Проблема происхождения хлорофилльного фотосинтеза (в дальнейшем - ХФС) имеет большое научное значение, так как именно этот тип усвоения свободной энергии в настоящее время является основным для современной земной биоты. Вместе с тем, насколько известно автору, до сих пор не существует более-менее общепринятой гипотезы его возникновения. Не претендуя, разумеется, на изложение полноценно проработанной теории, я, тем не менее, предлагаю вниманию читателей некоторые свои мысли на тему появления у земных организмов ХФС. Надеюсь, что они помогут хотя бы частично пролить свет на некоторые малопонятные в настоящее время факты.
Начнём с небольшого введения в проблему. Как известно, все системы ХФС можно разделить на 2 большие группы, отличающиеся между собой траекторией движения выбитого световым квантом из хлорофилла электрона. Это так называемые фотосистема I (ФС1) и фотосистема II (ФС2). Большинство бактерий-фотосинтетиков обладают лишь одной из указанных фотосистем, и лишь цианобактерии (или, как их некоторые продолжают называть по старинке, сине-зелёные водросли) обладают обоими указанными системами ХФС, что и позволяет им отщеплять электрон от молекулы воды, что, в конечном итоге, приводит к её распаду на кислород и водород. При этом, следует сразу отметить, что несмотря на различия в судьбе "заряженного" фотоном электрона, самые первые этапы работы обоих систем ХФС по существу идентичны. Так, в обоих случаях реакция начинается с того, что молекула хлорофилла, поглотив квант света, приходит в возбуждённое состояние, вследствии чего легко теряет электрон, перешедший на более высокую орбиту. Далее указанный электрон очень быстро передаётся соседней молекуле хлорофилла (или, в случае ФС2, её слегка модифицированному варианту без иона магния - феофитину), составляющей с первой единый молекулярный комплекс. Следующим переносчиком электрона, получающим его как бы в качестве эстафетной палочки от хлорофилльного комплекса, в обоих системах ХФС является та или иная разновидность особой довольно простой органической молекулы - хинона, например, пласатохинон (кстати, интересно отметить, что другая разновидность хинона - убихинон играет важнейшую роль и в разрабатываемом сейчас под руководством В.П.Скулачёва препарате, призванном замедлить процессы старения организма). Дальше пути электрона у разных фотосистем расходятся, энергия ещё несколько раз переводится из одной формы в другую, но для нас важно отметить две вещи, касающиеся особенностей ФС1. Во-первых, неотьемлимыми составляющими всего комплекса утилизации полученной в результате фотосинтеза энергии (его называют реакционным центром) являются уже хорошо знакомые читателям этого сайта по статье LUCA Минеральные корни современных ферментов железно-серные активные центры, в частности ферредоксины. Во вторых, в случае ФС1 некой промежуточной формой более-менее долгосрочного хранения энергии является одна из основных "энергетических подстанций" клетки молекула NADP, точнее, её восстановленная форма NADPH(см. так же в статью LUCA Реликтовые составляющие современного метаболизма. С чего он мог начаться?). В конечном итоге в обоих фотосистемах полученная энергия как правило используется для того, что бы вытолкнуть протоны против градиента их электростатического потенциала на наружную сторону плазматической мембраны, что бы их впоследствии можно было использовать для синтеза энергии с помощью АТФ-синтазы.
Примечательно, что фотосинтетики распределены между различными классами бактерий очень мозаично, и наиболее общепринятая в настоящее время версия филогенического дерева бактерий никак не объясняет этого странного обстоятельства. Более того, применяемые разными группами исследователей методы восстановления истории эволюции систем ХФС приводят к совершенно разным, практически не согласующимся между собой результатам. Гипотетические варианты истории эволюции ХФС, предложенные в своё время разными учёными, показаны на Рис. 1. Пунктирные линии на этих рисунках соответствуют предпологаемым актам горизонтального переноса генов цепочки ХФС между различными группами фотосинтетиков.

Рис. 1 Различные варианты эволюции систем ХФС (взято из обзора Xiong J, 2007).
(a) - филогеническое дерево фотосинтезирующих бактерий, основанное на сравнении малой субъединицы их РНК (16S RNA); (b) - филогеническое дерево фотосинтезирующих бактерий, построеное на основе анализа аминокислотных вставок в белках; (с) - предпологаемая последовательность событий в истории развития фотосинтетиков, восстановленная на основе анализа генов, участвующих в синтезе бактериохлорофилла; (d) - альтернативный сценарий истории развития фотосинтеза, предпологающий, что в основе их эволюционного дерева находится некая ныне вымершая бактерия, условно названная про-цианобактерией.
Ну что же, попробуем и мы вставить свои две копейки в обсуждение вопроса, по поводу которого разные учёные до сих пор яростно спорят между собой. Начнём с поисков истоков метаболической цепочки синтеза основного элемента любой системы ХФС - собственно молекулы хлорофилла. При этом нужно сразу отметить, что все биологически синтезируемые хлорофиллы делятся на две большие группы - собственно хлорофиллы и бактериохлорофиллы. Не вдаваясь в подробности, можно сказать, что разница между ними заключается в количестве добавляемых в уже почти готовую молекулу протохлорофилла дополнительных протонов (так называемое протонирование). С химической точки зрения это равносильно редукции, а осуществляющие её ферменты называются, соответственно, редуктазами. Так вот, хлорофиллы, содержащиеся в цианобактериях и растениях, редуцируются один раз, а бактериохлорофиллы, содержащиеся во всех остальных фотосинтетиках - дважды. Это очень важный момент, так как природа обладает сильным эволюционным консерватизмом, то есть, она очень не любит переделывать однажды найденные удачные решения. Учитывая это обстоятельство можно предположить, что повторная редукция молекулы хлорофилла появилась эволюционно позже, или, другими словами, что хлорофиллы, используемые цианобактериями, возникли раньше бактериохлорофиллов.
