Мотивации
Мотивации
http://med-stud.narod.ru/natur/n_fiz/motiv.html
Мотивация — вызванное той или иной потребностью эмоционально окрашенное состояние организма, избирательно объединяющее нервные элементы различных уровней мозга. На основе мотиваций формируется поведение, ведущее к удовлетворению исходной потребности.
Проблема мотивации занимает ведущее место среди других фундаментальных проблем высшей нервной деятельности. Если еще в середине нашего столетия мотивации находились целиком в сфере научных интересов психологов, то в последние годы мотивации стали предметом пристального исследования физиологов. В изучении мотиваций наметился значительный прогресс. Традиционный поведенческий и электрофизиологический подходы все активнее соединяются с изучением молекулярных механизмов мотиваций. Серьезному исследованию подверглись социальные мотивации человека.
Мотивация как основа целенаправленной деятельности. Каждый, кто внимательно наблюдал поведение живых существ, не раз обращал внимание на то, что среди разнообразных форм поведения встречаются такие, когда человек или животное не только реагируют на внешние воздействия, но активно ищут определенные объекты внешней среды. Поиск этот осуществляется весьма настойчиво, с большими затратами энергии и преодолением различных препятствий на пути к цели.
Такие формы активной целенаправленной деятельности живых существ были названы И. П. Павловым основными влечениями организма.
К числу основных влечений, направленных на обладание определенными раздражающими предметами, И. П. Павлов в первую очередь относил влечение голода и ориентировочно-исследовательскую деятельность. Нетрудно заметить, что основные влечения организма представляют собой именно такую форму поведенческой деятельности, которая в отличие от рефлекторной деятельности как реакции на внешние стимулы, напротив, направлена на поиск специальных раздражителей внешней среды.
Термин "влечение" в последние годы в отечественной литературе употребляется редко. Чаще используются его синонимы — "побуждение", или "мотивация" (movere — движение, побуждение). В зарубежной литературе часто применяется понятие "драйв", соответствующее понятию "основные влечения" организма.
КЛАССИФИКАЦИЯ МОТИВАЦИЙРазличают биологические и социальные мотивации.
Биологические мотивации, они же основные влечения, или низшие, простые, первичные мотивации. Биологические мотивации направлены на удовлетворение ведущих биологических потребностей индивидуумов по сохранению их вида или рода. К ним относятся мотивации голода, жажды, страха, агрессии, половые влечения, различные родительские, в частности материнские, температурные и другие влечения. Близко к этой группе мотиваций примыкают так называемые позывы, например к мочеиспусканию или дефекации.
Ведущими биологическими потребностями являются: 1) пищевая потребность, характеризующаяся уменьшением в организме уровня питательных веществ; 2) питьевая потребность, связанная с повышением осмотического давления; 3) температурная потребность — при изменении температуры тела; 4) половая потребность etc.
Поскольку общие потребности организма многопараметренны, животные могут одновременно испытывать несколько потребностей. Однако всегда имеется ведущий параметр общей метаболической потребности — доминирующая потребность, наиболее важная для выживания особи или ее рода, которая строит поведенческий акт, направленный на ее удовлетворение.
Социальные мотивации, они же высшие, или вторичные, строятся на основе врожденных биологических мотиваций путем общения индивидуумов со средой обитания, родителями и окружающими их живыми существами, а у человека — и с социальной средой. В формировании социальных мотиваций значительное место принадлежит воздействию различных факторов внешней среды, обучению и, следовательно, механизмам памяти. Все эти факторы могут значительно изменить природу лежащих в основе социальных биологических мотиваций. Достаточно сравнить голод животного с голодом воспитанного человека. В отличие от животного, которое будет использовать все свои биологические, в основном физические, способности для удовлетворения потребности пищи, голодный человек может долго стоять перед заполненной самыми привлекательными явствами витриной магазина, не производя никакого правонарушающего действия. На основе воспитания удовлетворение пищевых, половых и других биологических потребностей у человека, как правило, приурочено к определенному месту и времени.
Социальные мотивации человека, такие как стремление к образованию, определенного рода профессии, предметам искусства, литературы и быта, в отличие от биологических мотиваций также формируются в процессе общественного воспитания. Они обусловливаются общественными нормами морали и права и соответствующими законодательствами, присущими любому общественно-экономическому строю.
Как биологические, так и социальные мотивации определяют практически все формы выраженной целенаправленной деятельности живых существ, строящейся на основе иерархии различных биологических и социальных потребностей. Ведущая в биологическом или социальном плане потребность становится доминирующей, остальные выстраиваются по отношению к ней в определенной иерархической зависимости. После удовлетворения ведущей потребности доминирующей становится другая, наиболее важная в биологическом или социальном значении потребность etc. В формировании мотиваций и их иерархической смене ведущую роль играет принцип доминанты, сформулированный А. А. Ухтомским.
Ниже будут рассмотрены свойства и механизмы биологических мотиваций. Социальные мотивации человека являются предметом курса психологии.
ОБЩИЕ СВОЙСТВА БИОЛОГИЧЕСКИХ МОТИВАЦИЙБиологические мотивации характеризуются рядом общих свойств.
ГЕНЕТИЧЕСКАЯ ДЕТЕРМИНИРОВАННОСТЬБиологические мотивации, будучи тесно связанными с метаболическими потребностями организма, строятся на основе врожденных, генетически детерминированных механизмов. Их проявление не требует обучения, так как определяется активностью специальных генов, созревающих или освобождающихся от тормозных влияний на определенных стадиях онтогенетического развития. При этом в качестве основных факторов ресупрессии генетического аппарата выступают в первую очередь различные метаболические потребности и те информационные молекулы, которые эти потребности сопровождают.
СООТНОШЕНИЯ ВНУТРЕННИХ И ВНЕШНИХ ФАКТОРОВНепосредственной причиной биологических мотиваций являются преимущественно раздражители внутренней среды, связанные с изменением различных показателей гомеостаза, определяющих нормальное течение обменных процессов в организме, например, уровня питательных веществ, осмотического давления, состояния половых клеток, различных гормонов etc. Эти в различной степени выраженные изменения тех или иных метаболических показателей, будучи в значительной степени гуморальными изменениями, выступают в роли исходных стимулов биологических мотиваций.
Наряду с этим биологические мотивации могут активироваться или, наоборот, тормозиться специальными внешними "ключевыми" или "освобождающими" факторами внешней среды, например, видом или запахом противника или полового партнера, пищи или других удовлетворяющих различные потребности раздражителей. Например, накормленная обезьяна-самец может спокойно дремать в клетке, но поведение животного резко активизируется, если в клетку впускают рецептивную самку. Ее вид, запах и другие раздражители стимулируют у самца половую мотивацию. Вместе с тем следует иметь в виду, что в основе мотиваций, вызываемых внешними стимулами, всегда лежат врожденные, наследственно обусловленные внутренние механизмы. Например, агрессии способствуют определенные изменения нервных центров под влиянием нейрогормонов адреналина и норадреналина, половым реакциям — первичное воздействие половых гормонов на нервные клетки etc.
