К ВОПРОСУ О КРИТЕРИЯХ НАДЁЖНОСТИ И ФАКТОРАХ КОЭВОЛЮЦИИ ПАРТНЁРОВ ДИНАМИЧЕСКОЙ СИСТЕМЫ РАСТЕНИЕ-ХОЗЯИН - ПАРАЗИТ
Относится к разделу Биология
ВВЕДЕНИЕ
В настоящее время все больше внимания уделяется системному подходу в исследованиях биологических процессов. Наиболее общее определение понятия «система», принадлежащее Леону фон Берталанфи, гласит: «Система может быть определена как совокупность элементов, находящихся в определенных отношениях друг с другом и со средой» (57). Динамическая система, по Н.Ф.Реймерсу (38), - это саморазвивающаяся и саморегулируемая совокупность организмов, функционирующая как относительно устойчивое единое целое за счет взаимного обмена информацией и обеспечивающая преобладание внутренних связей над внешними.
Динамическая система растение-хозяин - паразит является составной частью глобальной экологической системы – аэробной биосферы, существование которой связано с потреблением кислорода. Для изучения жизнеспособности партнеров динамической системы хозяин-паразит, находящихся в определенных отношениях друг с другом (совместимость и несовместимость), и оценки функционального состояния системы необходим двусторонний анализ соотношения с фактором среды – кислородом: 1) потребление кислорода в окислительных деструктивных процессах; 2) эффективность утилизации кислорода и трансформации энергии в биосинтетических антиокислительных процессах.
Сохранение и активация энергетического обмена, компенсаторная способность мембранных структур, сопряженная с ферментными системами, ответственными за процессы дыхания, аккумуляции энергии и биосинтетические процессы, могут служить ключевыми моментами в механизмах устойчивости и надежности защитных реакций высших растений и системы хозяин-паразит в целом.
Физиолого-биохимические исследования иммунитета растений показали, что их устойчивость связана не только со структурной стабильностью мембран (6, 18, 48, 49), но и со способностью мембранных структур «узнавать» патоген, т.е. с чувствительностью к патогену или его метаболитам (12, 20, 31, 56, 61, 86).
Чувствительный ответ растения на воздействие патогена, скорость ответной реакции, несомненно, связаны с функциональным свойством мембран – реактивностью. Изучение совокупности мембранных процессов деструкции и синтеза – окислительных и антиокислительных – в динамике взаимоотношений партнеров системы растение-паразит поможет выявить кинетические закономерности механизмов защитных реакций в целях сохранения и развития функциональной целостности и структурной индивидуальности организмов – основной задачи фитоиммунитета.
Исследованию мембранных механизмов реактивности, структурно-функциональных критериев надежности фитоиммунитета и природы движущих факторов сопряженной эволюции партнеров динамической системы растение-хозяин - паразит и посвящена настоящая работа.
1.Окислительные и антиокислительные процессы в динамике взаимоотношений растения с патогеном.
Первичная реакция растения-хозяина при контакте с патогеном проявляется на уровне мембран (21, 26), а интенсивность ответа на патоген сопряжена с энергетикой клетки и характером мембранного потребления кислорода (15, 16, 26, 39, 58, 80).
Многочисленные исследования показали, что универсальной ответной реакцией растения на воздействие патогенов (вирусов, грибов, бактерий) является изменение дыхательной активности тканями растения-хозяина (18, 19, 40, 48, 49, 56, 77). Однако кинетические закономерности сдвигов в дыхании инфицированных растений – их направленность, интенсивность и продолжительность – оказались различными в адаптивных перестройках для толерантных организмов и в патологических – для восприимчивых растений. Кинетика изменения дыхательной активности, исследованная на ядрах из листьев различных по устойчивости видов табака в динамике развития вирусной инфекции, показала, что патогенез у восприимчивого вида табака, сопровождается снижением интенсивности дыхания ядер через 12 ч. после инфицирования вирусом табачной мозаики (ВТМ) и возрастанием уровня дыхания значительно выше контрольного на 7-10-е сутки после заражения. В динамике адаптации устойчивых растений табака к инфекционной нагрузке вирусом через 12 ч. после инфицирования имеют место повышение интенсивности дыхания ядер и последующее снижение потребления кислорода ниже контрольного значения (39). Дифференциальный анализ изменений дыхательной активности ядер табака на ранних этапах взаимодействия растений с патогеном обнаруживает зеркально-антибатное соотношение между кинетиками потребления кислорода в адаптивных и патологических процессах (рис.1).
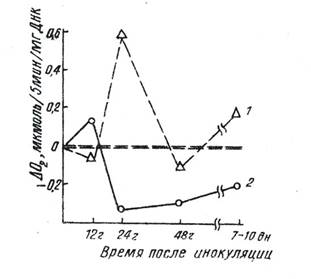
Рис.1. Кинетика изменений дифференциальных значений интенсивности дыхания ядерной фракции листьев восприимчивого (1) и устойчивого (2) видов табака по сравнению с контролем в динамике вирусной инфекции (по данным [39]).
Противоположная направленность характера потребления кислорода ядрами контрастных по устойчивости растений, по-видимому, может быть следствием различных функциональных, свойств структур клетки, т.е. их реактивности.
Потребление кислорода мембранными компонентами может также характеризоваться уровнем перекисного окисления липидов (ПОЛ) мембран и содержанием свободных радикалов (СР), концентрация которых свидетельствует об интенсивности деструктивных окислительных процессов (ДОП) и является специфической для поддержания стационарности ПОЛ определенных мембранных структур (10, 24).
Влияние фитопатогенов на интенсивность СР-окисления в растениях изучалось многими исследователями. Увеличение концентрации СР в тканях устойчивых растений по сравнению с восприимчивыми наблюдали при заражении табака вирусом табачной мозаики (23, 40), картофеля – возбудителем рака (25), пшеницы – ржавчинным грибом (29), гороха – возбудителем бактериального рака (54).
Исследование кинетики изменения концентрации парамагнитных центров (ПЦ) в различных по вилтоустойчивости сортах хлопчатника на протяжении трёх недель после инфицирования грибом VerticilliumdahliaeKI. показало, что адаптивная реакция устойчивых растений сопровождается повышением концентрации СР в первую неделю после заражения. В этот период в восприимчивых к заражению грибом растениях хлопчатника достоверно снижается уровень СР и парамагнитных комплексов двухвалентного марганца (ПК) Мn+2 (32, 33). Кинетика изменений концентрации ПК инфицированных растений имеет сходный характер как для пластосемихинонных СР (g=2,00), так и для ПК Мn+2 (g=2,157).
