|
НЕРВНАЯ
СИСТЕМА
Для слаженной деятельности
различных частей такой сложной системы, как организм человека, необходимо
координирующее устройство соответственной сложности. И в самом деле,
нервная система, интегрирующая деятельность всех частей тела, является,
несомненно, самой сложной из всех систем органов. Мышцы и железы животного
или человека носят общее название эффекторов; глаза, уши и другие органы
чувств называются рецепторами. Нервная система, состоящая из головного
мозга, спинного мозга и проводящих путей, соединяет рецепторы с
эффекторами и передает импульсы, или "сообщения", от первых ко вторым. Она
способна делать это таким образом, что при раздражении того или иного
рецептора должным образом реагирует надлежащий эффектор. Основными
функциями нервной системы являются проведение импульсов и интеграция
деятельности различных систем организма. Координирующие функции нервной
системы, эндокринная регуляция и собственные регуляторные механизмы
внутриклеточных ферментных систем (торможение и стимуляция активности
ферментов, индукция и репрессия их синтеза) - все это факторы,
способствующие гомеостазу, т. е. поддержанию постоянства внутренней среды
организма.
НЕЙРОНЫ
Хотя взаимоотношения
нейронов - клеток, составляющих нервную систему,- чрезвычайно
сложны, все эти клетки имеют общий основной план строения: они состоят из
тела клетки, содержащего ядро, и отростков - одного аксона и одного или
нескольких дендритов (рис. 25). Нейроны сильно различаются между
собой по форме клеточного тела и по длине, числу и степени ветвления
аксонов и дендритов. Нейроны подразделяются на чувствительные
(сенсорные), двигательные (моторные) и вставочные. У
чувствительных нейронов дендриты соединены с рецепторами, а аксоны - с
другими нейронами; у двигательных нейронов дендриты соединены с другими
нейронами, а аксоны - с каким-нибудь эффектором; у вставочных нейронов и
дендриты и аксоны соединяются с другими нейронами. Самый простой путь, по
которому может идти нервный импульс, состоит из трех нейронов: одного
сенсорного, одного вставочного и одного моторного.
Нервные стволы, или нервы,
состоят из большого числа аксонов и дендритов, объединенных в общей
соединительнотканной оболочке. Тела нейронов не разбросаны беспорядочно, а
образуют скопления, называемые ганглиями, если они расположены вне
головного и спинного мозга, и нервными центрами, если они находятся
в головном или спинном мозгу.
Аксон или
дендрит покрыты, помимо клеточной мембраны, еще одной или двумя
оболочками: наружной неврилеммой и внутренней миэлиновой
оболочкой (рис.
263). Неврилемма состоит из клеток. Миэлиновая оболочка состоит из
неклеточного жироподобного материала, благодаря которому покрытые ею нервы
кажутся белыми. Нервные волокна в спинном и головном мозгу имеют только
миэлиновую оболочку. Волокна, идущие к внутренним органам, покрыты
неврилеммой и тонкой миэлиновой оболочкой, так что они имеют скорее серую,
чем белую, окраску. Нервы, идущие к коже и скелетным мышцам, обладают
обеими оболочками. Б. Азмэн показала, что миэлиновая оболочка строится из
складок, образуемых оболочками клеток неврилеммы (рис. 264).
Положение миэлиновой оболочки заставляет
полагать, что она играет роль изолятора, т. е. препятствует переходу
нервного импульса с одного волокна на другое, что привело бы к возбуждению
несоответствующего эффектора; однако никаких данных, подтверждающих это
предположение, нет. Можно было бы предположить, что миэлиновая оболочка
служит резервом питательных веществ для нервного волокна, но имеющиеся
данные указывают на то, что волокно получает питание только из тела
клетки. Высказывалось также предположение, что миэлиновая оболочка
увеличивает скорость проведения нервных импульсов. У человека и у других
млекопитающих нервный импульс распространяется по миэлиновому волокну со
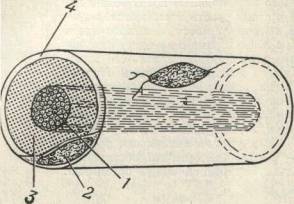
|
Рис. 263 Типичное нервное волокно и окружающие
его оболочки.
1 - нервное волокно;
2 - ядро клетки неврилеммы;
3 - миэлиновая оболочка;
4 - неврилемма.
|
скоростью около 100м/сек, а по
нервам, идущим к внутренним органам (эти нервы имеют очень тонкую
миэлиновую оболочку),- со скоростью 7-15 м/сек. Однако нет никаких
доказательств .того, что большая скорость проведения в первом случае
обусловлена миэлиновой оболочкой, а не каким-то другим свойством нерва.
Неврилемма, по-видимому, играет некоторую роль
в регенерации перерезанного нервного волокна (аксона или дендрита). В
случае разрыва волокон те их части, которые утратили связь с телом
нейрона, дегенерируют и исчезают в течение нескольких недель, оставляя
после себя пустые трубки неврилеммы. Если концы нервного ствола в месте
его перерезки соединить друг с другом при помощи скобы или шва, то нервные
волокна могут врасти из перерезанных концов нерва в неврилеммные трубки
дегенерировавших волокон и достигнуть органов, которые иннервировались
прежними волокнами. Таким путем могут восстановиться чувствительность и
способность управлять мышцами, утраченные вследствие перерезки нерва.
Время, необходимое для регенерации, зависит от того, насколько далеко
нерву приходится расти; иногда на это может потребоваться целых два года.
При рассечении ткани головного или спинного мозга, волокна которой лишены
неврилеммы, регенерации не происходит. Какова роль неврилеммы - направляет
ли она только рост регенерирующих волокон, способствует ли их питанию или
образует миэлиновую оболочку,- неизвестно.
НЕРВНЫЙ ИМПУЛЬС
Изучение природы нервного
импульса было связано с особыми трудностями, так как при прохождении
импульса по нерву никаких видимых изменений не происходит. Лишь недавно, с
развитием микрохимических методов, удалось показать, что во время
проведения импульса нерв расходует больше энергии, потребляет больше
кислорода п выделяет больше углекислоты, чем в состоянии покоя. Это
указывает на то, что в проведении импульса, в восстановлении исходного
состояния после проведения или в обоих этих процессах участвуют
окислительные реакции. Когда примерно 100 лет назад было установлено, что
нервный импульс сопровождается определенными электрическими явлениями,
возникло мнение, что сам импульс представляет собой электрический ток. В
то время было известно, что электрический ток распространяется очень
быстро, п поэтому высказывалось мнение, что скорость распространения
нервного импульса слишком велика, чтобы ее можно было измерить. Десять лет
спустя Гельмгольц измерил скорость проведения импульса, раздражая нерв,
идущий к мышце, на различных расстояниях от мышцы и измеряя время,
протекавшее между раздражением и сокращением. Таким способом он показал,
что нервный импульс распространяется гораздо медленнее электрического - в
нервах лягушки со скоростью около 30 м/сек. Это, конечно,
свидетельствовало о том, что нервный импульс не есть электрический ток,
подобный току в медном проводе. Кроме того, мертвый или раздавленный нерв
все еще проводит ток, но не проводит нервных импульсов, и, раздражаем ли
мы нерв током, прикосновением, приложением тепла или химическими
факторами, возникающий при этом импульс распространяется со скоростью
одного
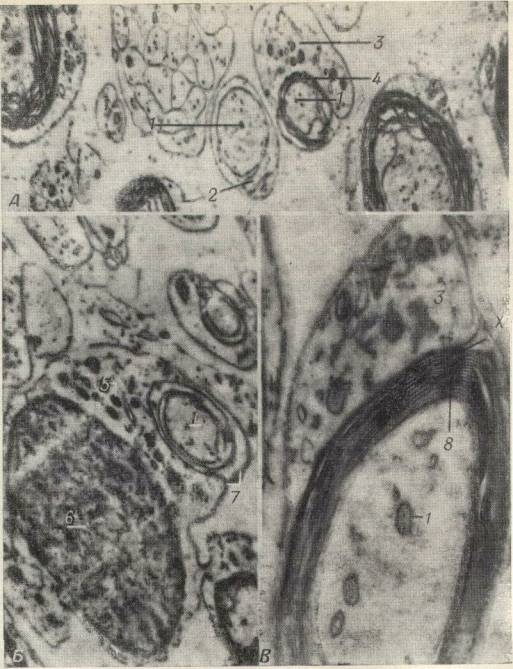
|
|
Рис. 264. электронные микрофотографии волокон
седалищного нерва мышонка в возрасте 7 дней, показывающие развитие
миэлиновой оболочки путем образования складок оболочками клеток
неврилеммы.
Д.Слева - ранняя стадия, на которой аксон окружен
клеткой неврилеммы; видна лишь небольшая часть этой
клетки.
Справа - несколько более поздняя стадия; виден большой
участок клетки неврилеммы, и слой, образованный складками оболочки
этой клетки, сделался толще (X 26 000).
Б. Большое темное
овальное образование представляет собой ядро клетки неврилеммы;
между ядром и аксоном видны митохондрии; можно видеть спирально
закрученные складки клеточной оболочки (X 28 000).
В. Более
поздняя стадия - аксон окружен толстой, плотной, многослойной
миэлиновой оболочкой. На участке, обозначенном буквой X, видно, что
слой миэлина переходит в оболочку клетки неврилеммы (X 83 000).
1
- аксон; 2 - складка оболочки клетки неврилеммы; 3 - клетка
неврилеммы; 4 - свернутые складки оболочки клетки неврилеммы; 5 -
митохондрии; 6 - ядро; 7 - спирально свернутая оболочка клетки
неврилеммы; 8 - миалин. |
и того
же порядка. Из этого мы заключаем, что нервный импульс представляет собой
не электрический ток, а электрохимическое возмущение в нервном волокне.
Вызванное раздражителем возмущение в одном участке нервного волокна
вызывает такое же возмущение в соседнем участке и так далее до тех пор,
пока импульс не дойдет до конца волокна. Таким образом, передача импульса
подобна горению бикфордова шнура: от теплоты, выделяющейся при горении
одного участка шнура, загорается следующий участок и т. д. В нерве роль
теплоты выполняют электрические явления, которые, возникнув в одном
участке, стимулируют следующий.
