Генез и эволюция ориентировочно-исследовательского поведения
РедактироватьОриентировочно-исследовательская активность предваряет любой поведенческий акт. Это основная форма адаптивного поведения, которая возникает в ответ на любое новое свойство окружающей среды. Такое поведение обнаружено у всех живых существ, имеющих нервную систему. Есть ли оно у их эволюционных предшественников одноклеточных организмов? Фактический материал, накопленный предыдущими исследованиями, приводит к следующему предположению: одноклеточные эукариоты имеют ориентировочно-исследовательское поведение, которое модифицирует их последующую активность. В проведенных опытах изучалось поведение одноклеточных Paramecium caudatum при появлении в окружающем пространстве новых свойств, а в электрофизиологических опытах регистрировались внутриклеточные показатели формирования нервной модели стимула при повторном действии электрических раздражений (привыкания). Результаты показали, что появление новых свойств в жизненном пространстве вызывает ориентировочно-исследовательское поведение микроорганизмов, выраженное изменениями двигательной активности, весьма разнообразными по форме проявления.
Ключевые слова: поведение, эволюция, парамеции, новизна, электрическая активность, осциллятор, пейсмекер, привыкание
Многочисленные эксперименты, направленные на изучение особенностей активности людей и животных, показали, что ориентировочно-исследовательское поведение является преддверием любого поведенческого акта, начальной фазой любого действия. Благодаря этой форме взаимоотношения со средой приобретаются знания, необходимые для адаптации к изменениям окружающего мира. К исследовательскому поведению относят активность, направленную на изучение окружающей среды и на поиск свойств, узнавание которых является врожденным. Именно поэтому ориентировочные реакции должны быть присущи живым организмам раннего уровня эволюции.
Ориентировочные реакции возникают в результате несовпадения воспринимаемых новых сигналов с формирующимися «нервными моделями стимула», что предполагает наличие памяти [Соколов, 1963]. Результаты экспериментов позволяют сделать вывод о том, что способность к накоплению индивидуального опыта появляется в эволюции на самых ранних ступенях развития – у донервных простейших организмов, в частности у парамеций [Тушмалова, 2008].
Одноклеточные эукариоты, имеющие ядро и митохондрии, появились примерно 2,5 млрд лет назад. Paramecium – род микроскопических (их размер около 100 мк) одноклеточных организмов, ведущих активный образ жизни. Показано, что парамеции имеют набор движений, позволяющий им осуществлять не менее 10 видов поведенческих реакций. Среди них реакция избегания, реакция ускорения плавательных движений, реакция «замирания» (отсутствие движения), специфические движения в контакте с ограничивающей поверхностью [Котов, 2001]. Комбинации этих «паттернов активности» позволяют парамециям решать самые разные двигательные задачи, связанные с поисками питания, избегания опасностей, противодействия хищникам, преодолением препятствий и т.д.
Изучение поведения инфузорий в различных ситуациях показывает, что чувствительность этих одноклеточных существ к различным свойствам окружающего мира высока. У парамеций описана масса таксисов – хемо-, фото-, гeo-, гравитационный и др.[Guevorkian, Valles, 2006; Van Houten et al., 2000; Schwab-Bonaventure, 1955.]. Среди сигналов, вызывающих активный отклик у парамеций, есть вещества, выполняющие нейромедиаторную функцию у многоклеточных организмов, например ацетилхолин и глутамат [Коштоянц и Кокина, 1962; Van Houten, 2000]. L-глутамат, добавленный во внеклеточную среду, более привлекателен для одноклеточных, чем L-глутамат в высокой концентрации в комбинации с KCl. Наличие чувствительности к различным свойствам окружающего мира является предпосылкой для развития ориентировочно-исследовательского поведения, результат которого выражен в формировании энграммы памяти (нервной модели стимула) и реорганизации активности. Изменения поведения, возникающие у этих существ при действии факторов, нейтральных по биологическому значению, но «новых» по физическим параметрам, мало изучены [Armus et al., 2006].
