Данная статья во многом разъясняет идеологию моих автопубликаций в той их части, которая посвящена наиболее ранним этапам возникновения жизни.
Нередко в научной литературе и в учебниках можно встретить утверждение, что все теории возникновения жизни можно свести к двум категориям - "сперва метаболизм" [Eschenmoser, A. 2007; Fernando, C., Rowe, J. 2007; Kaufman S.A. 1993; Lindahl P.A. 2004; Smith E., Morowitz H.G. 2004; Wächtershäuser G. 1988a, 1992, 1997] и "сперва репликация" [Delaye L., Lazcano A. 2005; Joyce G.F. 2002; Lazcano A., Miller S.L. 1999; Orgel L.E. 2004; Robertson A. et al. 2000].
Согласно одной группе теорий предполагается, что вначале "случайно" образовался репликатор – крупная полимерная молекула, способная к самовоспроизведению (например, РНК), которая и обеспечила существование ключевых признаков живых систем (ЖС) – наличие биологической памяти, способность к воспроизведению и дарвиновской эволюции. Данная модель основана на предположении, требующем довольно сильного допущения: воспроизведение было возможно благодаря тому, что в начальных условиях формирования жизни в окружающей среде существовал богатый органическими соединениями источник энергии и строительного материала для первых ЖС, того, что условно называется первичным бульоном. Часто допущение первичного бульона подразумевает, что в существовавших тогда анаэробных условиях источником энергии для таких первых систем должны были служить процессы брожения (ферментации) [De Duve C. 2002, 2005, см. также статью В.П. Скулачёва "Эволюция биологических механизмов запасания энергии"] и субстратного фосфорилирования, которые давали универсальную энергетическую валюту - АТP.
Брожение - это окислительно-восстановительные превращения органических веществ в анаэробных условиях, сопровождающиеся выходом энергии; все окислительно-восстановительные реакции в нём протекает, если можно так выразиться за счёт "внутренних" возможностей - он связан с такими перестройками органических соединений, которые связаны с перераспределением степени окисления углерода, то есть неким диспропорционированием степени окисления. Действительно, целый ряд фактов говорит в пользу того, что брожение по-видимому уже было присуще последнему универсальному общему предку (LUCA) на стадии прогенота (см. Древний фермент подтверждает гипотезу о зарождении жизни в горячих источниках). Однако LUCA - это далеко не самый примитивный возможный организм. Например, для него уже был присущ кодируемый синтез белков, который явно не был присущ для первых ЖС. Важно, что процесс гликолитического брожения (гликолиза), не смотря на то, что он может протекать просто в растворе, требует по крайней мере 11-12 сложных белковых ферментов. В настоящее время нет никаких данных, которые говорили бы в пользу того, что абиогенные аналоги данных ферментов могли бы присутствовать на самых ранних стадиях рождения и эволюции ЖС до формирования кодируемого пептидного синтеза, а структуры этих ферментов у двух главных прокариотных ветвей - архей и бактерий - существенно отличаются [Siebers B, Schönheit P. 2005]. Этот факт можно интерпретировать как независимое изобретение гликолиза на относительно поздних стадиях эволюции ЖС - непосредственно перед или после начала дивергенции.
Таким образом, в настоящее время нет каких-то даже очень косвенных данных, которые бы позволяли бы рассматривать процесс брожения не смотря на свою явную древность, как стоящий у истоков энергетики первых живых систем.
В настоящее время большинство исследователей биогенеза приняло эту точку зрения первичности репликатора в виде более конкретной гипотезы РНК мира, который существовал ещё до возникновения ДНК и кодируемого белкового синтеза.
Согласно другим теориям, сначала появился метаболизм, в процессе которого малые неполимерные или олигомерные (то есть состоящие всего из нескольких мономеров) молекулы образовали сеть химических реакций. В настоящее время трудно отрицать первичность РНКовой жизни по отношению к ДНКовой (см. Самое обоснованное утверждение, касающееся происхождения жизни- первичность молекул РНК по отношению к ДНК и сложным белкам). Но все полученные до сих пор данные свидетельствуют лишь о том, что РНК возникла раньше, чем ДНК и кодируемые ими белки. Некоторые положения мира РНК можно поставить под сомнение. Если представить, что первые ЖС были основаны преимущественно на полимерных (то есть что-то вроде порядка десятка и более молекул) РНК, которые были одновременно и ключевыми катализаторами, и наследственными матрицами, то необходимо представить одновременно и существование очень сложного метаболизма, связанного с синтезом компонентов этих РНК, который в отсутствие белков должен был катализироваться каким-то весьма разнообразным множеством ферментов. Тогда совершенно неясным остаётся вопрос о синтезе этого строительного материала. Если синтез строительного материала для РНК не контролировался самими РНК, а возник абиогенно, то есть в довольно богатом органикой первичном бульоне, то совершенно непонятным остаётся проблема накопления рибонуклеотидов в этом бульоне. Хотя такая точка зрения рассматривается и изучается всерьёз, и в последние годы достигнуты определённые успехи в экспериментальном моделировании синтеза компонентов РНК-молекул (см. Химики перодолели главное препятствие на пути к абиогенному синтезу РНК), в проблеме их геохимического накопления до сих пор не проглядываются перспективы их решения (систематическое изложение основных трудностей возникновения мира РНК см. в [Спирин А.С. 2007]). Таким образом, встаёт крайне сложная проблема снабжения первых ЖС таким сложным строительным материалом, как хиральные РНК-нуклеотиды. Непосредственный синтез биологических полимеров на основе имеющихся экспериментальных данных по моделированию абиогенного синтеза выглядит геофизически нереалистичным [Shapiro, R. 1987; Horgan J. 1991]. Критической трудностью возникновения такого богатого сложными органическими веществами первичного бульона является не только неизбежность накопления рацематов, но и стабильность компонентов РНК, которые должны были бы накапливаться в достаточно высокой концентрации, запас которой к тому же мог бы быстро истощился при репликации РНК. Проблема усугубляется также тем, что УФ облучение, необходимое для синтеза компонентов РНК, одновременно разрушала бы основную часть вновь синтезируемых компонентов. Следует также отметить так называемую концентрационную проблему, подчёркнутую Де Дювом [De Duve C. 1991].
