Насекомые имеют зрительную систему, сильно отличающуюся от аналогичной системы позвоночных, это видно даже визуально, стоит лишь сравнить наш камерный и фасеточный глаз. Поэтому часто первый вопрос, который приходит в голову: почему так? Почему у членистоногих эволюция пошла по другому пути, создав фасеточный, а не камерный глаз, и судя по всему, не собирается с него сворачивать? И почему в свою очередь у позвоночных не развился фасеточный глаз, если он так уж хорош? Американский физик-теоретик, Нобелевский лауреат Р. Фейнман на примере пчелы по этому поводу заметил:
Этому мешает несколько довольно интересных причин. Прежде всего, пчела слишком мала: если бы она имела глаз, похожий на наш, но соответственно уменьшенный, то размер зрачка оказался бы порядка 30 мкм, а поэтому дифракция была бы столь велика, что пчела все равно не могла бы видеть лучше. Слишком маленький глаз — это не очень хорошо. Если же такой глаз сделать достаточного размера, то он должен быть не меньше головы самой пчелы. Ценность сложного глаза в том и состоит, что он практически не занимает места — просто тоненький слой на поверхности головы. Так что, прежде чем давать советы пчеле, не забывайте, что у нее есть свои собственные проблемы!
Но я бы обозначил еще одну причину, как мне думается главную: фасеточный глаз эволюционировал как сенсорный орган зрительной системы животных, не имеющих достаточно сложной нервной системы, способной работать с «внутренними образами», «топографическими картами» и прочими абстрактными сущностями, которыми оперирует нервная система позвоночных. Поэтому там не требуется такой детализации, нервная система насекомого просто не сможет их обработать должным образом. По той же причине фасеточный глаз возник эволюционно раньше, но это не значит, что на его базе возник глаз позвоночных. Скорее это была параллельная ветвь, достаточно эффективно выполняющая задачи адаптации нервной деятельности для животных в их экологической нише (насекомых, ракообразных).
Задача глаза, как части нервной системы, способствовать максимально успешной адаптации животного в определенных границах, и фасеточный глаз наилучшим образом подходит членистоногим для этой задачи, иначе он бы не продержался так долго. И хотя насекомые стоят на более низкой ступени эволюции, устройство их зрительной системы, не смотря на кажущуюся простоту, вовсе не примитивно и до сих пор многое в ней не ясно. Но накопленный материал позволяет сделать интересные выводы и по-новому взглянуть на наш камерный глаз, глубже поняв его функциональность.
Строение фасеточного глаза
Фасеточные [axiom_id=636] (фр. facette — «грань»), или сложные глаза - основной, обычно парный орган зрения насекомых, ракообразных и некоторых других членистоногих. Фасеточные глаза состоят из особых элементов омматидиев [axiom_id=637], представляющих собой узкие вытянутые конуса, сходящиеся вершинами в глубине глаза, а основаниями образующих его сетчатую поверхность. Наружная часть фасетки омматидия имеет форму шестигранника, позволяющей поместить максимальное кол-во элементов на единице площади. В нижней части рабдома находится базальная мембрана, имеющая вид тонкой перепонки, выстилающей весь глаз изнутри. Базальная мембрана пронизана множеством отверстий, сквозь которые проходят трахеи и нервные отростки к зрительным долям головного мозга.
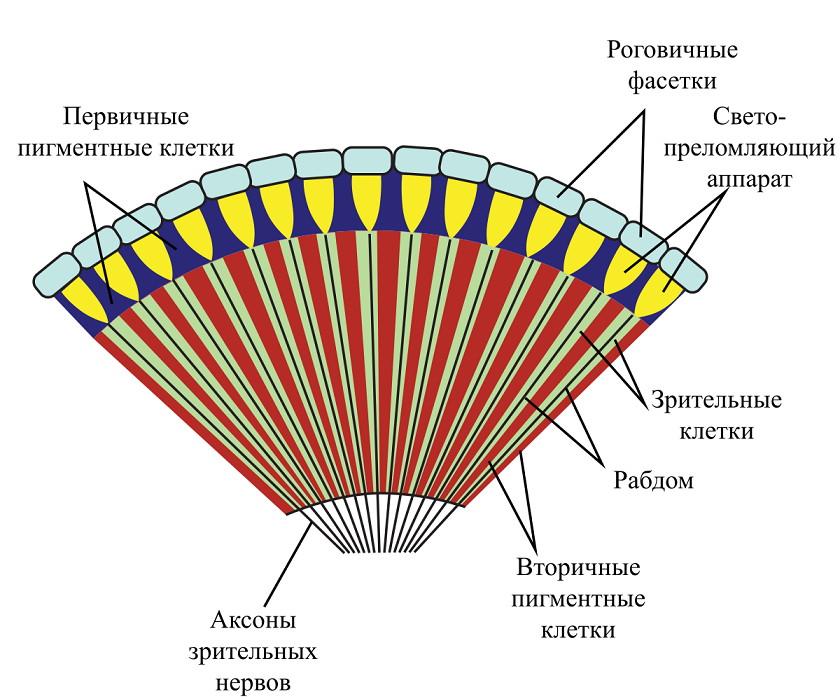
Омматидий имеет очень ограниченный угол зрения и воспринимает только тот крошечный участок находящегося перед ним предмета, на который направлена его оптическая ось. При этом ввиду отсутствия в омматидии зрачка, изображение получается прямым, а не перевернутым, как в камерном глазе. Так как омматидии расположены веерообразно, то фасеточный глаз в итоге получает почти полный круговой обзор, за исключением задней части. Можно представить, что в итоге фасеточный глаз воспринимает изображение в виде мозаики шестигранников, но это не совсем так. Дело в том, что функционально омматидий является аналогом рецепторной клетки (палочки/колбочки), а не фрагмента сетчатки, поэтому «видеть» он может только как отдельный пиксель. Более точной аналогией будет представлять изображение в виде разноцветных пятен, расположенных в углах гексагональной решетки. Но и это не даст верного представления, как воспринимает мир насекомое. Строго говоря, его вообще невозможно получить, так как оно всегда субъективно, и если экспериментально подтверждается, что насекомое реагирует, например, на красный цвет это вовсе не значит, что воспринимает оно его так же как мы. Поэтому, если задаваться вопросом о «видении» фасеточным глазом, то нужно исходить из его анатомических особенностей и способах организации нервных связей для передачи сигнала в зрительные ганглии. И тут оказывается, что у разных видов насекомых существуют три типа фасеточных глаз, которые различают в зависимости от анатомических особенностей омматидиев и их оптических свойств: аппозиционные (фотопические), оптикосуперпозиционные и нейросуперпозиционные (называемые в совокупности скотопическими). У некоторых насекомых (богомолы, подёнки) одна часть глаза может быть построена по аппозиционному типу, а другая — по суперпозиционному. Так проявляется широко распространенная функциональная неоднородность фасеточного глаза, к которой еще вернемся ниже.
