Колонки новой коры
Обобщение фактических данных исследований по кортикальным колонкам новой коры
Эта статья – дальнейшая детализация представлений модели МВАП . Ее задача – добиться достаточной ясности в организации, функции и механизмах такой структурной единицы новой коры – как кортикальная колонка. Все, кто изучает МВАП и вообще системные принципы организации новой коры, могут получить достаточно ясное понимание.
В первую очередь в статье приводятся наиболее хорошо изученные и описанные свойства кортикальных колонок, что в целом составит аксиоматическую базу для обобщений.
В конце статьи будет представлен вариант такого обобщения.
Каждая приводимая статья сохранена на сайте и есть ссылка не ее источник. Здесь же выделено самой существенное. Текст статьи выделен коричневым. Самое важное выделено жирным.
НОВАЯ КОРА (neocortex) составляет 96% от всей поверхности полушарий. По морфологическим особенностям в ней выделяют 6 основных слоев, однако в различных областях коры количество слоев варьирует.
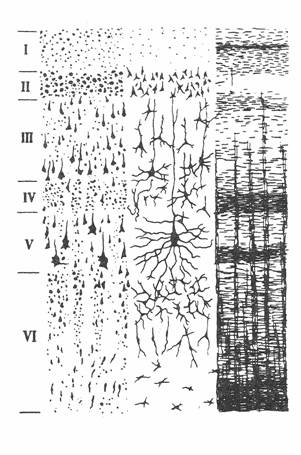
1 - МОЛЕКУЛЯРНЫЙ. Клеток мало, состоит в основном из горизонтальных волокон восходящих аксонов, в том числе неспецифические афференты от таламуса, а также в этом слое заканчиваются ветви апикальных (верхушечных) дендритов 4 слоя коры.
2 - НАРУЖНЫЙ ЗЕРНИСТЫЙ. Состоит из звездчатых и мелких пирамидальных клеток, аксоны которых заканчиваются в 3, 5 и 6 слоях, т.е. участвует в соединении различных слоев коры.
3 - НАРУЖНЫХ ПИРАМИД. Этот слои имеет два подслоя. Внешний- состоит из более мелких клеток, которые осуществляют связь с соседними участками коры, особенно хорошо развит в зрительной коре. Внутренний подслой содержит более крупные клетки, которые участвуют в образовании коммиссуральных связей (связи между двумя полушариями).
4 - ВНУТРЕННИЙ ЗЕРНИСТЫЙ. Включает клетки зернистые, звездчатые и мелких пирамид. Их апикальные дендриты поднимаются в 1 слой коры, а базальные (от основания клетки) в 6 слой коры, т.о. участвуют в осуществлении межкорковой связи.
5 - ГАНГЛИОЗНЫЙ. Его основу составляют гигантские пирамиды (клетки Беца). Их апикальный дендрит простирается до 1 слоя, базальные дендриты идут параллельно поверхности коры, а аксоны образуют проекционные пути к базальным ядрам, стволу и спинному мозгу.
6 - ПОЛИМОРФНЫЙ. В нем присутствуют клетки различной формы, но преимущественно веретенообразные. Их аксоны идут вверх, но в большей мере вниз и образуют ассоциативные и проекционные пути, переходящие в белое вещество головного мозга.
Клетки различных слоев коры объединены в "модули"-
структурно-функциональные единицы. Это группы нейронов из 10- 1000 клеток,
которые выполняют определенные функции, "обрабатывают" тот или иной
вид информации. Клетки этой группы преимущественно расположены перпендикулярно
поверхности коры и часто именуются "колонковые модули".
https://www.ok-t.ru/studopediaru/baza17/3804887305.files/image027.jpg
I. молекулярный
II. наружный зернистый
III. наружный пирамидный
IV. внутренний зернистый
V. ганглиозный ( гигантских пирамид)
VI. Полиморфный
В 1957 году Вернон Маунткасл, анализируя ответы клеток в соматосенсорной коре, обнаружил, что, если погружать микроэлектрод перпендикулярно поверхности коры, все встречаемые им нейроны отвечают на раздражитель одной и той же модальности (то есть одного вида чувствительности – например, кожной вибрации). Но как только он погружал электрод под углом к поверхности, на его пути попадались нейроны с другой сенсорной модальностью.
Из этого Маунткасл сделал вывод о том, что в коре есть некие вертикально ориентированные модульные структуры. Он назвал их колонками. В каждой такой колонке обычно присутствует около 100 нервных клеток (разных размеров и форм).
В центре колонок, как правило, находятся крупные пирамидные нейроны. Они представляют собой остов (основу) колонки. Считается, что крупные пирамидные нейроны отвечают за реакцию колонки в целом.
Над ними располагаются более мелкие клетки с отростками. Они называются короной. Внутри нее связи как бы замыкаются, формируя своеобразную память колонки. По замкнутым цепям короны нервный импульс может долго циркулировать, раздражая остов до тех пор, пока к нейронам не поступят тормозные импульсы.
Идущая в мозг информация через переключения в нижележащих глубинных структурах поступает в нижний слой коры. Здесь будут идентифицироваться самые простые свойства объектов: наклон линий, цвет, сила звука и так далее. Такая клетка может активно раздражаться, когда она видит линию или край, наклоненный под определенным углом. Этот край может быть частью любого объекта – клавиши пианино, стволом отдаленной сосны, стороной буквы «н».
Выше находятся нейроны, собирающие информацию от простых нейронов. Так происходит уточнение и обобщение получаемого сигнала внутри колонки. Но таким нейронам, например, важен не только контраст и наклон полоски, но и ее длина.
