Биохимическая (коэволюционная) модель эволюции генетического кода
Относится к разделу Молекулярная биология
Мы предполагаем, что кодируемый пептидный синтез, а, значит, и современный генетический код (ГК) возник на основе РНКовых форм жизни, в которых большинство канонических аминокислот отсутствовали.
В рамках модели постепенного включения аминокислот в код разумно считать, что в код включались в первую очередь включилось небольшое количество аминокислот, которые были доступны ещё в РНКовом мире - в первую очередь те, которые обнаруживаются в опытах по моделированию абиогенного синтеза (об этом будет отдельная статья). Другие аминокислоты могли появиться только после того, как возникли пути их синтеза.
В 1975 г. Вонг предположил, что главным фактором коэволюции ГК и аминокислот являлись их биосинтетические свойства [Wong J.T.F. 1975, 1981]. По его мнению, современные ферментативные пути биосинтеза аминокислот должны в значительной степени быть сходны с путями синтеза, возможно в том числе и абиогенного на стадии формирования кодируемого пептидного синтеза.
Такая сложная система, как ГК не могла возникнуть сразу полностью. Первоначально, вероятно, в кодируемый синтез были вовлечены лишь несколько аминокислот
Формирование этих первых аминокислот не обязательно должно было катализироваться в первых живых системах. Тем более, что первые биосинтетические пути в клетках могли обеспечиваться не белками, а рибозимами (РНКовыми ферментами). Относительно первых аминокислот, включёнными в пептидный синтез мы можем предположить, что уже до возникновения кодируемого синтеза они активно использовались в биохимических процессах, а значит быть доступными в первичном бульоне. Наиболее вероятными кандидатами на эту роль очевидно предположить те, что абиотически удалось синтезировать в опытах по экспериментальному воссозданию первичного бульона. Это прежде всего такие аминокислоты, как глицин, аланин, пролин и аспартат (обоснование этого факта будет приведено более подробно в других статьях).
Дальнейшая «энзиматизация» синтеза аминокислот привела к повышению скорости их образования. Поскольку гипотеза Вонга предполагает, что генерация новых аминокислот и формирования кодонов с новым смыслом происходили согласованно, данная теория получила название коэволюционной.
Важная особенность первоначального ГК – его вырожденность. Если на начальных этапах формирования ГК аминокислоты кодировались лишь небольшой частью кодонов, то актуальной была задача повысить вероятность нахожденя соответсвующих кодирующих триплетов на мРНК. Вероятность найти соответствующий дублет нуклеотидов на РНКовой матрице в несколько раз выше, чем триплет. Кроме того, вырожденность кода обеспечивает повышенную помехоустойчивость ГК. Поэтому не случайно, что глицин, аланин и пролин кодируются вырожденными кодонами.
В результате включения в код новых аминокислот становился менее вырожденным. Часть кодонов сильно вырожденного первоначального кода для первых аминокислот поменяла смысл и стала кодировать новые аминокислоты. Кодирующая машина «обучалась» более специфично узнавать старые и новые аминокислоты. Таким образом, аминокислоты, имеющие общие пути биосинтеза, должны образовывать связные группы кодонов.
Для удобства рассмотрения будем называть упорядоченную пару аминокислот биосинтетически сестринскими, если они имеют общего аминокислотного предшественника, и связанной отношением «предшественник-продукт», если первая аминокислота данной пары является биосинтетическим предшественником для второй. Ясно, что свойство быть сестринскими является симметричным, рефлексивным и транзитивным, и с его помощью множества аминокислот можно разбивать на классы эквивалентности. Отношение «предшественник-продукт» является асимметричным и транзитивным (это - чисто математический термир, см. значение в википедии), и с его помощью можно формировать упорядоченные множества.
