По современным научным представлениям Земля образовалась около 4.54 млрд. лет назад. Древнейшие породы возрастом более 4 млрд. лет обнаружены на северо-западе Канады и в Австралии. Однако, в столь древних породах никаких признаков жизни пока не обнаружено. Впрочем, это и не удивительно, так как они гораздо лучше сохраняются в осадочных породах, формирующихся в водоёмах, а указанные породы имеют отнюдь не осадочное происхождение. Долгое время считалось, что первые несомненные признаки существования земной биоты прослеживаются в гнейсах формации Исуа (Гренландия), возраст которой оценивается в 3.7-3.8 млрд. лет. Данное предположение основывалось на обнаружении в указанных гнейсах некоторого дефицита изотопов "тяжёлого" углерода 13С по отношению к его "лёгкому" изотопу 12C, по сравнению с их обычным для Земли соотношением, что обычно свидетельствует о признаках биологической активности. Хотя многие учёные до сих пор продолжают ссылаться на указанные гнейсы, как первое несомненное подтверждение существования жизни на Земле не менее 3.7 млрд. лет назад, в последнее время вышло несколько публикаций, ставящих под сомнение, что пласты породы, содержащие следы облегчённого углерода, имеют тот же возраст, что и основная формация, а не образовались позже (см, например, здесь или тут). Так же необходимо учитывать, что при определённых условиях фракционирование углерода возможно и за счёт небиологических процессов. Наконец, нужно так же иметь в виду, что вполне естественное желание многих исследователей открыть нечто "самое-самое древнее" иногда приводит к некоторым конфузам. Например, в течении примерно десятка лет считалось, что древнейшие следы эукариот и цианобактерий имеют возраст около 2.7 млрд. лет, однако, лишь относительно недавно выяснилось, что биомаркеры, обнаруженные в породах соответствующего возраста, просочились туда позже из более молодых геологических слоёв вместе с нефтью. Таким образом, не имея других подтверждений, весьма рискованно делать выводы о наличии в тех или иных породах признаков жизни соответствующего возраста, лишь на основе наличия в них облегчённого углерода или биомаркеров, так как это может оказаться следствием их позднейшего загрязнения или результатом активности небиологических процессов. Ниже мы ещё кратко вернёмся к вопросу о возможности появления жизни на Земле практически сразу же после окончания так называемой поздней тяжёлой бомбардировки, пока же ограничимся констатацией факта, что имеющиеся к настоящему времени данные не позволяют сделать однозначного вывода о том, присутствовала ли жизнь на Земле 3.7 млрд. лет назад.
Первые несомненные признаки жизни появляются в породах, датируемых возрастом 3.4-3.5 млрд. лет почти одновременно в западной Австралии и южной Африке. Есть достаточно надёжные данные, свидетельствующие о том, что некогда эти породы составляли единый древнейший материк времён архея - Ваальбара. Что же представляют из себя эти признаки жизни? Во-первых, это строматолиты - особые карбонатные (то есть, содержащие в своей основе соединения углерода) образования, которые большинством учёных считаются ископаемыми остатками цианобактериальных матов. Во-вторых, это предпологаемые отпечатки самих бактерий. В наиболее древних слоях преобладают отпечатки ниточных форм, размером и внешним видом напоминающие некоторые виды современных цианобактерий. Например, морфотип номер IV из обзора Шопфа (на сегодняшний день, это, пожалуй, один из наиболее систематизированных материалов по свидетельствам жизни в Архее) весьма напоминает современные цианобактерии рода Arthronema, располагающиеся, кстати говоря, вблизи основания эволюционного дерева цианобактерий, на первой же ветке, отделившейся от предпологаемого общего предка всех цианобактерий Gloeobacter violaceus (см. мою статью про происхождение хлорофильного фотосинтеза). В третьих, в обсуждаемых породах зафиксировано соотношение изотопов "лёгкого" и "тяжёлого" углерода, характерное для их фракционирования в результате биологических процессов, в частности, фотосинтеза. Как уже отмечалось ранее, само по себе это соотношение ещё не может считаться окончательным доказательством следов наличия жизни, но, принимая во внимание наличие и других вышеперечисленных "улик", можно сделать практически однозначный вывод - три с половиной миллиарда лет назад на Земле уже была жизнь, причём, в форме достаточно "продвинутых" бактерий, по внешнему виду весьма напоминающих