Информация, обработанная в нижнем слое, перемещается в вышележащий. При переходе от слоя к слою информация все больше обобщается. Часть ее отбрасывается. В конечном итоге наверху мы получаем целостный образ. Причем миниколонки объединяются в гиперколонки и за счет горизонтальных связей друг с другом обеспечивают еще более сложную комплексную обработку.
Давайте рассмотрим, как происходит восприятие зрительного образа.
В зрительной коре есть колонки, внутри которых находятся рецептивные (воспринимающие) поля нейронов. В сетчатке глаз есть палочки и колбочки со светочувствительными пигментами. Их можно считать рецепторами зрительной информации. Они собирают изображение по точкам. Рецептивное поле нейрона – это то, сколько рецепторов сетчатки отправляют сигнал.
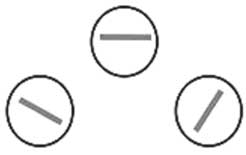
Рис. 17. Миниколонки, реагирующие на линии (полоски) определенной ориентации
Давайте вообразим, что мы смотрим на треугольный объект. В первичных областях зрительной коры найдется миниколонка, в которой нейрон среагирует на горизонтальную линию. В другой миниколонке нейрон откликнется на линию с наклоном под определенным углом. И так они вместе определят элементы треугольника. Схематично это показано на рис. 17. На самом деле нейроны в этих миниколонках будут различать не саму линию как таковую, а ее границу (за счет изменения контраста, теней и так далее). А вот целостную форму объекта и какие-то узоры (дополнительные линии, разные цвета, сложный объем и так далее) нейроны смогут «увидеть» только во вторичной области зрительной коры.
Во вторичных областях коры мы уже не встретим клеток, реагирующих на элементарные признаки (такие как граница или наклон линии). Здесь в колонках находятся нейроны, раздражающиеся только в ответ на определенный сложный комплекс стимулов. Известно, что в нижней височной доле существуют нейроны, которые активируются только при зрительном восприятии человеческого лица. Причем есть современные данные, указывающие на то, что это врожденная функция. Вероятно, именно поэтому на картинах, фотографиях и рекламных плакатах нас привлекают именно лица.
В свою очередь в третичных (ассоциативных) областях коры миниколонки, собираясь в модули, получают информацию от вторичных зон разных сенсорных систем. Здесь «смешиваются» зрение, обоняние, слух, осязание, вкус. При участии подкорковых структур мозга в ассоциативных областях формируется полноценное комплексное восприятие окружающего мира. Также здесь репрезентируются наши воспоминания. Они как бы собираются из элементов (представляются нам заново), поэтому мы и используем термин «репрезентируются». Формируется образ.
И тогда мы понимаем, что перед нами не просто треугольный объект, а египетская пирамида, а рядом с ней верблюды. И вообще, дело происходит в пустыне.
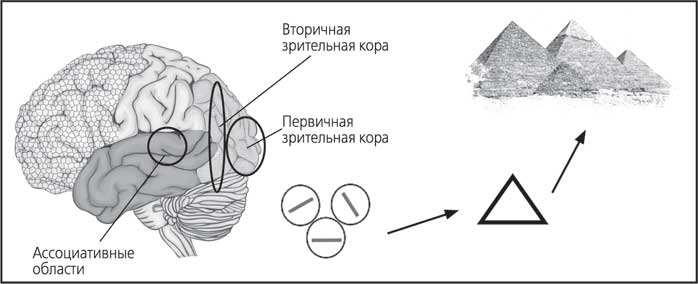
Рис. 18. Упрощенная схема поэтапной сборки изображения в коре мозга
Давайте подытожим. Мы выяснили, что кора имеет модульное
строение и состоит из миниколонок с сотней нейронов в каждой. Эти
миниколонки могут объединяться в гиперколонки и тем самым выполнять более
комплексные функции.
Мы также выяснили, что информация обрабатывается в колонках вертикально. Нижние
слои хранят самую примитивную информацию, тогда как верхние слои коры работают
с цельными образами. В первичных областях эти «цельные образы» отражают лишь
простые характеристики (наклон линий, движение линий, тени и так далее). Во
вторичных областях мы уже можем видеть объект более целостно, причем независимо
от его наклона (к примеру, мы можем распознать знакомое лицо, если даже голова
наклонена).
Таким образом, информация обрабатывается еще и горизонтально, перемещаясь по коре из первичных областей во вторичные, а оттуда – в ассоциативные. На каждом этапе перехода из одной области в другую происходит сборка все более цельного образа (пока из линий, кружочков, теней и прочего не соберется полноценная картина мира). Несомненно, это очень упрощенная схема восприятия.
… У нас есть нейроны-детекторы, которые отвечают за элементарные признаки воспринимаемого объекта. Они включаются в иерархически организованную нейронную сеть по типу пирамиды, вершина которой – сложный нейрон, синтезирующий воспринимаемый образ. Этот нейрон можно назвать главным. В первичных областях такой нейрон выполняет функцию попроще (контуры и тени линий), во вторичных областях он уже приобретает способность видеть целиком форму, цвет и даже лицо. А в третичных областях такой сложный нейрон будет собирать на себе уже информацию не только о форме, но и о запахах объекта, связанных с ним звуках и других характеристиках.
...Физиологи убедительно продемонстрировали, как формируется восприятие мира, в простом, но ошеломляющем своей показательностью эксперименте на котятах.

Рис. 19. Котенок, помещенный в цилиндрическую камеру с вертикальными полосами
Двухнедельных котят исследователи поместили в цилиндрическую камеру. Ее боковая поверхность была покрыта вертикальными полосами, снизу и сверху размещались зеркала. Это создавало иллюзию бесконечных вертикальных линий. Котята проводили в цилиндрической камере по пять часов каждый день. В остальное время они пребывали в темноте. Спустя пять месяцев выяснилось, что котята не воспринимают ничего, кроме вертикальных стимулов.
