Первая часть была написана более года назад, но сейчас новые данные появляются очень быстро, и по этой причине возникла необходимость уточнить некоторые из изложенных в ней гипотез. Для начала напомним, что на основе анализа паралогичных ДНК-полимераз эукариот был, в частности, сделан вывод о том, что по совокупности фактов наиболее вероятно, все эукариоты произошли от организма, напоминавшего современную микроспоридию. Однако, недавно был частично отсеквенирован геном паразитирующего одноклеточного организма Mitosporidium daphniae (отсеквенированный штамм получил название Microsporidia sp. UGP3). После построения соответсвующих филогенетических деревьев, был сделан вывод о том, что это представитель микроспоридий очень рано отделился от их общего предка. Данный организм интересен, в частности, ещё и тем, что, в отличии от остальных ("классических") микроспоридий, у него ещё не полностью редуцировался митохондриальный геном. После добавления к общему дереву эукариотных ДНК-полимераз организмов, геномы которых были отсеквенированы со времени написания первой части исследования происхождения эукариот, выяснилось, что ген дельта-полимеразы указанного выше организма ложится на соответствующем дереве отдельно от полимераз остальных микроспоридий, но, зато, рядышком с дельта-полимеразой криптомицеты (или, как транскриптируют их название некоторые другие учёные, криптомикоты) Rozella allomycis. Так как криптомицеты и по многим другим признакам являются сестринской группой по отношению к грибам, а так же родственны микроспоридиям, остаётся признать, что что у "классических" микроспоридий, утративших митохондрии, дельта-полимераза была просто-напросто замещена в результате горизонтального переноса вирусной. Это косвенно подтверждается ещё и тем, что у микроспоридии Encephalitozoon romaleae, геном который отсеквенирован полностью, по соседству с геном дельта-полимеразы расположен ген, имеющий гомологию с обратной транскриптазой ретровируса. Таким образом, мой вывод о близости микроспоридий к общему предку всех эукариот, сделанный на основе анализа дерева паралогов ДНК-полимераз, по-видимому, ошибочен.
Что же, попробуем зайти с другой стороны. Если не учитывать микроспоридии, то следующим наиболее древним таксоном на дереве паралогов ДНК-полимераз получаются экскаваты, а это уже хорошо согласуется с работами последних лет, в частности, с выводами из статьи Евгения Кунина и К "MitoCOGs: clusters of orthologous genes from mitochondria and implications for the evolution of eukaryotes". Или, вот, например, совсем свежая статья на ту же тему - "Bacterial proteins pinpoint a single eukaryotic root". Ради любопытства, можно попробовать на основе идеи использования в качестве внешней группы митохондриальные гены, построить дерево для цитохрома b. Данный ген удобен тем, что он, во первых, достаточно длинный, что, при прочих равных, повышает разрешающую силу при детализации графа, а с другой стороны, весьма широко распространён в большинстве царств эукариот. В итоге, получается картина, приведённая ниже.