Симбатность кинетических кривых накопления СР и ПК Мn+2 свидетельствует o сопряженной связи пула ПК Мn+2 с функциональной концентрацией СР (33, 41). Поэтому сигнал ПК Мn+2, как и СР, может служить маркером интенсивности ПОЛ и ДОП (35). Усиление ДОП, возможно, способствует сдвигу подвижного равновесия ди- и тетрамеров марганцевых кластеров со смешанной валентностью фотосистемы–2 с увеличением концентрации ПК Мn+2(88). Кинетические данные изменения уровня СР процессов на ранних этапах инфицирования растений патогеном подтверждают симбатность кинетических кривых потребления кислорода в процессах дыхания и ДОП, хотя структурная разобщенность дыхательных центров и центров локализации реакций СР перекисного окисления мембранных липидов, может обусловить временнỳю дистанцию в реализации их действия (46).
В механизмах взаимоотношений растений с фитопатогенами имеют значение не только характер потребления кислорода путем митохондриального дыхания, микросомального гидроксилирования, мембранного ПОЛ и СР-окисления, но и способы компенсации сдвигов антиокислительной (АО) системой клетки (8-10).
На ранних этапах взаимоотношений в системе растение-хозяин - патоген у растения хлопчатника, инфицированных вертициллёзом, были изучены динамика окислительных процессов по изменению концентрации ПК и характер их компенсации - по кинетике изменения уровня антиокислительной активности (АОА) (35). Взаимоотношения партнеров в системе устойчивое растение – патоген приводили к адаптивным перестройкам у растений, а в системе восприимчивое растение - патоген – к патогенезу хозяина. Согласно результатам изучения кинетики окислительых и АО-процесов, у растений восприимчивого сорта хлопчатника С-4727 (рис.2) и у толерантного сорта Кзыл-Рават (рис.3), несмотря на колебательный характер исследуемых показателей, существуют отчетливые различия: 1) в кратковременном эффекте последействия патогена (на 2-3-и сутки); 2) в долговременном эффекте (на 20-22-е сутки).
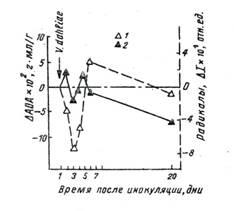
Рис.2. Дифференциальный анализ кинетики свободнорадикальных окислительных и антиокислительных процессов в растениях восприимчивого сорта хлопчатника на ранних этапах взаимодействия с грибом:
1 – концентрация СР(ΔI = Iопыт - Iконтроль); 2 – ΔАОА = АОАопыт - АОАконтроль
Кратковременный эффект последействия патогена в реакции совместимости (система восприимчивое растение - патоген) проявился в торможении ДОП на вторые сутки с синхронным повышением АОА. В случае же реакции несовместимости в системе устойчивое растение - паразит наблюдается усиление ПОЛ и ДОП на вторые сутки после заражения, синхронное со снижением уровня АОА (см.рис.3).
Парадоксальный факт усиления окислительных деструктивных процессов в адаптивных перестройках свидетельствует об ответной реакции на патоген как на хаотропный агент. Слабые возмущения в стационарном уровне СР и АОА мембранных структур в конечном итоге активизируют клетку для адаптивных, восстановительных процессов. Сверхпороговое же возмущение губительно действует и на клетку, и на организм в целом (24, 50).
Повышение интенсивности окислительных процессов с синхронным снижением АОА в клетках растений при контакте с патогеном правомерно отнести к качественному признаку адаптации растения к данной инфекционной нагрузке, в результате которой растительный организм обретает более высокий антиоксидантный потенциал в мембране, предохраняющий клетку от разрушения. Усиление мембранной пероксидации липидов с последующей мобилизацией синтеза АО может служить “триггером” адаптивных реакций растительного организма (35).
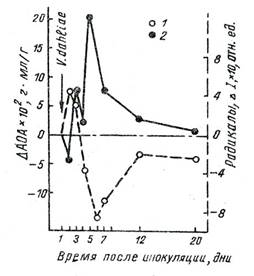
Рис.3. Дифференциальный анализ кинетики реципрокных процессов при заражении толерантного сорта хлопчатника [35]: 1 - концентрация СР – ΔI; 2 - ΔАОА
Следовательно, детерминантная фаза адаптивных процессов, т.е. фаза распознавания растением патогена (28, 82), выражается в усилении интенсивности ДОП и СР-процессов на ранних этапах, что согласуется с повышением интенсивности дыхания и ДОП в устойчивых сортах растений при инфицировании патогеном (51, 69, 74).
На ранних этапах реакции совместимости в системе восприимчивое растение - патоген наблюдается снижение интенсивности ДОП на второй день после заражения грибом, синхронное с локальным увеличением АОА и влекущее за собой ослабление биоантиоксидантного пула, итогом которого являются повышение окисляемости, разрушение мембран и гибель растительного организма. Тогда детерминантная фаза патологических процессов выражается снижением стационарного уровня ПОЛ и ДОП мембранных структур растений на ранних этапах взаимодействия с патогеном (см.рис.2). Долговременный контакт с паразитом, ведущий к патогенезу растения, вызывает снижение уровней СР м АОА ниже контрольных значений (см.рис.2). Снижение биоантиоксидантого пула может потенцировать локальное усиление ПОЛ, угрожающее целостности клетки. Патогенез, таким образом, характеризуется необратимой убылью антиоксидантов, обусловливающей неспособность мембранных структур компенсировать деструктивные сдвиги. Долговременным эффектом адаптивных перестроек является накопление биоантиоксидантов, компенсирующих возросший уровень СР и пероксидацию липидов (см.рис.3).
Установлено, что в норме устойчивые сорта по сравнению с восприимчивыми характеризуются сниженным уровнем потребления кислорода и вместе с тем повышенной фосфорилирующей активностью (16, 51, 77). Эти факты свидетельствуют o более высокой эффективности дыхания и прочности сопрягающего фактора в устойчивом организме, позволяющих растению не только устоять, но и компенсировать поражение. Изучение динамики окислительных и антиокислительных процессов в онтогенезе растений хлопчатника различной вилтоустойчивости показало, что для толерантных организмов характерен больший диапазон колебаний в уровнях СР и АОА (35). Несмотря на снижение потребления кислорода, выносливые к инфекции растения в процессе жизнедеятельности генерируют более высокий уровень СР, контролируемый более активной антиоксидантной системой. Следовательно, в толерантных организмах интенсифицируются как окислительные, так и антиокислительные процессы. Эти результаты согласуются с тем фактором, что способность генерировать свободные радикалы у липидов с повышенным уровнем АОА увеличена, т.е. у них существует потенциальная возможность более интенсивного окисления по сравнению с липидами с заниженной АОА (4, 13). В пользу наличия более мощной АО-системы в толерантных растениях свидетельствует также присущий им более высокий уровень оксидазной и пероксидазной активности (1, 43).
