Как профессор Савельев имитирует научную деятельность: разбор статей.
Как профессор Савельев имитирует научную деятельность: разбор статей.
https://g-equality.livejournal.com/10480.html
Автор статьи — Константин Лесков, Ph.D., Case Western Reserve University, Cleveland, OH.
Статья отредактирована членами команды EQUALITY: спасибо Анне Курановой, врачу-исследователю из Междисциплинарного Центра Психопатологии и Эмоционального Регулирования в Медицинском Центре Университета Гронингена.
Некоторое время назад мне стали попадаться интервью с Сергеем Вячеславовичем Савельевым, профессором, заведующим лабораторией развития нервной системы Института морфологии человека ФАНО. Одно из них было дано газете «Комсомольская Правда» от 22 июля 2010 года. В нем Савельев повествует о различиях в массе мозга между жителями разных стран, а также между мужчинами и женщинами, и о том, какие выводы из этих различий, по его мнению, следуют.
В то время о Савельеве мне было неизвестно абсолютно ничего. Никакие его работы по морфологии мозга или нейрофизиологии на семинарах не звучали. В интервью “КП” насторожили некоторые странности с тем, как Савельев подает факты и интерпретирует их. Впрочем, это я отнес на счет необходимости интервьюируемого общаться с журналистами.
Несколько лет спустя увидел Савельева на канале Дмитрия Пучкова oper.ru. Пучков публикует очень интересные интервью с историками, но биологов приглашает крайне редко, поэтому начал слушать с интересом. Однако по мере просмотра ролика стало нарастать ощущение, что в студии происходит что-то не то. Названный профессор нес редкостную чепуху из наукообразных терминов, «дней минувших анекдотов» и кухонных стереотипов вперемешку с обвинениями в адрес генетиков и молекулярных биологов в растрате государственных средств.
Ничего относящегося к нейрофизиологии, морфологии ЦНС или науке вообще в беседе с Савельевым не было. Странной казалась позиция Пучкова, который в своих «разведопросах» с историками обличал лженауку в лице Фоменко и Задорнова, здесь же открыто поддерживал Савельева и настоятельно рекомендовал слушателям его произведения с названиями типа «Церебральный сортинг».
Поскольку по ролику в YouTube не всегда можно судить о профессионализме научного работника, я разыскал публикации Савельева в рецензируемых изданиях. В большинстве своем они оказались русскоязычными. Поскольку Савельев утверждает, что он специалист по морфологии мозга и эмбриологии, я выбрал одну из его недавних работ, которая, судя по названию, имеет отношение к обеим областям: «Патология эмбрионального морфогенеза головного мозга человека» 1.
Начнем с абстракта. В нем вкратце должно сообщаться, что нам следует ожидать в статье: какая гипотеза, методы ее тестирования, результаты экспериментов и их интерпретация в соответствии с общепринятой научной методологией.
В кратком содержании Савельев заявляет, что в данной работе он произвел экспериментальные исследования и получил следующие результаты (курсивом выделен текст Савелева, нумерация моя, К. Л.):
1. Исследованы механизмы развития ранней эмбриональной патологии нервной системы у человека и экспериментальных животных.
Таким образом, ожидается, что автор предоставит в статье экспериментальные данные, включая материалы, методы и результаты, полученные на неких животных и человеческих модельных системах.
2. Установлены биомеханические принципы кодирования позиционной информации нейроэпителиальными клетками.
Раскрытие механизма «кодирования позиционной информации нейроэпителиальными клетками» явилось бы серьезным прорывом в биологии развития. Поэтому с нетерпением ждем экспериментальных данных Савельева, описывающих носителей этого «кода», его свойства, механизмы считывания и преобразования закодированной информации в реальные биологические структуры.
3. Экспериментальное обратимое изменение тангенциальных межклеточных взаимодействий в нейроэпителии животных приводило к прогнозируемым аномалиям развития нервной системы. Изменение проницаемости механозависимых ионных каналов является основным способом считывания позиционной информации и детерминации клеточной дифференцировки. Было проведено сравнение экспериментальных аномалий мозга у животных с уникальными аутопсиями патологического развития мозга человека, начиная от нейруляции и заканчивая 6-м мес внутриутробного развития.
Таким образом, ожидается, что Савельев, обладая соответствующей аппаратурой и методами, провел эксперименты по а) количественному измерению и б) контролированному изменению «тангенциальных межклеточных взаимодействий в нейроэпителии животных».
Согласно заявлению Савельева, он способен прогнозировать и воспроизводить аномалии развития нервной системы, направленно изменяя «тангенциальные межклеточные взаимодействия» (надеемся, что Савельев также даст определение, что же такое «тангенциальные межклеточные взаимодействия», какие белки их опосредуют, какими методами Савельев эти взаимодействия измеряет и изменяет).
На основе результатов экспериментов, описание которых мы ожидаем увидеть в статье, Савельев делает вывод, что проводимость механозависимых ионных каналов меняется вследствие механических взаимодействий между клетками. По утверждению Савельева, проницаемость механозависимых ионных каналов – это и есть механизм считывания «позиционной информации» (см. выше), которая преобразуется в дифференцировку клетки-предшественника.
Следует ожидать, что в статье автор предоставит экспериментальные данные по изменению проводимости механозависимых ионных каналов с количественным измерением этой проводимости с помощью соответствующего оборудования. Также следует ожидать, что автор путем контролируемого манипулирования механическими натяжениями в развивающемся эмбрионе продемонстрирует изменение проводимости ионных каналов, изменение дифференцировки клеток и экспериментально воспроизведет аномалии мозга на модельных организмах. Ожидается, что эти аномалии, согласно заявлению Савельева, будут аналогичны известным патологиям развития человеческого мозга.
4. Полученные результаты позволили установить, что при задержке движения постнейропорной волны замыкания нервной трубки формируются аномалии переднего мозга и этмоидной части лица. Обнаружено, что при остановке движения постнейропорной волны нейруляции формируются аномалии промежуточного и среднего мозга, а при нарушении каудальной волны — аномалии заднего и спинного мозга. Показано, что аномалии развития, диагностируемые на поздних сроках онтогенеза и после рождения, являются последствиями компенсаторных реакций нейроэпителия на изменение ранних позиционных сигналов в период формирования нервной трубки.
Здесь мы сталкиваемся с нетрадиционной терминологией Савельева. Под «постнейропорной волной замыкания нервной трубки», скорее всего, имеется в виду нейруляция – процесс замыкания нервной пластинки в нервную трубку, который у человека начинается в двух местах: посередине и в головной области эмбриона – и распространяется в антериальной области встречными волнами с образованием антериального нейропора (отверстия), а в каудальной (хвостовой) с образованием постериального нейропора. Ознакомление с предметом можно начать с обзора Copp, 2013 2.
Судя по его утверждению, Савельев экспериментально индуцировал задержку нейруляции и смоделировал «аномалии переднего мозга и этмоидной части лица». Интересно, что даже термины Савельев подбирает не слишком корректно. Взять ту же «этмоидную часть лица» - не сразу понятно, что вообще имеется в виду. Есть этмоидная кость, расположенная за носовой костью, и ассоциированный с ней этмоидный хрящ. Они входят в состав лицевого скелета. Судя по содержанию статьи, Савельев имеет в виду циклопию, при которой могут отсутствовать или быть редуцированы носовая и этмоидная кости.
Под «поздними сроками онтогенеза», скорее всего, имеются в виду поздние сроки эмбриогенеза или эмбрионального онтогенеза. Согласно определению Геккеля, онтогенез – это развитие организма от зачатия до конца жизни. Если следовать этому определению, то Савельев, очевидно, изучал половозрелых людей преклонного возраста, на поздних сроках их онтогенеза. Впрочем, на фоне остального это незначительное терминологическое замечание.
Из последнего предложения в абстракте следует, что Савельев экспериментально «изменил ранние позиционные сигналы», что привело к детектируемым «компенсаторным реакциям нейроэпителия» и, как следствие, к «аномалиям развития».
Пара слов об английской версии абстракта. Очевидно, что сам Савельев не владеет в достаточной степени языком и ему следовало воспользоваться услугами профессионального переводчика. В этом случае он избежал бы курьезных фраз, вроде: «The above anomalies resulted from local compensatory responses of the neuroepithelium due to the lack of intentions [sic!] that are characteristic of normal development of the neural tube».
В переводе это выглядит примерно так: «Аномалии выше были результатом локального компенсаторного ответа нейроэпителия из-за недостатка намерений (!) которые характеризуют нормальное развитие нервной трубки.»
Подведем итоги по абстракту. Савельев делает очень серьезные заявления о множестве собственных фундаментальных открытий в одной публикации. Такие заявления должны быть подкреплены результатами большого объёма очень хорошо спланированной экспериментальной работы. Посмотрим, что же на самом деле содержится в статье.
Введение
Обычно введение содержит постановку проблемы, обзор текущей литературы по данной теме, гипотезу и стратегию ее тестирования.
В качестве проблемы Савельев называет «необходимость поиска наиболее уязвимого и универсального механизма возникновения ранних отклонений в развитии нервной системы, которые обусловлены нарушением морфогенетических процессов и крайне плохо изучены».
Цель работы Савельев определяет как «поиск ранних эмбриональных межклеточных взаимодействий, детерминирующих формообразование головного мозга, нарушение которых вызывает необратимые аномалии развития человека». Иными словами, рабочая гипотеза автора состоит в том, что, нарушив каким-то образом межклеточные взаимодействия в эмбрионе на ранней стадии, он способен экспериментально воспроизвести некие неоговоренные «аномалии развития».
В качестве стратегии тестирования своей гипотезы Савельев называет «два параллельных подхода: изучение клинических случаев ранних эмбриональных патологий нервной системы человека и экспериментально-эмбриологический подход по моделированию наиболее распространенных аномалий развития.»
Таким образом, если Савельев стремится быть последовательным и отталкивается от своей гипотетической модели о роли межклеточных взаимодействий в морфогенезе мозга, нам следует ожидать, что в данной статье Савельев опишет 1) исследование белков, отвечающих за межклеточные взаимодействия, на клиническом материале и 2) экспериментальное изменение этих межклеточных взаимодействий с целью смоделировать некие неуказанные, но известные аномалии развития мозга.
Савельев не считает нужным указать модельную систему для второго подхода. Очевидно, это должно быть какое-то лабораторное животное. Общепринятой практикой считается указание модельного организма если не в заголовке, то хотя бы в абстракте. Савельев пренебрегает этим правилом.
Сразу настораживает крайняя расплывчатость проблемы и гипотезы. Когда речь идет о работе в области любой патологии, название патологии или группы патологий должно четко озвучиваться в виде диагноза в соответствии с международной классификацией заболеваний (ICD-10), которая используется в Российской Федерации как члене Международной Организации Здравоохранения (WHO). Врожденные аномалии нервной системы, которые, судя по всему, имеет в виду Савельев, включают около сотни диагностируемых патологий и их вариантов, которые по ICD-10 классифицируются статьями Q00-Q07. Отсутствие ссылки на конкретную группу кодифицированных диагнозов указывает на непрофессионализм Савельева.
Не приводя ссылок на соответствующие экспериментальные работы, Савельев утверждает, что «несмотря на разнообразие причин патологии развития, структурные изменения зародышей носят сходный характер. Однотипные морфогенетические реакции клеток ранних зародышей человека говорят о существовании универсальных механизмов рецепции различных типов патогенетических воздействий». Что за «однотипные морфогенетические реакции», в чем заключается «сходный характер» изменений зародыша и относится ли это ко всей сотне патологий из ICD-10-Q00-Q07, Савельев не удосуживается сообщить.
Савельев пишет: «К примеру, было неоднократно описано большое разнообразие аномалий при одном генетическом дефекте и одинаковые аномалии при изменении различных хромосом [1–4].» В профессиональной литературе это называют плейотропным действием гена в первом случае и мультигенным наследованием признака – во втором. Интерес представляет подбор ссылок, следующих за этим утверждением. Первой ссылкой Савельев цитирует свою собственную статью:
С. В. Савельев. Нейруляционные формы патологии нервной системы человека. // Архив патологии. — 1998. — №5. — С. 13—18.
При прочтении этой статьи оказывается, что никаких генетических исследований Савельев не провел. То есть ссылка была приведена некорректно. Вместо этого автор утверждает, что для «изучения аномалий развития головного мозга у эмбрионов в возрасте от 3 до 6 нед. и выявления общих принципов патогенеза этих аномалий мозга» он «исследовал» 47 абортивных эмбрионов.