некоторые виды своих современных собратьев. При этом, многие признаки (типичная для фотосинтетиков доля 13С, локализация бактериальных матов преимущественно на мелководье, то есть, в пределах прямой досягаемости солнечных лучей, внешнее сходство многих отпечатков с более молодыми отпечатками, уверенно классифицирующимися как цианобактерии) указывают на то, что как минимум значительная часть обитателей древнейших бактериальных матов получали необходимую им энергию с помощью хлорофильного фотосинтеза. Однако, у учёных до сих пор пока нет единого мнения о том, осуществлялся ли этот фотосинтез цианобактериями (то, есть, был ли он оксигенным), или другими, неоксигенными фотосинтетиками. Дело в том, что каждая из точек зрения поддерживается на первый взгляд достаточно вескими аргуменами. Так, в пользу того, что 3.4-3.5 млрд. лет назад фотосинтез был неоксигенным обычно приводят следующие аргументы:
- первые более-менее очевидные признаки наличия свободного кислорода в морской воде и атмосфере появляются не ранее, чем 2.7-2.8 млрд. лет назад (с резким увеличением его концентрации, так называемая "кислородная катастрофа", примерно 2.4 млрд. лет назад);
- для оксигенного фотосинтеза нужна весьма продвинутая система извлечения энергии фотонов с двумя различными скоординированно работающими реакционными центрами, появление которой на столь раннем этапе эволюции многим авторам представляется нереалистичным;
- на строматолитах возрастом более 3 млрд. лет не удаётся обнаружить признаков формирования крошечных пузырьков с кислородом (являющегося для них своеобразным отходом производства), в то время как на более молодых остатках цианобактериальных матов они обычно обнаруживаются;
- метод молекулярных часов даёт время появления цианобактерий не ранее 2.8 млрд. лет.
Послушаем теперь начальника транспортного цеха аргументы сторонников более древнего происхождения оксигенного фотосинтеза. Во-первых, это, естественно, уже упомянутое сильное сходство многих отпечатков в древнейших слоях с некоторыми видами современных цианобактерий (см., например, всё то же обзор Шопфа). Во-вторых, известно как минимум одно месторождение сланцевого керогена на северо-западе Австралии возрастом 3.2 млрд. лет, происхождение которого очень трудно объяснить как-либо иначе, кроме результата жизнедеятельности оксигенных бактерий. В третьих, кислород, выделяемый цианобактериями, в первое время мог сразу же идти на окисление железа, которого в океанах архея было достаточно много, формируя так называемы джеспилиты (известны со времён примерно 3.7 млрд. лет назад). Так же кислород в архее мог хотя бы частично нейтрализовываться биологическим путём в цикле жизнедеятельности матанотрофных бактерий, получающих энергию за счёт сжигания метана (с образованием углекислого газа и воды). Это косвенно подтверждается и совпадением периода, когда он, наконец, стал накапливаться в атмосфере, с резким падением в морской воде концентрации никеля, являющегося необходимым микроэлементом для бактерий-метаногенов. Указанными причинами можно объяснить и отсутствие пузырьков с кислородом в наиболее древних цианобактериальных матах (точнее, того, что от них осталось к настоящему времени). Образно говоря, кислород, как активный окислитель, был в восстановительной внешней среде палеоархея "на расхват", и вступал в химические реакции ещё задолго до того, как на поверхности матов могло накопиться такое его количество, которое необходимо для формирования макроскопических пузырьков. Аргумент о том, что сложная система оксигенного фотосинтеза не могла появиться столь рано, на самом деле тоже довольно шаток. С одной стороны, как я пытался показать в статье про эволюцию хлорофильного фотосинтеза, есть достаточно веские аргументы в пользу того, что из доживших до нашего времени бактерий именно цианобактерии являются наиболее древними. С другой стороны, когда говорят про недостаточность времени эволюции, неявно подразумевают, что жизнь зародилась именно на Земле. Но эта гипотеза ничуть не лучше подтверждается фактами, чем гипотеза внеземного абиогенеза, см. мою статью по этому поводу. Итак, остаётся последний аргумент, связанный с молекулярными часами. Но и он при ближайшем рассмотрении оказывается достаточно уязвимым. Не рассматривая подробно все проблемы, возникающие при калибровке молекулярных часов на таких больших промежутках времени для бактерий, живущих в очень широком спектре внешних