При исследовании окислительных и антиокислительных процессов в динамике взаимодействия растения с грибом установлено антибатное соотношение между кинетиками изменений ДОП и АОА (рис.2-3). Обнаруженная антибатность и повышенная интенсивность этих процессов в толерантных растениях могут проявляться одновременно только при единственном условии: различной локализации систем, отвечающих за эти функции, т.е. структурной компартментализации окислительных и АО-центров в мембранных и субклеточных фрагментах. Носителями этих двух противоположных свойств являются липиды мембран, амфипатические по своей природе, так как, с одной стороны, ввиду двойных связей в гидрофобных участках они могут легко окисляться, а с другой стороны, благодаря АО-свойствам полярных групп липидов и липорастворимых антиоксидантов могут тормозить окисление (3, 9, 22). Активные центры окислительных и антиокислительных процессов в силу их противоположной функции локализованы структурно в различных компартментах мембраны и клетки.
Компартментализация этих процессов и обнаруженная реципрокность могут свидетельствовать o наличии регуляторной связи и динамического равновесия между компартментами с противоположно ориентированными функциями в мембране и клетке. Воздействие хаотропного фактора приводит к нарушению естественного баланса этих двух функций, в первую очередь, на мембранном уровне. Действие экзогенных антиоксидантов направленно усиливает антиокислительный потенциал как для животных (7, 9), так и для растительных организмов (43, 76). По-видимому, двухкомпартментальная модель мембранных процессов с разделением реципрокных функций – окислительных и антиокислительных – может служить также базой для реализации механизмов клеточной ритмики многих биохимических параметров (53).
Таким образом, информационная роль колебаний уровня ДОП и АОА тесно сопряжена с чувствительным ответом, т.е. реактивностью мембран, регулирующей исход взаимоотношений партнеров в динамической системе растение-хозяин-паразит.
2. Мембранные механизмы реактивности.
Качественно различные реакции совместимых и несовместимых растений на воздействие одного и того же патогена могут быть обусловлены в первую очередь скоростью ответной реакции, т.е. реактивностью мембранных структур растительной клетки. Реактивность как показатель функционального состояния мембран отражается в кинетике ДОП и АОА (35, 42, 44).
Состояние мембранных липидов как переносчиков информации, их реактивность могут отразиться и на чувствительности клеток к гормональной регуляции, и на способности индуцирования иммунного ответа (8, 40).
Метаболиты патогена могут вступать во взаимодействие с компонентами клеточной стенки и плазмалеммы растений так, чтобы повышался стационарный уровень ПОЛ, что вызывает локальный “хаос”, микродеструкцию мембраны с увеличением доступности кислорода в клетку. Установлено (24), что действие разнообразных по природе физических и химических прооксидантных факторов, вызывающих разрыхление мембраны, - хаотропных агентов – обусловливает резкое возрастание скоростей генерации СР и липоперекисей. Инициирование ДОП и ПОЛ “хаотропным” воздействием патогена сопровождается кратковременным синхронным снижением АОА в мембранных компартментах (рис.3). По-видимому, ориентация мембранных структур растений на “хаотропность” чужеродного агента и высокая чувствительность ДОП к факторам внешней среды могут лежать в основе механизма реактивности мембран.
Принцип распознавания мембранами “свой - не свой” в случае вирусных, бактериальных и грибковых заболеваний растений также может сводиться к способности растительных мембран различать патоген как “хаотропный” агент.
Физико-химические механизмы распознавания чужеродной информации могут заключаться в активном взаимодействии рецепторных локусов плазмалеммы растений с метаболитами патогена как хаотропными агентами. В детерминантной фазе реакции совместимости, возможно, имеет место такой способ связывания метаболитов патогена с рецепторами мембраны, при котором ингибируется естественный уровень ДОП, снижая тем самым реактивность мембраны.
Таким образом, специфичность взаимоотношений низших и высших растений контролируется двойственным типом взаимодействия мембраноактивных метаболитов патогенов с рецепторами плазмалеммы растительной клетки.
В механизмах реализации реактивности имеют значение как величина стационарного уровня окисления липидов, так и активность пусковых систем, которые интенсифицируют ПОЛ и ДОП. При этом чем выше стационарный уровень ПОЛ и концентрации СР, тем легче подвергаются окислительным сдвигам липидные компоненты мембран, тем выше их реактивность (43).
В литературе распространена точка зрения, что в качестве инициирующих факторов ПОЛ в различных мембранных системах могут выступать активные формы кислорода (протонированный супероксидный радикал (НỎ2), гидроксильный радикал ỎН, синглетный кислород 1O2 (2, 45). Резервы повышения чувствительности мембранных структур к повреждающему хаотропному воздействию заключаются, по-видимому, не только в стационарной активности ферментативного и неферментативного ПОЛ, но и в интенсивной генерации активных форм кислорода. Синглетный кислород взаимодействует с полиненасыщенными жирными кислотами с константой скорости ~104-105М–1с-1. Супероксидный радикал наиболее активен в протонированной форме НỎ2; он может являться предшественником более активного гидроксильного радикала ỎН, возникающего, в частности, в катализированных реакциях типа Хабера-Вайса (Ō2 + Н2О2 → О2 + ỎН + ŌН) (52). Ферментативные системы генерации активных форм кислорода НỎ2, 1O2, ỎН способны многократно усилить слабое повреждение мембранной структуры, отражающееся в чувствительном изменении уровня ДОП. Повышение активности радикалообразования, иницирующего кооперативные деструктивные процессы в мембранных компартментах, увеличивает глубину и ареал поражения даже при сниженном потреблении кислорода в более устойчивых организмах (49). Образование тилл, пробок, защитных барьеров в виде раневой перидермы при физических воздействиях и механическом повреждении и некрозов при воздействии инфекции (71) свидетельствует o единстве механизма индукции реакции сверхчувствительности (РСЧ) при специфическом и неспецифическом воздействиях (48, 49, 80), заключающегося в триггерном усилении ДОП в мембранных фрагментах. Обработка ингибиторами дыхания (78, 84) и SH-блокирующими реагентами (28), снижающими АОА мембран, ослабляет, а внесение сульфгидрильных антиоксидантов индуцирует РСЧ (72). Эти факты подтверждают, что РСЧ индуцируется нарушением баланса окислительных и антиокислительных процессов в мембранных компартаментах.
Замечено, что образование фунгитоксичных соединений в растительных клетках – фитоалексинов (ФА) – более всего зависит от площади поражения и количества некротизированных клеток (28). При исследовании зависимости ФА-образования от скорости СЧ-ответа клеток клубней картофеля обнаружено, что синтез ришитина индуцировался через определенный лаг-период после СЧ-гибели клеток (83, 87). По-видимому, индукторами синтеза ФА могут быть продукты деструктивного распада мембраны самой растительной клетки, интенсивность которого обусловливается реактивностью мембранных структур.
