Данная статья является второй в серии автопубликаций, которую необходимо читать после ознакомления с первой статьёй Свойства генетического кода - след его возникновения. Крайне желательным для людей, плохо знакомым с основами молекулярной биологии, знакомство со статьёй О.О. Фаворовой "Строение транспортных РНК и их функция на первом (предрибосомном) этапе биосинтеза белков". Важно понимать, для того, чтобы понять КАК возник генетический код, необходимо понять, КАК он функционирует в современных организмах. А для этого необходимо вникнуть в молекулярные механизмы кодируемого синтеза белка. Для понимания данной статьи важно понимать, как устроена молекула РНК, чем она отличается от молекулы ДНК.
Разобраться в теме о происхождении жизни вообще, и возникновении генетического кода, в частности, просто невозможно без понимания основных молекулярных механизмов в живых организмах, в первую очередь двух аспектов - воспроизведения наследственных молекул (нуклеиновых кислот) и синтеза белка. Поэтому данная статья посвящена в первую очередь изложению того минимума знания, с помощью которого можно понять богатый и довольно интересный материал, связанный с происхождением генетического кода (ГК).
Знакомство с молекулярными механизмами синтеза белка лучше всего начинать с изучения структуры одного из ключевых компонентов и одной из самых древних структур в живых организмах - молекулы транспортной РНК (или тРНК). Молекула тРНК имеет необычайно консервативную структуру, которая сходна у всех живых организмах. Эта структура меняется в ходе эволюции настолько медленно, что позволяет нам извлечь немало информации о том, как могли выглядеть древнейшие белок-синтезирующие системы в период их начального формирования. Поэтому говорят, что молекула тРНК является молекулярным реликтом.
Молекулярный реликт, или молекулярное ископаемое - это абстракция, обозначающая древние механизмы и молекулярные и надмолекулярные структуры, встречающиеся в современных организмах, что позволяет нам извлекать информацию об устройстве древнейших живых систем. К молекулярным реликтам относятся молекулы рибосомной и транспортных РНК, аминоацил-тРНК-синтетаз, ДНК- и РНК-полимераз и сам генетический код, как способ кодирования, а также ряд других молекулярных структур и механизмов. Их анализ и является ключевым источником информации о том, как могла возникнуть жизнь, и генетический код, в частности. Рассмотрим подробнее структуру тРНК и те её участки, которые изменяются в ходе эволюции настолько медленно, что ещё содержат немало информации относительно древних тРНК, существовавших более 3,5 млрд. лет назад.
Молекула тРНК относительно небольшая, её длина варьирует от 74 до 95 нукелотидных остатков, наиболее часто - 76 нуклеотидов (см. рис. 1). В последовательности тРНК выделяют так называемые консервативные нуклеотидные остатки – это нуклеотидные остатки расположенные в строго определённых последовательностях почти у всех молекул тРНК. Кроме того выделяются полуконсервативные нуклеотидные остатки – это остатки, представленные только пуриновыми или пиримидиновыми основаниямив строго определённых последовательностях тРНК. Кроме того, различные участки тРНК изменяются с существенно различной скоростью.
До 25% всех нуклеотидных остатков представлены модифицированными нуклеозидами, часто называемых минорными. Минорных остатков описано уже больше 60. Они образуются врезультате модификации обычных нуклеозидных остатков с помощью специальных ферментов.
Среди модифицированных остатков часто встречаются псевдоуридин (5-рибофуранозилурацил, Ψ), 5,6- дигидроуридин (D), 4-тиоуридил и инозин. Структура некоторых модифицированный оснований и частично их роль изложены в статье Молекулярные механизмы обеспечения вырожденности кода
Наряду с первичной структурой (это просто последовательность нуклеотидов), молекула тРНК обладает вторичной и третичной структурой.
Вторичная структура обусловлена образованием водородных связей между нуклеотидами. Ещё в школе учат про водородные связи при комплементарным спариванием между нуклеотидами (A-U и G-C такой вид спаривания нуклеотидов называют каноническим), но в молекулах тРНК также образуется немалое количество неканонических связей, в частности, между G и U, которые будет несколько слабее и энергетически менее выгодная).