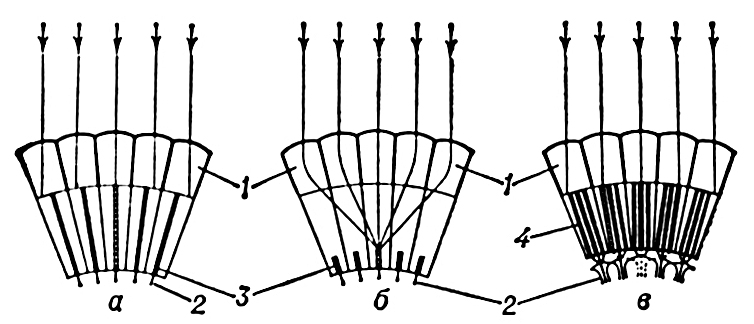
Схема возникновения сетчатого изображения в аппозиционных (а), оптикосуперпозиционных (б) и нейросуперпозиционных (в) фасеточных глазах: 1 — отдельные омматидии с единым или разобщённым светочувствительным элементом, сложенным рабдомерами; 2 — аксоны зрительных клеток. Заштрихованы те светочувствительные элементы, на которые попадают параллельно идущие лучи света (показаны стрелками).
Аппозиционные (фотопические) фасеточные глаза
В аппозиционных фасеточных глазах, свойственных обычно дневным насекомым, смежные омматидии постоянно изолированы друг от друга непрозрачным пигментом и рецепторы воспринимают только свет, направление которого совпадает с осью данного омматидия. При такой компоновке очевидна максимальная чувствительность глаза к деталям изображения, что подразумевает большее приближение к нашему камерному глазу. Но в действительности дневным насекомым такая дефрагментация изображения важна в первую очередь для максимально быстрого определения направления и скорости движения объекта, попавшего в поле зрения, а не детализации. Главное – успеть вовремя заметить опасность или жертву, для чего возникли специфические, связанные между собой особым образом нервные клетки, реагирующие на изменение яркости зрительных клеток омматидиев, что позволяет определять направление и скорость объекта. То есть уже от зрительного органа поступает информация, позволяющая сразу подцепить к нему рефлекторную цепочку. А для уточнения у некоторых, например хищников вроде богомола, часть омматидиев по центру глаза особо мелкого размера и направляет свои сигналы в зрительный ганглий по нейросуперпозиционной схеме для более глубокого анализа – определения начальных примитивов и контуров объекта. Этот участок фасеточного глаза аналогичен фовеоле в сетчатке камерного глаза, месту ее максимальной разрешающей способности. И такое разделение функциональности показывает, что у насекомых зрительная система такая же «многоканальная», как в камерном глазу. У насекомых часто встречается функциональная неоднородность глаза. Например, одна часть приспособлена для грубого определения изменения освещенности, движения и скорости объектов, другая более чувствительна к определенному спектру, третья различает поляризованный свет, небольшой участок служит для детализации. Визуально это можно заметить как разноцветные, замысловатые узоры на глазу с точками-вкраплениями. У летунов обычно верхняя часть специализируется на поляризации света, а нижняя служит для распознавания объектов. Кроме этого, у хищных типа стрекоз дорсальная половина глаза цветослепа, но обладает высокой чувствительностью к коротковолновым лучам: ультрафиолетовым, фиолетовым и синим, что облегчает стрекозе замечать мелкую добычу на фоне неба. Такая выборочная специализация показывает, что фасеточный глаз, так же как и камерный, обрабатывает в параллельном режиме несколько потоков информации, и точно так же как в сетчатке, многие задачи распознавания решаются схемотехнически, через специфические рецепторы и их компоновку, а не «программно».
Оптикосуперпозиционные фасеточные глаза
В оптикосуперпозиционных фасеточных глазах, характерных для ночных и сумеречных насекомых и многих ракообразных, изоляция омматидиев переменная (вследствие способности пигмента перемещаться [axiom_id=639]), и при недостатке света происходит наложение (суперпозиция) падающих под косым углом лучей, прошедших не сквозь одну, а сквозь несколько фасеток. Таким образом, при слабом освещении увеличивается чувствительность глаза. Можно предположить механизм такой регуляции с использованием обратной связи с интенсивностью возбуждения зрительной клетки. Это по сути аналог «зрачкового рефлекса» камерного глаза позвоночных, только более медленный.
У насекомых темновая/световая адаптация в отличии от позвоночных менее универсальна и требует наличия специально адаптированного глаза. Видимо потому, что насекомым проще переключиться на строго ночной/дневной образ жизни, чем пытаться адаптироваться одинаково хорошо в обоих режимах. И у них в меньшей степени встречаются эволюционные изменения образа жизни от ночного к дневному, или например от монохромного к цветовому зрению, скорей всего по тому, что основной принцип выживания насекомых - повышенная плодовитость, а не модификация имеющихся приспособлений.