Соотношение внешних и внутренних факторов в возникновении биологических мотиваций в разных условиях может меняться. Например, голод в одних случаях стимулируется первичными метаболическими изменениями внутри организма, в других — видом или запахом аппетитно приготовленной пищи.
Биологические мотивации нередко относятся к разряду сложных безусловных рефлексов, или инстинктов. Вместе с тем биологические мотивации в отличие от рефлекторных реакций на внешние стимулы представляют такую форму деятельности, когда животные и человек, стимулируемые внутренними потребностями или окружающими факторами, направленно ищут специальные внешние раздражители. Именно настойчивый поиск отличает мотивационные реакции от рефлекторных.
СИСТЕМНАЯ ОРГАНИЗАЦИЯ МОТИВАЦИЙМотивации как биологического, так и социального плана в целостной деятельности организма всегда являются компонентом системной архитектоники поведенческого акта. Мотивации играют существенную роль в организации таких ответственных системных механизмов, как стадия афферентного синтеза, принятия решения и предвидения потребного результата — акцептора результата действия. Не меньшее значение мотивация имеет в формировании эфферентной программы поведения — стадии эфферентного синтеза.
Системогенез мотиваций. Будучи неотъемлемым компонентом системной организации поведенческих актов, мотивации в своем становлении в развивающемся организме подлежат общим закономерностям развития функциональных систем — процессам системогенеза. В процессе онтогенетического развития отдельных функциональных систем мотивации, будучи их составным компонентом, созревают избирательно и последовательно. Так, у новорожденного ребенка уже к моменту рождения формируются мотивации голода и жажды; затем на основе обучения и влияния внешних факторов проявляются мотивации страха, температурные мотивации, формируются позывы; в период полового созревания проявляется половое влечение, и уже во взрослом состоянии — родительские мотивации.
ТЕОРИИ МОТИВАЦИЙОБЩИЕ ТЕОРИИ
Наиболее распространены две крайние общие теории о природе мотиваций. Авторы одной теории считают, что мотивация определяется внутренней, врожденной, энергией, которая реализуется в специальное поведение даже при отсутствии внешних, "ключевых", раздражителей. Другие авторы полагают, что внутренние мотивационные программы развертываются только при наличии определенных направляющих факторов внешней среды. Некоторые авторы допускают, что направляющими факторами мотиваций могут быть и определенные внутренние, врожденные и приобретенные в индивидуальной жизни животных модели поведения.
В последнее время большинство исследователей склоняются к компромиссной точке зрения, согласно которой любая мотивация имеет два аспекта. Один из них чисто энергетический, побуждающий фактор, а другой — фактор, направляющий животных к цели.
Среди общих теорий мотиваций следует отметить теорию "снижения влечений", согласно которой мотивации определяются стремлением человека и животных к уменьшению неприятных эмоциональных ощущений, сопровождающих ту или иную метаболическую потребность. Согласно этой теории, голодные влечения, например, рассматриваются как стремление субъектов избавиться от "голодных" болей, возникающих в эпигастральной области при сокращениях пустого желудка; жажда — как стремление избавиться от неприятных ощущений в ротовой полости и в глотке etc. В развитие этой точки зрения сформулировано представление о наличии в мозге двух реципрокных систем: системы поощрения и наказания. В указанной теории "снижения влечения" в происхождении эмоциональных ощущений ведущую роль отводят нервным импульсациям, поступающим в мозг от определенных периферических органов.
ФИЗИОЛОГИЧЕСКИЕ ТЕОРИИАмериканский физиолог У. Кэннон первый сформулировал представление о роли сигнализации различных отделов пищеварительного тракта в происхождении мотиваций голода и жажды и перешел от изучения чисто поведенческой стороны мотиваций к изучению их внутренних механизмов. Однако эта точка зрения не нашла убедительного экспериментального подтверждения. Отечественным физиологом Н.Ф. Поповым было показано, что при полной денервации пищеварительного тракта у животных сохраняются мотивации голода. Клинические наблюдения тоже указывали на то, что после тотальной резекции желудка больные по-прежнему испытывают чувство голода. Все эти наблюдения способствовали выявлению роли гуморальных факторов в возникновении биологических мотиваций. И.П. Павлов первый указал на роль "голодной крови" в происхождении мотивации голода. Тем не менее наблюдения, в первую очередь на сросшихся близнецах, имеющих общее кровообращение и различную иннервацию, поставили под сомнение ведущее значение гуморальных факторов в формировании биологических мотиваций. Попытки все более углубленного анализа мотивационных возбуждений привели к необходимости каким-то образом объединить нервные и гуморальные механизмы, участвующие в их возникновении, в единый нервно-гуморальный механизм. Так возникли концепции множественных факторов формирования мотиваций.
К середине нашего столетия интерес исследователей в изучении механизмов мотиваций постепенно переместился к изучению роли ЦНС.
По аналогии с "центральным возбудительным состоянием", которое было сформулировано Ч. Шеррингтоном, возникло представление о "центральном мотивационном состоянии". Французский исследователь П. Делл обосновал точку зрения, согласно которой "центральное мотивационное состояние" определяется неспецифическими восходящими активирующими влияниями ретикулярной формации на кору мозга. Близка к этим представлениям "теория активации эмоции" американского физиолога Р. Линдсли, который также считает, что эмоциональные реакции различного качества строятся на основе восходящих активирующих влияний ретикулярной формации ствола мозга.
Американский исследователь Е. Стеллар сформулировал теорию, которая связывает формирование мотиваций с деятельностью специальных мотивациогенных гипоталамических центров головного мозга, на которые оказывают влияние кора, тормозные гипоталамические центры, чувствительные стимулы, гуморальные факторы etc. Эта точка зрения нашла подтверждение в многочисленных исследованиях. Так, при электрическом или химическом раздражении заднелатерального отдела гипоталамуса в области свода оказалось возможным получить у коз выраженную питьевую реакцию. При стимуляции латерального отдела гипоталамуса у различных животных возникают направленные пищевые реакции. Эти и другие эксперименты позволили говорить о наличии на уровне гипоталамуса так называемых мотивациогенных центров голода и жажды, агрессии и страха etc.
Многими исследователями показано, что в формировании мотиваций различного биологического качества наряду с гипоталамическими структурами принимают участие лимбические и ретикулярные структуры мозга, включая различные отделы коры большого мозга.
МЕХАНИЗМЫ ФОРМИРОВАНИЯ БИОЛОГИЧЕСКИХ МОТИВАЦИЙВедущую роль в формировании биологических мотиваций играет гипоталамическая область мозга. Здесь осуществляются процессы трансформации биологической (метаболической) потребности в мотивационное возбуждение. Гипоталамические структуры мозга на основе их влияний на другие отделы мозга определяют формирование обусловленного мотивацией поведения.
