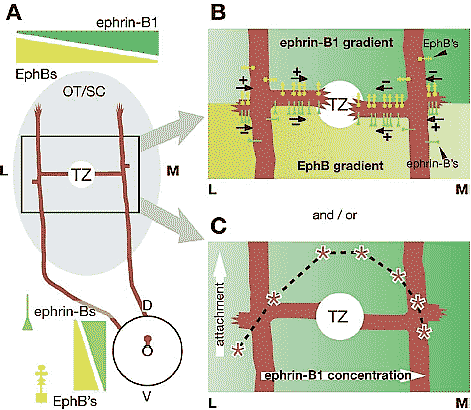
Figure 6 Potential bidirectional and bifunctional interactions resulting in topographic branch guidance along the LM axis of the OT/SC. (A) Representation of the projection from two RGCs in the same retinal location. One RGC axon has extended lateral (L) to the TZ and preferentially extends branches medially, toward the TZ. The other RGC axon has extended medial (M) to the TZ and preferentially extends branches laterally, toward the TZ. (B) EphBs and ephrin-Bs can cooperate via bifunctional and bidirectional signaling to guide branches appropriately both medially and laterally. The top half of this panel summarizes in vivo data demonstrating branch guidance by EphBs ( yellow icons) on RGC axons. Lateral to the TZ (left axon), EphB receptors encounter an ephrin-B1 level (green gradient) lower than that at their TZ (circle) and, at that relatively low level of ephrin-B1, branches are attracted up the ephrin-B1 gradient. Branches initially probing down the ephrin-B1 gradient from this axon have no incentive to do so and do not extend. Medial to the TZ (right axon), EphB receptors encounter a relatively high level of ephrin-B1 that repels branches down the ephrin-B1 gradient. The bottom half of this panel illustrates potential reverse signaling events. Lateral to the TZ, ephrin-Bs (green icons) on RGC axons encounter a higher level of EphBs ( yellow gradient) than that at the TZ and thus are repulsed down the gradient. Medial to the TZ, ephrin-Bs along RGC axons encounter a relatively low level of EphBs, and branches are attracted up the EphB gradient laterally toward the TZ. (C) Branches may also be guided to the TZ by forward signaling if EphBs act as ligand density sensors. Overlaid on the schematic are data from Huynh-do et al. (1999) showing EphB-expressing cell attachment ( y-axis) on substrates containing different concentrations of ephrin-B1 (x-axis). At low concentrations (i.e., lateral to the TZ), attachment is favored at a higher concentration (i.e., medial positions). At high concentrations (i.e., medial to the TZ) increased attachment is favored at a lower concentration (i.e., lateral positions). RGC axonal branches may be guided by a similar principle. The point of maximal attachment for each RGC axon is centered on the TZ and is dependent on its EphB concentration.
Одна из важных находок показала, что такое ростовое преимущество темпоральных аксонов не обусловлено аттрактантами или способствующей росту активностью, ассоциированной с передними тектальными мембранами, а обусловлено репеллентной активностью, ассоциированной с задними тектальными мембранами (Walter et al. 1987a,b). Задние тектальные мембраны также преимущественно вызывали коллапс ростовых конусов темпоральных аксонов – признак, который облегчал репеллентную активность к 33kDa - GPI-anchored белку (Cox et al. 1990, Stahl et al. 1990).