Фактический материал, накопленный предыдущими исследованиями, приводит к следующему предположению: одноклеточные эукариоты – парамеции – имеют ориентировочно-исследовательское поведение, которое модифицирует их последующую активность. В проведенных нами опытах изучалось поведение одноклеточных Paramecium caudatum при появлении в окружающем пространстве новых свойств, а в электрофизиологических опытах регистрировали внутриклетоточные показатели формирования нервной модели стимула при повторном действии электрических раздражений (привыкания).
Метод
Опыты выполнялись на поведенческом и электрофизиологическом уровне. За поведением парамеций наблюдали при помощи бинокулярного микроскопа МБС-9. Микроорганизмы помещали в каплю на предметном стекле и создавали различные ситуации, в которых они могли продемонстрировать изменения поведения. Можно было модифицировать форму капли, химический состав среды, уровень освещения, вносить различные объекты (крупинки химических веществ – например, сахара или соли, обломки микроэлектродов). В опыте участвовало от 1 до 7 парамеций. Продолжительность эксперимента не более 3–5 мин., так как могло произойти высыхание капли, в которой проходил опыт.
Регистрация электрической активности свободно плавающих микроорганизмов производилась в камере объемом 0,35 мл. При помощи микропипетки парамеций переносили в экспериментальную камеру, содержавшую специальный физиологический раствор (KCl – 4 mM, CaCl – 1 mM, MgCl2 – 5 mM, tris HCl – 1 mM. pH раствора – 7,2). Для регистрации применяли стеклянные микроэлектроды (МЭ), заполненные 0,1М КСl. Для электрической стимуляции использовали деполяризационные импульсы тока силой 0,1–1 нА, длительностью 1000–3000 мс. Раздражения подавали через второй введенный в микроорганизм микроэлектрод (МЭ). Так как интактные парамеции чрезвычайно подвижны, регистрации с двумя внутриклеточными МЭ (регистрирующим и стимулирующим) выполнены всего на трех особях. МЭ вводили в момент «отдыха» парамеции, чтобы не применять никаких способов для снижения двигательной активности. По этой причине время регистрации ограничивалось 3–15 мин. Результаты получены на 50 одноклеточных в различных ситуациях поведенческого и электрофизиологического эксперимента.
Результаты
Поведение
Так как в опыте наблюдали за небольшим количеством парамеций – не более 5–7 – это позволило получить данные о различии в поведении отдельных особей при действии одного и того же фактора в одной и той же среде.
Парамеции ведут себя при одинаковых изменениях внешней среды по-разному. Различия проявлялись сразу же после перенесения их из постоянной среды обитания в обстановку опыта (в каплю на предметном стекле). У одних двигательная активность повышается, у других снижается вплоть до «замирания» – парамеции перестают плавать. Когда микроорганизмы попадают из постоянного места обитания в каплю на предметном стекле, они немедленно начинают знакомиться с этим новым жизненным пространством: оплывают его сначала по краю, точно отслеживая форму, затем пересекают по диаметру, потом поднимаются в верхнюю часть к «куполу». Лишь после этого их плавание становится хаотичным и совершается по всем направлениям. У одноклеточных, проявляющих повышение двигательной активности, заметно меняется скорость плавания, наиболее активные передвижения происходят вблизи места, в котором локализован «очаг» изменений.
В опытах использовали несколько ситуаций, в которых рассматривалось исследовательское поведение парамеций. Например, если они находились в капле, имеющей куполообразную форму, то нарушение ее формы приводило к тому, что в течение двух-трех минут парамеции наиболее часто проплывали около места, которое подверглось деформации. Место изменения привлекает их, но плавание совершается по форме прежней капли, то есть «новый» кусок пространства вызывает любопытство, но не осваивается. Лишь подробное исследование постепенно приводит к тому, что парамеции начинают вплывать в новую часть своего мира и оплывать его вдоль границы.
Наблюдения за одноклеточными показывают, что любое изменение окружающей среды влияет на поведение, которое имело место до воздействия. Парамеции замечают «новизну» обстановки и, следовательно, демонстрируют память.
