Сначала разберёмся, какие молекулярные механизмы обеспечивают вырожденность ГК.
Вспомним, что кодон матричной РНК спаривается с антикодоном тРНК (первое основание кодона - с третьим основанием антикодона, или 36-м нуклеотидом на тРНК, второе - со вторым (35-м), третье - с первым (34-м)), как это схематически изображено на рис. 1.
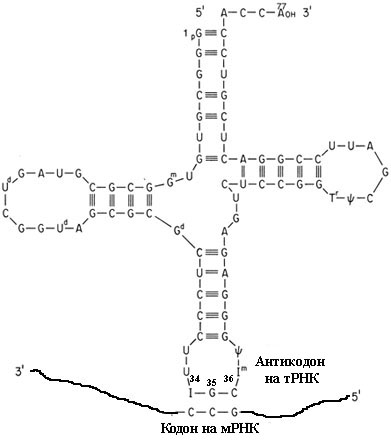
Рис. 1. Взаимодействие кодона и антикодона. На рисунке показано, что третий нуклеотид кодона взаимодействует с неканоническим основанием инозином (I). Цифры возле нуклеотидов антикодона соответтвуют стандартной общепринятой нумерации нуклеотидов на тРНК (см. рис. 1 в Транспортные РНК как молекулярные реликты)
Всего существуют 61смысловой кодон, однако в ГК эти кодоны читаются гораздо меньшим числом антикодонов на тРНК. И может быть не менее 20 (количество аминокислот), не более 61 (количество смысловых кодонов). Количество различных тРНК, читающих все аминокислоты, варьирует у разных организмов.
У бактерий обычно около 43 тРНК. У человека около 50 различных тРНК обеспечивают включение аминокислот в белок. Это объясняется тем, что:
Для считывания разных кодонов мРНК, соответствующих одной и той же аминокислоте, используются изоакцепторные тРНК с разными антикодонами. Для многих аминокислот число соответствующих им изоакцепторных тРНК с разными антикодонами гораздо меньше, чем общее число кодонов (например, 24 митохондриальных тРНК достаточно для узнавания 61-62 смысловых кодонов мРНК). Из этого следует, что одна и та же тРНК может узнавать несколько кодонов, кодирующих одну и ту же аминокислоту, но различающихся по одному нуклеотиду.
Правила, согласно которым происходит соответствие кодон-антикодон, суммированы в гипотезе неоднозначного соответствия (гипотеза качаний, Wobble-гипетеза). Эта гипотеза постулирует, что образование пары кодон-антикодон в двух первых положениях кодона происходит всегда по каноническим правилам, но в третьем положении возможно "качание" (неоднозначное соответствие). Объясняется это тем, что конформация антикодоновой петли тРНК допускает значительную подвижность первого основания антикодона. Некоторые пары оснований, предусмотренные гипотезой неоднозначного соответствия, приведены на рис. 2.