Критическими для модели первичности полимерного гена также являются многочисленные экспериментальные неудачи в создании РНК-кодируемого матричного синтеза РНК и, что очень важно, его отсутствие в современных ЖС. даже у РНК-вирусов. Действительно, в экспериментальных условиях получены некоторые матрично полимеризующие молекулы РНК (Рибозимы могут размножать друг друга), такого рода репликация является не только очень неточной, но и, что также существенно, неполной.
Кроме того, как будет аргументировано в ряде следующих статей, РНК должны были быть далеко не единственными ключевыми катализаторами.
Стоит отметить, что сторонники обоих точек зрения подвергли друг друга довольно суровой практически бескомпромиссной критике [Bada J.L., Lazcano A. 2002; Borgeson W. et al. 2002; Orgel L.E. 2008].
В рамках гетеротрофной модели происхождения репликатора одной из серьёзнейших проблем остаётся проблема источников энергии. Во всех современных формах жизни, и предположитльно у последнего общего предка всех живых организмов (LUCA, last univerasal common ancestor) поток энергии означает переход их в универсальные выскоэнергетические связи: тиоэфирные связи в Ацетил-СоА и ангидридные связи в ацетил-фосфате и АТР. Освобождение энергии такого рода связей должно обеспечивать синтез полимерного репликатора. Сам бульон должен был находиться в состоянии, близком к термодинамическому равновесию и с небольшой разницей окислительно-восстановительных потенциалов составляющих его молекул. Поэтому сам этот "суп" не мог быть источником энергии и необходимо допускать существование внешнего источника энергии.
Один из вариантов решения этой проблемы был предложен В.П. Скулачёвым - вариант абиогенного синтеза АТP с помощью ультрафиолетового облучения ("ультрафиолетовый фотосинтез"), описанный в статье Эволюция биологических механизмов запасания энергии. Не смотря на красоту и предсказательную силу данной модели, против неё говорит даже не только отсутствие экспериментальных данных, подтверждающих данную модель, но и также отсутствие сходных механизмов в современных экосистемах.
Если ни ДНК, ни РНК, ни белки, ни другие крупные и сложные молекулы не могли находиться у истоков жизни, то альтернативой является предположение, что первоосновой были небольшие органические молекулы. Первичность метаболизма подразумевает, что жизнь возникла из автокаталитических (продукты реакционного цикла увеличивают скорость реакций этого цикла) самоорганизующихся химических циклов [Shapiro R. 2006]. Эти циклы привели к увеличению концентрации одних веществ по сравнению с другими.
Для того, чтобы понять логику дальнейших рассуждений, необходимо чётко понимать, что без наследственной информации дарвиновская эволюция невозможна.
Не смотря на отсутствие информационных полимерных молекул, такие системы могли поддерживать определённые химические циклы, параметры которых могли служить наследственной информацией. Такого рода информация, косвенно поддерживающая гомеостаз, получило название "конформационный геном". Конформационный геном отличается от генома в обычном его понимании тем, что в обычном геноме наследственная информация записана на молекулах, представляющих собой полимеры (ДНК, РНК), определяющейся первичной структурой молекул (то есть последовательностью мономеров), тогда как в конформационном геноме информация записана на уровне концентраций альтернативных конформаций катализаторов и концентраций всех веществ, входящих в состав автокаталитических циклов.
Такого рода конформационные геномы по-видимому являются существенной частью современных организмов и ключевой основой для эволюционного усложнения видов (см. в качестве обзора "Кодирование и эволюция сложности биологической организации")
Уточним для определённости несколько терминов. С определённой степенью идеализации можно сказать, что для существенной части наследственной информации в современных организмах "мономеры ДНК, РНК и белков могут рассматриваться как символы базового алфавита, а макромолекулы задаются последовательностями символов, или генетическими текстами" (см. Ратнер В.А. 2002). Системы взаимодействующих генов характеризуются схемами молекулярных взаимодействий, называемых генетическими сетями, геномы - последовательностями символов генов, знаков пунктуации и управления [Ратнер В.А. 2002].
Немаловажным свойством генетических текстов является их определённая так называемая контекстная независимость. Это важнейшее свойство генетических текстов связано с тем, что кодирование одним мономером (или строго определённой группы мономеров), как правидо (с определёнными исключениями), не зависит от присутствия других мономеров (или групп мономеров). Фактически это означает, что, например, при репликации ДНК аденин одной цепи кодирует только тимин другой цепи, а гуанин - цитозин. При этом наличие соседних нуклеотидов в подавляющем большинстве случаев является несущественным.