условий, упомянем лишь одно обстоятельство, которое следует иметь в виду даже если предположить, что скорость хода часов откалибрована идеально. Дело в том, что все выводы о возрасте тех или иных таксонов в данном случае справедливы лишь в предположении, что соответствующее филогеническое дерево не только правильно построено, но так же правильно определено и положение его корня. Но как раз по вопросу о том, какие из ныне живущих на Земле бактерий являются самыми древними, то есть, филогенетически ближе всего к гипотетическому общему предку всех земных организмов, у учёных в настоящее время нет общего мнения! Вывод о том, что возраст цианобактерий не превышает 2.8 млрд. лет, получается в рамках предположения, что наиболее древними земными организмами являются термофильные бактерии. Однако, это лишь одна из гипотез (поддерживаемая в настоящее время большинством учёных, но и количество не согласных с ней тоже велико, причём, имеет тенденцию к постепенному увеличению). Автору ближе альтеренативная точка зрения заключающаяся в том, что корень дерева жизни ближе к грам-положительным бактериям, а из них, скорее всего, к клостридиям. Этому впоследствии будет посвящена отдельная статья, пока же, можно просто констатировать, что в рамках указанного альтернативного сценария перенос на эволюционном графе точки, соответствующей началу эволюции, от термофильных бактерий к клостридиям "состаривает" цианобактерии до возраста примерно в 3.9 млрд. лет, что уже не противоречит интерпретации древнейших отпечатков архея как останков цианобактерий. В общем, как видимо уже догадались наши читатели, сам автор больше склоняется к версии о том, что в древнейших слоях архея действительно запечатлены остатки цианобактерий, хотя вопрос, конечно, пока нельзя считать полностью закрытым.
Какие ещё бактерии обитали в океанах палеорхея? По-видимому, мы можем достаточно уверенно дополнительно назвать как минимум одну разновидность бактерий, составлявших тогда компанию цианобактериям, это сульфат-редуцирующие бактерии, которые были обнаружены в породах возрастом приблизительно 3.4 млрд. лет в западной Австралии. Исследованные останков бактерий в указанных породах, проведённое сразу по нескольким направлениям, начиная от величины фракционирования изотопов серы, характерных для его биологического происхождения, и заканчивая изучением под электронным микроскопом структур, ассоциирующихся с остатками бактериальной клеточной стенки, позволило практически наверняка исключить все остальные гипотезы кроме интерпретации найденных образований как остатков древних бактерий, живущих за счёт метаболизма соединений серы, причём, наиболее вероятно, восстановления сульфатов до сульфидов. Можно отметить, что подобные бактерии, точно так же как и цианобактерии, живут и процветают на Земле по сей день, причём, они, в принципе, способны жить в гордом одиночестве на многокилометровой глубине! Рассмотрим несколько подробнее детали их метаболизма. В качестве источника электронов для дыхания они, в отличии от цианобактерий, используют не воду, а водород или простейшие органические молекулы, в частности, ацетат. В качестве акцепотра электронов в данном случае выступает сульфат - SO42-. Следует отметить, что, судя по всему, сульфата, по крайней мере, в области геологически активных зон, в архее было достаточно много, так как он является конечным продуктом, появляющимся в результате серии химических реакций, вызванным попаданием во внешнюю среду в результате вулканических извержений сернистого ангидрида (SO2), причём, наиболее эффективно указанные реакции идут при поверхностных и мелководных извержениях, то есть, именно в тех областях, где преимущественно и располагались в архее бактериальные маты. Кстати, для того, что бы сульфат мог "принять на грудь" отвоёванный бактерией у внешней среды электрон, он вначале должен быть активирован. Делается это с помощью специального белка-фермента, а конечная схема реакции представлена на Рис. 1. Как нетрудно заметить, молекула с активированным сульфатом на борту (крайняя справа) до боли напоминает хорошо знакомую нам молекулу АДФ у которой один из атомов фосфора заменён на серу. Ещё одно подтверждение тому, сколь узок круг этих революционеров, как сильно канализированы биологические метаболические цепочки, как часто достаточно поменять всего одну деталь в биологическом Лего, что бы получить из того же набора элементарных кирпичиков новую функциональность.