Передача
нервного импульса сходна с горением бикфордова шнура и в некоторых других
отношениях. Скорость горения шнура не зависит от количества тепла,
затраченного при его зажигании, если только этого тепла достаточно, чтобы
шнур загорелся. Не имеет значения и метод зажигания. Так же обстоит дело и
с нервом. Нерв не будет реагировать, пока к нему не будет приложено
раздражение определенной минимальной силы, но дальнейшее увеличение силы
раздражения не заставит импульс распространяться быстрее. Это обусловлено
тем, что энергию для проведения импульса доставляет сам нерв, а не
раздражитель. Описанное явление отражено в законе "все или ничего":
нервный импульс не зависит от природы и силы вызвавшего его раздражителя,
если только раздражитель обладает достаточной силой, чтобы вызвать
появление импульса. Хотя скорость проведения не зависит от силы
раздражителя, она зависит от состояния нервного волокна, и различные
вещества могут замедлять передачу импульса или делать ее невозможной.
Сгоревший шнур нельзя использовать вторично,
нервное же волокно способно восстанавливать свое исходное состояние и
передавать другие импульсы. Оно, однако, не может проводить их непрерывно:
после проведения одного импульса проходит определенное время, прежде чем
волокно сможет передавать второй импульс. Этот промежуток времени,
называемый рефракторным периодом, продолжается от 0,0005 до 0,002 сек. В
это время происходят химические и физические изменения, в результате
которых волокно возвращается в первоначальное состояние.
Насколько нам известно, импульсы, передаваемые
нейронами всех типов - двигательными, чувствительными или вставочными, в
основном сходны между собой. То, что один импульс вызывает ощущение света,
другой - ощущение звука, третий-мышечное сокращение, а четвертый
стимулирует секреторную деятельность железы, всецело зависит от природы
тех структур, к которым приходят импульсы, а не от каких-либо особенностей
самих импульсов.
Хотя нервное волокно можно
стимулировать в любой его точке, в нормальных условиях возбуждение
вызывается только на одном его конце, от которого импульс идет вдоль
волокна до его другого конца1. Соединение между последовательными
нейронами называется синапсом. Нервный импульс передается с кончика аксона
одного нейрона на дендрит следующего через синаптическое соединение путем
выделения у кончика аксона определенного вещества. Это вещество вызывает
появление нервного импульса в дендрите следующего аксона. Передача
возбуждения через синапс происходит значительно медленнее, чем передача
его по нерву. В нормальных условиях импульсы проходят только в одном
направлении: в чувствительных нейронах они идут от органов чувств к
спинному и головному мозгу, а в двигательных - от головного и спинного
мозга к мышцам и железам. Направление определяется синапсом, так как
только кончик аксона способен выделять вещество, стимулирующее другой
нейрон. Каждое отдельное нервное волокно может проводить импульс в обоих
направлениях; при электрическом раздражении волокна где-либо в середине
возникают два импульса, один из которых идет в одном направлении, а другой
- в другом (эти импульсы можно обнаружить соответствующими электрическими
приборами), но лишь тот из них. который идет по направлению к кончику
аксона, может стимулировать следующий нейрон в цепи. Импульс, идущий к
дендриту, "остановится", достигнув его конца.
Химические и электрические процессы, с
которыми связана передача нервного импульса, во многом сходны с
процессами, происходящими при мышечном сокращении. Но проводящий импульсы
нерв расходует очень мало энергии по сравнению с сокращающейся мышцей;
теплота, образующаяся при раздражении нерва
1 Единственный нерв, раздражение которого
иногда происходит не на конце, а в некоторой промежуточной точке его
длины,- это локтевой нерв, лежащий в области локтя близко к поверхности
кожи. Этот нерв состоит из волокон, проводящих импульсы при ощущениях
боли, прикосновения, холода, давления и т. д. Вот почему при ушибе локтя
возникает неопределенное, "смешанное" ощущение.
в течение 1 мин, в расчете на 1 г ткани эквивалентна
энергии, выделяющейся при окислении 0,000001 г гликогена. Это означает,
что если бы нерв содержал в качестве источника энергии лишь 1% гликогена,
его можно было бы стимулировать непрерывно в течение недели и запас
гликогена не был бы исчерпан. При достаточном снабжении кислородом нервные
волокна практически неутомляемы. Какова бы ни была природа "умственного
утомления", это не может быть настоящим утомлением нервных волокон.
МЕМБРАННАЯ ТЕОРИЯ ПРОВЕДЕНИЯ
ВОЗБУЖДЕНИЯ
Наши современные знания о природе
нервного импульса получены главным образом в результате опытов с очень
крупными аксонами, имеющимися у кальмара, у речного рака и у некоторых
червей. Толщина гигантского аксона у кальмара (около 1 мм) позволяет
вводить микроэлектроды и микропипетки непосредственно в вещество нервного
волокна и измерять электрический потенциал нервной мембраны. Согласно
современной мембранной теории проведения возбуждения, электрические
явления в нервном волокне обусловлены различной проницаемостью нервной
мембраны для ионов натрия и калия, а эта проницаемость в свою очередь
регулируется разностью электрических потенциалов по обе стороны от нее.
Для возбуждения нервного волокна требуется определенное критическое
пороговое изменение. Возбуждение представляет собой освобождение
электрической энергии из нервной мембраны и распространяется вдоль волокна
в виде короткого электрического импульса типа "все пли ничего",
называемого потенциалом действия.
Для того
чтобы понять события, происходящие при прохождении нервного импульса по
волокну, мы должны сначала иметь ясное представление о нервном волокне в
состоянии покоя. Нервное волокно - это длинная цилиндрическая трубка,
поверхностная мембрана которой разделяет два раствора, имеющие различный
химический состав, но содержащие одинаковое количество ионов. В наружной
среде преобладают ионы натрия и хлора, а во внутренней - ионы калия и
различные органические анионы. Концентрация ионов натрия в наружном
растворе приблизительно в 10 раз выше, чем во внутреннем, а концентрация
ионов калия во внутреннем примерно в 30 раз выше, чем в наружном.
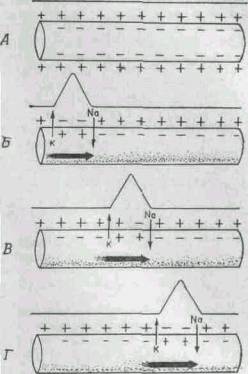
|
|
Рис. 265. Схема, поясняющая мембранную теорию проведения
нервного импульса.
А. Нерв в состоянии покоя; его мембрана
поляризована, наружная поверхность ее несет положительные заряды,
внутренняя - отрицательные.
Б - Г. Стадии прохождения импульса по
нерву; показана волна деполяризации и сопровождающий ее потенциал
действия, распространяющийся по мембране.
|
Эти
концентрации ионов Nа+ и К+ поддерживаются на относительно постоянном
уровне, но существует непрерывный поток ионов в клетку и из клетки.
Оболочка нервного волокна, подобно другим клеточным мембранам, активно
переносит одни ионы из внутренней среды в наружную, а другие - в обратном
направлении. В результате этого дифференциального распределения ионов по
обе стороны мембраны между ними существует разность потенциалов от 0,06 до
0,09 в (так называемый мембранный потенциал, или потенциал покоя}, причем
внутренняя поверхность заряжена отрицательно по отношению к наружной (рис. 265).
Эти различия в электрическом потенциале и
концентрации ионов поддерживаются в покоящемся (т. е. не проводящем
импульсов) нерве благодаря активному переносу ионов натрия из внутренней
среды в интерстициальную жидкость, окружающую нервное волокно. Этот
механизм получил название натриевого насоса. Для выведения ионов натрия из
клетки необходима энергия, которую доставляют процессы обмена,
происходящие в нерве. При отравлении нервного волокна (например, цианистым
калием) натриевый насос "выключается".
Выведение ионов натрия сопровождается
поглощением ионов калия, и хотя подробности этого процесса неизвестны,
повидимому, на клеточной поверхности происходит обмен катионами, в
результате которого на каждый выведенный ион натрия в клетку входит один
ион калия. Клеточная мембрана имеет относительно низкую ионную
проницаемость, так что даже если выключить натриевый насос (например, под
действием какого-либо метаболического яда), то пройдет немало часов,
прежде чем градиенты концентраций натрия и калия между наружной и
внутренней поверхностью мембраны исчезнут. Проницаемость мембраны для этих
двух ионов неодинакова; для калия она выше, чем для натрия. Когда нервное
волокно находится в нормальном покоящемся состоянии, то наблюдается
избыток положительно заряженных ионов на наружной поверхности мембраны (рис. 265). : .
Изучение "кабельных" свойств нервного волокна
показывает, что аксон едва ли представляет собой пассивный проводник,
поскольку происходящие при этом потери очень велики. При воздействии на
нервное волокно слабого сигнала, недостаточного для возбуждения
действующего в волокне релейного механизма, этот сигнал затухает в
нескольких миллиметрах от места его приложения. Нервный импульс не мог бы
распространяться на большие расстояния по нерву, если бы не было какого-то
процесса, усиливающего сигнал. Эту функцию выполняет процесс возбуждения,
который регенерирует и усиливает сигнал в каждой точке нервного волокна.
"Кабельные" свойства волокна обеспечивают распространение изменения
электрического потенциала по нервному волокну на короткое расстояние (хотя
это изменение быстро затухает) и тем самым - стимуляцию возбуждения в
соседних участках нерва.
При возбуждении
нервного волокна и возникновении нервного импульса происходит мгновенное
изменение проницаемости нервной мембраны, которое позволяет ионам натрия
проходить внутрь. Это приводит к деполяризации нервной мембраны: она
становится положительно заряженной изнутри и отрицательно заряженной
снаружи (возникает потенциал действия). Проницаемость нервной мембраны для
ионов натрия при обычном потенциале покоя очень низка, но по мере
уменьшения мембранного потенциала она возрастает. Ионы натрия могут теперь
просачиваться внутрь клетки в направлении концентрационного градиента. Это
ведет к дальнейшему уменьшению мембранного потенциала и к увеличению
проницаемости мембраны для натрия. Этот процесс прогрессивно нарастает (он
обладает способностью к самоусилению), что приводит к резкому подъему
потенциала действия (рис. 265). Когда
волокно изнутри становится заряженным положительно по отношению к наружной
поверхности, дальнейший реальный приток ионов натрия прекращается.