Контекстную независимость мы можем усмотреть и при кодировании последовательностью РНК определённых белков. Кодирование триплетом в некотором приближении (идеализации) на РНК не зависит от наличия других триплетов на этой же последовательности.
Данное допущение в действительности является определённой идеализацией. Например, кодирование селеноцистеина на мРНК является существенно контекстно зависимым, так как определяется не только данным триплетом, но и наличием определённой дополнительной последовательности, расположенной вблизи данного триплета (см. подробнее Свойства генетического кода - след его возникновения).
Не всегда и последовательности нуклеотидов однозначно определяют последовательность синтезируемой на них как на матрице цепи. Так, хорошо известен контекстно-зависимый эффект теломеры, когда краевые нуклеотиды матрицы не способны кодировать их синтез при матричном воспроизведении хромосом.
Тем не менее, определённая контекстная независимость генетических текстов - существенная их характеристика.
Компоненты конформационного генома являются существенно контекстно зависимыми. Так, даже функциональные свойства полимерных молекул (в частности, белков и РНК) зависят от замен мономеров, которые влияют на функциональные свойства других мономеров - то есть имеют контекстно зависимый характер.
Как полимерный, так и конформационный геном, мы можем рассматривать с информационной точки зрения: говорить об их условном размере (количестве мономеров, количестве молекулярных компонентов и др.) и помехоустойчивости.
Рассмотрим подробнее, какие ещё существенные аргументы делают предпочтительным первичность именно конфомационного генома, а не полимерного.
Немаловажным аргументом в пользу наличия автокаталитических метаболических циклов является проблема хиральной чистоты компонентов для биосинтеза. Дело в том, что в автокаталитических системах хиральная чистота могла уже поддерживаться автоматически. Конкуренция между автокаталитическими циклами с разными хиральными аналогами компонентов этой системы могла привести к сохранению только одного из вариантов энантиомеров как компонентов метаболизма. Оптическая чистота древних автокаталитических систем – один из важнейших компонентов конформационного генома.
Если же принять, что полимерная молекула, представлявшая собой древний ген, строилась и готовых синтезированных абиогенно веществ, то возникает трудность в объяснении использования оптических изомеров из хиральных смесей.
Свойство автокаталитичности обеспечивает воспроизводство химических циклов. Сами циклы могли расширяться и образовывать гиперциклы. Как было показано М. Эйгеном с соавторами [Эйген М., Шустер. П. 1973] гиперцикличность организации позволяет существенно увеличить ёмкость генома (не важно конформационного или полимерного), благодаря чему возможно дальнейшее адаптивное усложнение ЖС.
Чтобы можно было представить эволюцию таких циклов, нередко предполагается определённая физическая изоляция этих автокаталитических систем. Благодаря такой изоляции становился возможным отбор этих циклов, давая начало эволюции таких автокаталитических систем. Представления о таких изолирующих компонентах носят во многом умозрительный характер, и такого рода модели нельзя назвать детально разработанными, что, однако не отрицает их возможное отражение реальных событий.
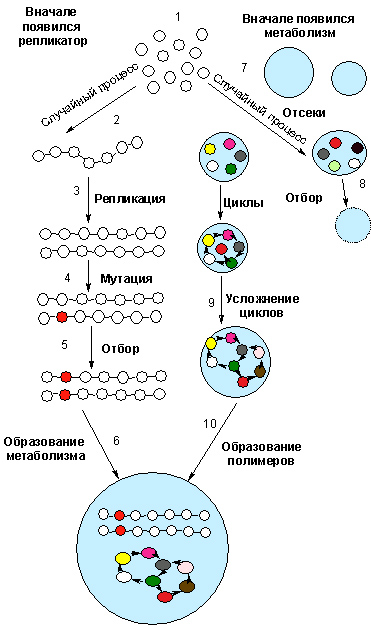
Рис.1. Происхождение жизни с точки зрения первичности репликатора (слева) или метаболизма (справа). Все научные теории происхождения жизни относятся к одной из двух категорий, утверждающих первичность репликатора или метаболизма. В обоих случаях все начинается с малых молекул (шарики с символами), образующихся абиогенным путем (1). Согласно модели «вначале был репликатор», некоторые молекулы случайно соединяются друг с другом в цепочку (возможно, прототип РНК), способную к самовоспроизведению (2). В отдельных копиях репликатора возникают мутации (3); мутантные копии тоже могут самовоспроизводиться (4). Те из них, которые лучше приспособлены к существующим условиям, вытесняют предшествующие версии (5). В конце концов, образуется некий отсек (протоклетка), в котором сформированы и метаболизм, и наследственная матрица (6). Идея рисунка взята из [Шапиро Р. 2007].
В рамках модели «вначале был метаболизм» все начинается со спонтанного образования отсеков (7). В некоторых из них содержится группа молекул, которые вступают в химические реакции, образующие циклы (8). Со временем циклы все более усложняются (9), и наконец в системе появляются полимерные молекулы – хранилища информации.