(1) Каплю жидкости, в которой находятся парамеции, соединяли канальцем с другой такой же каплей (рис. 1). Сначала парамеции «замечали», что появился дополнительный ход, – они часто и намеренно подплывали к устью канальца, образуя своеобразную толпу. Но ни одна не плыла по новому пути. Это «исследование» продолжалось в течение нескольких минут, после чего одна! – по-видимому, самая отважная – парамеция заплывала в канал, но проходила его только до трети или половины – и возвращалась назад.
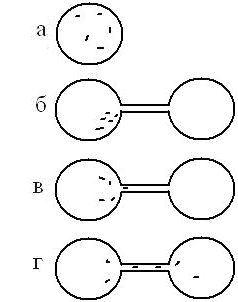
Рис. 1. Схема опыта с переходом в дополнительное пространство: а – капля с парамециями; б, в, г – новая капля, «дорога» и этапы освоения нового пути. Объяснения в тексте.
Потом она же снова отправлялась по пройденному пути и могла доплыть до конца, перейдя во вторую каплю, которая находилась с другой стороны канала. Затем она возвращалась обратно «домой», после чего такой же путь решались проделать и другие парамеции. Впоследствии парамеции равномерно распределялись в первой и второй капле, а канал постоянно применялся как «дорога».
(2). Появление нового предмета в среде обитания (в капле жидкости, где они находятся). В опыте таким незнакомым объектом был обломок кончика стеклянного микроэлектрода. Удалось сделать видеосъемку, которая запечатлела «знакомство» парамеции с этой диковинкой – она не только оплывала ее со всех сторон, располагалась на ней, медленно продвигаясь от конца к началу, долго кружила около отверстия. Наконец кульминацией исследовательской работы было заплывание внутрь трубочки.
Электрофизиологические опыты
Введение микроэлектрода производилось в момент отдыха парамеции – не применялось каких-либо обездвиживающих приемов, так как все способы травмируют двигательную систему этого одноклеточного. По этой причине опыты на единичной парамеции были кратковременны – не более 15 мин. Регистрация электрической активности внутриклеточным микроэлектродом у Paramecium caudatum показала, что уровень мембранного потенциала покоя (ПП) у этих существ отражает «покой» весьма условный: обнаружены изменения от –40–45 мВ до +20 мВ, периоды стабильного значения ПП чрезвычайно короткие – в условиях проведенных опытов не более 30–70 с.
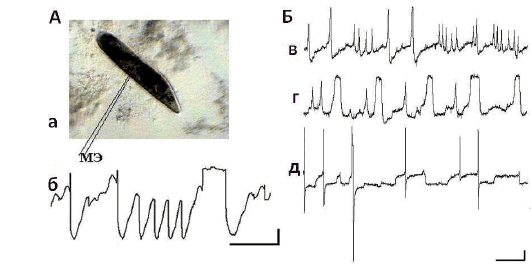
Рис. 2. Регистрация электрической активности от интактной парамеции. Примечания. А: а – Paramecium caudatum с введенным в нее микроэлектродом (МЭ); б – электрическая активность парамеции. Б: в–г – электрическая активность, зарегистрированная у одной и той же парамеции через 3, 7 и 15 мин. после введения МЭ. Калибровка: 10 мВ, 1 с.
Анализ электрической активности показывает, что, во-первых, в клетке регистрируются медленные и быстрые осцилляции, во-вторых, активны, как правило, одновременно несколько эндогенных осцилляторов, что совпадает с наблюдениями других авторов [Коштоянц и Кокина, 1962, Доронин и Зазулин, 1976]. На некоторых записях можно выделить два-три одновременно работающих пейсмекера (рис. 2, б–д). Увеличение амплитуды волн эндогенной активности может приводить к генерации потенциалов действия (ПД) разной амплитуды (рис. 2). Амплитуда ПД, по-видимому, зависит от функциональных особенностей вызвавшего его пейсмекера. На нейрограммах можно видеть ПД как одинаковой, так и различной амплитуды. Они отличаются друг от друга на единицы и даже десятки мВ (рис. 2, 3). Частота зарегистрированных колебаний от 0,1кол/сек до 20 кол/сек, амплитуда от 2–3 мВ до 50 мВ.