Без проведения каких-либо генетических тестов, не приводя никаких сведений из литературы, Савельев утверждает, что «корреляция между сомитами, числом позвонков и ганглиями является довольно точным критерием наличия или отсутствия генетических изменений у эмбриона. Если в цервикальном, торакальном, люмбальном, крестцовом или копчиковом отделе соотношение между осевыми структурами нарушается, то наиболее вероятно наличие геномных изменений у зародыша. Таких эмбрионов не использовали». На чем основано убеждение Савельева, что генетические изменения отсутствуют у эмбрионов, взятых им для «исследования», абсолютно непонятно, поскольку ни кариотипирование, ни более глубокое генотипирование проведено не было.
Далее Савельев пишет: «Для гистологических исследований применяли фиксаторы Буэна, Карнуа и Ценкера, 10% формальдегид. Из блоков готовили серийные срезы толщиной от 5 до 15 мкм, их монтировали на стекла, окрашивали гематоксилином и эозином, по Массону, Бионди, заключали в бальзам. Срезы использовали для создания трехмерных графических реконструкций». В данном случае имеет место серьезное нарушение методологии, заключающееся в некорректном сравнении препаратов, обработанных произвольно разными методами. На чем основывался выбор фиксатора, не указывается. Какие эмбрионы фиксировались каким фиксатором и как это могло отразиться на результатах исследования, Савельев не считает нужным указывать.
То же самое относится к окраске препаратов. Статья не содержит фотографий самих препаратов, лишь некие «трехмерные графические реконструкции» крайне низкого разрешения, сомнительного качества и, как следствие, абсолютно неинформативные. Почему Савельев не публикует фотографии гистологических срезов, как это принято в работах в области эмбриологии, остается непонятным.
Таким образом, статья Савельева 1998 г. является субстандартной и низкокачественной. Самоцитирование ее в статье 2012 г. очень похоже на наивную попытку поднять свой индекс.
Вторая ссылка относит нас в 1957 год:
Hugehes A. Development of the primary sensory system in Xenopus laevis (Daudin). J. Anat. 1957; 91: 323–328.
Как и в первом случае, статья не содержит никаких генетических исследований. Это работа по развитию органов чувств у шпорцевой лягушки. Никаких исследований аномалий развития мозга в статье не проводится. Почему Савельев цитирует ее в связи с генетическими патологиями у человека – абсолютно непонятно.
Следующая цитируемая Савельевым статья – это исследование нарушений ЦНС у куриц после кормления их органофосфатами:
Tanaka D.J., Bursian S.J. Degеneration patterns in the chicken central nervous system induced by ingestion of the organophosphorus delayed neurotoxin 3-orthototolylphosphate. A silver impregnation study. Brain Res. 1989; 484: 240–256.
По ее прочтении оказывается, что авторы работали не с эмбрионами, а с 8-месячными курицами породы белый леггорн. После кормления их 3-орто-толилфосфатом, куриц забивали в течение 4 недель и исследовали дегенерацию нейронов разных участков мозга. Ни эмбриологических, ни генетических исследований не проводилось.
То, что Савельев цитирует этo в связи с генетическими нарушениями морфогенеза головного мозга человека, говорит о том, что статью он не читал. Цитирует он ее лишь «для вида и объема», что является очевидной фикцией. Савельев уподобляется нерадивому студенту, который кидает какие попало ссылки в свой реферат в надежде, что преподаватель не обратит на них внимание. Это свидетельствует о серьезном непрофессионализме Савельева.
Четвертая ссылка: Simon E.M., Barkovich A.J. Holoprosencephaly: new concepts. Magn. Reson. Imaging. Clin. N. Am. 2001; 9 (1): 149–164. Эта работа опубликована в издании, ориентированном на использовании ядерного магнитного резонанса в клинических целях. Генетические исследования в статье не приводятся. Работа посвящена исключительно ЯМР. Один из разделов в конце статьи упоминает известные на тот момент работы молекулярных генетиков о роли мутаций SHH ZIC2 и SIX3 в развитии голопрозэнцефалии, однако по какой-то причине Савельев цитирует эту вторичную работу, а не оригинальные. Как увидим позднее, начиная с конца 1990-х годов, появляется всплеск экспериментальной литературы по этиологии голопрозэнцефалии, которые Савельев не цитирует вовсе, что указывает на то, что он с ней скорее всего не знаком.
Далее в введении Савельев уделяет много внимания работе Чарльза Стокарда 1921 года. Это одна из ранних работ по тератогенезу. За прошедшие 90+ лет появились более разработанные модели морфогенетических патологий. Исследования в области эмбриональной молекулярной генетики в последние 15 лет до написания Савельевым его статьи привели к тому, что работы Стокарда в настоящее время представляют лишь исторический интерес.
Читаем дальше. Савельев пишет: «По предварительным данным, реальная гибель зародышей человека в первые 10–14 дней после оплодотворения может составлять до 80%. Столь высокий уровень гибели обусловлен морфогенетической уязвимостью мигрирующего зародыша, нарушением имплантационных процессов и механизмов формообразования [6–8].»
Три приведенные ссылки – снова цитирование Савельевым собственных статей сомнительного содержания и научной ценности, не содержащих его собственных исследований по статистике преклинической мортальности человеческих эмбрионов. По оценкам Optiz, 19873, потери до первых пропущенных месячных, включая неделящиеся яйцеклетки, пре-имплантационные проблемы и отсутствие имплантации составляют около 40-55% от всех зачатий.
Далее Савельев повторяет описание «эмбриона темякопчиковой длиной 3,2 мм на 22–23-й день после оплодотворения» из своей публикации 1993 года. Этот эмбрион якобы имел доселе не описанную аномалию: «Аномалия возникла к концу нейруляции. На это указывает эктодерма, расположенная над дорсальной поверхностью головного и спинного мозга. Пластовое строение мозга этого эмбриона утрачено, а спинной мозг заканчивается на уровне брюшного стебелька. Спинной мозг не достигал своего нормального размера и был на треть короче, чем в норме».
Проблема в том, что у человека на 22-23-ий день нейруляция только начинается, и закрывается нервная трубка только к 24-му дню (ростральный нейропор) и 26-28-му дню (каудальный нейропор)4–6. Савельев показывает свою полную безграмотность в эмбриологии нервной системы.
Учитывая неопределенность с возрастом эмбриона (или просто ошибку Савельева с его определением), эмбрион мог быть фиксирован либо в самом начале нейруляции, либо в середине процесса. Делать какие-либо заключения о строении головного и спинного мозга в этот период бессмысленно. Поскольку никто, кроме Савельева, этого эмбриона не видел, сложно сказать, действительно ли имела место какая-либо аномалия, или, что более вероятно, Савельев выдал самую раннюю стадию нормальной нейруляции за придуманный им дефект.
О том же самом эмбрионе Савельев рассуждает: «Подобные реакции нейроэпителия наблюдали в экспериментах по сбросу осевых механических напряжений у амфибий [12–14]. Окклюзия нервной трубки и деструктурализация пластов характерны для релаксации механически ненапряженного нейроэпителия».
Здесь Савельев демонстрирует взгляды профессора кафедры эмбриологии биологического факультета МГУ Льва Белоусова. Белоусов и Савельев считают, что при морфогенезе главную роль играют «механические напряжения» клеточных пластов. Ни в классической работе Nusslein-Volhard, 19887о роли градиента экспрессии bicoid в морфогенезе D. Melanogaster, ни в последующем всплеске работ, исследовавших регуляцию морфогенеза градиентами различных мРНК и белков, ведущая роль «механических напряжений» в морфогенезе не рассматривается. Тем не менее, Савельев полностью игнорирует несколько тысяч статей по исследованиям роли градиентов морфогенных белков в период 1988 – 2012 гг. Судя по отсутствию цитат этих работ в его статьях, Савельев скорее всего не знаком с текущей литературой по молекулярным основам морфогенеза.
Давайте посмотрим, как Белоусов и Савельев тестируют эту гипотезу. Когда речь идет о таком физическом явлении как механическое напряжение, необходимо четко определить, какая конкретно величина имеется в виду, какова ее размерность? Какими методами ее измеряют и изменяют? Наконец, что очень важно, создается ли в реальном эмбрионе натяжение, соизмеримое с тем, которое авторы искусственно создают в эксперименте?
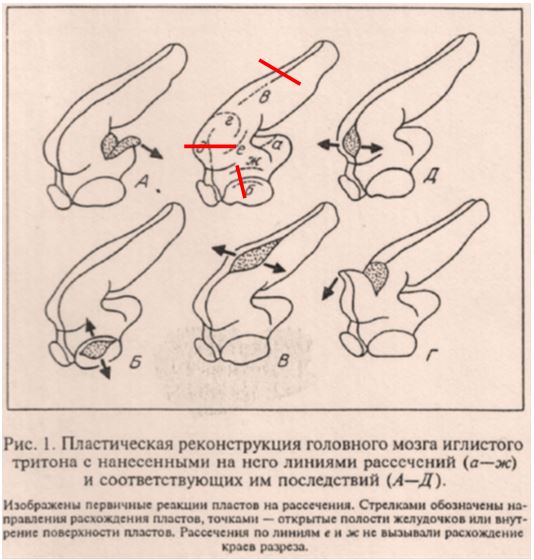
Рис.1. Иллюстрация из статьи Савельева9. Красным отмечены линии сечений, которые полагалось сделать в качестве минимального контроля. В целом, замысел эксперимента на уровне юного натуралиста. Ни фотографий, ни количественных данных Савельев не приводит. В тексте называет изображенное «эмбриональным мозгом человека».
Если мы имеем дело с классическим механическим напряжением, то это величина тензорная. Трехмерная, развивающаяся во времени система клеток эмбриона предполагает, что изменение ее геометрии описывается 4-тензором. Это в самой упрощенной модели, которая не учитывает, что клетки соприкасаются не мембранами, а фокальными и щелевыми контактами. Локальные механические взаимодействия между клетками не ограничены «тургором», а включают в себя активность цитоскелета, которую тоже следует учитывать в математической модели.
Знакомство с публикациями Белоусова и Савельева8 показывает, что авторы никогда не производили измерений «механических напряжений» и не указывали размерность этой величины. Никаких соответствующих расчетов и экспериментальных измерений в вышеуказанных публикациях Савельева и Белоусова не приводится. Вместо этого авторы использовали весьма странный метод углов между соседними клетками на плоском срезе. Соответствуют ли найденные ими этим методом величины реальным напряжениям в трехмерном пласте клеток авторы не комментируют, просто потому что неспособны это сделать.
Примечательно, что в тексте статьи Савельев пишет: «… обратимся к результатам простых опытов по сбросу механических натяжений в эмбриональном мозге человека при помощи микрорассечений (рис 1)». На рисунке же изображен мозг иглистого тритона, а вовсе не человека. Никаких рассечений человеческого мозга в статье не приводится. Прочие эксперименты, описанные в статье Савельева 2001 года, также не соответствуют принятым на то время методологическим стандартам. Например, использование рисунка вместо фотографии (рис. 2, Савельев, 2001) в качестве экспериментального результата неприемлемо.
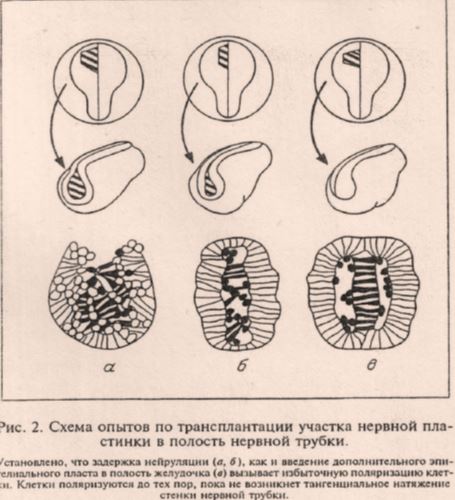
Рис. 2. «Схема опытов» Савельева по трансплантации нервной пластинки в нервную трубку с неясными целями и отсутствием каких-либо результатов в виде фотографий9. Савельев не предоставляет данные о морфологии, т.н. «поляризованных» клеток, ни тем более об их молекулярных маркерах. Интересно, что с начала 1990-х годов исследователи широко используют методику микроинъекции очищенных морфогенных мРНК и белков для оценки их влияния на нейруляцию и другие морфогенетические процессы41–45. Савельев явно незнаком с этими работами и не удосуживается тестировать транспланты на известные на момент написания статьи индукторы нейрогенеза, такие как Noggin, Follistatin, Chordin, Cerberus, and Xnr3.