Такой же эксперимент провели с горизонтальными линиями (исключив все вертикальные). По окончании эксперимента котята не то что не реагировали на вертикальные стимулы, они врезались в столбы, в ножки столов, совершенно их не замечая! Самое страшное, что эти изменения оказались необратимы. Котята (а потом выросшие кошки) так никогда и не научились видеть вертикальные линии.
По Рис. 18. (Упрощенная схема поэтапной сборки изображения в коре мозга) понятно, что на уровне колонок новой коры не происходит инвариантное обобщение образа, т.е. детекция линии независимо от наклона и размера и т.п., а все промежуточные детекторы используются просто как рецепторные детекторы, т.е. как выделение определенных сочетаний признаков воспринимаемого.
Слои новой коры, последовательность развития
Кора головного мозга млекопитающих состоит из шести слоёв
клеток. Нейроны, образующие эти слои, заметно отличаются друг от друга.
Самый нижний, внутренний слой коры
взаимодействует со спинным мозгом и стволом головного: вместе с ними нижний
слой коры помогает управлять такими процессами, как, например, дыхание,
выполнение движений и т. д. Самый верхний — и самый большой — слой собирает и
объединяет информацию от органов чувств, координирует совместную работу
полушарий и выполняет ещё ряд функций. Верхний слой — эволюционно самый
молодой, его бурное развитие случилось у приматов, и во многом благодаря
верхнему слою коры человек получил способность мыслить абстрактно, решать
проблемы и планировать свои действия.
Каждый слой формируется в строго определённый
момент индивидуального развития, и
довольно долго считалось, что судьба любого нейрона, входящего в состав
коры, зависит от времени его появления на свет.
Каждая кортикальная колонка представляет собой ориентированный перпендикулярно поверхности (т.е. вертикально) цилиндр диаметром 300-500 мкм. Нейроны одной колонки получают сходные афферентные сигналы и образуют синапсы преимущественно друг с другом. В колонках проекционной коры имеются две разновидности пирамидных нейронов, которые получили название простых и сложных. Нейроны вертикальной колонки получают от проекционных нейронов таламуса афферентные сигналы, характеризующие определенную субмодальность. Несколько вертикальных колонок, связанных с разными субмодальностями, образуют функциональный модуль, способный перерабатывать всю информацию, связанную с различными субмодальностями. Информацию от соседних областей перерабатывают другие функциональные модули, организованные по тому же принципу. Взаимодействующие друг с другом модули образуют распределенные системы коры.
Простые нейроны, расположенные поблизости от входного IV слоя, выполняют функцию детекторов элементарных признаков раздражителя. Например, простые нейроны зрительной коры сильнее всего реагируют на линейный зрительный стимул в виде полосы или грани между светлой и темной плоскостями, т.е. реагируют на контраст. Друг от друга они отличаются по чувствительности к разным углам наклона линейного стимула. Аксоны нескольких однотипных простых нейронов конвергируют к сложным, или комплексным, нейронам, поэтому комплексные нейроны реагируют на стимул позже простых клеток.
Рецептивное поле сложных нейронов состоит из небольших рецептивных полей простых нейронов, а эффективными раздражителями для комплексных нейронов становится уже не линейный раздражитель определенной ориентации, а характер и направление его движения через рецептивное поле. Так осуществляется переход от детекции простых сигналов к абстрагированию, обобщению. В этом процессе участвуют интернейроны локальной сети кортикальной колонки, а итогом преобразования становится уровень активности выходных нейронов, аксоны которых покидают колонку, чтобы обеспечить ее взаимодействие с другими колонками. Каждая кортикальная колонка представляет собой функциональную единицу коры, или микромодуль.
С помощью горизонтальных связей соседние колонки объединяются в гиперколонку, или модуль. Если микромодули предназначены для распознания элементарных признаков раздражителя или образа, то модуль служит для комплексного объединения элементарных признаков.
Каждый проекционный нейрон таламуса получает информацию от рецепторов одной модальности и возбуждает входные звездчатые нейроны преимущественно одной кортикальной колонки, выбор которой подчинен принципу топографической организации связей сенсорной системы. Соответственно этому, для любой из соседних с этой кортикальных колонок тоже существуют собственные проекционные нейроны таламуса. Аксоны входных звездчатых нейронов оканчиваются в пределах одной кортикальной колонки, тогда как выходные нейроны этой колонки, представленные пирамидными клетками, обеспечивают связи различных участков коры между собой и с другими структурами мозга, в том числе и с таламусом. Таким образом, связь между таламусом и корой является двусторонней, что обеспечивает регуляцию получения информации по принципу обратной связи.
Исследование колончатой организации коры
Нейрональная колонка — это совокупность из нескольких десятков нейронов коры головного мозга, которая представляет собой функциональную единицу мозга, служащую для обработки информации в коре. Внутри колонки функции нейронов различаются, но соседние колонки работают сходно (в зрительной коре они могут обрабатывать изображение, в моторной — обеспечивать управление движением рук, а в височной — формировать ассоциативные связи).
…Компьютерное моделирование позволило исследователям создать статистическую модель структуры коннектома небольшого участка коры (по-английски он называется microcircuit, что достаточно сложно однозначно и благозвучно перевести на русский). Этот участок состоял из 298 кортикальных нейронов шести типов (каждый был взят из базы смоделированных ранее нейронов), расположенных в объеме, соответствующем колонке коры мозга крысы.
…Оказалось, что использованный способ моделирования описывает почти 75% связей, обнаруженных в живом мозге. Получается, что значительная часть синапсов образуется просто там, где отростки разных нейронов встречают друг друга.