Нейросуперпозиционные фасеточные глаза
Для нейросуперпозиционных фасеточных глаз характерно суммирование сигналов от зрительных клеток, находящихся в разных омматидиях, но получающих свет из одной и той же точки пространства. При поглощении рабдомерами фотонов света в ретинальных клетках генерируются нервные импульсы, которые по аксонам направляются в парные зрительные доли головного мозга насекомых. В каждой зрительной доле имеется по три ассоциативных центра, где и осуществляется переработка потока зрительной информации, одновременно идущей от множества фасеток.
Вышеприведенное разнообразие схем оптической компоновки возникло в первую очередь как адаптация к слабому освещению, при этом нейросуперпозиционная подразумевает наличие более продвинутого анализатора, так как здесь уже явно происходит первичная обработка сигнала на уровне первого слоя нервных клеток. То есть здесь уже наблюдается попытка не просто передавать изображение в зрительные ганглии как «точка в точку», но определять «объекты» в виде простейших примитивов. Сама темновая адаптация необходима по той причине, что невозможно построить схему с диапазоном чувствительности рецепторов в сотни тысяч или даже миллионы раз. Поэтому все зрительные системы работают по принципу различения контрастов освещения, а не его абсолютной величины, а это обычно всего лишь несколько десятков раз. То есть определяется разница между освещенностью близлежащих рецепторов, что легко делается при помощи латерального торможения. Но для этого нужно изменять либо порог возбуждения рецепторов, либо величину светового потока на единицу площади. В сложных системах, таких как камерный глаз, присутствуют оба варианта, но насекомым из-за отсутствия перенастраиваемой оптической системы приходится использовать более простые механизмы, такие как миграция изолирующего пигмента. При этом единицей (пикселем) изображения служит отдельный омматидий, а не нервная клетка, как в камерном глазе. Поэтому стоит рассмотреть его строение более подробно.
Омматидий
Омматидии [axiom_id=637] сформировались в ходе эволюции из изолированных простых глазков - археомм при их интеграции в сложные, или фасеточные, глаза. Количество омматидиев в таком глазу различно: от горстки у примитивных древнечелюстных и щетинохвосток до 28 тыс. у стрекозы.
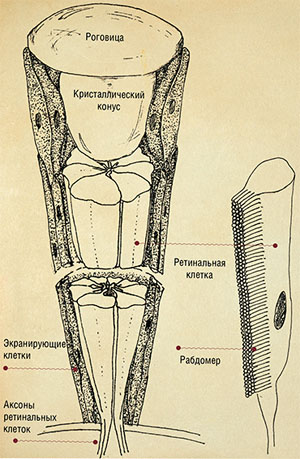 | Сложный фасеточный глаз насекомого состоит из отдельных единиц — фасеток (омматидий). Каждый омматидий является многоклеточным образованием, включающим в себя диоптрические структуры (роговицу и кристаллический конус), фоторецепторы — ретинальные клетки с фоточувствительным пигментом родопсином, а также экранирующие клетки со светопоглощающими пигментами. Родопсин находится в мембране множества микроскопических трубочек-ворсинок, составляющих рабдомер. Рис. Н. Крюковой (ИСиЭЖ СО РАН, Новосибирск) |
В омматидии можно различить три основные части: преломляющую (диоптрическую), воспринимающую (рецепторную) и изолирующую. Изолирующая часть состоит из пигментных клеток, содержащих чёрный пигмент, поглощающий световые лучи. Главную часть изолирующего аппарата составляют клетки, окружающие хрусталик. Благодаря им вокруг каждого омматидия образуется светонепроницаемая оболочка, защищающая рецепторы от попадания боковых лучей света. Но как уже говорилось выше, у некоторых насекомых развилась способность регулировать миграцию изолирующего пигмента как способ темновой адаптации к слабому освещению.
Преломляющая часть
Преломляющая часть омматидия состоит из двух линз с неизменным фокусным расстоянием. Наружная линза (хрусталик) имеет форму двояковыпуклую шестигранника и состоит из хитина. В углах соприкосновения смежных линз располагаются длинные прямые щетинки: они служат для защиты глаза от пыльцы и встречаются у всех насекомых, посещающих цветы. Под хрусталиком расположены две корнеагенные клетки, которые в процессе развития организма выделяют хрусталик, а затем преобразуются в пигментные клетки. Вторая линза — кристаллический или хрустальный конус, располагается под хрусталиком и представляет собой прозрачное тело конусообразной формы с основанием, прилегающим к хрусталику, и вершиной, обращённой вниз. Здесь видим принципиальное отличие от камерного глаза – отсутствие аккомодации, способности оптической системы глаза подстраиваться под наблюдение объектов на разном расстоянии. Такая система просто не нужна в омматидии, так как он воспринимает свой участок изображения в виде точки, и никакой детализации ему не требуется.
Рецепторная часть
Рецепторная часть омматидия, напоминающая вертикально стоящую расческу, состоит из сильно вытянутых зрительных (ретинальных) клеток, соединенных радиально в общий пучок. Чаще всего таких клеток насчитывается восемь. Вдоль оптической оси омматидия расположены рабдомеры зрительных клеток, образующие в совокупности рабдом. Различают замкнутый (спаянный) рабдом - когда отдельные рабдомеры тесно объединены друг с другом, и открытый - если отдельные рабдомеры (кроме двух) разобщены и оптически изолированы друг от друга. Каждый рабдомер сложен большим количеством микроскопических трубочек - микровилл - диаметром около 50 нм, лежащих перпендикулярно оптической оси омматидия. Микровиллы, образованные двойной мембраной толщиной около 5 нм, содержат зрительный пигмент, концентрация которого достигает 107 молекул на рабдомер. Поэтому рабдомер является собственно фоторецепторным элементом зрительной клетки, функциональным аналогом наружного членика палочек и колбочек сетчатки позвоночных.
