Природа устроена очень просто.
Иначе ничего бы не работало.
Вот только простоты этой много.
Отсюда и все сложности.
Информационная емкость мозга человека.
Шумилов В.Н.
Хотя сегодня о мозге и его строении известно много, на главный вопрос: «Как работает мозг?» пока нет ответа. Сегодня мозг представляется черным ящиком, на вход которого через рецепторы – органы чувств поступают сигналы, отображающие обстоятельства внешнего мира (в которых и находится организм с управляющим мозгом). Понятно, что на функционирование мозга влияют также обратные связи различной глубины через текущее физическое состояние и всего организма, и мозга. А также через сложившуюся к данному моменту структуру мозга, в которой отображен предыдущий опыт индивидуума в виде неких логических связей между элементами мозга.
До сих пор мы вынуждены были говорить о мозге, как о черном ящике, поскольку не было модели функционирования мозга, не было обоснованных представлений даже о принципах работы мозга, а не то, что о деталях его функционирования. В данной работе мы попытаемся разобраться в структуре этого до сих пор черного ящика. Хотя бы в принципах его функционирования.
На сегодня известно, что мозг состоит из отдельных логических элементов-нейронов, которые при определенных условиях могут возбуждаться (активизироваться) через свои входы сигналами с выходов предыдущих нейронов. И пребывать в возбужденном состоянии некоторое время, передавая при этом возбуждение дальше – к следующим логическим элементам-нейронам через связи от своих выходов (нейронов предыдущих звеньев) к входам последующих нейронов. Ясно, что, с точки зрения понимания логики работы мозга, не имеет значения, каким именно образом реализованы и как именно осуществляют анализ входных сигналов эти логические элементы-нейроны на уровне отдельного элемента – механически, химически или электрически. Главное – понять, какая логическая функция реализована на данном нейроне (или на группе нейронов). Поэтому мы не будем уделять особого внимания вопросам конкретного устройства логических элементов мозга – нейронов (их физиологии), а сосредоточимся на принципах, на логике их функционирования. Примеры же, для простоты, будем рассматривать в терминах электрических схем.
Прежде, чем говорить о возможном устройстве мозга, оценим информационную емкость мозга человека. Для этого воспользуемся некоторыми экспериментальными данными о мозге, существующими на сегодня и достаточно надежными. Для оценки информационной емкости мозга человека нам надо знать следующее:
_ нейроны связываются между собой (выход предыдущего на вход следующего),
_ N = количество нейронов в головном мозге,
_ L = среднее количество входов – дендритов у одного нейрона,
_ R = среднее количество нейронов, с которыми может связаться нейрон через один из своих дендритов; определяется средней длиной дендритов и аксонов,
_ D = различимое число значений веса логической связи между нейронами.
Отметим, что выход нейрона (аксон) представлен в единственном числе (хотя его единственность и не существенна для нашего рассмотрения). Множество всех остальных имеющихся на сегодня сведений о мозге человека (как фактов (результатов измерений), так и представлений (результатов рассуждений)) мы пока не будем рассматривать.
Сделаем естественное предположение, что информация в мозге отображается путем установления связей между нейронами головного мозга. Вообще-то, по большому счету, это предположение и предположением-то нельзя назвать. Можно сказать, что это уже устоявшееся и подтвержденное ([2]) представление.
Примем, что выходы нейронов работают не в аналоговом, а в чисто цифровом режиме, то есть, на выходе нейрона (логического элемента) сигнал или есть (уровень сигнала = 1), или его нет (вернее, сигнал нулевой, с уровнем 0). То есть, считаем, что сигналы с уровнем 0.25, 0.5, 0.75 на выходах нейронов не имеют места.