Основываясь на современных знаниях о биосинтезе аминокислот [Cohen G.N. 1968; Rodwell V.W. 1969] Вонг предложил следующие пары, связанные отношением «предшественник-продукт»:
Asp-Asn
Glu-Gln
Ser-Trp
Thr-Ile
Aln-His
Asp-Thr
Glu-Pro
Ser-Cys
Thr-Met
Val-Leu
Asp-Lys
Glu-Arg
Phe-Tyr
Лизин может быть синтезирован или из аспарагиновой кислоты через диаминопимелатный путь и из глутаминовой кислоты через альфа-аминодипатный путь. Первый путь встречается как в прокариотах, так и в эукариотах и может рассматриваться как более древний. Биосинтез метионина из аспартата может происходить в современных организмах более эффективно, однако из треонина метионин образуется за меньшее число стадий и поэтому может рассматриваться как более примитивный. Хотя серин и цистеин могут принимать участие в биосинтезе метионина из треонина, они не являются прямыми предшественниками его. Серин не является единственным возможным источником одноуглеродной группы метионина. Цистеин не является единственным возможным источником серы. Альфа-трансаминирование из-за своей относительной неспецифичности не может рассматриваться в анализе направлений биосинтетических путей. Глицин, аспартат и аланин являются взаимопревращаемыми через цикл трикарбоновых кислот; аланин, серин и глицин – через метаболизм пирувата, глицерата и глиоксилата (Greenberg 1969).
Эволюционная карта, изображенная на рис. 1, выделяет четыре основных центра биосинтеза – глициновый, аланиновый, аспарагиновый и сериновый, из которых могли в древних живых системах синтезироваться 11 других аминокислот.
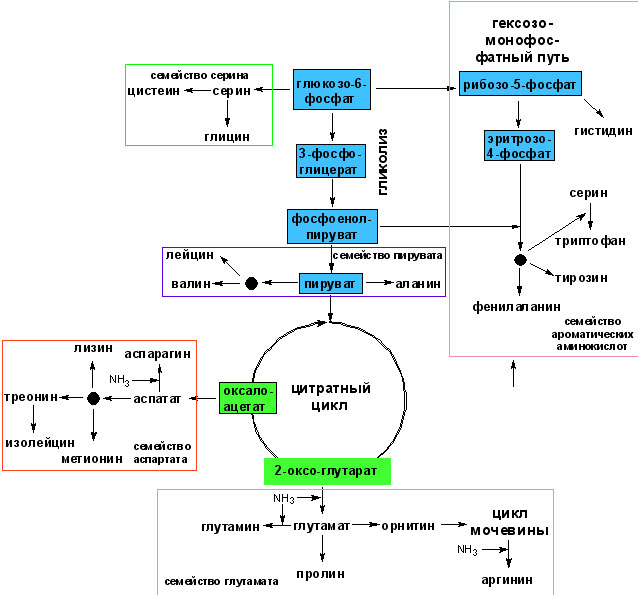
Рис. 1. Упрощенная схема биосинтеза аминокислот [Koolman J. K.-H. Rohm 1998; Кольман Я., К.-Г. Рём 2000]. По особенностям биосинтеза протеиногенные аминокислоты подразделяются на 5 семейств. Члены каждого семейства имеют общего предшественника из цитратного цикла или образуются при катаболизме углеводов.
Вонг выделил также два минорных центра – фенилаланиновый, давший начало тирозину, и валиновый, давший начало лейцину. Первоначально существовало только небольшое множество пребиотически синтезируемых аминокислот таких, как глицин, аланин и серин, кодирование которых обеспечивалось исключительно вырожденным ГК. Существенно, что эти аминокислоты (за исключением фенилаланина) обнаруживаются в опытах по абиогенному синтезу, причем, как правило, в наибольших по сравнению с другими аминокислотами относительных концентрациях (в некоторых опытах правда лейцин иногда может превосходить по концентрации аспартат). Аргументом в пользу существования этих первичных центров метаболизма является взаимопревращаемость всех 5 аминокислот главного метаболического центра и их вовлеченность в углеводный и липидный метаболизм. Существенно также, что аспартат, глутамат и глутамин сыграли важную роль в эволюционном формировании путей биосинтеза пуринов и пиримидинов (ключевых компонентов для синтеза РНК), что подчёркивает их важность уже на начальном этапе формирования ГК. Важность глутамата и аланина как формирующих центральное ядро для биосинтеза других аминокислот подчеркивается их повсеместным участием как доноров аминной группы в реакция трансаминирования, важной по-видимому при синтезе каждой аминокислоты, включая все аминокислоты минорных центров.