Разность потенциалов между наружной и внутренней поверхностями мембраны
может мгновенно достигнуть 0.04 в. причем теперь уже внутренняя
поверхность заряжена положительно по отношению к наружной. Вслед за этим
наступает период повышенной проницаемости мембраны для калия, к результате
чего ионы калия перемещаются в направлении градиента концентрации изнутри
клетки наружу.
Возврат мембранного потенциала
к исходному состоянию (отрицательный заряд внутри и положительный -
снаружи) происходит не под влиянием обратного движения ионов (ионы натрия,
вошедшие в клетку в период возрастания потенциала действия, не выходят из
нее), а под влиянием утечки наружу эквивалентных количеств ионов калия. Во
время прохождения импульса ионы натрия и ионы калия перемещаются по
соответствующим концентрационным градиентам. Однако количество участвующих
в этом ионов так мало, что при передаче одного импульса невозможно
обнаружить изменения в концентрации ионов натрия или калия в волокне.
Гигантский аксон кальмара может провести несколько сот тысяч импульсов,
даже если выключить его натриевый насос, прежде чем запасы
аккумулированных ионов будут исчерпаны.
После
прохождения каждого импульса наступает период невозбудимости - абсолютный
рефракторный период, в течение которого волокно не может передавать второй
импульс. Вследствие изменений в проницаемости, сопровождающих
деполяризацию нервной мембраны, нервное волокно не может реагировать на
второе раздражение. Способность к возбуждению появляется вновь после
восстановления нормальной проницаемости.
Таким
образом, нервный импульс - это волна деполяризации, проходящая вдоль
нервного волокна. Изменение мембранного потенциала в одном участке делает
соседний участок более проницаемым, и в результате волна деполяризации
распространяется по волокну. Полный цикл деполяризации и деполяризации
занимает всего несколько тысячных секунды.
ПЕРЕДАЧА В СИНАПСЕ
Механизм передачи нервного
импульса через синапс с одного нейрона на другой не вполне выяснен. Нет
никаких данных, которые бы указывали на непрерывность цитоплазмы между
последовательными нейронами. На электронных микрофотографиях видно, что
мембраны аксона и дендрита в синапсе сильно сближаются, но ни слияния, ни
непосредственного контакта между ними обнаружить нельзя. Промежуток между
ними составляет около 15 мили микрон , но это не исключает возможности
эффективной "кабельной" связи в синапсе.
Существуют две коренным образом различающиеся
теории синаптической передачи: электрическая и химическая. Теория
электрической передачи подразумевает, что, несмотря на кажущуюся
морфологическую обособленность двух нейронов, между ними должен
существовать эффективный местный контакт, позволяющий току проходить от
первого нейрона ко второму и возбуждать его. Это означало бы, что процессы
передачи по нервному волокну и через синапс в своей основе одинаковы.
Однако совершенно очевидно, что между ними существует какое-то различие,
потому что все синапсы проводят импульсы только в одном направлении, тогда
как нервные волокна обладают одинаковой проводимостью в обоих
направлениях; обычно же ввиду наличия синапсов импульсы в каждом данном
нервном волокне проходят только в одном направлении.
Согласно теории химической передачи, процесс,
происходящий в синапсах, в корне отличается от механизма передачи
возбуждения по волокну. Предполагается, что физическое разъединение
нервных волокон в синапсе препятствует "кабельной" передаче в месте
соединения и вместо нее вступает в действие химический медиатор. В кончике
аксона синтезируется специфическое вещество, которое освобождается под
действием приходящего нервного импульса. Оно диффундирует через
синаптическое пространство и связывается специальным хеморецептором на
поверхности дендрита соседней клетки. В результате соединения медиатора с
хеморецептором происходят изменения в мембране, вызывающие ее
деполяризацию и возникновение нового потенциала действия. Этот потенциал
распространяется по нейрону до следующего синапса, где в свою очередь
стимулирует выделение другого химического медиатора. Таким образом, между
потенциалом действия (волной деполяризации) в одном нейроне и потенциалом
действия в следующем нейроне имеется промежуточное звено - механизм,
включающий выделение специфического вещества одной клеткой и
взаимодействие этого вещества со специфическим хеморецептором другой
клетки.
Многие данные указывают на то, что
химическая передача в синапсе - общее явление, и некоторые исследователи
утверждали, что во всех синапсах действует химический механизм. Однако
опыты Фершпана и Поттера, проведенные в 1957 г.. показали, что передача
через гигантский синапс в брюшном ганглии речного рака имеет электрическую
основу. Контакт между двумя мембранами в этом особом синапсе действует как
выпрямитель и позволяет току легко проходить в одном направлении - от
аксона соединительного нейрона к дендриту двигательного нейрона. В этом
синапсе существует достаточная "кабельная" связь, которая может передавать
импульс в нормальном направлении.
Твердо
установлено, что передача в нервно-мышечном соединении, в точке, где
двигательный нерв соединяется с мышцей, происходит химическим путем. Под
действием импульса, приходящего по двигательному волокну, кончики этого
волокна выделяют особое вещество - ацетилхолин. Ацетилхолин обладает
сильным возбуждающим действием и вызывает местную деполяризацию мембраны
мышечного волокна, что ведет к возникновению импульсов, распространяющихся
по мембране, и вызывает мышечное сокращение. Яд кураре препятствует
передаче импульсов с нерва на мышцу через синапс этого типа; по-видимому,
он присоединяется к рецепторам ацетилхолина, лишая их возможности
нормально взаимодействовать с этим медиатором.
Существование нейрогуморальных веществ,
выделяемых кончиками нейронов, было открыто в опытах с сердцем - органом,
снабженным двумя группами нервов, из которых одна ускоряет, а другая
замедляет его сокращения. Если изолировать сердце лягушки и раздражать
идущий к нему блуждающий (тормозной) нерв, то оно перестает сокращаться.
Если затем промыть это сердце солевым раствором и промывную жидкость
ввести в другое сердце, то последнее тоже останавливается! Следовательно,
блуждающий нерв выделяет, очевидно, какое-то вещество, тормозящее
сердечные сокращения. Дальнейшее исследование показало, что это
сравнительно простое химическое соединение, называемое ацетилхолином.
Здесь химическая передача осуществляется с нерва на мышцу, но такой же
механизм действует по крайней мере в некоторых типах синапсов между двумя
нейронами. В аналогичных опытах показано, что симпатические
постганглионарные волокна, ускоряющие сокращение сердца, выделяют так
называемый симпатин, химическое вещество, тождественное или сходное с
гормоном адреналином. У вас может возникнуть вопрос: если кончик аксона
выделяет такое вещество, как ацетилхолин, почему это вещество не
стимулирует следующий дендрит или мышцу в течение длительного времени?
Гистохимическое изучение обнаружило в области
синапса высокую местную концентрацию весьма активного фермента -
ацетилхолин-астеразы, который гидролизует ацетилхолин и делает его
неактивным. Обнаружен также другой фермент, окисляющий симпатии.
Химическая передача в синапсе связана с двумя
процессами: 1) освобождение под действием нервного импульса специфического
вещества из места его хранения в кончике аксона в узкое пространство между
соседними нейронами и 2) процесс, путем которого специфическое вещество -
медиатор - присоединяется к специфическим рецепторам в дендрите и вызывает
изменение свойств его клеточной мембраны, приводящее к возникновению
нового импульса. Первый процесс - особый случай нейросекреции, второй -
особый случай хеморецепции, сходный с процессом, происходящим в органах
химического чувства, например в органах вкуса и обоняния. Передачу
химического медиатора от аксона к дендриту можно объяснить простой
диффузией. На таком малом расстоянии диффузия должна протекать достаточно
быстро, чтобы обеспечить скорость передачи, наблюдаемую в синапсе. Было
показано, что двигательные нервы освобождают ацетилхолин отдельными
"порциями", содержащими большое число молекул. Для функционирования
механизма выделения ацетилхолина необходимы ионы
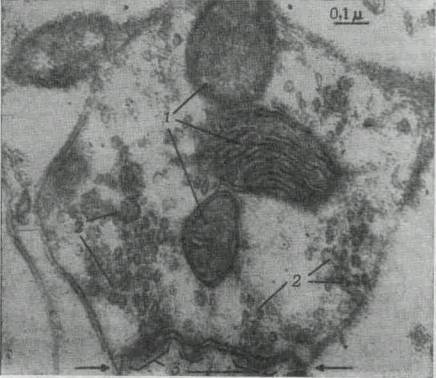
|
|
Рис. 266. Электронная микрофотография синаптического окончания
в возбужденной обонятельной луковице крысы.
Окончание содержит
три митохондрии (1) и несколько синаптических пузырьков (2). Зона
контакта между двумя нейронами показана двумя стрелками. Видно
утолщение мембран нервных волокон в зоне контакта (3}.
|
кальция, а ионы магния тормозят
его. Это позволяет предполагать, что медиатор хранится в нервных
окончаниях, в мельчайших внутриклеточных структурах, которые освобождают
все свое содержимое на поверхность клетки. На электронных микрофотографиях
кончиков нейрона (рис. 266) в синапсе видны скопления синаптических
пузырьков, которые, возможно, служат местом хранения ацетилхолина. Можно
представить себе, что каждый нервный импульс вызывает освобождение
содержимого одного из этих пузырьков в синаптическое пространство.
Еще не известно с достоверностью, как
происходит передача импульсов через синапсы в головном и спинном мозгу: с
помощью ли ацетилхолина, симпатина, какого-либо иного вещества или
электрического механизма.
Синаптическое
соединение оказывает известное сопротивление потоку импульсов в нервной
системе, и не каждый импульс, достигший синапса, передается на следующий
нейрон. Разные синапсы оказывают различное сопротивление, поэтому они
играют важную роль в определении пути импульсов через нервную систему и
реакции организма на тот или иной раздражитель.
В функциональном отношении вся нервная система
представляет собой единое целое, и импульс, возникший в любом рецепторе,
может быть передан на любой эффектор организма. Представим себе, что
человек обжег палец. Мышцы руки сокращаются, чтобы отвести палец от
горячего предмета, в головном мозгу возникает ощущение боли, человек может
вскрикнуть, возможна приостановка сокращений сердца, пищеварения и дыхания
- в сущности каждая мышца и железа может оказаться временно вовлеченной в
реакцию. Наши органы чувств получают непрерывный поток стимулов, но
избирательное сопротивление в синапсах препятствует постоянному
неконтролируемому сокращению мышц и выделению секретов железами.