В то же время, как показывает эксперимент, по входу нейрон ведет себя как пороговое устройство. То есть, входы нейронов имеют различные веса (что эквивалентно различию их электрических сопротивлений, меняющихся со временем). Так что одинаковые сигналы, приходящие на разные входы одного и того же нейрона, имеющие разные веса, вызывают несколько различающуюся реакцию этого нейрона. Различие весов различных связей несет некую информацию. Информационная емкость веса такой связи определяется количеством различимых значений веса связи. Если веса связей неразличимы – информации в весах этих связей ноль. Если можно различить 2, 4, 8, … 1024 состояния веса связи, то значение веса этой связи несет в себе, соответственно, 1, 2, 3 … 10 бит информации. Пока неизвестно, какова информационная емкость входа нейрона за счет различия весов связей. Но в технике для оцифровывания аналоговых сигналов обычно используется порядка 12 двоичных разрядов (бит). Иногда – 16. Предположим, что природа достаточно близка к техническому оптимуму, и количество различаемых весов связи D = 4096 = 212 . То есть, информативность веса связи =12 бит.
Примем, в соответствии с имеющимися на сегодня лабораторными данными, что в человеческом мозге содержится N=15 миллиардов нейронов (=1.5 * 1010 ). У каждого из этих N нейронов имеется L=10 000 = 104 входов – дендритов и один выход – аксон [3]. Средняя длина дендрита около 1 мм. Тогда как аксоны имеют длину в сотни мм.
Каждый из N нейронов посредством каждого из своих L входов – дендритов (через один или через несколько входов) может быть связан с некоторыми L нейронами из числа R близлежащих нейронов, (в том числе, и с самим собой). Пусть каждый нейрон может связаться через один из своих входов (длиной 1 мм) с одним из своих соседей – нейронов, находящихся в пределах досягаемости для дендритов данного нейрона, то есть, расположенных внутри сферы с радиусом, равным длине дендрита r = 1 мм. Объем этой сферы V = (4п/3) * r3 ≈ 4.2 мм3 , и в объеме этой сферы находится в среднем около 42 000 нейронов. Тогда как все 15*109 нейронов мозга располагаются приблизительно в 1.5 * 106 мм3 (= 1500 см3 ) объема всего мозга. Так что в пределах досягаемости дендритов рассматриваемого нейрона расположено около 42 000 нейронов-соседей (самих ядер – сом нейронов). Но дендриты контактируют с аксонами, простирающимися на сотни мм от своего ядра. Так что можно смело сказать, что нейрон через каждый из своих входов может связаться более, чем с 80 000 других нейронов. То есть, R > 80 000.
Подсчитаем теперь, какое количество состояний, различающихся той или иной конфигурацией связей между нейронами, может иметь человеческий мозг. Для этого изобразим схематически нейроны мозга и связи между ними.
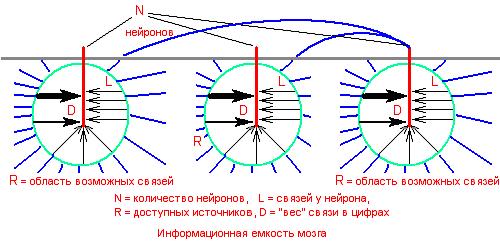
Произвольный нейрон через один из своих входов может установить с одним из R соседей связь, имеющую одно из 4096 возможных значений логического веса этой связи. То есть, каждая связь нейрона может находиться в одном их 320 000 000 состояний (R*D = 80 000 * 4096 = 216.3 * 212).
Так что при установлении связи через первый рассматриваемый вход (из L) с одним из R нейронов (или при неустановлении) рассматриваемый нами нейрон может реализовать один из (R*D) вариантов связей (с учетом веса связи). Второй вход этого же нейрона также может быть связан с одним из R нейронов (через связь с одним из D весов). Видим, что при задействовании двух входов может быть реализовано (R*D) * (R*D) вариантов соединений данного нейрона с другими. При задействовании следующего (третьего) входа может быть реализовано уже (R*D)*(R*D)*(R*D) = (R*D)3 вариантов. И так далее. Так что при задействовании всех L входов рассматриваемый нейрон может иметь одну из (R*D)L конфигураций связей через свои входы с другими нейронами. Точно так же каждый из (N-1) оставшихся нейронов может иметь одну из (R*D)L = ZL конфигураций входных связей независимо от конфигураций связей других нейронов. Что в итоге дает трудно вообразимое количество возможных конфигураций связей между нейронами системы из N нейронов (каждый из них с L входами, для каждого входа доступно (R*D) вариантов соединений) под названием «мозг человека»:
M = (ZL)*(ZL)*(ZL)*.....*(ZL) = (ZL)N = ((3.2*108 )10 000 )15 000 000 000
= ((3.2)(10 000 * 15 000 000 000) ) * (10(8 * 10 000 * 15 000 000 000) )
= ((3.2) 150 000 000 000 000 ) * (10 1 200 000 000 000 000 )
> 10 75 000 000 000 000 * 10 1 200 000 000 000 000
> 10 1 275 000 000 000 000 конфигураций!