ТРАНСФОРМАЦИЯ ВНУТРЕННЕЙ ПОТРЕБНОСТИ В МОТИВАЦИОННОЕ ВОЗБУЖДЕНИЕКак показывают физиологические наблюдения, внутренняя метаболическая потребность находит свое отражение прежде всего в деятельности определенных внутренних органов и изменении состава крови.
Нервные и гуморальные сигналы о доминирующей биологической потребности адресуются специальным зонам гипоталамуса. Например, нейроны, воспринимаюшие сигнализацию от пустого желудка, и факторы "голодной крови" обнаружены микроэлектродным методом в области латерального гипоталамуса (Б.В. Журавлев). Отдельные нейроны этой области проявляют отчетливые реакции на введение как пищи голодным животным в желудок, так и глюкозы в кровь.
СВОЙСТВА МОТИВАЦИОННЫХ ЦЕНТРОВЦентры гипоталамуса обладают рядом свойств.
1. Особенность нейронов гипоталамической области состоит в их тесных функциональных контактах с капиллярами и в специфике метаболических процессов. Последнее заключается в том, что каждая группа нейронов гипоталамической области использует в своем нормальном метаболизме только определенные гуморальные факторы и при изменении их содержания избирательно приходит в состояние возбуждения. Таким образом, эти нейроны обладают свойствами рецепции определенной внутренней потребности.
Благодаря специфическому метаболизму различные участки гипоталамической области и составляют так называемые мотивациогенные центры.
В настоящее время в области вентромедиальных ядер гипоталамуса и в латеральных его отделах обнаружены нейроны, избирательно чувствительные к содержанию в крови глюкозы. Эти нейроны проявляют подчеркнутую специализацию. Одни нейроны активируются при снижении уровня глюкозы в крови, другие, наоборот, при возрастании ее содержания. В области супраоптических и паравентрикулярных ядер гипоталамуса обнаружены нейроны, чувствительные к уровню осмотического давления крови, в области заднедорсального гипоталамуса — к норадреналину etc. Электрическая стимуляция именно этих областей гипоталамуса вызывает соответственно пищевые, питьевые мотивации и мотивации страха и агрессии.
На основе рецепторных свойств мотивациогенные центры гипоталамической области обладают способностью к трансформации внутренней, преимущественно гуморальной, потребности в процесс нервного возбуждения.
2. Процесс возбуждения мотивациогенных центров гипоталамуса осуществляется, как правило, ритмически. Возбуждение в составляющих эти центры клетках появляется не сразу при возникновении той или иной внутренней потребности, а благодаря постепенному возрастанию их возбудимости до критического уровня (триггерный механизм). При достижении этого уровня клетки начинают посылать ритмические разряды и проявляют свою специфическую активность до удовлетворения потребности (см. рис.).
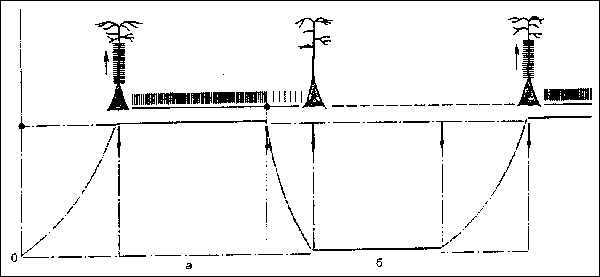
Рис. Триггерный механизм возбуждения нейрона гипоталамуса под влиянием метаболической потребности (а) и прекрашение импульсной активности при ее удовлетворении (б). Красной линией обозначено изменение возбудимости нейрона.
3. Гипоталамические мотивациогенные центры имеют обширные связи с другими отделами мозга и в первую очередь с лимбическими и ретикулярными образованиями, а через них — с корой большого мозга. Из этого следует, что возникшее первично в нейронах гипоталамической области мотивационное возбуждение может широко распространяться практически на все области мозга.
ОТРАЖЕНИЕ МОТИВАЦИИ В ЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ МОЗГАДоминирующая мотивация находит характерное отражение в электроэнцефалограмме. Особенно отчетливо биологические мотивации выявляются у животных, находящихся под уретановым наркозом.
Особенность уретана как наркотика состоит в том, что под уретановым наркозом у животных избирательно блокируется только ЭЭГ-активация бодрствующего состояния, но при этом сохраняются восходяшие активирующие влияния ретикулярной формации на кору мозга, например, при ноцицептивном раздражении животных. Точно так же под уретановым наркозом удается выявить активацию коры большого мозга, обусловленную голодным состоянием животных. У кошки после 2-суточного голодания при погружении в уретановый наркоз наблюдается избирательная ЭЭГ-активация передних отделов коры мозга, в то время как теменно-затылочные отделы мозга проявляют медленную высокоамплитудную ЭЭГ-активность, характерную для состояния наркотического сна. Тот факт, что эта активация ЭЭГ действительно отражает мотивацию голода, доказывают следующие опыты. После искусственного кормления животных, обнаруживающих под уретановым наркозом избирательную активацию ЭЭГ передних отделов коры мозга, при внутривенном введении раствора глюкозы или молока в ротовую полость и желудок активация ЭЭГ передних отделов коры мозга сменяется медленной высокоамплитудной активностью, и такая электрическая активность регистрируется во всех отделах коры мозга. Характерно, что у животных, погруженных в уретановый наркоз, немедленно после кормления во всех отделах коры мозга также регистрируется медленная высокоамплитудная электрическая активность.
Градуальность формирования мотивации. Эксперименты показали, что распространение мотивационного возбуждения на различные структуры мозга происходит градуально в зависимости от выраженности исходной потребности (см. рис.).
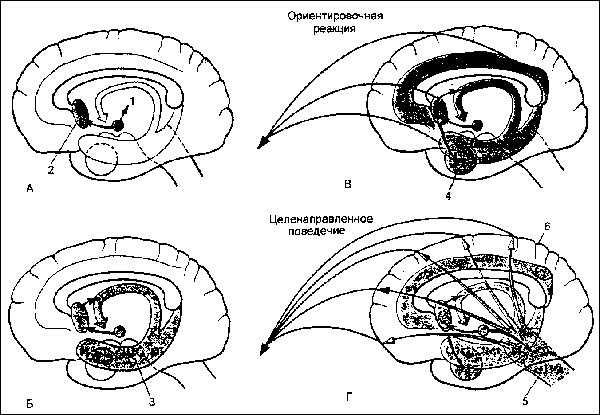
Рис. Динамика распространения возбуждения из мотивациогенного центра гипоталамуса (I) при нарастании силы его раздражения. А — возбуждение локализовано в мотивациогенном центре (1) и перегородке (2); Б, В, Г — возбуждение последовательно распространяется в гиппокамп (3), амигдалоидную область (4), ретикулярную формацию ствола мозга (5) и выходит на кору (6), формируя целенаправленное поведение.
