Введение
Одно из впечатляющих открытий в современной когнитивной нейробиологии, которое предвидели отцы-основатели электрофизиологии, заключается в том, что наш мозг никогда не находится в состоянии покоя. В периоды бодрствования, когда мы стараемся ничего не делать, глобальный метаболизм в мозге не снижается, и некоторые его отдельные области по-прежнему активны. Эти области образуют сети в состоянии покоя, или RSN. Более того, во время общего преднамеренного бездействия и невнимания к внешнему миру небольшое, но последовательное повышение активности происходит в определенном наборе регионов, называемых “Сетью режима по умолчанию” или DMN. Это основная часть ряда RSN, которые интенсивно исследовались в последние пару десятилетий. Были идентифицированы моторные, зрительные, слуховые сети и сети, связанные с языком, рабочей памятью и вниманием. Считается, что DMN, который был обнаружен одним из первых, тесно связан с функцией сознания.
Активность в режиме ожидания характерна не только для человека и нечеловекообразных приматов, но и для грызунов (Mantini et al., 2011; Mantini and Vanduffel, 2013). Таким образом, DMN можно рассматривать как одну из эволюционно старых консервативных систем (Mantini et al., 2011; Mantini and Vanduffel, 2013). DMN включает в себя заднюю поясную кору/преклинье (PCC), медиальную префронтальную кору (mPFC), двустороннюю нижнюю теменную дольку (IPL) и другие области, в том числе нижнюю височную извилину. Диффузионно-тензорная визуализация (DTI; van den Heuvel et al., 2008, 2009; Greicius et al., 2009) и функциональная магнитно-резонансная томография в состоянии покоя (fMRI) показывают, что эти области тесно взаимосвязаны (Greicius et al., 2003; Biswal et al., 2010). Большинство современных исследований, сочетающих фМРТ в состоянии покоя с крупномасштабным «сетевым анализом», сообщают о функциональной связности, которая отражает статистические зависимости (например, временные корреляции) между областями мозга. Но параметр корреляции не может ответить на вопрос о причинно-следственном влиянии одной нейронной системы на другую, и нам нужно найти способы понять эффективную связность. Как сообщают многие исследователи, области DMN участвуют в выполнении различных задач, таких как интроспекция и спонтанное познание (Эндрюс-Ханна и др., 2010), извлечение информации из памяти, эмоциональные процессы и социальное познание. Но функции каждого узла в сети DMN до сих пор чётко не определены. И ключевой идеей для определения функций определённого узла является изучение функциональности тех областей, с которыми этот узел связан.
Существует несколько методов измерения эффективной связности для фМРТ-исследований. К ним относятся моделирование структурными уравнениями (SEM; например, Bavelier et al., 2000), анализ причинно-следственных связей по Грейнджеру (GC; например, Goebel et al., 2003) и биологически правдоподобное динамическое причинно-следственное моделирование (DCM; см. Friston et al., 2003). С помощью первого метода можно анализировать только устойчивые паттерны мозговой активности: SEM не может работать с динамическими изменениями сигнала фМРТ. GC также является модельным подходом, в котором для оценки причинно-следственных взаимодействий между временными рядами фМРТ используется векторная авторегрессионная модель (VAR). Есть несколько исследований, в которых подход GC использовался для изучения причинно-следственных связей между узлами DMN, но их результаты едва ли можно назвать последовательными. Наконец, третий метод, DCM, работает с временными рядами фМРТ и явно моделирует динамику нейронов в соответствии с эффективной связью. По мере дальнейшего развития начинает преобладать мнение, что DCM является более последовательным и информативным подходом к выявлению причинно-следственных связей между областями мозга на основе данных фМРТ, чем другие (Дэвид и др., 2008; Фристон, 2009).
Байесовские подходы также используются для оценки взаимосвязей внутри DMN. В работе Wu et al. (2014) было показано, что DMN можно разделить на две подсистемы в зависимости от их функций. Эти подсистемы взаимодействуют друг с другом не только в состоянии покоя, но и при выполнении задачи на семантическое суждение. Причинно-следственные связи между ключевыми узлами DMN кардинально меняются при переходе от состояния покоя к состоянию выполнения задачи. Авторы предполагают, что DMN может быть стабильной функциональной структурой как в состоянии покоя, так и в состоянии выполнения задачи, но в разных состояниях информация в DMN обрабатывается по-разному. Ограничение подхода на основе байесовских сетей заключается в том, что этот метод не может выявить взаимные связи в DMN. Он также даёт представление о процессе в конкретный момент времени и не может выявить динамические изменения в связях между его частями. DCM лишён этих ограничений и может дополнить это исследование.
DCM — это байесовский подход, обычно используемый для объяснения изменений эффективной связи, лежащих в основе реакций мозга, связанных с выполнением задач (Friston et al., 2003; Шараев и Мнацаканян, 2014). Когда задаётся конкретная модель (включая активные области и направленные связи между ними), параметры связи оцениваются на основе структуры модели и наблюдаемых данных фМРТ. Различные модели сравниваются с помощью байесовского выбора модели (BMS; Penny et al., 2004). DCM оценивает эффективную связность, которая является мерой, определяющей влияние одной нейронной системы на другую. Существует множество примеров применения DCM к данным магнитоэнцефалографии и электроэнцефалографии (MEG/EEG) и фМРТ (см., например, Garrido et al., 2007; Moran et al., 2009; Razi et al., 2015).
DCM рассматривает мозг как «чёрный ящик», который получает входные данные и генерирует выходные. Поэтому при использовании DCM для изучения сети в состоянии покоя возникает проблема, заключающаяся в том, что модель DCM не может быть задана без каких-либо управляющих входных данных (Stephan and Friston, 2010). В таких случаях может быть полезно включить в модель стохастические термины (Daunizeau et al., 2009), но можно утверждать, что как спонтанное психическое состояние, так и сигналы фМРТ в состоянии покоя — это не просто случайный шум. Недавно была представлена новая версия DCM, основанная на детерминированной модели, которая генерирует прогнозируемые кросс-спектры (Friston et al., 2014) — спектральная DCM. В то время как стохастическая DCM оценивает зависящие от времени флуктуации в состояниях нейронов, создающих наблюдаемые данные фМРТ, спектральная DCM оценивает инвариантные во времени параметры их кросс-спектров. Это достигается путём замены исходного временного ряда его статистикой второго порядка (т. е. кросс-спектрами) при условии стационарности (Razi et al., 2015). Таким образом, спектральная DCM не рассматривает флуктуации нейронов как стохастический шум, поэтому для настоящего исследования был выбран спектральный подход DCM.
Цель данной работы — изучить эффективную связь между основными узлами DMN и оценить соответствующие параметры связи с помощью новой методики спектральной DCM в сочетании с моделированием нейронной активности в состоянии покоя у 30 здоровых испытуемых с использованием дискретного косинусного набора. Чтобы проверить только биологически правдоподобные гипотезы об эффективной связи, мы использовали данные о структурной связи для сокращения пространства модели.