Полученное количество конфигураций можно представить себе намного проще (привычнее в сегодняшнем мире), если сказать, что для представления такого количества конфигураций системы (для нумерации этих конфигураций) нужно иметь W двоичных разрядов – бит. Для определения количества бит, необходимых для указания на одну из этих возможных M конфигураций надо взять логарифм по основанию 2 от этого числа M.
То есть, количество информации, которое способен вместить в себя человеческий мозг за счет реализации тех или иных вариантов конфигурации связей между нейронами,
W = log2 ( M) = log2 ((ZL)N ) = log2 (((R*D)L)N ) = N * L * (log2 R + log2 D),
или
W = log2 ( M) = N * log2 ((R*D)L) = N * I,
Здесь через I мы обозначили среднее количество информации, которое может быть зафиксировано (размещено) в одном нейроне.
I = log2 ((R*D)L) = L * log2 (R*D) = 10 000 * log2 320 000 000 ≈ 10 000 * 28 бит.
Легко наглядно показать, что информационная емкость одного нейрона именно такая. Для этого отметим, что каждая из L=10 000 связей рассматриваемого нейрона установлена с одним из R=80 000 нейронов (связь указывает на один из R нейронов), каждый из которых как раз и адресуется приблизительно 16 битами (216 ≈ 64*103 ). Кроме того, информационная емкость веса каждой связи равна 12 битам (соответствует 4096 различимых значений веса связи).
Чтобы убрать возражения против учета информативности веса связи, примем пока, что информационная емкость каждой связи – входа нейрона определяется только возможностями адресации одного из 80 000 нейронов и составляет всего лишь 16 бит. То есть, I ≈ 10 000 связей * 16 бит/связь = 160 000 бит.
Поделим количество бит на 8, получим количество байт:
I ≈ 20 000 байт = 20 Кбайт,
Так что информационная емкость всего мозга W = N * I составляет не менее
W = N*I = (15*109)*20 000 = 300 * 1012 байт = 300 терабайт!
И это без учета весов связей! А с учетом весов связей I ≈35 Кбайт, W ≈500 терабайт.
Если принять, что на одной бумажной странице размещается 6 Килобайт информации, а 1000 страниц составляют один том (одну книгу), то емкость мозга человека (300 терабайт) составляет приблизительно пятьдесят миллионов таких тысячестраничных томов! Если мысленно поставить все эти тома (каждый толщиной порядка 3 см) на полку, то длина этой воображаемой книжной полки составит 1 500 км! Если бы человек прочитывал каждый день по одной такой книге, то за год он прочитал бы 365 томов примерно с 10 метров, так что за 100 лет были бы прочитаны книги примерно с 1 км этой воображаемой книжной полки. Конечно же, человек воспринимает информацию с гораздо большей скоростью, чем 1000 страниц в день.
Трудно представимое число (300 * 1012 байт) для наглядности можно свести еще к чему-нибудь, привычному на сегодняшний день. Предположим, что каждый день в мозге человека с предельной точностью фиксируется порядка 5*109 байт = 5 Гигабайт, что составляет приблизительно 4 часа фильмов на DVD. Для фиксации такого количества информации мозг должен был бы воспринимать ее непрерывно в течение суток (86400сек) со средней скоростью порядка 60 килобайт в секунду, что соответствует установлению приблизительно 30 000 новых связей между нейронами мозга за одну секунду.