При слабых раздражениях латерального гипоталамуса у накормленных животных сначала наблюдается активация электрической активности лимбических образований мозга — ядер перегородки, гиппокампа, миндалины. При этом медленная высокоамплитудная электрическая активность исходного состояния сменяется либо высокочастотной низкоамплитудной активностью (реакция десинхронизации), либо появлением упорядоченной тета-активности. При увеличении раздражения указанных центров наблюдаются регионарные изменения электрической активности преимущественно передних отделов коры большого мозга. Наконец, при еще более сильном раздражении латеральных отделов гипоталамуса, так же как и при длительном голодании, за счет восходящих активирующих влияний ретикулярной формации среднего мозга наблюдается генерализованная активация ЭЭГ всей коры большого мозга.
Характерно, что при распространении восходящих активирующих влияний гипоталамических мотивациогенных центров на лимбические структуры мозга у животных проявляются только ориентировочно-исследовательские реакции. При распространении этих влияний на кору мозга животные проявляют целенаправленные мотивационные реакции — употребляют находяшуюся перед ними пищу, осуществляют оборонительные реакции etc.
Таким образом, в основе биологических мотиваций любого качества лежат восходящие активирующие влияния специфических гипоталамических центров на кору большого мозга. Эти влияния устраняются после удовлетворения потребности. Именно эти восходящие активирующие влияния гипоталамических центров, обусловленные той или иной внутренней потребностью, составляют энергетический компонент мотивационного состояния.
МОТИВАЦИЯ КАК ОСОБОЕ СОСТОЯНИЕ МОЗГАВзаимоотношения подкорковых образований и коры головного мозга при мотивациях более сложные. Различные отделы коры и другие структуры мозга в свою очередь оказывают нисходящие влияния на инициативные мотивациогенные центры гипоталамуса. Кроме этого, между корой и подкорковыми образованиями постоянно циркулируют реверберирующие возбуждения.
Все это указывает на то, что мотивационное состояние представляет собой качественно особое, интегрированное состояние мозга, при котором на основе восходящих активирующих влияний гипоталамических центров каждая структура мозга вносит свои активирующие или тормозные влияния и тем самым создает специфическую для каждой мотивации интеграцию, приводящую в каждом случае к специфической форме целенаправленной деятельности.
Доминирующая мотивация представляет собой исходное, качественно особое состояние организма, которое определяет его целенаправленную деятельность и характер реагирования на действие многочисленных раздражителей внешней среды.
Мотивационное состояние можно рассматривать как активный "фильтр", определяющий подчеркнутую и избирательную реактивность животных только по отношению к тем раздражителям внешней среды, которые способствуют или препятствуют удовлетворению лежащей в основе каждой мотивации потребности.
МОТИВАЦИИ И ЭМОЦИИКаждая мотивация субъективно переживается, т. е. сопровождается специфической эмоциональной реакцией. Эмоциональные ощушения каждой мотивации подчеркнуто специфичны.
Биологический смысл субъективного переживания мотиваций заключается прежде всего в оценке индивидуумом лежаших в их основе потребностей. Ни одно живое сушество никогда не спутает субъективное ощущение голода с субъективным переживанием страха или полового возбуждения.
Субъективные переживания метаболических потребностей всегда носят неприятный (отрицательный) характер. Биологический смысл отрицательных эмоций заключается в стимулировании поиска потребных веществ.
Субъективное переживание, сопровождающее мотивацию, имеет важный информационный смысл, позволяя животным быстро и надежно, без анализа деталей оценивать каждую потребность.
Следует, однако, иметь в виду, что отрицательные эмоциональные ощущения в чистом виде сопровождают потребность только на ранних этапах онтогенетического развития и, возможно, только при первом возникновении соответствующей потребности. По мере неоднократного удовлетворения потребности, что как раз и имеет место в жизни каждого индивидуума, эмоциональное переживание потребности несколько изменяет свой знак. Согласно биологической теории эмоций П. К. Анохина, удовлетворение потребности всегда сопровождается эмоцией положительного знака, которая как бы санкционирует успех поисковой деятельности. По мере неоднократного удовлетворения однотипных потребностей и получения положительных эмоциональных ощущений каждое животное запоминает эти ощущения, и они по опережающему принципу начинают включаться в структуру мотивационного возбуждения. Формируется так называемый аппетит, или предвидение, субъектом, испытывающим ту или иную потребность, положительной эмоции, которая может быть получена при удовлетворении этой потребности.
Аппетит является мощным стимулом целенаправленной деятельности. Предвидение положительной эмоции заставляет животных порой даже более активно, чем на основе только отрицательных эмоциональных ощущений, преодолевать препятствия для удовлетворения ведущей потребности. Кроме того, аппетит определяет избирательную направленность мотивации. Эта избирательность влечения к определенным раздражителям обусловливается, с одной стороны, избирательностью метаболической потребности. Например, в случае осмотической потребности она может быть обусловлена недостатком солей или воды или различных их комбинаций, что приводит к поиску недостающих веществ. С другой стороны, избирательность аппетита обусловлена в значительной степени предвидением определенных свойств раздражителей, наиболее часто удовлетворяющих соответствующую потребность, т. е. определенными привычками. Особенно ярко демонстрируют это свойство аппетита национальные привычки к определенному виду пищи.
Итак, эмоции являются субъективным переживанием потребности и ее удовлетворения, средством оценки потребности и активации мотивационного поведения. Аппетит как субъективное предвидение положительной эмоции подкрепления определяет избирательность мотивационного поведения.
СВОЙСТВА МОТИВАЦИОННОГО СОСТОЯНИЯХИМИЧЕСКАЯ СПЕЦИФИКА
Мотивационные возбуждения различного биологического качества, например голод, жажда или страх, характеризуются одинаковыми изменениями электрической активности мозговых структур типа ЭЭГ-активации. Однако, несмотря на однотипность электрических характеристик, каждая мотивация обладает подчеркнутой спецификой. Эта специфика, как указывалось выше, проявляется прежде всего в специфике эмоциональных ощущений, сопровождающих каждую мотивацию. Кроме того, специфика мотивации заключается в особой корково-подкорковой интеграции возбуждений, при которой в каждую мотивацию в разной степени вносят вклад различные структуры мозга.
Специфика мотивационного возбуждения проявляется также и в особенностях химических механизмов различных мотиваций. Оборонительная мотивация страха у животных, например, избирательно блокируется альфа-адреноблокатором аминазином (В.А. Гавличек, А.И. Шумилина), а пищевые мотивации голодных животных — холинолитиками — атропином или амизилом. Подтверждением этого служит следующий эксперимент.