Поверхностный анализ показывает, что есть по крайней мере 8 генов, связанных с синтезом хлорофилла, присутствующих практически во всех без исключения фотосинтетиках. При этом, все эти гены имеют близких родственников (так называемые гомологи) среди генов архей, являющихся, как принято считать, одной из древнейших групп бактерий. Правда, археи не способны к хлорофилльному фотосинтезу, так что, данные гены выполняют у них другие функции. Например, три из указанных генов экспрессируют хелатазы. Так называются белки, вставляющие ион металла в уже заранее построенное для него "гнездо", состоящее из атомов азота - порфириновое или корриновое кольцо. Как уже отмечалось выше, природа крайне ленива (хотя и весьма любопытна). В данном случае, исходя из очень высокой степени гомологии соответствующих генов, это проявилось в том, что она попросту позаимствовала аналогичные гены из цепочки синтеза другой древнейшей органической молекулы - витамина B12 (подпольная кличка - кобаламин), являющегося важнейшим коферментом в одном из самых архаичных (если вообще не самом первом) метаболическом цикле, так называемом пути Вуда-Люнгдала (подробнее про этот путь см. замечательную статью LUCA на этом сайте - Как мог выглядеть архаичный метаболизм?). В ней указанные гены синтезируют протеины, вставляющие в корриновое кольцо кобаламина ион кобальта, но после небольшой "доводки" они, судя по всему, стали способны вставлять в порфириновое кольцо (оно отличается от корринового лишь некоторыми деталями) хлорофилла ион магния.
Ещё три гена, инициирующие добавление в местах двойных связей к порфириновому кольцу атома водорода (см. выше про протонирование) по-видимому были взяты из цепочки синтеза так называемого кофактора F-430 (Boyd et al., 2011), играющего ключевую роль в заключительном этапе основного метаболического пути бактерий-метаногенов. Отметим, что описанные выше заимствования стали возможны только потому, что набор "деталей конструктора", с которыми работает природа, весьма ограничен. Благодаря этому возможные пути эволюции оказываются сильно канализированными, и удачно найденные решения используются впоследствии многократно, причём, зачастую в областях, весьма далёких от их первоначального "внедрения". Желающие ещё раз убедиться в том, что природа активно претворяет в жизнь принцип "экономии мышления", могут ознакомится на Рис.2 с не блещущим большим разнообразием строением уже упомянутых выше витамина B12, хлорофилла c, кофермента F-430, а так же гема B, являющегося активным центром (коферментом) основного переносчика кислорода в организме животных - гемоглобина. Как нетрудно убедиться, по сути во всех четырёх случаях активная часть фермента состоит из иона металла, окружённого кольцом из четырёх атомов азота, в свою очередь, поддерживаемых снаружи неким полужёстким "панцирем", состоящим из атомов углерода. Снаружи к этой конструкции могут быть пришиты различные "побрекушки" (метильные группы, винилы, и т.д.). В зависимости от свойств конкретного металла (соответственно, кобальта, магния, никеля и железа) и особенностей упомянутых "побрекушек" сходные по строению своего скелета молекулы оказываются способными катализировать различные реакции, что, собственно, и позволяет природе широко использовать "наработки", достигнутые в одних метаболических цепочках для катализа реакций в совершенно других цепочках. Для этого часто оказывается достаточным просто скопировать соответствующий ген, а потом лишь слегка доработать его под новую задачу (кстати, у программистов такой метод написания нового кода называется copy-paste, и не очень приветствуется). Например, как уже отмечалось выше, реакционный центр (то есть, то место, где электрон, ранее оторванный от хлорофилла, преобразует свою энергию в другую, более надёжную в хранении и транспортировке форму) ФС1 представляет собой типичный ферредоксин, генов для кодирования которых в каждой клетке - легион.
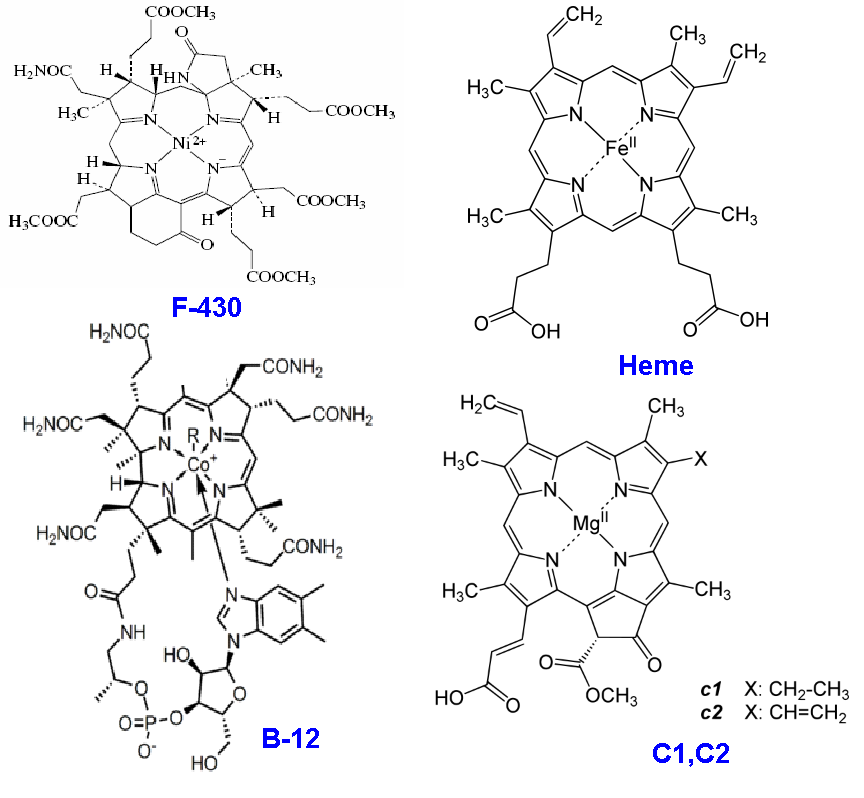
Рис. 2 Однотипные структуры органических молекул, представляющие собой ионы различных металлов, окружённые атомами азота и углерода. F-430 - важный кофермент в цепочке фиксации (усвоения) углерода археями-метаногенами; B12 - кобаламин, важнейший кофермент в цепочке фиксации водорода Вуда-Люнгдала; Heme - активный центр молекеулы гемоглобина; c1, c2 - типичные молекулы хлорофилла c.