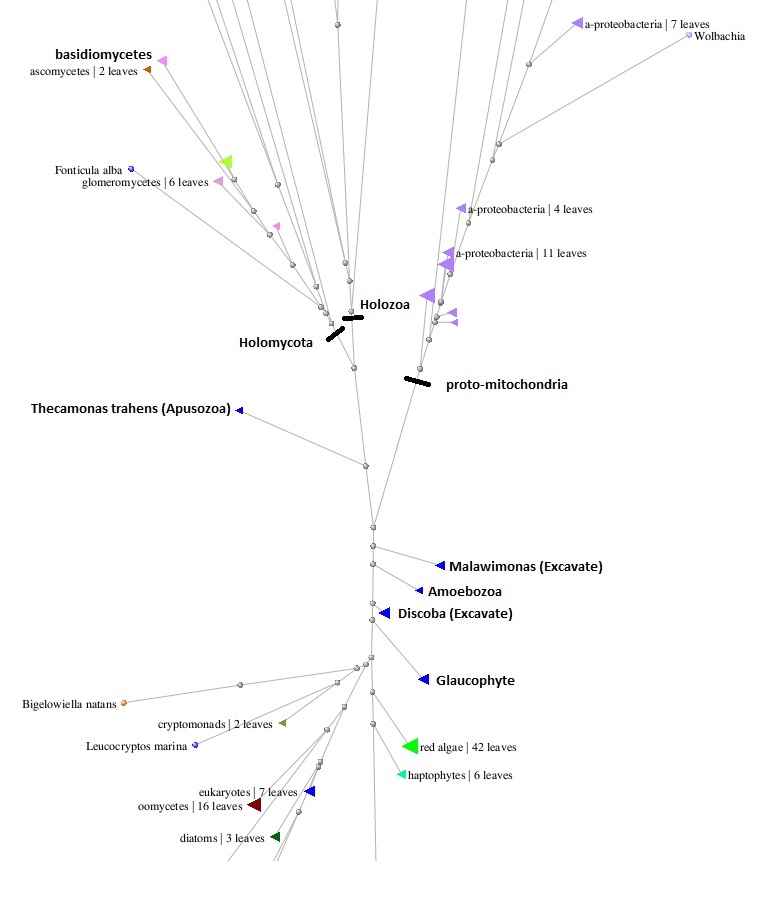
Рис. 1. Объединённое (геномы альфа-протеобактерий + митохондриальные геномы эукариот) филогенетическое дерево протеина цитохром b.
Как видно, от гена, доставшегося последнему общему предку эукариот (LECA) от альфа-протеобактерии, которую "приручили" его прешественники, превратив в митохондрию, пути двух его гомологов расходятся. Одна ветка, включающая, в частности, водросли и наземные растения, уходит вниз, а другая, включающая, в том числе, животных и грибы, вверх, тем самым, разделяя всё надцарство эукариот на две большие группы. В своё время, Кавальер-Смит предложил называть их соответственно, биконта и униконта, по типовому количеству жгутиков у их одноклеточных представителей (или их зооспор, или, в случае многоклеточных, у их мужских половых клеток). При этом, нужно иметь в виду, что, с одной стороны, у некоторых униконта (амёб, тех же микроспоридий и т.д.) жгутики в процессе эволюции исчезли вообще, а с другой стороны, у некотрых биконта может быть более двух жгутиков (например, многочисленные реснички у хорошо знакомой нам всем со школьной скамьи инфузории туфельки являются, по сути, сильно размножившимися модифицироваными жгутиками). По указанной причине надёжнее смотреть не столько на морфологические признаки, сколько на редкие события, происходящие на уровне генома. В частности, выяснилось, что у всех биконта гены дигидрофолат-редуктазы и тимидилат-синтазы в результате редкой генетической мутации объединились в один. Для группы униконта тоже есть характерный для неё уникальный генетический признак - вставка на определённой позиции в белке так называемого мышечнего миозина (миозин II) дополнительной аминокислоты (глицина). Данные признаки можно рассматривать в качестве своеобразных характерных генетических "шрамов", которые обычно навсегда остаются в геномах всех потомков организма, у которого произошла соответсвующая мутация. Ближе всего к общему корню у нас оказался класс Malawimonas, которого одни исследователи относят к царству Экскават, а другие, вместе с другим странным организмом, имеющим много архаичных черт, и носящим имя Collodictyon, предлагают выделить в отдельную группу протистов, сёстринскую по отношению к амёбозоям. Сами амёбозои, как видно из рисунка, тоже располагаются поблизости от точки разделения ветвей альфа-протеобактерий, биконта и униконта, что косвенно свидетельствует о их близости к последнему общему предку всех эукариот. В этой же области вблизи корня находятся и Discoba, являющиеся одним из типов Экскават.
