Этапы развития нервной системы
Этапы развития нервной системы
http://www.ngmu.ru/kafedri/normalnoi-fiziologii/umr/recom_uchebniki/Nozdrachev/index.htm
Фокусированное
представительство
Диффузное
представительство
Выделяют несколько видов функциональных асимметрий.
Неодинаковость двигательной активности рук. ног, лица, половин тела,
управляемой каждым полушарием мозга, называется моторной асимметрией.
Неравнозначность восприятия каждым из полушарий объектов, расположенных слева и
справа от средней плоскости тела, именуется сенсорной асимметрией.
Специализация полушарии мозга в отношении различных форм психической
деятельности обозначается как психическая асимметрия.
Так, человек с превалированием
левополушарных функций тяготеет к теории, имеет большой словарный запас и
активно им пользуется, ему присуща двигательная активность, целеустремленность,
способность прогнозировать события. Правополушарный» человек тяготеет к
конкретным видам деятельности, он медлителен и неразговорчив, наделен
способностью тонко чувствовать и переживать, он склонен к созерцательности и
воспоминаниям.
Совокупность индивидуальных особенностей психики и поведения
человека составляет тип высшей нервной деятельности, или темперамент человека
ин складывается из общих свойств нервной системы, которые характеризуются
экстра—интраверсией, эмоциональной
стабильностью — невротизмом, и подвижностью или инертностью нервных процессов.
И. П. Павлов выделял два типа высшей нервной деятельности человека,
которые сейчас могут рассматриваться с позиции межполушарной
асимметрии—мыслительный тип с преобладанием второй (речевой) сигнальной системы
—преимущественно левополушарный и художественный тип с преобладанием первой
сигнальной системы конкретных образов — преимущественно правополушарный
человек. В последнее время получает признание концепция о взаимодополняющем
сотрудничестве двух полушарий и преимуществе отдельного полушария лишь в
определенные стадии той или иной нервно—психической деятельности, а не всей
функции в целом. Правое полушарие быстрее, чем левое, обрабатывает поступающую
информацию, зрительно—пространственный анализ стимулов в правом полушарии
передается в левое полушарие (в моторный «центр речи»), где происходит
окончательный, высший семантический анализ и осознание раздражения.
Согласно концепции В. Л. Бианки, в процессе обучения правое
полушарие работает по принципу дедукции, т. е. сначала осуществляет
синтез, а затем анализ; левое же полушарие функционирует по принципу индукции,
сначала анализируя раздражители, а затем синтезируя их (рис. 3.57).
В эволюции человека речь первоначально лишь использовала, а
затем развила и качественно преобразовала уже имеющуюся у животных
функциональную межполушарную асимметрию.
|
|
|
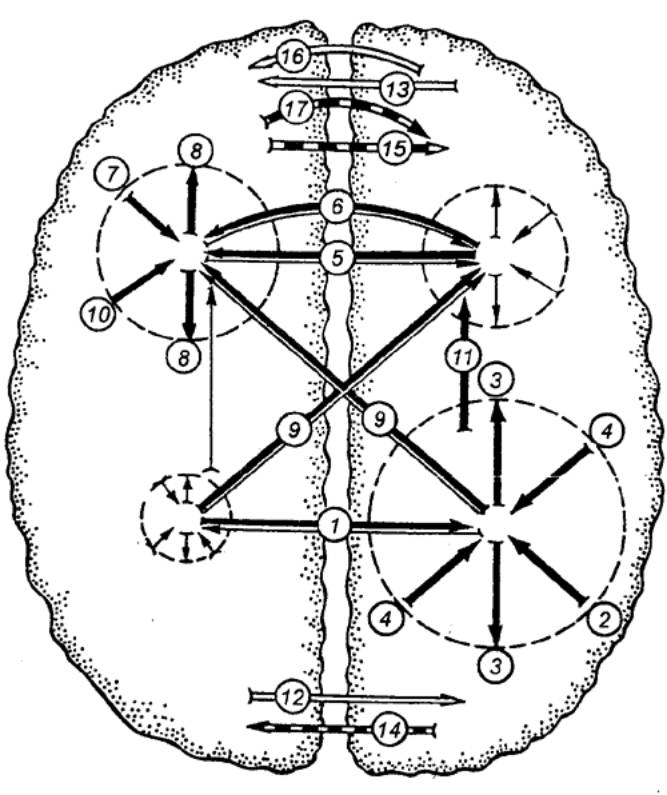
Рис. 3.57 Схема межполушарных взаимоотношений (по Бианки) Вверху — ассоциативная кора, внизу
— проекционная; слева — левое полушарие; справа — правое; жирные стрелки —
доминирующие влияния, тонкие — недоминирующие; белые стрелки — облегчающие
влияния; прерывистые — тормозящие; 1,5
— транскаллозальные (через мозолистое тело) влияния; 2,7, 10 — восходящие афферентные
влияния; 3, 8 — дивергенция
возбуждения; 4 —
конвергенция; 6 —
экстракаллозальные влияния; 9 —
межзональные транскаллозальные влияния; 11 — межзональные внутриполушарные влияния; 12, 13 — транскаллозальные
облегчающие влияния; 14, 15 —
транскаллозальные тормозящие влияния; 16
— экстракаллозальные облегчающие влияния; 17 — экстракаллозальные тормозящие влияния. |
3.21. ФОРМИРОВАНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ РЕБЕНКА
Современные представления о нервных
механизмах поведенческих и психических процессов строятся на признании определяющей
роли ассоциативных систем мозга. Эти образования мозга участвуют в
выполнении многих высших корковых функций: программировании целенаправленных
движений и действий, многих интеллектуальных операциях, познавательных
процессах, организации системы речевого высказывания, наглядных
пространственных и символических синтезов, формировании кратковременной
пространственной и речевой памяти.
Формирование высшей нервной деятельности ребенка тесно
связано с динамикой созревания ассоциативных корковых структур. Ребенок в
общении со взрослыми усваивает определенные знания, приобретает новые формы
отношений к внешнему миру. Все это строится на основе создания новых
функциональных связей, которые позволяют овладеть новыми формами восприятия и
запоминания, видами мышления, способами организации движения. Их структурной
основой выступают лобные и нижнетеменные области коры, которые в
онтогенезе человека увеличиваются по площади в 9 раз и у взрослого занимают
|
|
|
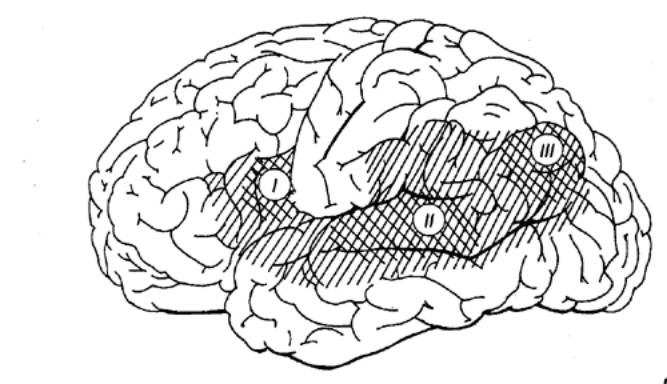
Рис. 3.58 Топографическое
расположение моторного (зона Брока, I), слухового (зона Вернике, II) и
зрительного (III) центров речи левого полушария мозга человека |
|
|
|
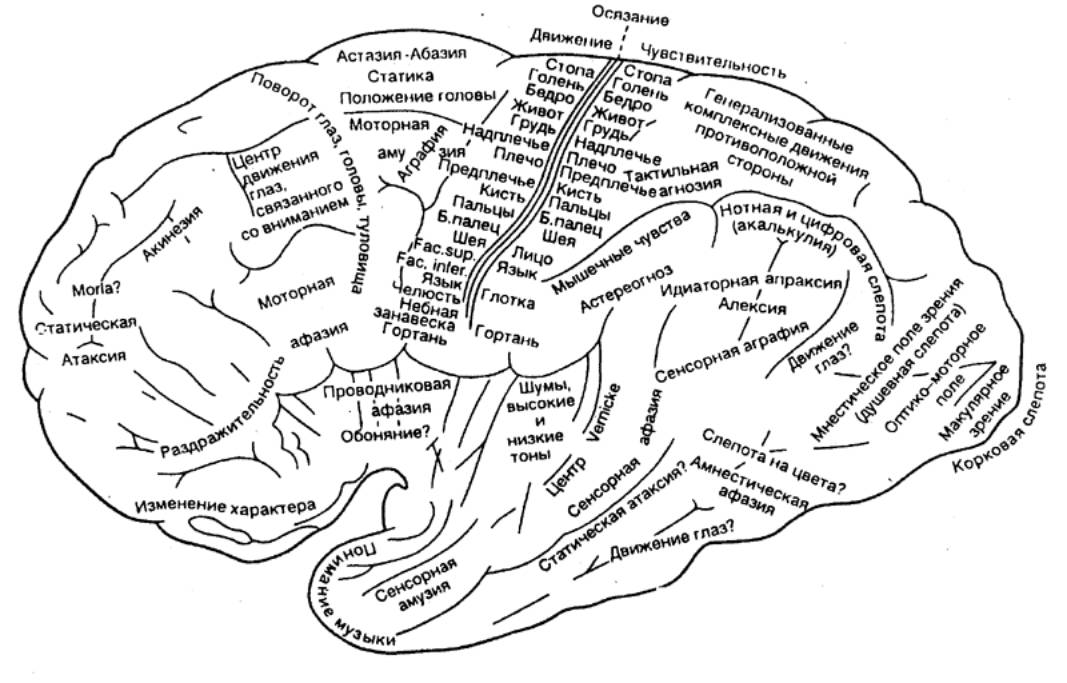
Рис. 3.59 Локализация
функций в коре большого мозга человека |
около 2/3 всей поверхности новой
коры. Кроме того, у человека в онтогенезе развиваются специфические
человеческие корковые формации, которые имеют специальное отношение к анализу и
синтезу раздражении, связанных с разными сторонами речевой деятельности (речедвигательной,
речеслуховой, речезрительной) (рис. 3.58, 3.59).
Установлено, что в течение первого года жизни ребенка
количество условных рефлексов на непосредственные раздражители постепенно
увеличивается. Вначале формируются только вегетативные условные рефлексы, затем
двигательные и, наконец, появляются речедвигательные. Наличие последних не
свидетельствует еще о появлении словесного мышления, так как нет еще отвлечения
и обобщения при помощи слов.
Исходно слово выступает как слабый компонент комплексного
раздражителя и имеет второстепенное значение. Однако постепенно при условии
частого повторения слово как наиболее постоянный компонент комплекса
приобретает значение сильного компонента и, наконец, самостоятельного
сигнала, хотя к концу 1—го года жизни ребенка первые воспринимаемые им
слова — это просто
.
|
|
|
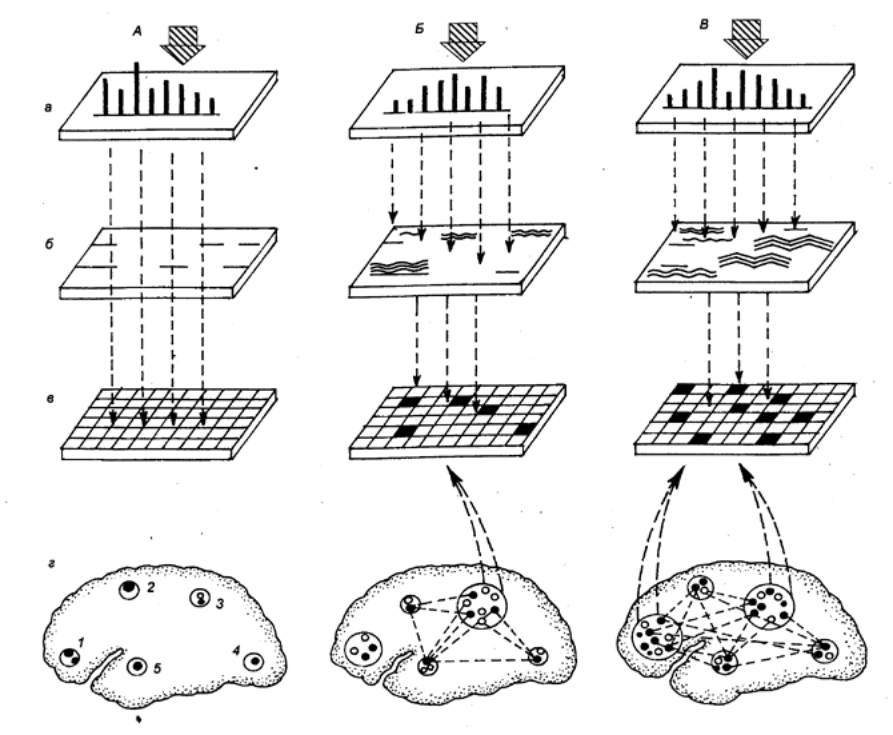
Рис. 3.60 Формирование механизмов восприятия слова у детей в процессе развития А —
новорожденные; Б—в возрасте 3—10 месяцев. В—в возрасте 2—3 лет: а — уровень рецептивных
преобразований, б — уровень
фонемного перекодирования слова, в — уровень семантического анализа слова, г — зоны мозга, участвующие в
семантическом анализе слова; 1 —
лобные зоны, 2 — моторные, 3
— нижнетеменные, 4 —
затылочные, 5 — височные; Заштрихованной стрелкой обозначено воздействие
слова. |
звуковые обозначения определенных
предметов и лиц. Позже, в следующем возрастном периоде, происходит превращение
слова в абстрактный сигнал, обозначающий совокупность предметов.
На 2—м году жизни возникают образы отдельных предметов.
В результате действия ребенка с предметами они вычленяются из обобщенного
недифференцированного мира. Формирование речевых реакций невозможно без
звукового словесного подражания. При этом особую роль играет установление связи
(ассоциации) слова с видами предмета, его осязанием и другими признаками.
Различают понимание речи (сенсорная речь) и ее воспроизведение (моторная речь).
Вначале возникает понимание речи, затем оба эти процесса теснейшим образом
увязываются.
Развитие обобщающей функции слова
проходит через следующие основные этапы.
I степень интеграции — слово
замещает чувственный образ определенного предмета, т. е. оно является
эквивалентом одного конкретного предмета («ляля» — только данная кукла; конец
1—го — начало 2—го года).
II степень интеграции — слово
замещает несколько чувственных образов однородных предметов («ляля» относится к
нескольким куклам; конец 2—го года).
Ш степень интеграции — слово
замещает ряд чувственных образов разнородных предметов («игрушка» — это и
куклы, и мячи, и кубики; не раньше 3—го года).
IV степень интеграции — в слове
связан ряд обобщений предыдущих степеней (например, слово «вещь» содержит
обобщение, даваемое словом «игрушка», «одежда», «еда» и др.; 5—й год жизни).
С помощью электроэнцефалографических
исследований установлена динамика функционального созревания основных корковых
зон у детей. Ведущим звеном в организации межцентральных отношений выступают
ассоциативные отделы неокортекса — нижнетеменные и лобные зоны (рис. 3.60). У
детей 1—го года жизни фокусами взаимосвязной активности являются нижнетеменные
зоны, которые устанавливают связи с затылочными, височными и моторными центрами
коры. У 2—летних детей фокусы взаимосвязной активности перемещаются в лобные
отделы, которые вступают в синхронные взаимоотношения с другими центрами коры.
У детей 4—5 лет отмечается усиление межцентральных взаимодействий нижнетеменных
полей. На более поздних этапах онтогенеза (6—7 лет) вновь происходит усиление
межцентральных связей лобных областей с другими отделами коры.
V годовалых детей
только при опознании знакомого слова усиливаются связи нижнетеменных и височных
областей коры левого полушария. На 2—м году жизни при опознании знакомого слова
усиливаются связи двух ассоциативных областей — нижнетеменных и лобных
преимущественно левого полушария.
У детей 4—5 лет при восприятии
знакомого слова связи левой лобной и левой нижнетеменной зон еще более
усложняются. Такая динамика отражает морфофункциональную зрелость высших
интегративных систем мозга ребенка.
Начиная с 6—летнего возраста ребенок
в состоянии управлять своим поведением на основе предварительной
словесно—обобщенной инструкции. К этому возрасту он способен удерживать
программу действий, состоящих из ряда движений. Поведение ребенка начинает в
большей степени определяться степенью вероятности тех или иных событий в
окружающей среде. К этому возрасту более ярко проявляются типологические
особенности высшей нервной деятельности (по признакам силы, уравновешенности и
подвижности процессов возбуждения и торможения). К 10—11—летнему возрасту
развитие коры головного мозга приближается к уровню взрослого человека, что
является важнейшим фактором формирования высших нервных и психических функций
ребенка.
Мышление — сложнейший вид мозговой
деятельности человека в процессе приспособления к новым условиям и при решении
новых жизненных задач. Процессы мышления сводятся к образованию общих
представлений и понятий, а также суждений и умозаключений. Помимо
словесно—логического, абстрактного существуют формы эмоционального мышления
(оценка), практического, или наглядно—действенного, мышления.
Среди множества теорий, пытающихся объяснить
природу мышления, наиболее распространены две: одна рассматривает мышление
как беззвучную речь, как выражение работы речевого механизма (и механизма
письма), а у глухонемого — механизма жестикуляции; вторая — считает мышление
своеобразным чисто психическим процессом, лишенным образности и не совпадающим
с внутренней речью. Полного подтверждения не получила ни одна из этих гипотез.
Раскрытие действительной сложности обоих процессов:
превращения мысли в речь и извлечения основного содержания (мысли) и речевого
сообщения — было связано с успехами психологии и лингвистики.
Мысль является сложнейшим обособленным отражением
действительности, направляющимся известными мотивами, т. е. это специфический
процесс, сформированный вещественно—историческим развитием на базе той роли,
которую играет язык в истории человечества. Поэтому мысль, которая на начальных
этапах истории сама являлась конкретной развернутой деятельностью и которая
лишь со временем превратилась в свернутый внутренний процесс, не может
рассматриваться как изначальный «духовный» акт.
В онтогенетических исследованиях установлено, что на ранних
этапах развития ребенка всякое препятствие, формирующее «задачу», вызывает
возникновение внешних проблем, в которые включается анализирующая и планирующая
внешняя речь. На последующих этапах развития эта развернутая внешняя речь,
включающаяся в действие и способствующая решению задачи, переходит в шепотную
речь, и именно эта внутренняя речь, еще более концентрируясь, становится
основой глубинного процесса, который на конечных стадиях превращается в мысль.
Восприятие речи является важнейшей стороной функционирования
мозга человека. Обработка речевого сигнала проходит по законам сенсорных
систем, хотя обладает специфическими особенностями, определяемыми сложной
иерархической структурой речи. Речь как сигнал имеет два класса признаков:
собственно физические признаки (фонемы, слоги) и информационные (просодические)
признаки, состоящие в интегрировании отдельных сегментов в более крупные
единицы — слова, фразы.
В физиологии речевой функции обнаруживаются основные
признаки отражения внутреннего (субъективного) и внешнего (объективного) мира.
Это должно включать в себя прежде всего механизм выделения существенных
признаков внешних стимулов и отношения этих признаков, которые формируют основу
для обобщения. Другой составной частью механизмов речевой функции является
обеспечение единства целенаправленной деятельности, сохранение задачи и
интеграции замысла, лежащих в основе внутренней свернутой речи, которая
является базой для реализации развернутой (экспрессивной) речи. Функция речи
выполняется при обязательном взаимодействии этих двух механизмов: гностического
и динамического.
Локальные поражения левого полушария у правшей приводят к
нарушению речевой функции в целом, а не к частичному выпадению какой—либо одной
ее формы. Распад экспрессивной речи, связанный с нарушением динамических схем
слов, неизбежно вызывает распад единства звука и смысла и, как следствие этого,
нарушение понимания речи.
