Оглавление
Доказательства эволюции
Наблюдаемая эволюция
Эволюция кишечной палочки в лаборатории
Опыты Г. Шапошникова по искусственной эволюции
Быстрая эволюция ящериц в новых условиях
Развитие устойчивости к антибиотикам и пестицидам
Примеры видообразования
Промежуточные этапы видообразования
Белый медведь
Наблюдаемое видообразование
Яблонная муха
Эволюционное дерево
Палеонтологические доказательства
Эволюция человека
Эволюция лошади
Морфологические доказательства
Гомологичные органы
Аналогичные органы
Рудименты
Атавизмы
Несовершенство строения организмов
Эмбриологические доказательства
Молекулярно-генетические и биохимические доказательства
Биохимическое единство жизни
Вторая хромосома человека
Эндогенные ретровирусы
Биогеографические доказательства
Дарвиновы вьюрки
Примечания
Доказательства эволюции
Доказательства эволюции - научные данные и концепции, подтверждающие происхождение всех живых существ на Земле от общего предка. Благодаря этим доказательствам основы эволюционного учения получили признание в научном сообществе, а ведущей системой представлений о процессах видообразования стала синтетическая теория эволюции.
Эволюционные процессы наблюдаются как в естественных, так и лабораторных условиях. Известны случаи образования новых видов. Описаны также случаи развития новых свойств посредством случайных мутаций. Факт эволюции на внутривидовом уровне доказан экспериментально, а процессы видообразования непосредственно наблюдались в природе.
Чтобы получить сведения об эволюционной истории жизни, палеонтологи анализируют ископаемые останки организмов. Степень родства между современными видами можно установить, сравнивая их строение, геномы, развитие эмбрионов (эмбриогенез). Дополнительный источник информации об эволюции - закономерности географического распространения животных и растений, которые изучает биогеография. Все эти данные укладываются в единую картину - эволюционное дерево жизни.
Наблюдаемая эволюция
Наблюдаемые сегодня изменения в популяциях доказывают не только существование эволюции, но и существование ряда механизмов, необходимых для эволюционного происхождения всех видов от общего предка. Было установлено, что геномы подвержены разнообразным мутациям, среди которых перемещение интронов, дупликация генов, рекомбинации, транспозиции, ретровирусные вставки (горизонтальный перенос генов), замена, удаление и вставка отдельных нуклеотидов, а также хромосомные перестройки. Известны такие хромосомные перестройки как дупликация генома (полиплоидия), неравный кроссинговер, хромосомная инверсия, транслокация, деление, слияние, дупликация и удаление хромосом. Также наблюдается изменение строения организмов и функциональные изменения - различные адаптации, появление способности усваивать новый вид пищи (в том числе - нейлон и пентахлорфенол, производство которых началось в 1930-х годах) и т.д. Кроме того, были обнаружены всевозможные промежуточные этапы возникновения новых видов, что свидетельствует о плавном характере видообразования.
Эволюция кишечной палочки в лаборатории
В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшества.
В ходе долгосрочного эволюционного эксперимента на бактериях E. coli в одной из 12 подопытных популяций после 31 000 поколений появился новый полезный признак - способность питаться цитратом в аэробных условиях. Как выяснилось, формирование эволюционного новшества проходило в три этапа. На первом этапе ("потенцирование") зафиксировались мутации, помогающие утилизировать цитрат, если он окажется в клетке. Повторные эксперименты с размороженными представителями предковых поколений показали, что потенцирующих мутаций было как минимум две: одна появилась после 15 000 поколений, другая после 20 000. На втором этапе ("актуализация") изменилась регуляция гена citT, ответственного за поглощение цитрата из внешней среды. В результате ген стал работать в присутствии кислорода, хотя в норме у E. coli он работает только в анаэробных условиях. Это привело к появлению слабо выраженной, "зачаточной" способности использовать в пищу цитрат. Наконец, на этапе "совершенствования" новая функция была многократно усилена благодаря нескольким дупликациям (удвоениям) фрагмента хромосомы, несущего активированный ген citT. Только после этого численность бактерий-мутантов увеличилась и они стали доминировать в своей популяции. Вероятно, такое поэтапное развитие характерно и для других эволюционных новшеств.
12 подопытных популяций кишечной палочки (E. coli) живут в аэробных условиях в жидкой прозрачной среде, где единственной пищей является глюкоза. С интервалом в сутки из каждой популяции берут небольшую часть и пересаживают в новую колбу с питательной средой. Там бактерии быстро размножаются, пока не исчерпают запасы глюкозы. Таким образом, в течение каждого суточного цикла начальный период изобилия, когда численность популяции быстро растет (численность оценивают по мутности среды), сменяется периодом голода, когда бактерии перестают размножаться и их численность стабилизируется.
По прошествии 31 000 поколений в одной из 12 популяций (ее условное обозначение - Ara-3) произошло нечто странное. Ее численность теперь продолжала расти и после того, как вся глюкоза в колбе была съедена. К концу суточного цикла численность популяции Ara-3 оказывалась намного выше, чем у остальных популяций. Очевидно, микробы научились использовать в пищу какое-то вещество, входящее в состав среды, но несъедобное для обычных кишечных палочек.
Вскоре Ленски и его коллеги выяснили, что бактерии из популяции Ara-3 приобрели способность питаться цитратом (лимонной кислотой). Цитрат добавляется в среду в качестве вспомогательного вещества (хелатирующего агента). Нормальные бактерии E. coli способны усваивать цитрат только в анаэробных условиях. Неспособность питаться цитратом в присутствии кислорода - один из определительных признаков данного вида бактерий. Таким образом, в популяции Ara-3 появилось настоящее "эволюционное новшество" - новый полезный признак (авторы обозначили его Cit+), изменивший взаимоотношения организма со средой и позволивший бактериям-мутантам увеличить свою численность и стать доминирующими в популяции. Любопытно, что бактерии Cit+ так и не вытеснили до конца бактерий Cit-, которые питаются только глюкозой. Возможно, их длительное мирное сосуществование объясняется разделением ниш, но это пока лишь гипотеза.
Дизайн долгосрочного эксперимента позволяет проследить формирование эволюционного новшества во всех генетических подробностях. Именно для решения подобных задач эксперимент и был поставлен. Часть представителей каждой популяции периодически замораживают. Это нисколько не вредит здоровью микробов. В любой момент можно их оттаять и использовать для повторных опытов. Это, в частности, позволяет оценить вероятность того или иного эволюционного события: насколько оно было случайным или закономерным.
Авторы отсеквенировали геномы 29 бактерий из популяции Ara-3, замороженных в разное время. Это позволило построить эволюционное дерево. Оказалось, что популяция оставалась генетически разнообразной на протяжении почти всей своей истории. В результате мутаций и отбора из одного предкового клона получилось несколько генетически различающихся линий (клад), но они не спешили вытеснять друг друга. Скорее всего, это значит, что большинство генетических различий между кладами были нейтральными, то есть не влияли на приспособленность (а в некоторых случаях, как уже говорилось, могло идти взаимное приспособление и раздел ниш).
Исследователи выявили конкретное генетическое изменение, которое дало бактериям возможность питаться цитратом. Микробы Cit- превратились в Cit+ благодаря дупликации (удвоению) фрагмента хромосомы, содержащего ген citT. Этот ген кодирует белок, транспортирующий цитрат из внешней среды в цитоплазму бактериальной клетки. В норме у E. coli ген citT активен только в анаэробных условиях. Однако в результате дупликации одна из двух копий citT попала под управление промотора другого, соседнего гена (rnk). При помощи генно-инженерных экспериментов авторы показали, что промотор гена rnk обеспечивает работу контролируемого им гена в аэробных условиях.
Разобравшись с природой ключевой мутации, авторы задались следующим вопросом: было ли появление фенотипа Cit+ в кладе 3 популяции Ara-3 полностью случайным? Могло ли это событие с тем же успехом произойти в другой кладе или в другой популяции, или оно было подготовлено предшествующей эволюцией именно этой клады? В поисках ответа авторы провели повторные эволюционные эксперименты с размороженными представителями клад 1, 2 и 3, а также с исходным (предковым) штаммом.
Выяснилось, что у предкового штамма вероятность появления фенотипа Cit+ пренебрежимо мала. Представители клады 1 выработали этот фенотип в двух повторных экспериментах из 55. Микробы из клады 2 научились питаться цитратом в двух случаях из 97. Наконец, бактерии из клады 3 приобрели эту способность в восьми случаях из 37 повторных опытов.
Приобретение фенотипа Cit+ во всех случаях было связано с тем, что ген citT начинал экспрессироваться в аэробных условиях. Но достигался этот результат разными путями. В части повторных опытов, как и в основном долгосрочном эксперименте, произошли тандемные дупликации, благодаря которым citT оказался под управлением аэробного промотора. Но границы дуплицированных участков в разных случаях были разными, а промотор, взявший на себя управление геном citT, не всегда был промотором гена rnk: в одном случае это был промотор гена rna. В нескольких повторных опытах тот же эффект был достигнут без дупликации - благодаря встраиванию мобильного генетического элемента IS3 в кодирующую часть гена citG, расположенного по соседству с citT. В этом мобильном элементе как раз имеется подходящий промотор.
Повторные эксперименты показали, что вероятность приобретения полезного признака Cit+ постепенно росла в ряду "предковый штамм - клады 1 и 2 - клада 3". Стало быть, появление данного признака именно в кладе 3 не было простой случайностью. Микробы были заранее подготовлены к этому - "потенцированы". Иными словами, у них в ходе предшествующей эволюции зафиксировались какие-то мутации, которые сами по себе не давали возможности питаться цитратом, но повысили вероятность развития этой способности в будущем. Разумеется, "потенцирующие" мутации зафиксировались не потому, что микробы планируют свою эволюцию наперед. Эти мутации либо были нейтральными и зафиксировались случайно, либо были полезны для чего-то другого. Подготовка к приобретению способности питаться цитратом была побочным эффектом, которого естественный отбор не мог предусмотреть.