Наверное, настало время вкратце описать, каковы морфологические признаки царства Экскават, представители которых (как и представители других групп протистов, имеющих с ними достаточно много общих черт) оккупируют нижние ветви дерева эукариот. Во-первых, большинство из них относятся к биконта, то есть, имеют два или более жгутика (либо произошедших от них ресничек). Во-вторых, в тельце их клетки имеется своеобразное углубление (карман), из дна которого, как правило, и выходят жгутики. Обычно данное углубление выполняет функции своеобразного "рта" одноклеточного, в который попадают его будущие жертвы - бактерии и более мелкие протисты. При этом, движения жгутиков обычно способствуют активной циркуляции воды внутрь "кармана", что повышает вероятность захватывания добычи. Как правило, Экскаваты движутся вперёд той стороной, на которой расположено углубление со жгутиками (примерно как самолёт с пропеллером), поэтому их ещё можно условно называть "переднежгутиковыми", в отличии от описанной выше группы униконта, у которых единственный жгутик, как винт у корабля, расположен сзади, за что их называют "заднежгутиковыми". В большинстве случаев жгутики у биконта попарно объединены в так называемые кинетиды, представляющие из себя два тесно связанных между собой жгутика вместе с цилиндрическими структурами, в которых они закреплены - кинетосомами. Обычно кинетосомы заякорены на клеточной или ядерной мембране и располагаются под прямым углом друг к другу. При этом, есть основания полагать, что, скорее всего, жгутики были организованы в кинетиды уже у последнего общего предка всех эукариот. Любопытно, что при делении клетки протиста каждая из двух кинетосом удваивается, после чего в каждой из вновь появившихся клеток появляется по кинетиде, в которой одна кинетосома (иногда вместе со жгутиком) досталась от материнской клетки, а другая является вновь созданной. Учитывая, что для многих Экскават характерна резкая ассиметрия жгутиков, один из которых (тот, который, собственно, и обеспечивает движение клетки) является "главным", а другой считается вспомогательным, при делении возникает дилемма, какой из жгутиков второй кинетосомы "назначить" главным. И здесь действует жёсткое правило, выполнявшееся, по видимому, ещё у LEСА - главным становится тот жгутик, который уже "оттрубил" в предыдущей жизни свой срок в качестве вспомогательного. Как видим, и у одноклеточных наблюдаются факты "дедовщины". :)
Ещё один интересный факт относительно кинетид заключается в том, что по своему строению они фактически полностью гомологичны таким классическим клеточным органеллам как центросомы (в частности, и те, и другие, состоят из пары расположенных перпендикулярно по отношению друг к другу цилиндров, состоящих, в свою очередь, из девяти триплетов тубулиновых микротрубочек), играющим очень важную роль в организации и синхронизации различных внутриклеточных процессов эукариот, в том числе, в делении их клеток. Более того, уже к концу 19-го века стало ясно, что эти структуры по мере надобности могут переходить одна в другую. С учётом сказанного, уже не вызывает удивления тот факт, что, как выясняется, у хламидомонад именно кинетосомы-центриоли после деления клетки тащат вновь образовавшееся ядро на то место в клетке, которое ему надлежит занять. После "отпочкавания" новой кинетосомы от родительской стратегия выращивания на её основе жгутика у разных групп протистов сильно варьирует. У одножгутиковых вторая кинетосома, как правило, остаётся пустой вплодь до этапа следующего деления клетки, а у биконта второй жгутик может отрастать сразу же после окончания процесса деления клетки (митоза). Но такое поведение наблюдается не всегда, иногда у вновь появившихся кинетосом сразу же вырастают новые жгутики, которые сохраняются в течении всего митоза и последующих этапов жизни протиста. Для многих групп Экскават типично тесное объединение двух кинетид с ядром эукариотической клетки, на котором они прочно заякорены. Такой морфологический комплекс (объединённые попарно 4 жгутика со своими кинетосомами, каждый из котрых заякорен на ядерной мембране) получили название кариомастигонты.