… Работа показала, что знание конкретного положения нейрона внутри определенного слоя коры не является необходимым для моделирования его связи с соседями. Достаточно расположить нейроны разных типов в правильных слоях, с подходящей плотностью и в необходимом количестве.
Колончатая организация коры больших полушарий
Клетки, предназначенные для все более поверхностных слоев, возникают в правильной временной последовательности: новая кора строится «изнутри кнаружи».
… В результате клетки коры располагаются радиально ориентированными тяжами, или колонками, пересекающими кору, и по существующему предположению клетки каждой такой колонки представляют собой единый клон.
1. Кортикальная колонка представляет собой обрабатывающее устройство с входом и выходом. Число других участков, передающих сигналы в традиционно определяемую область коры и получающих их от нее, может варьировать приблизительно от 10 до 30. Выборка, образуемая любой данной подгруппой модулей из этой области, как правило, гораздо меньше и варьирует от подгруппы к подгруппе, перекрываясь.
2. Расположение в виде колонок делает возможным картирование одновременно нескольких переменных на двумерной матрице.
3. Между упорядоченными группами колонок в разных областях коры и между группами корковых колонок и модулями подкорковых структур имеются специфические связи. Таким образом, при прохождении через такие области и между ними могут сохраняться топологические отношения с топографическим (географическим) картированием или без него.
4. Параметры, по которым идентифицируются колонки и упорядоченные группы колонок, могут варьировать внутри данной традиционно определяемой области коры резко различаться в разных областях.
5. Колончатая функциональная модель допускает частично сдвинутое перекрывание относительно топографического представительства, совместимое с динамической изоляцией активных элементов колонки посредством определенной формы латерального торможения вокруг колонки.
6. Внутренние пути колонки, дивергирующие к разным выходам, делают возможной селективную обработку («извлечение свойств») определенных параметров входного сигнала для определенного назначения на выходе.
Исследование первой соматосенсорной области у наркотизированных кошек принесло доказательства того, что основной функциональной единицей новой коры является вертикально ориентированная колонка (или цилиндр, или пластинка), идущая через все клеточные слои. Такая колонка способна выполнять функции вход — выход значительной сложности независимо от горизонтального распространения активности в сером веществе. Эта гипотеза подтвердилась и расширилась в исследованиях гомологичной области коры наркотизированных обезьян. Параметрами для идентификации колонок в соматосенсорной коре служат статические свойства нейронов: расположение их рецептивных полей в двух измерениях поверхности тела; характер стимулов, способных вызвать их возбуждение, называемый свойством «модальности»; скорость адаптации к постоянному стимулу — свойство, которое определяется на уровне сенсорного рецептора и таким образом является компонентом модальности.
…Пока что динамика нейронной переработки информации в колонках соматосенсорной коры исследована только предварительно. Имеются данные о том, что активность в одной колонке ведет к торможению нейронов в соседних колонках как той же, так и других модальностей.
…форма тела картирована на постцентральной соматосенсорной коре примата с сохранением топологической последовательности. Каждая маленькая часть сегментарной иннервации проецируется на длинную, иногда н извилистую, полоску коры, идущую в передне-заднем направлении; полоски варьируют как по длине (в передне-заднем направлении), так и по ширине (в медиально-латеральном направлении). Таким образом, любой локус на коре характеризуется параметрами места по своим осям X и Y. Кроме того, каждая колонка характеризуется статическим параметром модальности.
…Сочетая электрофизиологические методы с рядом экспериментальных анатомических подходов, Хьюбел и Визел показали, что колонки 17-го поля «образуют расположенные вертикально чередующиеся пластинки для левого и правого глаза. Эти пластинки на горизонтальных срезах имеют вид чередующихся полосок диаметром около 400 мкм, которые изредка дихотомически делятся и имеют слепые окончания». Приблизительно под прямым углом к этим полоскам доминантности глаза лежат полосы гораздо более узкие, клетки которых избирательно настроены на ориентацию коротких прямолинейных отрезков, таким образом, каждый член локального блока колонки доминантности содержит полную 180-градусную последовательность тонких ориентационных колонок (или полосок). Для каждого данного нейрона доминантность правого или левого глаза определяется афферентным входом, но степень этой доминантности и ориентационные свойства определяются внутрикорковыми процессами. Такая перекрестная комбинация бинокулярных и ориентационных групп занимает площадь около 800Х800 мкм. Я называю такую группу макроколонкой, это результат наложения друг на друга гиперколонок доминантности глаза и ориентационных, описанных Хьюбелом и Визелом. Таким образом, зрительная кора служит преимущественным примером того, как ряд переменных может быть картирован или представлен в двумерной матрице благодаря организации в форме колонок. Два измерения представительства пространства — поля зрения — картированы конгруэнтно с переменными доминантности глаза и ориентации и, несомненно, также с другими переменными (см. ниже).
Совместимость принципа колонок и принципа частично сдвинутого перекрывания проявляется в зрительной коре так же ясно, как в соматосенсорной. Здесь объединенное рецептивное поле каждой макроколонки перекрывает приблизительно наполовину поле соседних групп, а линейное отношение между фактором увеличения и размером рецептивного поля означает, что этот фактор перекрывания остается неизменным по всему представительству поля зрения в коре. Таким образом, любой ориентированный отрезок линии, предъявленный в поле зрения, будет максимально представлен в группе макроколонок, положение которых определяется местом в пространстве линейной протяженностью стимула. В пределах этой селективной группы макроколонок участок максимальной активности определяется ориентацией. Частично сдвинутое перекрывание представительства зрительного пространства приводит к тому, что по обеим сторонам ряда максимально активированных макроколонок будут находиться менее активированные группы — латеральное распространение активности будет резко ограничено торможением.