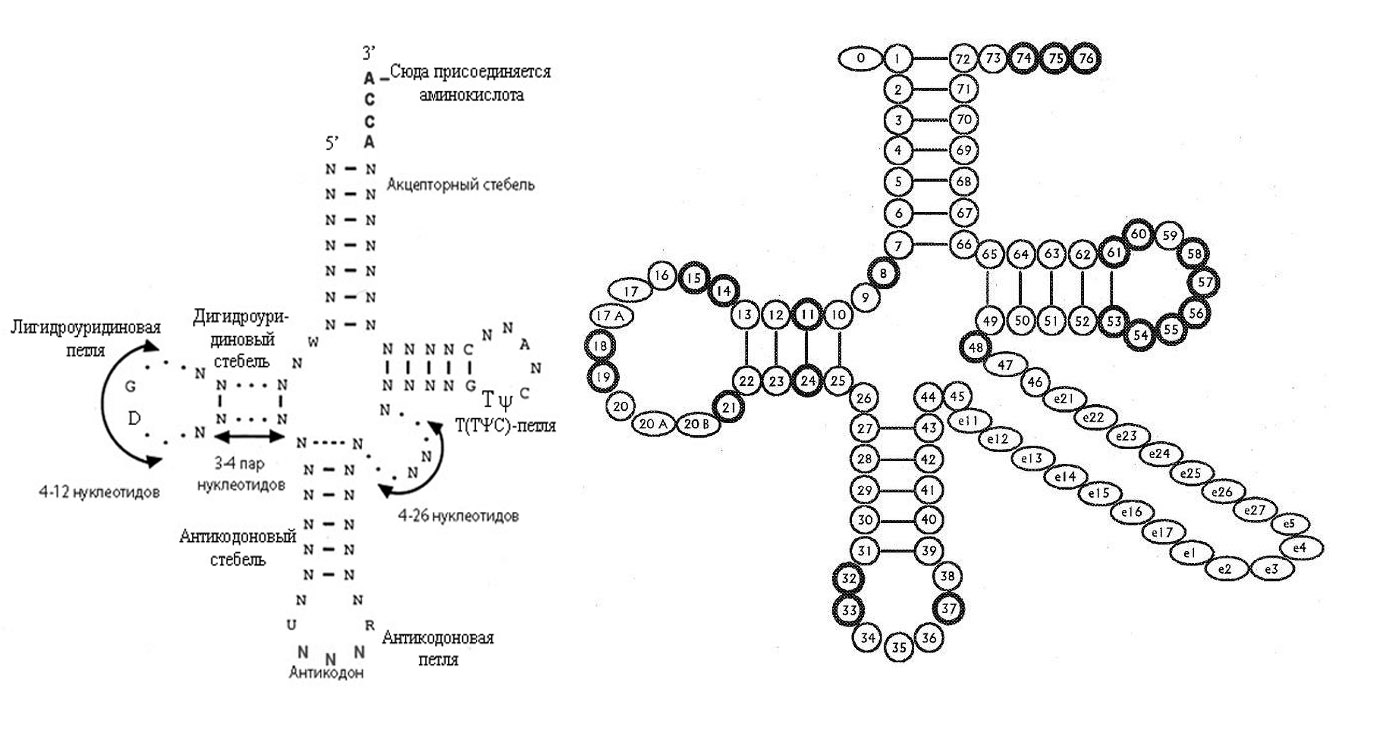
Рис. 1. Обобщённая вторичная структура тРНК (слева) и общепринятая нумерация нуклеотидов в тРНК (справа). Так она выглядит почти у всех живых организмов. На правом рисунке консервативные нуклеотиды выделены жирными кружочками.
Обозначения: N - любой нуклеотид, Т - тимин, D - дигидроуридин, Ψ - псевдоуридин, R - пуриновый нуклеотид.
В результате образуется так называемая структура клеверного листа. В структуре клеверного листа выделяют: акцепторный стебель и три ветви, или домена (arms): антикокодоновую (состоит из антикодонового двуцепочечного стебля (stem) и антикодоновой петли (loop), дигидроуридиновую, или D-ветвь, или D-домен, (также из дигидроуридиновой петли и стебля) и TΨC-ветви, или просто Т-ветви, или Т-домена, (Т-петли и Т-стебля). В дополнение к трём петлям клеверного листа выделяется также так называемая дополнительная, или вариабельная, петля. Длина вариабельной петли варьирует от4 до 24 нуклеотидов.