Лекарственное вещество стрихнин снижает синаптическое сопротивление; при
отравлении стрихнином самый слабый раздражитель вызывает у человека
секрецию всех желез и судорожное сокращение всех мышц тела.
Величина синаптического сопротивления может
изменяться под влиянием нервных импульсов, так что один импульс может
свести на нет действие другого. Это называется торможением. Обратное
явление, когда один импульс усиливает другой, называется усилением. Эти
два процесса играют важнейшую роль в интеграции активности различных
органов и частей тела. Мы уже говорили, что все мышцы тела все время
находятся в состоянии слабого сокращения, называемого тонусом, вследствие
непрерывного потока нервных импульсов. Но когда одной мышце, например '
трехглавой мышце плеча, предстоит сократиться, ее антагонист, в данном
случае бицепс, должен быть расслаблен. Это достигается одновременным
торможением нервных импульсов, идущих к бицепсу, и усилением импульсов,
идущих к трехглавой мышце. Торможение и усиление могут происходить только
в синапсе, поскольку после того, как импульс начал свой путь по нейрону,
распространение его не может быть ни остановлено, ни ускорено. Сможет ли
данный импульс пройти через синапс, зависит от того, будет ли он
заторможен или усилен другими импульсами.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА. СПИННОЙ
МОЗГ
Нервная система человека состоит
примерно из 10 млрд. нейронов; они делятся на две основные категории:
нейроны, принадлежащие к центральной нервной системе, которые образуют
головной и спинной мозг, и нейроны периферической нервной системы,
образующие черепно-мозговые и спинномозговые нервы.
Спинной мозг представляет собой трубку,
окруженную и защищенную невральными дугами позвонков, и выполняет две
важные функции: передает импульсы, идущие в головной мозг и из головного
мозга, и служит рефлекторным центром. На поперечном разрезе можно видеть,
что он состоит из двух типов ткани: внутренней массы серого вещества,
имеющей в разрезе форму бабочки и состоящей из тел нервных клеток, и
лежащего снаружи белого вещества, образованного пучками аксонов и
дендритов (рис.
267). Белый цвет этих пучков обусловлен миэлиновыми оболочками нервных
волокон; концы аксонов и дендритов, находящиеся в центральном сером
веществе, не имеют миэлиновых оболочек. "Крылья" серого вещества разделены
на два задних и два передних рога. Передние рога содержат тела нейронов,
аксоны которых направляются в составе спинномозговых нервов к мышцам; все
остальные нервные клетки спинного мозга являются вставочными нейронами.
Аксоны и дендриты белого вещества разделены на
пучки со сходными функциями: восходящие пути, которые проводят импульсы к
головному мозгу, и нисходящие пути, которые проводят импульсы от головного
мозга к эффекторам. Тщательно регистрируя симптомы, наблюдаемые у людей с
поврежденным спинным мозгом, и сопоставляя эти данные с картиной
разрушения определенных путей, обнаруживаемой при исследованиях мозга
после смерти больного, неврологи смогли составить карту расположения и
функций различных путей (рис. 267). Например,
задние столбы белого вещества передают возникающие в рецепторах мьппц,
сухожилий и суставов импульсы,
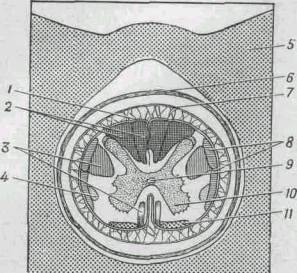
|
|
Рис. 267. Поперечный разрез спинного мозга, заключенного в
костную трубку из позвонков.
1 - мягкая мозговая оболочка; 2 -
проводящие пути для импульсов мышечного чувства; 3 - проводящие пути
для двигательных импульсов, идущих к мышцам; 4 - проводящие пути для
импульсов от рецепторов боли, тепла и холода; 5 - костное кольцо,
образованное невральной дугой позвонка; 6 - твердая мозговая
оболочка; 7 - паутинная оболочка; 8 - проводящие пути к мозжечку для
координации мышечной активности; 9 - центральный канал; 10 - серое
вещество; 11 - проводящие пути для импульсов от рецепторов
прикосновения и давления.
|
благодаря которым мы ощущаем
положение частей нашего тела. При далеко зашедшем сифилитическом процессе
задние столбы могут быть разрушены, так что больной не может сказать, где
находятся его руки и ноги, если он не видит их, и при ходьбе должен все
время смотреть на ноги.
При изучении
расположения и функции проводящих путей обнаружился один любопытный факт,
еще не получивший удовлетворительного объяснения. Все волокна спинного
мозга перекрещиваются, т. е. переходят с одной стороны тела на другую
где-либо на пути от рецептора к головному мозгу или от головного мозга к
мышце. Так, правая половина головного мозга контролирует левую половину
тела и получает сообщения от рецепторов левой стороны. Некоторые волокна
перекрещиваются , внутри самого спинного мозга, другие - в головном мозгу.
В центре серого вещества находится узкий
канал, проходящий вдоль всего спинного мозга и наполненный
цереброспинальной жидкостью, похожей на плазму крови. Спинной и головной
мозг покрывают три соединительно-тканные мозговые оболочки (тепшев).
Менингит - заболевание, при котором эти оболочки инфицируются и
воспаляются. Одна из них (твердая мозговая оболочка) прикреплена к костным
невральным дугам позвонков, другая (мягкая мозговая оболочка) лежит на
самой поверхности спинного мозга, а третья (паутинная оболочка) находится
между ними. Пространства между оболочками тоже наполнены цереброспинальной
жидкостью, так что спинной мозг (так же как и головной) плавает в этой
жидкости и защищен от ударов о твердую поверхность позвонков (или черепа)
при каждом движении.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА.
ГОЛОВНОЙ .МОЗГ
Головной мозг представляет собой
расширенный передний конец спинного мозга. У человека это расширение
настолько велико, что сходство со спинным мозгом в значительной степени
замаскировано, но у низших животных структурное родство головного мозга со
спинным ясно заметно. Детальная анатомия головного мозга чрезвычайно
сложна, и мы рассмотрим только шесть главных его отделов: продолговатый
мозг, варолиев мост, мозжечок, средний мозг, таламус и большие полушария
(рис. 268).
Продолговатый мозг - это самый задний отдел
головного мозга, лежащий непосредственно спереди от спинного мозга. Здесь
центральный канал спинного мозга расширяется и образует большую полость,
называемую четвертым мозговым желудочком (три остальных желудочка
находятся выше, в передних отделах головного мозга). Крыша четвертого
желудочка тонка и содержит сплетение кровеносных сосудов, выделяющих часть
цереброспинальной жидкости; остальную часть этой жидкости выделяют
аналогичные сплетения, находящиеся в других желудочках. В крыше четвертого
желудочка имеется три крошечных отверстия, через которые цереброспинальная
жидкость выходит в пространства между мозговыми оболочками. Стенки
продолговатого мозга толстые и состоят главным образом из нервных путей,
идущих к высшим отделам головного мозга. В продолговатом мозгу находятся
также скопления тел нервных клеток - нервные центры; это рефлекторные
центры, регулирующие ряд важнейших
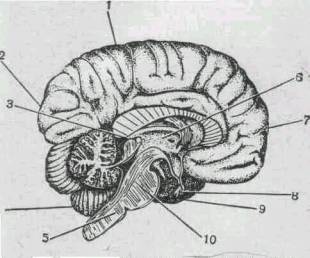
|
|
Рис 268. Срединный сагиттальныи разрез головного мозга
человека.
1 - теменная доля; 2 - затылочная доля; 3 - эпифиз 4 -
мозжечок; 5 - продолговатым мозг; 6 - таламус 7 - лобная доля; 8 -
гипофиз; 9 - височная доля 10 - средний мозг.
|
физиологических процессов:
дыхание, частоту сокращений сердца, расширение и сужение кровеносных
сосудов, а также глотание и рвоту.
Над
продолговатым мозгом расположен мозжечок, состоящий из средней части и
двух боковых полушарий, по форме напоминающих сосновые шишки. Серый
поверхностный слой мозжечка состоит из тел нервных клеток, а глубже
находится масса белой ткани, образованной волокнами, связывающими мозжечок
с продолговатым мозгом и с высшими отделами мозга. Величина мозжечка у
различных животных приблизительно соответствует их двигательной
активности. Мозжечок координирует движения и регулирует сокращение мышц;
поэтому у таких чрезвычайно активных животных, как птицы, он относительно
велик. Удаление или повреждение мозжечка не сопровождается параличом, но
ведет к расстройству мышечной координации. Птица после хирурги ческого
удаления мозжечка не способна летать. она может только беспорядочно
хлопать крыльями. При повреждении мозжечка у человека вследствие травм или
заболевания все движения становятся некоординированными и действия,
требующие тонкого согласования движений, например вдевание нитки в иголку,
оказываются невозможными.
На вентральной
стороне головного мозга под самым мозжечком лежит толстый поперечный пучок
волокон, так называемый варолиев мост, который проводит импульсы из одного
полушария мозжечка в другое, координируя движения мышц на обеих сторонах
тела.
Кпереди от мозжечка и варолиева моста
находится средний мозг, который имеет толстые стенки и узкий центральный
канал, соединяющий четвертый желудочек (в продолговатом мозгу) с третьим
желудочком (п таламусе). Толстые стенки среднего мозга содержат некоторые
рефлекторные центры и главные проводящие пути, ведущие к таламусу и
большим полушариям. На верхней стороне среднего мозга расположены четыре
невысоких округленных выступа, называемых четверохолмием, в которых
находятся центры некоторых зрительных и слуховых рефлексов. Рефлекторное
сужение зрачка при попадании в глаз яркого света регулируется центром,
находящимся в передних бугорках; в задних же бугорках лежит центр,
регулирующий настораживание ушей у собаки в ответ на звук. Средний мозг
содержит также группу нервных клеток, регулирующих мышечный тонус и позу.