…Исследования динамической активности зрительно-корковых нейронов позволяют думать, что в любой макроколонке обработка информации о разных свойствах стимула происходит в параллельных каналах. Общим правилом, по-видимому, является то, что в каком-либо пути, ведущем к выходному каналу, отбираются те свойства, которые дальше обрабатываются в той области, куда ведет этот канал: происходит комбинация обрабатывающей распределительной функции корковой колонки.
…Итак, в стриарной зрительной коре проявляются некоторые общие свойства колончатой организации.
1. В ее двумерной матрице картировано несколько переменных.
2. Ее макроколонки функционируют как такие устройства, ведущие обработку между входом и выходом, при которых параллельная обработка в каждом из них делает возможной селекцию некоторых динамических свойств стимула для формирования в ограниченных каналах, ведущих к определенным выходам (обычно эти свойства усиливаются в перерабатывающих механизмах, находящихся в областях куда направлены эти каналы).
3. Группы ее колонок избирательно связаны с упорядоченными группами колонок в других областях коры в модулях подкорковых структур, на которые она проецируется.
4. Зрительная кора обезьяны подчиняется своей геникулостриарной проекции, и этим она сходна с представительствами кисти и стопы в соматосенсорной коре по степени изоляции входа.
5. Колончатая организация зрительной коры совместима с принципом частично сдвинутого перекрывания в представительстве полей зрения в пространстве нервной системы.
…Общая концепция колонок не исключает возможности того, что в кору проникают и действуют там разные другие системы, в особенности такие, которые выполняют общие регуляторные функции, а не детальную обработку информации.
…Я обозначаю основную модульную единицу новой коры как мини-колонку. Это вертикально ориентированная цепь клеток, образованная миграцией нейронов из зародышевого эпителия нервной трубки вдоль, радиальных глиальных клеток к местам их назначения в коре, как это описано Ракичем. Если эта мини-колонка сравнима по величине с корковыми цилиндрами, в которых Рокел и др. производили подсчеты нейронов, то она содержит около 110 клеток. Эта цифра остается почти неизменной в разных областях новой коры и у разных видов млекопитающих, за исключением стриарной коры приматов, где она составляет 260. Такая цепь клеток занимает в пространстве коры слегка изогнутый, почти вертикальный цилиндр диаметром около 30 мкм. Общий объем новой коры человека составляет около 1000 см3. Если принять ее среднюю толщину равной 2500 мкм, то тогда новая кора головного мозга, человека имеет поверхность около 4000 см2 и содержит около 600 млн. мини- колонок и примерно 50 млрд. нейронов.
…Важным свойством внутрикорковых механизмов обработки является то, что я назвал торможением вокруг колонки. Это мощный механизм функциональной изоляции активных колонок от их соседей, который приводит понятие о колончатой организации в соответствие с понятием о частично сдвинутом перекрывании. Это последнее представляет собой общий принцип, согласно которому, стимул ограниченных размеров, сдвигаясь понемногу по рецепторному слою, приводит в действие ряд сдвинутых, но перекрывающихся популяций корковых элементов. Торможение вокруг колонки должно резко ограничивать латеральное распространение активности из групп колонок, наиболее сильно активированных таким локальным стимулом. Этот принцип, несомненно, сохраняет свое значение также и для других областей коры, помимо сенсорных.
Структура неокортекса достаточно однородна. Вертикально нейроны некортекса объединены в так называемые кортикальные колонки (синонимы колонки котрекса, гиперколонки, кортикальные модули). Нейроны, которые составляют кортикальную колонку, находящиеся на разных уровнях по высоте, выполняют схожие функции и объединены между собой, образуя горизонтальные слои неокортекса. У мозга крысы таких слоёв четыре, у высших млекопитающих в неокортексе 5 слоёв, у мозга человека количество слоёв максимально и равняется шести.
…существовало два взгляда на то, как устроен и функционирует головной мозг и его кора. Согласно первому, традиционному взгляду, связи между нейронами не являются случайными и при анализе деятельности мозга каждая из связей является функциональной и оказывает влияние на конечный результат. Такой взгляд возник не на пустом месте, поскольку у низших животных их нейронная сеть работает именно таким образом. Например, у улитки вся высшая нервная система, которая обеспечивает её достаточно сложное поведение, состоит всего из восьми нейронов.
С точки зрения изучения и создания искусственного мозга получить ответ на то, по какому из вариантов строится неокортекс, очень важно. Общее количество нейронов в головном мозге человека порядка 100 млрд., каждый из которых может образовывать до 10 тысяч связей с другими нейронами. Если верен первый вариант и каждая из связей функциональна, то мы получаем информационную емкость порядка 10 в степени 14 (1e14). Если же верен второй вариант, то это означает, что нейроны в коре образуют функциональные макроструктуры, каждая из которых включает в себя тысячи нейронов и миллионы связей между ними. Другими словами, каждая кортикальная колонна представляет собой аналог микросхемы в микроэлектронике, внутри которой могут находиться милионы полупроводниковых элементов. Но для проектирования и понимания того, как будет функционировать конечное устройство, в котором используются подобные микросхемы, нет необходимости знать и понимать то, как работает каждый транзистор в этой микросхеме. Переход на изучение функционирования мозга на уровне макроструктур неокортекса позволил бы на 4-5 порядков уменьшить информационный объём модели, то есть для мозга человека вместо 1e14 мы уже получаем 1e10 или даже 1e9. А это очень существенно, поскольку с точки зрения анализа и моделирования уровень 1e10 технологически достижим уже сейчас, в то время как до уровня 1e14 нам ещё достаточно далеко.
