Несколько лет назад на "Элементах" была опубликована статья А.Маркова про гипотезу Г. Гончикова о происхождения эукариот в результате симбиоза спорулирующих клостридий и архей-метаногенов. В ней, в частности, приводится ряд фактов, демонстрирующих некоторые общие черты, сближающие процессы размножения клеток эукариот без разрушения оболочки ядра (так называемый закрытый митоз) и образования спор у спорулирующих бактерий (особенно, у одной из их групп - эндоскелетных клостридий). Примечательно, что у некоторых клостридий, ведущих паразитический образ жизни, спорогинез используется не для того, что бы пережить неблагоприятные условия, а, как и у эукариот, для размножения. Хотя общий сценарий гипотезы Гончикова представляется автору не очень правдаподнобным, сама идея о связи эукариогенеза и споруляции, на мой взгляд, вполне имеет право на жизнь. Нельзя ли как-то проверить её независимым способом? Если одну из главных ролей в процессе происхождения эукариот действительно играли спорулирующие бактерии, то выглядит вполне логичным, что и первые эукариоты размножались с помощью спор. Таким образом, локализовав на филогенетическом дереве эукариот район предполагаемого корня, и определив, не находятся ли вблизи него размножающиеся с помощью спор эукариоты, мы можем найти аргументы за или против гипотезы о происхождении эукариот в результате модификации процесса споруляции. Итак, за дело!
Для целей предстоящего исследования выберем семейство ДНК-полимераз эукариот – альфа, дельта и дзета. Эти полимеразы есть у большинства эукариот – альфа отвечает за первоначальный этап удлинения праймера, дельта как правило является основной ДНК-полимеразой, а дзета служит для полимеризации проблемных (повреждённых) участков исходной ДНК. При этом, считается наиболее вероятным, что все они произошли в результате дубликации предковой архейной ДНК-полимеразы семейства B (можно отметить, что дублирование какого-нибудь гена с достаточно широким спектром функций с последующей специализацией каждой копии на более узкой задаче, это вообще "фирменный" метод эволюции эукариот). Это даёт нам шанс попытаться локализовать корень дерева эукариот, так как исходно указанные три гена должны были исходно обладать очень высокой степенью гомологии между собой, и лишь постепенно в процессе эволюции они дивергировали от точки «Большого Взрыва», коим я называю факт появления на эволюционной сцене эукариот. В качестве отправной точки можно выбрать ген дзета полимеразы многоклеточной красной водоросли Chondrus crispus, так как, насколько известно автору, отпечатки в породах возрастом 1.2 млрд. лет другой многоклеточной красной водоросли (Bangiomorpha) являются древнейшими из следов эукариот, по интерпретации которых у учёных ещё наблюдается единодушие. Результаты работы Бласта (это общедоступная система поиска гомологичных генов и построения соответствующих графов, отражающих степень их родства) по построению филогенетического дерева одной тысячи генов, наиболее похожих на исходный, приведены ниже.