Кпереди от среднего мозга центральный канал
снова расширяется и образует третий мозговой желудочек, крыша которого
содержит еще одно сплетение кровеносных сосудов, выделяющих
цереброспинальную жидкость. Толстые стенки третьего желудочка образуют
таламус. Это центр переключения сенсорных импульсов; волокна из спинного
мозга и низших отделов головного мозга образуют здесь синапсы с Другими
нейронами, идущими к различным сенсорным зонам больших полушарий. Таламус,
по-видимому, регулирует и координирует также внешние проявления эмоций;
так, раздражая таламус у кошки, можно вызвать у нее все внешние признаки
ярости: шерсть становится дыбом, кошка выпускает когти, выгибает спину
горбом и обнаруживает другие признаки враждебности. Но стоит только
прекратить раздражение, и проявления ярости исчезают.
На дне третьего желудочка (в гипоталамусе)
находятся центры, регулирующие температуру тела, аппетит, водный баланс,
углеводный и жировой обмен, кровяное давление и сон. Интересно, что
передняя часть гипоталамуса вступает в действие при повышении температуры,
а задняя - при понижении. Гипоталамус контролирует некоторые функции
передней доли гипофиза, например секрецию гонадотропных гормонов, и
вырабатывает гормоны, которые выделяет в кровь задняя доля гипофиза.
Рассмотренные до сих пор отделы головного
мозга управляют врожденными, автоматическими формами поведения, которые
определяются существенными чертами строения этих отделов; это строение в
основном одинаково у всех позвоночных - от рыб до человека. Большие
полушария мозга - самый передний и наиболее крупный из отделов головного
мозга - обладают совершенно иной функцией, заключающейся в регуляции
приобретенных форм. поведения. В основе сложных психологических явлений
сознания, умственной деятельности, памяти, понимания и истолкования
ощущений лежит активность нейронов большого мозга. Значение больших
полушарий у различных животных можно изучать, удаляя их хирургическим
путем. Лягушка после такой операции ведет себя почти в точности так же,
как нормальная лягушка, а голубь с удаленной корой больших полушарий
способен летать и, сидя на жердочке, сохранять равновесие, но склонен
часами оставаться в покое. Под действием раздражения он двигается, хотя и
бесцельно, как бы наудачу; так как он не способен есть предлагаемую пищу,
то при отсутствии искусственного питания он может умереть с голоду. Собака
после удаления коры больших полушарий способна ходить и проглатывать пищу,
вложенную ей в рот, но не обнаруживает никаких проявлений страха или
возбуждения. У человека иногда рождаются младенцы с неразвившейся корон
больших полушарий, и, хотя они способны к осуществлению вегетативных
функций дыхания и глотания, они ничему не научаются в результате опыта и
не делают никаких произвольных движений. Такие дети обычно умирают вскоре
после рождения.
В больших полушариях
сосредоточено несколько больше половины всех 10 млрд. нейронов, из которых
состоит нервная система человека. Большие полушария развиваются как
выросты переднего конца головного мозга. У человека и других млекопитающих
они растут назад, поверх остальных частей мозга и прикрывая их. Каждое
полушарие содержит полость, соединенную каналом с третьим желудочком (в
таламусе). Это первый и второй мозговые желудочки; в них, как и в
остальных желудочках, находятся сплетения кровеносных сосудов, выделяющих
цереброспинальную жидкость. Большие полушария состоят из серого и белого
вещества; белое вещество, образованное из нервных волокон, расположено
внутри, тогда как серое вещество, состоящее из тел нервных клеток,
находится на поверхности, образуя кору больших полушарий. Глубоко в
веществе больших полушарий лежат другие массы серого вещества - нервные
центры, служащие промежуточными станциями на пути в кору и из коры. У
низших позвоночных, у которых серого вещества мало, поверхность больших
полушарий гладкая, у человека же и у других млекопитающих она покрыта
извилинами. Таким образом, получаются валики, разделенные бороздами, что
увеличивает пространство, занимаемое серым веществом коры. Рисунок этих
извилин одинаков у всех людей независимо от их умственных способностей.
"География" коры больших полушарий подвергалась тщательному изучению (рис. 269).
Представление о том, что определенные участки головного мозга имеют особые
функции, возникло давно; еще "наука" френология основывалась на
предположении, что функции в мозгу локализованы определенным образом и что
если человек особо одарен в какой-либо области, то определенный участок
мозга должен быть увеличен и вызывает образование шишки на голове. Думали,
что анализ подобных "шишек" может показать, для какой деятельности больше
всего пригоден данный человек.
Экспериментальные данные позволили установить,
что функции в коре в значительной степени локализованы. Путем
хирургического удаления у подопытных животных отдельных участков коры
оказалось возможным строго локализовать многие функции; наблюдая параличи
или потерю чувствительности у людей с повреждениями или опухолями
головного мозга, а затем (после смерти больного) исследуя мозг и определяя
локализацию поражения, удалось составить "карту" также и человеческого
мозга. Во время операций на головном мозгу хирурги раздражали небольшие
участки электрическим током и наблюдали, какие мышцы при этом сокращаются;
поскольку мозговые операции могут производиться под местным
обезболиванием, больного можно было спросить, какие ощущения он испытывает
при раздражении того или иного участка. Интересно, что в самом мозгу нет
нервных окончаний, воспринимающих боль; поэтому раздражение коры
безболезненно. Новейший метод изучения активности головного мозга состоит
в измерении и регистрации электрических потенциалов, или волн, возникающих
в различных участках мозга (см. разд. 255).
Комбинируя данные, полученные различными
способами, исследователи смогли установить локализацию многих функций в
головном мозгу (рис.
269). В затылочной области находится центр зрения: удаление его
приводит к слепоте, а раздражение, даже в результате простого удара по
затылку, вызывает ощущение света. Удаление зрительной зоны на одной
стороне мозга вызывает слепоту на одну половину каждого глаза, так как
зрительный нерв каждого глаза расщепляется и половина волокон идет в одну
половину мозга, а другая половина - в другую. Центр слуха расположен в
височной доле мозга, над ухом. Раздражение его при ударе вызывает ощущение
звука; хотя удаление обеих слуховых зон приводит к глухоте, удаление одной
из них вызывает не глухоту на одно ухо, а двустороннее уменьшение остроты
слуха.
По наружной стороне полушария сверху
вниз проходит легко распознаваемая глубокая борозда (роландова борозда),
которая отделяет двигательную зону, контролирующую скелетные мышцы, от
лежащей позади нее области, ответственной за ощущение тепла, холода,
прикосновения и давления при раздражении рецепторов кожи. Внутри обеих зон
имеет место дальнейшая специализация участков вдоль борозды (от верхушки
мозга к его боковой стороне): нейроны верхнего участка контролируют мышцы
ступни, клетки последующих участков - мышцы голени, бедра, живота и т. д.,
а нейроны нижнего участка управляют мышцами лица. Величина корковой
двигательной зоны для той или иной части тела пропорциональна не
количеству мышечной ткани, а тонкости и сложности движений; особенно
обширны, например, зоны, управляющие кистью руки и лицом. Аналогичное
соотношение существует между частями сенсорной зоны и участками кожи, с
которых они получают импульсы. Таким образом, в связях между телом и
головным мозгом мы наблюдаем не только перекрещивание волокон, в
результате чего правая половина мозга контролирует левую половину тела-и
наоборот, но и еще одну инверсию, в результате которой самый верхний
участок коры управляет самыми нижними частями тела.
Зоны, функция которых известна, занимают у
крыс почти всю кору мозга, у собаки - значительную ее часть, у обезьяны -
несколько меньшую часть, а у человека - лишь небольшую долю всей
поверхности коры. Остальная поверхность, занятая так называемыми
ассоциативными зонами, состоит из нейронов, которые не связаны
непосредственно с органами чувств или мышцами,
|
|
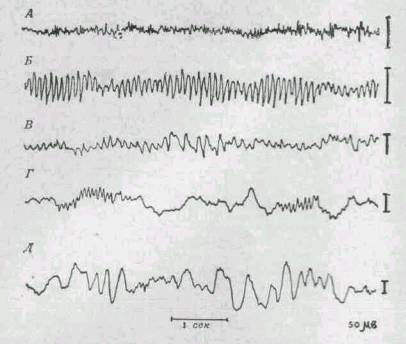
Рис. 270.
Электроэнцефалограммы человека,
зарегистрированные при возбужденни, в спокойном состоянии и на
различных зтапах сна.
На записи, сделанной во время возбуждения,
видны быстрые волны малой амплитуды, тогда как во время сна волны
более медленные и имеют большую амплитуду. Регулярные волны,
характерны для спокойного состояния, называются альфа-волнами.
А. Возбуждение, Б. Спокойное состояние. В. Сонливость. Г. Сон.
Д. Глубокий сон. |
а осуществляют
взаимосвязь между другими областями. Эти зоны лежат в основе высших
психических способностей (память, способность к логическому мышлению и
научению, воображение). Ассоциативные зоны каким то образом интегрируют
все разнообразные импульсы, непрерывно приходящие в мозг, и образуют из
них связное целое, обеспечивая возможность целесообразной реакции. Когда
вследствие заболевания или травмы функция одной или нескольких
ассоциативных зон выпадает, наступает так называемая афазия - состояние,
при котором утрачивается способность узнавать определенного рода символы.
При афазии одного типа больной не понимает написанных слов, хотя понимание
устной речи остается вполне нормальным. При афазии другого типа забываются
названия предметов, хотя знание и понимание функций этих предметов
сохраняются.
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ МОЗГА
Обмен веществ всегда сопровождается
электрическими явлениями; электрическую активность мозга можно регистрировать
при помощи прибора, называемого электроанцефалографом. Для этого прикрепляют
электроды к различным участкам кожи головы при помощи лейкопластыря и
исследуют активность лежащих под ними частей мозговой коры.