Микровиллы расположены строго упорядоченно внутри рамбдомера. Например, у медоносной пчелы микровиллы восьми наиболее крупных клеток расположены в двух взаимно перпендикулярных направлениях, при этом ориентация трубочек в рабдомерах соседних омматидиев остается одинаковой на больших участках глаза. У других насекомых картина ориентации микровилл может быть иной, но они всегда по-разному направлены в несимметричных рабдомерах.
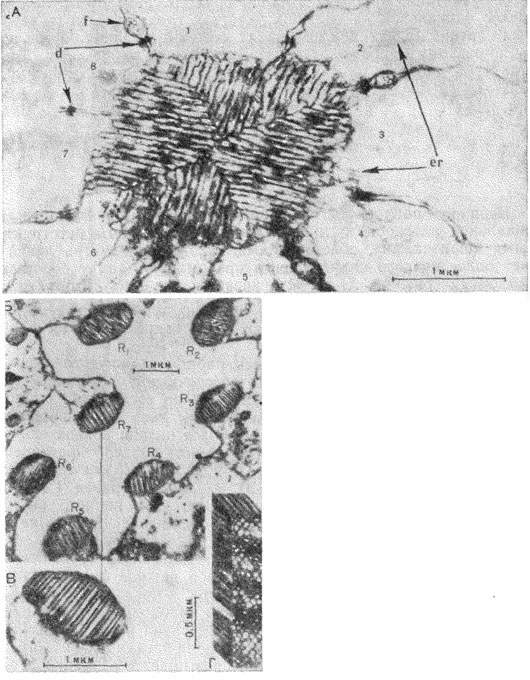 | Электронные микрофотографии поперечного среза рабдома. А - медоносная пчела Apis mellifera (пo Goldsmith, 1962); 1-8 - зрительные клетки; d - скопление десмосом; еr - эндоплазматический ретикулум; f - нити кристаллических клеток; Б, В, Г - муха Drosophila melanogaster (пo Wolken, 1961): R1-R7 - семь раб-домеров; Г-реконструкция участка рабдомера, рассеченного вдоль |
Такая структура рабдома при дихроизме поглощения света молекулами зрительного пигмента обеспечивает анализ поляризации лучей [axiom_id=643]. Это достигается тем, что при разбросе направлений ориентации микровилл в соседних рамбдомерах омматидиев всегда находится пара, направленных примерно в туже сторону, позволяя тем самым получить рецепторную активность в виде линии, направленной параллельно направлению поляризации света.
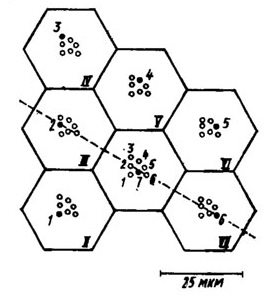 | Группа из семи (I-VII) омматидиев. Черным кружком изображен один из семи (1-7) рабдомеров каждого омматидия, смотрящий в одну и ту же точку пространства. |
Чувствительность к поляризации достигается тем, что в зрительных нейронах фоторецепторная мембрана микроворсинок рабдомера свернута в трубку круглого или гексагонального сечения. За счет этого часть молекул родопсина не участвуют в поглощении света из-за того, что дипольные моменты этих молекул располагаются параллельно ходу светового луча (Говардовский, Грибакин, 1975). В результате микроворсинка приобретает дихроизм — способность к различному поглощению света в зависимости от его поляризации. Повышению поляризационной чувствительности омматидия способствует и то, что молекулы зрительного пигмента не располагаются в мембране хаотично, как у человека, а ориентированы в одном направлении, да к тому же жестко закреплены.
Рассмотрим это на примере модели рамбдома пчелы. Восемь треугольников модели из поляроидной пленки сложены вершинами внутрь так, чтобы каждый сектор поляризовал свет параллельно своему основанию. Если смотреть через эту модель на белое облако - источник деполяризованного света, то все восемь секторов выглядят одинаковыми. При рассматривании же голубого неба на модели виден крестообразный узор из светлых и темных секторов, неодинаковый для разных участков небосвода.
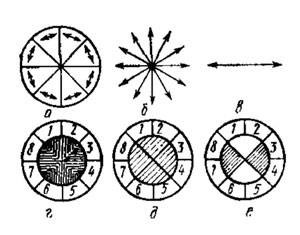 | Упрощенная схема механизма восприятия поляризации света
|
Чувствительность глаза к поляризации обеспечивает членистоногим не только широкие возможности астрономической ориентации, но и помогает опознавать некоторые земные объекты. Блики на разных предметах могут быть различными для глаза насекомых, и эта скрытая особенность бликов может служить для насекомого дополнительным признаком для распознавания объектов и их взаимного расположения. Существенно более развитая нервная система высших позвоночных позволяет составлять сложные "топологические карты местности", что намного эффективнее для ориентации, чем следование "по разметке" поляризованного света. Поэтому позвоночные практически не используют распознавание поляризации солнечного света.