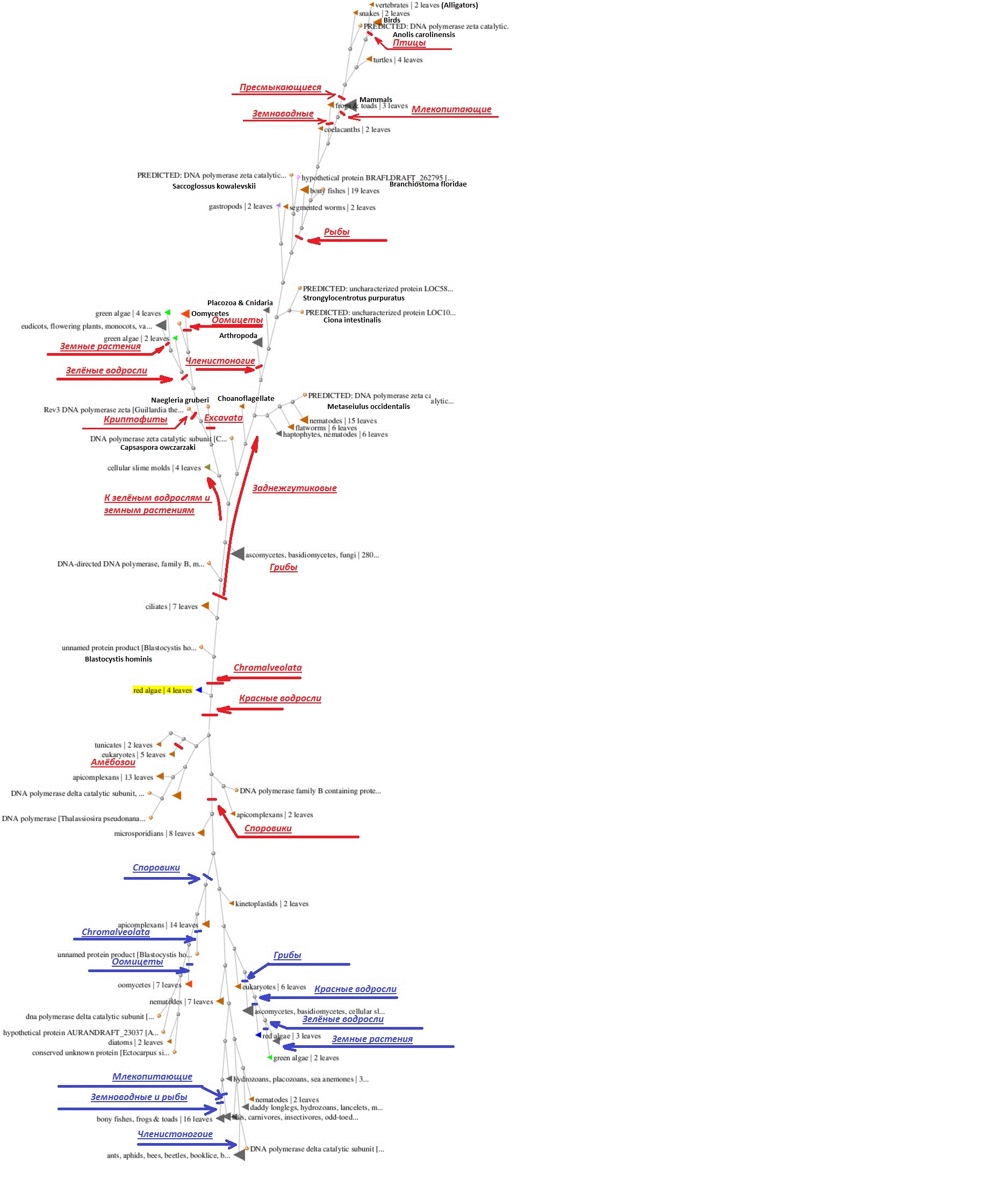
Рис. 1. Филогенетическое дерево дзета и дельта полимераз эукариот.
Так как гены дзета и дельта полимераз более похожи между собой по сравнению с геном альфа-полимеразы, в итоговое дерево вошли, в основном, лишь поддеревья соответствующих генов. В верхней половине рисунка располагается поддерево дзета-полимеразы, а в нижней – дельта полимеразы. Для наглядности я добавил некоторые пояснительные надписи, причём для поддерева дзета они сделаны красным цветом, а для поддерева дельта – синим. К концу я немного обленился, и пожамкал (схлопнул) часть поддеревьев в нижней части, поэтому она оказалась меньше по размеру. Кроме того, надо отметить, что (в виде исключения) у пресмыкающихся, и, как следствие, у птиц, была утрачена дельта-полимераза, поэтому соответствующие подветки в нижней части дерева отсутствуют. Но, в общем и целом, как видно по картинке, деревья получаются на удивление «правильными», по крайней мере, в общих чертах соответствующими сегодняшним представлениям об эволюции «кроны» эволюционного дерева эукариот, что даёт некую надежду, что и вблизи основания полученная картинка имеет ощутимую связь с реальностью, тем более, что обе половинки общего дерева по сути зеркально-симметричны не только в области короны, но и вблизи корня, как и можно было бы, в принципе, ожидать от дерева паралогичных генов. Теперь остаётся посмотреть, что же находится в районе Большого Взрыва (БВ), к которому в нашей расширяющейся Вселенной Генов сходятся «синяя» и «красная» половинки нашего дерева? В самом его центре мы обнаружим микроспоридий, окаймляемых близким к ним по многим признакам споровиками (они же апикомплексы). Смотрим на геном микроспоридий и обнаруживаем, что они являются одним из тех очень редких таксонов, у которых нет ген дзета-полимеразы! Конечно, она могла быть утрачена в результате массовой потери генов, сопровождающейся приспособлением к паразитическом образу жизни. Но не исключён и вариант, что у них ещё просто не успела произойти соответствующая дубликация гена. У близких к микроспоридиям споровиков во многих случаях уже обнаруживается три, или даже более, генов данного семейства, но в большинстве случаев все они гораздо сильнее похожи на гены дельта и альфа полимераз, чем на дзета полимеразу. Кстати говоря, возможно, это одна из причин небольшого размера геномов микроспоридий. Не имея хорошего способа репликации повреждённых участков ДНК трудно поддерживать в рабочем состоянии геномы больших размеров. Отсюда и их экологическая ниша паразитизма, так как с небольшими геномами гораздо проще живётся, когда во-первых, многие необходимые ферменты можно просто получать от хозяина в готовом виде (а от генов, кодирующих соответствующие белки, можно избавиться), а во-вторых внешние условия очень стабильны, так что многие гены, ответственные за адаптацию к изменению внешних условий, попросту не нужны.