Кролика кормят в определенной комнате, предлагая предпочитаемую им пищу: капусту и морковь. В результате неоднократных кормлений кролик охотно идет в экспериментальную комнату и проявляет в ней выраженную поведенческую пищевую активность. Картина совершенно изменяется после того, как кролику в комнате, где он предварительно получал пищу, было нанесено несколько электрокожных раздражений. После этого кролик перестает дотрагиваться до еды, несмотря на то что голоден, сидит постоянно в углу, сжавшись в комок. У кролика доминирует мотивация страха, которая полностью подавляет пищевую мотивацию. Однако после внутривенного введения кролику аминазина из расчета 0,5 мг/кг животное немедленно утрачивает мотивацию страха и устремляется к пище, которую тут же начинает активно поедать. Объяснение этого эксперимента очевидно и заключается в том, что аминазин избирательно блокирует у кроликов мотивацию страха и высвобождает ранее заторможенную страхом мотивацию голода. Следовательно, опыт указывает на то, что мотивации голода и страха строятся на различных химических механизмах.
Химическая избирательность пищевой и оборонительной мотивации проявляется и в следующем эксперименте.
Кошку после 2-суточной пищевой депривации погружают в уретановый наркоз. На ЭЭГ наблюдается избирательная активация передних отделов коры мозга, в то время как в теменно-затылочных отделах мозга регистрируется медленная высокоамплитудная активность, характерная для состояния наркотического сна. При нанесении голодным животным, находящимся под уретановым наркозом, электрического раздражения седалищного нерва наблюдается генерализованная ЭЭГ-активация всех отделов коры мозга. Введение аминазина блокирует только эту генерализованную активацию, вызванную ноцицептивным раздражением, и не действует на "голодную" активацию передних отделов коры мозга. "Голодная" активация исчезает только после введения холинолитиков — амизила или атропина.
Приведенные опыты указывают на специфику химических механизмов, лежащих в основе оборонительной и пищевой мотивации животных.
Фармакологические вещества позволяют осуществлять своеобразную "химическую препаровку" различных мотивационных состояний животных, избирательно блокируя множественные восходящие активирующие влияния подкорковых образований на кору мозга, каждое из которых определяет различное мотивационное состояние.
Механизмы химической специфики мотивационных возбуждений различного биологического качества сложнее.
Не у всех голодных животных холинолитики блокируют пищевую мотивацию; мотивация страха устраняется адренолитиками не у всех животных. Как пишевую мотивацию голодных кроликов, вызванную электрическим раздражением через вживленный электрод "центра голода" латерального гипоталамуса, так и оборонительную мотивацию, вызванную электрическим раздражением вентромедиального гипоталамуса, объединяют адренергические, холинергические и дофаминергические химические механизмы (В.Г. Зилов). При разных мотивациях указанные нейромедиаторы вовлекаются в различных комбинациях, локализуясь в различных структурах мозга. Иными словами, химическая специфика мотивационных возбуждений определяется специфической химической интеграцией различных физиологически активных веществ различных мозговых структур, объединенных в то или иное мотивационное состояние.
КОРКОВО-ПОДКОРКОВАЯ ИНТЕГРАЦИЯ В СТРУКТУРЕ МОТИВАЦИОННОГО ВОЗБУЖДЕНИЯКак указывалось выше, биологические мотивации строятся на основе восходящих активирующих влияний мотивациогенных структур гипоталамуса на кору большого мозга. В морфофункциональную архитектуру этих активирующих влияний избирательно включаются различные отделы мозга.
Избирательность активации корково-подкорковых структур наблюдается у голодных животных, находящихся под уретановым наркозом. В этих условиях наряду с активацией передних отделов коры мозга отмечается активация структур таламуса, ретикулярной формации и гипоталамуса. Таким образом, пищевое мотивационное возбуждение объединяет специфический комплекс избирательно взаимосвязанных корково-подкорковых образований с общими нейрохимическими свойствами, не блокируемый уретаном. Как указывалось выше, корково-подкорковая интеграция мотивационного состояния наряду со специфическими восходящими активирующими влияниями гипоталамических структур на другие отделы мозга, в том числе кору, включает нисходящие специфические влияния коры и других отделов мозга на инициативные центры гипоталамуса.
Так, например, в пищевом мотивационном возбуждении передние отделы коры мозга и дорсальный гиппокамп при их электрическом раздражении оказывают тормозное влияние на порог возбудимости "центра голода" латерального гипоталамуса. Затылочные отделы коры мозга при их электрическом раздражении, наоборот, снижают порог возбудимости пищевой реакции при раздражении латерального гипоталамуса. При оборонительной мотивации, наоборот, передние отделы коры мозга и дорсальный гиппокамп при электрическом раздражении оказывают облегчающие, а затылочные отделы — тормозные влияния.
Таким образом, каждое мотивационное состояние представляет собой специфический интегрированный комплекс избирательно объединенных корково-подкорковых образований, каждое из которых вносит свой особый вклад в формирование доминирующей мотивации.
ПЕЙСМЕКЕРНАЯ РОЛЬ ГИПОТАЛАМИЧЕСКИХ ЦЕНТРОВ В СТРУКТУРЕ ДОМИНИРУЮЩЕЙ МОТИВАЦИИСтруктуры мозга, избирательно вовлеченные в биологическое мотивационное состояние, обладают различными свойствами.
При раздражении или разрушении различных отделов коры или лимбических структур мозга биологические мотивации, как правило, приобретают измененный характер, проявляясь в ослабленной или усиленной форме. Например, при раздражении различных отделов амигдалоидного комплекса у животных может наблюдаться как афагия, так и гиперфагия. Точно такие же реакции наблюдаются при разрушениях различных отделов коры и других лимбических образований. В то же время разрушение гипоталамических центров полностью элиминирует биологические мотивации. Животное с двусторонне разрушенными латеральными отделами гипоталамуса может, например, лежать в окружении самой лакомой пищи и, не притронувшись к ней, погибнуть от истощения.
Таким образом, в структуре мотивационного возбуждения гипоталамическим центрам принадлежит особая, ведушая роль. Электрофизиологические исследования подтверждают это. Если кошкам, находяшимся под уретановым наркозом, после 2-суточной пищевой депривации разрушить, например, переднемедиальные структуры таламуса, то "голодная" активация исчезает только в соответствующих отделах коры большого мозга, но сохраняется в латеральных отделах гипоталамуса и в ретикулярной формации среднего мозга. В то же время двустороннее разрушение латерального гипоталамуса устраняет "голодную" активацию во всех отделах мозга.
Гипоталамические центры в структуре "голодного" мотивационного возбуждения гораздо чувствительнее к химическим веществам по сравнению с корой большого мозга.
Аппликация 0,01% раствора атропина на передние отделы коры мозга у голодных кошек локально устраняет "голодную" активацию только в коре мозга. В то же время инъекция 0,0001% раствора атропина через специальную иглу непосредственно в латеральный гипоталамус устраняет "голодную" активацию во всех структурах мозга (А.А. Панфилов).
Механизм формирования доминирующей биологической мотивации у животных очень напоминает возникновение возбуждения в синусном узле сердечной мышцы, где располагается специальный водитель — задаватель ритма сердечных сокрашений — пейсмекер.