Итак, как и следовало ожидать, основные гены, участвующие в синтезе хлорофилла, вовсе не с неба свалились, а, по сути, достались первым фотосинтезирующим бактериям, так сказать, по наследству от других уже существовавших в то время групп бактерий, причём, скорее всего, среди них были археи-метаногены. Им оставалось лишь довести их до ума. Но, всё-таки, какая-же из изображённых на Рис. 1 групп бактерий сделала это первой? По этому вопросу вот уже много десятков лет идут яростные споры и ломаются копья перья. Одна точка зрения состоит в том, что первыми были оксигенные фотосинтетики - цианобактерии. Эта гипотеза косвенно подтверждается, в частности, тем фактом, что цепочка синтеза хлорфилла у цианобактерий короче и проще, чем более более сложная цепочка синтеза бактериохлорофилла у аноксигенных фотосинтетиков (например, как отмечено выше, отсутствует этап вторичного протонирования порфиринового кольца). По существу, синтез бактериохлорофилла аноксигенными фотосинтетиками включает все метаболические шаги, свойственные цианобактериям, в качестве своего первого этапа. Эта более-менее очевидная мысль была высказана уже более 40-ка лет назад Граником (Granick, 1965) когда автор этих строк ещё ходил под стол пешком, и до сих пор (разумеется, в несколько модифицированном варианте) имеет своих сторонников (Mulkidjanian et al, 2006). В свою очередь, Гупта на основе своего метода анализа консервативных аминокислотных вставок в белках предлогает поместить в основание дерева фотосинтетиков гелиобактерий, принадлежащих к классу клостридий (Gupta et al, 1999). Другой известный специалист по истории древней жизни на Земле, Кавальер-Смит "продвигает на трон" группу хлорофлекси, которую он считает не только древнейшим фотосинтетиком, но и самой древней из ныне живущих бактерий вообще (Cavalier-Smith, 2006). Наконец, Хьонг на основе анализа предпологаемой эволюции реакционных центров фотосинтезирующих бактерий отдают пальму первенства в "изобретении" СХФ протеобактериям (Xiong et Bauer, 2002). Итого, среди претендентов на почётное звание первого хлорофилльного фотосинтетика мы находим практически все основные группы бактерий с системами ХФС за исключением хлороби, впору подавать заявку на рекорд в книге Гиннеса! Какая же гипотеза всё же ближе к истине? Попробуем зайти несколько с другого фланга. Достаточно логичным выглядит предположение, что аппарат фотосинтеза бактерий, лежащих у основания филогенического дерева хлорофилльных фотосинтетиков, должен быть устроен наиболее примитивно. Из всех подобных бактерий с секвенированными к настоящему времени (февраль 2012) геномами, наиболее просто система фотосинтеза устроена у двух из них - цианобактерии Gloeobacter violaceus и гелиобактерии Heliobacterium modesticaldum. У обеих указанных бактерий отсутствуют какие-либо выделенные специально для фотосинтеза клеточные органеллы, а всё необходимое для фотосинтеза "хозяйство" встроено непосредственно в их внутреннюю мембрану. Косвенным подтверждением древности указанных бактерий является так же тот факт, что хлорофилл a, используемый всеми цианобактериями, включая Gloeobacter violaceus, содержит больше всего общих с другими хлорофилльными фотосинтетиками компонентов (и, как уже было отмечено выше, является промежуточным этапом при их синтезе), а бактериохлорофилл g, используемый гелиобактериями, по существу отличается от него лишь дополнительным протонированием порфиринового кольца и небольшой модификацией служащего своеобразной антенной "хвоста" (см. рис. 3). Кроме всего прочего, следует отметить, что в построеннных на основе молекулярных данных эволюционных деревьях цианобактерий Gloeobacter violaceus как правило распологается у самого основания дерева.
Итак, по итогам "праймериз" мы имеем двух кандидатов, наиболее подходящих на роль последнего общего предка всех хлорофилльных фотосинтетиков, вышедших во второй тур наших "выборов". Что бы определиться, какой из них древнее, можно воспользоваться одним интересным свойством клостридий - их геномы крайне консервативны, так что, на многих участках длиной в десятки, а иногда и сотни генов, их последовательность практически не меняется. Конечно, и в этих участках иногда присутствуют следы более поздних генетических перестроек - выпадение, или, наоборот, внедрение одного или нескольких новых генов, но в нулевом приближении, по видимому, можно считать, что эти участки генома в основном сохранили структуру и последовательность генов общего предка всех клостридий. Этот факт даёт нам надежду понять некотрые особенности истории возникновения хлорофилльного фотосинтеза вообще, и взаимоотношения двух интересующих нас сейчас бактерий в частности, сравнивая участок генома Heliobacterium modesticaldum, содержащий кластер генов, ответственных за фотосинтез, с аналогичными участками других (нефотосинтезирующих) клостридий. Кстати говоря, сам по себе тот факт, что практически все гены, необходимые для фотосинтеза, у данной бактерии очень плотно сконцентрированы на очень небольшом участке её генома, весьма примечателен. Во многих случаях эти гены достаточно хаотично разбросаны по геному фотосинтетиков. Итак, рассмотрим более внимательно участок генома Heliobacterium, кодирующий белки, необходимые для фотосинтеза. В базе данных американского национального центра биотехнологий (NCBI) этот участок простирается от гена HM1_0654 до гена HM1_0701. В табл. 1 приведено сравнение этого участка вместе с "прилегающими территориями" у бактерии Heliobacterium с аналогичными участками других клостридий. Слева - направо в этой таблице указаны: номер гена, его аннотация в соответствии с данными из базы NCBI, группа, к которой принадлежит ген, в соответствии с данными базы о кластерах ортологичных генов (COG), факт наличия данного гена в данном месте оперона у соответствующей бактерии. При этом, в зависимости от того, на какой нити ДНК расположен этот ген, факт его наличия обозначен символами "+" или "-". Кроме данных для Heliobacterium, приведены так же (слева-направо) данные для клостридий Acetohalobium arabaticum DSM 5501, Symbiobacterium thermophilum IAM 14863, Thermincola sp. JR и Clostridium novyi NT. Гены из "классического" генома клостридий, вероятно представляющие последовательность генов в соответствующем месте генома у их последнего общего предка, представлены в таблице чёрным цветом, а гены, предположительно появившиеся в результате горизонтального переноса или последующей активности мобильный генетических элементов - красным цветом.