электроэнцефалография показала, что головной мозг постоянно находится в
состоянии активности, даже тогда, когда мы ни о чем не думаєм, и что наиболее
регулярные проявления активности - так называемые альфа-волны, -исходят из
зрительных зон затылочной доли, когда человек находится в покое и глаза его
закрыты. Эти волны возникают ритмически с частотой 9-10 в 1 сек и с
амплитудой около 45 мв (рис. 270). При открытых глазах альфа-волны исчезают и
вместо них появляются более быстрые, нерегулярные волны. Последние вызываются
видимыми объектами; это можно показать, воздействуя на глаза каким-нибудь
периодическим раздражителем, например светом, мигающим через правильные
промежутки времени: тогда на электроэнцефалограмме появляются волны с таким же
ритмом. Сон - единственное нормальное состояние, при котором электрическая
активность мозга резко изменена. Во время сна волны становятся медленнее и
выше (т. е. потенциал их возрастает), по мере того как человек впадает во все
более глубокое бессознательное состояние. При сновидениях регистрируются
нерегулярные волны.
При некоторых заболеваниях головного мозга характер
волн меняется. Например, у эпилептиков наблюдается своеобразный и легко
распознаваемый тип электроэнцефалограммы; подобные же изменения
обнаруживаются у людей, у которых ни разу не было эпилептического припадка,
но при известных условиях он мог бы произойти. Можно определить расположение
мозговых опухолей, установив, из какой части мозга исходят патологические
волны.
СОН
Нервные механизмы, с которыми связан сон,
неизвестны, и исследователям до сих пор не удалось выяснить, почему сон
необходим. Сон характеризуется уменьшением электрической активности коры
головного мозга (рис. 270), и с этим, возможно, связано его благотворное
действие на нервную систему. Спят только высшие позвоночные с хорошо развитой
корой головного мозга, причем животные, у которых большие полушария развиты
сильнее, по-видимому, больше нуждаются в сне.
Причиной сна люди обычно считают утомление, но
никаких экспериментальных данных, подтверждающих это мнение, нет. Одним из
важных факторов, вызывающих сон, служит отсутствие раздражителей; даже без
особенной усталости бывает легко заснуть, если нет ничего интересного, что
могло бы занять ум. Но хотя в присутствии раздражителей, привлекающих
внимание, мы не склонны засыпать, есть предел, за которым наступление сна
неизбежно. У всех высших животных жизнь характеризуется основным ритмом
чередования сна и бодрствования, и этот ритм регулируется гипоталамусом. В
передней части гипоталамуса находится центр сна, а в задней - центр
бодрствования. Полагают, что переход от бодрствования ко сну и обратно
регулируется механизмом с "обратной связью", в котором участвуют
эти два центра. У человека суточный ритм, включающий 8 час сна, несомненно,
является приобретенной привычкой; врожденный, естественный ритм состоит в
чередования сна и бодрствования через каждые 3 или 4 час, что можно видеть на
примере грудных детей.
ПСИХИЧЕСКИЕ БОЛЕЗНИ И НЕВРОЗЫ
Некоторые типы мозговых расстройств определенно
связаны с повреждением ткани мозга в результате болезни или ранения. Если
отверстие в крыше четвертого желудочка окажется закупоренным,
цереброспинальная жидкость не сможет выходить из желудочков и под ее
давлением начнется постепенное разрушение ткани. Или может лопнуть
кровеносный сосуд в мозговых оболочках, и вследствие давления скопившейся
крови будет разрушаться прилежащий участок мозга. Повреждение ткани может
произойти также в результате опухоли или инфекционных заболеваний, например
сифилиса. Симптомы (паралич, утрата чувствительности или других функций)
зависят от того, какая часть мозга поражена.
Причины нарушений другого типа, так называемых
функциональных расстройств -неврозов и психозов, менее понятны, так как при
этих заболеваниях патоморфологам до сих пор не удавалось обнаружить
каких-либо структурных или химических изменений в головном мозгу. Обычно
наблюдаются не столько изменения интеллекта, сколько эмоциональные нарушения.
Неврозы - сравнительно легкие и часто встречающиеся
расстройства с чрезвычайно разнообразными симптомами: беспокойством, страхом,
застенчивостью, чрезмерной обидчивостью и т. п. Эмоциональные расстройства
могут даже приводить к органическим нарушениям, например к нарушениям ритма
сердца и к расстройствам пищеварения. Причина этого рода психических аномалий
точно не установлена, и есть основания думать, что она в каждом отдельном
случае сложна и у разных людей различна. Существует теория, что неврозы
обусловлены внутренними конфликтами, и в некоторых случаях такое объяснение
кажется правдоподобным. Обычно, однако, неврозы бывают вызваны несколькими
факторами, действующими совместно (наследственность, окружающая обстановка, пережитые
в прошлом события, общее состояние здоровья). Во всяком случае, больной часто
совершенно не сознает причины своего неблагополучия. Не существует единого
метода лечения разнообразных неврозов; многие из них поддаются
психиатрическому лечению; другие неврозы постепенно исчезают спонтанно, без
лечения; третьи все более ухудшаются, а некоторые переходят в более серьезные
заболевания - психозы.
Психозы - это тяжелые психические заболевания, обычно
требующие госпитализации больного. Существует три основных типа психозов,
каждый из которых представляет собой преувеличенную форму определенных
нормальных тенденций. Маниакально-депрессивный психоз характеризуется
чередованием чрезмерного подъема с депрессией и иногда сопровождается
иллюзиями и галлюцинациями. Большинство страдающих им людей большую часть
своей жизни здоровы, но время от времени переживают повторные эпизоды
болезни. Паранойя - психоз, для которого характерны навязчивые идеи, обычно
мания величия или преследования. Для "dementia praecox" (один из
видов шизофрении) типичен уход от повседневной жизни в мир грез, который как
бы становится для больного реальным миром.
Лечить психозы гораздо труднее, чем неврозы, и в
большинстве случае не удается добиться стойкого излечения. Одним из самых энергичных
методов лечения является шоковая терапия, основанная на представлении о том,
что сильная встряска может возвратить такого 'больного в нормальное
состояние. Вводя больному инсулин или метразол или воздействуя электрическим
током, у него вызывают сильнейшие судороги. Такие способы лечения имеют много
недостатков, и механизм их действия не вполне выяснен, но в ряде случаев та
или иная разновидность шоковой терапии приводила к излечению. Во многих
случаях успешным оказывается лечение некоторыми новыми лекарственными
веществами, например хлорпромазином, и оно постепенно вытесняет люковую
терапию.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
От головного и спинного мозга отходят парные черепномозговые
и спинномозговые нервы связывающие мозг со всеми рецепторами и эффекторами
организма; эти нервы составляют периферическую нервную систему.
Черепно-мозговые и спинномозговые нервы состоят из пучков нервных волокон -
аксонов и дендритов. Что касается тел нервных клеток, то в периферической
нервной системе находятся только тела чувствительных нейронов, образующие
скопления (так называемые ганглии, или нервные узлы) вблизи головного и
спинного мозга, и тела некоторых двигательных нейронов вегетативной нервной
системы, которая будет рассмотрена позже.
Черепномозговые нервы. От различных отделов
головного мозга отходят 12 пар нервов, которые иннервируют главным образом
органы чувств, мышцы и железы, расположенные на голове. Эти же 12 пар,
иннервирующие сходные образования, мы встречаем у всех высших позвоночных - у
рептилий, птиц и млекопитающих; у рыб и амфибий имеются только первые 10 пар.
Подобно всем нервам, эти не вы состоят из отростков нейронов, хотя в одних
(I, II и VIII пары) эти отростки принадлежат только чувствительным нейронам,
в других (III, IV, VI, XI и XII пары) - почти исключительно двигательным, а в
третьих (V, VII, IX и Х пары) - как чувствительным, так и двигательным
нейронам. Названия черепномозговых нервов и иннервируемые ими структуры
указаны в табл. 11.
Один из самых важных черепномозговых нервов - блуждающий нерв, который
образует часть вегетативной нервной системы и иннервирует внутренние органы
грудной полости и верхней части брюшной полости.
Спинномозговые нервы. Все спинномозговые нервы
являются смешанными, т.е. содержат двигательные и чувствительные волокна в
примерно одинаковых количествах. У человека они отходят от спинного мозга
симметричными парами (всего 31 пара), и каждая пара иннервирует рецепторы и
эффекторы определенного участка тела. Каждый нерв начинается от спинного
мозга в виде двух тяжей, или корешков, которые вскоре соединяются, образуя
спинномозговой нерв. Все чувствительные волокна входят в спинной мозг через
задние корешки, а все двигательные волокна выходят из него через передние
корешки (рис. 271).
Если перерезать задний корешок, то иннервируемая данным нервом часть тела
полностью теряет чувствительность. но это не сопровождается параличом мышц.
Если же перерезан передний корешок, то наблюдается полный паралич мышц,
иннервируемых этим нервом, но ощущения прикосновения, давления и температуры,
кинестетические ощущения (чувство движения и положения частей тела) и болевая
чувствительность не нарушаются. Толщина каждого из спинномозговых нервов
соответствует величине участка тела, который он иннервирует; у человека
крупнее всех пара нервов, идущая к ногам. Каждый спинномозговой нерв вскоре
после слияния переднего и заднего корешков делится на три ветви: дорзальную
ветвь, иннервирующую кожу и мышцы спины, вентральную ветвь, идущую к коже и
мышцам боков и живота, и вегетативную ветвь, обслуживающую внутренности (рис. 271).
РЕФЛЕКСЫ И РЕФЛЕКТОРНЫЕ ДУГИ
Рефлекс - это автоматическая реакция на данный
раздражитель, зависящая только от анатомических взаимоотношений участвующих в
ней нейронов.