Какие именно мутации, характерные для клады 3, сыграли роль "потенцирующих", установить пока не удалось. Хорошим кандидатом является мутация в гене arcB, потому что повреждение этого гена активирует цикл трикарбоновых кислот - биохимический путь, позволяющий клетке эффективно утилизировать цитрат (если клетка вдруг сумеет его где-то раздобыть в больших количествах). Может быть, повреждение гена arcB превратило мутации, активирующие транспорт цитрата в клетку, из нейтральных или вредных в полезные? Предположение выглядит правдоподобным, но авторы пока не сумели подтвердить его экспериментально.
Однако им удалось подтвердить, что какие-то генетические особенности клады 3 действительно делают умение поглощать цитрат из внешней среды более полезным. Чтобы убедиться в этом, они вставляли в представителей предкового штамма и клад 1-3 по несколько копий гена citT под управлением аэробного промотора. Иными словами, они придавали микробам способность поглощать цитрат из внешней среды - и смотрели, пойдет ли эта способность им на пользу.
Представители всех генно-модифицированных штаммов приобрели способность подкармливаться цитратом, но в очень разной степени. Микробы из клады 3 делали это лучше всех: они быстро переключались с глюкозы на цитрат и хорошо росли на чистом цитрате. Микробы из клад 2 и 1 справлялись гораздо хуже: им требовалось больше времени на переключение, и росли на цитрате они медленнее. Предковый штамм использовал цитрат еще менее эффективно. Для того, чтобы после исчерпания запасов глюкозы переключиться на питание цитратом, ему требовалось около двух суток. Между прочим, это означает, что в условиях долгосрочного эксперимента он не получил бы никакого преимущества от мутации, которая создала фенотип Cit+ в кладе 3: никто не дал бы ему двух суток на раздумья, ведь бактерий пересаживали в новую среду с глюкозой каждые сутки.
Эти результаты, как и статистика повторных эволюционных экспериментов, показывают, что "потенцирование" проходило как минимум в два этапа. Вероятность формирования фенотипа Cit+ выросла при переходе от предкового штамма к кладам 1 и 2 и снова увеличилась при формировании клады 3.
Интересно, что первые представители популяции Ara-3, у которых после поколения №31 000 был зарегистрирован фенотип Cit+, использовали цитрат еще очень неэффективно, хотя у них уже были и "потенцирующие" мутации, и тандемная дупликация. Новая функция поначалу была крайне несовершенна и давала лишь едва заметное преимущество. В ходе дальнейшей эволюции эффективность использования цитрата микробами Cit+ быстро росла. Авторам удалось расшифровать один из механизмов усовершенствования: фрагмент ДНК с активированным геном citT просто-напросто подвергся еще нескольким дупликациям. Это, по-видимому, ускорило поглощение цитрата из внешней среды.
Таким образом, формирование эволюционного новшества происходило в три этапа. На первом этапе (потенцирование, potentiation) закрепились мутации, повысившие вероятность появления признака в будущем. На втором этапе (актуализация, actualization) появилась и была поддержана отбором ключевая мутация, превратившая микробов Cit- в Cit+. Правда, поначалу новый признак был слабо выражен и почти не приносил пользы. На заключительном, третьем этапе (усовершенствование, refinement) признак постепенно оптимизировался. В результате его полезность многократно выросла.
Усовершенствование - процесс, который может продолжаться долго. Однако события, происходившие в популяции Ara-3 после поколения №35 000, трудно анализировать. В это время среди микробов Cit+ распространилась мутация, повышающая скорость мутагенеза примерно в 20 раз (фенотип mutator). Такие "мутаторы" появились не только в Ara-3, но и в нескольких других популяциях. Найти среди сотен новых мутаций те, что связаны с усовершенствованием цитратного питания, - задача пока слишком трудная. Впрочем, замороженные бактерии никуда не спешат и будут спокойно дожидаться появления новых исследовательских методик и приборов.
Опыты Г. Шапошникова по искусственной эволюции
В конце 1950-х - начале 1960-х годов советским биологом Георгием Христофоровичем Шапошниковым была проведена серия опытов, в процессе которых проводилась смена кормовых растений у различных видов тлей. Во время опытов впервые наблюдалась репродуктивная изоляция использованных в эксперименте особей от исходной популяции, что свидетельствует об образовании нового вида.
Большинство тлей чрезвычайно специфичны к выбору кормового растения и обитают на растениях только одного рода и даже одного вида. По большей части это определяется их пищевой специализацией, хотя иногда имеет значение строение ног и хоботка, приспособленные к определенному типу поверхности. Тем не менее, среди тлей время от времени происходит смена кормового растения, сопровождаемая долгим периодом пищевой адаптации.
Г.Х. Шапошников стал одним из первых биологов, который экспериментально изучил этот процесс.
В 1940-х и 1950-х годах пищевую адаптацию тлей исследовали И.В. Кожанчиков, Е.С. Смирнов и З.Ф. Чувахина, однако они не довели опыты до логического завершения, проследив лишь небольшое количество поколений.
Методика исследования
Шапошников исследовал адаптацию при смене кормового растения для тлей подрода Dysaphis CB, s. str. Опыты проводились в 1954-1957 годах на хуторе Шунтук в районе Майкопа и в 1958 году в Ленинграде.
Первичными кормовыми растениями для всех видов тлей были яблони.
Шапошников использовал поиск случайных благоприятных сочетаний паразита и кормового растения. Во всех опытах использовались чистые клоны, произведенные партеногенетически от одной самки. В каждом случае формировалась популяция из 10 самок-клонов, которым предоставлялся выбор из нескольких вторичных хозяев, растущих в одном вазоне. Растения брались с разных стаций, отличных по почвенным и климатическим условиям.
Клоны, потомки которых лучше всего прижились на новом кормовом растении, были в качестве контрольной группы размножены на 2-3 старых хозяевах и 3-10 новых.
Совместимость с новыми хозяевами
Без адаптации участвовавшие в опытах насекомые были в малой степени совместимы с новыми кормовыми растениями. Только 6-9% насекомых могли питаться и не более 4% - размножаться.
Адаптация к новым хозяевам
Тли A. chaerophyllina в течение 4 поколений на новом хозяине вполне освоили его, хотя до 8-го поколения их жизнедеятельность была неустойчивой из-за резких колебаний смертности. После 8 поколений они не потеряли способность жить на старом хозяине.
У тли А. majkopica до 7-го поколения наблюдались резкие колебания плодовитости и смертности, однако начиная с 8-го поколения жизнедеятельность стабилизировались. Однако начиная с 8-го поколения они потеряли способность питаться и размножаться на старом кормовом растении.
Таким образом, опыты показали, что в 8-9 поколениях тли получили способность питаться, а с 10-11 поколений нормально размножаться на ранее совершенно непригодном кормовом растении.
Интенсивность естественного отбора
Средняя смертность личинок тлей на первичном кормовом растении была невелика. Во время периода адаптации к новому кормовому растению смертность увеличивалась в десятки раз и в некоторых случаях достигала 100%. По окончании периода адаптации смертность снижалась к уровню, характерному для видов, изначально специализировавшихся на данном растении.
Таким образом, естественный отбор интенсифицируется в период адаптации, в привычных же условиях он действует очень слабо. Процесс эволюции представляет собой чередование кратковременных периодов быстрых изменений с периодами относительного покоя и стабилизации.
У A. chaerophyllina адаптация шла в направлении использования нового, но пригодного корма. В результате интенсивность отбора была невелика, соответственно невелика была и глубина адаптации: освоив новое кормовое растение, тли не потеряли способность жить на старом.
У A. majkopica шел процесс освоения нового, ранее малопригодного корма. Интенсивность естественного отбора была чрезвычайно высока. В результате произошла глубокая адаптация, в процессе которой тли потеряли способность жить на старом кормовом растении.
Жизненный цикл тлей рода Dysaphis
Тли рода Dysaphis двудомны. На своем первичном хозяине (яблоне) они зимуют в виде яиц. Весной из яйца появляется самка-основательница и производит крылатых мигрантов, которые перелетают на вторичного хозяина - зонтичное растение. До осени мигранты размножаются путем партеногенеза и живорождения. Осенью на вторичном кормовом растении появляются ремигранты - самцы и так называемые полоноски, которые возвращаются к первичному хозяину. Здесь полоноски рождают бескрылых номальных (амфигонных) самок, которые спариваются с самцами и откладывают в трещины коры зимующие яйца.
Опыты по скрещиванию
В опытах по скрещиванию участвовали три популяции.
В опытах по скрещиванию нормальные самки одной формы скрещивались с самцами другой формы. Скрещивание производилось в прозрачных изоляторах, накрытых сетчатой тканью (мельничным газом). Кладка производилась либо в трещины коры (если спаривание происходило на яблоне), либо на кусочки бумаги, подкладываемые в изоляторы. Яйца зимовали в холодильнике, и весной из жизнеспособных яиц выходили самки-основательницы. Жизнеспособные яйца заметно отличались по внешнему виду - они были черными и блестящими в отличие от матовых, не черной окраски и часто сморщенных нежизнеспособных яиц.
Опыты по скрещиванию показали, что:
Между А. majkopica (m) и А. chaerophyllina (c) существует практически полная репродуктивная изоляция. При скрещивании m+c (самцы m, самки c) жизнеспособные яйца вообще не появляются, а при скрещивании c+m - в среднем 1 жизнеспособное яйцо на 100 самок.