Кроме различения яркости, омматидий может определять спектр излучения. Практически все насекомые ведущие дневной и сумеречный образ жизни прекрасно различают цвета, причем спектр чувствительности сдвинут в сторону коротких волн, благодаря чему им доступно ультрафиолетовое излучение. Это довольно интересное явление потому, что явно наблюдается симбиоз приспособляемости разных видов живых организмов к этой части спектра. Так многие цветы имеют специальную разметку, видимую только в ультрафиолетовом спектре, указывающую насекомым направление к месту расположения нектара. Сами насекомые активно используют аналогичные метки для различения особей разных полов. Кажущиеся нам одинаково белыми крылья бабочки в ультрафиолетовом спектре оказываются у особей одного из полов с хорошо различимыми темными/светлыми участками. Радужные хитиновые покрытия, буйство красок цветковых растений под ультрафиолетом для насекомых выглядят вполне лаконичными маркерами и указателями. Выбор ультрафиолета для этих целей думаю не случаен. Нижняя граница для ультрафиолетового излучения на поверхности Земли - около 300 нм. Однако у поверхности земли воздух достаточно прозрачен для коротковолнового излучения с меньшей длиной волны. Источниками же такого излучения могут быть делящиеся или отмирающие клетки, митогенетические лучи наблюдаются в диапазоне от 190 до 325 нм. Довольно интенсивное ультрафиолетовое излучение возникает при делении клеток дрожжей, субстраты с которыми ищут самки дрозофил для откладки яиц. Не исключено, что дрозофила видит развивающиеся клетки дрожжей сверкающими особым ультрафиолетовым светом. Возможно также, что другие насекомые видят точки роста растений, а также поврежденные листья, как излучающие собственный свет. То есть, возможно, изначально чувствительность к ультрафиолету возникла как адаптация к поиску специфического корма, а в дальнейшем стала использоваться для создания опознавательных меток, в том числе у растений, для привлечения насекомых. Например, ночные белые бабочки отражают от 20-65 % этих лучей, что делает их особенно заметными для других насекомых при свете ночного неба. Можно представить, что для насекомых ночью они выглядят как светлячки, со своими индивидуальными рисунками. Так же замечено, что пчелы легко отличают кажущиеся нам одинаково белыми две поверхности, одна из которых выкрашена цинковыми белилами, а другая - свинцовыми. Свинцовые белила гораздо сильнее отражают ультрафиолетовые лучи; они "действительно белые" для насекомых в отличие от цинковых белил, которые сильно поглощают ультрафиолетовые лучи и поэтому имеют для насекомых цвет, дополнительный к ультрафиолетовому. В частности, для пчел дополнительным к ультрафиолетовому является сине-зеленый цвет.
О способности различать цвета можно говорить только тогда, если имеется разная реакция зрительных рецепторов на излучения различного спектрального состава в не зависимости от их интенсивности. При этом нужно учитывать, что фоновое излучение тоже может иметь какой-то спектр, способный оказывать влияние на цветовосприятие. Поэтому обычно так же подразумевается и способность различать окраску предметов в не зависимости от цвета освещения. Это называется константным восприятием окраски, без которого операция различения отраженного света по спектральному составу была бы недостаточной для надежной ориентации в многообразии окружающих предметов. Можно предположить, что механизм такого различения использует антагонистическое противодействие рецептивных полей с разной цветовой специализацией, по аналогии с "цветовым противодействием" в сетчатке позвоночных, а так же сравнения отражаемого объектом спектра со спектром других предметов в поле зрения. Иначе трудно представить, как еще можно выделить один цвет на фоне других без контрастирования и сопоставления.
Для цветовосприятия нужно минимум 2 типа рецепторов (дихроматы), чувствительные к разным частям спектра, но чаще встречаются животные с тремя типами - трихроматы, хотя некоторые насекомые и членистоногие имеют 5 и более типов цветочувствительных рецепторов. Такими рецепторами являются рамбдомеры в омматидии, которые можно представить как аналог колбочки в сетчатке позвоночных. И так же как у колбочек их диапазон спектральной чувствительности довольно сильно плавает, что подразумевает обязательное наличие механизма контрастирования их активностей. Но так как первичной обработки сигнала, по аналогии с многослойной сетчаткой, внутри омматидия не происходит, поэтому остается только возможность обработки сигнала в зрительном ганглии, а для этого необходимо передавать туда сигнал отдельно от каждой клетки. Это подтверждается тем, что электрофизически зарегистрированы активности волокон, специфически реагирующих на свет разных длин волн, что было бы не возможно, если сигналы от зрительных клеток омматидия просто смешивались. И судя по всему, здесь присутствует "цветовое противодействие" характерное так же и для сетчатки позвоночных и реализованное интересным образом.
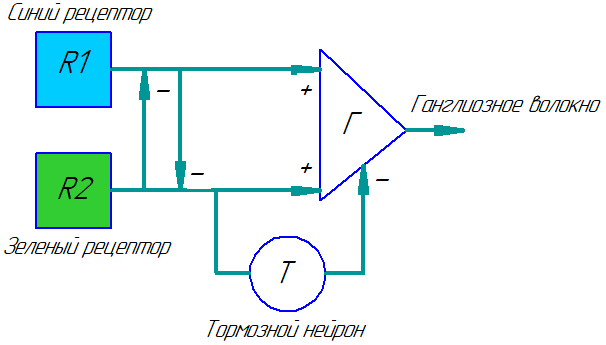
Модель передачи информации о цвете в зрительной системе бабочки Epargireus clarus (пo Swihart, 1969):
R1 - 'синий' рецептор; R - 'зеленый' рецептор; Г - цветное ганглиозиое волокно; Т - вставочный интернейрон с 'задержкой'.
У бабочки Epargyreus clarus замечен особенно интересный тип цветного "оппонентного" волокна. Это волокно интенсивно разряжается импульсами при освещении глаза синим светом, очень коротко отвечает на зеленые лучи, но не возбуждается "белыми" лучами. Здесь синий свет тормозит ответ волокна на зеленые лучи и наоборот, при этом ответ на синий свет возникает без задержки, а на зеленый - с задержкой. В результате на выходе получается либо одиночный, либо двойной импульс, что говорит о необходимости регистрировать разницу между двумя типами сигналов. Можно представить себе разные варианты таких схем, но мне не попадались конкретные описания.