На рис. 3 отсутствует какой-либо статистический анализ и элементарное описание наблюдаемых «аномалий».
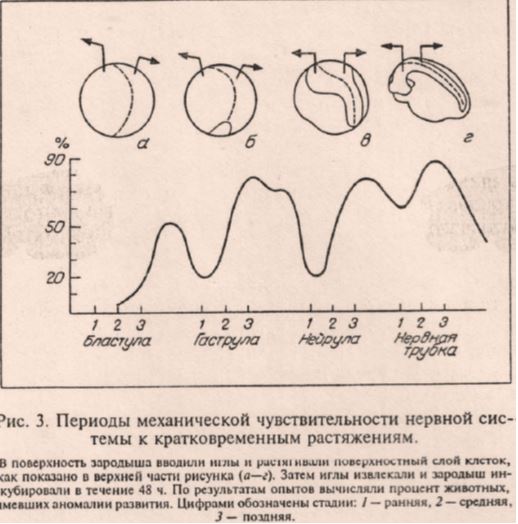
Рис.3. Типичная иллюстрация из статьи Савельева9. С отсутствием статистического анализа.
Фотографии эмбрионов на рис. 4 абсолютно неинформативны и низкого качества. Когда хотят продемонстрировать какую-либо патологию, минимальное требование – хотя бы сделать гистологические срезы аномального участка эмбриона.
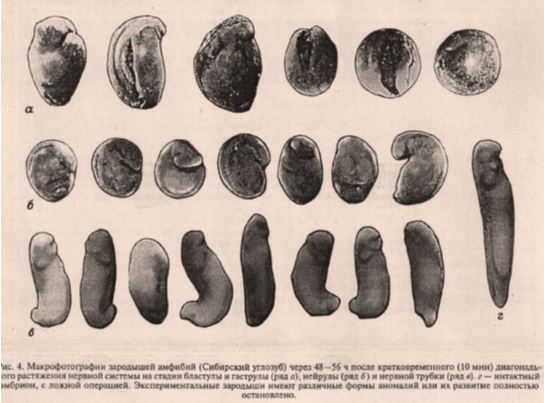
Рис.4. Савельев9демонстрирует «аномалии» развития после «диагонального растяжения нервной системы (sic!) на стадии бластулы и гаструлы ... нейрулы ... и нервной трубки.» Не очень понятно, где Савельев нашел нервную систему на стадии бластулы и гаструлы, чтобы ее «диагонально» растянуть. Отсутствует описание т.н. «аномалий», критериев, по которым эти аномалии определялись и отбирались, а также их гистологическое исследование и гистохимический анализ экспрессии известных морфогенов20,23,41–45и механосенсорных ионных каналов, на функции которых Савельев строит свою гипотезу. Возникает серьезный вопрос о физиологичности растяжения: генерируется ли такой стресс во время нормального эмбриогенеза или Савельев просто демонстрирует эмбрионы, погибшие от неспецифического механического воздействия. Отсутствуют контрольные эмбрионы, растянутые в других направлениях.
На рис. 5 непонятно, что конкретно Савельев измеряет по оси ординат в графике о содержании ионов в цитоплазме. Важный вопрос, возникающий при рассмотрении этого графика: не является ли изменение ионов хлора в цитоплазме.
Интересный комментарий дал старший научный сотрудник кафедры эмбриологии биологического факультета МГУ Станислав Кремнёв:
"Несмотря на наличие С.В. Савельева в списке авторов статьи Beloussov LV, Saveliev SV, Naumidi II, Novoselov VV. Mechanical Stresses in Embryonic Tissues: Patterns, Morphogenetic Role, and Involvement in Regulatory Feedback. 1994 p. 1–34, Савельев не участвовал в этой работе и не имеет отношение к экспериментам по выявлению роли механических напряжений в морфогенезе. Савельев также не внес никакого вклада в построение карт механических напряжений в эмбрионах (Beloussov, L. V., Dorfman, J. G., and Cherdantzev, V. G. (1975))"
Литература
1. Saveliev S V. Патология эмбрионального морфогенеза головного мозга человека. Вестник РАМН. 2012;8:40–46. http://vestnikramn.spr-journal.ru/jour/article/download/276/214
2. Copp AJ, Greene NDE. Neural tube defects--disorders of neurulation and related embryonic processes. Wiley Interdiscip Rev Dev Biol [Internet]. Europe PMC Funders; 2013 [cited 2017 Apr 5];2(2):213–27. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24009034 PMID: 24009034
3. Opitz JM. The Farber Lecture. Prenatal and Perinatal Death. Pediatr Pathol [Internet]. 1987 Jan 9 [cited 2017 Apr 29];7(4):363–392. Available from: http://www.tandfonline.com/doi/full/10.3109/15513818709161402
4. O’Rahilly R, Muller F. Neurulation in the normal human embryo. Ciba Found Symp [Internet]. 1994 [cited 2017 Apr 29];181:70-82–9. Available from: http://www.ncbi.nlm.nih.gov/pubmed/8005032 PMID: 8005032
5. O’Rahilly R, Muller F. The development of the neural crest in the human. J Anat [Internet]. Wiley-Blackwell; 2007 Sep [cited 2017 Apr 29];211(3):335–51. Available from: http://www.ncbi.nlm.nih.gov/pubmed/17848161 PMID: 17848161
6. Van Allen MI, Kalousek DK, Chernoff GF, Juriloff D, Harris M, McGillivray BC, Yong S-L, Langlois S, Macleod PM, Chitayat D, Friedman JM, Wilson RD, McFadden D, Pantzar J, Ritchie S, Hall JG. Evidence for multi-site closure of the neural tube in humans. Am J Med Genet [Internet]. 1993 Oct 1 [cited 2017 Apr 29];47(5):723–743. Available from: http://doi.wiley.com/10.1002/ajmg.1320470528
7. Berleth T, Burri M, Thoma G, Bopp D, Richstein S, Frigerio G, Noll M, Nusslein-Volhard C. The role of localization of bicoid RNA in organizing the anterior pattern of the Drosophila embryo. EMBO J [Internet]. European Molecular Biology Organization; 1988 Jun [cited 2017 Apr 29];7(6):1749–56. Available from: http://www.ncbi.nlm.nih.gov/pubmed/2901954 PMID: 2901954
8. Beloussov LV, Saveliev SV, Naumidi II, Novoselov VV. Mechanical Stresses in Embryonic Tissues: Patterns, Morphogenetic Role, and Involvement in Regulatory Feedback. 1994 [cited 2017 Apr 30]. p. 1–34. Available from: http://linkinghub.elsevier.com/retrieve/pii/S0074769608615351
9. Савельев СВ. Механизм кодирования позиционной информации в эмбриональном формообразовании мозга позвоночных. Вестник РАМН. 2001;4:49–61.
10. Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang D-S, Woolf CJ, Corey DP. TRPA1 Contributes to Cold, Mechanical, and Chemical Nociception but Is Not Essential for Hair-Cell Transduction. Neuron [Internet]. 2006 [cited 2017 May 9];50(2):277–289. Available from: http://www.sciencedirect.com/science/article/pii/S0896627306002698
11. Woo S-H, Ranade S, Weyer AD, Dubin AE, Baba Y, Qiu Z, Petrus M, Miyamoto T, Reddy K, Lumpkin EA, Stucky CL, Patapoutian A. Piezo2 is required for Merkel-cell mechanotransduction. Nature [Internet]. NIH Public Access; 2014 May 29 [cited 2017 May 9];509(7502):622–6. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24717433 PMID: 24717433
12. Shmukler BE, Huston NC, Thon JN, Ni C-W, Kourkoulis G, Lawson ND, Paw BH, Alper SL. Homozygous knockout of the piezo1 gene in the zebrafish is not associated with anemia. Haematologica [Internet]. 2015 Dec 1 [cited 2017 May 9];100(12):e483–e485. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26294733 PMID: 26294733
13. Delmas P, Hao J, Rodat-Despoix L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci [Internet]. Nature Publishing Group; 2011 Mar 9 [cited 2017 May 9];12(3):139–153. Available from: http://www.nature.com/doifinder/10.1038/nrn2993
14. Vitzthum C, Clauss WG, Fronius M. Mechanosensitive activation of CFTR by increased cell volume and hydrostatic pressure but not shear stress. Biochim Biophys Acta - Biomembr [Internet]. 2015 Nov [cited 2017 May 9];1848(11):2942–2951. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26357939 PMID: 26357939
15. Beloussov L V. Morphogenesis can be driven by properly parametrised mechanical feedback. Eur Phys J E [Internet]. Springer Berlin Heidelberg; 2013 Nov 25 [cited 2017 Apr 30];36(11):132. Available from: http://link.springer.com/10.1140/epje/i2013-13132-x
16. Plouhinec J-L, De Robertis EM. Systems Biology of the Self-regulating Morphogenetic Gradient of the Xenopus Gastrula. Cold Spring Harb Perspect Biol [Internet]. 2009 Aug 1 [cited 2017 Apr 30];1(2):a001701–a001701. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20066084 PMID: 20066084
17. Raftery LA, Sutherland DJ. Gradients and thresholds: BMP response gradients unveiled in Drosophila embryos. Trends Genet [Internet]. 2003 Dec [cited 2017 Apr 30];19(12):701–708. Available from: http://www.ncbi.nlm.nih.gov/pubmed/14642751 PMID: 14642751
18. Reversade B, Kuroda H, Lee H, Mays A, De Robertis EM. Depletion of Bmp2, Bmp4, Bmp7 and Spemann organizer signals induces massive brain formation in Xenopus embryos. Development [Internet]. 2005 Jun 23 [cited 2017 Apr 30];132(15):3381–3392. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15975940 PMID: 15975940
19. Plouhinec J-L, Zakin L, De Robertis EM. Systems control of BMP morphogen flow in vertebrate embryos. Curr Opin Genet Dev [Internet]. 2011 Dec [cited 2017 Apr 30];21(6):696–703. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21937218 PMID: 21937218
20. De Robertis EM. Spemann’s organizer and the self-regulation of embryonic fields. Mech Dev [Internet]. NIH Public Access; 2009 Dec [cited 2017 May 9];126(11–12):925–41. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19733655 PMID: 19733655
21. Stottmann RW, Berrong M, Matta K, Choi M, Klingensmith J. The BMP antagonist Noggin promotes cranial and spinal neurulation by distinct mechanisms. Dev Biol [Internet]. 2006 Jul 15 [cited 2017 Apr 30];295(2):647–663. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16712836 PMID: 16712836
22. Ybot-Gonzalez P, Gaston-Massuet C, Girdler G, Klingensmith J, Arkell R, Greene NDE, Copp AJ. Neural plate morphogenesis during mouse neurulation is regulated by antagonism of Bmp signalling. Development [Internet]. 2007 Aug 1 [cited 2017 Apr 30];134(17):3203–3211. Available from: http://www.ncbi.nlm.nih.gov/pubmed/17693602 PMID: 17693602
23. Bond AM, Bhalala OG, Kessler JA. The dynamic role of bone morphogenetic proteins in neural stem cell fate and maturation. Dev Neurobiol [Internet]. NIH Public Access; 2012 Jul [cited 2017 May 9];72(7):1068–84. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22489086 PMID: 22489086
24. Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature [Internet]. Nature Publishing Group; 1996 Oct 3 [cited 2017 May 10];383(6599):407–413. Available from: http://www.nature.com/doifinder/10.1038/383407a0
25. Bae G-U, Domene S, Roessler E, Schachter K, Kang J-S, Muenke M, Krauss RS. Mutations in CDON, encoding a hedgehog receptor, result in holoprosencephaly and defective interactions with other hedgehog receptors. Am J Hum Genet [Internet]. Elsevier; 2011 Aug 12 [cited 2017 May 10];89(2):231–40. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21802063 PMID: 21802063
26. Golden JA, Bracilovic A, McFadden KA, Beesley JS, Rubenstein JL, Grinspan JB. Ectopic bone morphogenetic proteins 5 and 4 in the chicken forebrain lead to cyclopia and holoprosencephaly. Proc Natl Acad Sci U S A [Internet]. National Academy of Sciences; 1999 Mar 2 [cited 2017 May 10];96(5):2439–44. Available from: http://www.ncbi.nlm.nih.gov/pubmed/10051661 PMID: 10051661
27. Roessler E, Belloni E, Gaudenz K, Jay P, Berta P, Scherer SW, Tsui L-C, Muenke M. Mutations in the human Sonic Hedgehog gene cause holoprosencephaly. Nat Genet [Internet]. 1996 Nov [cited 2017 May 10];14(3):357–360. Available from: http://www.ncbi.nlm.nih.gov/pubmed/8896572 PMID: 8896572
28. Chen JK, Taipale J, Cooper MK, Beachy PA. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev [Internet]. Cold Spring Harbor Laboratory Press; 2002 Nov 1 [cited 2017 May 11];16(21):2743–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/12414725 PMID: 12414725
29. BINNS W, JAMES LF, SHUPE JL, THACKER EJ. Cyclopian-type malformation in lambs. Arch Environ Health [Internet]. 1962 Aug [cited 2017 May 11];5:106–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/13869306 PMID: 13869306