После адаптации А. majkopica (n) на ранее непригодном для него кормовом растении возникает практически полная репродуктивная изоляция с исходными видом А. majkopica (m). При скрещивании m+n жизнеспособные яйца вообще не появляются, а при скрещивании n+m - не более 4 жизнеспособных яиц на 100 самок.
У насекомых А. majkopica (n), адаптировавшихся к новому хозяину, возникает репродуктивная совместимость с аборигенным для этого растения подвидом А. chaerophyllina (c). Скрещивание n+с по числу жизнеспособных яиц значительно превышает скрещивания с+с, n+n и m+m, однако скрещивание c+n уступает по этому критерию трем последним, что свидетельствует об определенной остаточной несовместимости.
Интерпретация результатов
Опыты Шапошникова вызвали сенсацию в среде биологов. Образование нового вида в течение нескольких поколений и в сроки, измеряемые месяцами, было явлением беспрецедентным.
Есть предположение, что быстрые эволюционные изменения в опытах Шапошникова связаны с ужесточением отбора только по одному признаку (непривычное питание) с одновременным ослаблением его по всем остальным (хищники, паразиты, болезни, неблагоприятные погодные условия). Аналогичные изменения наблюдаются у популяций, заселяющих обособленные территории (удаленные острова, озера), где отсутствует конкуренция со стороны других видов.
Быстрая эволюция ящериц в новых условиях
Согласно опубликованному в 2008 году исследованию, у популяции стенных ящериц всего лишь за 36 лет (ничтожный для эволюции период) изменились размер и форма головы, увеличилась сила укуса и развились новые структуры в пищеварительном тракте. Эти физические перемены произошли одновременно с серьезными изменениями в популяционной плотности и социальной структуре.
В 1971 году пять пар взрослых особей ящериц с острова Под Кописте были перевезены на соседний остров Под Маркару (эти хорватские острова находятся на юге Адриатического моря, недалеко от Ластово). Условия на новом месте были близки к привычной среде обитания, наземные хищники отсутствовали, и за три десятилетия ящерицы естественным образом распространились по острову, вытеснив при этом местную популяцию ящериц. Когда исследователи вернулись на Под Маркару, они обнаружили, что ящерицы на этом острове существенно отличаются от ящериц на Под Кописте. Сравнение митохондриальной ДНК показало, что популяции генетически неразличимы, но в то же время в популяции с Под Маркару средний размер особей увеличен, задние конечности короче, максимальная скорость бега меньше, а также отличается реакция на нападение хищников. Эти изменения строения и поведения объясняются меньшей активностью хищников на Под Маркару и наличием более густой растительности, способствующей маскировке. Кроме того, в новой популяции существенно изменились размер и форма головы, она стала более массивной, увеличилась сила укуса. Эти физические перемены соответствуют изменению рациона: с Под Кописте преимущественно насекомоядные, а диета ящериц на Под Маркару в зависимости от сезона почти на две трети состоит из растительной пищи. Растения на новом месте обитания жесткие и волокнистые, а мощные челюсти позволяют качественнее откусывать и размельчать растительную еду.
Наиболее неожиданным различием оказалось наличие у новой популяции илеоцекального (подвздошно-слепокишечного) клапана, предназначенного для замедления прохода пищи за счет создания в кишке камер брожения, в которых микробы могут разбивать трудно перевариваемые кусочки растительности. Эти клапаны никогда ранее не были обнаружены у этой разновидности ящериц, включая исходную популяцию с острова Под Кописте. Специалисты называют их "эволюционным новшеством, новым свойством, отсутствовавшим у предковой популяции и развившемся в ходе эволюции у этих ящериц". Кроме того, в кишечнике новой популяции присутствуют нематоды, отсутствующие у исходной популяции.
Изменения в диете также вызвали изменения в плотности популяции и социальной структуре. Из-за того, что растения - это намного больший, предсказуемый и надежный источник пропитания, чем насекомые, количество ящериц увеличилось. Кроме того, пропала необходимость гоняться за добычей, что привело к тому, что ящерицы перестали агрессивно защищать свои территории.
Развитие устойчивости к антибиотикам и пестицидам
Развитие и распространение устойчивости к антибиотикам у бактерий и устойчивых к пестицидам растений и насекомых является доказательством эволюции видов. Появление устойчивых к ванкомицину форм золотистого стафилококка и та опасность, которую они представляют для пациентов больниц, - это прямой результат эволюции путем естественного отбора. Еще один пример - развитие штаммов шигеллы, устойчивых к антибиотикам из группы сульфаниламидов. Появление малярийных комаров, устойчивых к ДДТ, и развитие у австралийских популяций кроликов устойчивости к миксоматозу демонстрируют действие эволюции в условиях давления отбора при быстрой смене поколений.
За последние 50 лет зарегистрировано более 2500 случаев адаптации насекомых-вредителей к различным пестицидам. Поэтому для борьбы с вредителями вместо обычных ядов стараются использовать биологические пестициды - энтомопатогенные вирусы. В частности, для борьбы с яблонной плодожоркой (личинки которой - это те самые червяки в червивых яблоках). Но в 2003 году были зафиксированы первые случаи снижения эффективности вируса, и к 2005 году появились популяции яблонной плодожорки, у которых устойчивость к вирусу выросла в 100 тысяч раз по сравнению с исходной. Установлено, что причиной устойчивости является мутация в половой Z-хромосоме.
Примеры видообразования
Согласно синтетической теории эволюции, образование новых видов происходит в результате разделения особей одного вида на группы, не скрещивающиеся между собой. Такое разделение может быть вызвано различными причинами, такими как наличие непреодолимых географических барьеров (аллопатрическое видообразование) или разделение по экологическим нишам внутри одного ареала (симпатрическое видообразование). После возникновения между популяциями различных типов изоляции, популяции начинают эволюционировать независимо, в результате чего между популяциями постепенно накапливаются генетические различия, и через некоторое время достигается генетическая несовместимость - скрещивание оказывается невозможным.
Кроме того, новые виды могут возникать практически мгновенно (в геологическом масштабе времени) путем полиплоидии (кратного увеличения количества хромосом в клетке) и гибридизации (скрещивания различных видов).
Наблюдения за современными видами показывают, что видообразование непрерывно происходит в существующих популяциях.
Промежуточные этапы видообразования
В определении понятия биологического вида существенное значение имеет практически полная репродуктивная изоляция в природных условиях. Если видообразование происходит постепенно, в результате небольших последовательных изменений, то и сегодня мы должны наблюдать все возможные стадии видообразования, а значит и все возможные стадии репродуктивной изоляции: свободное скрещивание между популяциями, затрудненное скрещивание, снижение плодовитости потомства, бесплодное потомство, и наконец - полную генетическую изоляцию. Если ничего подобного среди современных видов обнаружить не удается, то тем самым опровергаются основы эволюционного учения, то есть это пример фальсифицируемости эволюционного учения.
Существует множество примеров того, как разные виды могут скрещиваться в исключительных условиях. Например, у черной вороны и серой вороны разные ареалы, но на пересечении этих ареалов виды могут образовывать гибриды. Еще один пример - различные виды рыбы рода чукучанов из реки Платт, которые живут рядом, но редко скрещиваются.
Неполное видообразование можно наблюдать у различных кольцевых видов: представители вида обитают вокруг гор, водоемов и т.п., при этом соседние популяции скрещиваются между собой и незначительно различаются, а крайние формы различаются значительно и не скрещиваются. Пример кольцевого вида - безлегочная саламандра, обитающая на склонах гор близ тихоокеанского побережья Северной Америки. В зависимости от района обитания вокруг гор саламандры образуют различные формы, постепенно изменяющие свои морфологические и экологические характеристики. Крайние формы сосуществуют в узкой зоне гор на юге Калифорнии, однако между собой не скрещиваются. Другие примеры кольцевых видов - зеленая пеночка, оленья мышь, некоторые виды наземных улиток и дрозофил.
У многих гибридов снижена фертильность. У гибридов землероек из рода бурозубок размножение затруднено вследствие разницы хромосомных наборов. Тот же эффект наблюдался в лаборатории при скрещивании Drosophila pseudoobscura из Юты и Калифорнии. Фертильность также снижена у гибридов лягушек жерлянки краснобрюхой и жерлянки желтобрюхой.
Многие другие виды могут давать бесплодное гибридное потомство. В их числе - различные земноводные (например - некоторые виды лягушек) и млекопитающие (например, мул - гибрид от скрещивания лошади и осла - как правило, бесплоден). Гибриды гребенчатого тритона и мраморного тритона бесплодны вследствие наличия непарных хромосом.
При скрещивании ряда других видов достигается оплодотворение, но позднее зародыш погибает. В качестве примера можно привести леопардовую лягушку и лесную лягушку из рода настоящих лягушек. Аналогичная ситуация наблюдается у дрозофил и у некоторых растений, таких как хлопчатник обыкновенный и хлопчатник барбадосский.
Белый медведь
Конкретным примером крупномасштабных эволюционных изменений при неполном видообразовании является белый медведь, который, несмотря на родство с бурым медведем, очевидное в силу того факта, что эти виды могут скрещиваться и давать плодовитое потомство, приобрел значительные физиологические различия с бурым медведем. Эти различия позволяют белому медведю комфортно жить в условиях, в которых бурый медведь не выжил бы. В частности, белый медведь способен проплыть десятки километров в ледяной воде, его окраска сливается со снегом, он не замерзает в Арктике. Все это возможно благодаря конкретным изменениям: белая окраска способствует маскировке хищника при охоте на тюленей; полые волоски увеличивают плавучесть и сохраняют тепло; слой подкожного жира, толщина которого к зиме доходит до 10 сантиметров, обеспечивает дополнительную теплоизоляцию; удлиненная, по сравнению с другими медведями, шея позволяет легче держать голову над водой во время плавания; увеличенные лапы с перепонками действуют как весла; небольшие бугорки и полости-присоски на подошвах уменьшают опасность поскользнуться на льду, а плотная шерсть на подошвах защищает лапы от сильного холода и обеспечивает трение; уши меньше, чем у других медведей, и уменьшают потери тепла; веки действуют как солнечные очки; зубы острее, чем у других медведей, и больше подходят для полностью мясного рациона; увеличенный объем желудка позволяет голодному хищнику съесть сразу целого тюленя, кроме того, белый медведь способен обходиться без пищи до девяти месяцев за счет переработки мочевины.