Клинические исследования механизмов речепродукции и
речевосприятия указывают на существенную роль кортикальных формаций прежде
всего левого полушария и подкорковых образований — различных структур переднего
и промежуточного мозга. Существует предположение о конвергенции механизмов,
обеспечивающих раздельное восприятие и продукцию речи, которая усиливается от
нижних этажей мозга к высшим, достигая максимума на элементах лобного полюса
коры.
|
|
|
Рис.
3.61 Электроэнцефалографические
стадии сна, зарегистрированные у ребенка 12 лет А—Е — обозначения соответствующих стадий по классификации
Лумиса, Р — парадоксальный
сон. |

Для спокойного бодрствования
и дремоты (рис. 3.61) характерен α—ритм, амплитуда которого постепенно
уменьшается и может вообще исчезать (стадия А). При внешнем раздражении
наступает реакция активации, т. е. происходит смена α—ритма на
высокочастотный (β—ритм (реакция десинхронизации). При появлении легкой
сонливости (стадия В) возникает неустойчивая высокоамплитудная
активность. Стадию С связывают с поверхностным сном, когда начинают
появляться низкокоамплитудные медленные волны. Переход к стадии D и особенно Е свидетельствует
о глубоком сне, который сопровождается дальнейшим усилением
высокоамплитудной медленной активности (S—ритм). Типичная картина
парадоксального сна (ПС) — низкоамплитудная нерегулярная активность с
непостоянными редкими группами α—ритма и вспышками острых пилообразных волн.
Общая картина сна детей существенно
меняется с возрастом: если новорожденный бодрствует около 4 ч/сут, то к концу
первого года жизни время бодрствования возрастает до 10 ч/сут. У взрослых в
течение ночи отмечается 4—6 циклов ПС длительностью каждый по 20 мин. Если у
новорожденных общая продолжительность быстрого сна занимает около 10 ч, то у
взрослых — только 1,5 ч. Однако значение ПС для обеспечения продуктивного
бодрствования с возрастом увеличивается. Продолжительность сна взрослых людей
очень индивидуальна.
Сновидения. Сновидениями интересовались испокон
веков. Их трактовка дается еще в ведических текстах древней Индии. Мощнейший
импульс к экспериментальному изучению сновидений дало открытие ПС. Было
установлено:
1) каждый человек видит сны много
раз в течение ночи; 2) насыщенность сновидения событиями примерно соответствует
реальной длительности сна; 3) испытуемые отчитываются о сновидениях в 80%
случаев при пробуждении после ПС и лишь несколько реже — при пробуждении во
время МВС; 4) во время ПС доминируют яркие, сложные, странные, эмоционально
окрашенные и длительные сновидения, тогда как во время МВС — кратковременные
рациональные и реалистические сны, более сходные с мышлением в бодрствовании;
5) переживание сновидений возникает также при засыпании и при пробуждении,
ночные и дневные грезы близки и по характеру, и по содержанию; 6) при
пробуждении из глубокого МВС отмечаются сонливость, дезориентация и нарушение
памяти, тогда как при пробуждении из ПС — быстрая ориентация и готовность к
действию; 7) более одной трети сновидений содержат эмоции страха и тревоги;
поражения, неудачи и конфликты случаются в сновидениях чаще, чем
удовлетворение, успех и дружеские контакты; странности, фантастика и обыденные
ежедневные дела встречаются нечасто; большинство сновидений банальны и
неинтересны; 8) в сновидениях отсутствуют элементы воображения и трезвая оценка
невероятности развертывающихся событий; 9) сновидения быстро забываются, кроме
того, сновидения имеют особую атмосферу, которую трудно передать словами.
Гипотез о природе сновидений
множество. 3. Фрейд сравнивал сновидения с «работой сна» и рассматривал их как
особый важный язык мозга. Он был твердо убежден, что «толкование сновидений —
это основной путь к познанию бессознательного». Во многих гипотезах
подчеркивается, что состояние сновидений обусловливается в основном активностью
самого мозга. Необыкновенный, пестрый характер сновидений может быть
результатом синтеза мозгом противоречивой информации при одновременной
активации различных нейронных сетей. Ряд авторов рассматривает сновидения как
процесс пересмотра данных, хранящихся в памяти. Молекулярные же биологи
полагают, что сновидения способствуют «очистке» памяти от ненужной информации и
тем самым поддерживают эффективность работы нервной системы. Эти и многие
другие гипотезы трудно подтвердить и опровергнуть. Окончательный ответ о
биологическом значении сновидений еще не найден.
Проблема сновидений — наиболее
загадочная область человеческой психики. Сны преимущественно носят зрительный
характер. У слепорожденных в снах отсутствуют зрительные образы и преобладают
осязательные. На характер сновидений оказывает влияние профессиональная
деятельность человека: музыканты чаще переживают чисто слуховые сновидения.
Отмечается зависимость сновидений от уровня интеллектуальности, пола, возраста,
утомления.
В целом сновидения отражают
имеющиеся у личности проблемы, а материалом является конкретный опыт,
накопленный в течение жизни. И. М. Сеченов называл сновидения «небывалыми
комбинациями бывалых впечатлений». Установлено, что нет людей, не видящих сны.
Сновидения возникают в среднем 4—6 раз за ночь. Если пробуждение наступает в
стадии быстрого сна, 70—90% людей детально и достаточно эмоционально
рассказывает о своих сновидениях, а если в медленном — лишь 7—10%. При этом
психические процессы медленного сна скорее выступают как мышление, пересказ
событий прошедшего дня.
Сновидения иногда рассматривают как
следствие растормаживания нервных следовых процессов, связанных с прошлым
жизненным опытом и социально обусловленных по своему содержанию. Для понимания
механизмов извлечения из памяти и направленного характера сновидений может быть
использована теория доминанты. Скрытые корковые или подкорковые доминанты могут
выступать в качестве основных факторов сновидений, а сами последние могут
возникать со всеми признаками доминанты (стереотипные навязчивые сновидения).
В. М. Бехтерев при анализе
диагностического значения сновидений исходил из следующей позиции: в
сновидениях происходят явления, которые развиваются из того же источника, что и
внутренние переживания в состоянии бодрствования. Сновидения указывают на
наличие в прошлом или в настоящем определенных физиологических или психических
состояний.
Гипноз и внушение. Гипнотическое состояние является
сноподобным состоянием с сохранением речевой связи (раппорта) гипнотика с
гипнотизером. Различают три стадии гипноза: 1) сонливость, при которой
гипнотик может противостоять словесному внушению и открыть глаза; 2) гипотаксия
легкий сон, когда он не может открыть глаза и подчиняется внушению; 3)
сомнамбулизм — глубокий сон, когда гипнотик полностью подчинен гипнотизеру, у
него развивается амнезия (забывание предыдущих событий) после пробуждения.
Гипнабельность, т. е. подверженность гипнозу, степень
и вид внушаемости, зависит от возраста, пола, интеллекта, здоровья, т. е. от
многих индивидуальных качеств личности. Внушение под гипнозом — лишь пример из
общего учения о внушении, которое свойственно каждому человеку. При внушении
происходит вторжение в сознание посторонней идеи или приказа без участия воли и
внимания реципиента на подсознательном уровне его психики. Этим последним
внушение отличается от убеждения, которое формируется с участием сознания. В
отличие от внушения извне существует еще самовнушение, при котором в сознание
внедряется не только определенная мысль, но и эмоциональное ее переживание.
Чистого самовнушения без воздействия на личность извне не бывает, поэтому
источником процесса самовнушения всегда является внушение, т. е. внешние
факторы. Механизм гипноза остается неизвестным, а завершенной теории гипноза
еще не разработано, хотя целый ряд заболеваний успешно излечивают под гипнозом.
Гипноз иногда рассматривают как специфическую разновидность
бодрствования, полагая, что внушенные гипнотический сон и какие—либо действия —
явления одного порядка. Действительно, ЭЭГ человека, находящегося под гипнозом,
сходна с ЭЭГ человека в состоянии бодрствования; гипнотическое внушение
подчиняется закономерностям условнорефлекторной деятельности; у ряда лиц в
состоянии бодрствования можно получить все эффекты гипнотического внушения.
В психологическом плане в основе
гипноза лежит моральная капитуляция человека, который некритически подчиняется
приказу гипнотизера. Действительно, снижение волевых свойств личности,
повышенная тревожность усиливают внушаемость. Во время гипноза высшая нервная
деятельность существенно меняется. Это касается сенсорной сферы, когда можно
внушить отсутствие слуха, зрения, осязания и даже боли, можно получить извращенные
реакции (иллюзии), вызвать галлюцинации (восприятие несуществующих предметов и
явлений или исключить из восприятия определенные предметы). Гипнотику можно
внушить другой его возраст, различные эмоциональные состояния, воздействовать
на чувства, сферу его внимания, но нельзя изменить личность человека вопреки
его социальным и индивидуальным убеждениям.
3.24. ТРУДОВАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА—ОПЕРАТОРА
Современное производство оснащено
значительными техническими средствами, и на человека—оператора, входящего составной
частью в эту систему, ложатся задачи по контролю, управлению и регулированию
целыми комплексами. В то же время он освобождается от физических нагрузок, а
доля умственного труда существенно увеличивается. Оператор в такой системе
становится интегральным звеном управления, поэтому его характеристики
предопределяют успешность и надежность функционирования данной системы.
Ярким примером является водитель
автотранспорта, который воспринимает показания различных приборов в кабине,
сигналы от дороги, внешнего окружения, пешеходов. Он также воспринимает сигналы
от мышц собственного тела, шум двигателя и др. Основываясь на анализе этих
сигналов, разделяя их на значимые и незначимые, он принимает решение
относительно
|
|
|
Рис. 3.62 Система человек—машина,
работающая по замкнутому и открытому контурам управления |
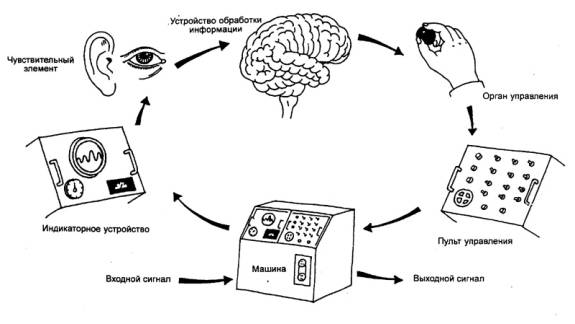
воздействия на органы управления
автомашиной. Последняя вновь подает информацию о новом режиме движений —
образуется так называемая замкнутая система человек—машина (рис. 3.62).
Такие системы составляют большинство и в других областях человеческой
деятельности.
Следовательно, задача
человека—оператора состоит в переработке за короткое время потока различного
вида информации и быстром принятии единственно правильного решения. Такая
ответственность повышает эмоционально—психическую напряженность работающего
человека, а также социально—экономическую значимость его труда для общества.
Обработка информации
человеком—оператором.
Если человека сравнить с компьютером, то у него больше недостатков, чем
преимуществ. Но достоинства человека оказываются решающими, особенно его
способность выполнять задачи логико—индивидуальными методами, что не требует в
каждом случае трудоемкого предварительного программирования. Практический опыт
и полученное образование уже предполагают наличие таких программ. Человек легко
ориентируется в непредвиденных ситуациях, мгновенно восстанавливает в памяти
предыдущие факты и методы решения проблем, ему не нужно специальное кодирование
информации и трансформации ее в числовую форму.
Вместе с этим человек имеет ряд
существенных ограничений, создающих значительные проблемы в его работе. Сюда
относится и ограниченность ресурсов памяти, и медленная скорость обработки
информации, и, что самое главное, подверженность влиянию утомления, измененного
функционального состояния, настроения и других психофизиологических факторов,
которые ухудшают его способность к оценке информации и быстрому принятию
правильного решения.
Диктуемая практикой необходимость
отражать на дисплее все больший объем информации вынуждает изыскивать
возможности расширения ассортимента размерностей и повышения плотности
размещения кодовых знаков. Легче всего кодируется цвет, затем форма, яркость,
размер. Звуковые сигналы оказываются наилучшими для привлечения внимания к
мгновенной опасности. Слуховая информация лучше передает простое, короткое
сообщение, требующее немедленного действия. Информация о последствиях тех или
иных действий оператора образует обратную связь, обеспечивающую саморегулирование
системы.
Рабочие движения человека—оператора. Рабочие движения являются
компонентами деятельности человека и включаются в структуру этой деятельности.
Физиологически они обеспечиваются тремя факторами: динамической формой
мышечной деятельности (собственно движение), статической мышечной работой
(поддержание рабочей позы) и координацией элементов статики и динамики в
необходимых соотношениях.
Эффективность деятельности оператора зависит больше от
совпадения характеристик сочетания сигнал—ответ, чем от специфических черт
отдельно взятых приборов и ответных движений. Сочетание сигнал—ответ должно
основываться на фундаментальных привычках, т. е. полностью согласовываться с
личными и групповыми стереотипами. Рациональная организация рабочих движений
создает условия для снижения утомления и повышения работоемкости, но вместе с
этим требует соответствующей конструкции органов управления.
Временная структура
деятельности оператора. Временная структура деятельности может быть либо свободной,
ориентированной на точностные, качественные показатели, либо навязанной,
когда главной становится скорость выполнения операции. Как правило, оба эти
показателя присутствуют совместно, неразрывно, требуя точности реагирования и
быстроты выполнения. Каждому оператору приходится работать в условиях
определенной степени риска, что вызывает эмоциональную напряженность, резко
возрастающую в аварийной ситуации. В этом случае временной фактор является
решающим, а правильное и отчетливое ориентирование во временном режиме имеет не
меньшее значение, чем точность выбора реакции. Трудовая деятельность строго
нормирована по объему труда и длительности ее осуществления. При разработке
режимов труда и отдыха учитывают периодические колебания работоспособности
человека. Особое значение это положение приобретает для организации сменного и
ночного труда, где суточная периодика работоспособности становится важным
фактором деятельности человека.
Индивидуальные
особенности человека—оператора. Эмоциональное напряжение, вызываемое условиями труда и
сложностью работы, а также сильная эмоциональная реакция, вызванная, например,
аварийной ситуацией, по—разному влияют на оператора. Адекватность его поведения
во многих отношениях будет определяться индивидуальными особенностями. Состояние
эмоционального стресса может проявляться в двух формах: возбудимой,
которая характеризуется повышенной двигательной активностью, суетливостью,
излишней говорливостью и т. д., и тормозной, для которой характерны
угнетение деятельности, замедление реакции, неспособность отвечать на различные
воздействия.
Положительное влияние
эмоционального стресса проявляется в мобилизации мозговой активности. Особенно
отчетливо это выражено в условиях, когда необходимо прогнозировать вероятный
исход своих действий и, соответственно, вероятность появления тех или иных
сигналов.
Исследования
прогностической деятельности человека в вероятностно организованной среде
показывают, что существует несколько принципиально различных тактик работы, а
успешность деятельности в значительной мере зависит от уровня мотивации и
психофизической напряженности оператора.
Основные требования к
условиям труда человека—оператора в сжатой форме сводятся к трем следующим
условиям. 1. Тщательный профессиональный отбору и хорошая подготовка оператора
к данному виду деятельности. 2. Детальный учет человеческого фактора при
проектировании оборудования и технических систем. 3. Учет экологического
фактора, связанного с социальной средой обитания человека и с непосредственными
условиями труда.
3.25. ЦЕНТРАЛЬНАЯ РЕГУЛЯЦИЯ ДВИЖЕНИЙ
Все многообразие видимых форм
движений животных и человека основывается на физических законах перемещения тел
в инерциальном пространстве. При классификации движений необходимо
учитывать конкретные целевые функции, которые должна выполнять двигательная
система. В самой общей форме таких функций четыре: 1) поддержание определенной
позы; 2) ориентация на источник внешнего сигнала для его наилучшего восприятия,
а также поддержания равновесия тела; 3) перемещение тела в пространстве; 4)
манипулирование. Иерархия уровней мозгового управления движениями также
находится в зависимости от требований к структуре движения. Если подкорковый
уровень связан с набором врожденных или автоматизированных программ, то корковый
уровень организует спинномозговой моторный аппарат для выполнения произвольных
и тонких движений.
3.25.1. Управление ориентационными движениями и позой
Ориентационные движения. Их система связана с ориентацией
тела относительно координат внешнего пространства и с установкой сенсорных
аппаратов в положение наилучшего восприятия внешнего стимула. Примером первого
может служить функция поддержания равновесия, второго — движения фиксации
взора.
При ориентации в пространстве относительно зрительных координат
одной из основных задач является фиксация взора, которая выполняется в основном
глазодвигательной системой. Изображение неподвижного или движущегося предмета
фиксируется в наиболее чувствительном поле сетчатки. Точной фиксации глаз или
стабильному их перемещению по определенной траектории противодействуют
различные помехи: движения головы, самого зрительного объекта и изменения
глазодвигательной системы. Компенсация этих помех достигается с помощью
специальных рефлекторных механизмов.
Быстрые скачки глаз (микросаккады), которые служат
для переноса точки фиксации, делят на произвольные и непроизвольные. Произвольные
саккады отражают сознательно запрограммированную смену фиксаций взора в
зависимости от общего плана исследовательского поведения и не зависят от характеристик
объекта. Непроизвольные саккады участвуют в механизмах фиксации,
корректируя отклонения сетчатки от нужного положения, возникающие вследствие
дрейфа глаз или смещения головы. Они регулируются характеристиками зрительного
объекта и сигналами от глазных мышц. Непроизвольные саккады могут служить для
квантования зрительного мира, стирая остаточные следы предыдущего раздражения
сетчатки. Каждый видимый объект точно локализован в пространстве по отношению к
некоторому «образу тела». Простое движение (например, быстрое касание
какого—либо предмета) полагает, что при формировании команды учитываются
положение цели на сетчатке, положение глаз относительно координатной системы и
положение головы относительно тела.
В организации непроизвольных саккад большую роль выполняет мозжечок,
который участвует в программировании таких быстрых движений, которые не
могут регулироваться по кольцу внешней обратной связи, т. е. с помощью
проприоцептивной афферентации. Однако это не исключает возможности
регулирования по «внутренней» обратной связи.
Ведущую роль в управлении произвольными саккадами играет кора
больших полушарий. Для затылочного глазодвигательного поля определяющими
будут
|
|
|
Рис. 3.63 Соматотопическая
организация корковой зоны SI человека Изображения
под поперечным срезом мозга (на уровне постцентральной извилины) и их
обозначения демонстрируют пространственное представительство поверхности тела
в коре, установленное путем локальной электрической стимуляции мозга
бодрствующих больных. |
параметры зрительного сигнала, а для
фронтального и теменного глазодвигательных полей — сигналы от мозжечка,
связанные с выполнением «внутренней» двигательной программы. Предполагают, что
глазодвигательная система вовлекается в работу не после полного описания
изображения в зрительной сенсорной системе, а непосредственно по ходу обработки
зрительной информации.
Координированное движение глаз и головы регулируется
специальной системой рефлексов: Например, при слежении за целью повороты
головы целиком определяются движением глаз, а при поиске источника звука в
темноте одновременно запускается движение глаз и головы к цели. При появлении
на свету неожиданного зрительного объекта вначале запускается саккадическое
движение к цели и спустя некоторое время начинается поворот головы в том же
направлении. Центральным звеном координированного движения глаз и головы
являются верхние холмики пластинки крыши среднего мозга и система шейных,
глазных и вестибулярных рефлексов.
Управление позой. Позу тела можно описать как совокупность некоторых
значений суставных углов, обладающих определенной ориентацией в поле тяготения.
Механизмы позы распадаются на две категории: фиксацию определенных положений
тела и конечностей, т. е. фиксацию движения в суставах, и ориентацию сегментов
тела относительно внешних координат, т. е. поддержание равновесия. Исходная
поза тела накладывает некоторые ограничения на последующее движение.
К низшим механизмам управления позой
относятся спинальные рефлексы, шейные установочные рефлексы и вестибулоспинальные
рефлексы. К высшим механизмам управления позой относятся механизмы формирования
«схемы тела».