Аналогично процессам распространения возбуждений по сердечной мышце возбуждение, первично возникающее в мотивациогенных центрах гипоталамуса, широко генерализуется в восходящем направлении вплоть до коры большого мозга.
В синусном узле сердца возбуждение возникает ритмически. Аналогичная картина имеет место и в мотивациогенных центрах гипоталамуса. Возбуждение в них в естественных условиях также возникает периодически, по "триггерному типу" по мере нарастания той или иной потребности до критического уровня. Оно сохраняется, пока существует эта потребность, и исчезает после ее устранения.
Пейсмекер сердца имеет повышенную по сравнению с другими образованиями сердца возбудимость к специфическим гуморальным или другим раздражителям. Точно так же гипоталамические структуры по сравнению с другими структурами мозга, вовлеченными в мотивационное возбуждение, обладают повышенной возбудимостью к электрическим и химическим раздражителям.
По отношению к гипоталамическим мотивациогенным образованиям, так же как и в сердечной мышце, другие структуры мозга выстраиваются по определенному градиенту возбудимости до коры большого мозга включительно.
Синусный узел сердца, как известно, держит в определенном подчинении другие центры автоматизма, которые обладают более низкой возбудимостью. Точно так же мотивациогенные центры гипоталамуса держат в морфологической и функциональной зависимости структуры других уровней мозга, Выключение гипоталамических центров приводит к распаду всей системы объединенных в мотивационное возбуждение элементов.
Все вышеизложенное составило концепцию о пейсмекерной роли гипоталамических центров в формировании основных биологических мотиваций (П. К. Анохин, К. В. Судаков). Согласно этой концепции, гипоталамическим центрам принадлежит ведущая, пейсмекерная роль в организации всей центральной архитектоники доминирующей мотивации.
Пейсмекерная роль гипоталамических центров в формировании биологических мотиваций определяет врачебную тактику влияния на них с помощью фармакологических препаратов. Согласно этим представлениям, фармакологические вещества в первую очередь и в меньших дозах действуют на пейсмекеры мотивационных состояний, обладающие наиболее напряженным метаболизмом. Тем самым разрушается вся корково-подкорковая интеграция соответствующей мотивации.
МОЛЕКУЛЯРНАЯ ИНТЕГРАЦИЯ МОТИВАЦИОННОГО ВОЗБУЖДЕНИЯУстановлено, что в структуре мотивационного возбуждения принимают активное участие различные олигопептиды — белковые молекулы с небольшим набором аминокислот. Показано (А. В. Котов, С. М. Толпыго), что введение в желудочки мозга голодным кроликам бета-липотропина тормозит их пищевые реакции. Этот же олигопептид активирует пищевые реакции накормленных животных. Особенно активны в формировании пищевых мотиваций животных пептиды пищеварительного тракта. Внутривенное введение накормленным животным пентагастрина вызывает у них появление выраженных пищевых мотиваций. Холецистокинин, наоборот, тормозит пищевые мотивации животных. Установлено также отчетливое влияние ангиотензина II, брадикинина и вазопрессина на реакции самораздражения и избегания у кроликов.
Участие многих олигопептидов в формировании мотиваций различного биологического качества свидетельствует о молекулярной интеграции мотивационного состояния. Каждая мотивация организует специфическое объединение белковых молекул на разных уровнях мозга.
МЕХАНИЗМЫ ТРАНСФОРМАЦИИ МОТИВАЦИИ В ЦЕЛЕНАПРАВЛЕННОЕ ПОВЕДЕНИЕНа уровне гипоталамических мотивационных центров, как указывалось выше, происходят два ответственных физиологических процесса: 1) трансформация нейрогуморальной сигнализации о метаболической потребности в процесс избирательного возбуждения структур мозга; 2) процессы энергетической активации структур мозга как основы формирования соответствующего поведения.
Мотивационное возбуждение, возникшее первично в структурах гипоталамуса на основе его восходящих активирующих влияний, распространяется до коры большого мозга, где происходит другой, не менее ответственный процесс трансформации мотивационного возбуждения, вызванного метаболическими потребностями, в механизм целенаправленного поведения. Тем не менее всегда следует иметь в виду, что как трансформация метаболической потребности в мотивационное состояние, так и трансформация доминирующей мотивации в поведение осуществляются без потери информационной значимости исходной потребности.
Доминирующая мотивация существенно изменяет свойства нейронов коры головного мозга. При этом повышается их чувствительность к различным раздражителям сенсорной и биологической модальности, увеличиваются конвергентные свойства нейронов, повышается их функциональная мобильность, изменяется чувствительность к нейромедиаторам и нейропептидам.
Доминирующая мотивация специфически отражается в рисунке межимпульсных интервалов у отдельных нейронов различных областей мозга. У голодных кроликов доминируют межимпульсные интервалы 10; 50 мс и иногда более 1000 мс, у воднодепривированных кроликов — 25 и 150 мс, у кроликов, находящихся в оборонительном состоянии, — 40 и 150 мс. После устранения доминирующей мотивации в деятельности этих нейронов выражен только один доминирующий интервал — 40—60 мс.
Доминирующая мотивация значительно повышает чувствительность соответствующих периферических рецепторов. При мотивации голода, например, возрастает чувствительность вкусовых рецепторов ротовой полости, при агрессивных мотивациях — рецепторов вокруг ротовой полости в тригеминальной области, при половых мотивациях избирательно активируются рецепторы половых органов. Все это расширяет взаимодействие субъектов, испытывающих ту или иную метаболическую потребность, с внешним миром и способствует более успешному удовлетворению исходных потребностей.
Указанные процессы в конечном счете определяют формирование на основе доминирующей мотивации целенаправленного поведения.
МОТИВАЦИЯ И ПАМЯТЬМотивации тесно взаимодействуют с механизмами памяти. Мотивации, формирующиеся под воздействием внутренних метаболических потребностей и факторов окружаюшей среды, обладают выраженной способностью по опережающему принципу извлекать из памяти генетический и индивидуальный опыт субъектов по удовлетворению лежащей в их основе доминирующей потребности.
Восходящие активирующие влияния, сформированные различными мотивациями, распространяются в протоплазме отдельных нейронов мозга до механизмов синтеза специальных белковых молекул с участием генетического аппарата (см. рис.). Механизмы синтеза белковых молекул определяются предшествующими подкреплениями, т. е. удовлетворением соответствующих потребностей. Экспрессируемые под влиянием доминирующей мотиваиии белковые молекулы в свою очередь участвуют в формировании соответствующего поведения. Синтез этих белковых молекул подавляется при введении блокаторов синтеза белка и вследствие этого мотивация утрачивает способность трансформироваться в поведение. Так, блокатор синтеза белка циклогексимид на несколько десятков часов подавляет пищевую мотивацию у кроликов при электрическом раздражении центра голода латерального гипоталамуса. Циклогексимид также блокирует у животных выработанную реакцию самораздражения. После введения циклогексимида сначала облегчается, а через несколько часов подавляется оборонительная реакция кроликов при электрическом раздражении центра страха вентро-медиального гипоталамуса.