Наблюдаемое видообразование
Судя по палеонтологической летописи и по измерениям скорости мутаций, полная несовместимость геномов, делающая невозможным скрещивание, достигается в природе в среднем за 3 млн лет. А значит, наблюдение образования нового вида в естественных условиях в принципе возможно, но это редкое событие. В то же время, в лабораторных условиях скорость эволюционных изменений может быть увеличена, поэтому есть основания надеяться увидеть видообразование у лабораторных животных.
Известны многие случаи видообразования посредством гибридизации и полиплоидизации у таких растений как конопля, крапива, первоцвет, редька, капуста, а также у различных видов папоротников. В ряде случаев видообразование у растений происходило без гибридизации и полиплоидизации (кукуруза, стефаномерия из семейства астровых).
Дрозофилы, также известные как плодовые мухи, входят в число наиболее изученных организмов. С 1970-х годов зафиксированы многие случаи видообразования у дрозофил. Видообразование происходило, в частности, за счет пространственного разделения, разделения по экологическим нишам в одном ареале, изменения поведения при спаривании, дизруптивного отбора, а также за счет сочетания эффекта основателя с эффектом бутылочного горлышка.
Видообразование наблюдалось в лабораторных популяциях комнатных мух, яблонных мух-пестрокрылок, мучных жуков, комаров и других насекомых.
Известны случаи, когда в результате давления отбора (в присутствии хищников) одноклеточные зеленые водоросли из рода хлорелла образовывали многоклеточные колониальные организмы, а у бактерий в аналогичных условиях менялось строение и увеличивались размеры (c 1,5 до 20 микрометров за 8-10 недель). Являются ли эти случаи примерами видообразования, зависит от того, какое используется определение вида (при бесполом размножении нельзя использовать критерий репродуктивной изоляции).
Видообразование также наблюдалось и у млекопитающих. Шесть случаев видообразования у домовых мышей на острове Мадейра за последние 500 лет были следствием исключительно географической изоляции, генетического дрейфа и слияния хромосом. Слияние двух хромосом - это наиболее заметное различие геномов человека и шимпанзе, а у некоторых популяций мышей на Мадейре за 500 лет было девять подобных слияний.
Яблонная муха
Яблонная пестрокрылка является примером наблюдаемого симпатрического видообразования (то есть видообразования в результате разделения по экологическим нишам). Первоначально вид обитал в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Однако с завозом в Америку яблонь (первое упоминание яблонь в Америке - 1647 год), открылась новая экологическая ниша. В 1864 году личинки были обнаружены в яблоках, тем самым зафиксирована яблонная раса этого вида. За полтора века наблюдений расы очень сильно разошлись. Они почти не скрещиваются друг с другом (уровень гибридизации не превышает 4-6%). Яблоневая раса спаривается почти исключительно на яблонях, а боярышниковая - на боярышнике, что, учитывая разное время созревания плодов, приводит к репродуктивной изоляции. В скором времени возможно превращение этих рас в самостоятельные виды.
У пестрокрылок известно еще несколько видов-двойников, которые живут на разных видах растений, - предположительно, видообразование у них протекало именно по описанной схеме.
Эволюционное дерево
Общепринятая в научном сообществе классификация живых организмов представляет собой многоуровневую иерархическую структуру: организмы делятся на царства, царства делятся на типы, типы - на классы, классы - на отряды, и так далее. В результате такого ветвления получается филогенетическое дерево. Существуют разногласия между биологами по поводу отнесения отдельных видов к конкретным группам, но эти противоречия имеют частный характер. Практика показывает, что биологические классификации, построенные на основе разных признаков, в тенденции стремятся к одной и той же древовидной иерархической схеме - естественной классификации. А значит, в основе этой классификации, возможно, существует закономерность.
Именно такой результат можно ожидать при эволюционном происхождении животных от общего предка. Ветвление филогенетического дерева соответствует делению популяций в процессе видообразования.
Как правило, объекты, возникшие не в ходе эволюции, не обладают этим свойством. Невозможно объективно построить иерархию элементарных частиц, химических элементов, планет Солнечной системы. Также не существует объективной иерархии таких сознательно созданных объектов как книги в библиотеке, дома, мебель, машины. Можно при желании объединять эти объекты в различные иерархии, но нет единственной объективной иерархии, принципиально лучшей, чем все остальные.
Существует ряд статистических методов для точной оценки того, насколько объекты с разными свойствами укладываются в ту или иную иерархию. Эти методы измеряют так называемый "филогенетический сигнал", позволяя отличить мнимые закономерности от объективных. Например, у любого "генеалогического древа" автомобилей будет низкий уровень филогенетического сигнала. У эволюционного дерева и у различных его частей, напротив, стабильно четкий филогенетический сигнал.
Есть несколько источников данных, на основе которых можно делать выводы о степени родства между видами. Если существует единое эволюционное дерево, объединяющее все виды в объективную генеалогию, то все данные должны подтверждать эту генеалогию. Филогенетические деревья, построенные независимо, должны соответствовать друг другу. Наиболее простой, хотя и не очень точный способ построить такое дерево - сравнение строения организмов животных: чем меньше различий между видами, тем меньше поколений отделяют их от общего предка. Палеонтологическая летопись позволяет установить родство между классами животных. Например, найденные останки пернатых динозавров свидетельствуют о родстве между рептилиями и птицами. Биогеография и эмбриология также дают информацию об эволюционной близости видов. Наиболее точный источник данных, недоступный во времена Дарвина - сравнительный анализ геномов различных организмов. Эволюционное дерево можно построить по каждому отдельно взятому гену, также исследователи рассматривают всевозможные некодирующие белки последовательности.
Все эти источники информации дают одинаковую картину с точностью до погрешности используемых методов. Тот факт, что эволюционные деревья, построенные по разным данным, соответствуют друг другу, объясняется эволюционной теорией. Другие объяснения отсутствуют: например, нет никаких причин, почему организмы, сходные по строению, должны иметь сходные некодирующие последовательности в геноме, если они не произошли от одного предка.
Как и любая модель, филогенетическое дерево соответствует действительности не абсолютно точно. Горизонтальная передача генов и гибридизация затрудняют использование деревьев для описания связей между видами. В тех редкихслучаях, когда необходимо учитывать эти факторы, приходится использовать более сложные модели.
Палеонтологические доказательства
Как правило, останки растений и животных разлагаются и исчезают без следа. Но иногда биологические ткани замещаются минеральными веществами, и образуются окаменелости. Обычно находят окаменевшие кости или раковины, то есть твердые части живых организмов. Иногда находят отпечатки следов животных или следы их жизнедеятельности. Еще реже находят животное целиком - вмороженным в лед в районах современной вечной мерзлоты, попавшим в окаменевшую позже смолу древних растений (янтарь) или в другую естественную смолу - асфальт.
Изучением ископаемых останков занимается палеонтология. Как правило, осадочные породы залегают слоями, поэтому более глубокие слои содержат окаменелости из более раннего периода (принцип суперпозиции). А значит, сравнивая ископаемые формы из последовательных напластований, можно делать выводы об основных направлениях эволюции живых организмов. Для оценки возраста окаменелостей используются многочисленные методы геохронологии.
При взгляде на палеонтологическую летопись можно сделать вывод, что жизнь на Земле существенно менялась. Чем глубже в прошлое мы смотрим, тем меньше видим общего с современной биосферой. Первые прокариоты (простейшие одноклеточные, не обладающие оформленным клеточным ядром) появляются приблизительно 3,5 миллиарда лет назад. Первые одноклеточные эукариоты появляются 2,7-1,75 миллиардов лет назад. Примерно через миллиард лет, 840 миллионов лет назад, в палеонтологической летописи появляются первые многоклеточные животные, представители хайнаньской фауны. Согласно опубликованному в 2009 году исследованию, вероятно, более 635 миллионов лет назад уже существовали многоклеточные, относящиеся к одному из современных типов - губки. В период "кембрийского взрыва", 540-530 миллионов лет назад, за геологически короткий промежуток времени, в геологической летописи появляются остатки представителей большинства современных типов, имеющих скелеты, а еще через 15 миллионов лет - первые примитивные позвоночные, похожие на современных миног. Челюстноротые рыбы появляются 410 миллионов лет назад, насекомые - 400 миллионов лет назад, и еще 100 миллионов лет на суше господствуют папоротниковидные, а основными группами наземной фауны остаются насекомые и земноводные. С 250 по 65 миллионов лет назад на Земле господствующее положение "верховных хищников" и крупных травоядных занимали динозавры и другие пресмыкающиеся, самыми распространенными растениями были саговники и другие группы голосеменных. Первые ископаемые остатки цветковых появляются 140-130 миллионов лет назад, а начало их широкого распространения относится к середине мелового периода (около 100 миллионов лет назад). Наблюдаемая картина соответствует происхождению всех видов от одноклеточных организмов и не имеет других научных объяснений.