…тот результат, который был получен, однозначно говорит о том, что структура коры головного мозга млекопитающих строится по второму типу. Это означает, что при дальнейшем изучении и моделировании деятельности мозга наша задача значительно упрощается, поскольку мы можем переходить на изучение и моделирование макроструктур, которые образуются нейронами, а не изучать поведение каждого нейрона в отдельности.
…форма и структура синапсов у тех нейронов, которые образуют неокортекс млекопитающих, не является случайной. Во время роста и образования связей данные нейроны подчиняются некому единому плану, воспроизводят на макроуровне определённую заранее заданную форму и структуру.
…в множество нейронов в одной кортикальной колонке выполняют одну и туже функцию, многократно дублируя и усиливая друг друга, как множество клеток в одной мышце, многократно дублируют друг друга и увеличивают общую силу мышцы, которую они совместно образуют.
Подобный способ построения из множества нейронов кортикальной колонки также говорит о том, что она является универсальным вычислительным элементом, перестройка структуры которой за счёт изменения связей между образующими её нейронами, либо очень сложна и длительна по времени, либо вообще невозможна в принципе, поскольку для изменения логики её работы необходимо, чтобы соответствующие связи были перестроены между всеми образующими её и дублирующими друг друга нейронами. В противном случае её функционирование в процессе перестройки связей будет невозможно ни согласно старой функции, которую данная колонка выполняла до начала перестройки связей, ни согласно новой функции, для выполнения которой связи должны принять новую конфигурацию.
Все виды сенсорных модальностей обрабатываются схожим образом: рецепторное поле проектируется на (внутреннюю) поверхность слоев коры причем так, что для каждого рецепторного сигнала оказывается больше количество входных нейронов обработчиков (сколько обычно исчисляется в одной микроколонке).
Совершенно не имеет значения в какую часть коры будет направлен пучок сенсорных аксонов для обработки потому, что важно только то, чтобы отклик активности коррелировал с реальным воспринимаемым стимулом, разложенным матрицей сенсоров со взаимным торможением и как-то обработанным.
В 30-х гг. XX столетия У. Пенфилд, базируясь на результатах, полученных во время нейрохирургических операций, установил наличиеправильной пространственной проекции соматических мышц различных отделов тела на двигательную область коры:
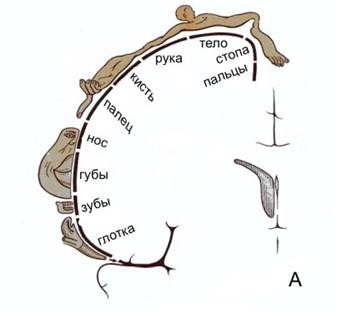
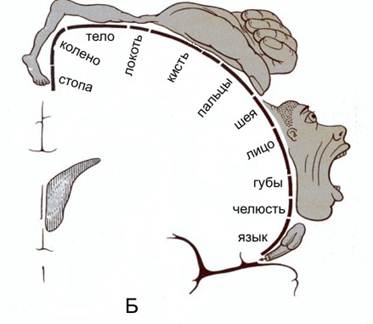
Если в случае проекции тела на кору все хорошо понятно т.к. каждый сенсор тела оказывает постоянно спроецирован на определенный участок коры, то в случае слуха и, тем более, зрения, активность сенсоров соответствует самым разным воздействующим сочетаниям элементов внешней среды: одна и та же матрица проекции на кору коррелирует с самыми разными сочетаниями распределения интенсивностей, цвета(высоты звука).
Рассмотрим данные о том, как проецируются на новую кору зрительные сигналы.
Зрительный путь – это система соединенных друг с другом нервных клеток, за счет которых человек видит. Основой органа зрения является рецептор, оснащенный фоторецепторными клетками. К клеткам можно отнести колбочки и палочки, от которых будет зависеть трансформирование светового пучка в электрический импульс.
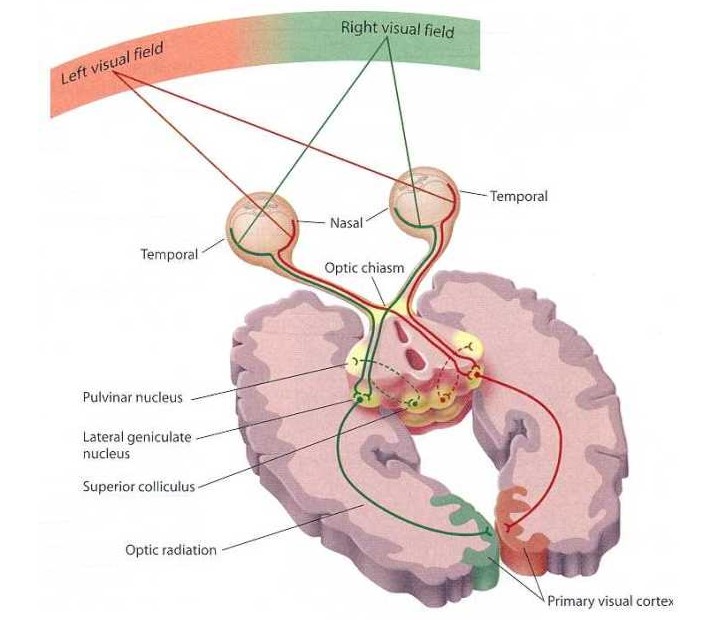 Строение зрительного пути
Строение зрительного пути
После своего образования импульсы будут проходить через промежуточные нервные клетки и попадают сначала в первичный зрительный центр. Зрительный центр обеспечивает образование (безусловно) рефлекторных реакций на внешнее световое воздействие глаз. Затем электрические импульсы проходят дальше и постепенно достигают коры головного мозга – ее центрального отдела. В этом отделе мозга происходит анализ поступающих импульсов. После этого работа всей нервной системы приведет к окончательному формированию действительности, то есть видимой картинки.