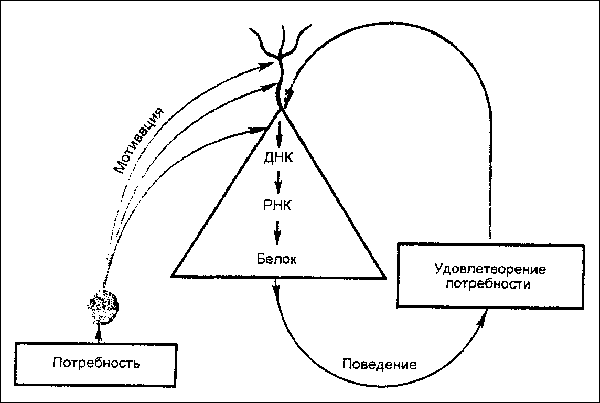
Рис. Молекулярные изменения в генетическом аппарате нейрона коры под влиянием восходящих активируюших мотивационных возбуждений. Повторное удовлетворение потребности инициирует синтез в теле нейрона и белка.
На фоне действия циклогексимида пентагастрин, введенный в боковые желудочки мозга, восстанавливает пищевую мотивацию при раздражении латерального гипоталамуса. Другой олигопептид — АКТГ4-10 — восстанавливает самораздражение. Брадикинин восстанавливает заблокированную циклогексимидом оборонительную реакцию.
Пищевую реакцию у кроликов при электрическом раздражении латерального гипоталамуса блокируют также иммуноглобулины к гастрину.
Имеется множество физиологических экспериментов, которые показывают, что доминирующая мотивация извлекает у животных опыт, ранее накопленный по удовлетворению соответствующих потребностей, по опережающему принципу до конечного подкрепляющего результата включительно и, таким образом, как бы "вытягивает" весь предшествующий опыт животного по удовлетворению соответствующей потребности.
В опытах кроликов предварительно перед получением пищи обучали совершать специальные ритуальные движения — поворот на 360°. В процессе выработки пищедобывателъных навыков каждый такой поворот подкреплялся порцией пищи. В результате уже к 30—40-му сочетанию животные демонстрировали прочно выработанный пишедобывающий навык: при помещении в экспериментальную камеру они совершали повороты на 360°, которые во всех случаях подкреплялись пищей. После того как у всех животных был упрочен пищедобывательный навык предварительного поворота, каждому из них в область латерального гипоталамуса вживляли биполярные электроды. Кончик электрода фиксировали в той точке латерального гипоталамуса, раздражение которой у накормленных животных вызывало отчетливо выраженную реакцию дополнительного приема находящейся перед ними пищи.
Вопрос заключается в следуюшем: способно ли искусственное раздражение мотивациогенного центра голода латерального гипоталамуса выявить не только пищевую реакцию приема пищи, но и то ритуальное движение на 360°, которому каждое из подопытных животных было обучено в предварительных экспериментах?
Опыты были проведены на кроликах в условиях той же экспериментальной камеры, в которой они предварительно обучались ритуальным пищедобывательным движениям.
Эксперименты показали следующее. Все животные, накормленные перед опытом, в экспериментальной клетке демонстрировали полное успокоение. Однако вслед за стимуляцией латерального гипоталамуса они оживлялись и совершали ритуальное движение — поворот на 360°, после чего устремлялись к кормушке и поедали находящуюся в ней порцию пищи. Опыты свидетельствуют о том, что доминирующая пищевая мотивация, вызванная у накормленных животных искусственным раздражением латерального гипоталамуса, привела к мобилизации всего того опыта, который в данной обстановке в предшествующем обучении предварял пищевое подкрепление. Это положение подтверждается опытом противоположного смысла. У животных другой группы с выработанным аналогичным ритуальным движением производили двустороннюю коагуляцию пищевых центров латерального гипоталамуса. Оказалось, что такая операция полностью элиминировала в экспериментальной камере как ритуальное движение, так и прием пиши. Опыты в этой серии с очевидностью указывают на ведущее значение доминирующей мотивации в извлечении навыка из памяти, причем извлечение опыта из памяти происходит до конечного результата включительно.
Мотивационное возбуждение выступает, таким образом, в роли ведущего фактора, формирующего предвидение животными конечного удовлетворяюшего исходную потребность результата.
Кроме извлечения опыта из памяти, доминируюшая мотивация определяет и быстроту фиксации промежуточных и конечных результатов действия по удовлетворению соответствующих потребностей.
В экспериментах Т. Н. Ониани с сотр. был проведен следуюший опыт. Голодных кошек помешали в специальную камеру, разделенную на два отсека. В первом отсеке животные не получали пищу, а должны были зафиксировать местоположение светового сигнала справа или слева, что соответствовало расположению кормушек с пищей во втором отсеке камеры. Вход во второй отсек преграждала дверь. Опыт проходил таким образом, что вслед за подачей правого или левого светового сигнала через определенный промежуток времени открывалась дверца во второй отсек и животноё получало пищу в кормушке, находяшейся на стороне сигнала. Путем тренировки у животных вырабатывали четкую линию поведения. При включении правой лампочки кошки при открывании двери во второй отсек устремлялись к правой кормушке, при зажигании левой лампочки — к левой кормушке. После этого замысел эксперимента усложнялся. Между подачей светового сигнала и открытием дверцы изменяли интервалы времени. Определялось максимальное время отсроченных реакций, при котором животные удерживали след сигнала и не совершали ошибок в выборе стороны расположения кормушки во втором отсеке. Оказалось, что в голодном состоянии при наличии выраженной пищевой мотивации время отсроченных реакций могло быть значительным. Однако оно существенно сокращалось по мере насыщения животных, удовлетворения их ведущей потребности и снижения доминирующей мотивации. Эксперимент отчетливо демонстрирует значение мотивацил для удержания следов памяти.
Итак, в системной организации целенаправленных поведенческих актов мотивации выступают как ведущий фактор фиксации опыта в памяти, способности его удержания при наличии препятствий к достижению цели и, наконец, обусловливают быстрое извлечение накопленного опыта из памяти.
Из этого следует важная практическая рекомендация: запоминание всегда успешно при наличии выраженной мотивации. Лучше запоминается тот предмет, который представляет для обучающегося наиболее значимый интерес.
С физиологической точки зрения, процессы фиксации доминирующей мотивации опыта по удовлетворению соответствующей потребности изучены недостаточно.