Повышение реактивности обнаруживается при обработке специфическими и ннеспецифическими агентами прооксидантной природы. К таким неспецифическим «индукторам» реактивности можно отнести хаотропные агенты физической природы, вызывающие разрыхление мембранной структуры (УЗ-, УФ-излучения, радиация, повышенная температура), химические соединения (ионы тяжелых металлов, антибиотики, пестициды и др.) (13, 21, 67, 70). К специфическим «индукторам» реактивности, так называемым «сенсибилизаторам», можно отнести поверхностно-активные метаболиты патогена, обработка которыми ориентирует реактивность мембран хозяина на узнавание их рецепторными участками с чувствительным повышением уровня ПОЛ (35, 68, 81). Вещества, подавляющие РСЧ и образование ФА в ответ на последующее инфицирование несовместимой расой патогена, получили название супрессоров (31, 37, 63, 64). Можно предположить, что функциональная роль супрессоров заключается в подавлении реактивности путем торможения стационарного уровня ПОЛ и ДОП. Возможно, что биосинтез подобных супрессоров обусловливает появление более вирулентных рас патогена и направляет эволюцию паразитирующих форм (11, 12).
Вполне допустимо, что воздействие малыми дозами вирулентных штаммов патогена (1, 73), либо инактивированными теплом клетками патогена (75), как и обработка специфическими поверхностно активными полиглюканами (68), либо БЛПК (1) и метаболитами патогена – «сенсибилизаторами» (28, 55, 67), приводит к повышению реактивности. Химическая иммунизация растений и обработка пестицидами, гербицидами и фунгицидами – мембраноспецифичными прооксидантными агентами – повышают реактивность, так как провоцируют усиление ДОП и ПОЛ в хлоропластах и листьях растений (53, 62, 67). Следует отметить, что с увеличением времени от момента иммунизации до повторной реинокуляции агрессивной расы патогена эффект последействия значительно увеличивается (1), по-видимому, вследствие более полной реализации компенсаторных функций и повышения антиоксидантного потенциала растительных мембран.
Парадоксальный факт существования в клетке ферментных систем, усиливающих деструктивные процессы, т.е. «несущих смерть», обретает смысл в повышении реактивного функционального состояния. Системы генерации высокореактивных форм кислорода и радикалов (2, 52), повышающих реактивность мембранных структур, так же необходимы клетке для реализации надежности функционирования защитных систем, целостности и индивидуальности клетки, как и системы детоксикации активных форм кислорода (супероксиддисмутазы, каталазы, пероксидазы), входящие в состав ферментативной антиоксидантной защиты клетки и повышающие ее устойчивость. Способность к усилению избирательного распознавания при повторном воздействии сигнала, сопряженная с чувствительностью систем генерации активных форм О2, и радикалов, может содействовать формированию иммунной памяти и развитию мембранной и иммунологической реактивности, тогда как тормозная система детоксикации радикальных форм служит ослаблению распознавания сигнала мембранной чувствительности и способствует выносливости, толерантности и соответственно устойчивости мембранных и клеточных структур. Возможно, что эволюция устойчивости предполагает развитие структурного разделения компартментов с окислительными центрами и пулом биоантиоксидантов, что обусловливает и увеличение временной дистанции в реализации этих противоположных функций в мембране и клетке. Однако целесообразность структурной организации устойчивости определяется в первую очередь функциональным критерием – реактивностью, поэтому взаимосвязь данных критериев и их соотношение определяют надежность иммунитета высших растений (36).
3. Устойчивость и реактивность – структурный и функциональный критерии биологической надежности растения.
В литературе по физиологии иммунитета существует неопределенность в понятии и терминологии устойчивости. Конституционная, или структурная, а по определению Реймерса (38), экологическая устойчивость – это способность экосистемы сохранять свою структуру и функциональные особенности при воздействии внешних факторов. Процессы распада, деструкции мембран и клеток замедлены у более устойчивых организмов. С другой стороны, активация синтетических процессов и таких катаболических ферментов, как пероксидаза, полифенолоксидаза, в сверхчувствительном (СЧ) ответе при воздействии патогена у так называемых «устойчивых» растений противоречит принципу устойчивости (21,39,71). Этот парадокс разрешим только при строгой интерпретации кинетики последействия внешнего воздействия на растения и четком разделении явлений устойчивости и реактивности. Реактивность мембранных структур, отвечающая за скорость ответной реакции, несомненно, является функциональным свойством иммунных растений, тогда как прочность, стабильность, устойчивость – структурным качеством мембран. Снижение интенсивности реактивного ответа у восприимчивых к патогену растений означает фактическое усиление прочности, жесткости, стабилизации мембраны, т.е. ее устойчивости. Очевидно, следует уточнить терминологию и ввести категорию биологической надежности, характеризующую ответные способности растения в динамической связи с воздействующей средой. Иммунитет – это не только свойства защиты жизнеспособности организма, внутреннего постоянства его состава от внешних воздействий и влияния (внедрения) генетически чужеродной информации, но и способность к адаптивным ответным реакциям в целях сохранения и развития целостности и индивидуальности клетки и организма в целом. Целостность клетки обеспечивается надежностью защитных реакций мембранных структур и поддерживается надежностью функционирования генома растений путем развития структурной устойчивости и функциональной реактивности (36). Однако если жизнеспособность означает индивидуальное выживание растений, то надежность защитных реакций обусловливает к тому же способность к воспроизведению полноценного потомства для обеспечения жизнеспособности вида в естественном отборе (17).
Кинетический анализ последействия гриба VerticilliumDahliaeKl. на различные сорта хлопчатника показал возможность прогнозирования надежности иммунитета по скорости ответной реакции, соответствующей критерию реактивности, и степени ее замедления, соответствующей критерию устойчивости (34, 36, 44).
Строгое разграничение и определение реактивности и устойчивости в кинетике ответной реакции позволят четко охарактеризовать биологическую надежность вегетирующих растительных систем, совпадающую по существу с полевой, или горизонтальной, устойчивостью. Правомерно допустить, что «полевой», или горизонтальной, устойчивостью обладают надежные сорта растений с оптимальной устойчивостью и реактивностью ответа мембранных структур на воздействие патогена.
Таким образом, физико-химические механизмы биологической надежности высших растений заключаются в развитии двух противоположных критериев: с одной стороны, структурной устойчивости молекулярной организации клеточных мембран, с другой – высокой реактивности и чувствительности мембранных структур к фитопатогенам как хаотропным агентам. Однако эволюция высших растений к более надежным формам немыслима вне сопряженной эволюции низших растений и микроорганизмов, приводящей к появлению более вирулентных рас патогена (11).
4. Надежность и изменчивость – факторы сопряженной эволюции партнеров динамической системы растение-хозяин – паразит.
Одним из центральных вопросов фитоиммунитета является проблема преодоления паразитом устойчивости растений, касающаяся механизмов совместимости и специфичности, видовой и сортовой. В результате повышения уровня деструктивных процессов и потребления О2 клетками растений в граничных с патогеном зонах малейшие отклонения в кислородной концентрации чувствительно отражаются на жизнедеятельности патогена. Так, если при росте в стационарной культуре у мутантных форм VerticilliumDahliaeKl. секреция пектолитических ферментов (ПФ) нарушается, то при культивировании на качалке изменяются состав клеточной стенки и характер роста гриба и восстанавливается продуцирование ПФ (28). Данные этих исследований показывают, что гибкость метаболизма и мутации, приводящие к нарушению механизмов биогенеза клеточной стенки и обусловливающие изменения в продуцировании ряда экстрацеллюлярных ферментов, могут быть связаны с ингибированием потребления О2, а вследствие этого и стационарного уровня ПОЛ и ДОП, восстанавливающихся при интенсивной аэрации. По-видимому, высокая чувствительность микроорганизмов высших растений к условия среды, в частности, к уровню О2, обусловливает высокую изменчивость, а способность к продуцированию множественных форм суперссорных агентов – совместимость и возможность паразитизма (59, 65).