Почему вторичная структура тРНК имеет Фому клеверного листа? Ответ на этот вопрос дал М.Эйген [Эйген М, Винклер Р. 1979]. Дело в том, что при длине РНКовой цепи 80 нуклеотидов со случайной последовательностью вторичная структура с 3-4 лепестками является наиболее вероятной. Хотя шпилька, имеющая только одну петлю имеет максимальное число спаренныхоснований, эта стуктура в случайных последовательностях является маловероятной. Именно поэтому разумно считать, что тРНК-подобные структуры (то есть структуры с 3-4 петлями) были наиболее распространёнными молекулами на стадии РНКовой и РНК-белковой жизни. Дополнительные доводы в пользу этого утверждения будут приведены в следующих статьях.
Третичная стуктура тРНК.
Третичная структура тРНК соответстует реальной пространственной структуре. Она получила название L-формы, из-за сходства третичной структуры с формой латинской заглавной буквы «L». Третичная структура образуется благодаря взаимодействию элементов вторичной структуры. Веё формировании принимают участие стэкинг-взаимодействия оснований. За счёт стэкинга оснований акцепторный и Т-стебель клеверного листа образуют одну непрерывную двойную спираль, формирующую одну из «палочек» L-формы. Антикодоновый и D-стебли образуют другую «палочку» этой буквы, D- и T-петли оказываются в такой структуре сближенными и скрепляются между собой путём образования дополнительных, часто необычных пар оснований, которые, как правило, образованы консервативными или полуконсервативными остатками. В свете такого участия консервативных и полуконсервативных оснований в образовании L-формы становится ясным их присутствие в T- и D-петлях. Формирование L-образной структуры и её взаимодействие с АРСазой схематически приведено на рис. 2.
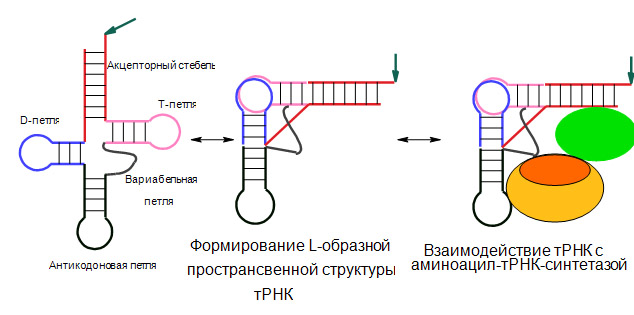
Рис. 2. Схема образования пространственной L-образной стуктуры тРНК и взаимодействия её с АРСазой.
Стрелкой обозначено место присоединения аминокислоты при аминоацилировании тРНК синтетазой. Красным цветом выделен акцепторный домен тРНК, синим – антикодоновый домен. Овалами обозначены домены АРСазы: зелёный – каталитический домен, содержащий домен связывания и аминоацилирования акцепторной области тРНК, жёлтым и оранжевым – вариабельныйдомен АРСазы. В зависимости от размера этого домена, АРСаза распознаёт вариабельным доменом антикодоновую область (домен обозначен жёлтым цветом), или не распознаёт (домен обозначен оранжевым цветом).
Основания антикодона обращены внутрь L-образной молекулы.
Транспортные РНК во всех живых организмах последовательно выполняют три фукнции, необходимые для осуществления синтеза белка:
1) акцепторную - с помощью белковых ферментов (аминоацил-тРНК-синтатаз) ковалентно присоединяет к аминоацильному остатку строго определённую аминокислоту (для каждой аминокислоты - строго своя одна или иногда несколько разных тРНК);
2) транспортную - транспортирует аминокислоту к специфическому месту на рибосоме;
3) адапторную - в комплексе с рибосомой способен специфически узнавать триплет генетического кода на матричной РНК, после чего присоединённая к тРНК аминокислота включается в растущую полипептидную цепь на рибосоме.
Статьи, связанные с темой:
Молекулярные механизмы обеспечения вырожденности кода
Строение транспортных РНК и их функция на первом (предрибосомном) этапе биосинтеза белков