Для более уверенных выводов хорошо бы повысить разрешение метода (образно говоря, улучшить разрешение нашего условного телескопа, через который мы рассматриваем центр нашей эукариотной "Вселенной"). Для этого надо перенести исходную точку координат как можно ближе к месту БВ, и оставить в запросе к Бласту лишь те группы организмов, которые находятся в эпсилон-окрестности предполагаемого корня. Я выбрал для этого ген дельта-полимеразы одного из видов микроспоридий, результат показан на рисунке ниже (за подписью eukaryotes там скрываются амёбозои).
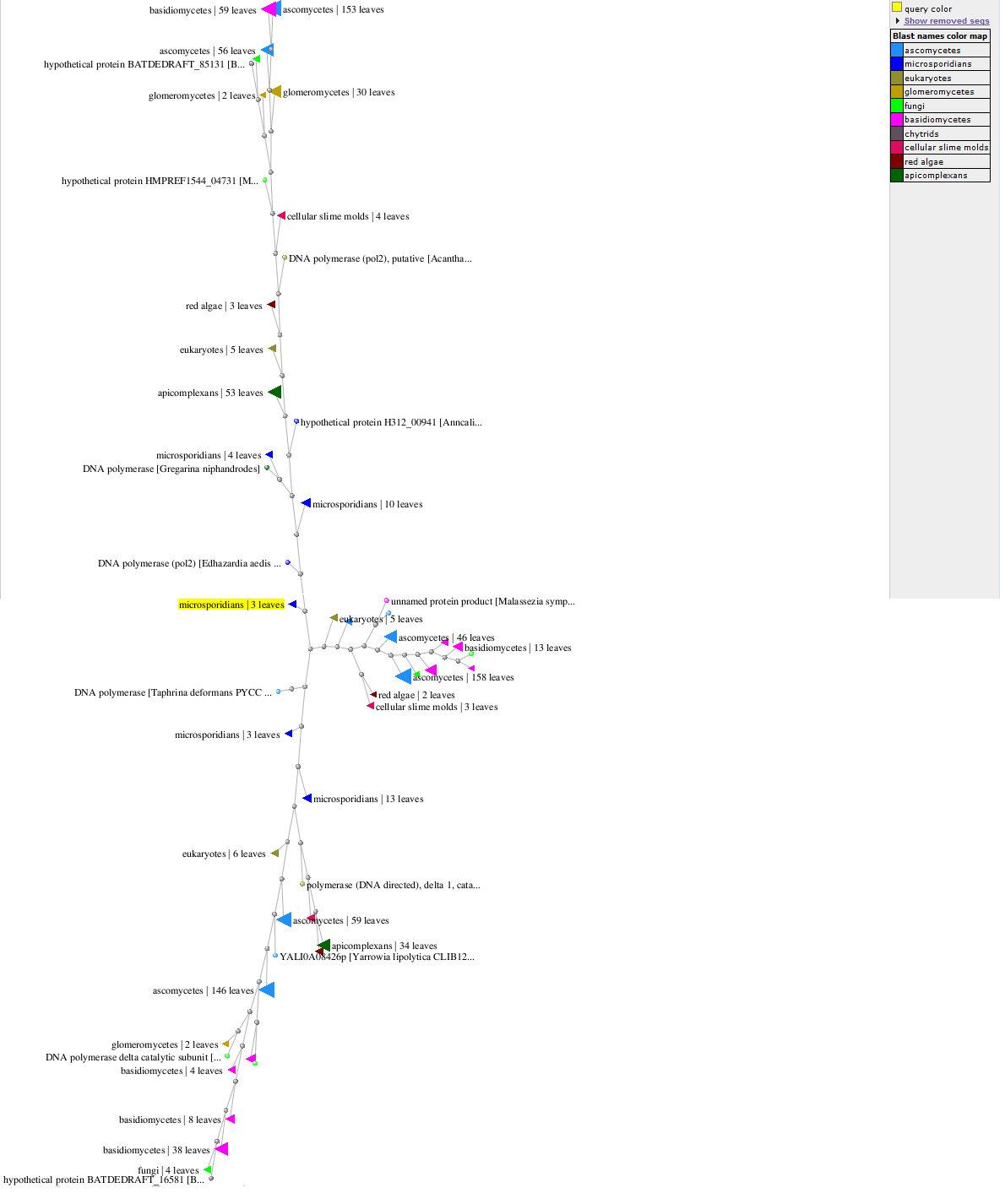
Рис. 2. Филогенетическое дерево альфа, дзета и дельта полимераз эукариот вблизи их общего корня.