Разрешающая способность фасеточного глаза
Острота зрения, или разрешающая способность, характеризует способность глаза различать степень расчлененности объектов, в том числе отличать две точки от одной более крупной. Но это не равнозначно способности различать минимальный объект [axiom_id=648], так как для этого важна контрастная чувствительность. Чтобы увидеть мелкий объект, например, в виде точки, достаточно одного рецептора, но чтобы отличить два - нужно минимум 3, при этом средний должен быть менее освещен, чем боковые, то есть служить "разделителем". Но так как контрастная чувствительность регулируется латеральным торможением, способность различать малые объекты зависит от количества и силы тормозных связей. Уменьшая величину омматидиев и плотность их упаковки, можно увеличить остроту зрения, но при этом будет возрастать дифракционная погрешность, возникающая на входном зрачке омматидия, которая в конечном итоге сведет на нет преимущества малого размера. В итоге получается зависимость теоретической возможности остроты зрения от размера омматидия, плотности их упаковки, дифракционной погрешности и величины взаимного торможения между ними. Стало быть, бесконечно увеличивать остроту зрения фасеточный глаз не может. Но есть и другое, скажем так, концептуальное ограничение.
Насекомым нет надобности иметь особо острое зрение в первую очередь потому, что это подразумевает более сложную организацию нервной системы, которая во многом зависит от сложности строения организма, а это в свою очередь определяется экологической нишей, занимаемым животным и сформированной стратегией выживания. Установлено, что насекомые успешно различают фигуры по размерам, но довольно грубо. Так пчелы способны различать два различных по площади круга с кратностью в 2 раза, а осы и мухи даже в 1,5. Поэтому понятие "разрешающая способность зрения" для членистоногих следует понимать в первую очередь как «достаточная для адаптации». При такой интерпретации возможен и другой способ различения малых объектов, когда последовательно, в разные моменты времени, сравнивается возбуждение одних и тех же зрительных единиц. Глаз, построенный на таком принципе, смог бы различать на движущейся картине даже больше деталей, чем это обеспечивает угловая плотность зрительных единиц. В зрительном анализаторе человека и, видимо, всех позвоночных животных этот принцип не реализован, но у насекомых, некоторых паукообразных и ракообразных, судя по всему есть зрительный механизм, основанный на трансформации пространственной последовательности стимулов во временную последовательность возбуждения одних и тех же рецепторных единиц. В первую очередь это касается летающих насекомых, которым критически важно не просто различать объекты, но делать это в полете, при быстрых маневрах. Как показали электрофизиологические измерения критической частоты мельканий и прямое наблюдение за поведением насекомых в оптомоторном барабане с периодическим узором полос, для хороших летунов - мух, пчелиных, стрекоз мелькания не сливаются до тех пор, пока их частота станет равной 250-300 в секунду. У человека мелькания начинают сливаться уже при 16 кадрах, именно такая частота использовалась в первых кинопроекторах.
Способность различать фрагменты объектов в движении развилась так же у некоторых паукообразных и ракообразных. Например, в глазу морского рачка Сорiliа позади сравнительно крупной линзы лежит всего четыре зрительные клетки, но они подвижны за счет ритмических сокращений специальных мышц. Такая сетчатка из четырех элементов непрерывно колеблется, сканируя ретинальное изображение с частотой 5-6 раз в секунду. Малое число рецепторов здесь компенсируется их подвижностью, что, очевидно, позволяет рачку различать детали и форму предметов. Сканирующая сетчатка известна также в глазах некоторых пауков и клещей. Так же описана особая мышца в глазах мух, периодически подергивающая проксимальную часть слоя рецепторов.
По поводу детектирования формы объектов [axiom_id=649] экспериментально определено, что пчелиных, бабочек, мух и некоторых других насекомых расчлененные фигуры спонтанно привлекают сильнее фигур с ровным сплошным контуром или однородным (нерасчлененным) внутренним полем. Колеблющиеся на ветру цветки насекомые посещают чаще, чем неподвижные. Многочисленные опыты показали, что насекомые не одинаково хорошо способны обучаться различать фигуры с простым контуром. Например, они достаточно легко отличают 8- и 16-лучевые звезды, а также крест и звезду от круга. Но попытки научить отличать круг от треугольника долгое время не приводили к успеху. Поэтому сначала предполагалось, что насекомые могут различать фигуры только по степени расчлененности их контура или внутреннего поля, т. е. путем сравнения частоты мельканий, возникающих в глазу при движении насекомого относительно объекта или самого объекта относительно глаза. Но дальнейшие опыты все же показали, что если контур фигур многократно повторен, это позволяет насекомому оценивать форму как издалека, так и вблизи, когда в поле зрения помещается только часть ее. Опознать эти фигуры по частоте и последовательности мельканий практически невозможно. Это дает основание полагать, что насекомым свойственны оба рассмотренных выше механизма анализа расчлененности поля зрения: по частоте мельканий и по картине возбуждения разных рецепторов одновременно.
Есть еще одно предположение, почему возникают проблемы с некоторыми фигурами. Дело в том, что фасеты упакованы по гексагональной схеме в виде шестигранников, а это накладывает ограничение на аппроксимацию кривых объекта. То есть фигуры с углами кратными 30, 60 градусам будут определяться максимально точно, а вот круг будет представлен в виде многогранника, причем все более приближающемся к шестиграннику по мере уменьшения размеров. В свою очередь, шестигранник - это 6 плотно упакованных треугольников. Поэтому у насекомого при виде мелкого круга активируются примитивы треугольников, что и вызывает первоначальный сбой в распознавании.Но в дальнейшем при формировании нейронных связей при обучении удается сгруппировать треугольники в самостоятельную макрофигуру.