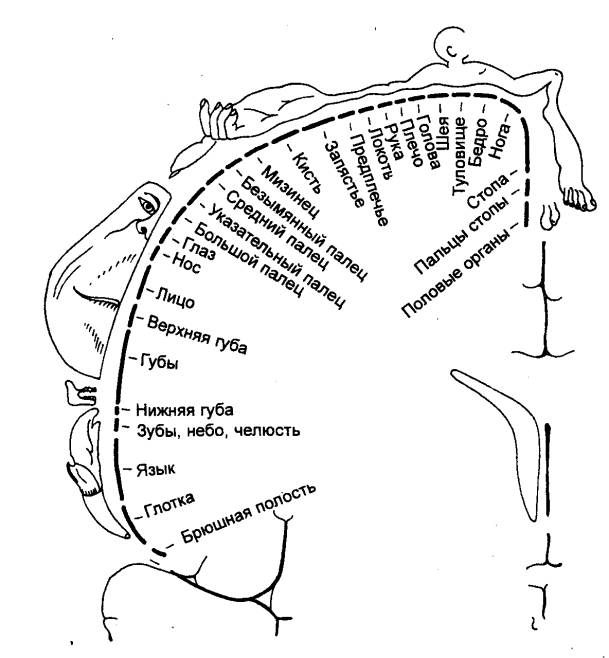
Термином схема тела обозначают систему обобщенной
чувствительности собственного тела в покое и при движении, пространственных
координат и взаимоотношений отдельных частей тела.
Детальное представительство отдельных частей тела, вплоть до
фаланг пальцев (соматотопия), характерно для соматосенсорной системы. Общая
«карта» тела для каждого полушария мозга представлена в виде «гомункулюса»
(рис. 3.63). Чувствительность всего тела, топографически распределенная по
поверхности коры, составляет ту основу, тот исходный материал, из которого
путем объединения формируются целостные функциональные блоки крупных отделов
тела. Эти интегративные процессы завершаются у взрослого организма и
представляют собой закодированное описание взаиморасположения частей тела,
которые используются при выполнении автоматизированных стереотипных движений.
|
|
|
Рис. 3.64 Высшие
механизмы регуляции позы. (Объяснение
см. в тексте). |
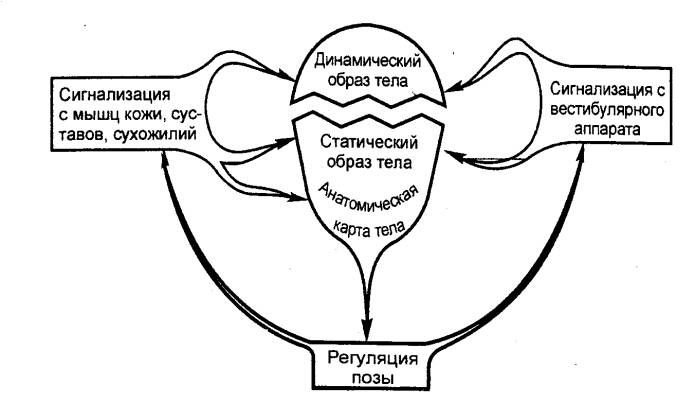
Базой для этих процессов служит анатомически закрепленная
«карта» тела, поэтому они составляют лишь основу статического образа тела.
Для его формирования необходимо соотнести эту информацию с положением тела по
отношению к силе земного притяжения и взаиморасположением функциональных блоков
тела в системе трех пространственных плоскостей. Вестибулярная система
воспринимает перемещения всего тела вперед—назад, вправо—влево, вверх—вниз, а
соответствующая информация поступает в теменные области коры мозга, где
происходит ее объединение с информацией от скелетно—мышечного аппарата и кожи
(рис. 3.64). Туда же поступает импульсация от внутренних органов, которая также
участвует в создании на бессознательном уровне статического образа тела.
Таким образом, статический образ тела представляет собой
систему внутримозговых связей, основанную на врожденных механизмах и
усовершенствованную и уточненную в индивидуальной жизни.
Осуществляя тот или иной род деятельности, человек меняет
взаиморасположение частей тела, приобретает в порядке научения новые
двигательные навыки, а значит, формирует новые трехмерные пространственные
модели тела, т. е. динамический образ тела.
В отличие от статического он имеет значение лишь для данного
конкретного момента времени, для определенной ситуации, при изменении которой
он сменяется новым. Динамический образ базируется на информации, поступающей от
кожи, мышц, суставов и вестибулярного аппарата.
В мозгу происходит постоянное взаимодействие того и другого
образов тела, осуществляется сличение динамического образа с его статическим
аналогом. В результате этого формируется субъективное ощущение позы,
отражающее не только положение тела в данный момент, но и возможные его
изменения в непосредственном будущем. Если согласование не достигнуто, то
вступают в действие активные механизмы перестройки позы. Итак, для того чтобы
субъективно оценить позу, необходимо сопоставить закодированный в памяти эталон
статического образа тела с его любой конкретной вариацией — динамическим
образом тела.
Не только схема тела является трехмерно пространственной, и
сам образ реальной пространственной ситуации воспринимается в том же
трехкоординатном пространственном измерении. Человек оценивает окружающее
пространство не абстрактно, а применительно к самому себе, т. е. включает свое
тело в качестве непременного компонента пространства.
Человек «вписывается» в окружающую ситуацию и воспринимает
ее, «привязывая», соотнося внешние объекты с положением собственного тела.
Внутренний и внешний образы описываются одним и тем же физиологическим языком,
в котором закодирован весьма ограниченный набор одних и тех же параметров.
Для подробного описания нужно представить себе наличие,
например, какой—то вертикальной плоскости, которая делит тело на две
симметричные половины. Тогда все участки тела и предметы окружающей среды,
расположенные
|
|
|
Рис. 3.65 Формирование
представлений о внешнем пространстве и «схеме тела» Объяснение см. в
тексте. |
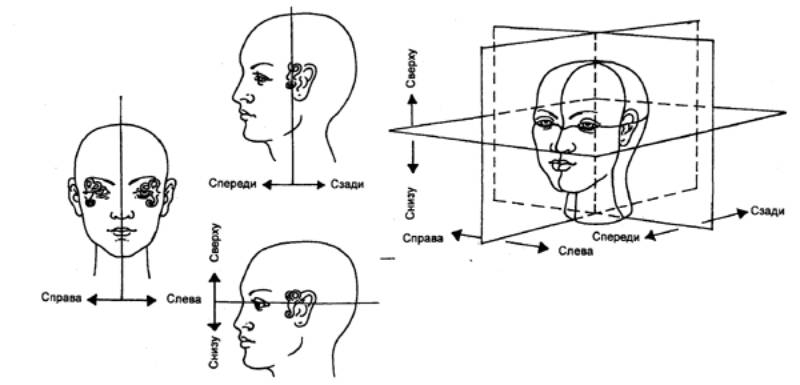
по одну сторону от этой плоскости, будут
субъективно восприниматься как «слева», а по другую — как «справа» (рис. 3.65).
Только информация о местоположении органов тела будет передаваться по
соматическим трактам, а информация о местоположении предмета по отношению к
организму — по зрительным трактам.
Эти два потока информации
встречаются и объединяются для последующего использования в произвольной
двигательной деятельности. Конвергенция и интеграция этих афферентных влияний
происходят в пределах таламопариетальной ассоциативной системы мозга.
Рассмотренное справедливо не только для описанной выше сагиттальной плоскости,
но и для фронтальной, которая проходит через центры вестибулярных аппаратов, и
горизонтальной, которая проходит через центры глазных яблок и вестибулярных
аппаратов.
Таким образом, и наклон головы, и
содружественный поворот глазных яблок в ту или иную сторону от этих плоскостей
способствуют восприятию внешнего и внутреннего пространства в одних и тех же
параметрах. В итоге создается целостный образ организма в среде, который служит
определенным сенсорным фоном для выполнения любого произвольного движения.
Под термином локомоция (от
лат. locus — место и motio — движение) понимают перемещение тела из одного места
пространства в другое, для чего необходимы некоторые силы, изменяющие исходное
состояние организма. Развиваемые при этом усилия должны преодолеть силу
тяжести, сопротивление окружающей среды и силы инерции тела. На локомоцию
влияют характер и рельеф местности. Во время локомоции необходимо непрерывно
поддерживать равновесие.
Наиболее распространенным примером
локомоции являются ходьба и бег, которые отличаются стереотипным движением
конечностей, причем для каждой из форм локомоции характерны две фазы шага:
опоры и переноса. Разные животные обнаруживают различные, свойственные только
им способы ходьбы, которые в то же время обладают общими чертами. Локомоция
характеризуется
|
|
|
Рис. 3.66 Модель спинального генератора
локомоции Приведены афферентные и эфферентные связи на спинальный генератор;
Ф — флексоры (сгибатели), Э —
экстензоры (разгибатели), α и γ — альфа— и гамма—мотонейроны. |
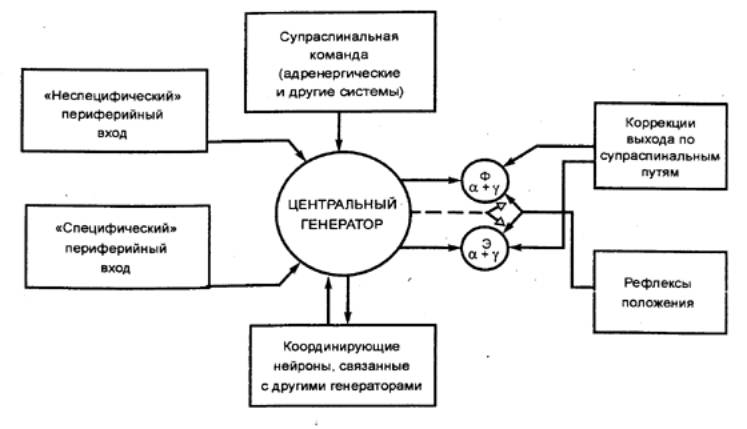
походкой — определенным способом передвижения
по поверхности, который оценивается по распределению во времени циклических
движений конечностей, длительностью опорной фазы и последовательностью
перемещения опорных конечностей.
Описана нейронная цепь, находящаяся в спинном мозгу, которая
называется спинальным генератором шагания. Она ответственна за
чередование периодов возбуждения и торможения различных мотонейронов и может
работать/в автоматическом режиме (рис. 3.66). Элементарной единицей такого
центрального генератора является генератор для одной конечности. Допускают
даже, что мышцы, управляющие одним суставом, могут контролироваться каким—то
собственным унитарным генератором. При формировании локомоторных движений эти
генераторы разных движений тесно связаны и оказывают друг на друга возбуждающее
влияние.
Структуры спинного мозга находятся под непрерывным супраспинальным
контролем. Установлено, что раздражение определенных участков ствола мозга
способно вызывать координированную локомоцию, обусловленную запуском
ритмической активности спинального генератора.
Супраспинальный контроль локомоции выполняет несколько
основных функций: 1) быстро запускает локомоцию, поддерживает постоянную
скорость или изменяет ее, если требуется, а также прекращает ее в нужный
момент; 2) точно адаптирует движение (и даже отдельный шаг) к условиям среды;
3) обеспечивает достаточно гибкую локомоторную позу, чтобы соответствовать
различным условиям передвижения, таким, например, как ползанье, плаванье, бег
по снегу, перенос груза и т. д. Афферентная сигнализация, идущая в спинной мозг
от мышечных веретен, сухожильных рецепторов Гольджи, кожи не является необходимым
условием возникновения локомоции, тем не менее она участвует в более точной
адаптации локомоторных движений к условиям среды.
Значительную роль выполняет мозжечок, который
обеспечивает коррекцию и точность постановки конечностей на основе сравнения информации
о работе спинального генератора и реальных параметров движений, причем
мозжечковые влияния направлены на регуляцию моторного выхода, а не на
активность самого спинального генератора. Мозжечок получает прямую информацию
по восходящим спинно—мозжечковым путям. Предполагают, что мозжечок
программирует каждый следующий шаг на основе информации о предыдущем.
|
|
|
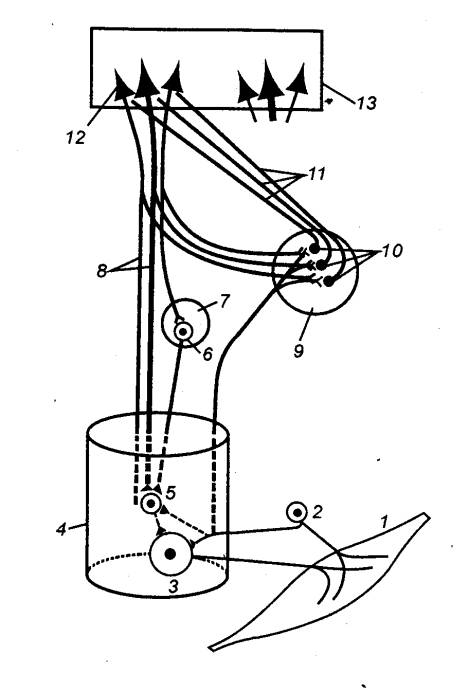
Рис.
3.67 Соотношение
прямых и внутренних обратных связей в ядрах дорсальных столбов 1 — мышца, 2 — чувствительный нейрон спинального ганглия, 3 — мотонейрон
спинного мозга, 4 — спинной
мозг, 5 — интернейрон спинного мозга, 6
— нейрон красноядерно—спииномозгового пути, 7 — красное ядро, 8 —
пирамидный путь, 9 — ядра
дорсальных столбов, 10 —
нейроны продолговатого мозга, дающие начало медиальной петле (11), которая переключается в
таламических ядрах и несет афферентацию в кору мозга. На этих нейронах (10) осуществляется
пресинаптическое торможение в момент эфферентного залпа по коллатералям
волокон пирамидного пути; последние берут начало от крупных пирамидных клеток
(12} сенсо—моторной области
коры (13). |
Другой важнейший уровень мозга, куда направляется информация
о характере выполнения движения, это полушария мозга с таламическими ядрами,
стриопаллидарной системой и соответствующими зонами коры головного мозга. В
быстрых движениях баллистического типа классическая соматическая обратная связь
не принимает участия потому, что само движение завершается раньше, чем успеет
поступить информация от двигательных аппаратов к соответствующим мозговым
центрам. Медленные движения постоянно корректируются благодаря показаниям с
соответствующих сенсорных датчиков, расположенных в скелетно—мышечном аппарате
и передающих информацию в разные отделы мозга вплоть до коры. Эти сенсорные
коррекции способны изменить характер движения по ходу его осуществления.
Движения, базирующиеся на врожденных координациях, в меньшей степени требуют
обратной связи от моторного аппарата, а движения, в основе которых лежит
формирование новых координационных отношений, всецело зависят от соматической
афферентации с двигательного аппарата. Кроме того, степень участия обратных
связей находится в зависимости и от уровня автоматизации двигательного акта.
Простые движения, осуществляющиеся без
проприоцептивной обратной связи (саккады и быстрые движения конечностей),
выполняются по жесткой «запаянной» программе. Любое же сложное движение требует
предварительного программирования. Для сложных движений существенным является
сличение обратной афферентации с тем сенсорным образом движения, который
формируется в составе программы. Результат сличения и является основным
фактором перестройки программы. Эти влияния передаются к аппаратам
программирования по каналам внутренней обратной связи, которая включает в себя
все процессы перестроек двигательной программы в зависимости от
внутрицентральных влияний.
Такими каналами внутренней обратной связи являются медиальные
петли (лемниски), по которым передается импульсация, интегрированная в
ядрах дорсальных столбов. Местом ее поступления в кору больших полушарий
оказываются не только соматосенсорные зоны, но и сенсомоторные области, куда,
кроме того, конвергирует гетеромодальная информация и где осуществляются
интегративные процессы.
Если перемещение конечности — это компонент
общекоординированного акта, то внутренняя (проприоцептивная) и внешняя (кожная
и телецептивная) афферентации выступают как содружественные звенья системы
сенсорного обеспечения движений. Соматическая афферентация от локомоторного
аппарата необходима для работы генератора инициации движений и их
координированного выполнения. Это в первую очередь касается медленных
компонентов двигательных актов. Существует обратная связь, с помощью которой
кора информируется не об отдельных параметрах движений, а о степени
соответствия предварительно созданной двигательной программы тому наличному
движению, которое достигается в каждый момент времени. Одним из важнейших
каналов такой внутренней обратной связи и выступают медиальные петли.
Известно, что пирамидный путь отдает
большое число коллатералей к ядрам дорсальных столбов в продолговатом мозгу.
Поэтому «пирамидальный залп» из коры достигает не только аппаратов спинного
мозга, но и ядер дорсальных столбов, нейроны которых в течение реализации
данного цикла движения находятся в тормозном состоянии благодаря развитию в них
пресинаптического торможения (рис. 3.67).
Следовательно, импульсация по
волокнам дорсальных столбов (тонкий и клиновидный пучки) оказывается
заблокированной, т. е. подпороговой. Превращение в надпороговую при отклонении
параметров реального движения от требуемых согласно команде вызывает поток
импульсов по системе медиальных петель в кору. Здесь будет проводиться
коррекция следующего цикла командного воздействия. Поэтому сенсомоторная кора →
коллатерали пирамидного пути → ядра дорсальных столбов →медиальные
петли — это важнейшее кольцо, по которому происходит непрерывная коррекция
двигательных команд сообразно параметрам предшествующего двигательного цикла.
Срочная соматическая информация
обеспечивает фильтрацию телецептивных и экстралемнисковых влияний на
корково—подкорковом уровне путем управления системой нисходящих связей. Этим
достигается организация точных выходных команд, придающих движению
координированный характер. Ядра дорсальных столбов являются одной из тех
мозговых структур, где осуществляется сличение результата двигательного цикла с
копией программы и последствия рассогласования поступают в сенсомоторную кору
для очередной коррекции двигательной программы. Следовательно, если внешняя
обратная связь от соматических рецепторов ограничивается спиннобульбарным
уровнем, то внутренняя обратная связь осуществляется по системе медиальных
петель.
3.25.3. Организация манипуляторных
движений
Манипуляторные движения — это наиболее яркий пример
произвольных движений, возникновение которых обусловливают внутренние
мотивационные влияния. Эти движения являются локальными, что ставит перед
двигательной системой следующие задачи: 1) выбор ведущего мышечного звена; 2)
компенсацию внешней нагрузки; 3) настройку позы; 4) соотнесение координат цели
и положения собственного тела.
Отличительной чертой манипуляторных движений является их
зависимость от центральной программы и телецептивных сенсорных входов. Поэтому ведущая
роль отводится следующим мозговым структурам: лобной коре, базальным ядрам
и мозжечку. Активность других супраспинальных цепей двигательного контроля в
большей степени зависит от периферической информации, поступающей вслед за
реализацией команд.
Ведущая роль в программировании быстрых движений принадлежит
мозжечковой системе, а в программировании медленных — базальным
ядрам. Выходы этих структур на спинальный уровень опосредованы в основном
через сенсомоторную кору, которая дополнительно к этому обладает своими собственными
функциями.
3.25.4. Корковая сенсомоторная
интеграция
Структурной единицей неокортекса
является модуль — микросистема тесно контактирующих между собой
пирамидных нейронов для их синхронного вовлечения в формирование эфферентного импульсного
залпа (рис. 3.68). Такое объединение рассматривают в качестве жесткого
структурного элемента организации коры, формирующегося в пренатальном
онтогенезе. Почти всем модулям свойственна широкая конвергенция гетеросенсорных
влияний. В моторную кору конвергируют те компоненты сенсорной информации,
которые являются жизненно важными для организма. Это информация о биологической
значимости того или иного сигнала либо его значимости, приобретенной в порядке
выработки условного рефлекса.
Топографическое распределение
соматических проекций участков тела на кору больших полушарий только частично
определяется анатомическими закономерностями. Не менее важную роль играют
функциональные механизмы, определяющим среди которых является внутрикорковое
торможение.