30. Keeler RF, Binns W. Teratogenic compounds of Veratrum californicum (Durand). I. Preparation and characterization of fractions and alkaloids for biologic testing. Can J Biochem [Internet]. 1966 Jun [cited 2017 May 11];44(6):819–28. Available from: http://www.ncbi.nlm.nih.gov/pubmed/5919283 PMID: 5919283
31. Taylor MD, Northcott PA, Korshunov A, Remke M, Cho Y-J, Clifford SC, Eberhart CG, Parsons DW, Rutkowski S, Gajjar A, Ellison DW, Lichter P, Gilbertson RJ, Pomeroy SL, Kool M, Pfister SM. Molecular subgroups of medulloblastoma: the current consensus. Acta Neuropathol [Internet]. Springer; 2012 Apr [cited 2017 Jan 21];123(4):465–72. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22134537 PMID: 22134537
32. Jain S, Song R, Xie J. Sonidegib: mechanism of action, pharmacology, and clinical utility for advanced basal cell carcinomas. Onco Targets Ther [Internet]. Dove Press; 2017 [cited 2017 May 11];10:1645–1653. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28352196 PMID: 28352196
33. Collier NJ, Ali FR, Lear JT. The safety and efficacy of sonidegib for the treatment of locally advanced basal cell carcinoma. Expert Rev Anticancer Ther [Internet]. 2016 Oct 2 [cited 2017 May 11];16(10):1011–1018. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27636236 PMID: 27636236
34. Tremblay MR, Lescarbeau A, Grogan MJ, Tan E, Lin G, Austad BC, Yu L-C, Behnke ML, Nair SJ, Hagel M, White K, Conley J, Manna JD, Alvarez-Diez TM, Hoyt J, Woodward CN, Sydor JR, Pink M, MacDougall J, Campbell MJ, Cushing J, Ferguson J, Curtis MS, McGovern K, Read MA, Palombella VJ, Adams J, Castro AC. Discovery of a Potent and Orally Active Hedgehog Pathway Antagonist (IPI-926). J Med Chem [Internet]. 2009 Jul 23 [cited 2017 May 11];52(14):4400–4418. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19522463 PMID: 19522463
35. De Smaele E, Ferretti E, Gulino A. Vismodegib, a small-molecule inhibitor of the hedgehog pathway for the treatment of advanced cancers. Curr Opin Investig Drugs [Internet]. 2010 Jun [cited 2017 May 11];11(6):707–18. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20496266 PMID: 20496266
36. Kaneko KJ, Kohn MJ, Liu C, DePamphilis ML. Transcription factor TEAD2 is involved in neural tube closure. genesis [Internet]. 2007 Sep [cited 2017 May 11];45(9):577–587. Available from: http://doi.wiley.com/10.1002/dvg.20330
37. Preiksaitiene E, Benusiene E, Ciuladaite Z, Sliuzas V, Mikstiene V, Kucinskas V. Recurrent fetal syndromic spina bifida associated with 3q26.1-qter duplication and 5p13.33-pter deletion due to familial balanced rearrangement. Taiwan J Obstet Gynecol [Internet]. 2016 Jun [cited 2017 May 11];55(3):410–414. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27343325 PMID: 27343325
38. Anderson MJ, Schimmang T, Lewandoski M. An FGF3-BMP Signaling Axis Regulates Caudal Neural Tube Closure, Neural Crest Specification and Anterior-Posterior Axis Extension. Wilkie AOM, editor. PLOS Genet [Internet]. 2016 May 4 [cited 2017 May 11];12(5):e1006018. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27144312 PMID: 27144312
39. Milunsky A, Jick H, Jick SS, Bruell CL, MacLaughlin DS, Rothman KJ, Willett W. Multivitamin/folic acid supplementation in early pregnancy reduces the prevalence of neural tube defects. JAMA [Internet]. 1989 Nov 24 [cited 2017 May 11];262(20):2847–52. Available from: http://www.ncbi.nlm.nih.gov/pubmed/2478730 PMID: 2478730
40. Roberts E, Hampshire DJ, Pattison L, Springell K, Jafri H, Corry P, Mannon J, Rashid Y, Crow Y, Bond J, Woods CG. Autosomal recessive primary microcephaly: an analysis of locus heterogeneity and phenotypic variation. J Med Genet [Internet]. 2002 Oct [cited 2017 May 11];39(10):718–21. Available from: http://www.ncbi.nlm.nih.gov/pubmed/12362027 PMID: 12362027
41. Lamb T, Knecht A, Smith W, Stachel S, Economides A, Stahl N, Yancopolous G, Harland R. Neural induction by the secreted polypeptide noggin. Science (80- ) [Internet]. 1993 Oct 29 [cited 2017 May 10];262(5134):713–718. Available from: http://www.sciencemag.org/cgi/doi/10.1126/science.8235591
42. Hemmati-Brivanlou A, Kelly OG, Melton DA. Follistatin, an Antagonist of Activin, Is Expressed in the Spemann Organizer and Displays Direct Neuralizing Activity. Cell [Internet]. 1994 [cited 2017 May 10];77:283–295. Available from: http://ac.els-cdn.com/0092867494903204/1-s2.0-0092867494903204-main.pdf?_tid=7806765c-35bd-11e7-940c-00000aacb361&acdnat=1494447533_a2c62319c30208326f4b1dc6dfb38b9f
43. Sasai Y, Lu B, Steinbeisser H, De Robertis EM. Regulation of neural induction by the Chd and Bmp-4 antagonistic patterning signals in Xenopus. Nature [Internet]. Nature Publishing Group; 1995 Jul 27 [cited 2017 May 10];376(6538):333–336. Available from: http://www.nature.com/doifinder/10.1038/376333a0
44. Bouwmeester T, Kim S-H, Sasai Y, Lu B, Robertis EM De. Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann’s organizer. Nature [Internet]. Nature Publishing Group; 1996 Aug 15 [cited 2017 May 10];382(6592):595–601. Available from: http://www.nature.com/doifinder/10.1038/382595a0
45. Hansen CS, Marion CD, Steele K, George S, Smith WC. Direct neural induction and selective inhibition of mesoderm and epidermis inducers by Xnr3. Development [Internet]. 1997 [cited 2017 May 10];124(2). Available from: http://dev.biologists.org/content/124/2/483.long
46. Cox J, Jackson AP, Bond J, Woods CG. What primary microcephaly can tell us about brain growth. Trends Mol Med [Internet]. 2006 Aug [cited 2017 May 11];12(8):358–366. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16829198 PMID: 16829198
Статья отредактирована членами команды EQUALITY: спасибо Анне Курановой, врачу-исследователю из Междисциплинарного Центра Психопатологии и Эмоционального Регулирования в Медицинском Центре Университета Гронингена.
Некоторое время назад мне стали попадаться интервью с Сергеем Вячеславовичем Савельевым, профессором, заведующим лабораторией развития нервной системы Института морфологии человека ФАНО. Одно из них было дано газете «Комсомольская Правда» от 22 июля 2010 года. В нем Савельев повествует о различиях в массе мозга между жителями разных стран, а также между мужчинами и женщинами, и о том, какие выводы из этих различий, по его мнению, следуют.
В то время о Савельеве мне было неизвестно абсолютно ничего. Никакие его работы по морфологии мозга или нейрофизиологии на семинарах не звучали. В интервью “КП” насторожили некоторые странности с тем, как Савельев подает факты и интерпретирует их. Впрочем, это я отнес на счет необходимости интервьюируемого общаться с журналистами.
Несколько лет спустя увидел Савельева на канале Дмитрия Пучкова oper.ru. Пучков публикует очень интересные интервью с историками, но биологов приглашает крайне редко, поэтому начал слушать с интересом. Однако по мере просмотра ролика стало нарастать ощущение, что в студии происходит что-то не то. Названный профессор нес редкостную чепуху из наукообразных терминов, «дней минувших анекдотов» и кухонных стереотипов вперемешку с обвинениями в адрес генетиков и молекулярных биологов в растрате государственных средств.
Ничего относящегося к нейрофизиологии, морфологии ЦНС или науке вообще в беседе с Савельевым не было. Странной казалась позиция Пучкова, который в своих «разведопросах» с историками обличал лженауку в лице Фоменко и Задорнова, здесь же открыто поддерживал Савельева и настоятельно рекомендовал слушателям его произведения с названиями типа «Церебральный сортинг».
Поскольку по ролику в YouTube не всегда можно судить о профессионализме научного работника, я разыскал публикации Савельева в рецензируемых изданиях. В большинстве своем они оказались русскоязычными. Поскольку Савельев утверждает, что он специалист по морфологии мозга и эмбриологии, я выбрал одну из его недавних работ, которая, судя по названию, имеет отношение к обеим областям: «Патология эмбрионального морфогенеза головного мозга человека» 1.
Начнем с абстракта. В нем вкратце должно сообщаться, что нам следует ожидать в статье: какая гипотеза, методы ее тестирования, результаты экспериментов и их интерпретация в соответствии с общепринятой научной методологией.
В кратком содержании Савельев заявляет, что в данной работе он произвел экспериментальные исследования и получил следующие результаты (курсивом выделен текст Савелева, нумерация моя, К. Л.):
1. Исследованы механизмы развития ранней эмбриональной патологии нервной системы у человека и экспериментальных животных.
Таким образом, ожидается, что автор предоставит в статье экспериментальные данные, включая материалы, методы и результаты, полученные на неких животных и человеческих модельных системах.
2. Установлены биомеханические принципы кодирования позиционной информации нейроэпителиальными клетками.
Раскрытие механизма «кодирования позиционной информации нейроэпителиальными клетками» явилось бы серьезным прорывом в биологии развития. Поэтому с нетерпением ждем экспериментальных данных Савельева, описывающих носителей этого «кода», его свойства, механизмы считывания и преобразования закодированной информации в реальные биологические структуры.
3. Экспериментальное обратимое изменение тангенциальных межклеточных взаимодействий в нейроэпителии животных приводило к прогнозируемым аномалиям развития нервной системы. Изменение проницаемости механозависимых ионных каналов является основным способом считывания позиционной информации и детерминации клеточной дифференцировки. Было проведено сравнение экспериментальных аномалий мозга у животных с уникальными аутопсиями патологического развития мозга человека, начиная от нейруляции и заканчивая 6-м мес внутриутробного развития.
Таким образом, ожидается, что Савельев, обладая соответствующей аппаратурой и методами, провел эксперименты по а) количественному измерению и б) контролированному изменению «тангенциальных межклеточных взаимодействий в нейроэпителии животных».
Согласно заявлению Савельева, он способен прогнозировать и воспроизводить аномалии развития нервной системы, направленно изменяя «тангенциальные межклеточные взаимодействия» (надеемся, что Савельев также даст определение, что же такое «тангенциальные межклеточные взаимодействия», какие белки их опосредуют, какими методами Савельев эти взаимодействия измеряет и изменяет).
На основе результатов экспериментов, описание которых мы ожидаем увидеть в статье, Савельев делает вывод, что проводимость механозависимых ионных каналов меняется вследствие механических взаимодействий между клетками. По утверждению Савельева, проницаемость механозависимых ионных каналов – это и есть механизм считывания «позиционной информации» (см. выше), которая преобразуется в дифференцировку клетки-предшественника.