Коэволюционная теория также может объяснить помехоустойчивость кода. Поскольку аминокислоты с общими путями биосинтеза, кодируемые связной группой кодонов, должны, в целом, быть близкими по физико-химическим свойствам. Таким образом, согласно коэволюционной теории, помехоустойчивость кода возникла бы даже без специальной селекции на нее, хотя и нельзя исключать дополнительных механизмов помехоустойчивости. Существенно, что связь «предшественник-продукт» не всегда обеспечивает близость по физико-химическим свойствам. Химически существенно отличаются следующие пары «предшественник-продукт»: Ser-Trp, Thr-Ile, Gln-His, Ile-Met, Asn-Lys и Cys-Trp. Кодоновые семейства для этих пар являются смежными, однако естественный отбор не привел к полной оптмизации помехоустойчивости кода. Этот факт может служить одним из косвенных доводов в пользу коэволюционной теории, которая рассматривает это отклонение от оптимальности как результат ведущей роли биосинтетической истории в формировании структуры ГК, а также в пользу того, что после своего формирования код не подвергался в ходе эволюции существенной ревизии.
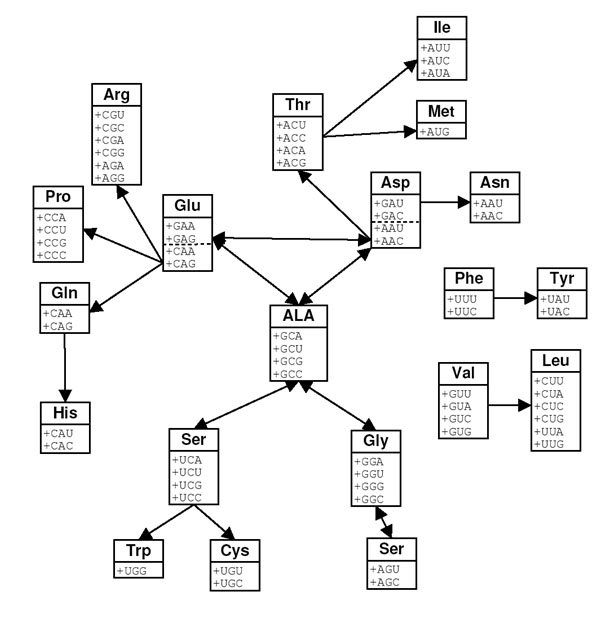
Рис. 2. Последовательность включения отдельных элементовкода согласно коэволюционной теории [Wong J.T.F. 1975]. Кодоны для каждой аминокислотыпредставлены отдельным блоком. Пунктирная линия отделяет кодоны, которыепервоначально кодировали Asp и Glu, но на более позднихэтапах эволюции кода изменили свой смысл (см.текст). Стрелки, направленные в одном направлении, соединяют пары аминокислот в направлении отпредшественника к продукту биосинтеза.Двойные стрелки соединяют аминокислоты, способные взаимопревращиться всовременных организмах.
Вонг показал, что в большинстве случаев пары аминокислот, связанных отношением «предшественник-продукт», образуют смежные серии. Тем не менее, существуют и исключения. Несмежными являются серии для глутамата-пролина, глутамата-аргинина, аспартата-треонина и аспартата-лизина. Эти несоответствия Вонг объяснил следующим дополнительным предположением:
Аминокислоты аспарагин и глутамин были включены в ГК позднее, чем большинство других аминокислот. До включения этих аминокислот их кодоны имели другой смысл. CAA и CAG кодировали глутамат; AAU и AAC были частью аспарагиновых кодонов [Wong J.T. 1975].
Из этого предположения следует, что серии для всех аминокислот связанных биосинтетически первоначально были смежными.
Почему аминокислоты, связанные отношением “предшественник-продукт” должны разделять смежные семейства? Вонг выделил три возможные причины.
Во-первых, продукт может быть по физико-химическим свойствам сходен с предшественником, что оказывается достаточным для того, чтобы он мог связываться с адаптором предшественника.
Во-вторых, химическое превращение предшественника в продукт могло происходить, когда предшественник был уже прикреплен к адаптору.
