Горизонтальные клетки
РедактироватьГоризонтальные клетки служат для организации взаимно-тормозного влияния между биполярными клетками, что обеспечивает латеральное торможение и контрастирование активностей биполяров. Кроме этого они имеют обратную положительную и отрицательную связь с фоторецепторами, еще больше усиливая эффект контрастирования. Между собой они связаны электрическими синапсами, что позволяет передавать друг другу возбуждение. Это увеличивает область активации и распространения латерального торможения.
Несмотря на большое разнообразие видов горизонтальных клеток у разных животных, их основное назначение - контрастирование активностей биполяров.
Вес уверенности: Вполне уверенно подтверждается независимыми исследователями
Дендриты горизонтальных клеток образуют контакты с фоторецепторами. От периферических ветвей дендритов берут начало аксоноподобные отростки, создающие тормозные контакты с биполярными нейронами. Функция горизонтальных клеток — торможение биполярных нейронов кнаружи от непосредственной области возбуждения. На выходе горизонтальные клетки всегда тормозные. Эта латеральная связь обеспечивает феномен латерального торможения, характерный для всех других сенсорных систем и важный для повышения надежности проведения визуального образа с соответствующим зрительным контрастом.
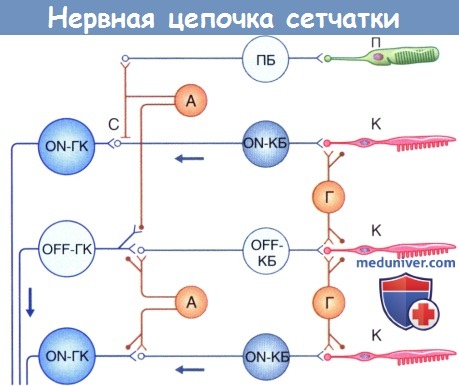
Схема нервной цепочки сетчатки: А—амакриновая клетка; К—колбочка; КБ—колбочковый биполярный нейрон; ГК—ганглиозная клетка; Г—горизонтальная клетка; С—соединение (щелевидный контакт); П—палочка; ПБ—палочковый биполярный нейрон.
Схема на рисунке является упрощенной, так как в реальности у разных видов позвоночных существует несколько видов горизонтальных клеток с более сложной схемой связей. Как интернейроны в других отделах нервной системы, горизонтальные клетки воспринимают сигналы от одних элементов — фоторецепторов и передают их различным структурам сетчатки — другим горизонтальным клеткам, обратно фоторецепторам, биполярным клеткам, опосредованно — ганглиозным клеткам.
Обнаружены так же значительные физиологические отличия горизонтальных клеток у разных видов животных. Например в сетчатке рыб найдено несколько типов горизонтальных клеток, раздельно воспринимающих палочковые и колбочковые сигналы [Parthe, 1972; Stell, Witkovsky, 1973]. В сетчатке черепахи (Chelydra serpentina) утолщенные аксонные терминали горизонтальных клеток первого слоя имеют как колбочковые, так и палочковые входы [Copenhagen, Ashmore, Schnapf, 1983]. В сетчатке кошки один и тот же нейрон воспринимает как палочковые, так и колбочковые сигналы [Steinberg, 1969; Niemeyer, Gouras, 1973; Kelson et al., 1975].
Существенно, что у различных позвоночных нет единообразия в структуре и числе типов горизонтальных клеток [Kolb. Mariani, Gallego, 1980; Gallego, 1982]. Kolb [1974], обобщая данные о структурных особенностях горизонтальных клеток у различных позвоночных, справедливо напомнила об отсутствии этих нейронов в сетчатке некоторых разновидностей глубоководных рыб, по данным Locket [1971]. Следовательно, наряду с другими нейронами сетчатки горизонтальные клетки весьма разнообразны, что, как можно думать, отражает особенности в развитии зрительной функции в зависимости от окружающей среды.
Обратная связь
Амакриновые и горизонтальные клетки секретируют такие медиаторы, как ГАМК, глицин, дофамин, ацетилхолин и индоламин, оказывающие тормозные эффекты , наиболее важным из которых является ГАМК. И амакриновые, и горизонтальные клетки используют этот медиатор для тормозного воздействия на биполярные клетки.