Следует ожидать, что в статье автор предоставит экспериментальные данные по изменению проводимости механозависимых ионных каналов с количественным измерением этой проводимости с помощью соответствующего оборудования. Также следует ожидать, что автор путем контролируемого манипулирования механическими натяжениями в развивающемся эмбрионе продемонстрирует изменение проводимости ионных каналов, изменение дифференцировки клеток и экспериментально воспроизведет аномалии мозга на модельных организмах. Ожидается, что эти аномалии, согласно заявлению Савельева, будут аналогичны известным патологиям развития человеческого мозга.
4. Полученные результаты позволили установить, что при задержке движения постнейропорной волны замыкания нервной трубки формируются аномалии переднего мозга и этмоидной части лица. Обнаружено, что при остановке движения постнейропорной волны нейруляции формируются аномалии промежуточного и среднего мозга, а при нарушении каудальной волны — аномалии заднего и спинного мозга. Показано, что аномалии развития, диагностируемые на поздних сроках онтогенеза и после рождения, являются последствиями компенсаторных реакций нейроэпителия на изменение ранних позиционных сигналов в период формирования нервной трубки.
Здесь мы сталкиваемся с нетрадиционной терминологией Савельева. Под «постнейропорной волной замыкания нервной трубки», скорее всего, имеется в виду нейруляция – процесс замыкания нервной пластинки в нервную трубку, который у человека начинается в двух местах: посередине и в головной области эмбриона – и распространяется в антериальной области встречными волнами с образованием антериального нейропора (отверстия), а в каудальной (хвостовой) с образованием постериального нейропора. Ознакомление с предметом можно начать с обзора Copp, 2013 2.
Судя по его утверждению, Савельев экспериментально индуцировал задержку нейруляции и смоделировал «аномалии переднего мозга и этмоидной части лица». Интересно, что даже термины Савельев подбирает не слишком корректно. Взять ту же «этмоидную часть лица» - не сразу понятно, что вообще имеется в виду. Есть этмоидная кость, расположенная за носовой костью, и ассоциированный с ней этмоидный хрящ. Они входят в состав лицевого скелета. Судя по содержанию статьи, Савельев имеет в виду циклопию, при которой могут отсутствовать или быть редуцированы носовая и этмоидная кости.
Под «поздними сроками онтогенеза», скорее всего, имеются в виду поздние сроки эмбриогенеза или эмбрионального онтогенеза. Согласно определению Геккеля, онтогенез – это развитие организма от зачатия до конца жизни. Если следовать этому определению, то Савельев, очевидно, изучал половозрелых людей преклонного возраста, на поздних сроках их онтогенеза. Впрочем, на фоне остального это незначительное терминологическое замечание.
Из последнего предложения в абстракте следует, что Савельев экспериментально «изменил ранние позиционные сигналы», что привело к детектируемым «компенсаторным реакциям нейроэпителия» и, как следствие, к «аномалиям развития».
Пара слов об английской версии абстракта. Очевидно, что сам Савельев не владеет в достаточной степени языком и ему следовало воспользоваться услугами профессионального переводчика. В этом случае он избежал бы курьезных фраз, вроде: «The above anomalies resulted from local compensatory responses of the neuroepithelium due to the lack of intentions [sic!] that are characteristic of normal development of the neural tube».
В переводе это выглядит примерно так: «Аномалии выше были результатом локального компенсаторного ответа нейроэпителия из-за недостатка намерений (!) которые характеризуют нормальное развитие нервной трубки.»
Подведем итоги по абстракту. Савельев делает очень серьезные заявления о множестве собственных фундаментальных открытий в одной публикации. Такие заявления должны быть подкреплены результатами большого объёма очень хорошо спланированной экспериментальной работы. Посмотрим, что же на самом деле содержится в статье.
Введение
Обычно введение содержит постановку проблемы, обзор текущей литературы по данной теме, гипотезу и стратегию ее тестирования.
В качестве проблемы Савельев называет «необходимость поиска наиболее уязвимого и универсального механизма возникновения ранних отклонений в развитии нервной системы, которые обусловлены нарушением морфогенетических процессов и крайне плохо изучены».
Цель работы Савельев определяет как «поиск ранних эмбриональных межклеточных взаимодействий, детерминирующих формообразование головного мозга, нарушение которых вызывает необратимые аномалии развития человека». Иными словами, рабочая гипотеза автора состоит в том, что, нарушив каким-то образом межклеточные взаимодействия в эмбрионе на ранней стадии, он способен экспериментально воспроизвести некие неоговоренные «аномалии развития».
В качестве стратегии тестирования своей гипотезы Савельев называет «два параллельных подхода: изучение клинических случаев ранних эмбриональных патологий нервной системы человека и экспериментально-эмбриологический подход по моделированию наиболее распространенных аномалий развития.»
Таким образом, если Савельев стремится быть последовательным и отталкивается от своей гипотетической модели о роли межклеточных взаимодействий в морфогенезе мозга, нам следует ожидать, что в данной статье Савельев опишет 1) исследование белков, отвечающих за межклеточные взаимодействия, на клиническом материале и 2) экспериментальное изменение этих межклеточных взаимодействий с целью смоделировать некие неуказанные, но известные аномалии развития мозга.
Савельев не считает нужным указать модельную систему для второго подхода. Очевидно, это должно быть какое-то лабораторное животное. Общепринятой практикой считается указание модельного организма если не в заголовке, то хотя бы в абстракте. Савельев пренебрегает этим правилом.
Сразу настораживает крайняя расплывчатость проблемы и гипотезы. Когда речь идет о работе в области любой патологии, название патологии или группы патологий должно четко озвучиваться в виде диагноза в соответствии с международной классификацией заболеваний (ICD-10), которая используется в Российской Федерации как члене Международной Организации Здравоохранения (WHO). Врожденные аномалии нервной системы, которые, судя по всему, имеет в виду Савельев, включают около сотни диагностируемых патологий и их вариантов, которые по ICD-10 классифицируются статьями Q00-Q07. Отсутствие ссылки на конкретную группу кодифицированных диагнозов указывает на непрофессионализм Савельева.
Не приводя ссылок на соответствующие экспериментальные работы, Савельев утверждает, что «несмотря на разнообразие причин патологии развития, структурные изменения зародышей носят сходный характер. Однотипные морфогенетические реакции клеток ранних зародышей человека говорят о существовании универсальных механизмов рецепции различных типов патогенетических воздействий». Что за «однотипные морфогенетические реакции», в чем заключается «сходный характер» изменений зародыша и относится ли это ко всей сотне патологий из ICD-10-Q00-Q07, Савельев не удосуживается сообщить.
Савельев пишет: «К примеру, было неоднократно описано большое разнообразие аномалий при одном генетическом дефекте и одинаковые аномалии при изменении различных хромосом [1–4].» В профессиональной литературе это называют плейотропным действием гена в первом случае и мультигенным наследованием признака – во втором. Интерес представляет подбор ссылок, следующих за этим утверждением. Первой ссылкой Савельев цитирует свою собственную статью:
С. В. Савельев. Нейруляционные формы патологии нервной системы человека. // Архив патологии. — 1998. — №5. — С. 13—18.
При прочтении этой статьи оказывается, что никаких генетических исследований Савельев не провел. То есть ссылка была приведена некорректно. Вместо этого автор утверждает, что для «изучения аномалий развития головного мозга у эмбрионов в возрасте от 3 до 6 нед. и выявления общих принципов патогенеза этих аномалий мозга» он «исследовал» 47 абортивных эмбрионов.
Без проведения каких-либо генетических тестов, не приводя никаких сведений из литературы, Савельев утверждает, что «корреляция между сомитами, числом позвонков и ганглиями является довольно точным критерием наличия или отсутствия генетических изменений у эмбриона. Если в цервикальном, торакальном, люмбальном, крестцовом или копчиковом отделе соотношение между осевыми структурами нарушается, то наиболее вероятно наличие геномных изменений у зародыша. Таких эмбрионов не использовали». На чем основано убеждение Савельева, что генетические изменения отсутствуют у эмбрионов, взятых им для «исследования», абсолютно непонятно, поскольку ни кариотипирование, ни более глубокое генотипирование проведено не было.
Далее Савельев пишет: «Для гистологических исследований применяли фиксаторы Буэна, Карнуа и Ценкера, 10% формальдегид. Из блоков готовили серийные срезы толщиной от 5 до 15 мкм, их монтировали на стекла, окрашивали гематоксилином и эозином, по Массону, Бионди, заключали в бальзам. Срезы использовали для создания трехмерных графических реконструкций». В данном случае имеет место серьезное нарушение методологии, заключающееся в некорректном сравнении препаратов, обработанных произвольно разными методами. На чем основывался выбор фиксатора, не указывается. Какие эмбрионы фиксировались каким фиксатором и как это могло отразиться на результатах исследования, Савельев не считает нужным указывать.
То же самое относится к окраске препаратов. Статья не содержит фотографий самих препаратов, лишь некие «трехмерные графические реконструкции» крайне низкого разрешения, сомнительного качества и, как следствие, абсолютно неинформативные. Почему Савельев не публикует фотографии гистологических срезов, как это принято в работах в области эмбриологии, остается непонятным.
Таким образом, статья Савельева 1998 г. является субстандартной и низкокачественной. Самоцитирование ее в статье 2012 г. очень похоже на наивную попытку поднять свой индекс.
Вторая ссылка относит нас в 1957 год:
Hugehes A. Development of the primary sensory system in Xenopus laevis (Daudin). J. Anat. 1957; 91: 323–328.
Как и в первом случае, статья не содержит никаких генетических исследований. Это работа по развитию органов чувств у шпорцевой лягушки. Никаких исследований аномалий развития мозга в статье не проводится. Почему Савельев цитирует ее в связи с генетическими патологиями у человека – абсолютно непонятно.
Следующая цитируемая Савельевым статья – это исследование нарушений ЦНС у куриц после кормления их органофосфатами:
Tanaka D.J., Bursian S.J. Degеneration patterns in the chicken central nervous system induced by ingestion of the organophosphorus delayed neurotoxin 3-orthototolylphosphate. A silver impregnation study. Brain Res. 1989; 484: 240–256.
По ее прочтении оказывается, что авторы работали не с эмбрионами, а с 8-месячными курицами породы белый леггорн. После кормления их 3-орто-толилфосфатом, куриц забивали в течение 4 недель и исследовали дегенерацию нейронов разных участков мозга. Ни эмбриологических, ни генетических исследований не проводилось.
То, что Савельев цитирует этo в связи с генетическими нарушениями морфогенеза головного мозга человека, говорит о том, что статью он не читал. Цитирует он ее лишь «для вида и объема», что является очевидной фикцией. Савельев уподобляется нерадивому студенту, который кидает какие попало ссылки в свой реферат в надежде, что преподаватель не обратит на них внимание. Это свидетельствует о серьезном непрофессионализме Савельева.
Четвертая ссылка: Simon E.M., Barkovich A.J. Holoprosencephaly: new concepts. Magn. Reson. Imaging. Clin. N. Am. 2001; 9 (1): 149–164. Эта работа опубликована в издании, ориентированном на использовании ядерного магнитного резонанса в клинических целях. Генетические исследования в статье не приводятся. Работа посвящена исключительно ЯМР. Один из разделов в конце статьи упоминает известные на тот момент работы молекулярных генетиков о роли мутаций SHH ZIC2 и SIX3 в развитии голопрозэнцефалии, однако по какой-то причине Савельев цитирует эту вторичную работу, а не оригинальные. Как увидим позднее, начиная с конца 1990-х годов, появляется всплеск экспериментальной литературы по этиологии голопрозэнцефалии, которые Савельев не цитирует вовсе, что указывает на то, что он с ней скорее всего не знаком.
Далее в введении Савельев уделяет много внимания работе Чарльза Стокарда 1921 года. Это одна из ранних работ по тератогенезу. За прошедшие 90+ лет появились более разработанные модели морфогенетических патологий. Исследования в области эмбриональной молекулярной генетики в последние 15 лет до написания Савельевым его статьи привели к тому, что работы Стокарда в настоящее время представляют лишь исторический интерес.
Читаем дальше. Савельев пишет: «По предварительным данным, реальная гибель зародышей человека в первые 10–14 дней после оплодотворения может составлять до 80%. Столь высокий уровень гибели обусловлен морфогенетической уязвимостью мигрирующего зародыша, нарушением имплантационных процессов и механизмов формообразования [6–8].»
