Дорсальные и латеральные глазки
РедактироватьГлазок служит не только для регистрации изменения освещения, но и для того, чтобы отличать неизменное освещение от темноты. Поскольку частота постоянной импульсации глазков градуально зависит от силы света, они могли бы давать информацию и об абсолютной освещенности. Наконец, благодаря глазкам центральная нервная система насекомого получает в темноте высокое тоническое возбуждение.
Вес уверенности: Вполне уверенно подтверждается независимыми исследователями
Дорсальные глазки
Кроме сложных глаз у многих взрослых насекомых имеются также фоторецепторы, состоящие из одного хрусталика (роговицы) и группы пигментных клеток, под которыми лежат ретинулярные клетки, образующие несколько рабдомов. Эти фоторецепторы именуются глазками (ocelli). Многие взрослые насекомые имеют дорсальные глазки, обычно в количестве трех, расположенных в виде треугольника на передней части головы. Иногда их бывает только один или два. Функция дорсальных глазков состоит в улавливании изменений интенсивности света.
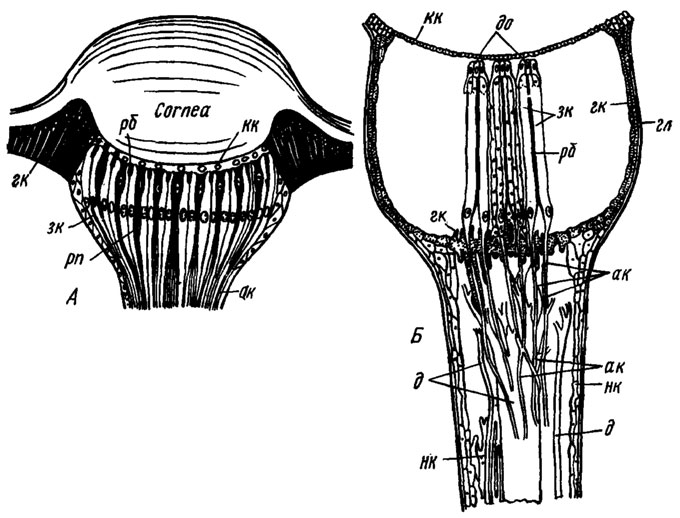 |
Строение дорсальных глазков насекомых (продольный разрез, схематично): А - схема типичного глазка (по Weber, 1949); Б - устройство бокового дорсального глазка стрекоз (роговичная линза удалена) (по Ruck, Edwards, 1964).
|
Из всех разновидностей глаз насекомых простые глазки обладают наиболее слабой способностью к зрению. По некоторым данным, они вообще не выполняют зрительной функции, и лишь отвечают за улучшение функции сложных глаз. Это, в частности, доказывается тем, что у насекомых практически не бывает простых глазков в отсутствии сложных. Кроме того, при закрашивании фасеточных глаз насекомые перестают ориентироваться в пространстве, даже если у них имеются хорошо выраженные простые глазки.
Оптический ганглий каждого глазка лежит вблизи его сетчатки в утолщенной части так называемой оцеллярной ножки, представляющей собой заключенную в футляр из непигментированных клеток совокупность нервных волокон, уходящих в межцеребральную часть переднего мозга. В этом ганглии аксоны зрительных клеток вступают в многочисленные синаптические контакты с дендритами нейронов второго порядка, тела которых лежат, видимо, в межцеребральной части мозга. Существенно, что число этих постсинаптических волокон значительно меньше числа пресинаптических волокон и что часть их отличается большой толщиной. В ножке медиального глазка саранчи Locusta migratoria содержится только шесть постсинаптических волокон: два более толстых, диаметром около 8 мкм, и четыре меньшей толщины - около 4 мкм. На обработанных хлоридом кобальта препаратах медиального глазка саранчи Schistocerca vaga обнаружено 17 больших билатерально симметричных глазковых интернейронов. В каждой ножке имеется по семь больших интернейронов, из которых четыре связаны сразу с двумя глазками. Один из интернейронов медиального глазка имеет в мозгу билатеральную связь, остальные ограничены каждый своей частью мозга. Обнаружена также связь ганглиозных интернейронов с оптическими ганглиями фасеточных глаз, в первую очередь с комиссурой между левым и правым оптическими ганглиями.