Структурный корковый
модуль выступает в качестве элементарной сенсо—моторной единицы. Различия между
отдельными модулями определяются характером соматической сенсорной проекции и
адресатом эфферентного выхода из того же модуля. Существует распределенная
система корковых модулей, выходы которых конвергируют на разных уровнях
ствола и спинного мозга, обеспечивая управление деятельностью определенной
мышечной группы. Распределенная система является частью более обширных
общемозговых систем, сформированных в раннем онтогенезе. Такие функциональные
структуры управляют более обширной частью скелетно—мышечного аппарата.
|
|
|
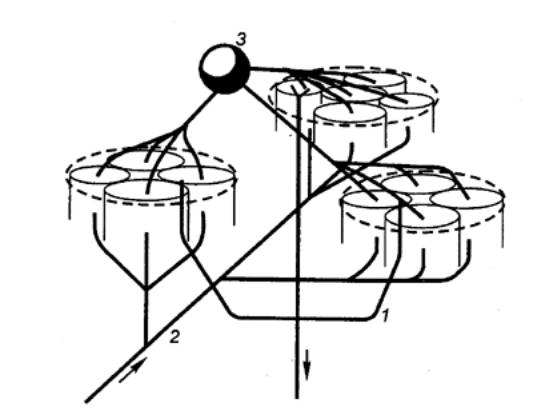
Рис. 3.68 Функциональные объединения нейронных
модулей Одиночный
структурный модуль имеет вид цилиндра; 4—6 модулей могут сгруппироваться в функциональные
концентрические объединения, между которыми . возможны как облегчающие, так и
взаимотормозные отношения. Обширные функциональные объединения из нескольких
концентрических модулей строятся на основе ветвлений аксонной коллатерали
пирамидных нейронов (1),
ветвлений таламического афферента (2)
и ветвлений аксона корзинчатой клетки (3); стрелками показано
направление восходящей и нисходящей импульсации. |
Вовлечение корковых систем в целостную деятельность мозга
определяется текущей мотивацией и конкретной двигательной программой,
направленной на удовлетворение этой мотивации. Двигательная программа, чтобы
носить приспособительный характер, должна учитывать все сигнально значимые
компоненты внешней среды, относительно которых совершается целенаправленное
движение, т. е. строиться на принципе мультисенсорной конвергенции.
Телецептивные входы на модулях сенсомоторной коры носят сравнительно диффузный
характер и адресуются ко многим пространственно разнесенным одноименным
модулям. Констелляции модулей основаны на синергичных и антагонистичных
межмодульных соотношениях, что обеспечивает динамичность их участия в различных
актах сенсомоторной интеграции.
3.25.5. Программирование движений
Основным элементом любого
целенаправленного движения является формирование предварительных программ, т.
е. процессов, позволяющих предвидеть динамику изменений факторов внешней среды
и придать будущему движению адаптивный характер. Результат сличения
афферентации о движении с двигательной программой, передающийся по системе
внутренней обратной связи, является основным фактором перестройки программы.
Последнее зависит от биологической мотивированности движения, его временных
параметров, моторной дифференцированности, сложности координационного состава и
уровня его автоматизированное.
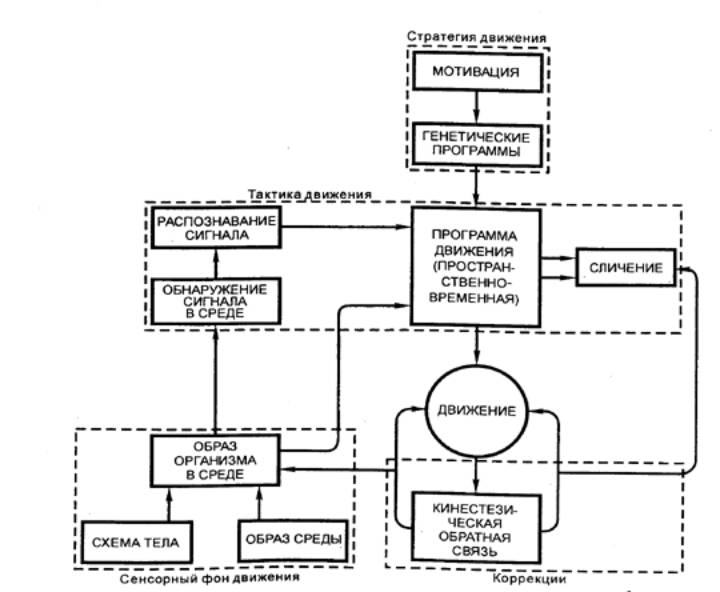
Мотивации определяют общую стратегию
движения. Каждый конкретный двигательный акт рассматривается как шаг к
удовлетворению мотивации, т. е. решает некоторую промежуточную задачу или цель
(рис. 3.69). Биологические мотивации приводят к запуску либо «запаянных»,
жестких моторных программ, либо формируют новые сложные программы. Если
мотивация определяет цель движения, его программу, то она же обусловливает ее
зависимость от сенсорных факторов. В качестве обратной связи здесь выступает
удовлетворение мотивации. Тактика движения, или двигательная команда,
определяет, как будет осуществляться требуемое запрограммированное движение, т.
е. каково распределение во времени тех эфферентных залпов к моторным ядрам
спинного мозга, которые определяют активацию различных мышечных групп.
В отличие от программы команды
движения должны быть адаптивными к функциональному состоянию самого
скелетно—двигательного аппарата как непосредственного исполнителя этих команд.
Предполагают, что срочная идентификация состояния мышечной системы
осуществляется путем оценки реакции
|
|
|
Рис.
3.66 Модель спинального генератора локомоции Приведены афферентные и
эфферентные связи на спинальный генератор; Ф — флексоры (сгибатели), Э — экстензоры (разгибатели), α
и γ — альфа— и гамма—мотонейроны. |
системы на небольшое точно
дозированное возмущение (тест—импульс), которое подается непосредственно
перед или с момента запуска команд.
Тест—импульс подается только на те мышцы, которые должны будут
участвовать в следующем движении. Реакция мышц будет не настолько значительна,
чтобы существенно изменить само движение, но достаточна для возбуждения
мышечных веретен при изменении длины интрафузальных волокон на 5—10 мкм.
Информация о реакции на тест—импульс конвергирует на некотором промежуточном
уровне с основным управляющим воздействием (командой), которое корректируется в
соответствии с состоянием самого двигательного аппарата. Такое тестовое
воздействие передается по быстропроводящим волокнам пирамидного пути и
адресуется к сегментам спинного мозга.
Непосредственное управление движением обусловливается
активностью сенсомоторной коры мозжечка и полосатого тела. Последнее участвует
в трансформации «намерения действовать» в соответствующие «командные сигналы»
для инициации и контроля движений.
Ассоциативные системы мозга и движение. Специфичность участия таламопариетальной
ассоциативной системы в организации движений определяется двумя моментами:
она участвует в формировании интегральной схемы тела, все части которого
соотнесены не только друг с другом, но и с вестибулярными и зрительными
сигналами, и регуляции внимания к текущим сигналам окружающей среды с учетом
ориентации всего тела относительно этих сигналов. Эта система «привязана» к
настоящему моменту времени и к анализу пространственных взаимоотношений
разномодальных признаков.
По таламофронтальной ассоциативной системе передается информация о
мотивационном состоянии и сопутствующих висцеральных изменениях. Фронтальная
ассоциативная кора опосредует мотивационные влияния на организацию поведения в
целом благодаря связям с теменно—височной корой, гипоталамусом, областями
среднего и промежуточного мозга. Поэтому лобная кора участвует в регуляции
процессов во внутренней среде организма, сенсорных и моторных механизмах мозга.
3.25.6. Функциональная структура произвольного движения
Среди общих принципов управления
движениями выделяют проблему организации единовременной команды к
исполнительным аппаратам. Если ее рассматривать безотносительно к стратегии и
тактике движения, то основной целью системы, формирующей эту команду, будет
координация всех компонентов команды.
Для того чтобы выполнить эту задачу, система должна
осуществлять следующие процессы: идентификацию состояния системы, определение
целевой функции ведущего кинематического звена, подстройку соотношения
активности мышц—антагонистов. Последнее производится автоматически благодаря
системе спинальных рефлексов. Идентификация же состояния двигательной системы
обеспечивается двумя путями: оценкой реакции системы на тест—импульс,
генерируемый корой, или на внешнее воздействие.
ЦНС располагает значительным числом генетически закрепленных
программ типа локомоторной программы, базирующейся на активности спинального
генератора шагания. Такие простые программы объединяются в более сложные
системы типа поддержания вертикальной позы, что происходит в процессе научения
при участии структур переднего мозга.
Одной из задач управления движением является выбор ведущего
кинематического звена. Этот выбор предопределен исходной позой, перестройка
которой всегда опережает локальное движение. Информация об исходной позе будет
перераспределять активность сенсомоторной коры. Подкорковые управляющие
структуры (полосатое тело и мозжечок) формируют новое движение почти исключительно
путем модификации состояния командных интернейронов во врожденных подсистемах
управления. Диапазон этой модификации очень широк и у многих видов животных
объемлет почти весь двигательный репертуар.
Для формирования новых координационных отношений, особенно в
случае манипуляторных движений, необходимо подавить активность командных
нейронов врожденных программ и сформировать параллельный управляющий канал.
Таковым является корково—спинномозговой путь, степень развития которого
независимо от положения животного в филогенетическом ряду определяет роль
манипуляторных движений в общем двигательном поведении. Морфологической основой
организации тонких манипуляторных движений является сенсомоторная кора с ее
структурными модулями и объединениями модулей разного порядка сложности. Все
элементы вышепоименованных систем одномоментно связаны между собой, поэтому
изменение любого звена влечет за собой сдвиг состояния всей двигательной
системы в целом.
Ориентационные движения и следящие движения глаз, головы,
конечностей представляют собой задачу более высокого порядка, чем координация
движений сегментов тела. Эти движения зависят от функционирования таламопариетальной
ассоциативной системы мозга.
Самой сложной, филогенетически самой молодой является
способность формировать последовательность движений и предвидеть ее реализацию.
Решение этой задачи связано с фронтальной ассоциативной системой,
которая запоминает и хранит в памяти такие последовательности движений.
Высшим отражением этого кодирования у человека является
вербализация основных понятий движения.
Одно из основных проявлений поражений лобных долей человека
состоит в нарушении грамматической структуры речи и невозможности использования
вербальных команд построения последовательности движений.
Всеобщей закономерностью работы системы управления
движениями является использование обратной связи. Сюда входит не только
проприоцептивная обратная связь от начавшегося движения, но и активация систем
поощрения или наказания. Кроме того, включается и внутренняя обратная связь, т.
е. информация об активности нижележащих уровней двигательной системы или
эфферентная копия самой двигательной команды. Этот вид обратной связи необходим
для выработки новых двигательных координации. Для движений разной сложности и
скорости обратная связь может замыкаться на разных уровнях. Поэтому оба типа
управления — программирование и слежение — могут сосуществовать в системе
управления одним и тем же движением.
3.26. ЭМОЦИИ КАК КОМПОНЕНТ ЦЕЛОСТНЫХ ПОВЕДЕНЧЕСКИХ РЕАКЦИЙ
Различные формы поведения человека и
животных направлены на удовлетворение многообразных потребностей, лежащих в
основе поддержания индивидуального и видового гомеостаза. Объективное описание
целостных поведенческих актов и сопутствующих им вегетативных реакций,
исследование центральной организации взаимоотношений организма с внешним миром,
возможность прогнозировать ответы на изменения во внутренней среде и на сигналы
из внешнего мира — необходимая основа физиологического осмысления механизмов
высшей нервной деятельности. Нельзя упускать из виду и процессы, отражающие
субъективные состояния, но поддающиеся объективному нейрофизиологическому
анализу.
К таким состояниям следует отнести
прежде всего мотивации и эмоции. Хотя представления о них пришли
в физиологию из психологии, современные исследователи не могут обойтись без их
использования при анализе механизмов высшей нервной деятельности. О мотивационно—эмоциональной
сфере человека судят по его поступкам, мимике и другим выразительным
движениям, вегетативным реакциям, электрическим явлениям в головном мозгу, а
также по словесному отчету о субъективном состоянии и собственному отношению к
нему.
3.26.1. Биологическая роль эмоций
Судить о мотивационно—эмоциональных
процессах у животных можно лишь по объективным показателям. Однако накопленные
сведения об эволюции головного мозга, сопоставление участия отдельных его
систем в организации поведения человека и животных дают возможность
постулировать существование у последних субъективных состояний как сформировавшихся
в процессе эволюции полезных признаков, необходимых для выживания особи и
сохранения вида.
В целостной поведенческой реакции
мотивация и эмоция проявляются в неразрывном единстве, но, как показывает
физиологический анализ, в экспериментальных условиях их можно разделить, так
как они отражают активность хотя и тесно взаимодействующих, но
специализированных отделов ЦНС. Особенно четко демонстрируется относительная
самостоятельность мотиваций и эмоций при разрушении этих отделов или
раздражении их электрическим током или фармакологическими веществами.
Мотивации и эмоции тесно связаны с
возникновением и удовлетворением потребностей организма — необходимым условием
его жизнедеятельности.
Мотивации (побуждения, влечения, драйвы)
определяются генетической программой и индивидуальным опытом, активизируются
потребностями и проявляются в избирательном возбуждении специализированных
мозговых структур, что приводит при благоприятной внешней ситуации к реализации
целенаправленного поведения.
Различают низшие (первичные,
простые, биологические) и высшие (вторичные, сложные, социальные)
мотивации. Примерами биологических мотиваций могут служить голод, жажда, страх,
агрессия, половое влечение, забота о потомстве. Не следует отождествлять
мотивации и потребности. Нередко последние не трансформируются в мотивационное
возбуждение и, следовательно, не могут быть удовлетворены посредством
реализации целенаправленного поведенческого акта.
Крысы, например, предпочитают жирную
безбелковую пищу безжировой белковой. Люди нередко также потребляют менее
полезную, а иногда и вредную пищу с высоким содержанием низкомолекулярных
Сахаров лишь потому, что она вкуснее, чем оптимальная по составу. У человека и
многих видов животных нет врожденного предпочтения питательных веществ, богатых
витаминами. Поэтому на протяжении многих веков отдельные группы населения
страдали болезнями, вызываемыми авитаминозами (цинга, пеллагра, бери—бери и
др.), причем развитие этих болезней не всегда определялось отсутствием
источника витаминов.
Вместе с тем для удовлетворения
многих биологических потребностей не обязательно существование врожденных
механизмов их трансформации в мотивационное возбуждение. Специализированные
поведенческие акты могут формироваться на основе неспецифического поиска,
инициируемого различными нарушениями гомеостаза. В частности, у крыс могут быть
выработаны витаминные, минеральные и другие аппетиты, а также предпочтение или
отвергание определенных пищевых продуктов. Подобные перестройки носят
условнорефлекторный характер и свидетельствуют о большой пластичности
биологических мотиваций. Естественно, в еще большей мере связаны с
индивидуальным опытом высшие, социальные мотивации.
Перестройки поведения
требуют оценки произведенных действий. Она основана на формировании
положительного или отрицательного эмоционального состояния. Первое отражает
благотворный эффект произведенного действия и создает предпосылку для его
продления или повторения, второе сигнализирует о вредном его влиянии на
организм и нецелесообразности повторения. Эмоции тесно связаны с подкреплением
— важнейшим компонентом обучения.
Таким образом, в
поведении отражаются отдельные, но тесно переплетающиеся явления. На основании
потребностей формируется мотивация, ее реализация в целенаправленных актах
поведения сопровождается эмоциями положительного или отрицательного знака.
Среди этих явлений
наиболее сложны для изучения именно эмоции. Анализ потребностей можно проводить
на основании консуматорных реакций. Так, потребность в воде или пище
определяется по их потреблению. Уровень биологической мотивации измеряется
многими способами, в частности скоростью целенаправленной побежки, преодолением
препятствий разной сложности, выбором действий в условиях конкурентных
отношений между удовлетворением разных биологических потребностей (пищевой и
половой, пищевой и водной и т. п.). В естественных условиях мотивации
проявляются как эмоционально окрашенные состояния, однако параллелизма между
уровнями мотивационного и эмоционального возбуждения не имеется. Поэтому нельзя
оценивать последнее по актам, нацеленным на удовлетворение биологических
потребностей.
Важной особенностью
эмоций является комплекс телесных реакций, сопутствующих отрицательным
или положительным переживаниям. У человека субъективные состояния удовольствия
или неудовольствия находят внешнее выражение в изменениях позы, походки,
выражения лица, жестов, голосовых реакций, частоты пульса, дыхания,
потоотделения, многих гуморальных показателей. По этим изменениям можно
косвенно судить о степени волнения испытываемого человеком. Однако условность
такого суждения очевидна. Мимические и прочие движения, характерные для
аффективных состояний, человек в состоянии затормозить или воспроизвести при
отсутствии соответствующих переживаний.
Во многих случаях внешние выражения чувств определяются социальными
стереотипами поведения. Тренировка позволяет подавлять или активизировать
некоторые вегетативные реакции, отражающие эмоциональные переживания. Наиболее
полно такая способность проявляется в игре актеров.
Таким образом, даже у человека на основании сопоставления
словесного отчета о субъективном состоянии и комплекса моторно—вегетативных
реакций можно лишь догадываться об испытываемых им переживаниях.
Еще более сложно оценить эмоциональное состояние у животных.
О нем судят по различным реакциям на действие внешних раздражителей или
стимуляцию некоторых отделов головного мозга. Вместе с тем общепринятое
положение о единых для человека
и животных закономерностях эволюции мозга позволяет считать, что первичные
эмоции сформировались задолго до появления человека как чувственное выражение
инстинктивной деятельности.
Именно с таких позиций
рассмотрел их Ч. Дарвин в работе «О выражении эмоций у человека и животных»
(1872). Им был приведен большой материал по сравнительному анализу мимики,
пантомимики, голосовых и вегетативных реакций при аффективных состояниях у
представителей разных отрядов млекопитающих и человека. Выразительные движения
рассматривались Ч. Дарвином как сформировавшиеся в процессе естественного
отбора приспособительные реакции, существенные для общения с особями своего или
других биологических видов. Он предположил, что мимико—жестикуляционные реакции
являются компонентами агрессивного, оборонительного или других форм поведения.
Например, отражающие аффект гнева оскаливание зубов и угрожающие жесты пугают
врага. Некоторые выражения аффективного состояния являются рудиментами. Так, в
состоянии гнева человек сжимает кулаки и скалит зубы, показывая готовность к
борьбе.
Ч. Дарвин оценивал
эмоциональные состояния в связи с побуждением к действию, не пытаясь
анализировать их субъективную сторону. Подобный подход был полезен для
подтверждения сходства в поведении животных и человека и обоснования
происхождения человека от обезьяны. Однако при этом игнорировались качественные
различия между психической сферой человека и животных.
3.26.2. Эмоции и психическая
деятельность
Периоду естественнонаучного подхода
к анализу эмоций предшествовало длительное осмысление их места в психической деятельности
человека. Еще в XVII в. Р. Декарт выдвинул положение, согласно которому под
влиянием эмоции организм осуществляет полезные для него действия и уклоняется
от совершения вредных. В работах философов XVIII — начала XIX вв. большое место
занимал вопрос о взаимоотношении аффективной сферы и телесных реакций.
Высказывались предположения о зависимости эмоциональных состояний от
органических изменений и о противоположных причинно—следственных отношениях.
Важно отметить, что некоторые философы пришли к заключению о связи эмоций с
конфликтом между представлениями (т. е. в соответствии с современной нам
терминологией — рассогласованием).