Сигналы фМРТ в состоянии покоя передают колебания в низкочастотном диапазоне, обычно в пределах 0,01–0,08 Гц (Biswal et al., 1995), поэтому для выявления узлов DMN и определения временных рядов для DCM состояние покоя было смоделировано с помощью обобщённой линейной модели, содержащей дискретный набор косинусоидальных базисных функций с частотами от 0,0078 до 0,1 Гц, в дополнение к индивидуальным регрессорам (Fransson, 2005; Kahan et al., 2014). После этой процедуры были определены модели DCM, каждая из которых состоит из четырёх основных областей DMN. Затем с помощью процедуры BMS, описанной в Stephan et al. (2009), была определена лучшая модель DCM.
Материалы и методы
Субъекты
Данные МРТ были получены от 30 здоровых испытуемых (10 мужчин и 20 женщин) в возрасте в среднем 24 года (от 20 до 35 лет). Было получено согласие каждого участника. Участникам было предложено закрыть глаза, лежать неподвижно и расслабиться. Каждого участника спросили, бодрствовал ли он во время исследования; те, кто заснул в сканере, были исключены из исследования. Разрешение на проведение этого эксперимента было получено этическим комитетом Института высшей нервной деятельности и нейрофизиологии РАН. Что касается спектрального DCM, то среднеквадратичная ошибка уменьшается по мере увеличения количества точек во времени. Основываясь на результатах Razi et al. (2015), мы решили получить 1000 точек во времени (с интервалом в 2 с), что заняло примерно 35 минут сканирования.
Параметры сканирования
Данные МРТ были получены с помощью магнитно-резонансного томографа SIEMENS Magnetom Verio с напряжённостью магнитного поля 3 Тесла. Сагиттальная трёхмерная последовательность быстрого градиентного эха с подготовкой к намагничиванию в Т1-взвешенном режиме была получена со следующими параметрами визуализации: 176 срезов, TR = 1900 мс, TE = 2,19 мс, толщина среза = 1 мм, угол наклона = 9°, время инверсии = 900 мс и поле зрения = 250 × 218 мм2. Данные фМРТ были получены со следующими параметрами: 30 срезов, TR = 2000 мс, TE = 25 мс, толщина среза = 3 мм, угол наклона = 90°, FOV = 192 × 192 мм2. Также мы получили данные, содержащие информацию о способах уменьшения пространственного искажения изображений EPI.
Анализ данных изображений
Данные фМРТ и анатомические данные были предварительно обработаны с помощью SPM12 (доступно бесплатно по адресу http://www.fil.ion.ucl.ac.uk/spm/software/spm12/) на основе Matlab. Предварительная обработка включала следующие этапы: импорт Dicom, приведение центра анатомических и функциональных данных к передней комиссуре, уменьшение пространственного искажения с помощью набора инструментов Field Map в spm12 (Friston et al., 2007). Затем была выполнена коррекция времени срезов для данных фМРТ (коррекция гемодинамической реакции в пространстве, а затем во времени, чтобы избежать выраженных артефактов движения; Sladky et al., 2011). Анатомические данные были сегментированы; как анатомические, так и функциональные данные были нормализованы. Функциональные данные были сглажены с помощью функции Гаусса с изотропным ядром 6 мм.
Мы использовали набор инструментов SPM — WFU pickatlas (доступен бесплатно по адресу http://uvasocialneuroscience.com/doku.php?id=uva_socia:wfu_pickatlas) для создания маски DMN. Маска содержит области правого и левого полушарий: области Бродмана 19 и 39, парагиппокампальную извилину, заднюю поясную извилину, медиальную лобную извилину (Jann et al., 2010; Di and Biswal, 2014).
Состояние покоя было смоделировано с помощью общей линейной модели с дискретным косинусным базисом (GLM-DCT), состоящей из 400 функций с частотами, характерными для динамики состояния покоя: 0,0078–0,1 Гц (Biswal et al., 1995; Deco et al., 2011), шести регрессоров, учитывающих движения головы, и смешанных временных рядов из внемозговых отделов. F-контраст был задан для всех частот DCT, что позволило создать SPM, которая выявила области, демонстрирующие колебания BOLD в пределах частотного диапазона. Полученные статистические параметрические карты затем были замаскированы маской DMN, основанной на ранее опубликованных координатах DMN Монреальского неврологического института (MNI) (Jann et al., 2010; Di and Biswal, 2014). Функциональная связность в DMN хорошо изучена, поэтому для нашего DCM мы взяли в качестве областей интереса (узлов), о которых чаще всего сообщают, четыре основные части DMN: mPFC (3, 54, -2), PCC (0, -52, 26), LIPC левой и правой внутриполушарной коры (-50, -63, 32) и RIPC (48, -69, 35) (Ди и Бисвал, 2014). В квадратных скобках указаны соответствующие MNI-координаты центров областей.
Для анализа DCM была вычислена главная собственная вариация сферы (радиусом 8 мм) (с поправкой на помехи) для каждой области с центром в пиковом вокселе вышеупомянутого F-контраста (см. рисунок 1). Чтобы ограничить количество возможных моделей, было принято допущение, что модель симметрична относительно левой и правой сторон. Действительно, никаких доказательств обратного найдено не было. Были определены следующие модели связей: полностью связная модель, три модели, в которых одни области преимущественно влияли на другие (mPFC, PCC и двусторонняя модуляция), и те же модели, но без прямых связей между двусторонними LIPC и RIPC, всего 4 × 2 = 8 моделей, см. рис. 2. Чтобы проверить стабильность модели в зависимости от интервала сканирования, мы также построили те же модели (с точки зрения узлов и рёбер DCM) на основе первых 500 («начальная» модель) и последних 500 («конечная» модель) сканирований. После оценки GLM у одного испытуемого не было выявлено значимой активности в префронтальной коре во время первых 500 сканирований (но она была выявлена во время последних), поэтому для первоначальной модели у нас было 29 испытуемых, а для окончательной — 30.