Известное доказательство эволюции - наличие так называемых промежуточных форм, то есть организмов, сочетающих в себе характерные признаки разных видов. Как правило, говоря о промежуточных (или переходных) формах имеют ввиду ископаемые виды, хотя промежуточные виды не всегда вымирают. На основе филогенетического дерева теория эволюции предсказывает, какие промежуточные формы могут быть найдены, а какие - нет. В соответствии с научным методом, сбывшиеся предсказания подтверждают теорию. Например, зная строение организмов пресмыкающихся и птиц, можно предсказать некоторые особенности переходной формы между ними. Можно прогнозировать возможность найти останки животных, подобных рептилиям, но с перьями, или останки животных, подобных птицам, но с зубами или с длинными хвостами со скелетом из несросшихся позвонков. При этом можно предсказать, что не будут найдены переходные формы между птицами и млекопитающими, например - ископаемые млекопитающие с перьями или подобные птицам ископаемые с костями среднего уха как у млекопитающих.
Вскоре после публикации "Происхождения видов" были обнаружены останки археоптерикса - промежуточной формы между рептилиями и птицами. Для археоптерикса характерно дифференцированное оперение (типичная птичья черта), а по строению скелета он слабо отличался от динозавров из группы компсогнатов. У него были когти на передних конечностях, зубы и длинный хвост со скелетом из несросшихся позвонков, а предполагаемые уникальные "птичьи" особенности скелета впоследствии были выявлены у других рептилий. Позднее были найдены и другие переходные формы между рептилиями и птицами.
Известно множество других переходных форм, в том числе - от беспозвоночных к рыбам, от рыб к четвероногим, от земноводных к рептилиям и от рептилий к млекопитающим.
В некоторых случаях ископаемые переходные формы обнаружить не удалось, например - нет следов эволюции шимпанзе (предположительно, это объясняется отсутствием условий для образования окаменелостей в лесах, где они живут), нет следов ресничных червей, а этот класс объединяет более 3500 видов.
Однако чтобы фальсифицировать теорию эволюции, недостаточно указать на подобные пробелы в палеонтологической летописи.
Были найдены миллионы окаменелостей около 250 тысяч ископаемых видов, и каждая находка - это проверка теории эволюции, а пройденная проверка подтверждает теорию.
Если находится скелет, не соответствующий филогенетическому дереву или не укладывающийся в хронологическую последовательность, то это означает не неправильность эволюционной теории, а всего лишь ее неполноту.
В тех случаях, когда палеонтологическая летопись оказывается особенно полна, появляется возможность построить так называемые филогенетические ряды, то есть ряды видов (родов и т.д.), последовательно сменяющих друг друга в процессе эволюции. Наиболее известны филогенетические ряды человека и лошади (см. ниже), также в качестве примера можно привести эволюцию китообразных.
Эволюция человека
Первый опознанный экземпляр неандертальца был найден в 1856 году, за три года до опубликования "Происхождения видов". На момент выхода книги не было известно никаких других ископаемых, подтверждающих эволюционное происхождение человека и человекообразных обезьян от общего предка. Хотя неандерталец является независимым видом из рода людей, с тех пор было найдено множество скелетов промежуточных форм между древними антропоидами и человеком.
Поскольку общий предок человека и шимпанзе передвигался на четырех конечностях, а мозг у него был не больше чем у шимпанзе, согласно теории Дарвина в процессе эволюции должно было развиться прямохождение, а также должен был увеличиться объем мозга. Таким образом, должен был существовать один из трех вариантов промежуточной формы:
- Промежуточный размер мозга, развивающееся прямохождение.
- Размер мозга примерно как у шимпанзе, развитое прямохождение.
- Большой мозг, прямохождение не развито.
В 1920-х годах в Африке были обнаружены останки существа, которое Раймонд Дарт назвал австралопитеком. Позднее были найдены и другие останки австралопитеков. Австралопитеки жили в восточной и северной Африке 4-2 миллиона лет назад. Объем мозга австралопитека был немного больше, чем у шимпанзе. Кости таза по строению наиболее близки к костям таза человека. Строение черепа характерно для прямоходящих животных, что можно определить в частности по отверстию в затылочной кости, соединяющему полость черепа с позвоночным каналом. Более того, в Танзании в окаменевшем вулканическом пепле были обнаружены следы, оставленные двуногими гоминидами. Возраст следов составляет 3,6 млн лет. Таким образом, австралопитеки представляют собой "переходную форму номер два": у них было развито частичное прямохождение, но мозг был примерно как у шимпанзе.
Позднее были найдены останки ардипитека, возраст которых составляет 4,5 миллиона лет. Анализ скелета показал, что по земле ардипитеки передвигались на двух задних конечностях, а по деревьям лазали на всех четырех. Прямохождение у ардипитеков было слабо развито по сравнению с последующими гоминидами, они не могли преодолевать большие расстояния. Ардипитек представляет собой переходную форму между общим предком человека и шимпанзе и австралопитеком.
Итак, в ходе эволюции у гоминид сначала развилось прямохождение, и только затем началось существенное увеличение объема мозга. У австралопитеков, живших 4-2 миллиона лет назад, объем мозга был около 400 куб.см., примерно как у шимпанзе. Человек умелый (Homo habilis) жил 2,4-1,4 миллиона лет назад, размер мозга у него был 500-640 куб.см. Человек работающий (Homo ergaster) жил 1,9-1,4 миллиона лет назад, размер мозга - 700-850 куб.см. Человек прямоходящий (Homo erectus) жил 1,4-0,2 миллиона лет назад, размер мозга составлял от 850 куб.см. у ранних особей до 1100 куб.см. у поздних. Гейдельбергский человек (Homo heidelbergensis) жил 600-350 тысяч лет назад, размер мозга составлял 1100-1400 куб.см. Неандертальцы жили 350-25 тысяч лет назад, размер мозга составлял 1200-1900 куб.см. 200 тысяч лет назад появился человек разумный (Homo sapiens), размер мозга людей составляет 1000-1850 куб.см. Согласно современным представлениям, не все перечисленные виды были непосредственными предками современных людей. В частности, неандертальцы развивались параллельно с Homo sapiens. Тем не менее, на примере этих видов можно наблюдать направление эволюции гоминид. Перечисленные виды хорошо представлены в палеонтологической летописи.
Согласно Дарвину, "в ряду форм, незаметно переходящих одна в другую от какого-либо обезьянообразного существа до человека в его современном состоянии, было бы невозможно точно указать, которой именно из этих форм следует впервые дать наименование „человека“. Именно с этой проблемой и столкнулись современные палеонтологи, вынужденные делить найденные останки гоминид на разные роды. Например, некоторые черепа в разное время классифицировались как австралопитек и как человек умелый, просто потому что они оказались на искусственной "границе", произвольно проведенной между практически неразличимыми родами.
Эволюция лошади
Эволюция лошади достаточно полно прослеживается в Северной Америке. Древнейший представитель лошадиных - гиракотерий - был размером с лисицу и жил в Северной Америке 54 миллионов лет назад (нижний эоцен), а затем распространился в Европу и в Азию. Это было животное стройного телосложения, с короткими ногами, но приспособленное к бегу. У него было четыре пальца на передних ногах и три пальца на задних, пальцы располагались практически вертикально. Резцы были маленькие, коренные зубы приплюснуты и покрыты сверху эмалью.
Вероятный путь развития лошадей от гиракотериев к современному виду включает не менее 12 родов и несколько сотен видов. При этом произошли следующие изменения:
- Увеличение размера (от 0,4 до 1,6 м)
- Удлинение ног и стопы
- Редукция боковых пальцев
- Удлинение и утолщение среднего пальца
- Увеличение ширины резцов
- Замещение ложнокоренных зубов коренными
- Удлинение зубов
- Увеличение высоты коронки коренных зубов
Судя по ископаемым растениям из различных слоев, болотистая местность, в которой обитал гиракотерий, постепенно становилась суше. Предки лошадей стали полагаться на улучшенный за счет поднятой головы обзор и на высокую скорость бега для спасения от хищников. Если при ходьбе по мягкой болотистой почве большое число пальцев на конечностях было оправдано, то в измененных условиях жизни для первобытных лошадей стало более выгодным, когда боковые пальцы у них стали постепенно отмирать, а средние - развиваться. Изменение строения зубов объясняется изменением рациона, а именно - переходом с мягкой растительности на траву и листья.
Иногда у лошадей развиваются один или два лишних пальца, чаще всего встречается 2-й (внутренний) палец с вполне развитыми пястными или плюсневыми костями, вполне развитыми суставами пальца и копытом, которое, впрочем, редко касается земли. В случае развития двух лишних пальцев нога получает сходство с ногами предковых видов.
Морфологические доказательства
В ходе эволюции каждый новый организм не проектируется с нуля, а получается из старого за счет последовательности небольших изменений. У образовавшихся таким образом структур есть ряд характерных особенностей, указывающих на их эволюционное происхождение. Сравнительно-анатомические исследования позволяют выявить такие особенности.
В частности, эволюционное происхождение исключает возможность целенаправленного заимствования удачных конструкций у других организмов. Поэтому у различных, не близкородственных видов для решения схожих задач используются различные органы. Например, крыло бабочки и крыло птицы развиваются из разных зародышевых листков, крылья птиц представляют собой видоизмененные передние конечности, а крылья бабочки - складки хитинового покрова. Сходство между этими органами поверхностно и является следствием их конвергентного происхождения. Такие органы называют аналогичными.
Противоположная ситуация наблюдается у близкородственных видов: для совершенно разных задач используются схожие по строению органы. Например, передние конечности позвоночных выполняют самые разные функции, но при этом имеют общий план строения, занимают сходное положение и развиваются из одних и тех же зачатков, то есть являются гомологичными. Сходство строения крыла летучей мыши и лапы крота невозможно объяснить с точки зрения полезности. В то же время, теория эволюции дает объяснение: единую структуру конечности четвероногие позвоночные унаследовали от общего предка.