Строение зрительного пути
Начало зрительного пути происходит в сетчатке глаза. Первыми его нервными клетками считаются фоторецепторы. Благодаря сложным химическим изменениям фоторецепторы переводят поступающие извне световые волны в электрические импульсы. Затем путь нервных импульсов лежит в ганглиозные и биполярные клетки. Они также расположены в сетчатке. Биполярные клетки в свою очередь относятся ко второму звену зрительного пути, а ганглиозные к третьему звену.
Ганглиозные клетки имеют длинные отростки. Их основной функцией считается сбор информации со всей поверхности сетчатки. Затем все аксоны объединяются между собой и их количество достигает 1 миллиона. Объединение аксонов приводит к формированию зрительного нерва.
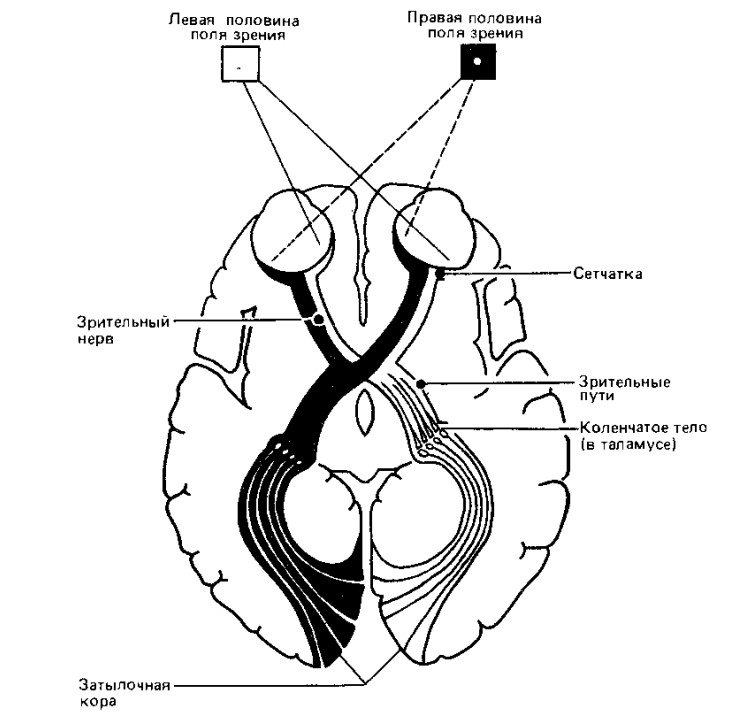 Особенности
строения зрительного пути
Особенности
строения зрительного пути
Отдельные аксоны, собранные в группы, располагаются по строгому порядку. Особое значение для зрительной функции имеет папилломакулярный пучок. Именно он переносит сигнал, собирая их с макулярной области. Папилломакулярный пучок располагается в наружной части зрительного нерва. Зрительный нерв попадает в область черепа через специальный зрительный канал. Нерв располагается над турецким седлом, где перекрест волокон двух зрительных нервов образует хиазму. В хиазме нервные волокна частично перекрещиваются.
Постепенно переходя в другую половину нервные волокна соединяются с волокнами, которые идут от наружных участков сетчатки, но уже от другого глаза. Хиазма снаружи имеет границу с внутренними сонными артериями.
Затем зрительные тракты огибают ножки мозга и оканчиваются в наружном коленчатом теле зрительного бугра и в переднем четверохолмии. Нервные клетки коленчатого тела относятся к первичному зрительному центру. Именно здесь формируется первичное ощущение света. Большей частью это неосознанное ощущение необходимо для осуществления рефлекторных реакций. Примером этого процесса также может служить поворот головы в одну из сторон в ответ на резкую вспышку яркого света. В наружном коленчатом теле также имеется определенная группа клеток, от которых начинается зрительная лучистость. Функция этой лучистости заключается в передаче информации, то есть в кору головного мозга. Отдел головного мозга, который полностью отвечает за зрение находится в шпорной борозде затылочной доли. Именно здесь располагается зрительный центр, нервные импульсы, идущие от сетчатки окончательно расшифровывается.
Так же см. Кортикальная проекция сетчатки:
…верхние квадранты сетчатки проицируются на дорзальную (со стороны спины) часть area striata (первичная зрительная зона), а нижние квадранты сетчатки — на ее вентральную (обращенный к брюшной поверхности) часть.
Обобщенные сведения
Слои созревают строго поочередно, в соответствии с эволюцией возникновения слоев у предков.
Колонки образуются поэтапно, с каждым новым созревшим слоем, дополняя и улучшая выделение совокупности признаков восприятия.
Вход колонки – самый нижний в коре (самый древний) слой. Выход – на самом верхнем (молодом) слое.
Из верхнего слоя аксоны возвращаются вновь вглубь мозга и проходят мимо первого слоя, что дает возможность замыкать выход на вход и самоподдерживать циркулирование активности. Так же аксоны идут к соседним колонкам и к соседним зонам мозга.
На уровне колонок новой коры первичной и вторичных зон не происходит инвариантного обобщения образа (т.е. детекции линии независимо от наклона и размера и т.п), а на уровне третичной теменной коры – происходит в виде рефлексов (когда к уже имеющемуся образу восприятия в данном контексте значимости добавляется еще один профиль входной активности – нового инварианта образа) и на уровне третичной лобной коры в виде автоматизмов (когда при осмыслении к уже имеющейся модели понимания добавляется новый инвариант).
У колонок оказываются две функции: выделение актуальных для окружения индивида совокупности восприятия и удержание их в активном состоянии на время доминирования по отношению с соседями, пока автоподдержание активности не будет прервано торможением соседями.