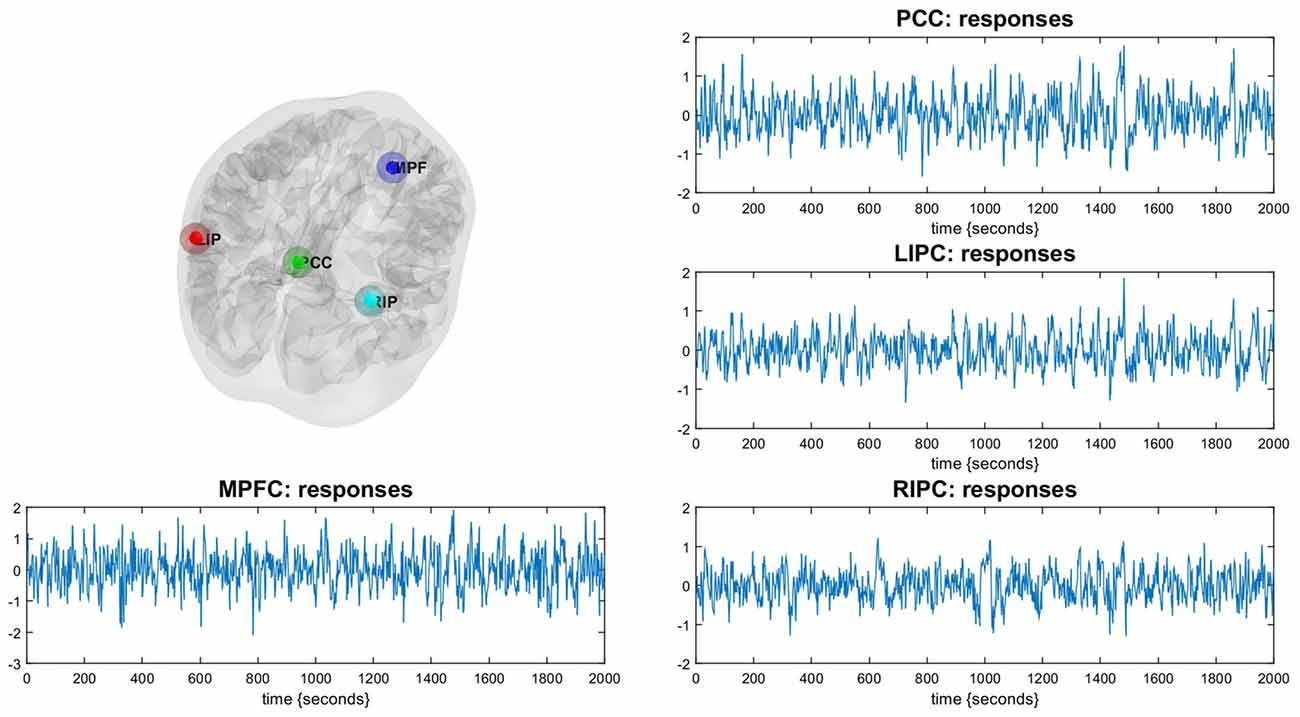
Рисунок 1. Иллюстрация сети режима по умолчанию (DMN). Регионы DMN идентифицируются с помощью обычного SPM-анализа. Соответствующие временные ряды являются основными собственными переменными регионов.
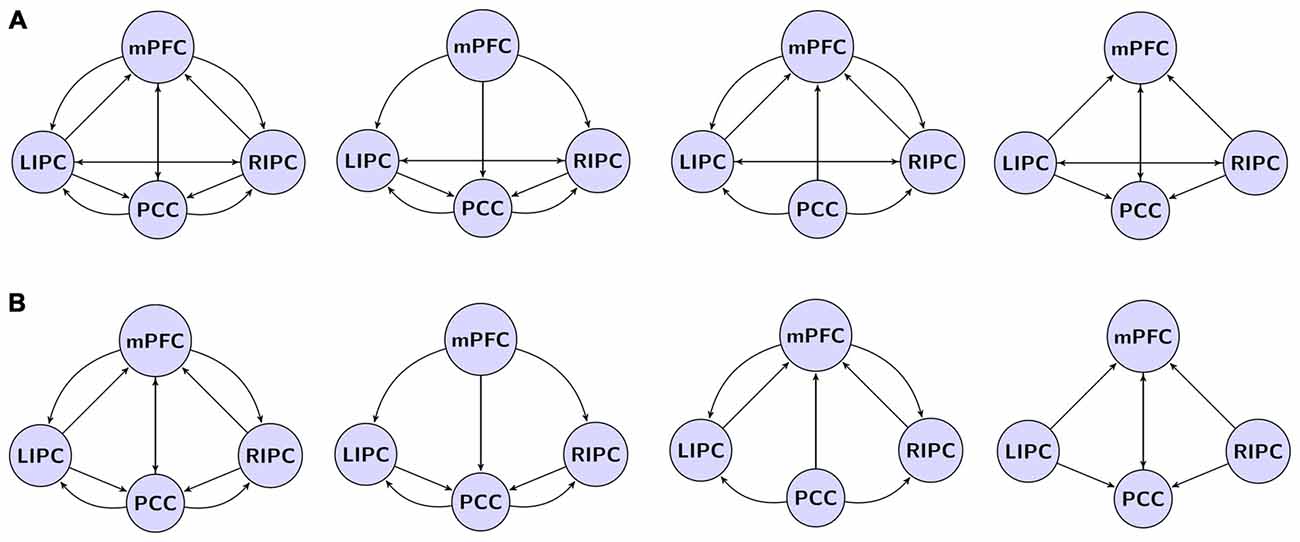
Рисунок 2. Исследуемое пространство моделей. (A) Модели с прямыми связями между двусторонними LIPC и RIPC, слева направо: полносвязная модель, mPFC, PCC, двусторонняя модуляция. (B) Модели без прямых связей между LIPC и RIPC. Двойная стрелка означает взаимные связи.
Для каждого участника эти схемы без внешних воздействий были инвертированы с помощью спектрального DCM. Поскольку мы предполагали, что все участники использовали одну и ту же модель. Для определения наилучшей модели, которая бы сочетала в себе соответствие данных и сложность модели, был использован метод фиксированных эффектов (FFX) BMS (Stephan et al., 2009). Учитывая наилучшую модель, параметры связи для каждого испытуемого были проанализированы количественно с помощью классической статистики и байесовского усреднения моделей (BMA; Пенни и др., 2010), чтобы определить, являются ли некоторые из них стабильными для группы испытуемых. Для проверки того, имеют ли эти параметры значимые ненулевые значения, были проведены t-тесты для одной выборки. Кроме того, было проведено BMA, чтобы получить взвешенные по вероятности значения параметров модели. Затем результаты t-теста и BMA были сопоставлены.
Результаты
BMS показала, что полностью связная модель является лучшей на уровне группы (восемь сравниваемых моделей, рис. 2), что согласуется с предыдущими аналогичными анализами (Li et al., 2012; Razi et al., 2015). Более того, эта модель была лучшей для 24 из 30 испытуемых.
Полностью подключённая модель показала наилучшие результаты на групповом уровне как для начальной (23 раза из 29), так и для конечной (26 раз из 30) моделей.
Результаты BMA и классического t-критерия приведены в таблицах ниже. Чтобы сосредоточиться на нетривиальных связях, мы указываем конкретную связь только в том случае, если её сила превышает 0,1 Гц, а вероятность — 0,95. Для простоты мы также не рассматриваем самосвязи в графах. Модель-победитель показана на рисунке 3.