В-третьих, интермедиат, ведущий от продукта к предшественнику мог связываться с адаптором предшественника, а затем уже превращаться в продукт. В любом из этих случаев новая аминокислота была способна захватить часть кодонов ее предшественника в биосинтезе.
Ди Гиулио конкретизировал коэволюционную гипотезу, предположив, что сами древние РНКовые шпильки – предшественники современных тРНК были активными участниками в биосинтезе аминокислот. Эти шпильки одновременно дали начало как биосинтезу аминокислот, так и генетическому коду.
В качестве одного из аргументов в пользу своей теории Вонг приводит данные о случайной смежности между сериями для аминокислот, связанных биосинтетически [Wong J.T. 1975]. Для любой группы кодонов существует группа из a кодонов, которые будут смежными с данной серией, а также группа из b кодонов, которые будут несмежными с ней. Если все продукты данной аминокислоты будут кодироваться n кодонами, то случайная вероятность того, что x из них будут смежными подчиняется гипергеометрическому распределению:
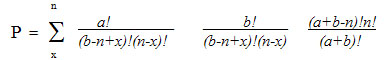
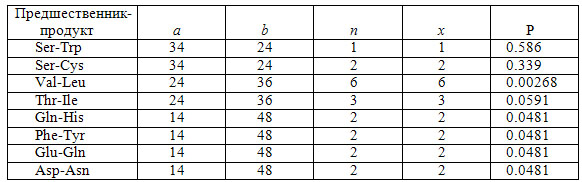
Вонг провел вычисление этих вероятностей для восьми пар «предшественник-продукт», результаты которых приведены ниже в таблице. Используя метод Фишера (Fisher R.A. 1950), просуммировав для восьми пар аминокислот значения –2lnP,получим значение c2=45,01 с 16 степенями свободы. Это значение соответствуетсовокупной вероятности того, что для всех этих восьми пар множеств кодоновбудет выполняться смежность, менее, чем 0,0002. Среди рассматриваемых восьмипар аминокислот пары Phe-Tyr и Val-Leu на стадии формирования ГК биосинтетического пути. Если исключить этидве пары из анализа, то общая вероятность одновременной смежности дляоставшихся шести пар составит 0,0075. Другие возможные биосинтетические связи – Glu-Pro, Glu-Arg, Asp-Thr, Asp-Lys, Thr-Met, Ala-Ser-Gly и Glu-Asp-Ala, также соответствующие смежным сериям – были исключены из анализ ввиду меньшей вероятности смежностей кодоновых серий для пар аминокислот«предшественник-продукт» [Wong J.T. 1975]. В таблице ниже параметры a,b,n и x определены указанным выше уравнением для гипергеометрического распределения.
С биосинтетической точки зрения интересно также сравнить пары аминокислот, кодоны которых отличаются только окончанием. Из семи таких пар по крайней мере три являются биосинтетически сестринскими (Ile-Met, Asn-Lys и Cys-Trp). Пара His-Gln связана отношением «предшественник-продукт»; пара Asp-Glu или является сестринской, или связана отношением «предшественник-продукт». Лишь две пары – Phe-Leu и Ser-Arg – являются биосинтетически неродственными.
Важный статистический тест провел Амирновин (Amirnovin R. 1997), который проанализировал, насколько случайным является сходство кодонов аминокислот с общими путями биосинтеза. Для этого было генерировано большое количество кодов, в которых случайным образом отдельные кодоны были сопоставлены двадцати каноническим аминокислотам, а также случайным образом выбирались три стоп-кодона. Амирновин показал, что значительная часть случайных кодов превосходят канонический код в корреляции степени близости кодонов и биосинтетических связей аминокислот. Таким образом, эти связи в естественном коде не могут рассматриваться как аргумент в пользу биосинтетической теории.