Секреция ГАМК не является единственным способом с помощью которого горизонтальные клетки влияют на фоторецепторы. Колбочки и горизонтальные клетки имеют реципрокное соединение. То есть согласованно и противоположно направлено, или в даном случае - взаимно-тормозящее. Такое соединение также лежит в основе латерального торможения и установления центрально-периферического антагонизма в сетчатке органа зрения. Колбочки передают на горизонтальные клетки возбуждающий сигнал и горизонтальные клетки отвечают на это тормозным сигналом. Для того, чтобы горизонтальные клетки "понимали" фоторецепторы, они обладают чувствительным к глутамату AMPA-рецептором. Реципрокное соединение осуществляется через 2 типа синаптических связей:
Положительный синапс
Горизонтальные клетки передают колбочкам сигнал, который приводит к повышению внутриклеточного уровня кальция, что вызывает ускорение секреции нейротрансмиттера (глутамата).
Отрицательный синапс
Были выдвинуты три гипотезы, призванные объяснить механизмы функционирования этих инвертирующих сигналы синапсов; а именно, каким образом деполяризация горизонтальных клеток подавляет высвобождение нейромедиаторов из колбочек.
- горизонтальные клетки секретируют ГАМК, гиперполяризуя мембраны колбочек.
- гипотеза связана с эфапсами (эфаптическими механизмами): она предполагает, что электрические токи через каналы в дендритах горизонтальных клеток локально изменяют трансмембранный потенциал на терминалях колбочек. Предполагается, что с помощью эфапсов опосредуется отрицательная обратная связь и модулируется мощностью сигнала колбочек.
- гипотеза связывает описанные эффекты с эффлюксом протонов, возникающим при деполяризации горизонтальных клеток, при котором происходит закисление межклеточной среды, что ингибирует мембранные потенциал-зависимые кальциевые каналы в колбочках.
На сегодняшний момент пока нет устоявшегося понимания, какой из трех механизмов играет наибольшую роль в проведении отрицательной обратной связи к колбочкам.
Межклеточная связь
Горизонтальные клетки соединены между собой электрическими синапсами [axiom_id=630] и образуют функциональный синцитий. Возбуждение одной горизонтальной клетки распространяется на другие. В центральной ямке сетчатки приматов одна горизонтальная клетка соединяется примерно с 6-ю колбочками, а на периферии — с 30-40. Дендритное поле горизонтальной клетки на периферии сетчатки достигает диаметра 250 мкм. В то же время, размер рецептивных полей горизонтальных клеток может изменяться в пределах 300мкм ÷ 5мм, что значительно превышает протяженность дендритных разветвлений. За счет этого горизонтальные клетки могут формировать периферию рецептивных полей биполярных нейронов. При освещении фоторецепторов горизонтальные клетки отвечают гиперполяризацией.
Фоторецепторы вместе с горизонтальными клетками и биполярами образуют так называемую триаду. Триада состоит из пресинаптической мембраны фоторецептора, в которую заходит дендрит биполяра, а по бокам от него заходят отростки горизонтальных клеток. Значение и принципы функционирования триады пока остаются неизвестными. За счет таких связей между фоторецепторами, горизонтальными клетками и биполярами, центр рецептивного поля биполяров формируется прямым путём при передаче сигнала от фоторецепторов, периферия формируется за счет непрямого пути через горизонтальные клетки за счёт возбуждающих или тормозных синапсов.
Источник:
Дополнительно:
В искуственной реализации сетчатки латеральное торможение может быть успешно реализовано программным способом.
Последнее редактирование: 02.11.2020
Чтобы оставить комментарии нужно авторизоваться.
Возможность управлять через латеральное торможение активностью целых областей биполяров означает по сути возможность получения конфигураций эффекторного рисунка. То есть на один раздражитель при разных уровнях торможения может быть сформирован свой уникальный ответный рисунок, а от него - рефлекторная цепочка. Это один из первичных, древних способов мгновенного реагирования. У простейших позвоночных как правило большая часть обработки зрительной информации происходит на уровне сетчатки, где и формируются конечные пусковые триггеры рефлексов.