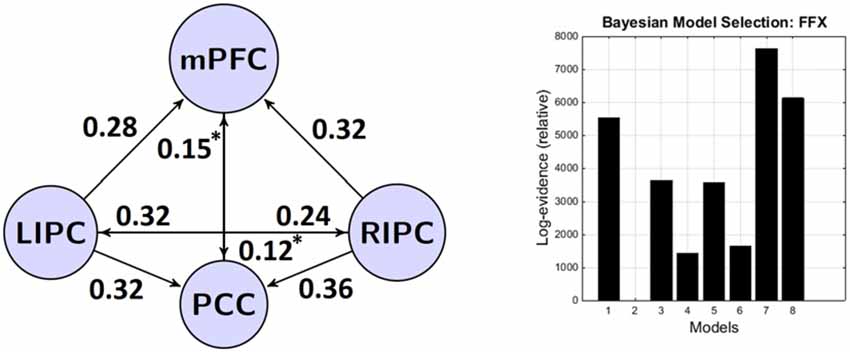
Рисунок 3. Модель-победитель на уровне группы и параметры её связности (в Гц). Слева: модель-победитель и её нетривиальные значимые (p < 0,05) связи. Справа: результаты байесовского выбора модели (BMS) — модели и их (относительные) логарифмические вероятности. Легенда моделей: 1 — латеральная модуляция с (w) прямыми связями между двусторонними LIPC и RIPC, 2 — латеральная модуляция без (wo) прямых связей между двусторонними LIPC и RIPC, 3 — модуляция mPFC (w), 4 — модуляция mPFC (wo), 5 — модуляция PCC (w), 6 — модуляция PCC (wo), 7 — полное подключение (w), 8 — полное подключение (wo). *Незначимо после поправки Бонферрони.
В таблицах 1, 2 представлены средние значения силы связи (в Гц) по результатам BMA и t-критериального анализа, в таблице 3 показаны соответствующие стандартные отклонения.
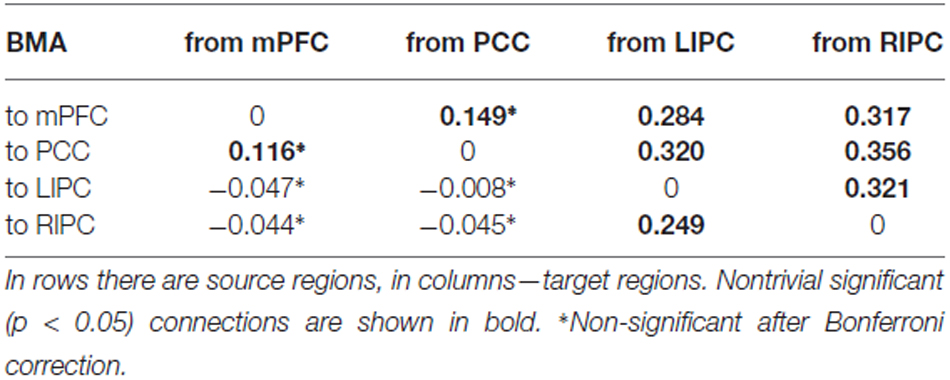
Таблица 1. Средняя сила связи (в Гц) по данным BMA.

Таблица 2. Средняя сила связи (в Гц) по результатам t-теста.
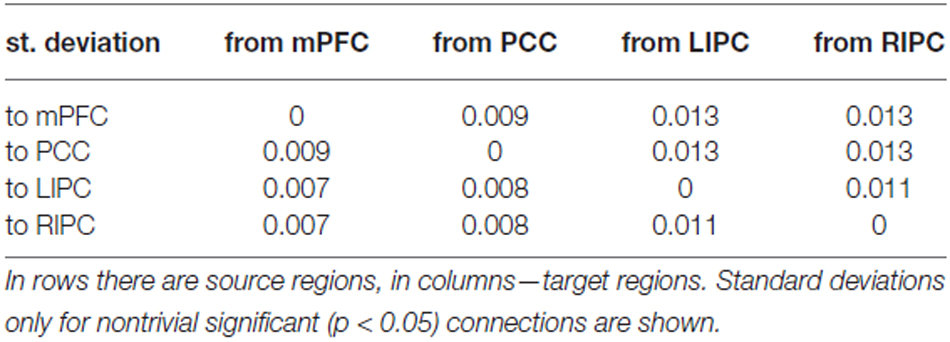
Таблица 3. Стандартные отклонения силы соединения.
Из таблиц 1, 2 видно, что результаты BMA и t-критерия практически идентичны, хотя классический t-критерий не учитывает предполагаемую точность каждой связи, рассчитанную с помощью DCM. Таким образом, для дальнейшего анализа мы можем использовать значения, полученные с помощью процедуры BMA.
Можно заметить, что связи между двусторонними IPC и mPFC и PCC довольно сильные, значимые, отличные от нуля (p < 0,01) и симметричные по силе (p < 0,05). Эти результаты отчасти схожи с теми, о которых ранее сообщали Ди и Бисвал (2014), которые обнаружили сильную межполушарную асимметрию в DMN с акцентом на правую сторону. Связи между mPFC и PCC являются двунаправленными, значимыми в группе и более слабыми, чем связи, исходящие из двусторонних IPC (p < 0,03). В целом, все связи от LIPC/RIPC к другим областям DMN намного сильнее на уровне группы, поэтому мы можем предположить, что эти области играют ведущую роль в DMN. Этот результат также согласуется с работой Di и Biswal (2014) и Razi et al. (2015).
В таблице 4 представлены средние значения силы связи (в Гц) для начальной/конечной моделей соответственно, в таблице 5 показаны соответствующие стандартные отклонения. Во-первых, из этих таблиц видно, что параметры связи в среднем имеют более низкие значения, чем параметры модели 1000 сканирований («общая»). Особенно это заметно по параметрам связи LIPC/RIPC: все они (кроме связи LIPC с PCC) значительно меньше (p < 0,05) в начальной и конечной моделях по сравнению с общей моделью. В начальной и конечной моделях эти параметры очень похожи, хотя некоторые из них не проходят коррекцию Бонферрони. Возможно, это связано с более коротким временем сканирования: оценки параметров меньше. В работе Razi et al. (2015) авторы показали, что среднеквадратичная ошибка оценки параметров при 512 сканах на 50% выше, чем при 1024. Тем не менее можно заметить, что нетривиальные параметры связности в общей, начальной и конечной моделях различаются только по величине. Эти различия не приводят ни к изменениям в структуре связности с точки зрения наличия/отсутствия конкретной связи, ни к изменениям в роли конкретной связи с возбуждающей на тормозящую. Это означает, что победившая модель стабильна в разные периоды времени с точки зрения своих параметров и отражает относительно стабильную структуру эффективной связности в DMN. Эта структура может незначительно меняться со временем, но основные области и связи между ними остаются прежними. Таким образом, мы можем предположить, что испытуемые находились примерно в одном и том же психическом состоянии во время первой и второй половины эксперимента.