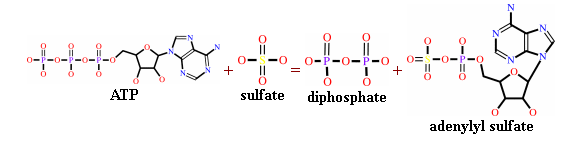
Рис. 1. Схема реакции активации сульфата перед тем, как он будет готов присоединить к себе дополнительный электрон (взято из Википедии).
В качестве отходов серные бактерии в процессе своего метаболизма "выдыхают" сероводород (SH2) аналогично тому, как животные выделяют при дыхании в качестве отработанного вещества углекислый газ. Существенная часть сероводорода тут же вступала в химическую реакцию с широко распространённым (в растворённом виде) в океанах архея железом, что в конечном итоге приводило к образованию нерастворимого в воде пирита (FeS2) по наличию залежей которого (вместе с наличием в породе останков бактерий) часто и удаётся идентифицировать древние сообщества серных бактерий. Так же интересно отметить, что в месторождениях пирита обычно наблюдается и повышенная концентрация двух других важнейших для живых систем химических элементов - кобальта и селена. Кстати говоря, тот факт, что серные бактерии могут использовать в качестве источника электронов не только водород, но и ацетат (хотя и с меньшим выходом свободной энергии) позволяет предположить любопытную схему взаимодействия различных групп бактерий в бактериальных матах. Дело в том, что самыми, так сказать, "архаичными" из доживших до наших дней бактерий, по мнению автора являются ацетогены - способные к спорогенезу бактерии, встречающиеся, в основном, среди клостридий, бацилл и актинобактерий (подробнее мы обсудим эту тему в одной из моих будущих статей). Ацетогены используют в своём метаболизме на входе водород в качестве источника электронов и углекислый (CO2) или угарный (CO) газ в качестве их приёмника, выделяя при этом во внешнюю среду уксусную кислоту (CH3СОOH). В свою очередь, серные бактерии могут использовать выделяемый ими ацетат (наравне с оставшимся от ацетогенов водородом) в качестве источника электронов для собственного метаболизма. Так же можно отметить, что в современных бактериальных матах в самом низу, там, где концентрация свободного кислорода в воде минимальна, часто располагаются бактерии-бродильщики, использующие в качестве основного источника энергии и материала для построения собственного тела органические остатки уже умерших бактерий, оседающие из верхних слоёв. В качестве основного побочного продукта их жизнедеятельности во внешнюю среду выделяется водород. Наконец, как уже было сказано в статье про хлорофилльный фотосинтез, большинство фотосинтетиков, включая цианобактерии, могут, в свою очередь, использовать выделяемый серными бактериями сероводород как источник свободных электронов для собственного метаболизма (окисляя его до серы или даже до сульфатов). В результате получается такая предположительная схема строения древнего бактериального мата. В самом низу могли бы располагаться ацетогены и бродильшики. Выделяемый ими ацетат (то бишь, по простому говоря, уксус) и водород могли бы использовать расположенные выше сульфаторедуцанты. Наконец, в самом верхнем слое могли располагаться фотосинтетики (включая цианобактерий), утилизирующие выделяемый серными бактериями сероводород. При этом, как уже отмечалось выше, некоторые фотосинтетики (например, часть пурпурных бактерий) способны окислять сероводород не только до молекулярной серы, но даже до сульфата, приготавливая, таким образом, "пищу" для своих расположенных на один уровень ниже сульфат-редуцирующих собратьев. Получается бактериальное сообщество, в котором все группы бактерий завязаны в своём метаболизме друг на друга, что так характерно и для большинства современных цианобактериальных матов! На первый взгляд в предложенной выше схеме может показаться странным, почему цианобактерии в палеоархее переключились с привычной им воды на сероводород? Но логически рассуждая, при условии наличия в окружающей среде достатачного количества сероводорода, переход на него как на основой поставщик электронов и протонов эволюционно как раз вполне оправдан. Ведь при этом цианобактериям приходится тратить практически на порядок меньше столь ценной для них свободной энергии, достигая де факто того же результата. Как говорится в известном рекламном ролике, если результат тот же, зачем платить