1 transсription-repair coupling factor COG1197LK + + + + +
2 ppic-type ppiase domain protein, putative COG0760O + + +
3 stage v sporulation protein t, abrb family transсriptional regulator COG2002K + + + + +
4 stage v sporulation protein b COG2244R + + + + +
5 mazg family protein COG3956R + + + + +
6 DNA-binding protein hu COG0776L + + + +
7 heat shock s4 domain protein COG1188J + + + +
8 hypothetical protein - +
9 hypothetical protein - +
10 septum formation initiator family protein - + + +
11 exopolyphosphatase ppx COG0248FP + + +
12 cobyric acid synthase cobq, putative COG3442R -
13 udp-n-acetylmuramyl tripeptide synthetase, putative COG0769M -
14 hypothetical protein - -
15 glutamyl-tRNA reductase 3 (hemA) COG0373H +
16 hypothetical protein - +
17 hypothetical protein (transposase IS4 family protein ?) - -
18 kda cytochrome c553 (cyhA) COG2010C +
19 hypothetical protein - +
20 rieske iron sulfur protein subunit of cytochrome bc complex (petC) COG0723C +
21 cytochrome b6 subunit of cytochrome bc complex (petB) COG1290C +
22 subunit iv of cytochrome bc complex (petD) COG1290C +
23 cytochrome c, diheme subunit of cytochrome bc complex (petA) COG2857C +
24 hypothetical protein - +
25 tryptophan-rich sensory protein tspo COG3476T +
26 bacteriochlorophyll 4-vinil resuctase (bchJ) COG1719R +
27 bacteriochlorophyll synthetase (bchG) COG0382H +
28 hypothetical protein (Prochlorococcus phage P-SSM7 ?) - -
29 reaction center core polypeptide psha - +
30 magnesium protoporphyrin o-methyltransferase COG2227H +
31 magnesium-protoporphyrin ix monomethyl ester anaerobic oxidative cyclase COG1032C +
31 light-independent protochlorophyllide reductase, iron-sulfur ATP-binding protein COG1348P +
33 light-independent protochlorophyllide reductase, n subunit COG2710C +
34 light-independent protochlorophyllide reductase, b subunit COG2710C +
35 magnesium chelatase ATPase subunit i COG1239H +
36 magnesium chelatase ATPase subunit d COG1239H +
37 magnesium chelatase, h subunit COG1429H +
38 diapophytoene dehydrogenase crtn COG1233Q +
39 stage ii sporulation protein e/serine phosphatase COG2208TK + + + +
40 tRNA(ile)-lysidine synthase COG0037D + + + + +
41 ATP-dependent metalloprotease ftsh COG0465O + + + + +
42 nucleoside diphosphate kinase COG0105F + +
43 abc1 family protein COG0661R +
44 hypothetical protein - +
45 nad dependent epimerase/dehydratase family protein COG1090R +
46 hypothetical protein COG5496R + + +
47 formate--tetrahydrofolate ligase COG2759F + + + + +
48 hypothetical protein COG1418R +
49 dihydropteroate synthase COG0294H + + +
50 folate synthesis, bifunctional protein COG0801H + + +
51 pantoate--beta-alanine ligase COG0414H + + +
52 aspartate 1-decarboxylase COG0853H + + +
53 quinolinate synthetase complex, a subunit (nadA) COG0379H + +
54 l-aspartate oxidase (nadB) COG0029H + +
55 nicotinate-nucleotide pyrophosphorylase (nadC) COG0157H + + +
56 biotin- COG0340H + + +
57 transсriptional activator, putative, baf family COG1521K + + + + +
58 dihydrouridine synthase tim-barrel nifr3 family protein COG0042J + + + +
59 isochorismatase hydrolase, putative COG1335Q +
60 chlorophyllide reductase iron protein, subunit x COG1348P -
61 hypothetical protein - -
62 hypothetical protein - -
63 hypothetical protein - -
64 chlorophyllide reductase, subunit z COG2710C -
65 chlorophyllide reductase 52.5 kda chain, subunit y COG2710C -
66 cytochrome c family protein - +
67 hydrolase, alpha/beta fold family, putative COG0596R +
68 hypothetical protein - +
69 hypothetical protein - +
70 hypothetical protein - +
71 sodium bile acid symporter family, ybas, putative COG0385R +
72 hypothetical protein COG4635CH -
73 hypothetical protein - +
74 hypothetical protein COG5322R + +
75 transсription elongation factor grea COG0782K + + + + +
76 lysyl-tRNA synthetase COG1190J +
77 lysyl-tRNA synthetase COG1190J + + + +
78 hypothetical protein - +
79 hypothetical protein - -
80 reverse transсriptase (RNA-dependent DNA polymerase) COG3344L +
81 hypothetical protein COG4929S -
82 hypothetical protein COG4872S -
83 helix-turn-helix domain protein COG1476K +
84 chaperone clpb COG0542O +
85 transсriptional regulator, ctsr family COG4463K + + + + +
86 uvrb/uvrc domain protein COG3880S + + + + +
87 ATP:guanido phosphotransferase, putative COG3869E + + + + +
88 clpc ATPase COG0542O + + + + +
89 hypothetical protein - -
90 DNA repair protein rada COG1066O + + + + +
91 DNA-binding protein, putative COG1623R + + + + +
Табл. 1 Сравнние участка генома, содержащие гены, необходимые для фотосинтеза, у клостридии Heliobacterium modesticaldum с аналогичными участками нефотосинтезирующих клостридий.