Для того чтобы набор малых абиотических молекул перешел в категорию живых систем Роберт Шапиро выделил 5 необходимых условий [Шапиро Р. 2007]:
1. Должна существовать граница раздела между локальной областью и окружением. Граница раздела необходима для осуществления эволюции отдельных популяций молекул. Природа таких перегородок неизвестна. Так, Дэвид Дример с соавторами обнаружили в составе упавших на Землю метеоритов мембраноподобные структуры [Deamer D. et al. 2002]. Возможно, перегородками служили мембраны из сульфида железа, поверхности каких-либо широко распространенных минералов (на них могли избирательно адсорбироваться заряженные молекулы) [Russell M.J. et al. 1993], аэрозоли и т.д.
2. Требуется источник энергии, в котором протекают процессы с высвобождением тепла (рис. 1 (2)). Энергия получается в ходе окислительно-восстановительных реакций. Одним из источников таких энергий могут служить минералы, которые многие микроорганизмы могут использовать в качестве источника энергии. Как будет описано ниже, в качестве этого "минерала" мог выступать древний аналог восстановительного цикла трикарбоновых кислот (ВЦТК) или фиксация через ацетил-коэнзим А путь, или же первые метаболические сети были представлены различными конкурирующими реакциями с различными вариантами фиксации углерода, составившие в будущем различные древнейшие варианты фиксации углерода (подробнее об этом см. Как мог выглядеть архаичный метаболизм?)
3. Энергия должна использоваться для запуска «движущей» химической реакции (рис.1 (3)). Сопряжение между процессами генерации энергии и ее потреблением осуществляется при участии общего для них промежуточного продукта и становится еще более эффективным в присутствии фермента. Одно из допущений теории возникновения жизни из малых молекул состоит в том, что сопряженных реакций и примитивных катализаторов достаточно для зарождения жизни.
4. Сеть химических превращений должна быть способна приспосабливаться к изменяющимся условиям и эволюционировать (рис.1 (4)). Адаптация химических реакций к окружающим условиям невозможна без цепи альтернативных путей.
"Помню, в детстве я завороженно следил, как вода из прохудившегося шланга, стекая под уклон, продвигается к ближайшему сточному колодцу. Если возникала преграда, струя терпеливо искала обходные маршруты. Точно так же, если колебание кислотности среды или какие-то другие обстоятельства «перекроют дорогу» от В к А, то поток вещества будет изменять направление, пока не отыщется другой путь к цели. Последовательно вводя аналогичные изменения, мы получим не замкнутый цикл, а сеть. Такое освоение химического ландшафта методом проб и ошибок может привести к появлению веществ, катализирующих важные этапы цикла, благодаря чему повысится эффективность использования энергии" [Шапиро Р. 2007].
5. Сеть должна разрастаться и воспроизводиться. Для сохранения и роста сети необходимо, чтобы вещества входили в сеть быстрее, чем выходить из нее, а отсеки – обладать способностью к делению (воспроизводству) (рис.1(5)). Должен был возникнуть механизм репродукции. Предположим, что система находилась в какой-либо полости внутри горной породы. Расширяясь, она занимала соседние пространства. Такое распределение по отдельным отсекам предотвращало полное исчезновение системы в целом под влиянием каких-то локальных деструктивных событий. Независимые подсистемы могли эволюционировать каждая по-своему и конкурировать за сырье. Дальше должен был произойти переход от жизни, возникшей абиогенным путем при участии какого-то имевшегося в те давние времена источника энергии, к форме, приспосабливающейся к окружению в соответствии с дарвиновскими законами эволюции.
В мире типа «сперва метаболизм», прежде чем генетические полимерные молекулы стали определять направление эволюции, отбор должен был направлять движение не в сторону лучшего репликатора, а в сторону химических реакций, наиболее эффективно создающих энергетический обмен, не позволяя энергии рассеиваться в побочных направлениях.
Таким образом, предположение о первичности репликатора, как правило подразумевает предположение о наличии богатого органическими веществами первичного бульона, поставляющего строительный материал и энергию для ЖС. Основой энергетического метаболизма в этом случае часто предлагается такой источник энергии, как брожение [Lazcano A., Miller S.L. 1999].
В случае первичности метаболизма предполагается отсутствие богатого органикой первичного бульона, делающего брожение невозможным. В этом случае источником энергии могут служить богатые энергией вещества, образованные в горячих источниках (CO, FeS/FeS2, H2 и др.), а также возможно и при облучении светом, которые участвуют в фиксации СО2 (подробнее об этом см. "Автотрофное термофильное происхождение жизни")
Следует, однако, отметить, что между вариантами «сперва метаболизм» и «сперва ген» нет чётко очерченной границы. Нельзя рассматривать оба этих варианта как абсолютно альтернативные, поскольку существуют и промежуточные варианты. Даже если представить, что первыми были автокаталитические системы, представленные малыми неполимерными молекулами, необходимо искать путь создания полимерных наследственных молекул, которые взяли бы под контроль метаболизм, то есть предопределяли бы существенные для выживания параметры автокаталитических систем.
Синтез таких полимерных наследственных молекул мог начаться только с олигомерных вариантов, то есть с молекул, состоящих только из небольшого числа мономеров: собственно мономеров, димеров, тримеров и других олигомеров. При таком рассмотрении фактически стирается разница между конформационным и классическим геномом. При таком подходе олигомеры могут рассматриваться как примитивные аналоги полимерного генома, в которых, однако немаловажную роль играет и контекст соседних нуклеотидов.
Какие молекулы могли бы служить кандидатами на роль молекул, занимающих промежуточное положение между классическим полимерным и конформационным геномом.