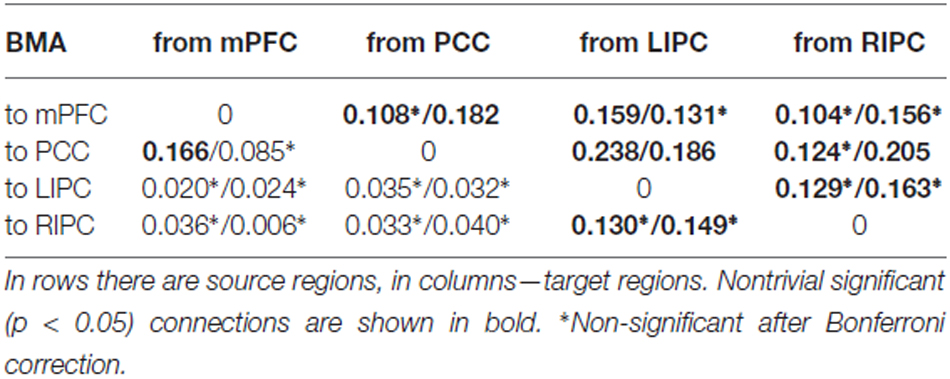
Таблица 4. Средняя сила связи (в Гц) для начальных (первые 500 сканирований) и конечных (последние 500 сканирований) моделей.
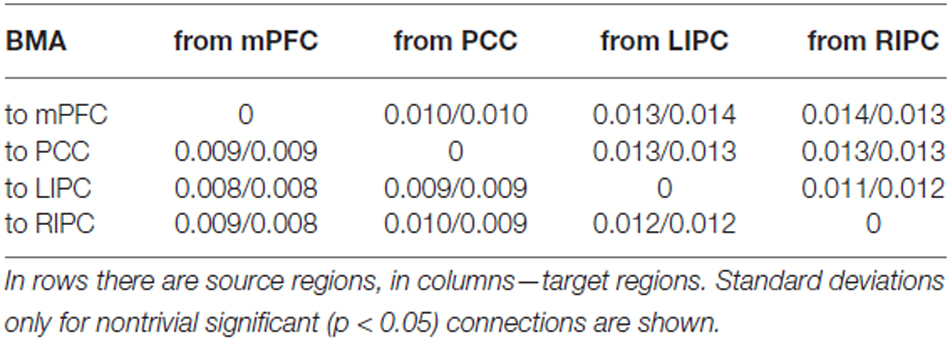
Таблица 5. Стандартные отклонения силы связи для начальных (первые 500 сканирований) и конечных (последние 500 сканирований) моделей.
Следует отметить, что, несмотря на то, что победившая модель является полносвязной, не все параметры связи проходят порог значимости. Это может быть связано с различием между BMS и BMA. BMS — это вывод о моделях; BMA — это вывод о соответствующих параметрах. В победившей модели могут быть параметры, которые по отдельности не проходят классический t-тест, но при этом каким-то образом влияют на модель. Эти параметры могут сильно различаться у разных испытуемых, иметь небольшие значения или и то, и другое.
Обсуждение
Мы использовали дискретный косинусный базис для моделирования низкочастотных колебаний и определения структуры связей, лежащей в основе DMN, с помощью спектральной DCM на основе данных фМРТ в состоянии покоя. Лучшая модель в группе из 30 испытуемых предполагала сильное влияние двусторонних IPC друг на друга, а также на mPFC и PCC, а также информационный поток от mPFC к PCC и наоборот.
Авторы, изучавшие связь между областями в рамках DMN, часто сообщали о противоречивых результатах. Например, Li et al. (2012) с помощью стохастической модели диффузионной связи показали влияние PCC на mPFC. Другие авторы (Jiao et al., 2011; Di and Biswal, 2014), используя частично другие методы, такие как GCA, обнаружили причинно-следственную связь между mPFC и PCC, но не наоборот. Мы видим обе связи в нашей модели-победителе. Некоторые исследования (Jiao et al., 2011; Di and Biswal, 2014) показали, что двусторонняя межполушарная передача импульсов влияет на префронтальную кору и миндалевидное тело, что также согласуется с нашим исследованием. Наконец, в нашей лучшей модели наблюдается сильная взаимная связь между двусторонней межполушарной передачей импульсов, о которой также сообщали Li et al. (2012). Чжоу и др. (2011) обнаружили влияние RIPC на LIPC, но не наоборот, в то время как такие авторы, как Ди и Бисвал (2014) и Цзяо и др. (2011), не обнаружили никакого взаимодействия между двусторонними IPC. Мы также обнаружили небольшую функциональную асимметрию в двусторонних IPC, что подтверждает аналогичные выводы Ди и Бисвала (2014). Мы обнаружили, что двусторонний IPC оказывает причинно-следственное влияние на области mPFC и PCC, но не наоборот. Таким образом, мы можем предположить, что LIPC и RIPC играют ведущую или модулирующую роль. Это открытие согласуется с предыдущими работами (Di и Biswal, 2014), частично согласуется с Razi et al. (2015), которые не обнаружили влияния RIPC на mPFC. Однако есть работы с другими результатами, например, исследование с использованием GC (Jiao et al., 2011; Zhou et al., 2011). Zhou et al. (2011) обнаружили причинно-следственную связь между LIPC и mPFC, в то время как Jiao et al. (2011) показали симметричную причинно-следственную связь между mPFC и двусторонним IPC и двусторонним IPC и PCC.
Это расхождение может быть связано с небольшим размером выборки в предыдущих исследованиях, из-за чего некоторые связи оказались незначительными, или с различными подходами к измерению причинно-следственных связей. В нашем исследовании у нас была довольно большая выборка (30 испытуемых), что соответствует лучшим практикам нейробиологии и экспериментальной психологии. Кроме того, следует отметить, что эмпирически было доказано, что модель DCM является более достоверным методом изучения эффективных связей, чем модель GC (David et al., 2008).
Префронтальная кора и поясная кора — два основных узла DMN, о которых чаще всего сообщается в различных подходах, таких как анализ независимых компонентов (ICA), корреляции на основе семян и другие. Наши результаты показали причинно-следственную связь между префронтальной корой и поясной корой, а также наоборот. Кроме того, стоит отметить, что связи между префронтальной корой и поясной корой, а также между поясной корой и префронтальной корой практически одинаковы по силе и вероятности.
PCC был описан как структурное ядро, которое связывается с основными структурами мозга по всему мозгу (Хагманн и др., 2008), выполняя важнейшие когнитивные функции (Козловский и др., 2012). GC-исследование всего мозга показало, что PCC является надежно управляемым центром, который получает информацию от всего мозга (Дешпанде и др., 2011; Ян и Хэ, 2011). Таким образом, PCC может быть особым региональным центром, который собирает информацию из других областей DMN, а также из всего мозга в целом.
Наибольшая доля «богатых» клубных коннекторов была обнаружена в DMN. Согласно одному из исследований, в общей структурной связности между периферийными сетями и DMN, а также в сетях, отвечающих за выполнение задач (Goulden et al., 2014), преобладают «богатые» клубные соединения. И большинство соединений связаны с периферийными сетями RSN.