Т А
Б Л И Ц А 11
Черепномозговые
нервы человека
|
№ пары
|
Название
|
Места окончаний чувствитеяьных волокон
|
Эффекторы, иннервируемые двигателышми волокнами
|
|
I
|
Обонятельный
|
Обонятельный эпителий носа (обоняние)
|
---------------
|
|
II
|
Зрительный
|
Сетчатка глаза (зрение)
|
---------------
|
|
III
|
Глазодвигательный
|
Проприоцепторы мышц глазного яблока (мышечное чувство)
|
Мышцы, двигающие глазное яблоко (совместно с IV и VI
парами); мышцы, изменяющие форму хрусталика; мышцы, сужающие зрачок
|
|
IV
|
Блоковый
|
То же
|
Другие мышцы, двигающие глазное яблоко
|
|
V
|
Тройничный
|
Зубы п кожа лица
|
Некоторые из жевательных мышц
|
|
VI
|
Отводящий
|
Пропрлодепторы мышц глазного яблока (мышечное чувство)
|
Другие мышцы, двигающие глазное яблоко
|
|
VII
|
Лицевой
|
Вкусовые почки передней части языка
|
Мышцы лица; подчелюстные и подъязычные железы
|
|
VIII
|
Слуховой
|
Улитка (слух) и полукружные каналы (чувство равновесия,
поступательного движения и вращения)
|
Околоушная железа; мышцы глотки, используемые при
глотании
|
|
IX
|
Языко-глоточный
|
Вкусовые почки задней трети языка; слизистая глотки
|
---------------
|
|
Х
|
Блуждающий
|
Нервные окончания во многих Внутренних органах: легких,
желудке, аорте. гортани
|
Парасимпатические волокна, идущие к сердцу, жедудку,
тонким кишкам, гортани, пищеводу
|
|
ХI
|
Добавочный
|
Мышцы плеча (мышечное чувство)
|
Мышцы плеча
|
|
ХII
|
Подъязычный
|
Мышцы языка (мышечное чувство)
|
Мышцы языка
|
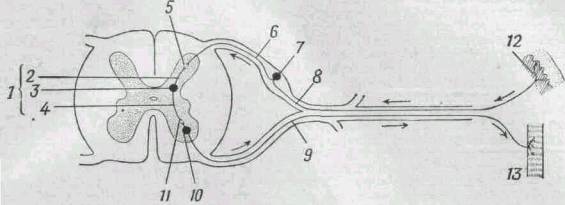
Нейроны, проводящие импульс, образуют
рефлекторную дугу·, простейшая дуга состоит из чувствительного, вставочного и
двигательного нейронов (рис.
272). Рефлексы являются функциональными единицами нервной системы, и
б6льшая часть нашей активности представляет собой результат рефлексов. Мы уже
видели, насколько важную роль играют рефлексы в регуляции ритма сердца. кровяного
давления, дыхания, слюноотделения, движений пищеварительного тракта и т. д. Еслп
мы наступили на что-нибудь острое или прикоснулись к чему-нибудь горячему, мы
не ожидаем, когда с участием головного мозга почувствуем боль и, подумав,
решим, что делать; наша реакция бывает мгновенной и рефлекторной. Мы
рефлекторно отдергиваем ногу или руку еще до того, как в головном мозгу
возникнет ощущение боли. Многие более сложные действия нашей повседневной
жизни, например ходьба, тоже в значительной мере регулируются рефлексами.
Врожденные рефлексы, общие всем людям, называются безусловными; рефлексы,
приобретенные позднее в результате опыта, называются условными.
Рефлексы классифицируют по числу участвующих в них
нервных путей. Типичным примером простого рефлекса, при котором раздражение
рецептора приводит к сокращению одной мышцы, может служить коленный рефлекс.
Если слегка ударить по сухожилию у коленной чашечки и тем самым растянуть
его, то рецепторы сухожилия подвергнутся раздражению, по рефлекторной дуге к
спинному мозгу и обратно пойдет импульс и мышца, прикрепленная к сухожилию,
сократится, что вызовет внезапное выпрямление ноги.
Важен, хотя и довольно прост, сгибательный рефлекс,
или рефлекс отдергивания конечности. При сильном раздражении кожи руки или
ноги происходит стимуляция мышц сгибателей и поврежденная конечность
отдергивается, прежде чем произойдет дальнейшее повреждение.
Многие рефлексы гораздо более сложны, чем
приведенные в этих примерах, но даже сложнейшие
|
|
|
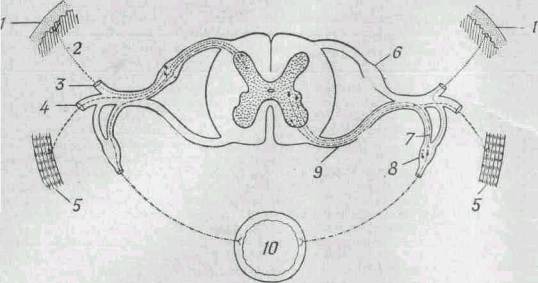
Рис 271. Схема основных типов чувствительных и двигательных нейронов спиномозговых
нервов и их связей со спинным мозгом.
Для удобства чувствительные нейроны показаны на левой стороне, а
двигательные - на правой, хотя на обеих сторонах тела имеются и те и
другие. 1 - кожа; 2 - нервные волокна от кожных рецепторов; 3 - задняя
ветвь; 4 - передняя ветвь; 5 - скелетная мышца; 6 - ганглий заднего
корешка; 7 - вегетативная ветвь; 8 - симпатический ганглий; 9 - передний
корешок; 10 - кишечник.
|
из врожденных рефлексов не требуют участия высших,
"мыслительных", процессов. Это можно продемонстрировать, если
разрушить у лягушки головной мозг, оставив в целости спинной мозг, и затем
прикладывать к спине животного кусочек бумаги, смоченный в кислоте. Сколько
бы раз мы ни прикладывали к коже кусочек бумаги, одна нога будет неизменно подниматься
и сбрасывать его. Эта реакция, требующая согласованного действия многих мышц,
является чисто рефлекторной и ясно демонстрирует одну из основных
особенностей всякого рефлекса - точную и безотказную повторяемость. Лягушка,
обладающая головным мозгом, может быть, повторила бы этот ответ два или три
раза, но в конце концов сделала бы что-нибудь другое - вероятно, ускакала бы
прочь. Большинство рефлексов полезно с точки зрения выживания животного;
анатомическая схема связей, обусловливающих рефлекс, закрепилась в эволюции
путем отбора именно благодаря своей полезности. Поведение только что
родившегося ребенка определяется врожденными унаследованными рефлексами; но
по мере того как ребенок растет, связь между раздражением и реакцией может
измениться н вместо прежнего раздражения ту же реакцию будет вызывать новый
раздражитель. Связь между новым раздражителем и прежней реакцией называется
условным рефлексом. Такие рефлексы обычно очень сложны, опосредованны и
трудны для изучения. Классические исследования русского физиолога И. И.
Павлова, изучавшего условные рефлексы на примере деятельности слюнных желез,
дали возможность объяснить их механизм и показали, в сколь больной степени
приобретенные формы поведения опираются на врожденное поведение.
В обычных условиях слюна рефлекторно выделяется лишь
тогда, когда пища находится во рту, но если одновременно с дачей пищи каждый
раз действует какой-нибудь другой раздражитель, например звонок, то
образуется условный рефлекс и слюна начинает выделяться в ответ на звонок
независимо от присутствия пищи. Способность к образованию условных рефлексов
связана с развитием коры больших полушарий;
|
|
|
Рис. 272, Схема рефлекторной дуги.
Путь импульса показан стрелочками.
1 - вставочный нейрон; 2 - дендрит; 3 - тело нейрона; 4 - аксон; 5 - синапс
между чувствительным и вставочным нейронами; 6 - аксон чувствительного
нейрона; 7 - тело чувствительного нейрона; 8 - дендрит чувствительного
нейрона; 9 - аксон двигательного нейрона; 10 - тедо двигательного нейрона;
11 - синапс между вставочными и двигательными нейронами; 12 - рецептор в
коже; 13 - мышца.
|
млекопитающие и птицы обладают этой способностью в большей степени, чем
низшие позвоночные, а у беспозвоночных она почти отсутствует. Если удалить
кору, то животное теряет все приобретенные ранее условные рефлексы и
способность к образованию новых. При прохождении одного импульса через синапс
синаптическое сопротивление понижается, так что импульс от нового
раздражителя следует по лути импульса в ранее выработавшемся рефлексе; затем
в результате облегчения, обусловленного повторением, новый путь закрепляется.
Один условный рефлекс может служить основой для другого, пока, наконец, не
выработается сложная иерархия условных рефлексов.
МЫШЛЕНИЕ, ПАМЯТЬ И НАУЧЕНИЕ
Пока еще невозможно дать полное физиологическое
объяснение психологическим явлениям мышления, памяти и научения. Полагают,
что в основе мышления лежат сложные физиологические процессы, связанные с
распространением нервных импульсов по определенным нейронным путям в мозгу. Возможно,
что та или иная мысль связана с прохождением нервных импульсов по замкнутой
нейронной цепи. Такого рода цепь могла бы включать от полудюжины до
нескольких сот или более нейронов и могла бы активироваться либо сенсорными
импульсами, либо импульсами, спонтанно возникающими в мозгу. Согласно этой
теории, мысли изменяются, когда нервные импульсы проходят по другой
"реверберирующей" цепи, образованной иной группой нейронов.
Воспоминания - это мысли, которые возникают
спустя некоторое время (исчисляемое минутами, месяцами или годами) после
первоначального появления этих мыслей и, по видимому, вызываются
возобновлением циркуляции импульсов в нейронной цепи. Было высказано
предположение, что непрерывное прохождение импульсов через синапсы данной
цепи ведет к уменьшению сопротивления в этих синапсах и повышает способность
цепи к проведению нервных импульсов. Процесс научения, возможно, связан с
многократным использованием определенного нейронного пути, приводящим в конце
концов к значительному уменьшению синаптического сопротивления.
В последние годы была выдвинута гипотеза о том, что
память связана с синтезом РНК и что каждое запоминаемое событие кодируется в
центральной нервной системе специфическими последовательностями нуклеотидов в
РНК. Установлены интересные корреляции между обменом РНК и нервной
деятельностью, однако физиологическая и биохимическая основа памяти остается
невыясненной.
Некоторые опыты, например такие, в которых животные
научаются проходить через лабиринт, чтобы получить пищу или избежать
электрических ударов, подчеркнули роль "проб и ошибок" в научении. У
высших млекопитающих - обезьян и человека - обнаруживается, кроме того,
феномен "понимания", или возникновения "идеи". После
немногих проб, сделанных наудачу, испытуемый схватывает сущность решения
задачи и затем решает ее с неизменным успехом.
Настроения и эмоции. Такие явления, как настроения и
эмоции, а также склад личности в целом тоже зависят от активности коры
больших полушарий, но лежащие в их основе нервные механизмы не выяснены. Эти
проявления, так же как и другие виды деятельности высших мозговых центров,
находятся под сильным влиянием физиологического состояния организма; на
состояние психики может резко влиять состояние желудка. Гормоны различных
эндокринных желез тоже влияют на работу головного мозга; например, у многих
женщин перед самой менструацией и во время ее бывают периоды душевной
депрессии, а менопауза (период, когда менструальный цикл прекращается обычно
в возрасте 40-50 лет) очень часто сопровождается глубокими эмоциональными и
психическими нарушениями.