Каждый вид наследует от предкового вида большинство его свойств - в том числе иногда и те, которые для нового вида бесполезны. Изменения обычно происходят за счет постепенного последовательного преобразования признаков предкового вида. Сходство гомологичных органов, не связанное с условиями их функционирования - свидетельство их развития в ходе эволюции из общего прототипа, имевшегося у предкового вида. Другие примеры эволюционных изменений морфологии - рудименты, атавизмы, а также многочисленные случаи специфического несовершенства строения организмов.
Гомологичные органы
Передние конечности млекопитающих представляют собой пример гомологичных органов, так как происходят из одного и тех же тканей зародыша по сходному механизму, состоят из одного и того же набора костей, но при этом выполняют самые разные функции.
Пятипалая конечность
Пятипалая конечность, характерная для четвероногих позвоночных - пример гомологии органов. Более того, прослеживается гомология пятипалой конечности и плавников некоторых ископаемых видов кистеперых рыб, от которых, по всей видимости, произошли первые земноводные. Конечности четвероногих различаются по форме и приспособлены к выполнению самых различных функций в самых разных условиях.
На примере млекопитающих:
- У обезьян передние конечности вытянуты, кисти приспособлены для хватания, что облегчает лазанье по деревьям.
- У свиньи первый палец отсутствует, а второй и пятый - уменьшены. Остальные два пальца длиннее и тверже остальных, концевые фаланги покрыты копытами.
- У лошади также вместо когтей копыто, нога удлинена за счет костей среднего пальца, что способствует большой скорости передвижения.
- Кроты имеют укороченные и утолщенные пальцы, что помогает при копании.
- Муравьед использует крупный средний палец для раскапывания муравейников и гнезд термитов.
- У китов передние конечности представляют собой плавники. При этом число фаланг пальцев увеличено по сравнению с другими млекопитающими, а сами пальцы скрыты под мягкими тканями.
- У летучей мыши передние конечности преобразовались в крылья за счет значительного удлинения четырех пальцев, а крючкообразный первый палец используется, чтобы висеть на деревьях.
При этом все эти конечности содержат сходный набор костей с одним и тем же относительным расположением. Единство структуры не может быть объяснено с точки зрения полезности, так как конечности используются для совершенно разных целей.
Части ротового аппарата насекомых
Основные части ротового аппарата насекомых - верхняя губа, пара жвал (верхних челюстей), подглоточник, две максиллы (нижние челюсти) и нижняя губа (сросшиеся вторые максиллы). У разных видов эти составные части различаются по форме и размеру, у многих видов некоторые из частей утрачены. Особенности строения ротового аппарата позволяют насекомым использовать различные источники пищи:
- В исходном виде (например, у кузнечика) сильные жвалы и максиллы используются для кусания и жевания.
- Медоносная пчела использует нижнюю губу для сбора нектара, а жвалами дробит пыльцу и разминает воск.
- У большинства бабочек верхняя губа уменьшена, жвалы отсутствуют, максиллы образуют хоботок.
- У самок комаров верхняя губа и максиллы образуют трубку, жвалы используются для протыкания кожи.
Аналогичные органы
Внешне схожие органы или их части, происходящие из различных исходных зачатков и имеющие неодинаковое внутреннее строение, называются аналогичными. Внешнее сходство возникает в ходе конвергентной эволюции, то есть в ходе независимого приспособления к сходным условиям существования.
Примеры:
- Крылья птиц - видоизмененные передние конечности, крылья насекомых - складки хитинового покрова.
- Жабры рыб - образования, связанные с внутренним скелетом, жабры многих ракообразных - выросты конечностей, ктенидиальные жабры моллюсков развиваются в мантийной полости, а жабры голожаберных моллюсков - выросты покровов спинной стороны тела.
- Обтекаемая форма тела у водных млекопитающих (китов, дельфинов) и у рыб.
- Колючки барбариса, кактуса - видоизмененные листья, колючки боярышника развиваются из побегов.
- Усики винограда образуются из побегов, усики гороха - видоизмененные листья.
- Форма различных суккулентов (растений, имеющих специальные ткани для запаса воды), таких как кактусы и молочай.
Полное отсутствие целенаправленного заимствования удачных конструкций отличает эволюцию от сознательного проектирования. Например, перо - это удачная конструкция, помогающая при полете, но у млекопитающих (в том числе и у летучих мышей) перья отсутствуют. Жабры чрезвычайно полезны для водных животных, но у млекопитающих (таких как китообразные) они отсутствуют.
Рудименты
Рудиментами называются органы, утратившие свое основное значение в процессе эволюционного развития организма. Рудименты также можно определить независимо от эволюционного учения как структуры, редуцированные и обладающие меньшими возможностями по сравнению с соответствующими структурами в других организмах. Если рудимент и оказывается функциональным, то он выполняет относительно простые или малозначимые функции с помощью структур, очевидно предназначенных для более сложных целей. Хотя многие рудиментарные органы совершенно не функциональны, отсутствие функций не является необходимым условием для рудиментарности. Такие органы крайне распространены в природе.
Например, птичье крыло - крайне сложная анатомическая структура, специально приспособленная для активного полета, но крылья страусов не используются для полета. Эти рудиментарные крылья могут использоваться для сравнительно простых задач, таких как поддержание равновесия на бегу и привлечение самок. Таким образом, специфическая сложность крыльев страуса неадекватна простоте задач, для которых эти крылья используются, и именно поэтому эти крылья называют рудиментами. В числе других нелетающих птиц с рудиментарными крыльями - галапагосские бакланы, киви и какапо. Для сравнения, крыло пингвина имеет большое значение, действуя в качестве плавника, а, значит, не может считаться рудиментом.
Примеры рудиментов:
- Глаза у некоторых пещерных и роющих животных, таких как протей, слепыш, крот, астианакс мексиканский (слепая пещерная рыба). Часто глаза скрыты под кожей.
- Малая берцовая кость у птиц.
- Остатки волосяного покрова и тазовых костей у некоторых китообразных.
- У некоторых змей, в частности у питона, имеются кости задних конечностей. Эти кости не крепятся к позвоночнику и относительно свободно перемещаются в брюшной полости.
- У многих видов жуков крылья лежат под сросшимися надкрыльями.
- У человека к рудиментам в частности относятся хвостовые позвонки, волосяной покров туловища, ушные мышцы, бугорок ушной раковины, морганиевы желудочки гортани.
- Червеобразный отросток слепой кишки (аппендикс) у некоторых травоядных животных используется для переваривания растительной пищи и имеет большую длину. Например, у коалы длина аппендикса составляет от 1 до 2 метров. Аппендикс человека имеет длину от 2 до 20 сантиметров и не участвует в расщеплении пищи. Вопреки распространенному убеждению, наличие у аппендикса второстепенных функций не означает, что он не является рудиментом.
Теория эволюции предсказывает, какие рудиментарные органы могут быть найдены, а какие - принципиально невозможны. Любой рудиментарный орган организма был полностью развит у его предков. А значит, каждый обнаруженный рудимент должен соответствовать эволюционному дереву, в противном случае основы теории эволюции пришлось бы пересматривать. Это очередной пример фальсифицируемости эволюционной теории, то есть свидетельство выполнения критерия Поппера для признания теории научной.
Атавизмы
Атавизмом называется появление у особи признаков, свойственных отдаленным предкам, но отсутствующих у ближайших. Появление атавизмов объясняется тем, что гены, отвечающие за данный признак, сохранились в ДНК, но в норме не формируют структуры, типичные для предков.
Примеры атавизмов:
- Хвостовидный придаток у человека;
- Сплошной волосяной покров на теле человека;
- Добавочные пары молочных желез;
- Задние ноги у китов;
- Задние плавники у дельфинов;
- Задние ноги у змей;
- Дополнительные пальцы у лошадей;
- Возобновление полового размножения у ястребинки волосистой и у клещей семейства Crotoniidae.
Аргументы в пользу эволюции аналогичны аргументам для рудиментов.
Несовершенство строения организмов
В ходе эволюции каждая новая конструкция получается из старой за счет последовательности приспособительных изменений. Эта особенность является причиной специфических несообразностей в строении живых организмов.
Например, возвратный гортанный нерв у млекопитающих идет (в составе блуждающего нерва) от мозга к сердцу, огибает (уже в качестве самостоятельного нерва) дугу аорты и возвращается к гортани. В результате нерв проходит гораздо более длинный путь, чем необходимо, а аневризма аорты может приводить к параличу левой голосовой связки. Особенно наглядно проблема видна на примере жирафа, у которого длина возвратного нерва может достигать 4 метров, хотя расстояние от мозга до гортани - всего несколько сантиметров. Такое расположение нервов и сосудов млекопитающие унаследовали от рыб, у которых шея отсутствует.
Желудочно-кишечный тракт млекопитающих пересекается с дыхательными путями, в результате мы не можем одновременно дышать и глотать, а кроме того существует опасность подавиться. Эволюционное объяснение данного обстоятельства заключается в том, что предками млекопитающих являются кистеперые рыбы, которые заглатывали воздух, чтобы дышать.
Упрощенная схема расположения нейронов сетчатки. Сетчатка состоит из нескольких слоев нейронов. Свет падает слева и проходит через все слои, достигая фоторецепторов. От фоторецепторов сигнал передается биполярным клеткам и горизонтальным клеткам. Затем сигнал передается амакриновым и ганглионарным клеткам. Эти нейроны генерируют потенциалы действия, передающиеся по зрительному нерву в мозг.