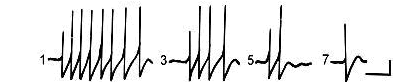
Рис. 3. Изменение ответа парамеции на повторное применение внутриклеточных деполяризационных импульсов тока силой 1,0 нА, длительностью 100 мс. Примечания. Цифры соответствуют номеру предъявляемого стимула. Калибровка: 500 мс, 10 мВ.
При многократном действии сигнала с постоянными физическими параметрами теряется качество новизны и развивается привыкание. Это показано в опытах на поведенческом и нейронном уровне [Соколов, 1969, Кандель, 2012]. В опытах с регистрацией электрической активности у парамеций эту ситуацию моделировали, предъявляя повторно внутриклеточные электрические импульсы. Их наносили через второй электрод, сила тока от 0,1 нА до 1 нА.
Действие электрического тока синхронизировало работу эндогенных осцилляторов и приводило к генерации ПД высокой амплитуды – до 70 мВ (рис. 3). При повторных предъявлениях импульсов постоянных параметров количество ПД уменьшалось. Они полностью исчезали через 7–15 предъявлений. При этом осцилляции сохранялись, хотя их состав фактически не отличался от исходной активности. Результаты этих опытов показали, что одноклеточный эукариот имеет внутриклеточный механизм следообразования.
Обсуждение
Анализ работ, посвященных изучению врожденных и приобретенных форм поведения у беспозвоночных, позволяет сделать вывод о том, что способность к накоплению индивидуального опыта появляется в эволюции на самых ранних ступенях развития – она есть у простейших организмов [Тушмалова, 2008]. Уровень эволюционного развития нервной системы определяет способы взаимодействия со средой [Кандель, 2012], но одноклеточные вообще не имеют нервной системы [Mitchell et al., 2009].
Тем не менее на одноклеточных организмах показан не только феномен привыкания, но и обнаружены способности к опережающему реагированию – по сути, формированию подобия «условных ответов» [Armus et al., 2006; Mirsky et al.,1958; Osborn et al., 1973; Hinkle,Wood, 1998]. Эти виды обучения найдены у парамеций, дрожжей и других микроорганизмов. Отличие в том, что вместо создания сложных систем, основанных на формировании новых синаптических связей между нейронами, у одноклеточных «знания» записываются в генетическую память, закодированную в последовательности нуклеотидов в ДНК.
Изменение поведения выражено соответствующими типами движений. У парамеций движение выполняется благодаря координированной работе многочисленных ресничек, равномерно покрывающих все тело парамеции.
Электрофизиологические исследования реснитчатых простейших, таких как Paramecium, позволили установить, что двигательная активность этих одноклеточных организмов регулируется возбудимостью мембраны. Во многих опытах подтверждено, что при изменении разности потенциалов на мембране простейшего изменяется характер биения ресничек. Анализ экспериментальных данных позволяет сделать предположение о том, что система управления движениями должна быть многоуровневой, при этом роль центрального интегратора и координатора двигательного поведения может выполнять ядро клетки, тесно связанное с периферией с помощью динамичных элементов цитоскелета [Свидерский и др., 2007].
Микроэлектродные регистрации показали взаимодействие нескольких эндогенных осцилляторов. Функция каждого из осцилляторов, по-видимому, связана с контролированием движений определенной группы ресничек парамеции. У одноклеточных существ механизм организации поведения – это механизм координации осцилляторов. Медленные, регулярные колебания потенциала известны у Opalina ranarum, они связаны с изменением направления биения ресничек [Коштоянц и Кокина,1962].
Подводя итог приведенным данным, можно сказать, что электрическая гетерогенность одноклеточных является отражением функциональной изменчивости, которая связана с существованием этой клетки как независимого организма [Доронин и Зозулин, 1976]. У одноклеточных формирование приобретенного поведения происходит за счет макромолекулярных и ультраструктурных перестроек внутри клетки в отличие от многоклеточных существ, у которых память в основном базируется на модификации эффективности синаптической передачи.