Итак, что же мы видим? Первое, что бросается в глаза, это наличие протяжённых участков генома, уникальных только для бактерии Heliobacterium, причём, если в "общих" для всех бактерий участках генома все гены расположены на одной и той же нити ДНК, то на уникальных для нашего фотосинтетика участках часть генов расположена на противоположной нити. Это сразу вызывает подозрения, что данные участки являются позднейшими вставками, связанными с активностью вирусов и ферментов мобильных генетических элементов бактерий, так называемых транспозаз. При более внимательном осмотре данные подозрения лишь усиливаются. Например, ген под номером 80 в табл. 1 является типичным для ретровирусов, а ген номер 17 при соответствующем запросе к базе NCBI оказывается очень близким гомологом транспозазы IS4. Ещё более интересное открытие ожидает нас при "пробивке" по базе NCBI гена номер 28. При ближайшем рассматрении с вероятностью более 99% этот ген оказывается очень отдалённым гомологом одного из генов в геноме фага (то есть, вируса) Prochlorococcus phage P-SSM7, кстати, почти целиком записанным на противоположной (anti-sense) нити гена, кодирующего реакционный центр фотосинтеза! Почему это важно? Дело в том, что данный фаг является одним из довольно многочисленных вирусов, поражающих цианобактерии, в геноме которого обнаружены гены, ответственные за фотосинтез. Таким образом, довольно естественным выглядит предположение, что по крайней мере в случае с Heliobacterium мы имеем дело с "заражением" изначально нефотосинтезирующей бактерии генами фотосинтеза. По какой-то причине бактерия после внедрения вирусной ДНК в свой геном не погибла, и, более того, даже сумела кое-как приспособить эти гены для своих повседневных нужд, что в итоге и спасло их от исчезновения в результате естественного отбора. Но, пожалуй, самый интересный результат получается для гена номер 22. Гомологи данного гена кроме нескольких клостридий, являющихся ближайшими родственниками Heliobacterium, встречаются лишь у одной альфа-протеобактерии и очень большого числа многоклеточных эукариотов, среди которых преобладают птицы, пресмыкающиеся и земноводные. Как же данный ген, участвующий в цепочке получения энергии путём фотосинтеза смог "просочиться" напрямую в геном высших многоклеточных животных? Возможно, разгадка заключается в том факте, что та единственная протеобактерия, у которой сохранился данный ген (её зовут Вольбахия) с одной стороны является облигаторным паразитом членистоногих и нематод, а с другой стороны, естественно, находится в близком родстве с другими альфа-протеобактериями, среди которых немало фотосинтетиков. Именно от них она могла получить "по наследству" указанный ген, а потом (уже потеряв способность к фотосинтезу) передать его одному из своих "хозяев" - червю нематоде, который, в свою очередь, во многих случаях является обязательным симбиотом высших животных, так что, на конце этой длинной цепочки серии горизонтальных переносов вполне могли оказаться позвоночные. Другой вариант предпологает получение этого гена многоклеточными эукариотами через геном митохондрий которые, как известно, тоже произошли от какой-то группы альфа-протеобактерий.
Итак, вполне резонно можно предположить, что бактерия Heliobacterium получила гены системы ХФС от какой-то группы цианобактерий, возможно, родственных прохлорофитам (в англоязычной литературе обычно используют название Prochlorococcus), путём их горизонтального переноса цианофагами. Но позвольте, зачем вирусам таскать с собой целый кластер генов, на копирование кода которых тратится драгоценная энергия? Если бы это не приносило фагам никакой ощутимой пользы, то, очевидно, данные гены были бы давно равнодушно отсеяны естественным отбором. Дело, скорее всего, в том, что энергия, потраченная на копирование данного кластера генов, впоследствии с лихвой окупается, так как после заражения клетки они могут быть использованы для производства белков, необходимых для извлечения энергии с помощью хлорофилльного фотосинтеза, даже если функционирование соответствующего аппарата был частично разрушено в процессе заражения, либо (в случае наличия полного набора генов для ХФС) даже если у клетки его вообще изначально не было! Очевидно, что доступность энергии является одним из главных лимитирующих факторов, ограничиивающих процесс размножения вирусов, так что, если возить с собой, так сказать, чертёж фабрики для налаживания на месте производства в больших количествах столь необходимой им энергии, это даёт обладающим им вирусам неоспоримое преимущество. Это должно было быть тем более актуальным во времена архея, когда ввиду отсутствия механизма окислительного фосфорилирования энергия была для биоты гораздо менее доступна, чем в наши дни. Это предположение косвенно подтверждается ещё одним фактом. Некоторые цианофаги, паразитирующие на всё той же группе Prochlorococcus, имеют гены для кодирования белков, необходимых для усиленного всасывания клеткой из внешней среды фосфора. Как известно, фосфор является одним из химических элементов, из которых состоит ДНК, так что, его возможный недостаток, точно так же как и недостаток энергии, может являться очень существенным фактором, лимитирующим процесс репликации вирусной ДНК.