Можно предположить, что эти молекулы следует искать в первую очередь среди универсальных компонентов современного метаболизма.
К таким универсальным компонентам современного метаболизма относятся всевозможные нуклеотидные кофакторы: ATP, GTP, CTP, UTP, ITP (инозинтрифосфат), CoA, NAD, NADP, FAD, SAM и другие. Не обязательно, чтобы все из перечисленных нуклеотидных кофакторов могли присутствовать в условиях первобытной Земли, однако это могли быть первые молекулы, взявшие под контроль (то есть, другими словами, составлявшими значительную часть конформационного генома) первые автокаталитические циклические химические реакции, сформировавшие метаболизм первых ЖС.
Эволюция такого рода конформационных геномов происходила не только в направлении синтеза новых мономерных компонентов, но и, что важно, в направлении создания компонентов конформационного генома, представленного димерами, тримерами и другими олигомерами, образованными из нуклеотидных предшественников. Фактически переход к полимерному геному (оцифровка генома) происходил с множеством промежуточных вариантов.
Эти олигомеры не обязательно могли быть представлены каноническими нуклеотидами, но и могли быть представлены нуклеотидами с неканоническими основаниями или связанными неканоническими связями.
Это означает, что в древнем РНК-мире и РНК-белковом мире в действительности существенная для выживания ЖС часть наследственной информации всё ещё могла быть закодирована в конформационной части генома.
Немаловажным моментом, связанным с включением рибонуклеотидов в метаболизм, является формирование уже практически современной формы древнейших метаболических компонентов промежуточного метаболизма – предположительно восстановительного цикла трикарбоновых кислот и (или) ацетилкоэнзимного пути, поскольку для протекания этих циклов рибонуклеотидные кофакторы являются абсолютно необходимы. Как могли выглядеть архаичные предшественники этих циклов, обсуждается в в статье Как мог выглядеть архаичный метаболизм?.
Резюмирующая схема начальных этапов происхождения жизни.
1. Возникновение первых циклических автокаталитических циклов реакций, один из которых обязательно должен был быть восстановительным связан с фиксацией углекислого газа и запасанием энергии. Остаётся под вопросом, какой именно способ фиксации углерода являлся первичным в первых каталитических циклах (см. Как мог выглядеть архаичный метаболизм?). Однако весьма правдоподобным выглядит предположение о том, что восстановительный цикл трикарбоновых кислот (ВЦТК), точнее его предковая форма без использования нуклеотидных кофакторов должен был возникнуть уже на наиболее ранних этапах начала эволюции первых ЖС ещё до формирования мира полимерных РНК и тем более кодируемого пептидного синтеза. Это утверждение основано на том, что данный цикл является необходимой частью, поставляющей строительный материал и энергию для дальнейшегно синтеза (абиогенного или же контролируемого конформационным геномом) аминокилот и рибонуклеотидов (см. "Реликтовые составляющие современного метаболизма. С чего он мог начаться?").
Кроме того, данный цикл имеет общие части с рядом других известных циклов фиксации углерода (Как мог выглядеть архаичный метаболизм?), что говорит, во-первых, об общности происхождения различных автотрофных метаболических циклов, а во-вторых, о возможности формирования различными автотрофными циклами метаболической сети, обладающей большей устойчивостью, чем отдельный циклы, в рамках которой возможна конкуренция и отбор различных элементов этой сети.
Кроме ВЦТК в качестве компонентов первых метаболических сетей также рассматривается прообраз другого безусловно древнейшего способа фиксации углерода - по ацетил-коэнзим А -пути (пути Вуда-Люнгдала) [Martin W., Russell M.J. 2003] или же других способов, рассмотренных подробнее в статье Как мог выглядеть архаичный метаболизм?.
Однако существенно, что в современных метаболических путях синтез аминокислот и в дальнейшем компонентов РНК протекает именно с участием компонентов цикла трикарбоновых кислот, что указывает на принципиальную важность именно древнего аналога ВЦТК и его возникновение до или одновременно с биосинтезом аминокислот и компонентов РНК.
Немаловажным обстоятельством также является тот факт, что непременными компонентами различных типов фиксации углерода является вещество рибонуклеотидной природа - как минимум, ацетил-коэнзим А (в ацетил-коэнзим А-пути), а в других известных способах фиксации углерода - также NAD, ATP, FAD и FADP, которые, как предполагается ниже, должны были возникнуть позднее, уже на основе первичных циклов фиксации углерода и обеспечиваться существованием более продвинутых автокаталитических рибонуклеотидных циклов.
Поэтому можно говорить, что на данном этапе не существовали современные варианты путей фиксации углерода, а только похожие на них архаичные аналоги, которые обеспечили возможность биосинтеза аминокислот, и, что особенно важно, нуклеотидов. Только появление нуклеотидов, как ключевых кофакторов биосинтеза, позволило сформировать метаболические пути, уже очень похожие на современные.
2. Ключевую роль в начале дарвиновской эволюции первых ЖС сыграли первые железо-серные компартменты, обеспечивавшие изоляцию конкурирующих метаболических циклов (сети), локальное концентрирование метаболитов и образование первой универсальной энергетической валюты - протонного градиента (об этом см. подробнее Автотрофное термофильное происхождение жизни).