В конце XIX в. американский психолог
и философ У. Джеме предпринял попытку создания физиологической теории эмоций,
вторичных, по его мнению, по отношению к вызванным различными стимулами
телесным изменениям. Согласно «периферической теории» У. Джемса, эмоция
является осознанием изменений в работе внутренних органов и мышц. Близкую
позицию занял датчанин К. Ланге, полагавший, что эмоциональное состояние
отражает органические изменения, связанные преимущественно с кровообращением.
Вопреки представлению об эмоции (психическом состоянии) как первопричине
изменения деятельности различных физиологических систем было предложено считать
эти изменения источником эмоций. С этой точки зрения человек печален, потому
что он плачет, испытывает ярость или страх, потому что бьет другого или дрожит,
и т. д. Попытка превратить эмоции в объект, доступный объективному анализу, не
была подкреплена физиологическими опытами.
Пытаясь проверить теорию
Джемса—Ланге в эксперименте, Ч. Шеррингтон перерезал у собак блуждающие нервы и
спинной мозг, устраняя сигнализацию от внутренних органов. При этом
эмоциональные реакции на действие внешних раздражителей не исчезали. В работах
других физиологов было показано, что даже после полной изоляции головы (в
условиях перекрестного кровообращения) у кошек или собак сохраняется гамма
мимических проявлений эмоционального состояния.
Несмотря на то что теория Джемса—Ланге
низводила эмоцию до остаточных эффектов органических изменений, она явилась
толчком для многих физиологов к объективному исследованию этих изменений.
Американский физиолог У. Кеннон, первоначально сочувственно относившийся к
«периферической теории», в результате экспериментов на кошках пришел к
заключению о центральном происхождении эмоций. Он показал, что после
удаления симпатических ганглиев эмоциональные реакции сохраняются. Например,
симпатэктомированная кошка, увидев собаку, издавала угрожающие звуки,
оскаливала зубы, поднимала лапу с выпущенными когтями. Один из выводов этих
опытов был связан с признанием неспецифичности органических изменений, в
частности однонаправленности вегетативных реакций при таких различных
эмоциональных состояниях, как ярость и страх.
Как будет видно из дальнейшего
изложения, развитие физиологии показало правильность центральной теории эмоций.
Были описаны эмоциогенные структуры мозга, связанные с положительными и
отрицательными переживаниями. Вместе с тем появились новые факты,
свидетельствующие о влиянии периферических стимулов на эмоциональную сферу.
Установлена корреляция между положительными переживаниями и сокращениями мышц
лица у улыбающегося человека. Показано, что мимика изменяет кровоснабжение
мозга, т. е. может оказывать влияние на структуры, регулирующие эмоциональное
состояние. Проприоцептивная обратная связь этих рецепторов лицевой мускулатуры
также вносит вклад в его формирование. Модулирующее влияние оказывает на него и
интероцептивная сигнализация, преимущественно по афферентным системам
блуждающих нервов.
Таким образом, эмоциональное
состояние, являясь отражением активации ряда структур ЦНС, в определенной
степени зависит от висцеральных раздражителей. ,
В ряде случаев возбуждение
эмоциогенных центров вовлекает в реакцию структуры мозга, связанные с
регуляцией висцеральных процессов. Состояние последних при этом изменяется,
что, в свою очередь, оказывает модулирующее влияние на эмоциональное состояние.
Большую роль в формировании взаимодействия между центрами эмоций и интеро— и
проприоцептивными системами играет условнорефлекторная деятельность.
Наличие в структуре условного рефлекса прямых и обратных связей позволяет
понять сопряженность моторных и сенсорных функций и участие их в формировании
эмоциональных реакций.
Зависимость переживаний от
деятельности висцеральных органов наиболее рельефно проявляется при различных
патологических процессах. Нарушение кровоснабжения миокарда при спазме
коронарных сосудов сердца часто сопровождается страхом смерти. Это переживание
является рефлекторным и может быть устранено лекарственными препаратами,
расширяющими коронарные сосуды. Изменения в эмоциональной сфере часто
сопутствуют сдвигам в эндокринной системе. Больные гипотиреозом редко смеются или
плачут, у них возникает подавленность, преобладание плохого настроения.
Различные нарушения эмоциональной сферы характерны и для больных гипертиреозом.
Гуморальные раздражители могут
влиять на эмоциогенные системы мозга непосредственно и опосредованно, изменяя
состояние мотивациогенных центров. При этом положительная или отрицательная
направленность переживаний в значительной мере определяется
условнорефлекторными факторами. Например, у человека чувство голода, которое
может быть удовлетворено, окрашено положительными переживаниями, тогда как в
условиях отсутствия пищи метаболические и нервные стимулы, сигнализирующие о
потребности в ней, приводят к формированию отрицательного эмоционального
состояния.
В зависимости от
текущего возбуждения мотивационно—эмоциогенных центров мозга одни и те же
сигналы могут вызывать различные реакции. Так, периодические сокращения пустого
желудка могут инициировать или усиливать чувство голода при соответствующей
возбудимости гипоталамических отделов пищевого центра. Если же она снижена, эти
сокращения не приводят к формированию голодного побуждения. Например, в
зависимости от инструкций, полученных испытуемыми, введение им адреналина
вызывает либо гнев, либо эйфорию (повышенное благодушное настроение,
сочетающееся с беспечностью), либо не вызывает никаких эмоций.
Таким образом, хотя
периферические сигналы и участвуют в формировании эмоционального состояния,
теорию Джемса—Ланге, из которой следует, что эмоции вторичны по отношению к
органическим изменениям, нельзя считать правильной.
3.26.3. Вегетативные реакции, сопутствующие эмоциональному
состоянию
Вегетативные реакции являются важными компонентами
целостных актов, имеющих приспособительный характер, поэтому они традиционно
исследуются физиологами и психологами. Ниже перечислены некоторые из этих
реакций.
Одним из самых популярных
показателей деятельности автономной нервной системы при аффективных состояниях
у человека является кожно—гальваническая реакция (феномен Тарханова).
Она зависит от состояния потовых желез, проявляется в снижении электрического
сопротивления между двумя участками поверхности кожи. Эту реакцию используют
для определения ложных показаний, раскрытия симуляции, анализа состояния
эмоциональной сферы испытуемого.
У человека и животных аффективные
реакции отражаются также в изменениях артериального давления,
электрокардиограммы, различных показателей дыхания, температуры кожи, зрачковой
реакции, секреции слюны, пиломоторных реакций, дермографизма (изменение окраски
кожи при ее механическом штриховом раздражении), моторики желудка и кишки,
мышечного напряжения, мигания, движения глаз.
Об эмоциональной сфере судят по
содержанию катехоламинов, пептидов и глюкозы в крови, слюне и моче. Показателем
эмоционального напряжения является тремор мышц. На фоне аффекта резко меняются
различные характеристики ЭЭГ. Хотя по ЭЭГ трудно дифференцировать знак эмоции,
установлено, что отрицательные эмоциональные возбуждения характеризуются
длительным последействием и могут суммироваться; положительные —
кратковременным последействием, но устраняющим картину суммарной электрической
активности, присущей отрицательным эмоциональным состояниям. Тесно связан с
эмоциональным состоянием, преимущественно с отрицательным, гиппокампальный
Θ—ритм, однако его изменения сочетаны не только с аффективной сферой, но и
с другими сторонами деятельности мозга. Попытки связать определенную эмоцию с
возбуждением одного из отделов автономной нервной системы не увенчались
успехом. Некоторые исследователи привели доказательства связи эмоций умеренной
интенсивности, если они приятны, с парасимпатическим отделом автономной нервной
системы, если неприятны — с симпатическим.
3.26.4. Участие различных структур мозга в формировании
эмоциональных состояний
В прошлом веке было принято относить
эмоции к функции высших отделов мозга. Однако еще В. М. Бехтерев предположил,
что центром эмоций является таламус. В XX в. У. Кеннон и Ф. Бард создали
теорию, в соответствии с которой эмоционально окрашенное восприятие
определяется первоначально активизацией таламических структур, вторично
влияющих на кору больших полушарий. Д. Линдсли развил основные положения этой
теории, включив в состав эмоциогенных систем ретикулярную формацию мозга.
Согласно его «активационной теории», основанной на данных ЭЭГ, внешние и
внутренние стимулы возбуждают ретикулярную формацию ствола мозга, откуда
импульсы распространяются на гипоталамус, таламус, гиппокамп, кору больших
полушарий. Активационная теория связывает воедино эмоции, кортикальную реакцию
пробуждения, сон и бодрствование, некоторые формы патологического поведения.
Дж. Папес (1937) выдвинул иную
теорию эмоций, приписав главную роль в их формировании лимбической системе
мозга: от гиппокампа импульсы распространяются через свод в сосцевидные
тела, далее в поясную извилину, от нее — на другие области коры больших
полушарий. Значительный вклад в разработку теории эмоций внесли отечественные
физиологи (И. С. Бериташвили, П. К. Анохин, П. В. Симонов, Н. П. Бехтерева и
др.).
Наблюдения в нейрохирургической и
неврологической клиниках и опыты на животных показали, что эмоции нельзя
связать с функцией ограниченного круга мозговых структур, так как каждая из
этих структур может быть связана с положительным и с отрицательным состояниями.
В клинических случаях было
обнаружено, что электрическая стимуляция (проводимая с лечебной целью)
образований среднего мозга ведет к активации с положительной или отрицательной
эмоциональной окраской или к дезактивации, характеризующейся успокоением и
безразличием. Раздражение передней и нижней поверхностей височной доли вызывает
чувство страха; переднего и заднего отделов гипоталамуса — тревоги и ярости;
перегородки конечного мозга — наслаждения; миндалевидного тела — страха,
ярости, гнева, а в некоторых случаях и Удовольствия.
Изменения мотивационно—эмоциональной
сферы наблюдали в многочисленных опытах, проведенных на животных. Еще в конце
прошлого века Ф. Гольтц описал реакцию ярости на самые разные стимулы у собаки
с удаленной корой больших полушарий. Подобные изменения в поведении наблюдали позднее
у декортицированных и децеребрированных животных. Так как полноценное
аффективное поведение невозможно без участия в нем высших отделов мозга,
описанные особенности поведения (опущенная голова, прижатые уши, оскаленные
зубы, выпускание когтей) обозначили как «ложную ярость». Действительно,
комплекс аффективных реакций, вызываемых локальной стимуляцией подкорковых
структур, нельзя отождествлять с полноценным эмоциональным состоянием. Эти
реакции следует рассматривать лишь как отдельные элементы естественного аффекта
(рис. 3.70).
Благодаря
избирательному удалению различных структур мозга удалось определить их участие
в формировании целенаправленного поведения. Так, после разрушения латеральных
отделов гипоталамуса было обнаружено подавление мотивации к потреблению
пищи и воды, сочетающееся с изменениями в эмоциональной сфере животного.
Извращение эмоциональных реакций наблюдали у макак—резус с удаленными височными
долями, миндалевидным телом и передним гиппокампом. Особое место в этих
изменениях играло удаление миндалевидного тела. Функционально оно тесно связано с вентромедиальной
и задней областями гипоталамуса и перегородкой. Повреждение миндалевидного тела
снижает агрессивность животных, но последующее повреждение вентромедиальной области
гипоталамуса вызывает у них проявления злобности. Разрушение зоны перегородки
ведет к агрессивности, которую можно предотвратить, повредив миндалевидное
тело.
Согласно
представлениям П. Маклина, в формировании мотивационно—эмоциональных состояний
играют роль два круга: амигдалярный (миндалевидный) и септальный
(перегородочный). Первый связан с пищевыми и защитными реакциями, второй
—преимущественно с половыми.
Подкорковые
эмоциогенные структуры оказывают активирующее влияние на кору головного мозга.
Она же, в свою очередь, на основании оценки сигналов из внешней и внутренней
среды определяет формирование эмоционально окрашенных целостных поведенческих
актов.
У человека и животных существует
функциональная межполутарная асимметрия. Специализация больших полушарий
головного мозга проявляется ив эмоциональной сфере. Более эмоциогенным является
правое полушарие, играющее преимущественную роль на ранних этапах овладения
навыками. У человека положительные эмоции связаны в основном с левым
полушарием, отрицательные — с правым. Соответственно различается и оценка
афферентации от периферических рецепторов: различные психосоматические
проявления чаще связаны с левой половиной тела. Левая половина лица в большей
степени отражает отрицательные, правая — положительные эмоции. Эти различия
проявляются уже у новорожденных, в частности, в асимметрии мимики при
восприятии веществ, обладающих сладким и горьким вкусом.
Трудности в
определении локализации эмоциогенных структур мозга у животных были в
значительной степени преодолены после открытия в 1954 г. Дж. Олдсом и П.
Милнером феномена внутримозгового самораздражения
|
|
|
Рис. 3.70 Возникновение пищевых и эмоциональных реакций у животного
при поражениях ядер гипоталамуса 1 —
таламус, 2 — путь от таламуса
к гипоталамусу (перерезка), 3, 4 —
гипоталамические ядра, 5 — латеральная область гипоталамуса, 6 — супраоптическое ядро
гипоталамуса, 7 —
вентромедиальные ядра гипоталамуса; поражение вентромедиальных ядер
гипоталамуса вызывает «волчий» аппетит и ярость (А); поражение латеральной области гипоталамуса — потерю
аппетита и исхудание (Б). |

(самостимуляции). Крыса получала возможность, нажимая на педаль, замыкать
электрическую цепь и таким образом стимулировать через вживленные в мозг
электроды различные его отделы. Высокий уровень положительного подкрепления в
тех случаях, когда электрод находился в положительной эмоциогенной структуре
(«зоны награды», «центры удовольствия», «система поощрения»), приводил к
выработке инструментального условного рефлекса в течение нескольких минут.
«Зоны награды» созревают очень рано
и уже в первые дни после рождения включаются в регуляцию поведения. Система
отрицательного подкрепления («зоны наказания») также готова к функционированию на
самых ранних этапах постнатальной жизни. Если электрод вживлен в «зону
наказания», животное, замкнув один раз электрическую цепь, в дальнейшем
избегает нажатий на педаль.
Таким образом, по стремлению
животного к максимизации и минимизации электростимуляции данной «точки» можно
отнести ее к системам положительного или отрицательного подкрепления.
3.26.5. Эмоциогенные системы мозга
У крысы 35% клеточного пространства
мозга связано с положительной эмоциогенной системой, только 5% — с
отрицательной, 60% — с отсутствием побуждения к повторению или избеганию
стимуляции. Принципиальная организация эмоциогенных систем мозга оказалась
весьма сходной у представителей разных отрядов млекопитающих. На рис. 3.71
представлена топография эмоциогенных систем мозга.
В процессе обучения животные выбирают силу тока и ритм
нажатий на педаль, ведущие к наибольшему «вознаграждающему» эффекту. Как и
естественное аффективное поведение, внутримозговая самостимуляция
сопровождается комплексом вегетативных реакций симпато—парасимпатической
природы.
|
|
|
Рис.
3.71.Топография
эмоциогенных зон (заштриховано) мозга у крысы (А), кошки (Б), обезьяны (В) и
человека (Г) |
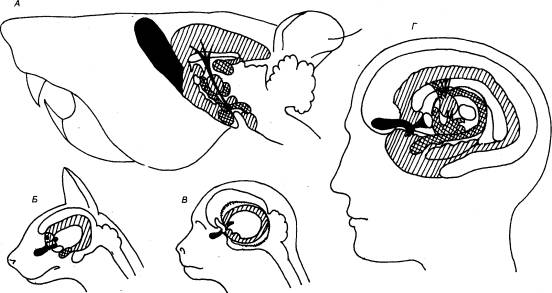
«Зоны награды» находятся
в тесном соседстве с мотивациогенными структурами мозга. В некоторых случаях
слабое принудительное раздражение определенной «точки» вначале вызывает
потребление пищи, воды или другое поведение, направленное на удовлетворение
биологической потребности. При увеличении силы тока животное переходит к
самостимуляции. В других случаях можно выделить «точки», изолированно связанные
только с определенной мотивацией или только с реакцией самораздражения.
Следовательно, мотивациогенные и эмоциогенные структуры представляют собой хотя
и функционально взаимозависимые, но дискретные нейроанатомические системы.
Опыты, проведенные на щенках,
начиная с периода новорожденности, показали, что на разных этапах онтогенеза наблюдается
различное взаимодействие мотивациогенных и эмоциогенных систем мозга.
Свойственное взрослому животному единство мотивационно—эмоционального поведения
является результатом формирования сложных условнорефлекторных стереотипов,
отражающих опыт существования в определенной внешней среде, т. е. наиболее
целесообразных для приспособления к конкретной обстановке.
Пластичность поведения хорошо прослеживается в
экспериментах с телестимуляцией различных мозговых структур (X. Дельгадо). Раздражение определенных
участков таламуса или центрального серого вещества вызывало у вожака колонии
обезьян повышенную агрессивную реакцию, однако это проявлялось лишь по
отношению к сопернику; он никогда не нападал на маленькую самку, свою любимую
подругу. Показано, что в зависимости от ситуации стимуляция одной и той же
«точки» может вызывать различные целенаправленные действия (пищевые, питьевые,
половые, защитные, агрессивные реакции) с присущей им эмоциональной окраской. В
соответствии с подобной пластичностью мотивационно—эмоциональных реакций
телеметрическая стимуляция одной и той же «точки» мозга вызывает самые разные
формы поведения в зависимости от того, исследуется ли оно в лабораторных
условиях или в экологически адекватной среде.
Эмоциональные состояния строятся на
основе определенных нейрохимических механизмов. Фармакологический анализ
показал, что настроение у человека и самостимуляция «зон награды» у животных
зависят от нейромедиаторного баланса в ЦНС. Однако попытка связать определенное
эмоциональное состояние с уровнем какого—либо медиатора потерпела неудачу.
«Точки» мозга, в которых можно
вызывать самораздражение, связаны с дофаминергическими и норадренергическими
системами, снижение их активности подавляет самостимуляцию «зон награды».
Возможно, однако, что положительное эмоциональное возбуждение определяется
норадреналином, тогда как дофамин лишь активирует систему побуждения к
самораздражению.
По другим предположениям,
подкрепляющая функция эмоций зависит от дофамин— и серотонинергических систем,
а мотивационные реакции — от адренергической.
Несомненно доказано участие в
формировании положительного эмоционального состояния эндогенных опиатов. В свою
очередь, эндорфины модулируют состояние серотонинергической системы. По мнению
некоторых исследователей, серотонинергическая и холинергическая системы
обслуживают отрицательные эмоции. Существуют, однако, сведения о подавлении
агрессивных тенденций серотонином.
Таким образом, можно предполагать,
что в реализации различных эмоциональных состояний участвуют все
нейромедиаторные системы мозга.
3.26.6. Влияние эмоциональных состояний на научение и память
Связь мотивациогенных и эмоциогенных
систем мозга проявляется в существенной роли, которую играет уровень
мотивационно—эмоционального возбуждения в процессах научения. Оно затруднено
при низком уровне мотивации и подкрепления. Высокий уровень потребности
формирует выраженное эмоциональное возбуждение. Оно наиболее четко проявляется
на первой стадии выработки условного рефлекса — стадии генерализации, по мере
его упрочения постепенно снижается. Как можно видеть по ряду вегетативных
проявлений (например, частоте сердцебиений), даже оборонительные условные
рефлексы после их упрочения реализуются автоматически без каких—либо проявлений
эмоционального возбуждения.
Оборонительные условные рефлексы,
связанные с отрицательным подкреплением, формируются значительно быстрее и
длительнее сохраняются, чем условные рефлексы, вырабатываемые при положительном
подкреплении (например, пищедобывательные). Так, условнорефлекторные вкусовые
аверсии — отказ от пищи, потребление которой вызвало отравление, —
вырабатываются при однократном сочетании вкусовых сигналов с последущим
дискомфортным состоянием, причем этот рефлекс может сохраняться в течение
многих месяцев, а иногда и лет.