НАПРАВЛЯЮЩИЙ КОМПОНЕНТ ДОМИНИРУЮЩЕЙ МОТИВАЦИИПрактически все основные биологические мотивации (пищевые, питьевые, половые, оборонительные, агрессивные etc.) генетически программируют ведущие свойства соответствующих подкрепляющих раздражителей. Только благодаря этому новорожденные животные проявляют столь поразительное избирательное отношение к биологически значимым раздражителям окружаюшей среды. Программирование доминирующей мотивацией свойств потребных результатов совершенствуется в процессах индивидуального обучения. Опережающее программирование свойств подкрепляющих раздражителей осуществляется в аппарате акцептора результата действия. Программирование свойств потребного результата в системной организации поведенческих актов составляет направляющий компонент (вектор) доминирующей мотивации. С этим формируюшимся по опережающему принципу направляюшим компонентом доминирующей мотивации постоянно происходит сравнение параметров реально достигнутых результатов. Этот механизм в конечном счете и направляет животных к полноценному удовлетворению их ведущих потребностей и позволяет им оценивать и исправлять ошибки поведенческой деятельности.
МОТИВАЦИИ И ПОДКРЕПЛЕНИЕВ системной организации поведения доминирующая мотивация и подкрепление тесно взаимодействуют, причем часто на одних и тех же нейронах мозга. Подкрепляющее возбуждение изменяет активность нейронов, вовлеченных в исходную мотивацию. При этом пачкообразная активность этих нейронов сменяется упорядоченной, регулярной (см. рис.).
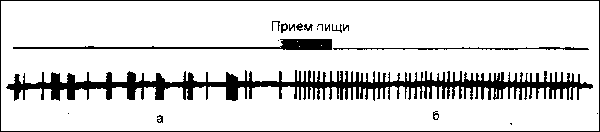
Рис. Смена пачкообразной активности нейрона, вовлеченного в доминирующую мотивацию голода (а), на регулярную после приема пищи (б).
Реакции других нейронов мозга, не вовлеченных в доминирующую мотивацию, на подкрепляющее возбуждение могут быть самыми разнообразными — в виде учащения или торможения исходной активности.
Электрическое раздражение мотивациогенных центров гипоталамуса приводит к тому, что отдельные клетки коры и подкорковых образований, ранее не реагировавшие на подкрепляющие воздействия, начинают на них отчетливо реагировать. Двусторонняя коагуляция или анодическая поляризация пейсмекеров мотивационного возбуждения, наоборот, приводит к тому, что клетки различных областей мозга, ранее отвечавшие на подкрепляющее воздействие, перестают на него реагировать. Следовательно, мотивационное доминирующее возбуждение как бы "настраивает" нейроны различных областей мозга на подкрепляющее воздействие. В то же время подкрепленке изменяет также реакции нейронов мозга на мотивационные воздействия.
Взаимодействие мотивационных и подкрепляющих возбуждений на отдельных нейронах мозга строится по комплементарному принципу. На них существенное влияние оказывают иммуномодуляторы и нейропептиды.
Доминирующее мотивационное возбуждение, формирующееся на основе той или иной потребности как организационный специфический корково-подкорковый комплекс, представляет собой функциональную канву избирательно возбужденных синаптических и нейрональных образований мозга. На этой канве подкрепляющие этапные и завершающие возбуждения в определенной временной последовательности пишут своеобразный узор, или энграмму. Этот узор по мере неоднократных подкреплений отшлифовывается в форме специфической корково-подкорковой архитектуры, в которой синаптические и нейрональные элементы объединены в пространственно-временных соотношениях.
При каждом очередном возникновении соответствующей потребности доминируюшее мотивационное состояние активирует элементы выработанной на основе предшествующего опыта энграммы, возбуждая их до конечного пункта, связанного с получением необходимой информации об удовлетворении соответствующей потребности. Этот комплекс избирательно возбужденных корково-подкорковых аппаратов, представляющий нейрофизиологическую архитектуру акцептора результатов действия, и направляет поведение живого существа через постоянное сравнение поступающей к нему с периферии обратной афферентации, вызванной действием раздражителей внешней среды, к достижению цели, т. е. к удовлетворению доминирующей на каждый данный момент времени потребности.
БИОЛОГИЧЕСКИЕ МОТИВАЦИИ В ФОРМИРОВАНИИ ЛИЧНОСТИВыраженная мотивация человека обусловливает целеустремленность личности, ее способность активно действовать для достижения целей в соответствии с юридическими, правовыми и моральными законодательствами общества.
Врожденные биологические мотивации также участвуют в формировании личности, определяя ее индивидуальные и общественные интересы, а также черты характера уже в раннем детстве.
По доминированию пищевых, агрессивных, оборонителъных, половых и других биологических мотиваций выявляются индивидуальные характеры.
Тем не менее, несмотря на важное значение биологических мотиваций, ведущее значение в формировании личности человека, его характерологических особенностей принадлежит социальным мотивациям, формирующимся под влиянием окружающей, в частности социальной, среды. Вопросы формирования личности человека широко освещаются в соответствуюшей психологической литературе.
ПАТОЛОГИЧЕСКИЕ МОТИВАЦИИК патологическим относятся такие искусственно создаваемые влечения, как наркомании, алкоголизм и курение. Имеются и другие патологические влечения, которые рассматриваются в курсе психиатрии.
В случае употребления алкоголя и наркотиков в гипоталамических структурах мозга на основе изменения метаболических реакций формируются искусственные пейсмекеры, создаюшие в отсутствие наркотиков активное возбуждение структур мозга, приводящее к выраженному влечению к их употреблению (см. рис.).
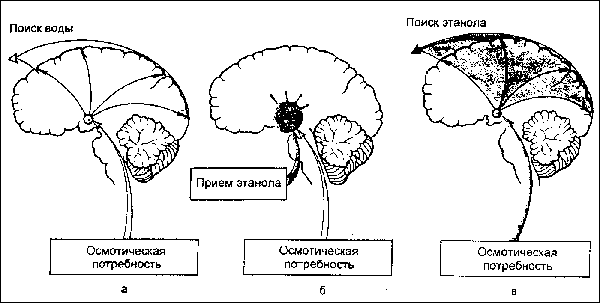
Рис. Динамика формирования алкогольной мотивации на основе биологической мотивации жажды. а — осмотическая потребность приводит к формированию мотивации жажды, к поиску и приему воды; б — под влиянием приема этанола изменяются свойства "центров жажды" гипоталамуса: они прекращают оказывать активирующие влияния на кору; в — осмотическая потребность формирует патологическую алкогольную мотивацию.
Расстройства мотиваций проявляются в их усилении (булимия, гиперсексуальность, полидипсия) или значительном подавлении (афагия, адипсия, импотенция). В случаях избирательного расстройства какой-либо одной, преимущественно врожденной, мотивации нередко происходит формирование психопатической личности.
Представление о формировании мотиваций по пейсмекерному принципу определяет возможность избирательно направленного воздействия на патологические их формы. Фармакологические и другие виды воздействий должны быть направлены прежде всего на метаболизм пейсмекеров, имеющих, как указывалось выше, повышенную чувствительность по сравнению с другими элементами мотивационного возбуждения. Именно путем таких направленных воздействий на самое уязвимое звено патологической мотивации можно разрушить всю систему мотивационного возбуждения, включая связанные с ней формы патологических поведенческих реакций и патологические навыки.