больше? Кстати говоря, и сегодня, например, цианобактерии вида Oscillatoria limnetica прекрасно растут и размножаются в озёрах с высоким содержанием сероводорода за счёт бескислородного фотосинтеза. И ещё одна примечательная деталь - именно цианобактерии порядка Oscillatoriales, к которому и принадлежит указанный вид, судя по оставшимся от них отпечаткам, и преобладали в цианобактериальных матах 3.4-3.5 млрд. лет назад (см. уже неоднократно упомянутый выше обзор Шопфа). Таким образом, в рамках указанной гипотезы можно хотя бы частично примерить сторонников и противников существования цианобактерий уже в палеоархее. Цианобактерии могли действительно существовать на Земле уже в тот ранний период её геологической истории, но при этом в процессе своего метаболизма они, возможно, использовали не оксигенный, а преимущественно аноксигенный фотосинтез!
Наконец, нельзя обойти вниманием ещё один важный вопрос. В настоящее время большинство исследователей считают одной из древнейших форм жизни своеобразную группу бактерий, выделенную в отдельный домен археи, а первым ветвлением на дереве жизни - разделение архей и "обычных" бактерий (их, что бы не путать с археями, часто называют эубактериями). Подобный вывод основан, в первую очередь, на достаточно сильных отличиях аппарата репликации их ДНК и характерных особенностях строения их мембраны и клеточной стенки, не встречающихся среди эубактерий. Кроме того, анализ филогенетического дерева рРНК тоже показывает очень раннее отделение архей от основного ствола дерева (хотя аналогичные филогенетические деревья, построенные для большинства белков, задействованных в стандартных метаболических цепочках, обычно демонстрируют для архей не большую удалённость от корня дерева, чем все остальные группы "обычных" бактерий). В то же время, среди учёных есть и немало "оппозиционеров", придерживающихся альтернативного взгляда, утверждающего, что архей среди самых первых обитателей Земли не было, они появились несколько позже, "отпочковавшись" от грам-положительных бактерий. Что можно сказать о возможности существования архей в слоях возрастом 3.2 млрд. лет и более? Никаких отпечатков, которые можно было бы классифицировать как возможные останки арехеобактерий, в этих породах пока не обнаружено. В настоящее время существует лишь одна публикация, авторы которой претендуют на обнаружение в породах возрастом более 3.2 млрд. лет (кратон Пилбора в Австралии) пузырьков газа с примесью метана, соотношение изотопов углерода которого характерно для матаболизма архей-метаногенов. Однако, это косвенное свидетельство нельзя считать неопровержимо достоверным. Во-первых, опять же, существуют абиогенные процессы (идущие, например, в железно-никелевых сплавах), которые генерируют метан с изотопным соотношением углерода, характерным для биологических процессов. А то, что такие процессы по-видимому реально происходили на древней Земле, косвенно подтверждается обнаружением в частицах циркона возрастом более 4.2 млрд. лет углерода с типично "био-метаногенным" отклонением отношения 13C/12С от его стандартного значения (его в научных публикациях обычно обозначают как δ13) - до -58-ми промилле. К тому же, по-видимому, нельзя априори полностью исключить и возможное загрязнение указанных пород проникшими в них много позже из более высоких геологических слоёв археями. Во-вторых, следующие (и, на этот раз, многочисленные) следы метаболизма метаногенов появляются лишь спустя примерно 700 млн. лет (см. например, здесь). Такой большой временной провал в палеогеологической летописи не может не настораживать. Например, временное расстояние между самым ранним свидетельством существования сульфаторедуцантов и следующей по времени находкой составляет величину порядка 200 млн. лет, а свидетельств существования цианобактериальных матов в породах древнее 3.2 млрд. насчитывается более десятка. Соответственно, не отвергая, в принципе, возможность существования архей уже в палеоархее (такой вот выходит каламбур), с точки зрения автора по совокупности имеющихся на сегодняшний день фактов убедительных подтверждений этому пока нет. Что, впрочем, тем более верно для ацетогенов и бродильщиков.