Множественность морфологических форм зооспорангиев паразитических грибов также обусловлена высокой изменчивостью их метаболизма (47). Поэтому изменения уровня потребления О2 в ДОП могут провоцировать соответствующую мутацию, которая позволяет патогену модифицировать распознаваемую часть молекулы индуктора так, что она теряет способность активизировать ПОЛ и ДОП мембран хозяина и приобретает такую комплементарность к рецептору, при которой происходит снижение стационарного уровня ДОП, и таким образом становится «неузнаваемой». Такие мутанты паразитов в результате высокой изменчивости и лабильности приобретают ген вирулентности (V1), способный преодолевать ген реактивности R2 хозяина.
Следовательно, можно предположить, что совместимость хозяина и патогена возникает с приобретением способности патогена вырабатывать не только общие антигены (79), но и видо- и сортоспецифичные супрессоры реактивности, снижающие при контакте стационарный уровень ПОЛ и ДОП мембран хозяина и подавляющие при заражении реактивный ответ растения-хозяина. Тогда несовместимость партнеров может иметь место по двум причинам: либо паразит не способен продуцировать супрессоры, комплементарные рецепторам хозяина, подавляющие его реактивность, либо у хозяина могут отсутствовать рецепторные участки, комплементарные метаболитам патогена и способные устанавливать такое «корректное» состояние (31, 60), при котором хозяин отвечает тушением стационарного уровня СР на каждом этапе установления совместимости, в то время как для несовместимости достаточен реактивный ответ хотя бы на одном этапе. Можно допустить, что в ходе сопряженной эволюции системы хозяин – паразит, характеризующейся взаимосвязями «ген-на-ген» (66), у растений усиливаются гены сортовой реактивности, продуктом которых являются рецепторные участки, «узнающие» супрессор, т.е. связывающие его таким образом, чтобы вызвать локальное повышение стационарного уровня ПОЛ и СР. Тогда бывшие супрессоры становятся индукторами реактивности хозяина, а растение приобретает чувствительность и распознает патоген.
Некоторые авторы считают, что микроэволюция лежит вне поля зрения генетики. Первичными механизмами молекулярной микроэволюции биосистем являются, по-видимому, процессы возникновения комплементарных структур на внешних мембранных поверхностях клеток партнеров динамических систем растение-хозяин - паразит, микроб-жертва и пр., способных служить первым звеном в цепи реакции совместимости и прогрессивного адаптациогенеза (35).
Оптимальная реактивность обеспечивает партнерам динамической системы растение-хозяин - патоген не только целостность и индивидуальность, но и возможность прогрессивного адаптациогенеза и сопряженной эволюции с партнером. Взаимоадаптация значительно повышает надежность системы высших и низших растений – этим объясняется долговечность очень древнего симбиоза водоросли и гриба – лишайников (27). При снижении реактивности хозяина мутуалистический симбиоз партнеров системы переходит в паразитизм с преждевременным увяданием и гибелью растения-хозяина. Однако даже при подходящих климатических и почвенных условиях патоген будет заселять только физиологически мощные растения, способные удовлетворить потребности паразита в его питании для осуществления полного репродуктивого цикла (49); взамен потери иммунности (невосприимчивости) паразит обеспечивает хозяину возможность прогрессивного адаптациогенеза. Меньшие дозы либо отсутствие фактора адаптациогенеза могут ослабить реактивность хозяина и усилить жесткость, т.е. устойчивость его мембран.
Избыточная жесткость, стабильность мембранной организации будут препятствовать гибкости метаболизма, поливариантности компенсаторных решений в критических экстремальных условиях, снижать жизнеспособность и биологическую надежность высших растений. Максимальная доза внешнего фактора определяет порог устойчивости и толерантности мембранных структур, превышение которого гибельно для мембраны и целостности клетки. Эти выводы совпадают с законом толерантности Шелфорда, согласно которому лимитирующими факторами процветания организма, вида являются как максимум, так и минимум экологического фактора, причем оба определяют скорее диапазон биологической надежности, нежели «величину толерантности» организма к данному фактору (38).
Более вирулентные расы патогена препятствуют адаптациогенезу хозяина, провоцируемые ими патологические процессы снижают жизнеспособность растений в ущерб самому паразиту.
Стабилизирующий отбор, способствующей большей выживаемости менее вирулентных рас по сравнению с более вирулентными (12), свидетельствует o наличии диапазона оптимальной изменчивости, превышение которого так же ослабляет жизнеспособность паразитирующих форм, как и низкая изменчивость. Поэтому оптимальное соотношение критериев реактивности и лабильности (неустойчивости) может определить диапазон оптимальной изменчивости, обеспечивающей жизнеспособность и адаптивную эволюцию низшим растениям и микроорганизмам. Следовательно, реактивность и чувствительность мембранных структур как хозяина, так и паразита является движущим фактором жизнеспособности, надежности и сопряженной эволюции партнеров динамической системы растения-хозяина и патогена.
Взаимосвязь категорий биологической надежности надёжности высших растений и изменчивости низших организмов с критериями реактивности и устойчивости показана на рис.4. Оптимальное соотношение критериев устойчивости и реактивности определяет диапазон высокой надежности биологических свойств системы высших растений.
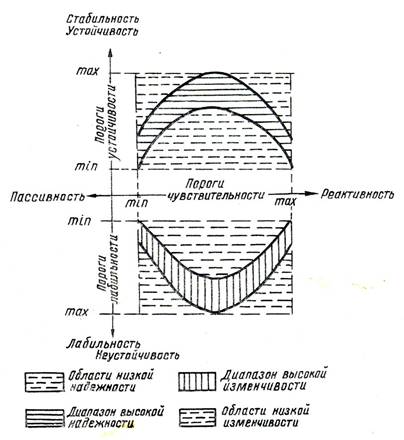
Рис.4. Схема взаимосвязи категорий биологической надёжности с критериями реактивности и устойчивости.
Естественный отбор закрепляет у растений механизмы надежной защиты, реактивности и устойчивости к паразитам, а у последних – механизмы изменчивости, позволяющие преодолевать эту защиту (11, 31). Таким образом, движущим фактором эволюции низших растений и микроорганизмов может быть высокая изменчивость и лабильность мембранных структур и генов вирулентности, способных продуцировать все новые молекулы супрессоров или изменять до неузнаваемости старые, способные снижать при контакте стационарный уровень ПОЛ и реактивности мембран хозяина.