Как видно, от «центрального перекрёстка» (он же индикатор места БВ) теперь отходят уже три ветви, соответствующие альфа, дельта и дзета полимеразам. Ветвь альфа-полимераз уходит вверх, ветвь дельта-полимераз - вниз, дзета полимераз – вправо. Таким образом, и при более сильном «увеличении» в центре нашей Вселенной генов эукариот находятся микроспоридии (ген ДНК полимеразы гриба-аскомицета Taphrinia появился там, по видимому, в результате ГПГ). Дальше, по мере удаления от центра, появляются, соответственно, споровики, амёбозои, слизевики, красные водоросли, грибы.
Итак, в непосредственной близости от общего корня эукариот мы наблюдаем микроспоридий и споровиков, являющихся размножающимися с помощью спорогенеза облигаторными паразитами многоклеточных животных. На первый взгляд, такой результат выглядит неожиданно. Но, возможно, паразитический образ жизни это как раз то, что позволяет выжить в изменяющихся внешних условиях (одна только серия сильнейших оледенений незадолго перед Кембрием чего стоит) архаичным видам, которым иначе было бы трудно к ним быстро адаптироваться. А жизнь внутри хозяина гарантирует более-менее стабильные условия, не требующие от генома быстрых адаптивных реакций (см. выше). Как уже было упомянуто ранее, некоторые клостридии, ведущие паразитический образ жизни, тоже размножаются через спорогинез, при этом, из одной материнской клетки образуется не одна, а сразу несколько спор. Возможно, у них это когда-то был один из основных способов размножения, который потом у большинства бактерий выродился в метод пережить "трудные времена" в анабиозе. Во всяком случае, до сих пор у большинства спорулирующих бактерий вначале образуются две септы на концах клетки, и лишь потом одна из них (случайно выбранная) разбирается.
Любопытно, что микроспоридии в различных исследованиях уже несколько раз оказывались вблизи предполагаемого корня эукариот (из недавних можно упомянуть, например, вот эту статью), но каждый раз это списывали на дефекты метода, в частности, эффект притяжения длинных ветвей. Однако, по крайней мере от последнего эффекта, данный способ, по идее избавлен, так как корни паралогичных генов по определению должны располагаться примерно в одной и той же точке.
Кстати говоря, по-видимому, одним из самых древних из ныне вымерших многоклеточных эукариот была Tappania, первые отпечатки которой находят в слоях возрастом в 1.4-1.6 млрд. лет. По поводу её классификации ещё идут споры, но всё же большинство палеонтологов относит её к вымершим ныне грибам (возможно, имевшим некое отдалённое родство с современными аскомицетами и зигомицетами). Факт любопытный, особенно в свете того, что микроспоридии сейчас принято относить к грибам. Наконец, можно упомянуть и про отпечатки таинственных акритархов, которые встречаются начиная с пластов архея. Некоторые исследователи считают, что акритархи являются спорами ныне вымерших эукариот.
Итак, есть основания полагать, что в появлении эукариот могли играть активную роль некие организмы, родственные спорулирующим бактериям. Как же они дошли до такой жизни? Обязательно ли для этого предполагать симбиоз архей и бактерий? Об этом и многом другом мы попытаемся порассуждать в следующей статье.
PS.
Примерно через год после окончания данной статьи выяснилось, что оригинальные ДНК-полимеразы микроспоридий, по-видимому, были замещены в результате горизонтального переноса генов вирусными. Таким образом, вывод о их близости к последнему общему предку эукариот, скорее всего, ошибочен. Подробнее об этом см. в моей новой статье.