С учётом вышеизложенного, можно ещё раз под несколько иным углом зрения вернуться к вопросу о возрасте цианобактерий. Давайте совместим на одном графике интенсивность процесса образования в различные геологические эпохи архея пород вулканического происхождения, и количество остатков (циано?)бактериальных матов, обнаруженных в них палеонтологами. Результат приведён на рис. 2, где вулканические породы обозначены чёрным, а строматолиты зелёным. Как нетрудно заметить, наблюдается явная положительная корреляция между интенсивностью процессов извержения магмы и активностью биологических процессов в соответствующий период. В частности, локальные пики обоих процессов в большинстве случаев почти совпадают, а в период между 3.2 млрд. лет и 3.0 млрд. лет не наблюдается ни пород вулканического происхождения, ни наличия строматолитов. О чём это может говорить? Например, о том, что древнейшие очаги жизни на Земле остро нуждались в подпитке многими важными для них химическими веществами, которые могли поступать в местах повышенной геологической активности вместе с магмой и вулканическими газами. Без этих своеобразных "костылей" древняя биота поначалу чувствовала себя неуверенно и имела склонность к исчезновению в условиях, когда эти процессы начинали ослабевать. Особенно существенным могло быть отсутствие водорода в форме, удобной для утилизации бактериями. Во-первых, водород является самым распространённым в ДНК, РНК и белках химическим элементом. Во-вторых, как отмечает Федонкин, "Многие группы прокариот используют молекулярный водород в качестве донора электрона и источника энергии, и, фактически, конкурируют между собой за доступ к водороду". В этой же работе автор ссылается на эксперименты, подтверждающие, что только что изверженные вулканические породы продолжают эмитировать во внешнюю среду водород на протяжении многих дней даже после их остывания. С другой стороны, молекулярный водород является очень лёгким веществом, и благодаря этому весьма эффективно диффундирует из атмосферы в космическое пространство. Всё это приводит к тому, что без постоянной подпитки за счёт процессов дегазации магмы он оказывается в большом дефиците. Например, можно отметить, что несмотря на то, что при разложении цианобактериями молекулы воды на каждую молекулу кислорода выделяется одна молекула водорода, в настоящее время концентрация свободного кислорода в земной атмосфере примерно на 6 порядков больше, чем концентрация свободного водорода. Таким образом, можно предположить, что в периоды ослабления вулканической активности, когда, в частности, водород в удобной для усвоения биотой того времени форме (молекулярный водород, сероводород, метан и т.д.) переставал поступать в ощутимых количествах в мировой океан и атмосферу, как раз и складывались условия, необходимые для переключения цианобактерий с аноксигенного на оксигенный фотосинтез. Соответственно, к рубежу 3.2 млрд. лет, когда геологическая активность Земли упала до локального минимума, что совместно с резким уменьшением количества сулфаторедуцантов существенно уменьшило концентрацию в окружающей среде свободного водорода и сероводорода, мы и фиксируем первые достаточно явные сигналы оксигенного фотосинтеза (де факто, переключаясь на оксигенный фотосинтез, цианобактерии спасали от вымирания не только себя, но и всю земную биоту, успевшую уже основательно "подсесть" на водород!). До этого времени он мог быть попросту энергетически невыгоден, и если и включался, то лишь эпизодически. Именно это могло быть одной из причин, почему многие сотни миллионов лет свободный кислород не накапливался в атмосфере и океанических водах. Таким образом, тезис о наличии на Земле уже 3.4-3.5 млрд. лет назад цианобактерий представляется не столь уж абсурдным, хотя он до сих пор и оспаривается многими учёными, особенно, не являющимися практикующими специалистами в области палеонтологии бактерий.