Принято считать, что
запоминание и длительность сохранения памятного следа пропорциональны
эмоциональному напряжению. Однако, как показали эксперименты на добровольцах,
низкий и высокий его уровни отрицательно сказываются на обучении и процессах
памяти. Наиболее благоприятным для них оказался умеренный уровень
эмоционального напряжения, соответствующий умеренной активации ЦНС. Наблюдения
на школьниках разного возраста показали, что в I—III, IX и Х классах положительное
подкрепление улучшало, а в IV—VIII — ухудшало запоминание. Полагают, что
разнонаправленные влияния эмоционально—положительной и отрицательной систем на
мнестические процессы данных возрастных групп определяются гормональными
факторами, соответствующими .этим периодам развития.
В опытах на животных
было показано, что условнорефлекторные реакции, подкрепляемые
непосредственной электрической стимуляцией эмоциогенных структур мозга,
формируются очень быстро. При этом можно переделать значимость исходно
отрицательных раздражителей. Так, у собак вырабатывали положительную реакцию на
вкус горького раствора, сочетаемого со стимуляцией «зон награды». Этот же
механизм, по—видимому, лежит в основе условнорефлекторного формирования
предпочтения человеком веществ с исходно индифферентными или отрицательными
вкусовыми свойствами, если они оказывают положительный эмоциональный эффект.
Привычка потреблять эти вещества может быть чрезвычайно сильной, а в некоторых
случаях перейти в патологическое влечение. Доказано, что начальные стадии
формирования наркомании определяются эйфорией, связанной с активацией «зон
награды».
Влияние эмоциональных
состояний на научение и память определяется также модулирующим действием
подкрепляющих систем мозга на сенсорные процессы (зрительная, слуховая,
вкусовая, болевая и другие системы). В частности, стимуляция «зон награды»
изменяет электрические характеристики нейронов ряда анализаторов на разных
уровнях мозга, т. е. эмоциогенные системы мозга оказывают модулирующее влияние
на обработку сенсорной информации.
Влияния, оказываемые
эмоциональными состояниями на сенсорные процессы, научение, память,
характеризуются разной степенью напряженности.
В ряде случаев преимущественно
обращают внимание на аффекты, крайние эмоциональные состояния,
характеризующиеся выраженными субъективными и объективными показателями. Однако
эмоциональная окраска присуща любой деятельности и не обязательно проявляется
как ярко окрашенное переживание. Состояния общего комфорта или дискомфорта,
отражающие баланс между уровнем активности положительных и отрицательных
эмоциогенных систем, являются основой организации целостных поведенческих
актов.
Наряду с исследованием
физиологических механизмов аффективных состояний было обращено внимание на
разработку общетеоретических представлений о роли эмоций в организации
поведения. Согласно биологической теории П. К. Анохина, эмоции возникли в
процессе эволюции как средство быстрой оценки потребностей и их удовлетворения
в соответствии с состоянием внешней ситуации. Если параметры достигнутого
эффекта соответствуют свойствам акцептора результатов действия, возникает
положительная эмоция, если не соответствуют — отрицательная.
Потребностно—информационная теория П. В. Симонова рассматривает эмоцию
как отражение мозгом человека и высших животных качества и величины определённой
потребности и вероятности ее удовлетворения. Регуляторная роль эмоций
проявляется в их отражательно—оценочной, переключающей, подкрепляющей,
компенсаторной (замещающей) функциях. Подобный подход к рассмотрению эмоций
сближает физиологические, психологические и философские аспекты изучения этого
важнейшего феномена в поведении человека и животных.
Другая сторона исследования эмоций
связана с разработкой вопросов их нарушения. Ослабление деятельности
систем положительного подкрепления или усиление деятельности системы
отрицательного подкрепления приводит к гипотимии (от греч. hypo — понижение и thymos — настроение, чувство) —
депрессивному состоянию с проявлением тревоги, страха, апатии. При этом
происходит ослабление мотивационных процессов, снижение массы тела, нарушение
работы внутренних органов.
При гипертимии наблюдаются
повышенное настроение, усиление двигательной активности, легкая отвлекаемость.
Нередко фазы гипотимии и гипертимии перемежаются. У человека описаны и другие
разновидности нарушения эмоциональной сферы, причины которых различны. Как
правило, в основе глубоких нарушений эмоциональной сферы лежат генетические
факторы. Обычно гипо— и гипертимия сопровождаются значительными отклонениями в
балансе нейромедиаторов. Так, при маниакальных и депрессивных состояниях
изменяется синтез дофамина, норадреналина и серотонина в разных отделах мозга.
Ряд симптомов этих заболеваний, в частности, тревожность и депрессию, можно
воспроизвести на животных с помощью введения нейротоксинов или фармакологических
веществ, подавляющих синтез норадреналина, серотонина и опиатов.
При взаимодействии человека и животных с внешней средой
возникают ситуации, характеризующиеся конфликтом между потребностями и возможностями
их удовлетворения. Они ведут к формированию состояния напряжения — эмоционального
стресса, приспособительное значение которого проявляется в мобилизации
защитных сил, направленных на преодоление конфликта. Невозможность его
разрешения приводит к формированию длительного застойного эмоционального
возбуждения, проявляющегося в нарушениях мотивационно—эмоциональной сферы и в
различных соматических заболеваниях.
Эмоциональный стресс может привести
к развитию ишемической болезни сердца, гипертензии, язвообразованию, дисфункции
эндокринной системы. При этом наблюдаются глубокие изменения баланса
нейромедиаторов и нейропептидов в ЦНС. У человека такие нарушения связаны
преимущественно с социальными конфликтами. Некоторые из этих проявлений могут
быть смоделированы на животных. Так, у вожака стада обезьян, изолированного, но
имеющего возможность наблюдать последующие иерархические изменения во
взаимоотношениях ранее подчинявшихся ему животных, развивается артериальная
гипертензия, а в некоторых случаях и инфаркт миокарда.
Степень устойчивости животных к действию стрессорных
факторов различна и может быть установлена в специальных экспериментах. Так,
менее устойчивые к стрессу животные отвечают на стимуляцию отрицательных
эмоциогенных зон вентромедиального гипоталамуса преимущественно прессорными
сосудистыми реакциями, а более устойчивые — прессорно—депрессорными. Снижение
эмоционального напряжения и, соответственно, предотвращение висцеральных
нарушений достигается стимуляцией «зон награды» или введением фармакологических
препаратов, нормализующих баланс медиаторов в ЦНС.
Исследование различных проявлений
эмоционального стресса оказалось полезным для разработки проблемы неврозов —
группы обратимых функциональных расстройств психогенной природы. В отечественной
литературе принято выделять три формы невроза: истерический, навязчивых
состояний и неврастению. Возникновение невроза и его форма определяются
взаимодействием психотравмирующей ситуации с исходными характеристиками
личности. В ряде случаев невротическое состояние развивается на фоне
недостаточности структур лимбико—ретикулярного комплекса. Эмоциональные
нарушения занимают в симптоматике неврозов одно из ведущих мест.
И. П. Павловым было введено понятие экспериментальные
неврозы. — функциональные нарушения (срыв) высшей/нервной деятельности в
результате перенапряжения основных нервных процессов. Учение И. П. Павлова об
экспериментальных неврозах оказалось плодотворным для понимания ряда сторон
патологии психической деятельности у человека. Однако нельзя отождествлять
сложную клиническую картину невроза у человека с нарушениями поведения у
животных, на которых можно лишь моделировать определенные симптомы заболевания.
Тенденция к отождествлению большей части функциональных нарушений поведения
животных с экспериментальными неврозами создала положение, которое затрудняет
прямое сопоставление клинических и экспериментальных данных. Вместе с тем
возможность исследовать у животных отдельные проявления невроза у человека, а
также проводить их фармакологический анализ, приносит существенную пользу
неврологической клинике.
Основными приемами выработки экспериментального невроза И.
П. Павловым и его сотрудниками было создание конфликтной ситуации при
столкновении разнонаправленных мотиваций, например, пищевой и оборонительной,
или же при необходимости решения трудной задачи (дифференцирование близких по
параметрам условных сигналов и т. п.).
Массовое проявление невротических реакций у собак,
находившихся в виварии, затопленном во время ленинградского наводнения 1924 г.,
позволило И. П. Павлову сделать ряд важных обобщающих выводов о закономерностях
развития патологии высшей нервной деятельности. Он постулировал зависимость
невротизации от силы, подвижности и уравновешенности корковых
процессов возбуждения и торможения. В соответствии с ними была предложена
классификация темпераментов у человека.
Эксперименты, проведенные на
собаках, показали, что неврозы легче возникают у животных слабого и сильного
неуравновешенного типов высшей нервной деятельности. Анализируя происхождение
истерии, И. П. Павлов рассматривал ее как следствие слабости нервной системы.
Современные методы исследования усложнили эту схему.
Оказалось, что при экспериментальном неврозе функциональные изменения возникают
прежде всего в лобных отделах новой коры, лимбических структурах и ретикулярной
формации среднего мозга. Нарушается баланс нейромедиаторов в ЦНС и в крови, в
частности, между катехоламинами и ацетилхолином. Хотя неврозы и рассматривают
как функциональные заболевания, выяснилось, что они сопровождаются развитием
реактивных и дегенеративных процессов в различных отделах мозга. Показана роль
информационных факторов в развитии патологии мотивационно—эмоционального
поведения. Большую роль при этом играют нарушения памяти.
Изучение патологии высшей нервной деятельности подтвердило
роль обратной связи от соматических и висцеральных систем в поддержании
отрицательного эмоционального состояния. Были приведены факты,
свидетельствующие в пользу существования эмоциональной памяти
(«аффективной памяти», «памяти чувств»), на которую впервые обратил внимание в
конце прошлого века психиатр С. С. Корсакова, а И. С. Бериташвили выделил ее
как особую форму памяти.
Она особенно четко проявляется при неврозах, в основе
которых лежат инертные очаги возбуждения (травматические неврозы,
патологические влечения — алкоголизм, наркомания).
Эмоциональные условные реакции могут усиливаться или
тормозиться внешними раздражителями, преимущественно ситуационными. Так, уже
упоминавшиеся вкусовые аверсии могут быть резко ослаблены в привычной
обстановке и в полной мере проявиться в непривычной. У человека и животных
отрицательные эмоциональные реакции со всем комплексом вегетативных проявлений
могут воспроизводиться при столкновении с ситуацией, сопровождавшейся
травматизирующими факторами, даже если со времени их воздействия прошло много
лет.
Эмоциональное состояние определяется генотипическими
характеристиками и условиями, в которых проходило развитие животных. Особо
важную роль при этом играют ранние воздействия. Так, изоляция от сверстников
или от взрослых особей в раннем онтогенезе повышает неустойчивость к
невротизирующим стимулам. Обогащенная или обедненная раздражителями внешняя
среда сказывается на морфофункциональном развитии мозга, в частности, его
эмоциогенных систем. Эти данные, полученные на животных, позволяют разработать
рекомендации для создания условий воспитания детей, начиная с периода
новорожденности, способствующих оптимальному развитию их
мотивационно—эмоциональной сферы.
3.27. ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР
Из пищеварительного тракта в кровь
поступают продукты расщепления пищи, и некоторые из них в случае проникновения
в мозг могли бы нарушить его функции. Например, переваривание белков приводит к
повышению уровня в плазме крови аминокислот глицина и глутамата, которые в
мозге играют роль медиаторов. Высокая двигательная активность также нарушает
постоянство внутренней среды организма. Поведение низших животных, не имеющих
гематоэнцефалического барьера (ГЭБ), сильно зависит от состава пищи и
равномерности ее поступления в организм, что привязывает каждый вид к
конкретному источнику питания и делает невозможными активные перемещения в
пространстве для эффективного поиска пищи или половых партнеров.
Возникновение в ходе эволюции
способности к зоосоциальному поведению и простейшим формам научения было
связано с усложнением структуры и функций мозга, что потребовало поддержания в
нем более строгого постоянства внутренней среды. Поскольку достичь такой
степени гомеостатирования во всем организме невозможно, у высокоорганизованных
животных вокруг ЦНС выделилось особое пространство, окруженное барьером,
непроницаемым для большей части веществ, содержащихся в крови (гемолимфе) и
тканевой жидкости. К двум хорошо известным пространствам — внутриклеточному и
внеклеточному — добавилось третье пространство, содержащее
цереброспинальную (спинномозговую) жидкость, или ликвор. Так эволюция сделала
шаг сопоставимый по значению с появлением внутренней среды организма.
Создание особой внутренней среды
мозга, во—первых, защитило его нейроны от колебаний внутренней среды организма,
связанных с потреблением пищи и двигательной активностью, и обеспечило
стандартные условия для интегративной деятельности нейронов и синаптической
передачи; во—вторых, позволило нейронам мозга при передаче друг другу сообщений
гуморальным путем экономно использовать запасы нейросекретов и уменьшило
искажение передаваемой информации.
Потребность в барьере
была настолько сильной, что он независимо возник не менее чем в трех ветвях
эволюции, выделившихся 500—600 млн. лет назад, и имеется у всех таксонов,
способных к сложному поведению: у позвоночных головоногих моллюсков и
членистоногих (у насекомых, некоторых паукообразных и отчасти у ракообразных).
Структура барьера. Мысль о существовании барьера между
кровью и мозгом впервые высказал Пауль Эрлих в 1885 г. К этому времени
накопились данные о том, что красители (например, трипановый синий) после
введения их в кровь не обнаруживаются в мозге. В 1913 г. Голдманн показал, что
если краситель вводить не в вену, а в ликвор, то мозг окрашивается. Тем не
менее долгое время существовала и противоположная точка зрения: барьера нет,
красители не проходят в мозг потому, что мозг не содержит соединительной ткани
т. е. для красителя в мозге просто нет места. Только в 60—е годы мнение о
существовании ГЭБ стало общепринятым.
Термин «гистогематические барьеры» предложила наша соотечественница Л. С. Штерн в 1929 г. Сейчас он понимается как общее название для барьеров: гематоэнцефалического (ГЭБ),
—ликворного (ГЛБ), —неврального, —офталъмического, —тестикулярного,
—лабиринтного, —плеврального, —синовиального
и др. В 1933 г. Вальтер и Шпац
ввели понятие «гематоэнцефалический
барьер».
Как известно, капилляры мозга принципиально отличаются от
капилляров всех других областей тела отсутствием в их стенке водных каналов,
пор, или фенестр. Под электронным микроскопом отчетливо видно, что
эндотелиоциты мозговых капилляров соединены между собой так называемыми плотными
контактами, которые не позволяют водорастворимым веществам проходить из
крови в головной мозг (и обратно). На электронограммах также видны и отростки астроцитов,
охватывающие всю (или почти всю) наружную поверхность мозговых капилляров (рис.
3.72).
Эти отростки не составляют механического препятствия
проникновению веществ в мозг, как думали прежде, но они секретируют некие пока
недостаточно изученные, вещества, повышающие плотность контактов между
|
|
|
Рис. 3.72 Ножки отростков астроцитов (А)
окружают мозговой капилляр (К) |
|
|
|
Рис. 3.73 «Энзимный барьер» МАО — моноаминооксидаза, КОМТ —
катехол—0—метил—трансфераза, ЭП — эпендима, ЭН — эндотелий. |
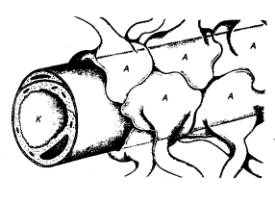
эндотелиоцитами. Известно, что по
крайней мере часть этих веществ имеет молекулярную массу менее 4 кДа и,
следовательно, не относится к белкам.
В середине 60—х годов было открыто существование «энзимного
барьера», т. е. ферментов в тканях, лежащих между просветом капилляров и
нейронами мозга. Эти ферменты расщепляют вещества, которые в противном случае
могли бы проникнуть из крови в мозг. Примерами таких «барьерных» энзимов могут
служить катехол—0—метилтрансфераза (КОМТ), моноаминоксидаза (МАО),
тирозингидроксилаза и аминопеппгидаза. Энзимный барьер в большей
степени препятствует проникновению в мозг моноаминов (потенциальных
нейротрансмиттеров), чем их непосредственных предшественников (рис. 3.73).
Феномен ГЭБ пытались объяснить еще и
тем, что введенный в кровоток краситель связывается с белками плазмы и это
делает его неспособным проникнуть сквозь барьер. Однако связывание с белками
плазмы не препятствует окрашиванию всех прочих (немозговых) тканей, поэтому
сегодня фактору связывания придается небольшое значение.
Итак, ГЭБ состоит, по меньшей мере,
из трех главных компонентов: (1) плотных контактов в эндотелии капилляров; (2)
веществ, секретируемых отростками астроцитов и поддерживающих функцию плотных
контактов; и (3) «барьерных энзимов».
Проницаемость барьера и транспортные системы. В 1902 г. Эрлих заметил, что
красители, поглощаемые мозгом, поглощаются и жировой тканью тоже. Значение
этого факта было оценено только в 70—е годы, после создания высокоточного
метода количественной оценки проницаемости ГЭБ.
Для жирорастворимых веществ ГЭБ барьером вообще не является: они
простой диффузией легко проходят сквозь эндотелиоциты: сначала растворяются в
липофильных участках люминальной (т. е. обращенной в сторону просвета
сосуда) мембраны, затем диффундируют через всю толщу цитоплазмы и, наконец,
растворяясь в аблюминальной (расположенной с другой, наружной по
отношению к просвету сосуда стороны) мембране, выходят из эндотелиоцита в ткань
мозга (рис. 3.74, путь 1А).
Никакие живые клетки не являются препятствием для веществ,
обладающих высокой липофильностью (или, что то же самое, гидрофобностью).
Наиболее высокой «проходимостью» обладает углекислый газ, легко
|
|
|
Рис. 3.74 Пути проникновения веществ из крови в мозг ГЭБ — гематоэнцефалический барьер;
ЦВО — циркумвентрикулярные органы; ВСС — внутрижелудочковое сосудистое
сплетение; ЦСЖ — цереброспинальная жидкость (ликвор). 1 — сквозь нефенестрированный эпителии: А — простой диффузией;
Б — активным транспортом; 2 —
через фенестры эндотелия капилляров в ЦВО и далее: А — внутриаксональным
транспортом; Б — сквозь эпендиму простой диффузией в ликвор; В — вдоль
наружной границы эпендимы желудочков до ее участков, где нет плотных
контактов (гипотетический путь); 3 — сквозь стенки сосудистого сплетения в
ликвор (простой диффузией); 4 —
со слизистой оболочки полости носа (конкретный путь неясен); 5 —
внутриаксональным транспортом (например, по чувствительным нейронам). Для
жирорастворимых веществ наибольшее значение имеют пути 1А, 2Б и 3, для
водорастворимых — 2А, 2В, 4 и 5. Для водорастворимых веществ, нужных мозгу в
значительных количествах (глюкоза, аминокислоты и др.) существуют системы
переносчиков. |
проникающий
из крови в нейроны
дыхательного центра. Довольно высокой растворимостью, соответственно,
«проходимостью» обладают героин, алкоголь и никотин, что объясняет их быстрое (после
попадания в кровь) действие на ЦНС. Следовательно, липофильность — главное из
качеств, определяющих возможность простои диффузии вещества сквозь ГЭБ. На «проходимость» вещества
диффузией также влияют степень его ионизации, молекулярная масса и некоторые
другие факторы.
Мозг нуждается в некоторых веществах, не обладающих
способностью растворяться в жирах, например, в глюкозе — для обеспечения своих
энергетических потребностей, и в аминокислотах — для синтеза белков. Поскольку
гидрофильные вещества простой диффузией пройти в мозг не могут, для их доставки
существуют транспортные системы со специфическими белками—переносчиками (рис. 3.74, путь 1Б). Сейчас известны
транспортные системы для: (а) D—глюкозьг; (б) крупных нейтральных аминокислот;
(в) основных и кислых аминокислот; (г) электролитов (K+, Mg2+, Ca2+, I— и др.); (д)
водорастворимых витаминов; (е) нуклеозидов и др. Все эти системы обладают
общими свойствами: селективностью, стереоспецифичностью, конкурентным
ингибированием и насыщаемостью,
Мелкие нейтральные аминокислоты не транспортируются, так как
мозг синтезирует их сам. Транспортная система, переносящая глюкозу, не требует
для своей работы присутствия инсулина. Необычно то, что в ГЭБ инсулин регулирует транспорт не
глюкозы, а аминокислот.