3. Различные метаболические циклы (сети) конкурировали между собой за ресурсы, обеспечиваемые предположительно флюидами в щелочных гидротермалях. Основные вещества абиогенной природы, служащие ресурсами, за которые конкурировали различные метаболические сети были: CH4, HCHO, CH3OH, CH3COO-, HS-, CH3CH2S-, NH3, CN-, а также возможно простейшие абиогенно синтезируемые аминокислоты, в первую очередь глицин и аланин.
4. Первыми катализаторами, запустившими архаичные метаболические реакции, были железо-серные минералы (подробнее об этом см. Минеральные корни современных ферментов), катализировавшие ключевые для первых метаболических циклов окислительно-восстановительные реакции и обеспечивавшие создание протонного градиента как первой универсальной энергетической валюты в минеральных железо-серных компартментах (об этом см. подробнее Автотрофное термофильное происхождение жизни).
Конкурирующие за общие ресурсы (молекулярный водород, аммиак, метан и др. вещества) различные метаболические циклы вырабатывали различные интермедиаты, часть из которых становилась лигандами (например, тиолы, аминокислоты, короткие пептиды и др.) для минеральных железо-серных, жилезо-никелево-серных и других катализаторов. От этих лигандов существенно зависела эффективность катализа метаболических реакций, а, значит, и исход конкуренции между альтернативными метаболическими циклами.
Таким образом, уже на самых первых этапах возникновения ЖС существенную роль для начавшейся дарвиновской эволюции конкурирующих метаболических циклов играла обратная положительная связь между эффективностью катализа железо-серными минералами и лигандами, связывавших эти минералы и обеспечивавшими образование протоферредоксинов, (де)гидрогеназ и других катализаторов.
5. Наряду со способами фиксации углерода, дающими строительный материал и источники энергии, должны были функционировать, и другие составляющие сети автокаталитических циклов, представленные последовательными превращениями рибонуклеотидов и их производных, компоненты которых и представляли собой ключевую часть конформационного генома. Конкретными материальными носителями конформационного генома могли быть ATP, GTP, CTP, UTP, ITP (инозинтрифосфат), CoA, NAD, NADP, FAD, SAM и другие нуклеотидные кофакторы современного и древнего метаболизма (подробнее будет изложено в отдельной статье).
Система «рибонуклеотидные химические автокаталитические сети», включавшая древний прообраз восстановительного цикла трикарбоновых кислот, как раз и представляла собой древнейшую живую систему, способную к эволюции. Это - одновременно наиболее спорное и важное положение в представленном изложении. Причина - в настоящее время, не смотря на бурный рост фактов, касающихся РНК-метаболизма и РНК-химии, мы не можем построить достаточно полную правдоподобную картину возникновения первых рибонуклеотидных каталитических циклов.
Тем не менее, данное предположение выглядит привлекательным прежде всего потому, что оно подразумевает существенно большую структурную простоту компонентов этих циклов по сравнению с канонической гипотезой РНК-мира, где ключевыми должны были быть именно полимерные молекулы. Фактически согласно данному предположению нуклеотидные кофакторы не просто являлись важными компонентами мира РНК, а были частью его конформационного генома. На данной стадии развития не имеет смысла различать ферменты и их кофакторы, поскольку существенная часть ферментативной активности должна была быть представленной малыми молекулами рибонуклеиновой природы. Эти вещества должны были контролировать собственный биосинтез (на данной стадии уже уместно говорить именно о биологическом синтезе, поскольку уже предполагаются эволюционные изменения конформационного генома). Для биосинтеза этих кофакторов предположительно могли использоваться компоненты ВЦТК.
Подборка фактов, которые могут детализировать данное предположение будет в отдельной статье.
6. Не смотря на отрицание роли первичного бульона как среды, исключительно богатой строительным материалом для ЖС, исключать абиогенное образование сложных органических веществ на самых ранних этапах бессмысленно (без них невозможно было бы возникновение дарвиновски эволюционирующих автокаталитических циклов). Поэтому предполагается, что на ранних этапах существенно важным был абиогенный синтез отдельных компонентов РНК - азотистых оснований, нуклеозидов и нуклеотидов. В свете того, что только в последние годы был получен ряд сущетвенно важных фактов, делающих более правдоподобным абиогенный синтез компонентов РНК (см. "Химики преодолели главное препятствие на пути к абиогенному синтезу РНК", "Химикам удалось стабилизировать абиогенный синтез сахаров"), можно ожидать в дальнейшем новых экспериментальных данных, которые дадут возможность детализировать данную модель или же опровергнуть её.
7. Дальнейшее направление эволюции свелось не только и не столько к разветвлению автокаталитических сетей, но и, в первую очередь, полимеризации нуклеотидных компонентов конформационного генома. Конформационный геном стал всё больше предопределяться не наличием тех или иных мономерных или димерных компонентов конформационного генома, но в первую очередь последовательностью рибонуклеотидов в олигонуклеотидных последовательностях. Это явление уместно назвать оцифровкой генома.
Оцифровка генома – это переход кодирования наследственной информации от физико-химических параметров автокаталитических сетей к более узкому параметру – первичной структуре олиго- и полимерных молекул, в котором информация становится существенно контекстно-независимой. Контекстная независимость на ранних стадиях могла быть применима только к олиго- и в дальнейшем к полинуклеотидным последовательностям, структура которых могла предопределяться преимущественно конкретными нуклеотидами первичной структурой родительских матриц.
