Темновая адаптация
РедактироватьУ насекомых темновая/световая адаптация в отличии от позвоночных менее универсальна и требует наличия специально адаптированного глаза. Видимо потому, что насекомым проще переключиться на строго ночной/дневной образ жизни, чем пытаться адаптироваться одинаково хорошо в обоих режимах. И у них в меньшей степени встречаются эволюционные изменения образа жизни от ночного к дневному, от монохромного к цветовому зрению скорей всего по тому, что основной принцип выживания насекомых - огромная плодовитость, а не модификация имеющихся приспособлений.
Вес уверенности: Вполне уверенно подтверждается независимыми исследователями
Изображение, сформированное хрусталиком и кристаллическим конусом, попадает на светочувствительные ретинальные (зрительные) клетки, представляющие собой нейрон с коротким хвостиком-аксоном. Несколько ретинальных клеток образуют единый цилиндрический пучок — ретинулу. Внутри каждой такой клетки на стороне, обращенной внутрь омматидия, расположен рабдомер — особое образование из множества (до 75–100 тыс.) микроскопических трубочек-ворсинок, в мембране которых содержится зрительный пигмент. Как и у всех позвоночных, этим пигментом является родопсин — сложный окрашенный белок. Благодаря огромной площади этих мембран фоторецепторный нейрон содержит большое количество молекул родопсина (например, у плодовой мушки Drosophila это число превышает 100 млн!).
Рабдомеры всех зрительных клеток, объединенные в рабдом, и являются светочувствительными, рецепторными элементами фасеточного глаза, а все ретинулы в совокупности составляют аналог нашей сетчатки.
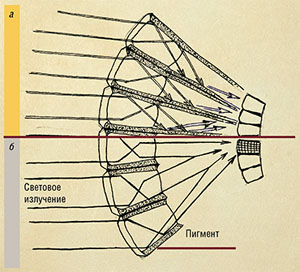 | Глаза насекомых, ведущих сумеречный или ночной образ жизни, отличаются особыми скотопическими омматидиями. В их экранирующих клетках пигменты могут мигрировать: при достаточном количестве света они распределяются равномерно (а), а при недостатке — скапливаются в верхней части клеток (б). В результате в темное время световое излучение с одного омматидия может попадать на рецепторные клетки соседних омматидиев. Рис. Н. Крюковой |
Светопреломляющий и светочувствительный аппарат фасетки по периметру окружают клетки с пигментами, которые играют роль световой изоляции: благодаря им световой поток, преломляясь, попадает на нейроны только одного омматидия. Но так устроены фасетки в так называемых фотопических глазах, приспособленных к яркому дневному свету.
Для видов, ведущих сумеречный или ночной образ жизни, характерны глаза другого типа — скотопические. Такие глаза имеют ряд приспособлений к недостаточному световому потоку, например, очень большие рабдомеры. Кроме того, в омматидиях таких глаз светоизолирующие пигменты могут свободно мигрировать внутри клеток, благодаря чему световой поток может попадать на зрительные клетки соседних омматидиев. Этот феномен лежит в основе и так называемой темновой адаптации глаз насекомых — увеличении чувствительности глаза при недостаточном освещении.
При поглощении рабдомерами фотонов света в ретинальных клетках генерируются нервные импульсы, которые по аксонам направляются в парные зрительные доли головного мозга насекомых. В каждой зрительной доле имеется по три ассоциативных центра, где и осуществляется переработка потока зрительной информации, одновременно идущей от множества фасеток.
Другой механизм адаптационной подстройки чувствительности глаза основан на изменении волноводных свойств рабдома, регулирующих в конечном счете угловое поле зрения рецептора. Так, у саранчи Locusta migratoria при световой адаптации палисад из вакуолей эндоплазматического ретикулюма вокруг рабдома распадается, и вместо него здесь скапливаются митохондрии, коэффициент преломления которых ближе к таковому рабдома, чем у палисада. В результате ухудшаются условия светопроведения, меньше света удерживается рабдомом при многократном отражении лучей на границе с митохондриями, а поле зрения рабдома при этом сужается. В итоге падает уровень светочувствительности глаза. И наоборот, образование палисада в темноте повышает светочувствительность глаза, поскольку уменьшается угол полного внутреннего отражения рабдома, что приводит к расширению его поля зрения.
Сходное явление описано у мухи Musca domestica. Оно наблюдалось за счет миграции гранул экранирующего пигмента в цитоплазме зрительных клеток. При освещении глаза гранулы экранирующего пигмента быстро, в течение нескольких секунд, мигрируют к рабдомерам, вследствие чего в последних нарушаются условия для полного внутреннего отражения и свет частично рассеивается, частично поглощается экранирующим пигментом. Это влечет за собой сужение поля зрения рецепторов, что показано по уменьшению угла, в пределах которого свет вызывает электрический ответ зрительной клетки. Таким способом фоторецепторы мухи защищаются от избытка света. Регуляция осуществляется, по-видимому, на основе петли обратной связи между интенсивностью возбуждения зрительной клетки и количеством мигрирующего в ней защитного пигмента, т. е. она похожа по своей сущности на так называемый "зрачковый рефлекс" камерного глаза позвоночных.
Источник:
Последнее редактирование: 07.12.2020
Чтобы оставить комментарии нужно авторизоваться.