«Дефекты» в
барьере и пути в обход барьера. В головном мозгу есть области, в которые, в
отличие от основной его массы, краситель легко проходит из крови. Впервые это
обнаружил Шульманн в 1912г.: трипановый синий, введенный мышам внутривенно,
проникал в нейрогипофиз. Количество областей, обладающих такими особенностями,
постепенно росло, и сейчас включает:
нейрогипофиз,
медиальное возвышение, самое заднее поле, субфорникальный орган, шишковидное
тело, зрительное углубление, сосудистое сплетение и сосудистый орган конечной
пластинки. Поскольку все эти структуры расположены по периферии желудочковой
системы, было предложено называть их циркумвентрикулярными органами (рис.
3.75). Только один из них не обладает названными особенностями: в
субкомиссуральном органе эндотелиоциты соединены плотными контактами.
|
|
|
Рис. 3.75 Циркумвентрикулярные органы Э — шишковидное тело (эпифиз), ВСС — сосудистое сплетение,
СКО — субкомиссуральный орган, СФО — субфорникальный орган, MB — медиальное возвышение, СОКП — сосудистый
орган конечной пластинки, НГ — нейрогипофиз, СЗП — самое заднее поле. |
|
|
|
Рис. 3.76 Принципиальное различие
устройства гематоликворного (А) и гематоэнцефалического (Б) барьеров Плотные
контакты (ПК) соединяют либо клетки эндотелия капилляров (К), либо клетки
эпендимы (Э), выстилающей стенки желудочков мозга. ИП — интерстициальное
пространство; Н — нейрон; Г — глия; Ф — поры (фенестры); ЦСЖ —
цереброспинальная жидкость (ликвор); S — площадь барьера. Стрелками
показано проникновение водорастворимых веществ. |
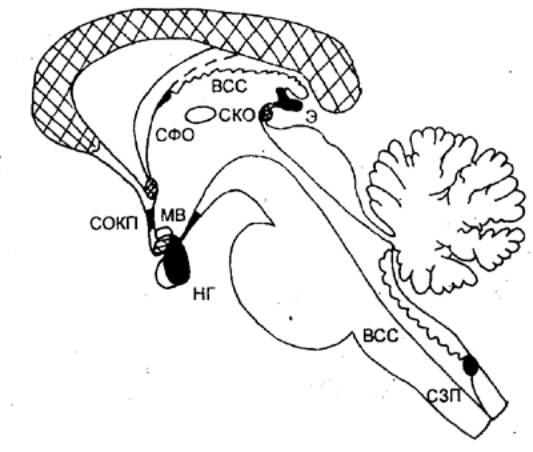
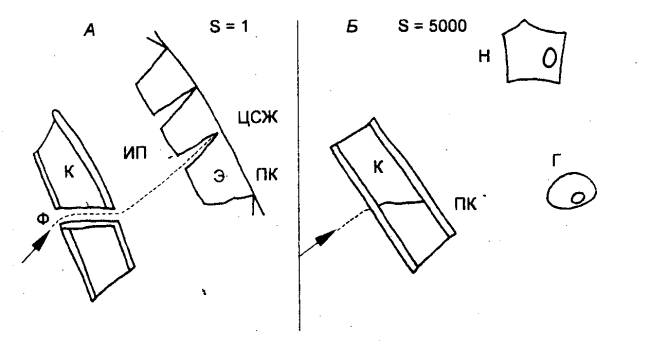
В циркумвентрикулярных органах переносимые кровью вещества относительно
свободно покидают просвет капилляров (рис. 3.74., путь 2) и доходят до наружной
границы желудочков мозга, где их задерживают плотные контакты, которыми соединены
клетки выстилающей желудочки эпендимы. Эту особую организацию барьера в
циркумвентрикулярных органах часто ошибочно принимают за «дефекты» в барьере. В
действительности, барьер в этих областях не менее эффективен, только
разделительную функцию выполняет не эндотелий капилляров, а эпендима желудочков
мозга. Таким образом, барьер здесь просто отнесен чуть далее вглубь мозговой
ткани и носит название гематоликворного (ГЛБ). Площадь ГЛБ составляет
1/5000 общей площади ГЭБ (рис. 3.76).
Полагают, что некоторая часть молекул веществ, оказавшихся в
циркумвентрикулярных органах, может проникнуть в собственно мозг:
— через нервные окончания и далее внутриаксональным
транспортом (см. рис. 3.74, 2А);
— сквозь клетки эпендимы — в ликвор,
в соответствии с растворимостью данного вещества в липидах (рис. 3.74, 2Б);
— перемещением вдоль наружной границы эпендимы до тех участков,
где между ее клетками есть водные каналы, и по ним — в ликвор (рис. 3.74, 2В).
Некоторая часть сосудов мозга выпячивается в желудочковую систему
и образует в ней сосудистое сплетение — орган, продуцирующий ликвор.
Капилляры этих сплетений имеют фенестры, сквозь которые вещества легко покидают
кровь, но в дальнейшем задерживаются хориоидным эпителием, составляющим единое
целое с остальной эпендимой. Дальнейшее продвижение (в ликвор) возможно только
простой диффузией, в соответствии с липофильностью конкретного вещества (рис.
3.74, путь 3).
Возможно,
единственный путь, по которому водорастворимые вещества в значительных
количествах без помощи специальных транспортных систем попадают в мозг,
начинается на слизистой оболочке полости носа (рис. 3.74, путь 4). Пероксидаза
хрена (белок с молекулярной массой 40 кДа), введенная интраназально, за
несколько минут достигает пиальной поверхности обонятельных луковиц. Водные
растворы нейропептидов вызывают ЦНС—зависимые эффекты (например,
интраназальное введение человеку вазопрессина приводит
|
|
|
Рис. 3.77 Пути воздействия веществ на мозг без их
проникновения сквозь гематоэнцефалический барьер 1—3 — связывание веществ со специфическими рецепторами в
циркумвентрикулярных органах приводит к: увеличению проницаемости ГЭБ для
другого вещества (1)—образованию в циркумвентрикулярных органах
жирорастворимых веществ, проникающих далее сквозь эпендиму желудочков простои
диффузией (2А); образованию в самом мозгу гипотетических водорастворимых
веществ—посредников (2Б); передаче возбуждения в виде потенциалов действия
(3); 4 — связывание с пресинаптическими рецепторами эфферентов имитирует
центральное действие; 5 — воздействие на периферические органы через
возбуждение центральных афферентов изменяет состояние ЦНС. Остальные
обозначения, что и на рис. 3.74 |
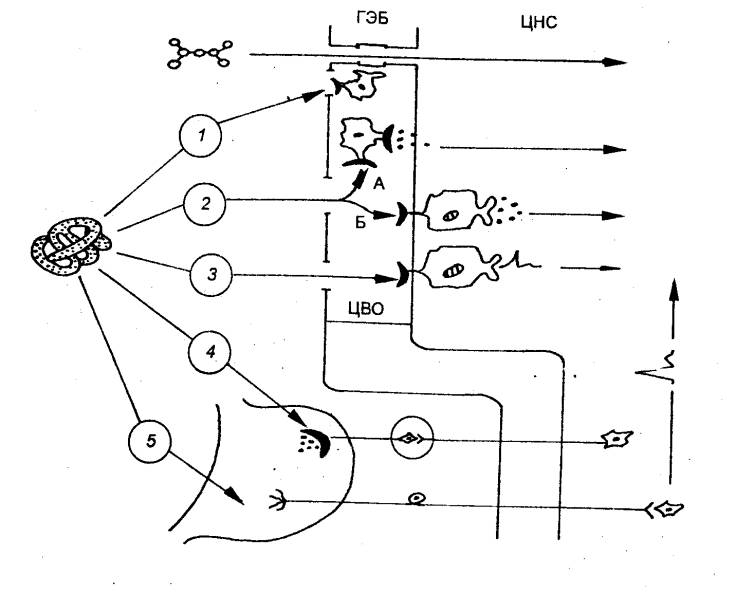
к отчетливому повышению настроения).
В этом случае диффузия происходит либо внутри аксонов обонятельных нейронов,
либо через подслизистый слой связанный с подпаутинным пространством в области
обонятельных луковиц. Нарушение целостности слизистой оболочки (например, при
рините) способствует проникновению веществ в мозг именно этим путем.
Гидрофильные
молекулы из крови могут попадать в мозг внутриаксональным транспортом,
например, по волокнам чувствительных нейронов (рис. 3.74, путь 5)
Рецепторы
наружной поверхности барьера. Есть несколько механизмов посредством которых
водорастворимые вещества, находящиеся в плазме крови могут воздействовать на
мозг без проникновения в него (рис. 3.77), и часть этих механизмов связана с
существованием на «внешней» стороне барьера специфических рецепторов к многочисленным
гормонам белково—пептидной природы
В 50—е годы было
показано, что если разрушить отдельные циркумвентрикулярные органы, то
некоторые гормоны, переносимые плазмой крови теряют способность действовать на
мозг. Позднее были открыты расположенные в циркумвентрикулярных органах
рецепторы к соматостатину, кортикотропину (адренокортикотропному гормону),
пролактину, вазопрессину, окситоцину паратирину (паратгормону), кальцитонину,
инсулину и инсулиноподобным факторам роста, эндотелину, панкреатическому полипептиду,
ангиотензину II атриальному и мозговому натрийуретическим пептидам,
брадикинину, вазоактивному интестинальному полипептиду, субстанции Р и
нейропептидам У и YY
Возбуждение
«внешних» рецепторов молекулами одного вещества может увеличить проницаемость
барьера для другого находящегося в плазме крови вещества (рис. 3.77, путь 1).
Это может выражаться в стимулировании работы
одной из систем активного транспорта (например, внутривенное введение
вазопрессина увеличивает транспорт в мозг лейцина, некоторые гормоны регулируют
проникновение в мозг моноаминов) или в снижении сопротивления барьера простой
диффузии веществ (так, вазопрессин увеличивает проницаемость ГЭБ для молекул
воды). Взаимодействие лиганда с рецептором
на наружной стороне барьера может также приводить к образованию
веществ—посредников, способных воздействовать непосредственно на нейроны мозга
(рис. 3.77, путь 2). Место синтеза таких посредников определяется их
способностью проникать сквозь барьер. Жирорастворимые вещества—посредники синтезируются
на внешней стороне барьера, как правило, в циркумвентрикулярных органах (см.
рис. 3.77, путь 2А). Так, образующиеся в периферических тканях интерлейкины —
полипептиды с большой молекулярной массой (примерно от 8 до 26 кДа) и низкой
растворимостью в жирах, попадая в сосудистый орган конечной пластинки,
стимулируют образование в нем липофильных молекул простагландинов, которые
проникают в мозг и, воздействуя на центры терморегуляции, вызывают повышение
температуры тела — характерное проявление лихорадки.
Водорастворимые вещества—посредники,
не обладающие способностью проникать сквозь ГЭБ, могут синтезироваться и
высвобождаться только «за барьером», в самом мозге (рис. 3.77, путь 2Б). Иногда
в качестве посредника выступает то же самое вещество, которое воздействовало на
«внешние» рецепторы. Так, в ответ на поступление пищи в кишку расположенные в
ее слизистой оболочке эндокринные клетки высвобождают в кровь холецистокинин
(ХЦК). Одновременно возрастает и концентрация ХЦК в мозге. Однако ХЦК, как
и большинство других пептидов, не обладает сколько—нибудь заметной
растворимостью в жирах, поэтому простой диффузией пройти в мозг он не может.
Специальной транспортной системы для ХЦК в барьере также не обнаружено.
Полагают, что молекулы ХЦК, не проникая в ЦНС, связываются с
холецистокининовыми рецепторами на наружной стороне ГЭБ и активируют
неизвестный пока механизм, способствующий высвобождению «по ту сторону барьера»
таких же молекул ХЦК, но синтезированных в самом мозге. Этот механизм играет
важную роль и в возникновении такого чувства, как насыщение.
Предполагают также, что
чувствительные окончания некоторых нейронов мозга выходят на внешнюю сторону
ГЭБ и через них возбуждение от перечисленных выше рецепторов может передаваться
в мозг виде потенциалов действия (рис. 3.77, путь 3).
Таким образом, с помощью
циркумвентрикулярных органов мозг отвечает на колебания концентраций в крови
многочисленных биологически активных веществ. При этом циркумвентрикулярные
органы могут быть столь же непроницаемы, как и другие части ГЭБ.
Вещества, циркулирующие с кровью,
могут воздействовать на ЦНС и опосредованно, через действие на периферические
органы (рис. 3.77, путь 4). Например, пептидный гормон бомбезин
связывается со специфическими рецепторами в гладких мышцах пилорической части
желудка, что приводит к повышению мышечного тонуса привратника и способствует
задержке пищи в желудке. Импульсация от механорецепторов стенки заполненного
пищей желудка по чувствительным волокнам блуждающего нерва передается в ЦНС, в
результате чего частично подавляется пищевое поведение. Так воздействие
вещества на периферический орган вызывает поведенческий (т. е. центральный)
эффект.
Гидрофильные вещества, растворенные
в плазме крови, могут не только действовать на ЦНС, но и имитировать такое действие,
усиливая или ослабляя высвобождение медиаторов из периферических нервных
окончаний (рис. 3.77, путь 5). Так, тиролиберин и ангиотензин II,
связываясь с пресинаптическими рецепторами симпатических нервных окончаний,
облегчают высвобождение ими норадреналина, имитируя влияние на тонус
симпатических нейронов спинного мозга.
Пути выхода веществ из мозга в
кровь. Специальные
транспортные системы выносят из мозга в кровь метаболиты самого мозга (их
накопление могло бы
|
|
|
Рис. 3.78 Неизбирательный механизм
выноса веществ из мозга в кровь. Паутинная оболочка в виде ворсинки (В)
проникает сквозь мозговую оболочку в венозный синус (С). Каналы, имеющиеся в
ворсинке, позволяют небольшим количествам ликвора проникать из подпаутинного
пространства (П) в кровь. Э — эндотелий. |
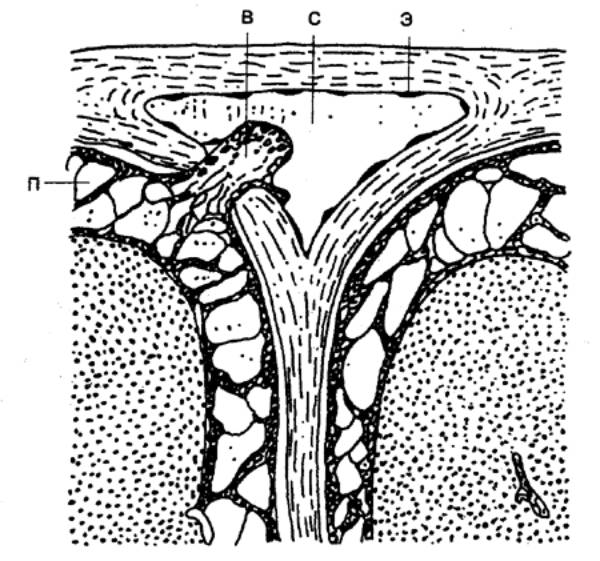
нарушить его работу) и токсические
вещества, которые все—таки в небольших количествах проникают в мозг сквозь ГЭБ.
Самый простой,
неизбирательный механизм выноса состоит в том, что избыток ликвора периодически
сбрасывается в кровь через ворсинки паутинной оболочки, выдвинутые в
просвет венозных синусов (рис. 3.78).
Ворсинки выполняют
роль клапанов, регулирующих отток: быстрое поступление ликвора в венозный синус
повышает давление в нем, ворсинка сдавливается, и отток ликвора прекращается. У
человека за 1 сут образуется примерно 500 мл ликвора, следовательно,
сопоставимое количество его оттекает ежедневно в венозную кровь.
Существуют и специфические
транспортные системы,
каждая из которых выводит из мозга против градиента концентрации конкретное
вещество или группу структурно родственных веществ:
ионы К+; мелкие
нейтральные аминокислоты; глицин, аспарагиновую и глутаминовую кислоты; пептиды
из семейства вазопрессина/окситоцина; меланостатин и энкефалины; нуклеозиды.
Эти транспортные системы необходимы для поддержания гомеостаза мозга. Так,
например, выведение глицина и K+ нормализует уровень
возбудимости ЦНС, удаление избытка меланостатина повышает активность опиоидной
системы мозга.
Создание в ходе
эволюции механизмов, выносящих в кровь пенициллины, цефалоспорины и некоторые
цитостатики, в нормальных условиях отсутствующие в мозге и применяемые при
лечении воспалительных процессов ЦНС, объясняют тем, что небольшие количества
этих веществ синтезируются микрофлорой кишечника, а также попадают в организм с
растительной пищей и проникают сквозь ГЭБ в мозг.
Важную роль в
системах, выводящих вещества из мозга в кровь, играет особый белок — гликопротеин
Р (Pgp).
Обычно он присутствует в ГЭБ в небольших количествах. При наличии в мозге
очагов воспаления или опухолей проницаемость барьера возрастает, введенные в
кровь антибиотики и цитостатики легко проникают в ЦНС. В ответ на это
происходит экспрессия гена, ответственного за синтез Pgp, его количество в
структуре ГЭБ возрастает и он быстро выводит названные вещества из мозга.
Попытки проведения
различных веществ сквозь барьер. Первый из возможных подходов состоит в том, чтобы
воздействием на ГЭБ неизбирательно и обратимо повысить его проницаемость для
любых веществ. Этого добиваются следующими нефизиологическими мерами: (а)
повышением артериального давления (в том числе локальным, в сонной артерии) на
60—90 мм рт. ст. и более, что приводит
к резкому расширению сосудов мозга; (б) повышением осмотического давления крови
путем введения в сонную артерию маннитола, арабинозы, мочевины или
гипертонического раствора NaCI, что вызывает сморщивание эндотелиоцитов; (в)
вдыханием газовой смеси, содержащей 25% СО2, что обусловливает
расширение артериол мозга и переполнение капилляров. Во всех перечисленных
случаях происходит временное нарушение плотных контактов между эндотелиоцитами
и образование водных каналов. Очевидно, что такие способы повышения
проницаемости ГЭБ не годятся для проведения лекарств в мозг человека.
Еще один подход — модификация
молекул с целью повышения их «проходимости» сквозь неизмененный барьер. Поиск
таких форм ведется по следующим направлениям: (а) повышение жирорастворимости
вещества, например, соединением его молекулы с молекулой холестерина; (б)
повышение устойчивости вещества к воздействию «энзимного барьера», например,
для пептидов это создание циклических структур или замещение L—форм аминокислот
на D—изомеры; (в) соединение препарата с веществом, для которого в ГЭБ
существует транспортная система, например, с глюкозой (создание так называемых химер);
и (г) включение изучаемого вещества в липосомы. Пока эти разработки
имеют лишь ограниченный успех.
Рассматривая механизм действия на
ЦНС любого вещества — эндогенного, присутствующего в крови, или вводимого извне
в кровь, необходимо понять, проходит ли оно сквозь ГЭБ или воздействует на мозг
иным образом, без проникновения в него.
Краткое знакомство со структурой и
функцией различных отделов центральной нервной системы позволяет сформулировать
несколько общих положений, касающихся принципов организации ее работы.
Одно из них состоит в том, что в
основе деятельности нервной системы лежит универсальный физиологический процесс
распространяющегося возбуждения — нервный импульс. Нервный импульс,
распространяясь по поверхности нервной клетки и ее отростков, выполняет в
нервной системе функцию носителя информации. Передача информации осуществляется
с помощью изменения либо последовательности нервных импульсов во времени, либо
распределения между различными клетками. Это так называемое пространственно—.