Судя по электрофизиологическим данным, сигналы с глазков влияют на активность нервных клеток не только переднего, но и среднего и заднего мозга головы. При освещении глазков спонтанная активность одних нервных клеток усиливается, других - тормозится. Замечена также импульсация в шейных коннективах нервной цепочки в ответ на возбуждение глазков: включение света или движение в их поле зрения предметов, контрастирующих с фоном.
Таким образом, дорсальные глазки связаны с мозгом чрезвычайно малым числом анатомически обособленных нервных каналов, и трудно ожидать, чтобы они передавали много информации. Но влияние сигналов с глазков прослеживается на многих мозговых центрах и даже на "исполнительной" части центральной нервной системы за пределами головы.
Электрическая активность глазка зависит от реакции его зрительных клеток и интернейронов оцеллярной ножки. Зрительные клетки реагируют на увеличение интенсивности света нарастающим деполяризационным потенциалом до 40 мВ. Крупные интернейроны ножки отвечают на свет гиперполяризационным потенциалом, максимальная амплитуда которого достигает 30 мВ. Для того чтобы вызвать ответ такого интернейрона, достаточно одной десятой интенсивности света, вызывающего реакцию отдельной зрительной клетки. Благодаря множеству контактов с разными рецепторами сетчатки динамический диапазон ответов интернейронов глазка, по сравнению с его зрительными клетками, сдвинут на более низкие интенсивности света.
В темноте интернейроны оцеллярной ножки мух, пчелиных, саранчовых и других насекомых спонтанно генерируют 40-70 импульсов в секунду. Освещение сначала сильно подавляет частоту импульсации, а при достаточно ярком свете активность интернейрона совсем прекращается. Но по истечении двух-трех минут устанавливается новый уровень импульсации, который тем ниже, чем выше освещенность. При выключении света частота импульсов возрастает, но через две-три минуты возвращается к стационарному темновому уровню. Оба переходных процесса, видимо. отражают адаптационные изменения, протекающие в глазке достаточно быстро.
Следовательно, по данным электрофизиологии, глазок служит не только для регистрации изменения освещения, но и для того, чтобы отличать неизменное освещение от темноты. Поскольку частота постоянной импульсации глазков градуально зависит от силы света, они могли бы давать информацию и об абсолютной освещенности. Наконец, благодаря глазкам центральная нервная система насекомого получает в темноте высокое тоническое возбуждение.
Если судить ,по поведению насекомых и результатам электрофизиологических экспериментов, глазки физиологически не равноценны фасеточным глазам и не заменяют последние, а только дополняют их. Мухи, пчелы, муравьи, саранчовые и другие насекомые с закрашенными фасеточными глазами и интактными дорсальными глазками ведут себя как слепые, не ориентируясь даже по обычному источнику света. Вместе с тем закрашивание глазков, хотя бы одного из трех, более или менее существенно нарушает фотокинетические реакции: они замедляются, теряют четкость и требуют для своего осуществления более интенсивного освещения. Поэтому уже давно сложилось мнение, согласно которому глазки регулируют фотокинетические реакции организма, обычно усиливая их при оптимальном или пониженном освещении и тормозя их при чрезмерном освещении. Предполагается, что сигналы с глазков влияют в первую очередь на чувствительность интернейронов фасеточного глаза, хотя стимулирующий эффект глазков замечен даже на сердечной активности насекомого: их освещение вызывает тахикардию у неподвижно сидящей мухи.