Заключение
Опыты на одноклеточных эукариотах Paramecium caudatum показали, что появление новых свойств в окружающем их жизненном пространстве вызывает ориентировочно-исследовательское поведение. Оно выражено изменениями двигательной активности, которая у этих микроорганизмов весьма разнообразна по форме проявления. Физиологический механизм, обеспечивающий ориентировочно-исследовательское поведение, у парамеций может быть только эндонейрональным по причине одноклеточности этих живых орагнизмов. Внутриклеточная регистрация электрической активности парамеций доказывает это предположение. У одноклеточных существ механизм организации поведения базируется на координации эндогенных осцилляторов.
Финансирование Исследование выполнено при поддержке Российского гуманитарного научного фонда, проект 11-06-00917а и проект 12-06-00952а.
Литература
Доронин В.К., Зозулин С.В. Гетерогенность внутриклеточных потенциалов инфузории Spirostomum ambiguum. Журнал эволюционной биохимии и физиологии, 1976, 12(6), 539–43.
Котов Н.В. Двигательная активность Paramecium caudatum: автореф. дис. … д-ра биол. наук. Казан. гос. университет, Казань, 2001.
Коштоянц Х.С, Кокина Н.Н. О действии антихолинэстеразы и ацетилхолина на ритмическую электрическую активность инфузории Opalina ranarum. Журнал общей биологии, 1962, 23(1), 74–76.
Кандель Э. В поисках памяти. М.: Астрель, 2012.
Свидерский В.Л., Лобзин Ю.В., Горелкин В.С, Плотникова С.И. Двигательная активность инфузорий: теоретические и прикладные аспекты. Журнал эволюционной биохимии и физиологии, 2007, 43(5), 379–390.
Соколов Е.Н. Восприятие и условный рефлекс. М.: Моск. гос. университет, 1958.
Соколов Е.Н. Механизмы памяти. М.: Моск. гос. университет, 1969.
Тушмалова Н.А. Память донервных организмов (о физиологических механизмах поведения простейших). В кн.: Соколов Е.Н., Филиппов В.А., Черноризов А.М. (Ред.), Нейрон (обработка сигналов, пластичность, моделирование). Тюмень: Тюмен. гос. университет, 2008. С. 60–72.
Armus H.L, Montgomery A.R., Gurney R.L. Discrimination learning and extinction in paramecia (P. caudatum). Learning and Behavior, 2006, 34(4), 361–365.
Guevorkian K., Valles J.M., Jr. Aligning Paramecium caudatum with static magnetic fields. Biophysical Journal, 2006, 90(8), 3004–3011.
Hinkle D.J., Wood D.C. Is tube-escape learning by protozoa associative learning? Behavioral Neuroscience, 1994, 108 (1), 94–99.
Mirsky A.F., Katz M.S. Avoidance conditioning in paramecia. Science, 1958, 127(3313), 1498–1499.
Mitchell A., Romano G.H., Groisman B., Yona A., Dekel E., Kupiec M., Dahan O., Pilpel Y. Adaptive prediction of environmental changes by microorganisms. Nature, 2009, 460 (7252), 220–224.
Osborn D., Blair H.J, Thomas J., Eisenstein E.M. The effects of vibratory and electrical stimulation on habituation in the ciliated protozoan Spirostomum ambiguum. Behavioral Biology, 1973, 8(5), 655–664.
Schwab-Bonaventure N. The action of chemical substances inverting the sign of galvanotropism in Paramecia. Comptes Rendus des Seances de la Societe de Biologie et de ses Filiales, 1955, 149(34), 395–398.
Van Houten J.L., Yang W.Q., Bergeron A. Chemosensory signal transduction in Paramecium. Journal of Nutrition, 2000, 130(4S Suppl), 946S-949S.
Поступила в редакцию 22 марта 2013 г. Дата публикации: 15 апреля 2013 г.
Сведения об авторе
Греченко Татьяна Николаевна. Доктор психологических наук, ведущий научный сотрудник, Институт психологии Российской академии наук, ул. Ярославская, д. 13, 129366 Москва, Россия. E-mail:
grecht@mail.ru
Источник:
Чтобы оставить комментарии нужно авторизоваться.