| EPH FAMILY OF RECEPTOR TYROSINE KINASES The Eph family of receptor tyrosine kinases (RTKs) is the largest known family of RTKs, composed of 14 Ephs and 8 ephrins in mouse and 15 Ephs and 9 ephrins in chick. Signaling through Ephs and ephrins has been implicated in a wide variety of processes including axon guidance, cell migration, vascular development, synapse development, structure and plasticity, and midline fusion events (Himanen & Nikolov 2003, Murai&Pasquale 2003, Palmer&Klein 2003, Poliakov et al. 2004, Rossant & Hirashima 2003, Surawska et al. 2004). Ephs and ephrins are separated into two subclasses on the basis of homology, the EphA/ephrin-As and EphB/ephrin-Bs, within which receptor-ligand binding and activation are promiscuous. In addition, some cross talk occurs between subclasses and may be functionally relevant in some systems. Ephs and ephrins are membrane bound, Ephs and ephrin-Bs are transmembrane proteins, and ephrin-As are GPI-linked to the plasma membrane, allowing for complex and precise patterns of expression within a single tissue or organ and between a projecting set of neurons and their targets (e.g., the retinocollicular/retinotectal projection; Figure 3). Eph-ephrin binding initiates signal transduction cascades by both Ephs and ephrins, resulting in bidirectional signaling. Additionally, Eph/ephrin signaling can be bifunctional, resulting in opposing responses (i.e., attraction or repulsion) to the same cues, depending on context (e.g., level of signaling). These features combine to make the Ephs and ephrins major players in many intricate problems of development, including the development of topographic maps described here. |
RGM имеет градуированный паттерн экспрессии в ОТ, сходный с ephrin-As, и инактивация RGM, используя CALI (chromophore assisted laser inactivation) метод привела к потере селективного репеллентного эффекта задних ОТ мембран на темпоральные RGS аксоны (Muller et al. 1996). Недавно клонированы куриные RGM, а рекомбинантные RGM, экспрессированные в 293Т клетках, имели репеллентный эффект на куриные RGC аксоны (Monnier et al. 2002). RGC аксоны, трансфицированные для экспрессии неогенина (neogenin) - anRGMreceptor, также отторгались RGM (Rajagopalan et al.2004). Предположили, что RGM играет роль в в ретинотопическом картировании. Однако мыши с таргетированной делецией RGMa не имели никаких дефектов топографического картирования в ретиноколликулярной проекции (Niederkofler et al. 2004), возможно из-за функциональной избыточности, например, других членов RGM семейства или ephrin-As.
MECHANISMS OF MAP FORMATION
Детерминация процессов, посредством которых RGCs образуют топографические связи, крайне важна для выявления роли graded guidance молекул в становлении карты и создания моделей этого процесса. Для решения этого вопроса исследования проводили, главным образом, на лягушках, рыбах, цыплятах, и грызунах – моделях развития ретинотопических карт у позвоночных. Эти виды различаются по развитию зрительной системы и ретинотопических карт, а также по абсолютным размерам OT/SC. Например, AT ось OT куриного эмбриона в 50 раз больше, чем у лягушек и рыб (РИС.1). Эти виды имеют и существенные различия в механизмах, управляющих RGC аксонами к мишеням их точных зон терминации (termination zone - TZ) и, следовательно, в функциях «управляющих» молекул в контроле топографического таргетирования RGC аксонов.
Развитие ретинотектальной топографии у цыплят (Nakamura & O’Leary 1989, Yates et al. 2001) и грызунов (Hindges et al. 2002; Simon & O’Leary 1992a,b,c) является многоэтапным процессом, включающим «выброс» (overshoot) аксонов и их промежуточное ветвление. Тщательный количественный анализ показал, что это единственный механизм образования карты и в настоящее время начала вырисовываться относительная важность направления роста аксонов, их ветвления и роль «управляющих» молекул в контроле создания топографических карт (Hindges et al. 2002, McLaughlin et al. 2003b, Yates et al. 2001). Авторы рассматривают различия в направлениях роста аксонов (первичных ростовых конусов) и их промежуточного ветвления у разных моделей – цыплят и грызунов и у лягушек и рыб. Подробности достижения аксонами своих TZ см. на РИС.1. Авторы отмечают важную роль интерстициального ветвления аксонов, специфичное вдоль AP оси – наиболее высокая степень ветвления найдена в участке АР, являющимся будущей зоной терминации (Yates et al. 2001). Промежуточные ветви располагаются перпендикулярно по отношению к первичному аксону и распространяются, главным образом, по LM оси в направлении их будущих TZ (Hindges et al. 2002, McLaughlin et al. 2003b, Nakamura et al. 1989). Такое ветвление является единственным способом с помощью которого RGCs формируют постоянные упорядоченные синаптические связи (Yates et al. 2001).
Далее авторы рассматривают распространение аксонов во время AP картирования, механизмы височно-назального ретинотопического картирования с участием АР-специфического промежуточного ветвления, АР градиенты (параллельные и оппозитные) промотеров и результаты подавления ветвления аксонов, роль EphBs и Ephrin-Bs в молекулярном контроле DV и LM картирования у разных моделей, роль и механизмы «управляющих» молекул в росте первичных аксонов и промежуточном ветвлении, видоспецифические различия в развитии топографических карт у разных моделей, роль Eph семейства receptor tyrosine kinases (RTKs), идентификацию дополнительных генов-кандидатов, участвующих в ретинотопическом картировании, обсуждаются перспективы развития этого направления исследований.
Источник:
Последнее редактирование: 13.03.2017
Чтобы оставить комментарии нужно авторизоваться.