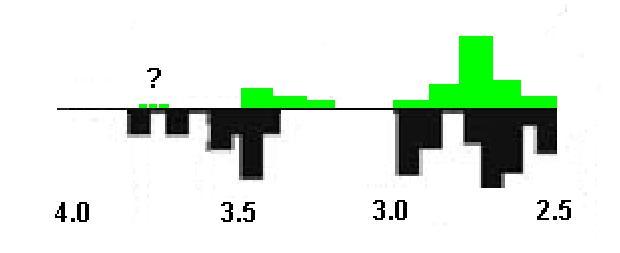
Рис. 2. Мощность пород вулканического происхождения (чёрные прямоугольники ниже горизонтальной оси) и количество обнаруженных в слоях соответствующего возраста строматолитов (зелёные прямоугольники выше горизонтальной оси). Числа внизу соответствуют изменению возраста пород вдоль оси координат. Пунктиром обозначены возможные гипотетические бактериальные маты, соответствующие породам с "облегчённым" углеродом возрастом 3.7-3.8 млрд. лет. Данные по магматическим породам взяты отсюда, данные по строматолитам из обзора Шопфа.
Итак, в сухом остатке вырисовывается примерно следующая картина. 3.4-3.5 млрд. лет назад в районах повышенной геологической активности, например, около подземных вулканов и у геотермальных источников существовали бактериальные сообщества, напоминающие современные цианобактериальные маты. Эти сообщества активно использовали солнечные лучи и исходящие из недр Земли и находящиеся в далёком от состояния термодинамического равновесия вещества, что бы извлекать энергию, встраиваясь в циклы химических реакций, идущих в направлении установления термодинамического равновесия (то бишь, в направлении увеличения энтропии). Из магматических же источников они получали многие необходимые им химические макро и микроэлементы. В самом верхнем слое, располагавшемся вблизи поверхности воды, обитали цианобактерии, и, возможно, некоторые другие фототрофные бактерии. Они могли черпать энергию, необходимую для своего метаболизма, непосредственно от Солнца. В более нижних слоях, по видимому, располагались сульфаторедуцирующие бактерии. Не исключено, что там же могли присутствовать и ацетогены. Наконец, исходя из логических рассуждений, в самом нижнем слое, по аналогии с современными бактериальными матами, можно было бы ожидать преобладания перерабатывающих оседающую сверху органику и выделяющих водород бактерий-бродильщиков. Для матов, располагающихся вблизи подводных вулканов и выхода на поверхность разогретых до температуры 100С и более струй подземных вод, не исключено так же существование термофильных архей-метаногенов. Другими словами, уже 3.4-3.5 млрд. лет назад Земля, судя по всему, была заселена сложными бактериальными сообществами включая такие их высокоорганизованные типы, как цианобактериальные маты. Относительно существования жизни на Земле в ещё более древние времена, отстоящие от нас на 3.7-3.8 млрд. лет в настоящее время пока нет достаточных оснований ни для того, что бы уверенно принять, ни для того, что бы уверенно отвергнуть такую возможность. Имеются как факты, косвенно подтверждающие существование в то время бактериальной жизни (обнаружение облегчённого углерода в одной из осадочных пород, сформировавшейся в этот период, формирование в этот же период первых джеспилитов, наличие признаков геологической активности, обычно способствующей расцвету бактериальных матов того времени), так и заставляющие в этом усомниться (отсутствие отпечатков бактерий и строматолитов, возможное объяснение появления следов облегчённого углерода более поздними загрязнениями).