Информационная емкость каждой связи (без учета ее логического веса) равна приблизительно 2 байта = 16 бит адресации одного из 80 000 нейронов, связывающегося посредством этой связи с нашим рассматриваемым нейроном. Так что 60 000 (байт/сек) / 2 (байта/на одну связь) ~ 30 000 (связей/сек). Эти 30 000 связей соединяют множество пар нейронов в цепочки (последовательно), а также отображают мозаику свойств объекта, попавшего в «поле зрения» мозга через входные рецепторы (параллельно). Можно сказать, что эти 30 000 отдельно установившихся связей эквивалентны установлению всех связей 3 нейронов в секунду.
При таком предположенном темпе фиксации информации мозг человека будет полностью заполнен примерно за 60 000 дней ((емкость мозга ≈ 300 * 1012 байт) / 5*109 (байт в день)). Или за 160 лет (точнее, 164).
Понятно, что мы запоминаем в день гораздо меньше, чем по 5 Гигабайт. Мозг наш не работает с механической точностью. С одной стороны, вполне возможна ситуация, когда одна и та же информация многократно дублируется (устанавливается множество параллельных связей). С другой стороны, какая-то информация утрачивается: то какие-то фрагменты картины окружающей действительности не дошли до места фиксации, то какие-то дошли, но не были зафиксированы. Какие-то были зафиксированы, но потом разрушились. И так далее. Именно поэтому об одном и том же событии 10 свидетелей сообщают 10 различных версий.
Видим, что вычисленная нами информационная емкость мозга приблизительно соответствует нашим интуитивным представлениям об объеме человеческой памяти.
Отметим, что все цифры, полученные в этой главе – оценочные. Демонстрирующие, в основном, методику их расчета. Но кроме методики, они позволяют также судить (в меру достоверности использованных экспериментальных данных) о порядке величины объема нашей памяти и скорости восприятия информации человеком.
Можно, конечно, заявить о возможной неразличимости возможных конфигураций идентичных связей нейронов, связанной с нумерацией их входов-дендритов. И поэтому информационная емкость каждого нейрона меньше, чем было принято выше. По той причине, что не имеет значения порядок, в каком различные дендриты одного нейрона связаны с выходами других нейронов. То есть, реально различимых состояний каждого нейрона (вариантов конфигурации связей) в L! (=L факториал) раз меньше, чем мы подсчитали (это из комбинаторики). Если согласиться с этим доводом, то получится, что количество различимых состояний связей нейрона не (R*D)L, а (R*D)L)/(L!). Так что информационная емкость одного нейрона I равна не 35 Кбайт, а на log2(L!) бит меньше. Воспользовавшись асимптотической формулой Стирлинга для значения факториала
k! ≈ (2πk)1/2 (k/e)k , получаем
_ I = – log2(L!) = –(0.5log2(2πL) + L log2 (L/e) )/8 байт = –(0.5*18 + 10 000 * 12)/8 б
_ I = – 15 000 байт,
То есть, если возможные состояния нейрона (конфигурации его связей) неразличимы при замене одного его входа-дендрита на другой, мы должны были бы принять информационную емкость одного нейрона равной приблизительно 20 Кбайт (35Кб-15Кб), что составляет примерно 57% от значения, полученного ранее.
Однако по поводу того, что состояния нейрона, различающиеся только «номерами» дендритов, нужно считать одинаковыми, неразличимыми, надо сказать следующее: дендриты не могут быть идентичными, уже хотя бы потому, что логические веса связей – дендритов различны. То есть, длины, а также электрические характеристики разных дендритов (время задержки, электрическое сопротивление и т.д.) различны. И поэтому состояния нейрона, различающиеся порядком соединения двух дендритов с двумя другими нейронами, существенно различаются. Да и вообще, весь мозг вряд ли можно считать чисто цифровым устройством с двумя возможными состояниями уровней сигналов (0 или 1) и на входах, и на выходах всех составных элементов. Мозг, скорее всего, эквивалентен гигантской логической схеме, реализованной в виде аналоговой (по входам) электрической схемы с очень изменчивыми порогами срабатывания элементов, составляющих ее. Хотя сигналы на аксонах–выходах нейронов имеют при этом цифровой характер. То есть, электрические потенциалы на выходах имеют всего два значения: «0» или «1». Аналоговая (даже только по входу) электрическая схема (по сравнению с дискретной, цифровой) имеет существенно большее количество возможных состояний каждого элемента и всей схемы – нейрона и мозга в целом, хотя имеет при этом существенно меньшую устойчивость, повторяемость, воспроизводимость результатов.