Три приведенные ссылки – снова цитирование Савельевым собственных статей сомнительного содержания и научной ценности, не содержащих его собственных исследований по статистике преклинической мортальности человеческих эмбрионов. По оценкам Optiz, 19873, потери до первых пропущенных месячных, включая неделящиеся яйцеклетки, пре-имплантационные проблемы и отсутствие имплантации составляют около 40-55% от всех зачатий.
Далее Савельев повторяет описание «эмбриона темякопчиковой длиной 3,2 мм на 22–23-й день после оплодотворения» из своей публикации 1993 года. Этот эмбрион якобы имел доселе не описанную аномалию: «Аномалия возникла к концу нейруляции. На это указывает эктодерма, расположенная над дорсальной поверхностью головного и спинного мозга. Пластовое строение мозга этого эмбриона утрачено, а спинной мозг заканчивается на уровне брюшного стебелька. Спинной мозг не достигал своего нормального размера и был на треть короче, чем в норме».
Проблема в том, что у человека на 22-23-ий день нейруляция только начинается, и закрывается нервная трубка только к 24-му дню (ростральный нейропор) и 26-28-му дню (каудальный нейропор)4–6. Савельев показывает свою полную безграмотность в эмбриологии нервной системы.
Учитывая неопределенность с возрастом эмбриона (или просто ошибку Савельева с его определением), эмбрион мог быть фиксирован либо в самом начале нейруляции, либо в середине процесса. Делать какие-либо заключения о строении головного и спинного мозга в этот период бессмысленно. Поскольку никто, кроме Савельева, этого эмбриона не видел, сложно сказать, действительно ли имела место какая-либо аномалия, или, что более вероятно, Савельев выдал самую раннюю стадию нормальной нейруляции за придуманный им дефект.
О том же самом эмбрионе Савельев рассуждает: «Подобные реакции нейроэпителия наблюдали в экспериментах по сбросу осевых механических напряжений у амфибий [12–14]. Окклюзия нервной трубки и деструктурализация пластов характерны для релаксации механически ненапряженного нейроэпителия».
Здесь Савельев демонстрирует взгляды профессора кафедры эмбриологии биологического факультета МГУ Льва Белоусова. Белоусов и Савельев считают, что при морфогенезе главную роль играют «механические напряжения» клеточных пластов. Ни в классической работе Nusslein-Volhard, 19887о роли градиента экспрессии bicoid в морфогенезе D. Melanogaster, ни в последующем всплеске работ, исследовавших регуляцию морфогенеза градиентами различных мРНК и белков, ведущая роль «механических напряжений» в морфогенезе не рассматривается. Тем не менее, Савельев полностью игнорирует несколько тысяч статей по исследованиям роли градиентов морфогенных белков в период 1988 – 2012 гг. Судя по отсутствию цитат этих работ в его статьях, Савельев скорее всего не знаком с текущей литературой по молекулярным основам морфогенеза.
Давайте посмотрим, как Белоусов и Савельев тестируют эту гипотезу. Когда речь идет о таком физическом явлении как механическое напряжение, необходимо четко определить, какая конкретно величина имеется в виду, какова ее размерность? Какими методами ее измеряют и изменяют? Наконец, что очень важно, создается ли в реальном эмбрионе натяжение, соизмеримое с тем, которое авторы искусственно создают в эксперименте?
Рис.1. Иллюстрация из статьи Савельева9. Красным отмечены линии сечений, которые полагалось сделать в качестве минимального контроля. В целом, замысел эксперимента на уровне юного натуралиста. Ни фотографий, ни количественных данных Савельев не приводит. В тексте называет изображенное «эмбриональным мозгом человека».
Если мы имеем дело с классическим механическим напряжением, то это величина тензорная. Трехмерная, развивающаяся во времени система клеток эмбриона предполагает, что изменение ее геометрии описывается 4-тензором. Это в самой упрощенной модели, которая не учитывает, что клетки соприкасаются не мембранами, а фокальными и щелевыми контактами. Локальные механические взаимодействия между клетками не ограничены «тургором», а включают в себя активность цитоскелета, которую тоже следует учитывать в математической модели.
Знакомство с публикациями Белоусова и Савельева8 показывает, что авторы никогда не производили измерений «механических напряжений» и не указывали размерность этой величины. Никаких соответствующих расчетов и экспериментальных измерений в вышеуказанных публикациях Савельева и Белоусова не приводится. Вместо этого авторы использовали весьма странный метод углов между соседними клетками на плоском срезе. Соответствуют ли найденные ими этим методом величины реальным напряжениям в трехмерном пласте клеток авторы не комментируют, просто потому что неспособны это сделать.
Примечательно, что в тексте статьи Савельев пишет: «… обратимся к результатам простых опытов по сбросу механических натяжений в эмбриональном мозге человека при помощи микрорассечений (рис 1)». На рисунке же изображен мозг иглистого тритона, а вовсе не человека. Никаких рассечений человеческого мозга в статье не приводится. Прочие эксперименты, описанные в статье Савельева 2001 года, также не соответствуют принятым на то время методологическим стандартам. Например, использование рисунка вместо фотографии (рис. 2, Савельев, 2001) в качестве экспериментального результата неприемлемо.
Рис. 2. «Схема опытов» Савельева по трансплантации нервной пластинки в нервную трубку с неясными целями и отсутствием каких-либо результатов в виде фотографий9. Савельев не предоставляет данные о морфологии, т.н. «поляризованных» клеток, ни тем более об их молекулярных маркерах. Интересно, что с начала 1990-х годов исследователи широко используют методику микроинъекции очищенных морфогенных мРНК и белков для оценки их влияния на нейруляцию и другие морфогенетические процессы41–45. Савельев явно незнаком с этими работами и не удосуживается тестировать транспланты на известные на момент написания статьи индукторы нейрогенеза, такие как Noggin, Follistatin, Chordin, Cerberus, and Xnr3.
На рис. 3 отсутствует какой-либо статистический анализ и элементарное описание наблюдаемых «аномалий».
Рис.3. Типичная иллюстрация из статьи Савельева9. С отсутствием статистического анализа.
Фотографии эмбрионов на рис. 4 абсолютно неинформативны и низкого качества. Когда хотят продемонстрировать какую-либо патологию, минимальное требование – хотя бы сделать гистологические срезы аномального участка эмбриона.
Рис.4. Савельев9демонстрирует «аномалии» развития после «диагонального растяжения нервной системы (sic!) на стадии бластулы и гаструлы ... нейрулы ... и нервной трубки.» Не очень понятно, где Савельев нашел нервную систему на стадии бластулы и гаструлы, чтобы ее «диагонально» растянуть. Отсутствует описание т.н. «аномалий», критериев, по которым эти аномалии определялись и отбирались, а также их гистологическое исследование и гистохимический анализ экспрессии известных морфогенов20,23,41–45и механосенсорных ионных каналов, на функции которых Савельев строит свою гипотезу. Возникает серьезный вопрос о физиологичности растяжения: генерируется ли такой стресс во время нормального эмбриогенеза или Савельев просто демонстрирует эмбрионы, погибшие от неспецифического механического воздействия. Отсутствуют контрольные эмбрионы, растянутые в других направлениях.
На рис. 5 непонятно, что конкретно Савельев измеряет по оси ординат в графике о содержании ионов в цитоплазме. Важный вопрос, возникающий при рассмотрении этого графика: не является ли изменение ионов хлора в цитоплазме.
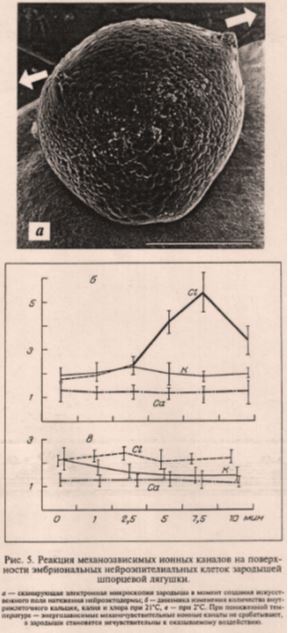 Рис. 5. Савельев9 демонстрирует эксперимент по измерению внутриклеточной концентрации ионов в эмбрионе после растяжения. Непонятно, измерялись ли концентрации ионов в эмбрионе, изображенном на верхней панели, после СЭМ (который очевидно проводился под вакуумом). Если так, то данные не имеют физиологического значения. Не проведен анализ дисперсии (ANOVA), что является характерным для статей Савельева пренебрежением статистическим анализом данных. Не указана величина на оси ординат. Все вышесказанное ставит способности Савельева как экспериментатора на уровень начинающего юного натуралиста, а отнюдь не профессионала, за которого он себя выдает. |
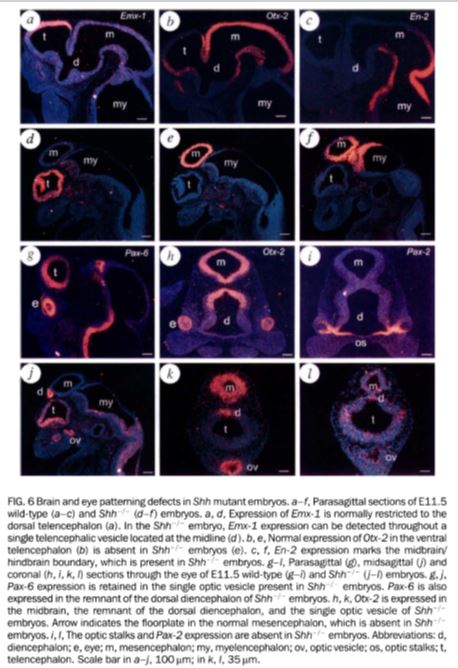
К слову о механозависимых ионных каналах. Они играют важную роль в органах чувств и метаболизме, однако их роль в раннем морфогенезе не прослеживается. Ни один из нокаутов известных генов механозависимых рецепторов не привел к нарушению нейруляции или других аспектов морфогенеза мозга10–12. В качестве обзора на эту тему могу посоветовать Delmas et al. 201113. Мутации в гене CFTR, кодирующем Cl--канал с потенциально механозависимой регуляцией14, хоть и приводят к муковисцидозу, но никак не отражаются на нейруляции. Надо сказать, что Белоусов продолжает публиковаться в журналах, правда с низким Impact Factor15. Псевдонаучность его статей даже не в том, что гипотеза о роли механических межклеточных взаимодействий в морфогенезе и дифференцировке клеток не поддерживается экспериментальными данными, а в отсутствии адекватной методологии у Белоусова и его соавторов, а также их неспособности поставить эксперименты для ее тестирования. Интересно, что как Белоусов, так и Савельев полностью игнорируют изменения градиентов и экспрессии известных морфогенов, таких, которые могут происходить под действием процедур рассечения или растяжения эмбриона. Работы группы De Robertis и других16–19демонстрируют наличие механизма обратной связи в экспрессии белков BMP, Sizzled, Tolloid таким образом, что после рассечения эмбриона Xenopus их градиенты восстанавливаются (в качестве обзора: De Robertis, 200920). Савельев не комментирует роль белков BMP2, BMP4, Noggin, Chordin, Xolloid, SHH и других в индукции нейруляции и регуляции дорзo-вентральной оси, что было уже известно на момент написания его статьи21,22. (В качестве обзора на эту тему см. Bond et al. 201223.) Таким образом, прослеживается закономерность, что, за исключением своих собственных публикаций, которые, как мы видим, крайне низкого качества, Савельев не цитирует работы по молекулярной эмбриологии, опубликованные с конца 1990-х до момента выхода статьи в 2012 году. Это свидетельствует о том, что автор либо не читает текущей литературы, либо сознательно игнорирует все работы, чьи выводы не согласуются с его воззрениями. В обоих случаях это свидетельство крайнего непрофессионализма и дилетантства Савельева. В силу вышеуказанных причин заявления Савельева об «открытии» им «механизмов кодирования позиционной информации в эмбриональном формообразовании мозга» являются ложью. Вернемся к статье Савельева 2012 года1. Автор вдается в пространное описание якобы описанных им эмбриональных аномалий головного мозга без каких-либо иллюстративных и гистологических данных о специфике этих аномалий. Происхождение врожденных аномалий головного мозга Савельев «объясняет» механическими напряжениями следующим образом:«Для выяснения роли механических напряжений в мозге ранних эмбрионов человека были исследованы патологические случаи развития. При локализации зародыша человека в маточной трубе на голову зародыша давление со стороны стенок маточной трубы действует неравномерно. По лобно-височно-затылочной линии мозг зародыша испытывает тангенциальное сжатие, в результате которого стенка мозга в теменной области утрачивает свою механическую напряженность. Пласты теменной области свободны от натяжений, существующих в нормальном развитии, и могут демонстрировать реакции, характерные для ненапряженных пластов [15, 39]. С одной стороны, при усилении механического давления на стенки мозга происходит увеличение поляризации клеток радиальной глии, снижается интенсивность пролиферации и увеличивается площадь внутренней поверхности стенок мозга, с другой — свободные края пластов подвергаются гиперплазии, а пролиферация нейробластов быстро возрастает. Анализ патологических случаев развития эмбрионов вне матки показывает, что механическое напряжение нейроэпителиальных пластов является необходимым условием для их нормального развития.» При этом Савельев не приводит никаких количественных данных о т.н. «механических напряжениях», испытываемых эмбрионом в матке vs. вне матки, при патологии vs. при ее отсутствии. Никаких данных об «интенсивности пролиферации», сведений о «гиперплазии» и ее молекулярных маркерах (например, Ki67), митотическом и апоптотическом индексах он не демонстрирует, очевидно, в силу того, что никаких исследований он просто не проводил, а вышесказанное является плодом его натурфилософствования. Теория хороша лишь тогда, когда способна предсказывать воспроизводимые экспериментальные результаты. Ни тестируемых предсказаний, ни результатов Савельев не предоставляет, демонстрируя полное отсутствие профессионализма. Как и раньше, цитирует Савельев или сам себя, или статьи 1980-х годов. А зря. В 1990-х годах публикуется серия исследований о молекулярно-генетической природе тех самых аномалий, которые Савельев «исследует» в данной статье. Одна из первых значительных работ была сделана группой Philip Beachy, John Hopkins University, которая обнаружила, что двойной нокаут гена Shh приводит к голопрозэнцефалии в виде циклопии у мышей24. На рис. 7 приведена одна из иллюстраций к этой статье, демонстрирующая циклопию у эмбрионов Shh-/- мышей. Сравните методологию и качество данных группы Beachy с тем, что публикует Савельев. Дальнейшие исследования нескольких групп продемонстрировали роль Shh и других компонентов сигнального пути Shh (BMP4, BMP5, CDON) в голопрозэнцефалии у мышей и кур25,26. Как можно заметить, голопрозэнцефалию вызывают мутации в тех же генах, продукты которых необходимы для индукции нейруляции у Xenopus20. Наконец, подтверждение роли Shh в голопрозэнцефалии у человека было сделано после обнаружения мутаций в гене Shh в том же 1996-м году27. 