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Сердце, легкие, пищеварительный тракт и другие
внутренние органы иннервированы особым комплексом периферических нервов, в
совокупности называемых автономной, или вегетативной, нервной системой.
Система эта в свою очередь состоит из двух частей: симпатической и
парасимпатической.
Вегетативная нервная система в целом содержит как
чувствительные, так и двигательные нервы, но она отличается от остальной
нервной системы рядом особенностей. Волевой контроль над этими нервами со
стороны больших полушарий невозможен; мы не можем произвольно ускорить или
замедлить ритм сердечных сокращений или действие мускулатуры желудка и кишок.
Далее, связь между чувствительными нервами и большими полушариями является
менее прямой, так что нормальное раздражение этих нервов не вызывает
ощущений. Еще одна важная особенность вегетативной системы состоит в том, что
каждый внутренний орган получает двойной набор волокон: одна группа их
подходит к органу через симпатические нервы, а другая - через
парасимпатические. Импульсы с симпатических и парасимпатических нервов
оказывают на иннервируемый орган противоположное действие. Если первые,
например, усиливают какую-либо активность, то вторые ослабляют ее. Эти
действия суммированы в табл. 12.
Т А Б Л И Ц А 12
Действие вегетативной
нервной системы
|
Нннервируемый орган
|
Действие симпатических нервов
|
Действие парасимпатических нервов
|
|
Сердце
|
Усиливают и ускоряют сокращения сердца
|
Ослабляют и замедляют сокращения сердца
|
|
Артерии
|
Вызывают сужение артерий и повышают кровяное давление
|
Вызывают расширение артерий и понижают кровяное давление
|
|
Пищеварительный тракт
|
Замедляют перистальтику, уменыиают активность
|
Ускоряют перистальтику, повышают активность
|
|
Мочевой пузырь Мускулатура бронхов
|
Вызывают расслаблеиие пузыря Расширяют бронхи, облегчают
дыхание
|
Вызывают сокращение пузыря Вызывают сокращение бронхов
|
|
Мышечные волокна радужной оболочки
|
Расширяют зрачок
|
Суживают зрачок
|
|
Мышцы, поднимающие волосы
|
Вызывают поднятие волос
|
Вызывают прилегание волос
|
|
Потовые железы
|
Повышают секрецию
|
Понижают секрецию
|
Следующая особенность вегетативной системы состоит в
том, что двигательные импульсы идут от головного или спинного мозга до
органа-эффектора не по одному нейрону, как импульсы ко всем другим частям
тела, а через два или большее число последователъных нейронов. Тело первого
нейрона этой цепи, так называемого преганглионарного нейрона, находится в
головном или спинном мозгу, а тело второго нейрона - постганглионарного - в
ганглии, лежащем где-либо вне центральной нервной системы (рис. 271). Тела
симпатических постганглионарных нейронов расположены вблизи спинного мозга,
ганглии парасимпатическнх нервов - вблизи иннервируемых органов или даже в их
стенках. Афферентные волокна от внутренних органов входят в центральную
нервную систему вместе с соматическими нервными волокнами.
Симпатическая система. Симпатическая нервная
система состоит из волокон, клеточные тела которых лежат в боковых столбах
серого вещества спинного мозга. Их аксоны выходят через передние корешки
спинномозговых нервов вместе с двигательными волокнами, идущими к скелетным
мышцам, а затем отделяются от этих волокон и образуют вегетативную ветвь
спинномозгового нерва, идущего к симпатическому ганглию. Эти ганглии парные;
с каждой стороны спинного мозга лежит цепочка из 18 ганглиев, которая тянется
от шеи до области живота (рис.
273). В каждом ганглии аксон первого нейрона образует синапс с дендритом
второго нейрона. Тело этого второго нейрона находится внутри ганглия, а его
аксон направляется к иннервируемому органу.
Кроме волокон, идущих от каждого спинномозгового
нерва к соответствующему ганглию, имеются волокна, идущие от одного ганглия к
следующему. Аксоны некоторых из вторичных нейронов идут от симпатического
ганглия обратно к спинномозговому нерву и проходят в его составе к
иннервируемым потовым железам, мышцам, поднимающим волосы, и мускулатуре стенок
кровеносных сосудов. Аксоны других вторичных нейронов направляются от шейных
симпатических ганглиев вверх к слюнным железам и к радужной оболочке глаза.
Чувствительные волокна симпатической системы проходят внутри тех же нервных
стволов, что и двигательные, но вступают в спинной мозг через задние корешки
вместе с другими чувствительными нервами, не принадлежащими к вегетативной
системе.
Парасимпатическая система. Это система состоит
из волокон, начинающихся в головном мозгу и выходящих в составе III, VII, IХ
и особенно Х (блуждающего) черепномозговых нервов и из волокон, начинающихся
в крестцовом отделе спинного мозга и выходящих со спинномозговыми нервами
этого отдела (рис. 273).
Блуждающий нерв берет начало в продолговатом мозгу и спускается через область
шеи в грудную и брюшную полости, где иннервирует сердце, дыхательную систему
и пищеварительный тракт. Толстые кишки, мочевая система и половые органы
иннервируются парасимпатическими волокнами через тазовые спинномозговые
нервы. Радужная оболочка глаза, подъязычные и подчелюстные железы и
околоушная железа иннервированы соответственно III, VII и IX парами черепномозговых
нервов. Все эти нервы содержат аксоны первых нейронов в цепи; ганглии
парасимпатической системы расположены в иннервируемых ими органах или около
них, так что все аксоны вторичных нейронов сравнительно короткие.
НЕРВНАЯ СИСТЕМА НИЗШИХ ЖИВОТНЫХ
Одноклеточные животные, как, например, амеба и
парамеция, не имеют нейронов, так как их тело состоит, всего лишь из одной
клетки. Амеба отлично обходится без какого-либо приспособления для
интеграции, но парамеция, тело которой покрыто тысячами мельчайших ресничек,
нуждается в каком-то механизме, который координировал бы биение этих
ресничек, чтобы она могла передвигаться. Это осуществляется системой
тончайших нитей, так называемых нейромоторных волокон, которые тянутся от
переднего конца тела ко всем ресничкам. Волокно можно перерезать, произведя
хирургическую операцию под микроскопом; после этого биения ресничек уже не
координируются и происходят беспорядочно.
Наиболее низко организованные многоклеточные животные
- губки - тоже не имеют нервной системы; специализированные нервные клетки
впервые появляются у гидры и других кишечнополостных. Нервные клетки этих
животных не отделены друг от друга синапсами и не объединены в истинную
нервную систему, а либо представляют собой отдельные разветвленные клетки,
либо образуют нервную сеть, состоящую из многих клеток с соединенными между
собой ветвистыми отростками, так что импульс, возникший в одной части тела,
может распространяться по всем направлениям во псе· остальные части. Нервные
клетки гидры не
|
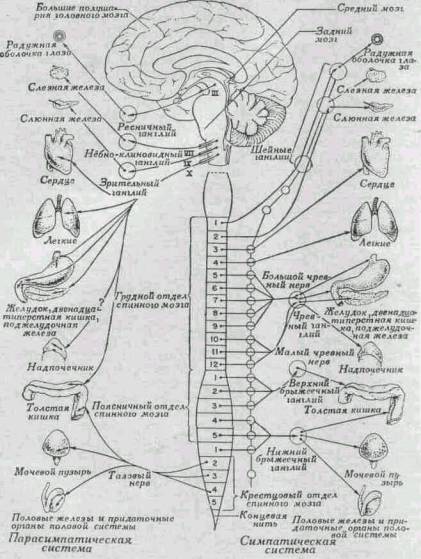
|
|
Рис. 273. Схема вегетативной нервной системы.
Парасимпатическая система показана слева, симпатическая - справа. Римскими
цифрами обозначены порядковне номера черепномозговых нервов.
|
дифференцированы на чувствительные, вставочные и двигательные нейроны, а
просто одни ветви нервной сети направляются к рецепторным клеткам, а другие -
к сократимым. Степень реакции у гидры зависит от силы раздражения: слабый
укол иглой в щупальце может вызвать свертывание одного лишь этого щупальца,
более же сильный укол заставит сократиться все щупальца и все тело, и
животное сожмется в шарик.
У большинства беспозвоночных нервная система в
высокой степени централизована. Особенно это относится к членистоногим
(насекомые, пауки, крабы, раки), моллюскам (кальмары, улитки) н кольчатым
червям (например, дождевые черви). Дождевой червь имеет настоящую нервную
систему с аксонами и дендритами, образующими хорошо выраженные нервные тяжи и
волокна. Эта система дифференцирована на центральную и периферическую системы
с отдельными чувствительными, вставочными и двигательными нейронами,
соединенными между собой синапсами, так что нервный импульс может идти только
в одном направлении. Это позволяет центральной нервной системе играть роль
интегрирующего механизма, который отбирает некоторые из подходящих импульсов
и передает их на эффекторы, затормаживая или подавляя другие импульсы. У
дождевого червя имеется центральный нервный тяж, идущий по всей длине тела,
что позволяет отдельным сегментам тела осуществлять координированные
движения. Самая передняя часть нервного тяжа представляет собой расширение,
иногда называемое "головным мозгом", которое посылает импульсы
вдоль всего тяжа с целью координации движений. После удаления этого
"мозга" животное может двигаться почти так же энергично, как и
раньше, но при встрече с препятствием оно не свернет в сторону, а будет
продолжать бесплодные попытки продвинуться вперед. Таким образом,
"мозг" необходим для движений приспособительного характера. Нервные
системы других высших беспозвоночных более или менее сходны с нервной
системой дождевого червя. У всех этих животных нервная цепочка расположена на
брюшной стороне, под пищеварительным трактом н не имеет полости внутри.
У позвоночных нервный тяж расположен на спинной
стороне тела и имеет центральную полость. Основной план строения нервной
системы у всех позвоночных сходен; различия касаются главным образом развития
отдельных частей головного мозга и размеров последнего по отношению к
размерам спинного мозга.
|