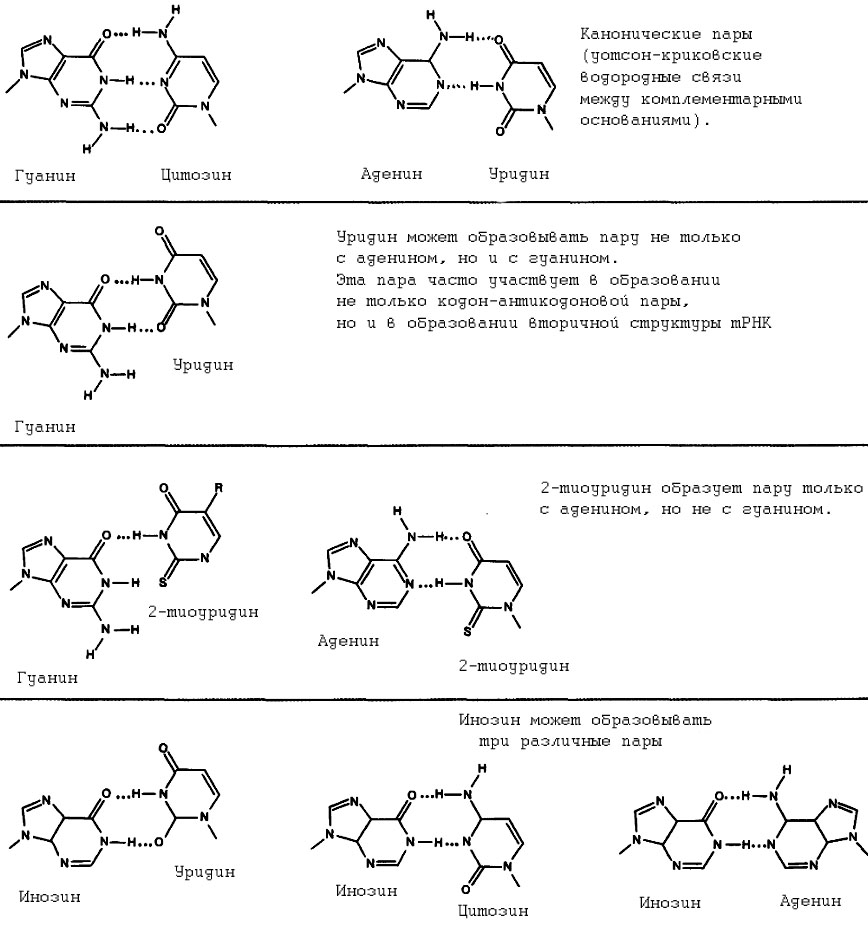
Мы видим, в частности, что U в антикодоне узнаёт как А, так и G в кодоне; С в антикодоне узнаёт только G в кодоне; А в антикодоне узнаёт только U в кодоне; G в антикодоне узнаёт С или U в кодоне. Данный факт накладывает ограничения на множество возможных кодонов с вырожденностью 1: однозначно узнавать уникальные кодоны возможно тогда, когда третьим основанием является G или U. К таким кодонам относятся UGG и AUG, хотя аналогичных кодонов в случае U не существует. Дело в том, что в действительности некоторые варианты пар в первом положении антикодона (соответствует третьему положению кодона) некоторые обычные ванианты обычно не встречаются в силу того, что ряд оснований всегда модифицируется. В частности, в первом положении антикодона не используются U и A, что сужает круг уникальных кодонов (с вырожденностью 1) до реально существующих в природе UGG и AUG. В первом положении антикодона А никогда не обнаруживается, так как обычно он превращается в инозин (I), способный при этом образовывать пары с тремя основаниями - U,C и A. Это особенно важно для изолейциновых кодонов, когда AUA кодирует изолейцин, не смотря на то, что AUG кодирует метионин. Когда А находится в третьем положении кодона, при помощи обычных оснований невозможно образовать однозначную пару, так как любая тРНК, у которой имеется в антикодоне U будет узнавать AUG так же, как и AUA. Проблема может быть решена только благодаря использованию инозина в первом положении антикодона. Ограничение, не достигаемое при использовании обычных правил, возможно при наличии в антикодоне 2-тиоуридина (см. рис.2). Эта модификация эффективно узнаёт А, но в отличие от U не узнаёт G. Модифицированные нуклеотиды по-видимому существенно увеличивают точность считывания кодонов в аппарате трансляции. Однако, как увидим в далее, в ряде сильно редуцированных геномов митохондрий и других прокариот далеко не всегда используются модифицированные основания и набор тРНК в таких организмах представлен минимально возможным числом по одной на каждую аминокислоту. В этом случае можно говорить, что взаимодействие первого антикодонового основания с третьим кодоновым вообще не играет никакой роли в случае сильных основ кодонов. Можно предположить, что этот менее точный, но более простой механизм взаимодейстия является более древним по сравнению с механизмом, обеспечиваемым модифицированными основаниями. Рассмотрение этого механизма в следующей приведёт нас к неизбежному выполнению симметрических преобразований Румера.
Рис. 2. Неоднозначное соответствие при образовании кодон-антикодоновой пары позволяет некоторым основаниям в первом положении антикодона узнавать более чем одно основание в третьем положении кодона.