Высокая плотность связей между DMN, сетями, отвечающими за значимость, и исполнительными сетями позволяет предположить, что PCC может быть значимым структурным и функциональным ядром, которое накапливает информацию из когнитивных, мультимодальных сетей и может играть очень важную роль в коммуникации с другими сетями RSN, в основном через их представителей в «богатом клубе».
Наши результаты и предыдущие исследования показывают, что PCC управляется всеми основными областями DMN и, возможно, другими областями мозга. Однако из нашей работы и предыдущих исследований неясно, какие области являются целевыми для информационного потока, исходящего из PCC. Одна из возможных гипотез заключается в том, что PCC может быть связующим звеном между функциональными сетями (Di and Biswal, 2014). Это согласуется с теорией о том, что DMN — это корковая система более высокого порядка, которая осуществляет взаимный обмен информацией с подчинёнными ей системами мозга (Кархарт-Харрис и Фристон, 2010). Таким образом, необходимы дальнейшие исследования, чтобы понять, как сети мозга, отвечающие за конкретные задачи, зависят от DMN.
Анализ полученных данных показывает, что наилучшая модель имеет структуру взаимодействий областей DMN (отмечена толстыми стрелками на рисунке 4A), частично аналогичную моделям, полученным в других исследованиях (рисунки 4B,C), но также содержит связи, объединяющие другие модели взаимодействий (отмечены тонкими стрелками на рисунке 4A). Как и в других моделях, присутствуют эффективные управляющие связи областей LIPC и RIPC, соединяющие друг друга напрямую и через области mPFC и PCC. Полученные данные несколько отличаются от результатов других исследовательских групп (см. рисунок 4), что может быть связано с изменением весовых коэффициентов причинно-следственных связей в нейронных сетях из-за активного влияния дополнительных областей DMN. Одной из возможных причин расхождений между нашими результатами и работой Razi et al. (2015) является количество сканов, использованных для моделирования. Когда мы провели BMA для начальной и конечной моделей (первые и последние 500 сканов), некоторые связи (например, от RIPC к mPFC) стали незначимыми после поправки Бонферрони. Несмотря на то, что наша модель связей не может быть с уверенностью распространена на всю популяцию, наши выводы о стабильности модели во времени могут служить дополнительным доказательством её надёжности.
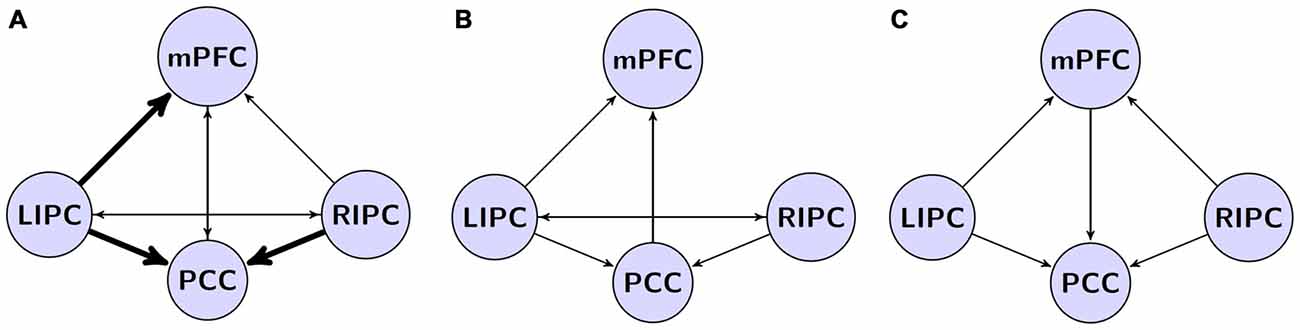
Рисунок 4. Наша модель в сравнении с моделями из предыдущих исследований. (A) Модель-победитель из текущего исследования. Эффективные связи, общие для всех существующих моделей, показаны толстыми стрелками. (B) Модель Рази и др. (2015) на основе спектрального динамического причинно-следственного моделирования (DCM); (C) модель Ди и Бисвал (2014) на основе детерминированного DCM.
Наша следующая задача — проверить стабильность полученной модели с добавлением новых областей. Например, известно, что область PCC имеет проекционные связи с передней поясной корой, префронтальной корой, латеральной теменной корой и парагиппокампальной корой (Mantini et al., 2011). Было показано (Mantini and Vanduffel, 2013), что парагиппокампальные области, играющие ключевую роль в механизмах извлечения информации из памяти, также относятся к DMN.
Мы предполагаем, что добавление новых функционально важных областей, таких как парагиппокампальная кора, может изменить связи в ранее полученных моделях. Это имеет особое значение для обсуждения роли сетей DMN в модуляции когнитивных состояний как объективного параметра процессов на базовом уровне функционирования нейронных сетей (Raichle and Snyder, 2007), роли мПФК в интеграции эмоциональных и когнитивных процессов (Raichle et al., 2001) и в оценке вклада мПФК в ментальное моделирование поведенческих реакций на основе информации из памяти. Полученные данные подтверждают гипотезы об участии DMN в реализации двух функций — базовой спонтанной активации когнитивного уровня (включая механизмы «сверху вниз») и мониторинга интермодального восприятия контекста окружающей среды (Морком и Флетчер, 2007; Мантини и Вандуффель, 2013).
Особый интерес представляет изучение причинно-следственных взаимодействий между крупномасштабными сетями. Например, функциональная связь всего мозга у здоровых людей (эксперимент с магнитоэнцефалографией) выявила сложные сетевые взаимодействия в нескольких физиологических частотных диапазонах (особенно в альфа-диапазоне; Schmidt et al., 2014). Сочетание ICA и DCM было использовано для подтверждения вывода Шридхаран и др. (2008) о том, что сеть значимости (SN) является ключевым элементом для переключения между центральной исполнительной сетью (CEN) и DMN (Гоулден и др., 2014). Этот новый подход к извлечению временных рядов ICA для представления всей сети, а не отдельных интересующих нас областей, недавно помог нам решить давнюю проблему нисходящего влияния на визуальное восприятие (Верхлютов и др., 2014). В том же ключе можно использовать DCM для изучения взаимосвязи между большим количеством RSN. Сокращая исходное пространство, можно лучше понять, как эти сети модулируют переключение, когда меняются психические состояния, постановка задачи или окружающая обстановка.
Авторский вклад
MGS: математические методы, программирование, анализ данных, моделирование, подготовка текста статьи (введение, методы, результаты) и иллюстраций, ответственность за все аспекты работы. VVZ: планирование эксперимента, сбор и предварительная обработка данных, окончательное утверждение статьи, ответственность за все аспекты работы. VLU: концепция работы, текст статьи (обсуждение), окончательное утверждение статьи, ответственность за все аспекты работы. SIK: сбор данных, текст статьи (введение), окончательное утверждение статьи, ответственность за все аспекты работы. BMV: общее руководство, идея проекта, текст статьи (введение, нейропсихологические аспекты обсуждения), окончательное утверждение статьи, ответственность за все аспекты работы.