временное кодирование.
Ведущей формой нервной деятельности
являются рефлексы — закономерные реакции, возникающие в ответ на
раздражение рецепторов изменениями в окружающей или внутренней средах
организма. Рефлекторная деятельность представляет собой сложный комплекс
взаимодействующих и взаимовлияющих рефлексов. В основе рефлекторной
деятельности лежит ряд закономерностей. Это зависимость характера реакции от
типа раздражаемых рецепторов и силы действующего раздражителя, иррадиация
возбуждения, обусловленная нефункционирующими межнейронными связями ЦНС,
которые дают возможность активироваться одновременно большому числу нервных
связей.
Одна из основных закономерностей
заключается в том, что характер рефлекторной реакции способен изменяться в
зависимости от состояния тех нервных структур, через которые данная реакция
реализуется. При этом изменение возбудимости нервных структур может изменить не
только количественную, но и качественную направленность реакции. Примером тому
может служить открытое А. А. Ухтомским состояние доминанты, т. е.
появление очага возбуждения (в частности, под действием гормонов), которое приводит
к тому что стимуляция различных рецептивных полей вызывает возникновение или
усиление рефлекторной деятельности нервных структур именно этого очага.
Разные типы рефлексов осуществляются
с помощью одних и тех же мотонейронов, с использованием одних и тех же
эффекторов, т. е. мотонейроны представляют собой в этом случае общий
конечный двигательный путь. При этом правилом взаимодействия рефлексов
является у однонаправленных рефлексов взаимное усиление, у
противоположных рефлексов — взаимное торможение.
Общие закономерности тормозного
взаимодействия аналогичны закономерностям взаимного усиления. Облегчающие и
тормозные взаимные влияния между нервными центрами в процессе сочетанной
рефлекторной деятельности описаны Ч. Шеррингтоном на примере спинного мозга в
виде положительной и отрицательной одновременной индукции.
При сочетанной рефлекторной
деятельности помимо пространственного взаимодействия в нервных центрах
возникают еще и сложные явления, разворачивающиеся во времени. Еще в прошлом
столетии И. М. Сеченовым было установлено, что прекращение торможения
рефлекторной реакции сменяется так называемой отдачей — последующим
усилением по сравнению с исходной величиной. Явление отдачи рассматривают
сейчас как результат дополнительного включения более сложных центральных и
периферических путей.
Каждый рефлекторный акт всегда
является компонентом комплексной системы нервных процессов, направленных на
более эффективное выполнение определенной функции организма. Это объединение не
бывает жестко фиксированным и представляет собой динамическое взаимодействие.
Объединение относительно простых
рефлекторных механизмов и процессов в комплексы (функциональные системы),
способные уже оценивать результат рефлекторной деятельности и таким образом
регулировать эту деятельность, было изучено П. К. Анохиным. Любая часть нервной
системы является многоуровневой и построена по иерархическому принципу.
Одиночный нейрон, нейрональная цепь, нервный центр представляют собой
соподчиненные уровни системной организации, обладающей определенной
направленностью действия. Этот системный принцип встречается везде
независимо от того, идет ли речь о регуляции висцеральных функций или
организации комплексных систем корковых модулей. Системный подход наиболее
перспективен при исследовании общих аспектов поведения, таких как
взаимодействие сознания и подсознания, эмоций, научения и памяти.
Следующее положение касается
универсальности некоторых общих принципов организации нервной системы. В
качестве примера можно привести принцип доминанты, одинаково приемлемый для
спинного мозга и для колонок сенсомоторной коры. Доминанта как общий принцип
работы нервных центров определяет биологическую направленность поведения, без
учета которой глубинный анализ механизмов интегративной деятельности мозга
практически невозможен.
Сравнительно—физиологические
исследования нейронных сетей у позвоночных и беспозвоночных животных показали
принципиальное сходство их организации. Такие физиологические феномены, как суммация,
синоптическая потенциация, реципрокное торможение и ряд других имеют место
и в простых нервных системах беспозвоночных, и в ЦНС высших позвоночных.
Не меньшее число аналогий дает и
клеточный уровень организации. Достаточно напомнить, что все основные постулаты
мембранной теории разработаны на гигантских аксонах кальмара, затем повторены
на нейронах позвоночных. Сходство морфофункциональной организации нейронов и
нейронных сетей различных животных свидетельствует о возможно большем
консерватизме в области филогенетического преобразования молекулярных и клеточных
структур по сравнению с системным уровнем организации, где мы встречаемся с
явлениями параллелизма и специфичности.
Регулирующая и управляющая деятельность характеризует все
отделы нервной системы. Она распространяется на все области тела, его отделы,
органы, ткани. Относительно простыми формами такой деятельности характеризуются
структуры спинного мозга, первично управляющие соматическими и
висцеральными реакциями. Сигналы для соматических моторных реакций
(собственные, защитные, ритмические, позные и другие рефлексы мышц) формируются
спинальными мотонейронами, для висцеральных — симпатическими и
парасимпатическими преганглионарными нейронами, локализующимися в грудном и
крестцовом отделах спинного мозга. Из висцеральных реакций наиболее изучены механизмы,
обеспечивающие регуляцию уровня кровяного давления за счет изменения просвета
сосудов. По такому функциональному признаку в спинном мозгу разделяются лишь
эфферентные нейроны. Что касается чувствительных и особенно вставочных, то они
способны активизировать и соматические двигательные нейроны, и висцеральные,
управляющие сосудодвигательной функцией, регуляцией сокращений мочевого пузыря
и др.
Значительно сложнее деятельности спинного мозга рефлекторная
деятельность продолговатого мозга и мозжечка (ромбовидный мозг). Их нервные
образования (ретикулярная формация ствола, подкорковые ядра, кора мозжечка) не
получают сенсорных сигналов непосредственно от рецепторных структур и не отдают
также эфферентных сигналов непосредственно исполнительным органам. Они связаны
только с другими центрами мозга и являются, строго говоря, надстройкой над
первичной сегментарной структурой ЦНС. Осуществляемые ядрами расположенных
здесь черепных нервов рефлекторные реакции, как и в спинном мозгу, по характеру
конечного эффекта разделяются на соматические и висцеральные.
Первые направлены главным образом на поддержание позы, вторые — на обеспечение
последовательности восприятия, жевания, глотания и продвижения пищи по
пищеварительному каналу.
Ряд важнейших висцеральных функций связан непосредственно с ретикулярной
формацией продолговатого мозга. В ней имеются комплексы клеток, ритмичность
деполяризации мембран которых у одной части соответствует фазе вдоха, у другой
— фазе выдоха («вдыхательные» и «выдыхательные» нейроны). На деятельность этих
групп нейронов специфическое действие оказывает парциальное давление CO2 в крови. Чем оно выше, тем чаще
ритм, и наоборот. Эта автоматическая деятельность может изменять свою
активность под влиянием импульсов из различных рецептивных зон, а также в
определенной мере произвольным волевым усилием.
Кроме того, в продолговатом мозгу находятся и комплексы
нейронов, связанных с регуляцией сосудистого тонуса: одни из них учащают
свою деятельность при повышении артериального давления, другие — при его
понижении («сосудосуживающие» и «сосудорасширяющие» нейроны). Нисходящие пути,
образованные такими клетками, направляются в спинной мозг, где синаптически
контактируют с преганглионарными симпатическими нейронами, обладающими
сосудодвигательными функциями. Подобно дыхательным нейронам, клетки
сосудодвигательного центра обладают высокой чувствительностью к изменениям
химического состава крови.
Функции сосудодвигательного центра тесно связаны с
деятельностью группы нейронов, регулирующих работу сердца. Эти нейроны
находятся в постоянной тонической активности, которая модулируется влиянием со
стороны сосудодвигательного центра. Возбуждение клеток сердечной, области через
соответствующие ганглионарные нейроны (главным образом метасимпатической
природы) вызывает торможение деятельности сердца.
Нейронные механизмы среднего мозга обслуживают в
основном дистантные функции организма — зрение, слух, а также регулируют
определенные двигательные процессы. Нейроны последней специализации находятся
под непосредственным контролем и влиянием коры больших полушарий и подкорковых
ядер. Они образуют систему экстрапирамидных двигательных путей.
Рефлекторная деятельность промежуточного мозга,
который включает большое число различных структур, локализующихся вокруг III желудочка (главными среди них
являются таламус и гипоталамус), исключительно сложна. Все
сенсорные системы, за исключением обонятельной, проходят через ядра таламуса
перед вступлением в кору. Помимо проекционных нейронов ядер таламуса, в нем
имеются клетки, аксоны которых следуют в другие подкорковые структуры.
Комплексы таких нейронов обозначают как неспецифические ядра таламуса.
Посредством двусторонних связей с другими подкорковыми структурами таламус
регулирует взаимодействие ряда сложнорефлекторных функций, требующих интеграции
соматических и вегетативных реакций.
В этом отношении исключительно важное значение принадлежит гипоталамусу.
За счет связей со стволовыми структурами мозга и ведущей эндокринной железой — гипофизом
— гипоталамус выполняет функцию высшего центра интеграции висцеральных
функций, которые далее регулируются симпатическими, парасимпатическими и
метасимпатическими частями автономной нервной системы, а также эндокринными
влияниями. В этом случае центральные процессы, направляющие деятельность организма
на поиск удовлетворения или избегания неблагоприятных ситуаций, обозначают
понятием мотивация.
Самой новой в эволюционном отношении структурой нервной
системы является конечный мозг. Все поверхности его полушарий покрыты
корой большого мозга, образованной серым веществом. Вместе с другими
структурами нервной системы кора участвует в регуляции и координации функций
организма, играет исключительную роль в психической или высшей нервной
деятельности.
Высшая нервная деятельность представляет собой интегративную
деятельность высших отделов мозга, обеспечивающую индивидуальное поведенческое
приспособление человека и высших животных к изменяющимся условиям внутренней и
окружающей среды. При введении этого понятия И. П. Павлов определил его отличие
от низшей нервной деятельности. Последняя объединяет совокупность безусловных
рефлексов, обеспечивающих координированную деятельность организма, направленную
на поддержание постоянства внутренней среды, например уровня кровяного,
осмотического, онкотического давления.
Следовательно, высшая нервная деятельность представляет
собой аналитико—синтетическую деятельность коры и ближайших подкорковых
образований головного мозга. Она проявляется в способности выделять из
окружающей среды ее отдельные элементы и объединять их в комбинации, которые
точно соответствуют биологической значимости явлений окружающего мира.
Главная структурная особенность коры — огромное число (более 10 млрд.)
нейронов. Клетки всех слоев, которые расположены перпендикулярно по отношению к
поверхности коры и тесно связаны между собой и подкорковыми образованиями,
образуют комплекс, называемый колонкой клеток. Каждая подобная колонка
отвечает за восприятие по преимуществу одного вида чувствительности. В коре
выделяют сенсорные зоны, к клеткам которых проецируются основные
сенсорные системы (вкусовая, слуховая, зрительная и др.), а также моторные
зоны, которые посылают команды по пирамидным и экстрапирамидным путям в
ствол мозга и в спинной мозг. Клетки этих зон также посылают сигналы к
эффекторным нервным клеткам.
Одним из узловых вопросов физиологии высшей нервной
деятельности является научение. Сейчас принято различать ряд его форм:
1) облигатное, стимулзависимое (суммация, привыкание, импринтинг, подражание);
2) факультативное, эффектзависимое (условные рефлексы, классические и
инструментальные); 3) когнитивное (основанное на образной памяти, элементарной
рассудочной деятельности, вероятностном прогнозировании).
Важнейшими принципами интегративной деятельности мозга
являются принцип условного рефлекса (И. П. Павлов) и принцип
доминанты (А. А. Ухтомский),
сочетание которых обеспечивает активно—творческий и высокоадаптивный характер
целостного поведения. В качестве главного субстрата управления интегративной
деятельностью мозга сейчас рассматривают высшие ассоциативные системы; что
касается психики человека, то биологические предпосылки ее формирования зависят
от трех видов потребностей — витальных, социальных и идеальных.
Человеческий мозг характеризуется функциональной и
межполушарной асимметрией: правое полушарие работает по принципу
дедукции, т. е. сначала осуществляет синтез, а затем анализ, левое — по
принципу индукции, т. е. сначала анализирует, а затем синтезирует информацию. В
формировании речевой деятельности и мышления человека особую роль играют
активно развивающиеся в онтогенезе ассоциативные системы больших полушарий
мозга.
Барьер между кровью (гемолимфой) и ЦНС независимо возник не
менее чем в трех ветвях эволюции: у позвоночных, головоногих моллюсков и высших
членистоногих в связи с усложнением функций мозга и возросшей потребностью в
чрезвычайно строгом поддержании постоянства его внутренней среды.
У позвоночных основу гематоэнцефалического барьера
(ГЭБ) составляют эндотелиоциты капилляров мозга, соединенные между собой особого
рода контактами. Важную роль играют также вещества, секретируемые отростками
астроцитов, и энзимы, находящиеся в тканях между кровью и нейронами мозга.
Способность веществ проникать сквозь ГЭБ определяется в
первую очередь их растворимостью в жирах; для некоторых других веществ,
необходимых мозгу (например, для глюкозы и аминокислот), в барьере существуют
системы переносчиков.
На границе между кровью и мозгом есть небольшие по площади,
но очень важные участки, в которых функцию барьера выполняет не эндотелий
капилляров, а эпендима желудочков мозга. Существуют также пути «обхода»
барьера, самым известным (и, тем не менее, малоизученным) у человека является
путь, начинающийся на слизистой оболочке полости носа.
ГЭБ не только обеспечивает поддержание особого гомеостаза
мозга, но и является сложной коммуникационной системой, связывающей мозг и
периферические органы. Эти связи могут осуществляться и без проникновения
каких—либо веществ сквозь барьер (например, гормоны могут связываться с
рецепторами на внешней стороне барьера). Проникновение сквозь барьер
осуществляется в обе стороны (из крови в мозг и из мозга в кровь). Транспортные
системы, выносящие вещества из мозга, играют важную роль в поддержании его
гомеостаза. Средства, которые применяют для повышения проницаемости ГЭБ (резкое
повышение напряжения CO2 или осмотического давления плазмы
крови и т. п.), нельзя считать физиологическими — они приводят не к вскрытию,
но скорее к взлому барьера.
Айрапетянц М. Г., Вейн А. М. Неврозы в эксперименте и в клинике.
М., 1982.
Альтман Я. А. Психофизиологический анализ
поэтического вдохновения. СПб., 1994.
Анохин П. К. Биология и нейрофизиология
условного рефлекса. М., 1968.
Батуев А. С. Высшая нервная деятельность. М.,
1991.
Батуев А. С. Высшие интегративные системы мозга.
Л., 1981.
Бериташвили И. С. Избранные труды: Нейрофизиология и
нейропсихология / И. С. Бериташвили. Л., 1975.
Бернштейн Н. А. Очерки по физиологии движений и
физиологии активности. М., 1966.
Бехтерева Н. П. Здоровый и больной мозг человека.
Л., 1988.
Бехтерева Н. П. Магия мозга и лабиринты жизни.
СПб., 1999.
Бехтерева Н. П. О мозге человека: Размышления о
главном. СПб., 1994.
Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение. М., 1988.
Брёже М. Электрическая активность нервной
системы. М., 1979.
Буреш Я., Бурешова О., Хьюстон Дж. Методики и основные эксперименты по
изучению мозга и поведения. М., 1991.
Вартанян Г. А., Пирогов А. А. Эмоциональный анализатор мозга.
СПб., 1994.
Вейн А. М., Хехт К. Сон человека: Физиология и
патология. М., Берлин, 1989.
Гельгорн Э. Луфборроу Дж. Эмоции и эмоциональные
расстройства. Нейрофизиологическое исследование. М., 1966.
Громова Е.А. Эмоциональная память и ее
механизмы. М., 1980.
Дельгадо X. Мозг и сознание. М., 1971.
Дьюсбери Д. Поведение животных: Сравнительные
аспекты. М., 1981
Карамян А. И. Эволюция конечного мозга
позвоночных. Л., 1976.
Кассиль В. Г. Пищевое поведение в онтогенезе. Л.,
1990.
Кассиль Г. Н. Наука о боли. М., 1975.
Ковбаса С. И., Ноздрачев А. Д. Информационные характеристики
систем нейронов. Л., 1990.
Ковбаса С. И., Ноздрачев А. Д.,
Ягодин С. В. Анализ
взаимосвязи нейронов. Л., 1984.
Костюк П. Г. Физиология центральной нервной
системы. Киев, 1977.
Котляр Б. И., Шульговский В. В. Физиология центральной нервной
системы. М., 1979.
Кругликов Р. И. Нейрохимические механизмы обучения
и памяти. М., 1981.
Крушинский Л. В. Биологические основы рассудочной
деятельности. М., 1986.
Куффлер С., Николе Дж. От нейрона к мозгу. М., 1979.
Кэндел Э. Клеточные основы поведения. М.,
1980.
Лапицкий В. П. Головные ганглии и двигательная
активность насекомых. Л., 1990.
Мак—Фарленд Д. Поведение животных: Психобиология,
этология и эволюция. М., 1988.
Милейковский Б. Ю., Ноздрачев А. Д. Торможение двигательной активности.
Стволовые механизмы. СПб., 1998.
Ноздрачев А. Д., Поляков Е. Л. Анатомия кошки. СПб., 1998.
Ноздрачев А. Д., Поляков Е. Л. Анатомия лягушки. М., 1994.
Ноздрачев А. Д., Поляков Е. Л„
Гнетов А. В.
Исследование функций головного мозга: некоторые современные методы. Л., 1987.
Ноздрачев А. Д., Поляков Е. Л.,
Лапицкий В. П., Осипов Б. С., Фомичев Н. И. Анатомия беспозвоночных: пиявка, прудовик,
дрозофила, таракан, рак.
Ноздрачев А. Д., Чумасов Е. Н. Периферическая нервная система.
СПб., 1999.
Ноздрачев А. Д.. Чернышева М. П. Висцеральные рефлексы. Л., 1989.
Поляков Г. И. Проблема происхождения рефлекторных
механизмов мозга. М., 1964.
Раус С. Устройство памяти. От молекул к
сознанию. М., 1995.
Серков Ф. Н. Корковое торможение. Киев, 1986.
Симонов П. В. Мотивированный мозг: высшая нервная
деятельность и естественнонаучные основы общей психологии. М.,1987.
Симонов П. В. Эмоциональный мозг. М., 1981.
Слоним А. Д. Инстинкт: загадки врожденного
поведения организмов. Л., 1967.
Соколов Е. Н. Нейронные механизмы памяти и обучения.
М., 1981.
Сравнительная физиология животных. В
3 т. / Под ред. Л. Проссера. М., 1977.
Ухтомский А. А. Доминанта. М.—Л.,
1966.
Физиология поведения.
Нейробиологические закономерности / В. П. Бабминдра и др. Л., 1987.
Физиология поведения. Нейрофизиологические
закономерности / А. С. Батуев, А. Д. Солоним, П. В. Симонов и др. Л., 1986.
Функциональные системы организма / Под ред. К. В. Судакова.
М., 1987.
Хайнд Р. Поведение животных. Синтез этологии
и сравнительной психологии. М., 1975.
Хьюбел Д. Глаз, мозг, зрение. М., 1990.
Черниговский В. Н. Интероцепция: Сборник. Л., 1985.
Шаде Д., Форд Д. Основы неврологии. М., 1976.
Шаповалов А: И. Механизмы синаптической передачи.
Избранные труды. СПб., 1997.
Шеперд Г. Нейробиология. В 2 т. М., 1987.
Эволюционная физиология. В 2—х ч. /
Отв. ред. Е. М. Крепе. Л., 1983. Ч. 2. Руководство по физиологии.