Причастность глазков к ориентации непосредственно замечена по поведению мух Sarcophaga и нескольких видов шмелей. Эти насекомые с закрашенными фасеточными глазами, но не тронутыми глазками реагируют на поворот плоскости поляризации света. Глазки отличаются от фасеточных глаз намного большей светосилой и могут функционировать даже в сумерках, когда света недостаточно для работы фасеточных глаз. В результате суточный период активности насекомого удлиняется благодаря глазкам. По крайней мере, на перепончатокрылых замечено, что при увеличении размера глазков расширяется диапазон освещенностей, в которых насекомое активно. Различные виды пчел и ос прекращают фуражировку при определенной пороговой интенсивности освещения, которая оказывается тем ниже, чем крупнее глазки, причем виды с глазками более 0,37 мм в диаметре летают даже ночью. Если у медоносной пчелы закрасить дорсальные глазки, то она утром вылетает позднее, а вечером возвращается в улей раньше, чем контрольные интактные пчелы. Следовательно, глазки действительно позволяют насекомому оценивать абсолютную освещенность.
Латеральные глазки
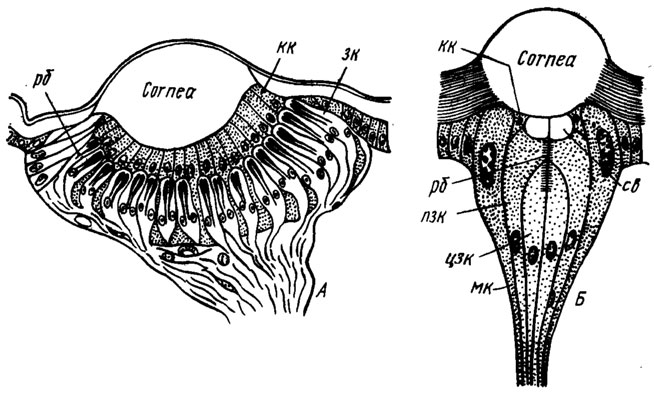 | Латеральные глазки личинок (продольный разрез схематично):
|
У личинок насекомых с полным превращением сложных глаз нет вообще, а есть латеральные глазки, или стеммы. Стеммы бывают самого разного строения. У гусениц они устроены как омматидий — с хрусталиком, кристаллическим конусом и группой ретинулярных клеток, образующих один рабдом. С помощью таких стемм гусеница различает форму предметов, цвет и движение. У других насекомых стеммы сходны по строению с дорсальными глазками.
Предполагается, что глазки могут служить анализаторами движения крупных объектов, учитывающими направление их движения относительно стрекозы, в частности приближение или удаление. Так, медиальный глазок Sytnpetrutn flaveolum сигнализирует о движении крупного предмета в основном сверху вниз, а латеральные глазки - о движении вверх-вниз.
Роговичная линза и кристаллическое тело глазка функционируют как единая оптическая система. Эта система чрезвычайно короткофокусна (f≈20 мкм), поэтому ее относительное отверстие превышает 1:1,0 и, следовательно, светосила глазка очень велика. Благодаря малой величине фокусного расстояния создаваемое оптической системой изображение незначительно перемещается по глубине, когда объект приближается к глазку или удаляется от него. Поэтому резкое изображение объекта всегда остается в плоскости рабдома, вытянутого вдоль оптической оси. Поле зрения отдельного глазка такого типа, измеренное у личинки муравьиного льва электрофизиологическим способом, равно примерно 47°, а его чувствительность даже под углом 8° к оптической оси не превышает 50% от максимальной.
В глазках с трехраздельной роговицей образуются соответственно три изображения одной и той же точки. Последнее обстоятельство, а также узкое поле зрения и вытянутый "двухэтажный" рабдом свидетельствуют в пользу того, что отдельный глазок гусеницы не предназначен для восприятия деталей предметов и функционирует подобно одиночному омматидию сложного глаза. Шесть глазков расположены так, что каждый имеет обособленное, практически независимое поле зрения. Поэтому гусеница одновременно видит с каждой стороны головы, очевидно, только шесть независимых точек, каждая из которых есть совокупность всех лучей, попадающих в один глазок. Мозаика этих точек слишком груба, чтобы различать детали и форму предмета. Но малочисленность зрительных единиц, возможно, компенсируется их движением: "привычкой" большинства гусениц раскачивать переднюю часть тела из стороны в сторону и таким способом сканировать окружающие предметы.
Источник:
Последнее редактирование: 13.12.2020
Чтобы оставить комментарии нужно авторизоваться.