На примере обсужденной возможной неразличимости некоторых конфигураций связей между нейронами, видим, что, в принципе, возможны какие-то уточнения оценки информационной емкости мозга. Но в этом уточнении (размером в десятки процентов) нет особого смысла, поскольку экспериментальные оценки количественных характеристик мозга имеют разброс в разы (в сотни процентов).
Так, некоторые источники утверждают, что N – количество нейронов в головном мозге человека – намного больше, чем 15 миллиардов – до 100 миллиардов ([4]). И каждый из них может иметь до 20 000 связей с другими нейронами, что увеличивает расчетную информационную емкость мозга W ≈ 300*1012 байт (300 терабайт), вычисленную в соответствии с нашими допущениями, примерно в 12 раз. К тому же, как мы увидим позже, следует непременно учитывать различие логических весов связей между нейронами, что увеличит вычисленную нами при N = 15*109 и L = 10 000 информационную емкость мозга до 500*1012 байт или даже больше.
Так что не будем ничего уточнять, а примем пока, что информационная емкость каждого из 15 миллиардов нейронов составляет 20 Кбайт. Иначе, если мы начнем уточнять, то утонем в деталях. Придется говорить и о пороге чувствительности входов различных нейронов, и об изменении этого порога в зависимости от различных обстоятельств, как внутренних, так и внешних. А также и о других деталях, пусть даже очень существенных. А такое отклонение от главной темы не будет способствовать пониманию нами сути работы нашего мозга.
Мы подсчитали количество состояний мозга, различающихся конфигурациями связей между нейронами. Поэтому, даже если запоминание в мозге происходит не за счет установления связей между нейронами (а за счет чего-то другого, что сомнительно), информационная емкость мозга не меньше подсчитанной нами выше, вытекающей из количества различимых состояний мозга (количества возможных конфигураций связей между нейронами). Эти состояния мозга различаются, тем самым несут информацию, хотя мы можем и не знать, как извлечь и воспользоваться этой информацией.
Конечно, можно сказать, что сама по себе некоторая конфигурация связей между нейронами мозга в информационном смысле подобна конфигурации контактов между отдельными песчинками в куче песка, то есть, совершенно бессмысленна и бесполезна. Лишь тогда можно будет сказать, что в конфигурации связей нейронов содержится полезная для организма информация, когда будет показано, что существуют механизмы записи (фиксации), чтения (воспроизведения) и использования этой информации (фиксируемой в виде определенной конфигурации связей между нейронами мозга в нашем случае). Еще лучше, если эти механизмы будут известны и промоделированы. Эти механизмы записи, воспроизведения и полезности описаны в последующих главах.
Подтверждением того, что связи между нейронами действительно содержат актуальную информацию, является экспериментальный факт, состоящий в том, что через межнейронные связи нейроны возбуждают друг друга, так что сигнал (мысль, раздражение) проходит по тому или иному пути, определяемому конфигурацией связей между нейронами. Это и есть прямое доказательство того, что в конфигурации связей между нейронами отображена информация. Причем эта информация совершенно точно задействована в функционировании организма – путь прохождения сигнала через ту или иную цепочку связанных нейронов, вне всякого сомнения, самым существенным образом влияет на функционирование организма. Поскольку возбужденные на том или ином участке траектории сигнала нейроны в конечном итоге управляют соответствующими мышцами и подсистемами-органами. В свете сказанного становится очевидным, что в конфигурации межнейронных связей отображена (зафиксирована) жизненно важная информация, проявляющаяся (считывающаяся) в момент прохождения сигнала по той или иной цепочке нейронов (в зависимости от конфигурации возбуждающих сигналов).