Насекомые, отличающиеся быстрым полетом или хорошо прыгающие, а также большинство хищников воспринимают также объемность предметов и хорошо оценивают глубину пространства. Стереоскопичность зрения достигается тем, что поля зрения обоих глаз впереди и внизу частично перекрываются: в правом и левом глазах есть омматидии, оптические оси которых сведены или хотя бы параллельны друг другу. Такие (корреспондирующие) пары омматидиев "видят" предмет в одной и той же точке пространства. Для разных точек пространства по мере удаления от глаза корреспондирующими окажутся разные пары омматидиев, что в принципе достаточно для улавливания изменения глубины. Корреспондирующие омматидии, например, у богомолов лежат в специальной зоне глаза причудливой формы, одностороннее ослепление которой ведет к утрате способности насекомого безошибочно схватывать добычу. В норме же богомол, как и охотящаяся личинка стрекозы, никогда не выбрасывает ловчий аппарат, когда расстояние до жертвы больше последнего. Стереоскопичность зрения важна не только охотникам за добычей, но и всем быстро летающим и быстро бегающим формам для обеспечения посадки, своевременного маневра при облете препятствий или преследования полового партнера. В случае посадки сигнал для своевременного вытягивания ног посылают особые детекторы, описанные на примере зрительной системы мух.
Дорсальные глазки
Кроме сложных фасеточных глаз, у насекомых имеются более примитивные дорсальные глазки [axiom_id=641]. Многие взрослые насекомые имеют дорсальные глазки, обычно в количестве трех, расположенных в виде треугольника на передней части головы. Иногда их бывает только один или два.
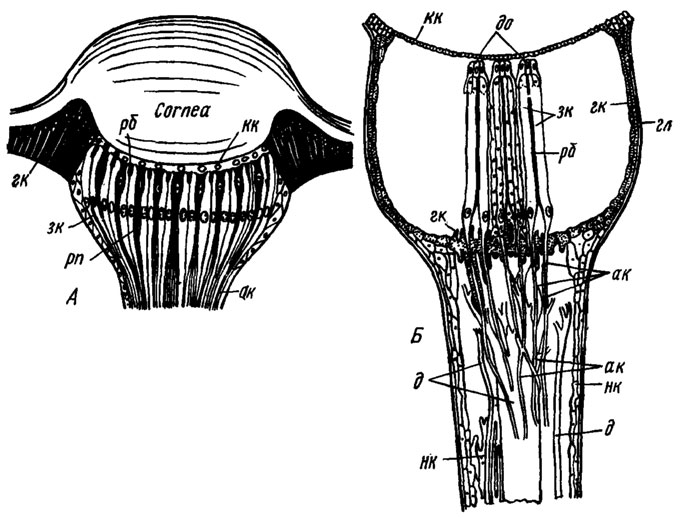
Строение дорсальных глазков насекомых (продольный разрез, схематично):
- А - схема типичного глазка (по Weber, 1949)
- Б - устройство бокового дорсального глазка стрекоз (роговичная линза удалена) (по Ruck, Edwards, 1964)
- ак - аксоны зрительных клеток в области их синаптического контакта с интернейронами
- гк - эпителиальные (гиподермальные) пигментные клетки
- д - дендриты интернейронов; до - дистальные окончания клеток тапетума
- зк - зрительные клетки, сгруппированные в ретинулы
- кк - корнеагенные клетки
- рб - рабдом
- нк - непигментированные клетки оцеллярной ножки, часть которых футляром одевает нервные волокна
- гл - расширенная часть глазка
- рп - ретинальные пигментные клетки
Эксперименты показали, что при закрашивании фасеточных глаз, насекомые перестают ориентироваться в пространстве, даже если у них имеются хорошо развитые дорсальные глазки. Это указывает на то, что такие глазки выполняют лишь вспомогательную функцию и не могут даже частично заменить основные фасеточные глаза. При этом сигналы с глазков влияют на активность нервных клеток не только переднего, но среднего и заднего мозга головы, причем одни клетки возбуждаются, другие тормозятся, что говорит о наличии двух типов связей (возбуждающей и тормозной) или специализированных связующих клеток с ONN/OFF ответом, подобных биполярам в сетчатке позвоночных. Такие связи могут указывать на то, что активность глазков служит фоновым контекстом при обработке сигналов с омматидиев. По этой причине они не могут самостоятельно служить полноценным детектором даже простейшего определения изменения освещенности, что показали опыта с закрашиванием фасеточных глаз, за исключением животных, где они являются единственными органами зрения. Так же замечена импульсация в шейных отлелах нервной цепочки в ответ на возбуждение глазков: включение света или движение в их поле зрения предметов, контрастирующих с фоном. Это опять подтверждает основную роль глазков как дополнительных детекторов для запуска вспомогательных реакций, в данном случае скорей всего для активации мышц шейного отдела при захвате в поле зрения объекта.
В темноте интернейроны глазка спонтанно генерируют 40-70 импульсов в секунду. При освещении частота пульсации сначала резко падает, а при слишком ярком свете вообще прекращается. Но затем через несколько минут устанавливается новый уровень пульсаций, тем ниже, чем выше освещенность. При выключении происходит обратный процесс: сначала частота импульсов возрастает, затем возвращается к базовому темновому уровню. Это наглядно показывает, что глазок не просто регистрирует изменение освещенности, но может отличать статическое освещение от темноты. А это уже ясное указание на детектирование новизны ситуации, что критически важно для процесса адаптации нервной системы. Таким образом можно сделать вывод, что глазок, несмотря на свою простоту, служит пусковым триггером для запуска и поддержания критически важных процессов в нервной системе насекомого. Уже давно сложилось мнение, согласно которому глазки регулируют фотокинетические реакции организма, обычно усиливая их при оптимальном или пониженном освещении и тормозя их при чрезмерном. Это доказывается тем, что закрашивание глазков, хотя бы одного из трех, более или менее существенно нарушает фотокинетические реакции: они замедляются, теряют четкость и требуют для своего осуществления более интенсивного освещения. Кроме всего вышесказанного, благодаря пульсации глазков центральная нервная система насекомого получает в темноте высокое тоническое возбуждение, что препятствует ее угнетению в отсутствии раздражителей.