Изложенные выше обстоятельства позволяет сделать вывод о большей древности цианобактерий по сравнению с гелиобактериями и объявить, таким образом, победителем нашего конкурса претендентов на звание максимальной близости к последнему общему предку всех фотосинтетиков цианобактерию Gloeobacter violaceus. Кстати говоря, указанная бактерия обладает ещё одной любопытной особенностью. Кроме извлечения солнечной энергии с помощью хлорофилльного фотосинтеза, она способна добывать её и с помощью второго известного в природе способа утилизации энергии солнечных лучей - родописнового фотосинтеза (Balashov et al, 2010), являя собой, таким образом, уникальный пример реализации двух разных типов фотосинтеза, так сказать, "в одном флаконе".
С учётом истории появления фотосинтеза у гелоиобактерий не исключено, что по крайней мере и часть других фотосинтетиков получила соответствующие гены аналогичным образом, что может объяснить отмеченную выше мозаичность их распределения среди различных групп бактерий. Действительно, взять, например, группу бактерий Bacteroidetes-Chlorobi. Она, как легко догадаться, состоит их двух подгрупп, соответственно, Bacteroidetes и Chlorobi, которые при построении филогенических деревьев (это один из способов представить степень родства различных бактерий и предпологаемую историю их эволюции) неизменно отказываются рядом друг с другом, что, вероятно, говорит о том, что у них был общий предок, однако бактерии Bacteroidetes не способны к фотостинтезу, а для бактерий из группы Chlorobi он является их обязательным атрибутом. Если предположить, что одна из бактерий их общего предка подобно Heliobacterium приобрела гены для фотосинтетической "кухни" путём горизонтального переноса от другой группы фотосинтетиков посредством фагов, то это может объяснить появление как бы из ничего фотосинтезирующей подгруппы Chlorobi. Примерно такая же ситуация и с группой Chloroflexi, где часть видов обладает способностью к фотосинтезу, а другая часть - нет. Среди группы любящих кислые среды бактерий (Acidobacteria) недавно был открыт ещё один вид с хлорофилльным фотосинтезом, который пока недостаточно изучен (по этой причине его даже нет на рис.1). Среди самой по себе очень большой группы протеобактерий фотосинтетики представлены достаточно широко, но, опять-таки, не во всех из их подгрупп. Например, среди дельта и эпсилон протеобактерий, которые обычно принято считать наиболее древней их подгруппой, способных к фотосинтезу видов пока не обнаружено. С учётом всего, изложенного выше, наиболее правдоподобным по мнению автора является предположение, что хлорофилльный фотосинтез зародился среди цианобактерий, возможно, близких к современному виду Gloeobacter violaceus. Не исключено, что ещё до этого события данная бактерия уже обладала более примитивным родопсиновым фотосинтезом, что помогло ей быстрее освоить и его хлорофилльную разновидность благодаря, например, наличию средств защиты от ультрафиолетового излучения, что позволило ей находиться очень близко к поверхности воды, тем самым, максимально используя возможности ещё весьма несовершенного в то время хлорофилльного фотосинтеза. Впоследствии гены системы ХФС были разнесены бактериофагами и среди ряда других групп бактерий.
Обсудим теперь вопрос, реакционным центром какого типа обладал общий предок всех современных хлорофилльных фотосинтетиков? В настоящее время считается более менее общепринятым, что имеющая более простое строение ФС1 появилась первой, а ФС2 возникла путём её дупликации и постепенного последующего усложнения. Первичность ФС1 дополнительно подтверждается ещё парой фактов. Во-первых, и её более простой укороченный вариант, не предпологающий запасения в итоге энергии в виде межмембранного протонного потенциала, а ограничивающийся передачей добытой энергии молекуле NADPH, тоже позволяет в какой-то мере удовлетворять энергетические запросы клетки. Во-вторых, многие цианобактерии, обладающие обоими фотосистемами, способны в случае выхода по какой-то причине из строя ФС2, видимо, так сказать, "по старой памяти" переходить на использование только ФС1, в то время как противоположных примеров, когда цианобактерии использовали бы только ФС2, неизвестно.
Следующая важная проблема, с которой желательно разобраться для лучшего понимания эволюции систем ХФС, заключается в выяснении исходного донора электронов для фотосинтеза. Как уже отмечалось в начале статьи, в процессе фотосинтеза от молекулы хлорофилла отрывается электрон. Что бы она после этого продолжала оставаться работоспособной, нужно восстановить статус-кво, вернув ей электрон обратно. Самый очевидный способ это сделать заключается в том, что бы запустить электроны, так сказать, по кругу, возвращая хлорофиллу его же собственные электроны после того, как они передадут запасённую в них энергию системам жизнеобеспечения клетки. Это так называемая циклическая циркуляция электронов действительно по видимому достаточно широко применяется в наиболее примитивных системах хлорофилльного фотосинтеза, например, у гелиобактерий и альфа и бета протеобактерий. Однако, такое решение имеет свои недостатки. Во-первых, КПД никакого реального процесса не может достигать 100%, таким образом, в процессе циркуляции часть электронов на каждом цикле неизбежно теряется, уходя, например, во внешюю среду, что приводит к необходимости заимствовать недостающую часть из других метаболических цепочек. Во-вторых, свободные электроны сами по себе являются в клетке достаточно дефицитным ресурсом, так что, дополнительное пополнение их запаса непосредственно в процессе фотосинтеза вещь очень нужная и полезная. Видимо именно по этой причине многие бактерии перешли от механизма