Эволюция высших растений заключается в надежности реализации механизмов устойчивости и реактивности мембранных структур, мобилизующих реактивный ответ.
Для обеспечения сопряженной эволюции симбионтов динамической системы растение-хозяин - паразит необходимо ослабить агрессивность патогена, либо повысить надежность растения-хозяина. Для повышения биологической надежности растения возможны два пути: 1) отбор генов и селекция надежных сортов растения на базе диких высокореактивных видов, мало контактировавших с современными агрессивными расами паразитов и сохранившими доминантные гены реактивности («вертикальной устойчивости») – это пути селекции и генной инженерии; 2) последовательная стратегия повышения реактивности восприимчивых сортов растений с хозяйственно ценными признаками путем обработки индукторами и сенсибилизаторами реактивности различной природы. Так, использование мембран-селективных краунэфиров имеет положительный эффект, заключающийся в снижении заболеваемости хлопчатника вилтом (1), и открывает большие перспективы для применения синтетических мембранадресованных сенсибилизаторов в народном хозяйстве. Реабилитация реактивности культурных растений на всех фазах онтогенеза позволит не только ослабить эпифитотийные агрессии, но и повысить жизнеспособность и надежность иммунитета высших растений на обширных площадях, занятых монокультурами.
Заключение
Кинетический анализ и оценка физико-химических свойств мембранного аппарата высших и низших растений различных генотипов имеют безусловную ценность в выяснении пусковых механизмов молекулярной микроэволюции с позиции адаптивных закономерностей сопряженной микроэволюции партнеров динамической системы растение-хозяин - паразит.
Таким образом, физико-химические исследования динамической системы растение-хозяин-паразит позволили выдвинуть постулат o наличии динамического равновесия двух ассиметричных функций в мембране и клетке, предложить двухкомпартментальную модель мембранных процессов и возможные механизмы реактивности мембранных структур. Кинетический анализ реципрокных процессов – окислительных и антиокислительных – на ранних этапах взаимодействия партнеров динамической системы хозяин-паразит дал возможность подойти к выяснению функциональной организации двухкомпартментальных систем, распознающих хаотропный сигнал, определить критерии надежности высших и изменчивости низших растений, направляющих процессы молекулярной микроэволюции комплементарных мембранных структур партнеров. По-видимому, именно оптимизация реактивности мембранных структур партнеров может явиться ускоряющим фактором эволюции облигатного паразитизма к мутуалистическому симбиозу, при котором взаимоадаптивные перестройки развивают и совершенствуют жизнеспособность, целостность и индивидуальность каждого симбионта, обеспечивая надежность сопряженной эволюции партнеров экологических систем.
Список использованной литературы:
Авазходжаев М.Х. Физиологические основы вилтоустойчивости хлопчатника и пути ее индуцирования. Автореф. дис. на соиск. учен. степени докт. биол. наук. Баку, 1984, 38с.
Ажипа А.И. Медико-биологические аспекты применения электронно-парамагнитного резонанса. М.: Наука, 1983, 527 с.
Аристархова С.А., Бурлакова Е.Б., Шелудченко Н.И. Влияние лецитина на перекисное окисление липидов в микросомах печени // Биохимия, 1979. Т.44. Вып.1. С. 125-129.
Архипова Г.В., Бурлакова Е.Б. Нарушение механизма выхода и переноса липидов мембран при лучевых поражениях и действии радиопроекторов. Некоторые теоретические аспекты противолучевой химической защиты. М.: Наука, 1980. С.65-71.
Бабков В.В. Система основных понятий микроэволюции // Московская школа эволюционной генетики. М.: Наука, 1985. С. 158-160.
Бахтиярова Р.Г. К вопросу устойчивости тонковолокнистого хлопчатника к фузариозному увяданию и физиологии больного растения. Автореф. дис. на соиск. учен. степени канд. биол. наук. Душанбе, 1974. 23 с.
Бречко В.В. Показатели перекисного окисления липидов и антиоксидантной системы при экспериментальной гипокинезии у крыс и их коррекция глутатионом / Тезисы II Всес. Конф. «Биоантиоксидант». М., 1986. Т.1 С. 72-73.
Бурлакова Е.Б., Архипова Г.В., Голощапов А.Н., Молочкина Е.М., Хохлов А.П. Мембранные липиды как переносчики информации // Биоантиокислители в регуляции метаболизма в норме и патологии / Труды МОИП. 1982. Т57. С. 74-83.
Бурлакова Е.Б., Алесенко А.В., Молочкина Е.М., Пальмина Н.П., Храпова Н.Г. Био-антиоксиданты в лучевом поражении и злокачественном росте. М.: Наука, 1975. 214 с.
Бурлакова Е.Б., Храпова Н.Г. Перекисное окисление липидов мембран и природные антиоксиданты // Усп. хим. 1985. Т.54. №9. С. 1540-1558.
Вавилов Н.И. Законы естественного иммунитета растений к инфекционным заболеваниям // Изв. АН СССР, Сер.биол. 1961. №1. С.117-156.
Вандерпланк Я. Генетические и молекулярные основы патогенеза у растений. М.: Мир, 1981. 236 с.
Веселовский В.А. О роли биоантиоксидантов в устойчивости растений к неблагоприятным условиям существования // Биоантиокислители в регуляции метаболизма в норме и патологии. М.: Наука, 1982. С. 150-162.
Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в биологических мембранах. М.: Наука, 1972. 252 с.
Воронков Л.А., Гужова Н.В., Перова И.А., Овчинникова Т.А. Структура и функции энергетического аппарата хлрпчатника, пораженного вилтом // Тезисы докл.XII Международного ботанического конгресса. Л., 1975. Т.2. С.497.
Воронков Л.А., Гужова Н.В. Энергообмен хлопчатника при заболевании вилтом // Патологическая физиология и иммунитет растений. М.: Изд-во МГУ, 1976. С.164-172.
Гродзинский Д.М. Надежность растительных клеток, Киев: Наукова думка, 1983. 368 с.
Губанов Г.Я. Вилт хлопчатника. М.: Колос, 1972. 335 с.
Гужова Н.В., Лебедева А.Ф., Бывалова Т.А. Энергообмен у хлпчатника в связи с его устойчивостью к фузариозному вилту // Сельскохозяйственная биология. 1975. Т.10 №4. С.623-625.
Деверолл Б.Дж. Защитные механизмы растений. М.: Колос, 1980. 128 с.
Дьяков Ю.Т. Мембранные аспекты фитопатологии. М.: ВНИИИТЭИСХ, 1976. 32 с.
Журавлев А.И. Развитие идей Б.Н. Тарусова o роли цепных процессов в биологии / Труды МОИП. Т.57. Биоантиокислители в регуляции метаболизма в норме и патологии. М.:Наука, 1982. С.3-36.
Каличава Г.С., Ванин А.Ф. Исследование методом ЭПР здоровых и пораженных ВТМ листьев табака / Сообщ. АН ГССР, 1971. Т.63. №3. С.717-720.