Другое дело, что на сегодня пока неизвестно, как эта информация отображается, записывается (фиксируется), каков механизм прокладывания связей между нейронами. Ниже, в главах «Эволюция организмов», «Мозг – орган предвидения» и «Принципы функционирования мозга» мы опишем (представим) возможный способ возникновения таких связей между нейронами, то есть, покажем, каким именно образом может фиксироваться жизненно важная для организма информация в связях между нейронами.
Некоторое представление о принципах установления связей между парами нейронов мы можем получить чисто логически, исходя из самых общих соображений. Поскольку связи парные, то есть, каждая связь соединяет только два нейрона, то для установления этой связи имеет значение только состояние именно этих двух нейронов (при условии, что эта связь возможна, то есть, существует физическая возможность установления такой связи; скажем, геометрия расположения нейронов допускает образование этой связи).
Рассмотрим пример. Пусть в мозге между несколькими нейронами (A, B, C, D, E, R), отображающими следующие объекты-события внешнего мира:
А = на улице А состоялась, D = демонстрация,
В = на улице В состоялась, E = прогулка слона,
С = на улице С состоялась, R = церемония старта ралли,
в принципе, могут установиться такие связи, отвечающие реальности:
AD, AE, AR,
BD, BE, BR,
CD, CE, CR.
И не могут установиться связи AB, AC, BC, DE, DR, ER (хотя возможны связи между другими проекциями этих событий).
Пусть в какой-то момент времени устанавливается одна из возможных связей, отображающая реальную связь между событиями, имеющими место в данный момент, скажем,
АЕ = (на улице А состоялась) (прогулка слона).
То есть, в тот момент, когда одновременно были возбуждены два нейрона, отображающие события А и Е, между ними установилась связь. Тогда как другие допустимые, возможные связи, не установились, поскольку не было такого составного события, чтобы одновременно были возбуждены А и R, или А и D, В и D и так далее. Из этого рассмотрения делаем предварительный вывод, что из принципиально возможных связей между двумя нейронами устанавливаются только те, и только в тот момент, когда оба эти нейрона возбуждены. Иначе очень быстро (при первых же возбуждениях нейронов, или даже, вообще, без возбуждения) установились бы все допустимые по географическому расположению нейронов связи (но в подавляющем большинстве своем не отражающие реальных событий), и, тем самым, связи уже не несли бы в себе информации, отображающей действительность.
Можно утверждать, что связи между нейронами являются главным носителем информации, накопленной в мозге человека. Этот вывод вытекает уже из того факта, что любая возможность, любой механизм требует каких-то ресурсов, затрат. Ресурсы организма вполне ограничены, а ему для успешного своего функционирования необходимо иметь самые разные возможности. И, если какая-то функция – возможность в организме уже реализована, то при попытке дублирования этой функции у организма может уже не хватить ресурсов на другие возможности. Поэтому в организме, как и в любой самоорганизующейся системе, устанавливается баланс между затратами и полученными (имеющимися) возможностями (выгодами, полезностью). А поскольку на нейроны человеческого мозга и связи между ними затрачено около 1.5 кг, что составляет не менее 1.5% массы человека, то на другие виды памяти (на дублирование функции памяти) у организма уже просто нет ресурсов. Остальная масса человеческого организма (кроме 1.5 кг мозга) пошла на реализацию других систем жизнеобеспечения: движения, энергоснабжения (питания), доставки питательных веществ к различным системам организма и отвода от них использованных веществ, терморегулирования и т.д.
Литература
[1]. Дирак П. А. М. Принципы квантовой механики: Пер. с англ. 2-ое изд. — М.: Наука. Гл. ред. физ.-мат. лит., 1979. — 440 с.
[2]. А. Торгашев Голова процессора. http://www.ogoniok.com/4908/17/
[3]. Развитие и организация. Строение мозга.
http://www.brainer.ru/a4.html
[4]. Проничев И. В. Лекции по физиологии центральной нервной системы. Биолого-химический факультет УдГУ. http://www.distedu.ru/edu4/p_2