Финансирование
Эта работа была частично поддержана грантом Российского научного фонда, проект РНФ № 14-28-00234 (структура DMN и ее связь с психическими состояниями) и грантом Российского фонда фундаментальных исследований, проект РФФИ № 15-29-01344 (частотный анализ в нейрокогнитивных исследованиях).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Эндрюс-Ханна Дж. Р., Рейдлер Дж. С., Сепулькре Дж., Пулен Р. и Бакнер Р. Л. (2010). Функционально-анатомическое разделение сети пассивного режима работы мозга. Нейрон 65, 550–562. doi: 10.1016/j.neuron.2010.02.005
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Бавелье, Д., Томанн, А., Хаттон, К., Митчелл, Т., Корина, Д., Лю, Г. и др. (2000). Зрительное внимание к периферии усилено у людей с врождённой глухотой. J. Neurosci. 20:RC93, 1–6.
Бисвал, Б. Б., Элдрет, Д. А., Мотес, М. А. и Рипма, Б. (2010). Зависимые от задачи индивидуальные различия в связях префронтальной коры. Cereb. Cortex 20, 2188–2197. doi: 10.1093/cercor/bhp284
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Бисвал Б., Йеткин Ф. З., Хотон В. М. и Хайд Дж. С. (1995). Функциональная связь в моторной коре головного мозга человека в состоянии покоя с использованием эхо-планарной МРТ. Magn. Reson. Med. 34, 537–541. doi: 10.1002/mrm.1910340409
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Кархарт-Харрис, Р. Л., и Фристон, К. Дж. (2010). Режим по умолчанию, эго-функции и свободная энергия: нейробиологический взгляд на идеи Фрейда. Brain 133, 1265–1283. doi: 10.1093/brain/awq010
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Даунизо, Дж., Фристон, К. Дж. и Кибель, С. Дж. (2009). Вариационная байесовская идентификация и прогнозирование стохастических нелинейных динамических причинно-следственных моделей. Physica D 238, 2089–2118. doi: 10.1016/j.physd.2009.08.002
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Дэвид О., Гийом И., Сайе С., Рейт С., Дерансар К., Сегебарт К. и др. (2008). Выявление нейронных водителей с помощью функциональной МРТ: электрофизиологическая проверка. PLoS Biol. 6:e315. doi: 10.1371/journal.pbio.0060315
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Деко, Г., Джирса, В. К. и Макинтош, А. Р. (2011). Новые концепции динамической организации активности мозга в состоянии покоя. Nat. Rev. Neurosci. 12, 43–56. doi: 10.1038/nrn2961
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Дешпанде, Г., Сантханам, П. и Ху, К. (2011). Мгновенная и причинно-следственная связь в сетях мозга в состоянии покоя, полученная на основе данных функциональной МРТ. Neuroimage 54, 1043–1052. doi: 10.1016/j.neuroimage.2010.09.024
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Ди, К. и Бисвал, Б. (2014). Определение структуры сети по умолчанию с помощью динамического причинно-следственного моделирования на основе функциональной магнитно-резонансной томографии в состоянии покоя. Neuroimage 86, 53–59. doi: 10.1016/j.neuroimage.2013.07.071
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Франссон, П. (2005). Спонтанные низкочастотные колебания сигнала BOLD: исследование с помощью фМРТ гипотезы о режиме работы мозга в состоянии покоя. Hum. Brain Mapp. 26, 15–29. doi: 10.1002/hbm.20113
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Фристон, К. (2009). Причинно-следственное моделирование и связи в мозге при функциональной магнитно-резонансной томографии. PLoS Biol. 7:e33. doi: 10.1371/journal.pbio.1000033
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Фристон К. Дж., Эшбернер Дж. Т., Кибель С. Дж., Николс Т. Э. и Пенни У. Д. (ред.). (2007). Статистическое параметрическое картирование: анализ функциональных изображений мозга. Лондон: Эльзевир.
Фристон К. Дж., Харрисон Л. и Пенни У. (2003). Динамическое причинно-следственное моделирование. Neuroimage 19, 1273–1302. doi: 10.1016/s1053-8119(03)00202-7
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Фристон, К. Дж., Кахан, Дж., Бисвал, Б. и Рази, А. (2014). DCM для фМРТ в состоянии покоя. Neuroimage 94, 396–407. doi: 10.1016/j.neuroimage.2013.12.009
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Гарридо, М. И., Килнер, Дж. М., Кибель, С. Дж., Стефан, К. Э. и Фристон, К. Дж. (2007). Динамическое причинно-следственное моделирование вызванных потенциалов: исследование воспроизводимости. Neuroimage 36, 571–580. doi: 10.1016/j.neuroimage.2007.03.014
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Гобель, Р., Робрук, А., Ким, Д. С. и Формисано, Э. (2003). Исследование направленных корковых взаимодействий в данных фМРТ с временным разрешением с помощью векторного авторегрессионного моделирования и картирования причинно-следственных связей по Грейнджеру. Magn. Reson. Imaging 21, 1251–1261. doi: 10.1016/j.mri.2003.08.026
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Гулден Н., Хуснулина А., Дэвис Н. Дж., Брейсвелл Р. М., Бокде А. Л., Макналти Дж. П. и др. (2014). Сеть, отвечающая за значимость, отвечает за переключение между сетью режима ожидания и центральной исполнительной сетью: репликация из DCM. Neuroimage 99, 180–190. doi: 10.1016/j.neuroimage.2014.05.052
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Грейсиус, М. Д., Краснов, Б., Рейсс, А. Л. и Менон, В. (2003). Функциональная связь в мозге в состоянии покоя: сетевой анализ гипотезы режима ожидания. Proc. Natl. Acad. Sci. U S A 100, 253–258. doi: 10.1073/pnas.0135058100
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Грейсиус, М. Д., Супекар, К., Менон, В. и Догерти, Р. Ф. (2009). Функциональная связь в состоянии покоя отражает структурную связь в сети пассивного режима работы мозга. Cereb. Cortex 19, 72–78. doi: 10.1093/cercor/bhn059
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Хагманн П., Каммун Л., Жиганде К., Меули Р., Хани К. Дж., Ван Ведин Дж. и др. (2008). Картирование структурного ядра коры головного мозга человека. PLoS Biol. 6:e159. doi: 10.1371/journal.pbio.0060159
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Янн К., Коттлоу М., Диркс Т., Бош К. и Кёниг Т. (2010). Топографические электрофизиологические признаки сетей фМРТ в состоянии покоя. PLoS One 5:e12945. doi: 10.1371/journal.pone.0012945
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Цзяо, К., Лу, Г., Чжан, З., Чжун, Ю., Ван, З., Го, Ю. и др. (2011). Причинно-следственное влияние Грейнджера предсказывает уровни активности BOLD в сети пассивного режима работы мозга. Hum. Brain Mapp. 32, 154–161. doi: 10.1002/hbm.21065