циркуляции электронов внутри фотосистемы к варианту фотосинтеза, при котором кроме производства энергии попутно осуществляется и извлечение столь необходимых клетке электронов непосредственно из внешней среды. В качестве внешнего донора электронов вначале могли выступать те вещества, у которых "оторвать" электрон в энергетическом смысле наиболее просто. Например, водород, сера, некоторые простые органические молекулы. Но по мере уменьшения их концентрации на первый план стала выходить доступность самих веществ-доноров. Вначале наиболее универсальным поставщиком электронов для большинства фотосинтезирующих бактерий стал сероводород, являющийся неким компромиссным вариантом между доступностью и энергозатратностью. Кроме всего прочего, отъём электрона у сероводорода позволяет бактерии дополнительно получить и доступный водород, во многих случаях так же являющийся дефицитным ресуросом. В настоящее время сереводород с удовольствием используют и гамма-протеобактерии, и хлорофлекси, и хлороби, и даже цианобактерии, у которых по каким-то причинам не работает ФС2. Так что, можно предположить, что переход к использованию в качестве донара электронов сероводорода был следующим важным шагом в эволюции хлорофилльного фотосинтеза, позволив кроме решения энергетической проблемы параллельно решить и проблему обеспечения бактериальной клетки дефицитными электронами и ионами водорода. Такое решение можно было бы считать уже почти идеальным, если бы не одно "но". Дело в том, что и сероводород на нашей планете доступен далеко не всегда и не везде. И если на ранних этапах эволюции, в период архея он, по всей видимости, был достаточно широко распространённым веществом, то в дальнейшенм его доступность начала постепенно уменьшаться. Возможно именно это, вкупе с постепенно ослабевающим потоком органики из вулканических и геотермальных источников и заставило биоту искать новые способы добычи столь необходимых для неё свободных электронов и водорода. И здесь сразу по крайней мере по двум пунктам вода оказалась вне конкуренции. Во-первых, она является едва ли не самым доступным вблизи поверхности Земли веществом, запас которого огромен. Во-вторых, кроме того, что она "всегда под рукой", строение её молекул в геометрическом плане очень сильно напоминает строение молекул сероводорода, что, по-видимому, существенно облегчило перенастройку предыдущих систем фотосинтеза, основанных на сероводороде, на воду. Однако, что бы освоить столь привлекальный ресурс, природе пришлось решить и ряд проблем. Одна из них заключалась в том, что для того, что бы растащить на составляющие молекулу воды, нужно затратить примерно в 7 раз больше энергии, чем на разложение сероводорода. При этом, по физико-химическим причинам такую операцию принципиально невозможно реализовать в одном и том же центре хлорофилльного фотосинтеза, нужны минимум два разных центра с разными характеристиками и возможностью передачи энергии между ними. Данную проблему природа решила, объединив в одной клетке две разные фотосистемы, и наладив взаимодействие между ними. Другая проблема заключается в том, что побочным продуктом данной реакции является кислород, являющийся сильнейшим окислителем, вследствие чего он активно разрушает многие важнейшие компоненты клетки, включая ДНК. Для нейтрализации многочисленных соединений, содержащих активные формы кислорода, цианобактериям пришлось вооружиться большим количеством всевозможных антиоксидантов.
Ещё один интересный вопрос, который трудно обойти обсуждая эволюцию систем ХФС, заключается в том, когда и как произошло разделение на хлорофилльный и бактериохлорофилльный фотосинтез. Как уже отмечалось выше, бактериохлорофиллы отличаются от хлорофиллов наличием в процессе их синтеза дополнительной операции протонирования, что меняет спектр поглощения молекулы, позволяя, тем самым, другим бактериям избегать конкуренции с цианобактериями за фотоны, имеющие одинаковые длины волн. Это особенно важно в случаях, когда разные бактериии совместно обитают в тесных бактериальных сообществах, так называемых цианобактериальных матах. Так как вероятность одновременного появления второй редукции молекулы хлорофилла независимо сразу в нескольких группах фотосинтетиков представляеться весьма малой, логично предположить, что это событие произошло в процессе эволюции лишь однажды, а бактериохлорофиллы во всех остальных группах бактерий являются "потомками" той молекулы, в которой это событие благодаря соответствующей мутации в геноме произошло в первый раз. Так как выше мы пришли в выводу, что цианобактерии являются древнейшей группой хлорофилльных фотосинтетиков (что, кстати говоря, в определённой степени подтверждается и палеонтологическими данными), то можно предположить, что общим предком всех бактериохлорофиллов была соответствующая бактерия с наиболее простым фотосинтезирующим аппаратом, бактериофилл которой может быть получен их "базового" для цианобактерий хлорофилла a с помощью минимальной модификации соответствующей метаболической цепочки. Обоим условиям удовлетворяет уже упомянутая выше группа гелиобактерий. По существу, как уже было отмечено выше, не считая дополнительного протонирования кольца, бактериохлорофилл g отличается от хлорофилла a лишь отсутствием протонов в паре стандартных элементов "хвоста" (см. рис. 3.). Все остальные бактериохлорофиллы имеют отличия и в "побрекушках" базового кольца.
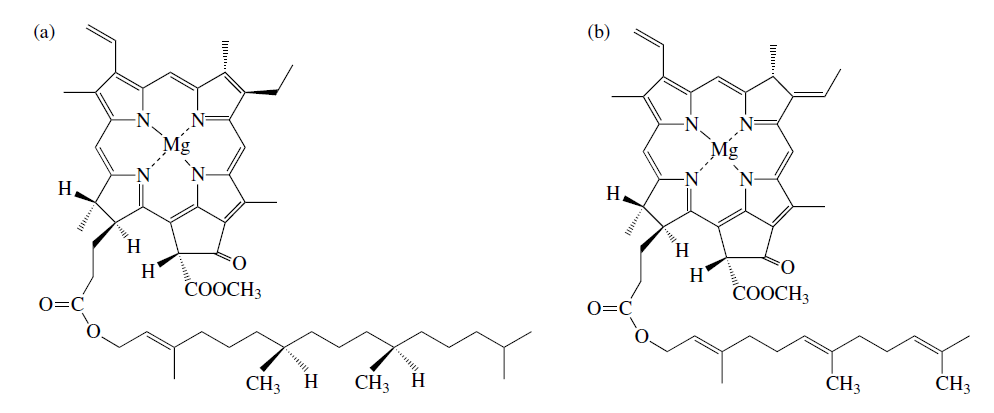
Рис. 3. Сравнение строения предположительно древнейших хлорофилльных молекул. (a): хлорофилл а, непременный (и, в большинстве случаев, основной) хлорофилл цианобактерий; (b): бактериохлорофилл g, единственная светопоглащающая молекула гелиобактерий.