Латеральные глазки
У личинок с полным превращением глазки являются единственным органом зрения, поэтому их функциональность более сложная. Такие глазки называют латеральными, или стемами. В отличие от дорсальных глазков они способны самостоятельно детектировать объекты, поэтому часто имеют схожее с омматидием строение: хрусталик, конус и группу клеток, образующих общий рандом. Личинка гусеницы с такой зрительной системой способа различать форму предметов, их цвет и движение. Очевидно, что это развилось как необходимая адаптация к ее образу жизни, где нужно более сложное реагирование: вовремя заметит опасность, определять направление движения на открытой местности и т.д. У других личинок ведущих более примитивный образ жизни этого может быть и не нужно, поэтому их глазки во многом подобны дорсальным.
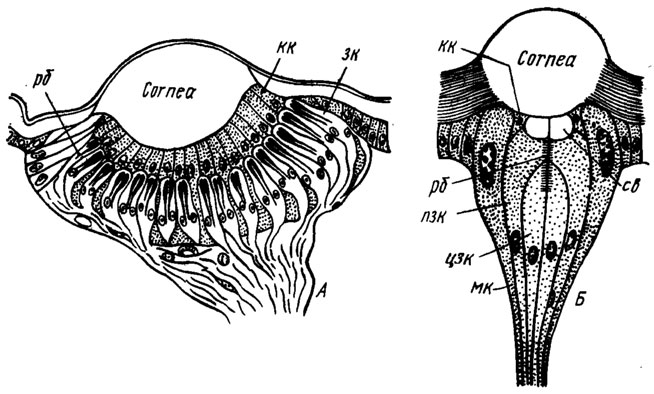 | Латеральные глазки личинок (продольный разрез схематично):
|
Заключение
Фасеточный глаз специализирован в первую очередь для восприятия движения и обеспечивает очень широкое поле зрения, что позволяет максимально быстро реагировать простейшими рефлексами на изменение визуальной картинки с любой стороны. Сравнивая фасеточный и камерный глаз по строению и функциональности можно найти как принципиально похожие, так и совершенно не похожие элементы. Логично сделать вывод: если в обеих зрительных системах что-то собрано по одинаковому плану, значит это принципиальное решение, иначе схема не будет работать. Различия же указывают на обратное – решение лишь частный случай, и критически не важно, как оно реализовано. В первом случае принципиальным является:
- Распознавание цвета при помощи специализированных сенсоров, реагирующих на определенную длину спектра. Таких рецепторов нужно минимум 2, обычно 3, но может быть и больше, смотря по специализации животного. Природа не смогла придумать иного способа различения цвета.
- Простое сведение сигналов от настроенных на разную длину волны рецепторов не способно получить итоговый суммарный цвет. Обязательно наличие контрастирования сигналов от рецепторов, для выделения наиболее яркого, и антагонистическое цветовое противодействие перекрывающихся областей рецепторной матрицы. Иначе точность распознавания будет очень низкой.
- Определение яркости/цвета области рецепторной матрицы происходит за счет сравнения разницы активности соседних рецепторов, которая обычно отличается не более чем в несколько десятков раз. Определение яркой точки на черном фоне происходит за счет контрастирования активных рецепторов на фоне менее активных. Это значит, что не нужно наличие рецепторов с огромным диапазоном чувствительности, например от 1 до 1 000 000 условных единиц.
- Зрительные образы определяются при помощи контрастных границ, состоящих из базовых примитивов, от простейших в виде точки до сложных многогранников.
- Глаз представляет собой многоканальный сенсорный орган, параллельно выдающий сигналы в разные отделы нервной системы, что позволяет формировать как простые, так и сложные иерархические рефлекторные реакции.
- Для организации режима многоканальности глаз функционально не однороден: рецепторы сгруппированы не равномерно по поверхности рецепторной матрицы в зависимости от своих свойств, образуя специфические зоны распознавания.
- Необходима предварительная обработка активности рецепторов на уровне глаза для «форматирования» информации в виде простейших примитивов восприятия, иначе, если переложить это на специализированные зрительные отделы, это потребует существенных вычислительных мощностей и быстро упрется в технический потолок реализации. Поэтому даже в простейших дорсальных глазках сигнал, прежде чем попасть в зрительные отделы проходит первичную обработку контрастирования на первом слое связанных с рецепторами нейронах. А в сетчатке это уже представлено в виде сложно связанного, многослойного массива нервных клеток.
Не принципиальным мне видятся следующие решения:
- Формирование итогового «цветного пикселя» при помощи двух типов рецепторов: палочек и колбочек, чувствительных к свету и цвету соответственно. Так сделано в сетчатке позвоночных, что позволяет им иметь 2 типа зрения: цветное и сумеречное черно-белое. Но это частный случай адаптации к двум фазам светового дня. Насекомым проще оказалось вести дневной/сумеречный/ночной образ жизни, создав разные типы компоновки фасетчатого глаза и простые механизмы темновой адаптации, не усложняя свою зрительную систему.
- Аккомодация зрительной системы, подстройка под различение близко и далеко расположенных объектов зависит от сложности организации нервной системы и необходимости получать такую информацию. «Внутренний мир» насекомых намного проще, им нет надобности в точном детектировании далеко расположенных объектов.
- Детализация объекта так же важна лишь относительно специализации животного в своей экологической нише. Например, богомолу не важно, какой конкретно вид насекомого или мелкого позвоночного станет ему добычей, важны лишь его размеры и некоторые общие признаки для оценки возможности справиться с ним.
- Ширина поля зрения, чувствительность к области спектра или особая специализация для восприятия поляризованного света – все это частные случаи адаптации животного к его среде обитания и образу жизни. Кому то достаточно зрительно различать лишь темные/светлые пятна, а кому то нужно с высоты нескольких сотен метров увидеть мышь в траве.