Первоначально предложенная схема биосинтетических связей между аминокислотами [Wong J.T.F. 1975] неоднократно пересматривалась. Одной из причин этого является различной количество отдельных энзиматических превращений, которые требуются в современных организмах для превращения одной аминокислоты в другую. Так, аргинин может быть получен из глутамина с помощью шести энзиматических реакций, тогда как из аспарагина - только с помощью двух. Некоторые пары аминокислот нельзя рассматривать как равноправно взаимопревращаемые, поскольку превращение в одном из направлений может требовать затрат энергии, причем в современных организмах расходуется энергия гидролиза ATP. Раннеберг с соавт. пересмотрели схему биосинтенических взаимоотношений аминокислот с учетом этих требований и показали, что кодовые корреляции, вычисленные на основе биосинтетического родства аминокислот в каноническом коде пренебрежительно малы [Ronneberg T.A. Landweber L.F., Freeland S.J. 2000].
Для проверки коэволюционной теории довольно привлекательной задачей может стать сравнение структур тРНК биосинтетически родственных и неродственных аминокислот. Поскольку различные части тРНК эволюционируют с разной скоростью, важную роль может здесть сыграть анализ отдельных частей тРНК. Так, сравнение двуцепочечных участков антикодоновых петель, показанное в таблице , тРНК E. coli показало определенное сходство для тРНК биосинтетически родственных аминокислот (Wong J.T. 1975). Эти сходства нельзя назвать строго связанными с биосинтезом аминокислот. Так, общая последовательность стебля антикодонового домена (подробней о структуре тРНК вообще и антикодоновом стебле, в частности, будет отдельная статья), у E. coli для тРНК1Ser и тРНКTrp встречается также в тРНКGln. Сходство в первичной последовательности антикодонового стебля тРНК биосинтетически родственных аминокислот может охватывать только часть из изоакцепторных тРНК. Так, тРНКTrp E.coli разделяет общую последовательность с тРНК1Ser, но не с тРНК3Ser, в стебле антикодонового домена которой последовательность будет CUCCC……GGGAG. Такого рода факты говорят о том, что при захвате кодонов новой аминокислотой использоваласть лишь часть из гетерогенного пула изоакцепторных тРНК.
Последовательность стебля антикодоновой петли для тРНК некоторых пар биосинтетически родствевенных аминокислот:

Коэволюционная теория качественно может объяснить особенности вырожденности кода на основе двух факторов. Первый – чем позднее появилась аминокислота, тем меньше у нее оставалось возможности захватить новые кодоны. Поэтому аминокислоты, появившиеся одними из самых последних – метионин и триптофан – «захватили» только по одному кодону. С другой стороны такая древнейшая аминокислота, как аспартат, в современном каноническом коде использует только два кодона. Причина этого объясняется другим фактором – химической реактивностью аминокислоты. Чем больше возможных аминокислотных продуктов может дать какая-то аминокислота, тем больше ее кодонов могут «захватить» новые аминокислоты. Поэтому такие реактивные аминокислоты, как аспарагиновая иглутаминовая кислоты имеют только по два кодона, тогда как такие относительно инертные аминокислоты, как лейцин и аргинин кодируются каждая шестью кодонами.
Литература:
1. Wong J.T. A Coevolution theory of the genetic code. // Proc. Natl. Acad. Sci. USA. 1975. V.72. P.1909-1912.
2. Wong J.T. Coevolution of genetic code and amino acid biosynthesis. // Trends Biochem. Sci. 1981. V.6. P.33-35.
Статьи, связанные с темой:
Древний дублетный генетический код был предопределён путями синтеза аминокислот
Кодазы помогают понять, как возник генетический код
Реликтовые составляющие современного метаболизма. С чего он мог начаться?
Свойства генетического кода - след его возникновения.
Список публикаций >>
Оценить статью можно после того, как в обсуждении будет хотя бы одно сообщение.
Тест: А не зомбируют ли меня? Тест: Определение веса ненаучности
Создан синаптический коммутатор с автономной памятью и низким потреблением
Ученые Северо-Западного университета, Бостонского колледжа и Массачусетского технологического института создали новый синаптический транзистор, который имитирует работу синапсов в человеческом мозге.
Тематическая статья: Как образумить Beast?
Рецензия: Статья П.К.Анохина ФИЛОСОФСКИЙ СМЫСЛ ПРОБЛЕМЫ ЕСТЕСТВЕННОГО И ИСКУССТВЕННОГО ИНТЕЛЛЕКТА