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Кахан Дж., Урнер М., Моран Р., Фландин Г., Маррейрос А., Манчини Л. и др. (2014). Функциональная МРТ в состоянии покоя при болезни Паркинсона: влияние глубокой стимуляции мозга на «эффективную» связь между нейронами. Brain 137, 1130–1144. doi: 10.1093/brain/awu027
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Козловский, С. А., Вартанов, А. В., Никонова, Е. Ю., Пясик, М. М. и Величковский, Б. М. (2012). Поясная кора и процессы памяти у человека. Психол. Россия, гос. арт. 5, 231–243. doi: 10.11621/pir.2012.0014
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Ли, Б., Ван, К., Яо, С., Ху, Д. и Фристон, К. (2012). Зависимая от задачи модуляция эффективной связи в сети режима ожидания. Front. Psychol. 3:206. doi: 10.3389/fpsyg.2012.00206
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Мантини, Д., Геритс, А., Нелиссен, К., Дюран, Ж.-Б., Жоли, О., Симоне, Л. и др. (2011). Режим работы мозга по умолчанию у обезьян. J. Neurosci. 31, 12954–12962. doi: 10.1523/JNEUROSCI.2318-11.2011
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Мантини Д. и Вандуффель У. (2013). Новые роли дефолтной сети мозга. Нейробиолог 19, 76–87. doi: 10.1177/1073858412446202
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Моран, Р. Дж., Стефан, К. Э., Зайденбехер, Т., Пейп, Х. К., Долан, Р. Дж. и Фристон, К. Дж. (2009). Динамические причинно-следственные модели устойчивых реакций. Neuroimage 44, 796–811. doi: 10.1016/j.neuroimage.2008.09.048
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Морком, А. М., и Флетчер, П. К. (2007). Есть ли у мозга базовый уровень? Почему мы должны сопротивляться отдыху. Neuroimage 37, 1073–1082. doi: 10.1016/j.neuroimage.2006.09.013
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Пенни, У. Д., Стефан, К. Э., Даунизо, Дж., Роза, М. Дж., Фристон, К. Дж., Шофилд, Т. М. и др. (2010). Сравнение семейств динамических причинно-следственных моделей. PLoS Comput. Biol. 6:e1000709. doi: 10.1371/journal.pcbi.1000709
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Пенни, У. Д., Стефан, К. Э., Мечелли, А. и Фристон, К. Дж. (2004). Сравнение динамических причинно-следственных моделей. Neuroimage 22, 1157–1172. doi: 10.1016/j.neuroimage.2004.03.026
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Райкл, М. Э., Маклеод, А. М., Снайдер, А. З., Пауэрс, У. Дж., Гаснард, Д. А. и Шульман, Г. Л. (2001). Инаугурационная статья: режим работы мозга по умолчанию. Proc. Natl. Acad. Sci. U S A 98, 676–682. doi: 10.1073/pnas.98.2.676
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Райкл, М. Э., и Снайдер, А. З. (2007). Режим работы мозга по умолчанию: краткая история развивающейся идеи. Neuroimage 37, 1083–1090. doi: 10.1016/j.neuroimage.2007.02.041
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Рази А., Кахан Дж., Рис Г. и Фристон К. Дж. (2015). Проверка конструкции DCM для фМРТ в состоянии покоя. Neuroimage 106, 1–14. doi: 10.1016/j.neuroimage.2014.11.027
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Шмидт, Б. Т., Гьюман, А. С. и Хапперт, Т. Дж. (2014). Функциональная связь всего мозга с использованием показателей фазовой синхронизации в магнитоэнцефалографии в состоянии покоя. Front. Neurosci. 8:141. doi: 10.3389/fnins.2014.00141
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Шараев М. Г. и Мнацаканян Е. В. (2014). Динамическое причинно-следственное моделирование электрических реакций мозга, вызванных простыми стимулами в парадигме визуального парадоксального отклонения. Ж. Всс. Нерв. Деят. И П Павлова 64, 627–638. doi: 10.7868/S0044467714060100
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Сладкий Р., Фристон К. Дж., Трёстл Дж., Каннингтон Р., Мозер Э. и Виндишбергер К. (2011). Эффекты синхронизации срезов и их коррекция в функциональной МРТ. Neuroimage 58, 588–594. doi: 10.1016/j.neuroimage.2011.06.078
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Шридхаран Д., Левитин Д. Дж. и Менон В. (2008). Критическая роль правой лобно-островковой коры в переключении между сетями центральной исполнительной системы и сетью режима ожидания. Proc. Natl. Acad. Sci. U S A 105, 12569–12574. doi: 10.1073/pnas.0800005105
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Стефан К. Э. и Фристон К. Дж. (2010). Анализ эффективной связи с помощью функциональной магнитно-резонансной томографии. Wiley Interdiscip. Rev. Cogn. Sci. 1, 446–459. doi: 10.1002/wcs.58
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Стефан К. Э., Пенни У. Д., Даунизо Дж., Моран Р. Дж. и Фристон К. Дж. (2009). Выбор байесовской модели для групповых исследований. Neuroimage 46, 1004–1017. doi: 10.1016/j.neuroimage.2009.03.025
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
ван ден Хёвел, М. П., Мандл, Р. К. У., Кан, Р. С. и Хулшофф Пол, Х. Э. (2009). Функционально связанные сети в состоянии покоя отражают базовую структурную архитектуру связей в человеческом мозге. Hum. Brain Mapp. 30, 3127–3141. doi: 10.1002/hbm.20737
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
ван ден Хёвел, М., Мандл, Р., Луйгес, Дж. и Хулшофф Пол, Х. (2008). Микроструктурная организация поясной извилины и уровень функциональной связности в режиме по умолчанию. J. Neurosci. 28, 10844–10851. doi: 10.1523/JNEUROSCI.2964-08.2008
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Верхлютов, В. М., Ушаков, В. Л., Соколов, П. А. и Величковский, Б. М. (2014). Крупномасштабный сетевой анализ воображения выявляет расширенные, но ограниченные компоненты нисходящего потока в зрительном восприятии человека. Психол. Россия, гос. арт. 7, 4–19. doi: 10.11621/pir.2014.0401
Ву, К., Ю, К., Яо, Л. и Ли, Р. (2014). Анализ байесовской сети выявил различия в связях сети пассивного режима в состоянии покоя и в состоянии выполнения задачи. Front. Comput. Neurosci. 8:118. doi: 10.3389/fncom.2014.00118
Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Ян, К. и Хэ, Ю. (2011). Управляющие и управляемые архитектуры направленных функциональных сетей человеческого мозга в малом мире. PLoS One 6:e23460. doi: 10.1371/journal.pone.0023460
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый
Чжоу, З., Ван, К., Клар, Н. Дж., Лю, В., Ариас, Д., Лю, Х. и др. (2011). Условная модель причинно-следственных связей Грейнджера для группового анализа в функциональной магнитно-резонансной томографии. Magn. Reson. Imaging 29, 418–433. doi: 10.1016/j.mri.2010.10.008
Опубликованный Реферат | Перекрестная ссылка на Полный текст | Google Scholar - Ученый