Козлов Ю.П. Структурно-функциональные аспекты перекисного окислителя липидов в биологических мембранах // Липиды, структура, биосинтез, превращения и функции. М.: Наука, 1977, С.80-93.
Круглякова К.Е., Долягин А.Б., Симонян Г.Г., Воловик А.С., Сухова Л.С. Противоопухолевые свойства химических соединений и кинетические особенности изменения концентрации свободных радикалов и некоторых природных фенолов при раке картофеля // Патологические новообразования у растений. Черноголовка, 1982. С 29-37.
Липсиц Д.В. Биохимические основы болезнеустойчивости картофеля. М.: ВНИИИТЭИСХ, 1972. 60 с.
Маргелис Л. Роль симбиоза в эволюции клетки. М.: Мир, 1984, 351 с.
Метлицкий Л.В. и др. Биохимия иммунитета, покоя, старения растений. М.: Наука, 1984. 264 с.
Мочалкин А.И., Озолина И.А. Исследование послесвечения и свободнорадикальных состояний пшеницы при заражении ржавчинным грибом // Сверхслабые свечения в биологии / Труды МОИП, 1972. Т.39. С.96-98.
Мустафа М., Дьяков Ю.Т. Влияние химических веществ на взаимоотношения Phytophthorainfestans с растениями-хозяевами. II. Влияние фунгицидов и антибиотиков // Микология и фитопатология. 1979. Т.13. Вып.1. С.33-36.
Озерецковская О.Л., Васюкова Н.И., Леонтьева Г.В., Метлицкий Л.В. Биохимические механизмы специализации фитопатогенов // Усп. соврем. биол. 1985, т.100 Вып.2(5). С.230-232.
Пирназарова Ф., Сагдуллаев Х.Х., Исмаилов А.И., Тыщенко А.А., Садыков А.С. Исследование свободнорадикальных состояний в листьях хлопчатника при поражении вертициллезным вилтом // Докл. АН УзССР. 1978. №2. С.46-49.
Пирназарова Ф., Сагдуллаев Х.Х., Исмаилов А.И., Абраров А.А. Исследование кинетики свободнорадикальных состояний в различных по вилтоустойчивости растениях хлпчатника // Физиология и биохимия культурных растений. 1978. Т.10. №5. С.541-546.
А.с. 728783 (СССР). Способ определения устойчивости растений к фитопатогенным поражениям / Пирназарова Ф., Сагдуллаев Х.Х., Исмаилов А.И., Гимашева Н.Ф., Садыков А.М. – Опубл. В Б.И. 1980, № 15.
Пирназарова Ф., Сагдуллаев Х.Х., Исмаилов А.И., Салахутдинов Б.А., Садыков А.С. Окислительные и антиокислительные процессы в динамике взаимоотношений растение-хозяин - патоген // Физиология растений. 1983, Т.30. №1. С.108-117
Пирназарова Ф.Н., Сагдуллаев Х.Х., Исмаилов А.И., Садыков А.С. Антиокислительная активность для оценки надежности фитоиммунитета / Тезисы Всес. совещ. “Биоантиоксидант”. Москва-Черноголовка, 1983. С. 177-178.
Платонова Т.А., Акеньшина Г.В., Озерецковская О.Л., Метлицкий Л.В. Торможение внутриклеточных защитных реакций клубня картофеля под действием супрессоров, выделенных из возбудителя фитофтороза // Прикладная биохимия и микробиология. 1985. Т.21. Вып.1. С. 101-106.
Реймерс Н.Ф. Азбука природы. М.: Знание, 1980. 207 с.
Рубин Б.А., Ладыгина М.Е. Энергообмен и иммунитет растений // Итоги науки и техники. Т.2. Физиология растений. М.: ВИНИТИ, 1976. С. 41-84.
Рубин Б.А., Ладыгина М.Е. Физиология и биохимия дыхания растений. М.: Изд-во МГУ, 1974. 512 с.
Сагдуллаев Х.Х., Пирназарова Ф.Н., Исмаилов А.И. Исследование динамики изменений концентрации парамагнитных центров листьев хлопчатника в норме и при вертициллезном увядании / Тезисы Всес. симпозиума “Магнитный резонанс в биологии и медицине”. Москва – Черноголовка, 1977. С. 113-114.
Сагдуллаев Х.Х., Пирназарова Ф.Н., Исмаилов А.И., Садыков А.С. Реактивность мембранных структур как фактор надежности защитных систем клетки // Докл. АН УзССР. 1982. № 4. С. 45-46.
Сагдуллаев Х.Х. Роль свободнорадикальных окислительных процессов в вилтоустойчивости хлопчатника. Автореф. дис. на соиск. ученой степени канд. биол. наук. Ташкент, 1983. 22 с.
Сагдуллаев Х.Х., Рустамова Ф.Н. Водорастворимые антиоксиданты в оценке реактивности и устойчивости сортообразцов хлопчатника / Тезисы II Всес. конф. “Биоантиоксидант”. Москва – Черноголовка, 1986. С. 152.
Свободные радикалы в биологии / Под ред. А.Прайора. Т.1. М.: Мир, 1979. 318 с.
Сейланов А.С., Попов Г.А., Конев В.К. Связь перекисного окисления и клеточного дыхания // Биофизика. 1982. Т.27. Вып.5. С. 906-908.
Сурнакова Н.Е. Определение жизнеспособности зооспорангиев возбудителя рака картофеля по их морфологическим, биохимическим и биофизическим особенностям. Автореф. дис. на соиск. ученой степени канд. биол. наук. М., 1984. 23 с.
Таирбеков М.Г. Структурные и функциональные аспекты устойчивости растительной клетки // Усп. совр. биол. 1973. Т.75. № 3. С. 406-419.
Тарр С. Основы патологии растений. М.: Мир, 1976. 587 с.
Тарусов Б.Н. Устойчивость – надежность биологических систем при экстремальных воздействиях // Надежность клеток и тканей. Киев: Наукова думка, 1980. С. 16-18.
Тухтаева Г.М., Павловская Н.Е., Ходжаева А.С. Дыхание и фотосинтез хлопчатника, инфицированнго грибом VerticilliumdahliaeKleb. // Узб. биол. журн. 1974, №2 с. 16-19.
Фридович И. Радикалы кислорода, пероксид водорода и токсичность кислорода // Свободные радикалы в биологии. М.: Мир, 1979. С. 272-291.
Эйдус Л.Х., Векслер А.М., Ганасси Е.Э., Плотникова Е.Д. Проблемы защиты, сенсибилизации и восстановления // Биофизика сложных систем и радиационных нарушений. М.: Наука, 1977. С. 188-201.
Эквтимишвили Н.И., Зоз Н.Н., Каюшин Л.П. Кинетика изменения содержания парамагнитных центров при бактериальном раке гороха / Материалы симпозиума “Магнитный резонанс в биологии и медицине”. Москва-Черноголовка, 1981. с 223-233.