Интересное исследование недавно проделали авторы статьи, в которой они проанализировали всё многообразие центров организации микротрубочек (ЦОМТ), являющихся частным случаем центросом, в различных царствах эукариот. Как уже говорилось выше, в большинстве случаев центросомы (ЦОМТ), по существу, составляют с жгутиковой системой эукариотической клетки одно целое, так что, изучая различные варианты заякоривания кинетид (базальных телец) на мембране с помощью микротрубочек, можно попытаться реконструировать наиболее вероятную структуру ЦОМТ (вместе с соответствующей системой жгутиков) LECA. Это было сделано, и в результате авторы статьи пришли к выводу, что система жгутиков, и, соответственно, ЦОМТ последнего общего предка всех эукариот больше всего напоминала соответствующие структуры сегодняшних Экскават, а генеральной тенденцией эволюции жгутиковой системы в большинстве линий эукариот было её упрощение, которое наиболее далеко продвинулось у заднежгутиковых, к которым принадлежим и мы с вами. Таким образом, два разных метода (филогенетический анализ и морфологический анализ жгутиковых систем) независимо приводят к одному и тому же выводу - из сохранившихся до наших дней царств эукариот наиболее близком к LECA являются Экскаваты. В частности, у авторов уже упоминавшейся выше статьи "Bacterial proteins pinpoint a single eukaryotic root" именно они оказываются наиболее близкой к LECA группой протистов со стороны биконта. Если же смотреть со стороны униконта, то наиболее рано отделившейся ветвью будут тоже весьма близкие в морфологическом плане к Экскаватам Malawimonas и Collodictyon.
Особенно интересен в плане исследования возможных характерных черт LECA Collodictyon, в котором причудливо смешаны типичные признаки Экскават и Амёбозой. Некоторые исследователи считают это семейство самым архаичным протистом из тех, что дожили до наших дней. Одним из его любопытных свойств является то, что его жертвы (бактерии и одноклеточные водоросли) после поглощения хищником, по-видимому, погибают отнюдь не сразу. Более того, те же водоросли не просто какое-то время продолжают как ни в чём не бывало жить внутри Коллодиктиона, но нередко им удаётся даже "сбежать" от своего врага, проделав брешь в его мембране. Данный факт, возможно, служит намёком на то, каким образом некоторые группы эукариот заполучили цианобактерий в качестве своих симбиотов.
Как видно, в общем и целом получается, что у LECA были два или четыре передних жгутика, питался он, видимо, бактериями, обладал митозом закрытого типа, осуществляемым с помощью ассоциированных с клеточным центром (центросомой) микротрубочек, укоренённых в двух находящихся под прямым углом друг к другу (по существу, по полностью аналогичной с парой кинетосом схеме) центриолей. Кроме участия в делении клеточного ядра, клеточный центр так же служил для формирования микротрубочек другого типа, служащих для формирования кинетосом и крепящихся к ним жгутиков. Вообще, стоит отметить, что появление у эукариот такой структуры, как ЦОМТ (наравне с появлением ядра, приобретением митохондрий, способности к фагоцитозу, и т.д.), является одним из важнейших моментов в их эволюции. Кроме того, в жизненном цикле LECA могла присутствовать характерная для многочисленного семейства современных слизевиков фаза плазмодия, что можно при желании рассматривать как преадаптацию к появлению многоклеточности. Вполне вероятно, что у LECA в передней части клетки так же имелось углубление, выполнявшее функции своеобразного "рта", наподобие того, что и сегодня существует у многих Экскават. Будем надеяться, что благодаря быстрому прогрессу молекулярной биологи, тайна происхождения эукариот в ближайшие годы если и не будет раскрыта полностью, то, во всяком случае, количество белых пятен в истории этого важнейшего эволюционного события существенно уменьшится.