Еще один пример - сетчатка позвоночных и слепое пятно. У позвоночных нервные волокна и ганглиозные клетки сетчатки расположены поверх светочувствительных клеток, и свет должен пройти через несколько слоев клеток, прежде чем попадает на палочки и колбочки. Слепое пятно - это участок сетчатки, лишенный фоторецепторов, от которого отходит к мозгу зрительный нерв. Для решения многочисленных проблем, вызванных таким устройством сетчатки, у позвоночных есть ряд адаптаций. В частности, у нервных волокон, идущих поверх сетчатки, отсутствует миелиновая оболочка, что повышает их прозрачность, но снижает скорость передачи сигнала. Схожие по строению глаза головоногих лишены этого недостатка. Это наглядно показывает, что и у позвоночных, возможно, могли бы в ходе эволюции сформироваться глаза без слепых пятен. Предполагается, что такое строение глаза связано со способом его развития в онто- и филогенезе. Вероятно, у общих отдаленных предков хордовых фоторецепторы находились на спинной поверхности тела. Затем, при формировании нервной трубки, они оказались на ее внутренней поверхности, т. е. на стенке нервного канала (как у ланцетника). Глаза современных позвоночных образуются как выпячивания стенок нервной трубки (глазные пузыри), а фоторецепторы по-прежнему формируются в их внутренней стенке.
Такие частые патологии как выпадение матки, частое воспаление носоглотки, боли в пояснице у людей отчасти связаны с тем, что мы используем для прямохождения тело, сформированное эволюцией в течение сотен миллионов лет для передвижения на четырех конечностях.
Эмбриологические доказательства
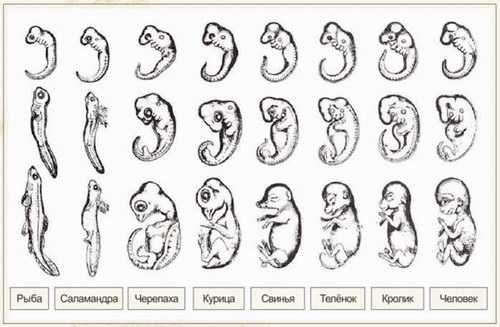
У всех позвоночных животных наблюдается значительное сходство зародышей на ранних стадиях развития: у них похожая форма тела, есть зачатки жаберных дуг, имеется хвост, один круг кровообращения и т.д. (закон зародышевого сходства Карла Максимовича Бэра). Однако по мере развития сходство между зародышами различных систематических групп постепенно уменьшается, и начинают преобладать черты, свойственные их классам, отрядам, семействам, родам, и, наконец, видам.
При этом не следует путать эмбриологические доказательства с "биогенетическим законом" Эрнста Геккеля. Печально известна иллюстрация из работы Геккеля, на которой, согласно распространенной точке зрения, различия между эмбрионами искусственно занижены. Карл Бэр не был эволюционистом, не использовал рисунки Геккеля, открыл свой закон задолго до Геккеля и за 30 лет до публикации "Происхождения видов". Сегодня эмбриологические доказательства эволюции признаны научным сообществом, а "биогенетический закон" - отвергнут.
Эволюционные изменения могут касаться всех фаз онтогенеза, то есть могут приводить к изменениям не только зрелых организмов, но и эмбрионов, даже на первых этапах развития. Тем не менее, более ранние фазы развития должны отличаться большим консерватизмом, чем более поздние, так как изменения на более ранних этапах развития, в свою очередь, должны привести к большим изменениям в процессе дальнейшего развития. Поэтому изменения, проявляющиеся на ранних этапах, гораздо чаще оказываются летальными, чем изменения, касающиеся более поздних периодов онтогенеза.
Таким образом, ранние стадии развития изменяются относительно редко, а значит, изучая эмбрионы разных видов, можно делать выводы о степени эволюционного родства.
В 1837 году эмбриолог Карл Райхерт выяснил, из каких зародышевых структур развиваются квадратная и суставная кости в челюсти рептилий. У зародышей млекопитающих обнаружены те же структуры, но они развиваются в молоточек и наковальню среднего уха. Палеонтологическая летопись также подтверждает происхождение частей уха млекопитающих из костей челюсти рептилий.
Есть многие другие примеры того, как эволюционная история организма проявляется в ходе его развития. У эмбрионов млекопитающих на ранних стадиях есть жаберные мешки, неотличимые по строению от жаберных мешков у водных позвоночных. Это объясняется тем, что предки млекопитающих жили в воде и дышали жабрами. Разумеется, жаберные мешки зародышей млекопитающих в ходе развития превращаются не в жабры, а в структуры, возникшие в ходе эволюции из жаберных щелей или стенок жаберных карманов, такие как евстахиевы трубы, среднее ухо, миндалины, паращитовидные железы и тимус.
У эмбрионов многих видов змей и безногих ящериц (например, веретеницы ломкой) развиваются зачатки конечностей, но затем они рассасываются. Аналогично, у китов, дельфинов и морских свиней нет задних конечностей, но у эмбрионов китообразных начинают расти задние ноги, развиваются кости, нервы, сосуды, а затем все эти ткани рассасываются.
Дарвин в качестве примера приводил наличие зубов у зародышей усатых китов.
У человеческих эмбрионов на ранних этапах развития (4-5 недель) есть заметный хвост с 10-12 развивающимися хвостовыми позвонками, длина которого составляет более 10% длины эмбриона. Развиваются не только позвонки, но и спинной мозг, и ряд других структур. К восьмой неделе шесть крайних позвонков отмирают в результате гибели клеток по механизму апоптоза. 5-й и 4-й хвостовые позвонки уменьшаются, прочие структуры также отмирают и уменьшаются. Макрофаги утилизируют разрушающиеся клетки. У взрослых людей остается 4 хвостовых позвонка, которые образуют копчик.
Молекулярно-генетические и биохимические доказательства
Дарвин опубликовал "Происхождение видов" почти за 100 лет до расшифровки структуры ДНК. Новые знания, полученные с тех пор, могли бы однозначно опровергнуть эволюционное учение, если бы оно было ложно. Вместо этого, анализ ДНК дает доказательства теории эволюции. Сам факт наличия наследственной изменчивости необходим для эволюции, и если бы оказалось, что ДНК устойчива к изменениям, это означало бы конец теории. Но ДНК постоянно мутирует, причем эти мутации соответствуют различиям между геномами разных видов. Например, основные отличия генома человека от генома шимпанзе включают 35 миллионов замен отдельных нуклеотидов, 5 миллионов делеций (удалений) и инсерций (вставок), слияние двух хромосом и девять хромосомных инверсий. Все эти мутации наблюдаются и сегодня, в противном случае версию об эволюционном происхождении от общего предка пришлось бы пересматривать.
Различия между геномами видов должны соответствовать не только наблюдаемым мутациям, но и филогенетическому дереву, и палеонтологической летописи. Подобно тому, как анализ ДНК может установить степень родства между двумя людьми, сравнение геномов позволяет выяснить степень родства между видами, а зная количество накопленных различий, исследователи определяют время расхождения двух видов, то есть время, когда жил их последний общий предок. Например, согласно данным палеонтологии общий предок человека и шимпанзе жил примерно 6 миллионов лет назад. Для того чтобы получилось наблюдаемое число различий между геномами, на каждый миллиард нуклеотидов должно было приходиться в среднем 20 изменений за одно поколение. Сегодня у людей скорость мутаций составляет 10-50 изменений на каждый миллиард нуклеотидов за одно поколение, то есть данные палеонтологии согласуются с результатами анализа ДНК, в строгом соответствии с теорией эволюции.
Биохимическое единство жизни
Носителем наследственной информации во всех клетках являются молекулы ДНК, у всех известных организмов в основе размножения - репликация этой молекулы. В ДНК всех организмов используются 4 нуклеотида (аденин, гуанин, тимин, цитозин), хотя в природе встречаются не менее 102 различных нуклеотидов. Кроме того, в природе встречается 390 различных аминокислот, но белки всех организмов составляются из одного и того же набора, в котором всего 22 аминокислоты. Кроме 20 стандартных аминокислот в редких случаях используются селеноцистеин и пирролизин. При этом возможно 1,4^1070 различных информационно эквивалентных генетических кодов, использующих те же самые кодоны и аминокислоты.
Если не учитывать эволюционное происхождение всех организмов от общего предка, то ничто не мешает каждому виду иметь собственный генетический код. Такое положение вещей было бы крайне выгодным, так как при этом исключалось бы преодоление вирусами межвидовых барьеров. Тем не менее, ничего подобного не наблюдается, и теория эволюции исключает такую возможность: изменения генетического кода приводят к изменению большинства белков организма, такая мутация практически всегда оказывается летальной, поэтому код не мог значительно измениться со времен последнего общего предка, что гарантирует его универсальность.
Ученые, которые внесли значительный вклад в установление структуры генетического кода в 1950-х и 1960-х годах, в своих исследованиях активно использовали предположение, что код практически универсален. Фрэнсис Крик, Сидней Бреннер, Георгий Гамов и другие сделали это предположение исходя из версии об эволюционном происхождении от общего предка, не имея никаких прямых доказательств универсальности кода. Полагаясь на универсальность кода, Бреннер в 1957 году пришел к выводу о неперекрываемости кода (один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов). Работа имела большое значение, так как до нее большинство исследователей считали код перекрывающимся.
В 1961 году, за пять лет до установления структуры генетического кода, Фрэнсис Крик сослался на работу Бреннера в статье "Общая природа генетического кода для белков". Основываясь на эволюционном предсказании об универсальности кода (в частности - на том, что у бактерий, табака и людей код устроен одинаково), Крик установил такие важнейшие свойства генетического кода как триплетность (значащей единицей кода является сочетание трех нуклеотидов - триплет, или кодон), неперекрываемость и наличие "стартовых" кодонов, с которых начинается чтение (трансляция).