Таким образом, колонки, являющиеся элементом обработки данной модальности признаков восприятия, и являются источников все более разнообразных образов восприятия, которые оказывается способен различить мозг и там самым, может полагаться на каждый из них в поведенческой регуляции. И это универсально происходит в любой зоне новой коры.
Увеличение числа слоев новой коры эволюционно остановилось на шести, что говорит об отсутствии качественных преимуществ при большем числе для одной модальности восприятия, а дальнейший выигрыш возникает от сведения модальностей уже разных видов сенсорики на уровне вторичной коры и, затем на уровне третичных (ассоциативных) зон коры.
Разработка второй версии прототипа системы индивидуальной адаптивности показало конкретную причину фиксированного числа слоев. Оказалось, что использование уникальных идентификаторов образов восприятия различного уровня иерархии позволяет распознавать текущее состояние условий с помощью фиксированного числа узлов в каждой ветке дерева распознавания. При использовании самих образов с различным числом признаков на входе каждого, дерево бы получилось в неопределенным числом узлов в каждом случае и его сложность сделала бы затруднительным и медленным распознавание. Так что фиксированное число слоев коры четко коррелирует с применением уже сформированных видов детекторов образов при распознавании актуального их сочетания.
Становится достаточно убедительной картина того, что кора имеет функциональность именно древовидной функции распознавания.
Самоподдерживающаяся активность на уровне отдельных колонок обеспечивает некоторое “послесвечение” стимула, пока оно не будет погашено конкурирующими стимулами. Это делает более уверенным распознавание примитивов восприятия и есть время на то, чтобы на них как-то отреагировать, что дает эволюционное преимущество.
На примере зрения прослеживается общая системная последовательность эволюционного наращивания эффективности восприятия.
Первые одиночные фоторецепторы позволяли определять только степень яркости в очень размытом направлении. При этом яркость быстро истощала фоторецептор (потому, что нет соседей с латеральным торможением, обеспечивающим определение относительной яркости).
Появление соседних фоторецепторов позволило лучше различать яркость и, возможно, улучшалась избирательность к длине волны света (цвет).
Если бы у слепого был хотя бы один фоторецептор, различающий красное и зеленое, то он уже мог бы ориентироваться какой сигнал светофора светит. Это дает представление о том, насколько расширяет возможности каждое новое качество восприятия.
Появление фасеточных глаз позволило определять направление светящегося объекта и выделять разности яркости между фасетами, что позволяет формировать первые детекторы примитивов восприятия.
Появление фокусирующий линзы дало новое качество в детализации.
Далее прогресс идут уже не столько в видах сенсорных органов зрения, сколько изощренности обработки уже достаточно детализованных изображений – на уровне слоев новой коры мозга. И с каждым таким слоем возникало новое качество возможности такой обработки. Это соответствует принципу схемотехники: все возможности устройства в первую очередь лимитируются разрешением используемых сенсоров. Поэтому задача эволюции сенсорных систем – разбить параметр воздействия внешней среды на как можно большее число деталей (элементарных точек) и обеспечить их максимальное соответствие воздействию среды.
Итоговые выводы
У разных видов животных очень по-разному используются доступные им модальности восприятия и, соответственно, участие сформированных примитивов восприятия. При этом могут очень по-разному пролегать нервные пути, проецирующие на кору имеющиеся модальности. Это значит, что, собственно, нет принципиальной разницы, что именно используется и как именно реализовано это использование.
Важно только одно: способность улавливать значимые для жизни особи сочетания воздействующих признаков восприятия и использовать их как “внутренние органы” восприятия: органы, распознающие зрительные, слуховые, тактильные, запаховые образы и их сочетания, соответствующие элементам внешней реальности.
Системным (независимым от особенностей вида животных) оказывается лишь сам принцип постепенного образования иерархии примитивов восприятия, усложняющимися образами, адекватными проявлениям реальности, - выделение коррелятов реальности или абстрагирование.
Это значит, что при системном обобщении можно не особенно заморачиваться вопросами как именно реализовать схемы таких выделений коррелятов реальности, а важно лишь обеспечение их индивидуальное формирование у особи, обеспечивающие ей максимальное соответствие особенностям ее окружения.
В контексте дополнения уже имеющегося каркаса модели индивидуальной адаптивности МВАП , данное обобщение не меняет уже сформированные системы принципов построения и функционирования модели, хотя и привносит следующее:
1. Особенности организации параллельной обработки признаков восприятия в иерархию примитивов – с определенными количественными характеристиками, которые, видимо, хорошо оптимизированы эволюцией, то, что в текстах МВАП звучало как “пятно” за счет перекрытия соседей распознавателей данных примитивов – альтернативное привычной схемотехнике, где все цепи никак не дублируются.
Один аксон сенсора проецирует активность на целую колонку с множеством нейронов и еще наполовину соседние колонки. Потеря одного такого аксона так же мало значит из-за того, что эти аксоны, в свою очередь, так же избыточны. Для целей искусственной реализации на более надежных элементах такой избыточности не требуется потому как она не дает лучшее выделение примитивов, хотя возникает некое статистическое усреднение откликов нейронов в колонке, - это похоже на то, как избавляются от зернистости и дрожи в фотографии, делая большое число снимков, а потом усредняя их в фотошопе (накладывая слои всех фото).
2. Полезность самоподдержания активности примитивов восприятия на время пока ее не погасят конкурирующие соседи.
3. Оптимизированное эволюцией число слоев усложнения примитивов – не более шести, а дальнейшая обработка переносится на следующие (по эволюционному порядку возникновения) зоны, в которых точно так же используются колонки.