Дополнительным свидетельством нарушения сигнального пути Shh в эмбрионах с циклопией явилось обнаружение того, что алкалоид циклопамин напрямую связывается и ингибирует белок Smoothened (Smo) – важный компонент пути Shh28. Циклопамин содержится в растениях рода чемерица (Veratrum). Его роль во вспышке циклопии среди ягнят, матери которых кормились на калифорнийской чемерице, была выявлена в 1960-х годах29,30. Странно, что Савельев не упоминает эти важные работы, когда пишет о причинах голопрозэнцефалии. Способность циклопамина ингибировать Smo и, таким образом, путь Shh нашла применение в онкологической фармакологии, поскольку путь Shh аномально активирован в ряде опухолей мозга, таких как медуллобластома31, а также базальной клеточной карциноме и ряде других опухолей. Были разработаны, прошли клинические испытания и были одобрены FDA синтетический аналог циклопамина – сонидегиб32,33и другие ингибиторы Shh пути саридегиб34и висмодегиб35. То, что Савельев 16 лет спустя после выявления роли сигнального пути Shh в голопрозэнцефалии пишет о механических напряжениях как причине нарушения нейруляции и возникновении голопрозэнцефалии, равносильно наивной попытке возродить теорию флогистона через много лет после опытов Лавуазье.
Этиология других врожденных аномалий головного мозга, которые, согласно Савельеву, якобы возникают вследствие механических стрессов, в настоящее время интенсивно изучается. Например, причины анэнцефалии, по всей вероятности, включают в себя недостаток фолиевой кислоты во время беременности35. Исследование 2007 года показало, что эмбрионы мышей, нокаутированные по гену транскрипционного фактора TEAD2, играющего важную роль в закрытии нервной трубки, имеют экзенцефалию, т.е. их головной мозг расположен вне черепной коробки36. Экзенцефалия является ранней стадией анэнцефалии. Интересно, что мыши, беременные tead2-/- эмбрионами и инъецированные фолиевой кислотой, рожали нокаутированных мышат без признаков анэнцефалии. Другой упомянутый Савельевым дефект, spina bifida, также зависит от приема матерью фолиевой кислоты и, возможно, частично опосредован генетическими причинами37–39. Савельев игнорирует вышеуказанные работы о причинах анэнцефалии и spina bifida. Возможно, будет интересно рассмотреть неупомянутый Савельевым дефект развития мозга – микроцефалию. Классический генетический анализ семей из северного Пакистана с часто встречающейся микроцефалией идентифицировал 5 локусов, ассоциированных с синдромом40. Клонирование генов, содержащих мутацию у пациентов с микроцефалией (MCPH1, CENPJ, ASPM, CDK5RAP2), выявило, что все они так или иначе участвуют в делении апикальных и базальных предшественников нейронов и глии. Мутации в генах этих белков приводили к нарушению ориентации веретена деления и расхождению хромосом в митозе. Это приводило к уменьшению количества базальных предшественников и, как результат, к уменьшению количества нейронов и глии в коре головного мозга (в качестве обзора Cox et al., 200646). В настоящее время обнаружено 17 локусов, ассоциированных с аутосомно-рецессивной микроцефалией40. Позволю себе опустить подробный разбор чепухи Савельева в оставшейся части его статьи. Основные пункты несостоятельности его гипотезы о механической природе аномалий нейруляции мы уже разобрали и сделали краткий обзор современной исследовательской литературы по генетическим и фармакологическим причинам голопрозэнцефалии, анэнцефалии и сходных патологий. Методологическая несостоятельность Савельева очевидна на фоне гигантского количества качественно сделанных работ в области молекулярной генетики развития нервной системы. Заключение Как мы видим, ни по одному из пунктов, заявленных в абстракте, Савельев не в состоянии предоставить мало-мальски приемлемых экспериментальных данных. Гипотеза, заявленная во введении, тестирована не была. Утверждение автора, что им были открыты «механизмы кодирования позиционной информации», не соответствует действительности. Утверждение Савельева о роли механических напряжений в этиологии врожденных аномалий развития мозга не подкреплено экспериментальными данными и не согласуется с имеющимися на момент выхода статьи исследованиями. По ознакомлении с вышеуказанными публикациями создается впечатление, что Савельев вообще не получил должного образования, которое позволило бы ему вести самостоятельную экспериментальную работу в медико-биологических науках на современном уровне. Отсутствие должного базового образования в сочетании с практикой под руководством таких псевдоученых как Лев Белоусов и нежелание знакомиться с современными методами и результатами исследований ведущих лабораторий привело к крайней маргинализации Савельева. Остается загадкой, как такой человек защитился и почему ему доверена лаборатория в государственном институте. Его докторская диссертация, несомненно, ждет детального анализа на соответствие заявленному уровню. Если разобранные выше статьи являются отчетом о проделанной им работе на занимаемой должности, то иначе как систематическим обманом и лженаукой деятельность Савельева на посту завлаба назвать нельзя. Дмитрий Пучков в одном из интервью с Савельевым говорит: «Настоятельно рекомендую [книгу Савельева «Церебральный сортинг», прим. К.Л.] к прочтению. Даже если со взглядами автора в чём-то не согласны, вызывает неприятие личность, как это обычно бывает, сведения исключительно полезные, знать надо, и знать надо обязательно». Позволю себе прокомментировать цитату: 1. Неприятие вызывает не личность, а, как видно из его собственных публикаций, полная несостоятельность Савельева как ученого, которая заключается в его невежестве, незнании литературы по нейробиологии и эмбриологии за последние 30 лет, в полном отсутствии у Савельева знания научной методологии, принципов построения эксперимента и интерпретации результатов. 2. Если вам действительно интересна биология развития и функционирование нервной системы, то, прежде чем покупать опусы Савельева и тем самым финансово поддерживать его коммерческий проект с издательством «Веди» по выпуску псевдонаучной литературы, абсолютно необходимо ознакомиться со следующими учебниками:
|
Интересный комментарий дал старший научный сотрудник кафедры эмбриологии биологического факультета МГУ Станислав Кремнёв:
"Несмотря на наличие С.В. Савельева в списке авторов статьи Beloussov LV, Saveliev SV, Naumidi II, Novoselov VV. Mechanical Stresses in Embryonic Tissues: Patterns, Morphogenetic Role, and Involvement in Regulatory Feedback. 1994 p. 1–34, Савельев не участвовал в этой работе и не имеет отношение к экспериментам по выявлению роли механических напряжений в морфогенезе. Савельев также не внес никакого вклада в построение карт механических напряжений в эмбрионах (Beloussov, L. V., Dorfman, J. G., and Cherdantzev, V. G. (1975))"
Литература
1. Saveliev S V. Патология эмбрионального морфогенеза головного мозга человека. Вестник РАМН. 2012;8:40–46. http://vestnikramn.spr-journal.ru/jour/article/download/276/214
2. Copp AJ, Greene NDE. Neural tube defects--disorders of neurulation and related embryonic processes. Wiley Interdiscip Rev Dev Biol [Internet]. Europe PMC Funders; 2013 [cited 2017 Apr 5];2(2):213–27. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24009034 PMID: 24009034
3. Opitz JM. The Farber Lecture. Prenatal and Perinatal Death. Pediatr Pathol [Internet]. 1987 Jan 9 [cited 2017 Apr 29];7(4):363–392. Available from: http://www.tandfonline.com/doi/full/10.3109/15513818709161402
4. O’Rahilly R, Muller F. Neurulation in the normal human embryo. Ciba Found Symp [Internet]. 1994 [cited 2017 Apr 29];181:70-82–9. Available from: http://www.ncbi.nlm.nih.gov/pubmed/8005032 PMID: 8005032
5. O’Rahilly R, Muller F. The development of the neural crest in the human. J Anat [Internet]. Wiley-Blackwell; 2007 Sep [cited 2017 Apr 29];211(3):335–51. Available from: http://www.ncbi.nlm.nih.gov/pubmed/17848161 PMID: 17848161
6. Van Allen MI, Kalousek DK, Chernoff GF, Juriloff D, Harris M, McGillivray BC, Yong S-L, Langlois S, Macleod PM, Chitayat D, Friedman JM, Wilson RD, McFadden D, Pantzar J, Ritchie S, Hall JG. Evidence for multi-site closure of the neural tube in humans. Am J Med Genet [Internet]. 1993 Oct 1 [cited 2017 Apr 29];47(5):723–743. Available from: http://doi.wiley.com/10.1002/ajmg.1320470528
7. Berleth T, Burri M, Thoma G, Bopp D, Richstein S, Frigerio G, Noll M, Nusslein-Volhard C. The role of localization of bicoid RNA in organizing the anterior pattern of the Drosophila embryo. EMBO J [Internet]. European Molecular Biology Organization; 1988 Jun [cited 2017 Apr 29];7(6):1749–56. Available from: http://www.ncbi.nlm.nih.gov/pubmed/2901954 PMID: 2901954
8. Beloussov LV, Saveliev SV, Naumidi II, Novoselov VV. Mechanical Stresses in Embryonic Tissues: Patterns, Morphogenetic Role, and Involvement in Regulatory Feedback. 1994 [cited 2017 Apr 30]. p. 1–34. Available from: http://linkinghub.elsevier.com/retrieve/pii/S0074769608615351
9. Савельев СВ. Механизм кодирования позиционной информации в эмбриональном формообразовании мозга позвоночных. Вестник РАМН. 2001;4:49–61.
10. Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang D-S, Woolf CJ, Corey DP. TRPA1 Contributes to Cold, Mechanical, and Chemical Nociception but Is Not Essential for Hair-Cell Transduction. Neuron [Internet]. 2006 [cited 2017 May 9];50(2):277–289. Available from: http://www.sciencedirect.com/science/article/pii/S0896627306002698
11. Woo S-H, Ranade S, Weyer AD, Dubin AE, Baba Y, Qiu Z, Petrus M, Miyamoto T, Reddy K, Lumpkin EA, Stucky CL, Patapoutian A. Piezo2 is required for Merkel-cell mechanotransduction. Nature [Internet]. NIH Public Access; 2014 May 29 [cited 2017 May 9];509(7502):622–6. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24717433 PMID: 24717433
12. Shmukler BE, Huston NC, Thon JN, Ni C-W, Kourkoulis G, Lawson ND, Paw BH, Alper SL. Homozygous knockout of the piezo1 gene in the zebrafish is not associated with anemia. Haematologica [Internet]. 2015 Dec 1 [cited 2017 May 9];100(12):e483–e485. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26294733 PMID: 26294733
13. Delmas P, Hao J, Rodat-Despoix L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci [Internet]. Nature Publishing Group; 2011 Mar 9 [cited 2017 May 9];12(3):139–153. Available from: http://www.nature.com/doifinder/10.1038/nrn2993
14. Vitzthum C, Clauss WG, Fronius M. Mechanosensitive activation of CFTR by increased cell volume and hydrostatic pressure but not shear stress. Biochim Biophys Acta - Biomembr [Internet]. 2015 Nov [cited 2017 May 9];1848(11):2942–2951. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26357939 PMID: 26357939
15. Beloussov L V. Morphogenesis can be driven by properly parametrised mechanical feedback. Eur Phys J E [Internet]. Springer Berlin Heidelberg; 2013 Nov 25 [cited 2017 Apr 30];36(11):132. Available from: http://link.springer.com/10.1140/epje/i2013-13132-x
16. Plouhinec J-L, De Robertis EM. Systems Biology of the Self-regulating Morphogenetic Gradient of the Xenopus Gastrula. Cold Spring Harb Perspect Biol [Internet]. 2009 Aug 1 [cited 2017 Apr 30];1(2):a001701–a001701. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20066084 PMID: 20066084
17. Raftery LA, Sutherland DJ. Gradients and thresholds: BMP response gradients unveiled in Drosophila embryos. Trends Genet [Internet]. 2003 Dec [cited 2017 Apr 30];19(12):701–708. Available from: http://www.ncbi.nlm.nih.gov/pubmed/14642751 PMID: 14642751
18. Reversade B, Kuroda H, Lee H, Mays A, De Robertis EM. Depletion of Bmp2, Bmp4, Bmp7 and Spemann organizer signals induces massive brain formation in Xenopus embryos. Development [Internet]. 2005 Jun 23 [cited 2017 Apr 30];132(15):3381–3392. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15975940 PMID: 15975940
19. Plouhinec J-L, Zakin L, De Robertis EM. Systems control of BMP morphogen flow in vertebrate embryos. Curr Opin Genet Dev [Internet]. 2011 Dec [cited 2017 Apr 30];21(6):696–703. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21937218 PMID: 21937218
20. De Robertis EM. Spemann’s organizer and the self-regulation of embryonic fields. Mech Dev [Internet]. NIH Public Access; 2009 Dec [cited 2017 May 9];126(11–12):925–41. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19733655 PMID: 19733655
21. Stottmann RW, Berrong M, Matta K, Choi M, Klingensmith J. The BMP antagonist Noggin promotes cranial and spinal neurulation by distinct mechanisms. Dev Biol [Internet]. 2006 Jul 15 [cited 2017 Apr 30];295(2):647–663. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16712836 PMID: 16712836
22. Ybot-Gonzalez P, Gaston-Massuet C, Girdler G, Klingensmith J, Arkell R, Greene NDE, Copp AJ. Neural plate morphogenesis during mouse neurulation is regulated by antagonism of Bmp signalling. Development [Internet]. 2007 Aug 1 [cited 2017 Apr 30];134(17):3203–3211. Available from: http://www.ncbi.nlm.nih.gov/pubmed/17693602 PMID: 17693602
23. Bond AM, Bhalala OG, Kessler JA. The dynamic role of bone morphogenetic proteins in neural stem cell fate and maturation. Dev Neurobiol [Internet]. NIH Public Access; 2012 Jul [cited 2017 May 9];72(7):1068–84. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22489086 PMID: 22489086
24. Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature [Internet]. Nature Publishing Group; 1996 Oct 3 [cited 2017 May 10];383(6599):407–413. Available from: http://www.nature.com/doifinder/10.1038/383407a0
25. Bae G-U, Domene S, Roessler E, Schachter K, Kang J-S, Muenke M, Krauss RS. Mutations in CDON, encoding a hedgehog receptor, result in holoprosencephaly and defective interactions with other hedgehog receptors. Am J Hum Genet [Internet]. Elsevier; 2011 Aug 12 [cited 2017 May 10];89(2):231–40. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21802063 PMID: 21802063
26. Golden JA, Bracilovic A, McFadden KA, Beesley JS, Rubenstein JL, Grinspan JB. Ectopic bone morphogenetic proteins 5 and 4 in the chicken forebrain lead to cyclopia and holoprosencephaly. Proc Natl Acad Sci U S A [Internet]. National Academy of Sciences; 1999 Mar 2 [cited 2017 May 10];96(5):2439–44. Available from: http://www.ncbi.nlm.nih.gov/pubmed/10051661 PMID: 10051661
27. Roessler E, Belloni E, Gaudenz K, Jay P, Berta P, Scherer SW, Tsui L-C, Muenke M. Mutations in the human Sonic Hedgehog gene cause holoprosencephaly. Nat Genet [Internet]. 1996 Nov [cited 2017 May 10];14(3):357–360. Available from: http://www.ncbi.nlm.nih.gov/pubmed/8896572 PMID: 8896572
28. Chen JK, Taipale J, Cooper MK, Beachy PA. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev [Internet]. Cold Spring Harbor Laboratory Press; 2002 Nov 1 [cited 2017 May 11];16(21):2743–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/12414725 PMID: 12414725
29. BINNS W, JAMES LF, SHUPE JL, THACKER EJ. Cyclopian-type malformation in lambs. Arch Environ Health [Internet]. 1962 Aug [cited 2017 May 11];5:106–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/13869306 PMID: 13869306
30. Keeler RF, Binns W. Teratogenic compounds of Veratrum californicum (Durand). I. Preparation and characterization of fractions and alkaloids for biologic testing. Can J Biochem [Internet]. 1966 Jun [cited 2017 May 11];44(6):819–28. Available from: http://www.ncbi.nlm.nih.gov/pubmed/5919283 PMID: 5919283
31. Taylor MD, Northcott PA, Korshunov A, Remke M, Cho Y-J, Clifford SC, Eberhart CG, Parsons DW, Rutkowski S, Gajjar A, Ellison DW, Lichter P, Gilbertson RJ, Pomeroy SL, Kool M, Pfister SM. Molecular subgroups of medulloblastoma: the current consensus. Acta Neuropathol [Internet]. Springer; 2012 Apr [cited 2017 Jan 21];123(4):465–72. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22134537 PMID: 22134537
32. Jain S, Song R, Xie J. Sonidegib: mechanism of action, pharmacology, and clinical utility for advanced basal cell carcinomas. Onco Targets Ther [Internet]. Dove Press; 2017 [cited 2017 May 11];10:1645–1653. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28352196 PMID: 28352196
33. Collier NJ, Ali FR, Lear JT. The safety and efficacy of sonidegib for the treatment of locally advanced basal cell carcinoma. Expert Rev Anticancer Ther [Internet]. 2016 Oct 2 [cited 2017 May 11];16(10):1011–1018. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27636236 PMID: 27636236
34. Tremblay MR, Lescarbeau A, Grogan MJ, Tan E, Lin G, Austad BC, Yu L-C, Behnke ML, Nair SJ, Hagel M, White K, Conley J, Manna JD, Alvarez-Diez TM, Hoyt J, Woodward CN, Sydor JR, Pink M, MacDougall J, Campbell MJ, Cushing J, Ferguson J, Curtis MS, McGovern K, Read MA, Palombella VJ, Adams J, Castro AC. Discovery of a Potent and Orally Active Hedgehog Pathway Antagonist (IPI-926). J Med Chem [Internet]. 2009 Jul 23 [cited 2017 May 11];52(14):4400–4418. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19522463 PMID: 19522463
35. De Smaele E, Ferretti E, Gulino A. Vismodegib, a small-molecule inhibitor of the hedgehog pathway for the treatment of advanced cancers. Curr Opin Investig Drugs [Internet]. 2010 Jun [cited 2017 May 11];11(6):707–18. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20496266 PMID: 20496266
36. Kaneko KJ, Kohn MJ, Liu C, DePamphilis ML. Transcription factor TEAD2 is involved in neural tube closure. genesis [Internet]. 2007 Sep [cited 2017 May 11];45(9):577–587. Available from: http://doi.wiley.com/10.1002/dvg.20330
37. Preiksaitiene E, Benusiene E, Ciuladaite Z, Sliuzas V, Mikstiene V, Kucinskas V. Recurrent fetal syndromic spina bifida associated with 3q26.1-qter duplication and 5p13.33-pter deletion due to familial balanced rearrangement. Taiwan J Obstet Gynecol [Internet]. 2016 Jun [cited 2017 May 11];55(3):410–414. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27343325 PMID: 27343325
38. Anderson MJ, Schimmang T, Lewandoski M. An FGF3-BMP Signaling Axis Regulates Caudal Neural Tube Closure, Neural Crest Specification and Anterior-Posterior Axis Extension. Wilkie AOM, editor. PLOS Genet [Internet]. 2016 May 4 [cited 2017 May 11];12(5):e1006018. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27144312 PMID: 27144312
39. Milunsky A, Jick H, Jick SS, Bruell CL, MacLaughlin DS, Rothman KJ, Willett W. Multivitamin/folic acid supplementation in early pregnancy reduces the prevalence of neural tube defects. JAMA [Internet]. 1989 Nov 24 [cited 2017 May 11];262(20):2847–52. Available from: http://www.ncbi.nlm.nih.gov/pubmed/2478730 PMID: 2478730
40. Roberts E, Hampshire DJ, Pattison L, Springell K, Jafri H, Corry P, Mannon J, Rashid Y, Crow Y, Bond J, Woods CG. Autosomal recessive primary microcephaly: an analysis of locus heterogeneity and phenotypic variation. J Med Genet [Internet]. 2002 Oct [cited 2017 May 11];39(10):718–21. Available from: http://www.ncbi.nlm.nih.gov/pubmed/12362027 PMID: 12362027
41. Lamb T, Knecht A, Smith W, Stachel S, Economides A, Stahl N, Yancopolous G, Harland R. Neural induction by the secreted polypeptide noggin. Science (80- ) [Internet]. 1993 Oct 29 [cited 2017 May 10];262(5134):713–718. Available from: http://www.sciencemag.org/cgi/doi/10.1126/science.8235591
42. Hemmati-Brivanlou A, Kelly OG, Melton DA. Follistatin, an Antagonist of Activin, Is Expressed in the Spemann Organizer and Displays Direct Neuralizing Activity. Cell [Internet]. 1994 [cited 2017 May 10];77:283–295. Available from: http://ac.els-cdn.com/0092867494903204/1-s2.0-0092867494903204-main.pdf?_tid=7806765c-35bd-11e7-940c-00000aacb361&acdnat=1494447533_a2c62319c30208326f4b1dc6dfb38b9f
43. Sasai Y, Lu B, Steinbeisser H, De Robertis EM. Regulation of neural induction by the Chd and Bmp-4 antagonistic patterning signals in Xenopus. Nature [Internet]. Nature Publishing Group; 1995 Jul 27 [cited 2017 May 10];376(6538):333–336. Available from: http://www.nature.com/doifinder/10.1038/376333a0
44. Bouwmeester T, Kim S-H, Sasai Y, Lu B, Robertis EM De. Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann’s organizer. Nature [Internet]. Nature Publishing Group; 1996 Aug 15 [cited 2017 May 10];382(6592):595–601. Available from: http://www.nature.com/doifinder/10.1038/382595a0
45. Hansen CS, Marion CD, Steele K, George S, Smith WC. Direct neural induction and selective inhibition of mesoderm and epidermis inducers by Xnr3. Development [Internet]. 1997 [cited 2017 May 10];124(2). Available from: http://dev.biologists.org/content/124/2/483.long
46. Cox J, Jackson AP, Bond J, Woods CG. What primary microcephaly can tell us about brain growth. Trends Mol Med [Internet]. 2006 Aug [cited 2017 May 11];12(8):358–366. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16829198 PMID: 16829198