Также можно отметить крайне похожие пути метаболизма в клетках всех организмов. Так, гликолиз у всех эукариот (организмов, в клетках которых содержится ядро) и у большинства прокариот (безъядерных организмов) проходит за 10 одинаковых шагов, в одной и той же последовательности, с использованием одних и тех же десяти ферментов (при том что возможны тысячи различных, но термодинамически эквивалентных путей гликолиза). У всех изученных видов основным переносчиком энергии в клетке является аденозинтрифосфат (АТФ), хотя эту роль могли бы выполнять сотни других молекул.
Вторая хромосома человека
У всех представителей семейства гоминид по 24 хромосомы, за исключением людей, у которых лишь 23 хромосомы. Человеческая вторая хромосома, является результатом слияния двух хромосом предков.
Доказательства слияния основываются на следующих фактах:
- Хромосома человека соответствует двум хромосомам обезьян. Ближайший человеческий родственник, бонобо, имеет практически идентичные находящимся во 2-й хромосоме человека последовательности ДНК, но они расположены на двух отдельных хромосомах. То же самое верно и для более дальних родственников: гориллы и орангутана.
- На хромосоме человека имеются рудиментарные центромеры. Обычно хромосома имеет только одну центромеру, но на длинном плече 2-й хромосомы наблюдаются остатки второй.
- На хромосоме человека имеются рудиментарные теломеры. Обычно теломеры находятся только на концах хромосомы, но последовательности нуклеотидов, характерные для теломер, наблюдаются еще и в середине 2-й хромосомы.
2-я хромосома, таким образом, представляет собой убедительное доказательство эволюционного происхождения людей и других обезьян от общего предка.
Эндогенные ретровирусы
Эндогенные ретровирусы представляют собой следы древних вирусных инфекций в ДНК. Ретровирусы (такие как ВИЧ и Т-лимфотропный вирус человека, вызывающий лейкоз и лимфому) встраивают собственный код в геном зараженного организма. Обычно после этого клетка начинает продуцировать новые копии вируса, но в этом процессе возможны сбои: копирование встроенной вирусной последовательности подавляется клеткой хозяина, но последовательность остается в структуре хромосомы. Если этот процесс произошел в стволовой половой клетке, то такую последовательность будут наследовать потомки инфицированной особи. Ретровирусы встраиваются в геном случайным образом, вероятность независимой встройки одинаковых вирусов на одинаковые позиции пренебрежимо мала. А значит, генетический код одного и того же ретровируса может присутствовать у двух животных на одной и той же позиции в ДНК только в том случае, если эти животные произошли от общего предка.
Около 1% человеческого генома занимают эндогенные ретровирусы, всего таких последовательностей в ДНК каждого человека около 30 000. Некоторые из этих ретровирусов встречаются только у человека. Другие последовательности встречаются только у шимпанзе и у человека (тем самым подтверждается происхождение человека и шимпанзе от одного предка). Также есть последовательности, встречающиеся у горилл, шимпанзе и человека, у орангутанов, горилл, шимпанзе и человека, и так далее. Распределение эндогенных ретровирусов в точности соответствует филогенетическому дереву.
Также можно привести пример из семейства кошачьих. У малых кошек (точнее - у таких животных как камышовый кот, европейская лесная кошка, степная кошка и домашняя кошка) найдена общая ретровирусная вставка. Ни у каких других хищников этот ретроген не обнаружен.
Биогеографические доказательства
Кактус астрофитум звездчатый из Техаса и молочай из Южной Африки. Два вида живут в сходных природных условиях и приобрели сходные формы за счет конвергентной эволюции. При этом они относятся не только к разным семействам, но и к разным порядкам. Несмотря на благоприятные условия, кактусы практически полностью отсутствуют в Африке.
Биогеография - наука, изучающая закономерности географического распространения животных и растений, а также характер фауны и флоры отдельных территорий.
Если два вида сравнительно недавно произошли от одной популяции, то они, как правило, обитают недалеко от ареала этой исходной популяции, а значит и недалеко друг от друга. Таким образом, географическое распределение видов должно быть совместимо с филогенетическим деревом. Если не принимать во внимание теорию эволюции, то разумно предположить, что виды живут в наиболее подходящих для них условиях. Теория эволюции же предсказывает, что должно быть много благоприятных для вида мест, в которых представители вида тем не менее отсутствуют, в связи с наличием географических барьеров.
Именно так дело и обстоит на практике. Среди млекопитающих Австралии преобладают сумчатые. Плацентарные млекопитающие представлены китообразными, ластоногими и рукокрылыми, которые могли перебраться в Австралию сравнительно легко, а также грызунами, которые появляются в палеонтологической летописи в миоцене, когда Австралия приблизилась к Новой Гвинее. При этом природные условия Австралии благоприятны и для других видов млекопитающих. Например, завезенные на континент кролики быстро размножились, широко расселились и продолжают вытеснять аборигенные виды. В Австралии и на Новой Гвинее, на юге Южной Америки и в Африке встречаются нелетающие бескилевые птицы, свистуны (зубастые жабы) и двоякодышащие, в других частях света они отсутствуют. Условия обитания в пустынях Африки, Америки и Австралии очень похожи, и растения из одной пустыни хорошо растут в другой. Тем не менее, кактусы были обнаружены только в Америке (за исключением одного, по всей видимости занесенного в Старый Свет перелетными птицами). Многие африканские и австралийские суккуленты (то есть растения, имеющие специальные ткани для запасания воды) внешне напоминают кактусы за счет конвергентной эволюции, но относятся к другим порядкам.
Морские обитатели восточных и западных берегов Южной Америки различны, за исключением некоторых моллюсков, ракообразных и иглокожих, но на противоположных берегах Панамского перешейка живет около 30% одних и тех же видов рыб, что объясняется недавним возникновением перешейка (около 3 миллионов лет назад). На большинстве океанических островов (то есть островов, которые никогда не были соединены с материком) отсутствуют наземные млекопитающие, земноводные и другие животные, не способные преодолевать значительные водные преграды. Видовой состав фауны таких островов беден и является результатом случайного заноса некоторых видов, обычно птиц, рептилий, насекомых.
Географическое распределение видов в прошлом, которое можно частично восстановить по ископаемым останкам, также должно соответствовать филогенетическому дереву. Теория дрейфа материков и теория эволюции позволяют предсказать, где следует искать те или иные ископаемые останки. Первые окаменелости сумчатых найдены в Северной Америке, их возраст составляет около 80 миллионов лет. 40 миллионов лет назад сумчатые уже были распространены в Южной Америке, но в Австралии, где они сейчас доминируют, сумчатые появились только около 30 миллионов лет назад. Теория эволюции предсказывает, что австралийские сумчатые произошли от американских. Согласно теории дрейфа материков, 30-40 миллионов лет назад Южная Америка и Австралия еще оставались частью Гондваны, крупного континента в южном полушарии, а между ними находилась будущая Антарктида. На основании двух теорий исследователи предсказали, что сумчатые мигрировали из Южной Америки в Австралию через Антарктиду 30-40 миллионов лет назад. Это предсказание сбылось: начиная с 1982 года на острове Сеймур, расположенном недалеко от Антарктиды, были найдены более десяти ископаемых сумчатых возрастом 35-40 миллионов лет.
Наиболее близкие родственники современных людей - гориллы и шимпанзе - обитают в Африке. Исходя из этого, в 1872 году Чарльз Дарвин предположил, что и древних предков человека следует искать в Африке. Многие исследователи, такие как Луис, Мэри и Ричард Лики, Раймонд Дарт и Роберт Брум, последовали совету Дарвина, и начиная с 1920-х годов в Африке было найдено множество промежуточных форм между человеком и человекообразными обезьянами. Если бы ископаемых австралопитеков обнаружили, например, в Австралии, а не в Африке, то представления об эволюции гоминид пришлось бы пересматривать.
Дарвиновы вьюрки
Во время кругосветного путешествия на корабле "Бигль" Чарльз Дарвин описал 13 видов вьюрков, обитающих на Галапагосских островах. Наблюдение за этими птицами натолкнуло Дарвина на идею происхождения видов за счет изменчивости и естественного отбора. Все галапагосские вьюрки происходят от общего предка, случайно попавшего сюда из Южной Америки. От предковой формы, питавшейся семенами, произошли три группы птиц: семяноядные земляные вьюрки, насекомоядные древесные вьюрки и славковые вьюрки, которые тоже питаются мелкими насекомыми. В результате приспособления к разным источникам пищи, вьюрки стали сильно отличаться друг от друга строением клюва. Три обычных вида земляных вьюрков - большой, средний и малый - встречаются на большинстве островов. В этом случае они хорошо различаются по размерам клюва и, соответственно, по величине предпочитаемых семян. На одном из островов обитает лишь средний земляной вьюрок, и здесь у птиц клювы меньше - в отсутствие конкурента средний земляной вьюрок занимает отчасти нишу малого вьюрка.
Примечания
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не "участок ствола". (Александр Марков. Ископаемые рыбы в очередной раз подтвердили правоту Дарвина.)
Карл Везе высказал гипотезу, согласно которой на первых этапах развития жизни, во время формирования первых клеток, основную роль играл горизонтальный перенос генов. В этом случае возможно, что последний универсальный общий предок представлял собой не единственную клетку, а группу примитивных клеток, свободно обменивающихся генетическим материалом.
Происхождение от общего предка - генетический процесс, в котором "будущее" не зависит от "прошлого" при известном "настоящем": эволюция потомков некоторой фиксированной популяции не зависит от того, какие предки были у этой исходной популяции. А значит, постепенное эволюционное происхождение от общего предка должно соответствовать таким моделям как марковские процессы и цепи Маркова. Используя эти модели, можно строго доказать, что ветвящиеся марковские системы образуют многоуровневые иерархические структуры. Поэтому биологи систематически используют цепи Маркова для моделирования эволюционных процессов, в числе которых - сложные генетические процессы, изменение частотного распределения фамилий со временем и поведение патогенных микроорганизмов при эпидемиях.