Первыми этапами оцифровки генома являлось образование димерных и олигомерных молекул, представленных рибонуклеотидами. Фактически значительная часть нуклеотидных кофакторов как раз и представлена димерными олигонуклеотидами: NAD, NADP, FAD и многие другие (подробности в одной из следующих статей).
Оцифоровка генома на данном этапе фактически означает возникновение матричного синтеза РНК-олигонуклеотидов, в котором впервые синтез олигонуклеотидов был осуществлён не косвенно на автокаталитических циклах, а непосредственно с участием родительской матрицы. Обычно олигонуклеотидами весьма условно называют последовательности нуклеиновых кислот длиной до 40-50 нуклеотидов. Мы будем подразумевать под этим термином малое, но неопределённое число мономеров в рибонуклеотидной последовательности, в которой не обязательно будут только канонические основания.
8. В автокаталитических гиперциклах существенную роль играли не только нуклеотидные кофакторы и компоненты ВЦТК, но и также аминокислоты, которые на начальных этапах формирования первых живых систем могли быть доступны из абиогенного источника, но на этапе динуклеотидной жизни уже частично синтезировались на основе компонентов ВЦТК (подробнее об этом см. "Древний дублетный генетический код был предопределён путями синтеза аминокислот").
Сильным аргументом в пользу этого утверждения является объяснение данной моделью связи свойств аминокислот с центральным нуклеотидом кодирующего их триплета, и одновременно связь первого нуклеотида кодона с путями биосинтеза соответствующих аминокислот.
Немаловажным доводом в пользу этого предположения является также то, что модель древнего синтеза аминокислот, представленная в статье о древнем дублетном коде решает также проблему древнего специфического аминоацилирования древних предковых тРНК.
Согласно наиболее распространённой точке зрения древние тРНК должны были специфически аминоацилироваться рибозимами. Однако против этого предположения говорит, во-первых, отсутствие каких-либо следов этих рибозимов в современных организмах, а, во-вторых, все искуственно синтезирванные РНК, проявляющие аминоацилирующую активность, являются исключительно низкоспецифическими ферментами и способны использовать лишь активированные формы аминокислот [Lee N. et. al. 2000].
В этой связи хорошей альтернативой данной модели является предположение о том, что большинство аминокислот на ранних этапах входили в состав аминоацил-РНК не в результате специфического аминоацилирования, а в результате специфического их синтеза на РНК.
Данное предположение выглядит ещё более правдоподобным, если учесть, что в определённой мере сходная картина биосинтеза аминокислот, аминоацилированных на РНК, уже встречается у современных организмов. Так, биосинтез глутамина и аспарагина может происходить у ряда организмов именно на тРНК из соответствющих предшественников, которые перед этим аминоацилировали тРНК - глутамата и аспартата. Кроме того, синтез селеноцистеина по-видимому у всех организмов происходит на тРНК, которая перед этим аминоацилируется серином, служащим основой для синтеза селеноцистеина. Затем селеноцистеиновая синтаза, использующая в качестве кофермента пиридоксальфосфат (рибонуклотидный кофактор!), удаляет гидроксильную группу серина, превращая его в аминоакриловый интермедиат, который служит акцептором. для активированного селена. Донором селена является моноселенфосфат, образованный из селенида и АТФ (опять же рибонуклеотидный кофактор!) и селенофосфатсинтазы.
Также немаловажным аргументом, говорящем о правдоподобности данного утверждения, является хорошая обоснованность модели происхождения кодируемого пептидного ситнтеза (генетического кода), в которой первыми аминокислотами были именно те, синтез которых хорошо воспроизводится в опытах по абиогенному синтезу (глицин, аланин, валин, аспартат и некоторые другие). Один из этих доводов уже приведён в статье "Верхняя половина тРНК более древняя. Протокодоны и первые аминокислоты в древнем пептидном синтезе", и ряд новых доводов в пользу того, что именно эти аминокислоты должны были быть первыми, будет приведён в других статьях.
9. В ходе дальнейшей эволюции РНК-олигонуклеотиды обеспечили биосинтез новых аминокислот, что предопределило следующий этап возникновения жизни - возникновение кодируемого пептидного синтеза, который будет резюмирован позднее в отдельной статье.
10. Только возникновение сложных кодируемых белков позволило увеличить точность матричного воспроизведения РНК-геномов, а, значит, увеличить размеры РНК-генома с нескольких нуклеотидов до величины порядка нескольких тысяч нуклеотидов.Тем не менее предельный размер значащей части РНК-геномов не должен был превышать размер значащей части генома современных РНК-вирусов вследствие низкой (относительно ДНК) точности копирования. Поэтому подобно РНК-геномам современных вирусов, в геномах РНК-прогенота использовались всевозможные ухищрения, позволявшего при малом размере геномов генерировать максимальное количество различных генов. Об одном из таких ухищрений уже было обоснованно рассказано в статье "Древние тРНК с комплементарными антикодонами кодировались комплементарынми РНК-парами". О ряде других ухищрений будет написано позднее.
Эти два процесса (увеличение сложности кодируемых белков и рост размера РНК-генома) следует рассматривать как взаимно усиливающие друг друга, что в дальнейшем дало возможность перейти к новому на пять и более порядков более точному кодированию на уровне ДНК.
Литература.