То, что гелиобактерии были первыми фотосинтетиками с бактериохлорофиллом в какой-то мере подтверждает и анализ генома бактерии Heliobacterium modesticaldum. Гены, отвечающие за вторичную редукцию порфиринового кольца (гены номер 60, 64, 65 в Табл. 1) лежат на некотором расстоянии от основного кластера генов фотосинтеза, отделённые от него участком в пару десятков генов из, так сказать, "стандартного генома клостридий" (участок от гена номер 39 до гена номер 58 в Табл. 1), причём, на противоположной по отношению к основному пулу генов фотосинтеза нити ДНК. Это обстоятельство позволяет предположить, что данные гены исходно не принадлежали "заразившему" бактерию аппаратом фотосинтеза бактериофагу, а были перенесены из другого участка генома этой же бактерии (кодирующего, например, редуктазу нитрогенезы, с которой редуктаза хлорофилла имеет достаточно высокую степень гомологии) с помощью мобильных генетических элементов типа транспозаз. По счастливой случайности эти гены в данном месте пришлись ко двору, так как позволили избежать прямой конкуренции вновь появившегося фотосинтетика с цианобактериями за счёт изменения спектра поглащения хлорофилла. Скорее всего, исходно у всех бактерий бактериохлорофилл отличался от цианобактериального хлорофилла a лишь дополнительым протонированием, и лишь впоследствии в него были внесены дополнительные изменения - небольшая модификация "хвоста" у гелиобактерий и добавление к порфириновому кольцу одного дополнительного атома кислорода (с получением, соответственно, бактериохлорофилла a) в случае протеобактерий. Остальные бактерии с более сложными системами ХФС (Chlorobi, Chloroflexi, Acidobacteria) получили их, по видимому, несколько позже, тоже благодаря многочисленным актам горизонтального переноса вирусами, причём, отдельные составляющие системы фотосинтеза могли переноситься и независимо. Так, например, ФС1 проэволюционировала в ФС2, по видимому, у протеобактерий, после чего гены, кодирующие её реакционный центр, были независимо перенесены в группы Chloroflexi и цианобактерий (некоторые цианобактерии до сих пор не имеют ФС2, хотя это может быть и результатом редукции). Специфические структуры, хорошо приспособленные для увеличения эффективности фотосинтеза - хлоросомы, по видимому, были тоже "изобретены" какой-то одной группой бактерий, а потом это полезное изобретение было разнесено вирусами по другим группам.
Итак, в заключение кратко обрисуем возможный сценарий эволюции систем ХФС. Наиболее вероятно, что примитивный ХФС в виде набора из достаточно простого ферродоксина, хлорофилла, выполнявшего изначально функцию защиты бактериальной клетки от разрушительного ультрафиолетового излучения, филлохинонов (типа витаминов К), являющихся одним из основных перносчиков электронов и протонов в схемах анаэробного дыхания, а так же ферредоксин-NADPH редуктазы появился у древних цианобактерий (в то время, ещё не способных к оксигенному фотосинтезу). Некоторые гены, впоследствии адаптированные для ХФС, цианобактерии могли получить от архей-метаногенов. Указанный горизонтальный перенос мог быть облегчён тем обстоятельством, что цианобактерии могут использовать для добычи энергии в том числе, и процессы брожения. При этом в качестве побочного продукта во внешнюю среду выделяется водород, абсолютно необходимый для существования метаногенов. Таким образом, тесное сожительство с цианобактериями в одних и тех же бактериальных матах должно было быть для них крайне выгодно. В то же время, столь тесное сосуществование обычно существенно облегчает горизонтальный перенос генов даже между не родственными группами бактерий. В качестве донора электрона на первых порах могли выступать вначале органические молекулы, водород, сера, а потом и сереводород. ХФС, как весьма ценное изобретение у цианобактерий быстро "хакнули" бактериофаги, для которых возможность иметь "на борту" практически готовую фабрику по производству большого количества энергии, в которой они остро нуждаются для синтеза своих многочисленных копий, оказалась очень кстати. В свою очередь, наличие в геноме многих вирусов кассеты генов для хлорофилльного фотосинтеза привело к многочисленным актам её горизонтального переноса к другим, изначально нефотосинтезирующим бактериям. По видимому, это было основной движущей силой дальнейшей эволюции фотосинтеза. В частности, протеобактерии существенно усложнили устройство реакционного центра, создав прообраз будущей ФС2. Основные гены новой фотосистемы, в свою очередь, были перенесены обратно в геном цианобактерий. Наличие сразу двух реакционных центров фотосинтеза позволило цианобактериям совершить прорыв, перейдя на использование в качестве основого источника электронов широкодоступной воды вместо более дефицитного сереводорода. По видимому, такой переход был облегчён тем обстоятельством, что строение молекул воды и сереводорода достатночно схожее, так что, кардинальной перестройки ФС2 не потребовалось. Далее эволюция фотосинтеза шла, в основном, за счёт совершенствования специальных органелл, в которых энергия фотонов использовалась бы с максимальным КПД. В частности, у цианобактерий появились фикобилосомы, а у какой-то группы бактерий, использующих бактериофилл - хлоросомы, гены кодирования которых впоследствии посредством всё того же горизонтального переноса вирусами быстро распространились и в геномах других "хлоросомных" фотосинтетиков - Chloroflexi, Chlorobi, Acidobacteria.
Литература.
Granick S: Evolution of heme and chlorophyll. In Evolving Genes and Proteins. Edited by Bryson V, Vogel HJ. New York: Academic Press; 1965:67-68.
Всё остальное доступно по прямым интернет-ссылкам. 