Anderson – Prouty A.J., Albersheim P. Host – Pathogen interactions. VIII. Isolation of the pathogen – synthesized fraction rich in glucan that elicit defense response in the pathogens host. // Plant Physiology. 1975. Vol.56. № 2. P. 286-291.
Bell A.A., Stipanovic R.D. Biochemistry of disease and pest resistance in cotton // Mycopathologia. 1978. Vol.65. N 1-3. P. 91-106.
Bertalanffy L. von. General system Theory: Foundations, Development, Applications. London: Allen Lane the Penguin Press. 1971. XXII. 311 p.
Brennemann I.A., Black L.L. Respiration and terminal oxidases in tomato leaves by Phitophthora // Physiol. Plant Pathol. 1979. Vol.14. N 3. P.281-290.
Byrde R.J.W., Willets H.J. The brown root fungus: their biology and control // Oxford: Pergamon Press, 1977. 171 p.
Daly I.M. The use of near-isogenic lines in biochemical studies of the resistance of wheat to stem rust // Phytopathology. 1972. Vol.62. P.392-400.
Day P.R. Genetics of Host-Parasite Interactions // San-Francisco, 1974. 238 p.
Dodge A.D., Harris N. The mode of action of paraquat and diquat. // Biochem. J. 1970. Vol. 118. P.43-F.
Doke N., Garas N.A. Partial characterization and aspects of the mode of action a hypersensitivity-inhibiting factor (HIF) isolated from Phytophthora infestans // Physiol. Plant Pathol. 1979. Vol.15. N 2. P.127-140.
Doke N., Tomiyama K. Effect of hyphal wall components from Phytophthora infestans on protoplasta of potato tuber tissues // Physiol. Plant Pathol. 1980. Vol.16. N.2. P.169-176.
Ersek T., Kirally Z., Dobrovolsky A. Lack of correlation between tissue necrosis and phytoalexin accumulation in tuber of potato cultivars // J. Food Safiety. 1977. Vol.1. P.77-85.
Flor H.H. Current status of the gene-for-gene concept // Ann. Rev. Phytopathology. 1971. Vol.9. P.275-296.
Frank I.A., Paxton J.D. An inducers of soybean phytoalexins and its role in the resistance of soybeans to Phytophthora Rot // Phytopathology. 1971. Vol.61. N 8. P.954-958.
Hahn M.G., Darvill A.G., Albersheim P. Host – pathogen interactions. XIX. The endogenous elicitor, a fragment of a plant cell wall polysaccharide that elicit phytoalexin accumulation in soybeans // Plant Physiology. 1981. Vol.68. N 68. P. 1161-1169.
Heath M.C., Heath I.B. Ultra structure of an immune and susceptible reaction of cowpea leaves to rust infection // Physiology. Plant Pathol. 1971. Vol.1. P. 277-287.
Keen N.T. Specific elicitors of plant phytoalexin production determinants of race specificity in pathogens // Science. 1975. Vol.187. P. 74-75.
Kirally Z., Barna B., Erzek T. Hypersensitivity as a consequence, not the cause, of plant resistance to infection // Nature. London. 1972. Vol.239. P.456-458.
Klement Z., Goodman R.N. The hypersensitive reaction to infection by bacterial plant pathogens // Ann. Rev. Phytopathology. 1967. Vol.5. P.17-44.
Kuc J., Richmond S. Aspects of the protection of cucumber against Colletotrichum lagenarium by Colletotrichum Laenarium // Phytopathology. 1977. Vol.67. N 4. P. 533-536.
Lakshmanan M. Respiratory changes in Fusarium infected cotton // Phytopathol. Z. 1959. Vol.36. N 4. P. 406-418.
Loebenstein G., Loverkovich L. Interference with tobacco mosaic virus local lesion formation in tobacco by injecting beat-killed of Pseudomonas syringae // Virology. 1966. Vol. 30. P. 587-591.
Lupu R., Grossmann S., Cohen Y. The involvement of lypoxygenase and antioxidants in patogenesis of powdery mildew on tobacco plants // Physiology. Plant Pathol. 1980. Vol.16. N 2. P. 241-248.
Millerd A., Scott K.J. Respiration of the diseased plants // Ann. Rev. Plant Physiol. 1962. Vol.13. P. 559-574.
Nozue M., Tomiyama K., Doke N. Effect of adenosine-5-triphosphate on hypersensitive death of potato tuber cells infected by Phytophthora infestans // Phytopathology. 1978. Vol.68. N 6. P. 873-876.
Schnathorst W.C., De Vay J.E. Common antigens in Xanthomonas malvasearum and Gossypium hirsutum and their possible relationship to host specificity and diseased resistance // Phytopathology. 1963. Vol.53. P. 1142.
Smedegaared – Peterson V. Increased demand for respiratory energy of barley leaves reacting hypersensitively against Erysiphe graminis, Pyrenospora teres and Pyrenospora graminis // Phytopathol. Z. 1980. Vol.99. P. 54-62..
Stoessl A. Inermin associated with pisatin inoculated with the fungus Nonilia fructicola // Can. J. Biochem. 1972. Vol.50. P. 107-108.
Talboys P.W. Resistance to vascular wilt fungus // Proc. R. Soc. Long. 1972. B.181. P. 107-108.
Thyamos E.C., Smith I.M. The role of phytoаlexins in the resistance of tomato to Verticillium Wilt // Physiol. Plant Pathol. 1974. Vol.4. N 2. P. 249-252.
Tomiyama K., Doke N., Nozue M., Ishiguri Y. Recognition and specificity in plants host-parasite interaction // Bd. J. M. Daly, J. Uritani. Tokyo. Japan scientific press, 1979. P. 69.
Uritani J. Protein changes in diseased plants // Ann. Rev. Phytopathology. 1971. Vol.9. P. 211-234.
Ward H.M. On the relations between host and parasite in the browse and their brown rust Puccinia dispersa (Eriks) // Ann. Bot. 1902. Vol.16. P. 233-315.
West C.A. Fungal elicitors of the phytoalexins response in higher plants// Naturwissenschaften. 1981. Vol.68. P. 447-451.
Zimmermann J., A.W. Research of manganese clusters in plants by electron paramagnetic resonance spectroscopy // Physiol. Veg. 1985. Vol.23. N 4. P. 425-430
Список произведений >>
Список публикаций >>
Оценить статью можно после того, как в обсуждении будет хотя бы одно сообщение.
Тест: А не зомбируют ли меня? Тест: Определение веса ненаучности
Создан синаптический коммутатор с автономной памятью и низким потреблением
Ученые Северо-Западного университета, Бостонского колледжа и Массачусетского технологического института создали новый синаптический транзистор, который имитирует работу синапсов в человеческом мозге.
Тематическая статья: Как образумить Beast?
Рецензия: Статья П.К.Анохина ФИЛОСОФСКИЙ СМЫСЛ ПРОБЛЕМЫ ЕСТЕСТВЕННОГО И ИСКУССТВЕННОГО ИНТЕЛЛЕКТА