Колчанов Н.А., Суслов В.В. Кодирование и эволюция сложности биологической организации. // В сб. "Эволюция биосферы и биоразнообразия" Под ред. Леоновой Т.Б. и др. Москва, 2006.
Маракушев С.А., Белоногова О.В. Метаболический дизайн и биометрический катализ архаического хемоавтотрофного цикла фиксации СО2. // Вестник Моск. Универ. 2010а. Сер. Химия. Т.51. С. 254-260.
Маракушев С.А., Белоногова О.В. Эволюция архаической хемоавтотрофной системы фиксации двуокиси углерода в гидротермальных системах. // Доклады Академии Наук. 2010б. №2. С.262-268.
Ратнер В.А. Генетика, молекулярная кибернетика. Личности и проблемы. Новосибирск, "Наука", 2002.
Спирин, А.С. Где, когда и в каких условиях мог возникнуть и эволюционировать мир РНК? // Палеонтологический журнал 2007. №5, С.11-19.
Шапиро Р. У истоков жизни. // В мире науки. №10. 2007. С.22-29.
Эйген М., Шустер П.. Гиперцикл. Принципы организации макромолекул. М.: Мир. 1973.
Bada J.L., Lazcano A. Some like it hot, but not the first biomolecules. // Science. 2002. V.296. P.1982–1983.
Borgeson, W. et al. Discussing the origin of life. // Science. 2002. V.298. P.747–749.
Deamer D., Dworkin J.P, Sandford S.A., Bernstein M.P, Allamandola L.J. The first cell membranes. // Astrobiology. 2002. V.2. P.371-381.
311–341.
De Duve. C. Blueprint for a Cell: The Nature and Origin of Life. Neil Patterson Publishers. 1991.
De Duve C. 2002. Life Evolving. New York: OUP
De Duve C. 2005. Singularities. Cambridge: CUP.
Delaye L., Lazcano A. Prebiological evolution and the physics of the origin of life. // Phys. Life Rev. 2005. V.2. P.47–64.
Eschenmoser, A. Question1: commentary reffering to the statement ‘‘The origin of life can be traced back to the origin of kinetic control’’ and question ‘‘Do you agree with tsis statement; and how would you envisage the prebiotic evolutionary bridge between thermodynamic and kinetic control?’’. // Origins Life Evol. Biosphere. 2007 V.37. P.309–314.
Fernando, C., Rowe, J. Natural seleсtion in chemical evolution. J. Theor. Biol. 2007. V.247. P.152–167.
Joyce G.F. The antiquity of RNA-based evolution. // Nature. 2002. V.418. P.214–221.
Horgan J. In the beginning. Scientific American 1991. V.264 P.100–109.
Kaufman S.A., The origin of order: Self-organization and seleсtion in evolution. Oxford University Press, New York.1993.
Lazcano A., Miller S.L. On the origin of metabolic pathways. // J. Mol. Evol. 1999. V.49. P.424–431.
Lindahl P.A. Stepwise evolution of nonliving to living chemical system. // Origins Life Evol. Biosphere. 2004. V.34. P.371–389.
Lee N., Bessho Y., Wei K., Szostak J.W., Suga H. Ribozymecatalyzed tRNA aminoacilation. // Nature Struct. Biol. 2000. V.7. P.28-33.
Marakushev S.A, Belonogova O.V. The parageneses thermodynamic analysis of chemoautotrophic CO2 fixation archaic cycle components, their stability and self-organization in hydrothermal systems. // J. Theor. Biol. 2009. V.257. P.588-597..
Martin W., Russell M.J. On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Phil. Trans. R. Soc. Lond. В. 2003. V.358. P.59-85.
Orgel L.E.The implausibility of metabolic cycles on the prebiotic earth. // PLoS Biol. 2008. V.6: e18, 10.1371/journal.pbio.0060018.
Orgel L.E. Prebiotic chemistry and the origin of the RNA world. Crit. Rev. Biochem. Mol. Biol. 2004. V.39. P.99–123.
Robertson A., Sinclair A.J., Philp D. Minimal self-replicating systems. // Chem. Soc. Rev. 2000. V.29. P.141–152.
Russell M.J., Daniel R.M., Hall A. On the emergence of life via catalytic iron-sulphide membranes. // Terra Nova 1993. V.5. P.343–7.
Siebers B, Schönheit P. Unusual pathways and enzymes of central carbohydrate metabolism in Archaea. // Curr. Opin. Microbiol. 2005. V.8. P.695–705.
Smith E., Morowitz H.G. Universality in intermediary metabolism. // Proc.Natl. Acad. Sci. USA 2004. V.101. P.13168–13173.
Wächtershäuser G. Before enzymes and templates: theory of surface metabolism. Microbiol. Rev. 1988a. V.52. P.452–484.
Wächtershäuser G. Groundworks for an evolutionary biochemistry: the iron–sulfur world. // Prog. Biophys. Mol. Biol. 1992. V.58. P.185–201.
Wächtershäuser G. The origin of life and its methodological challenges. // J. Theor. Biol. 1997. V.187. P.483–494.
Статьи, связанные с темой.
Автотрофное термофильное происхождение жизни
Древний дублетный генетический код был предопределён путями синтеза аминокислот
К вопросу о происхождении жизни
Минеральные корни современных ферментов
Реликтовые составляющие современного метаболизма. С чего он мог начаться?