Раздел «Тема 6. Базовый механизм индивидуального поддержания жизнеспособности»
Модель произвольной адаптивности МВАПДоступ для всех
Темактика: Лекторий для «Модель произвольной адаптивности МВАП»
Лекторий школы «Модель произвольной адаптивности МВАП»
Тема 6. Базовый механизм индивидуального поддержания жизнеспособности
Назрел момент, чтобы сказать самое главное про МВАП. Это – не описание того, как природа замутила эволюцию разума и какими вариантами она разродилась при этом. Это – не описание природной реализации нейросети, - этим занимаются нейрофизиологи, собирая фактические данные. МВАП – каркас системы принципов в их взаимосвязи причин и следствий индивидуальной системы адаптивности, проработанный до некоторого уровня детализации ее узлов.
В природе нет ни одного одинакового нейрона так же, как нет ни одной одинаковой снежинки. И эти снежинки в зависимости от температуры, ветра, влажности и т.п. приобретают различимо разные виды форм. Одно дело – изучать разнообразие этих форм, а другое – общие закономерности их формирования. Если оставаться на уровне первого, то никогда не получится понять второе из-за невероятной сложности отдельных компонентов и разнообразия их взаимодействия.
Представим, что марсиане перехватили транспортный корабль, перевозящий реквизиты музея истории часов. Там, каждый экспонат хранился в одинаковых ящичках, но в каждом ящичке - очень разные часы, от древних ювелирных, инкрустированных каменьями, до современных изысков из драгоценного дерева, огромное разнообразие механических и электронных из разных.
Это было ни на что не похоже, и ценность находки не вызывала сомнения: наверняка здесь скрывалась величайшая тайна чужой цивилизации, сулящая наконец-то понять, где и как правильно нести яйца.
Марсианские ученые изощренным методом научного тыка, перепортив четверть всей добычи, научились запускать часы, но так и не смогли понять их назначение, ведь эти штуковины не совершали ничего определенно полезного. Есть ли у них вообще какое-то общее назначение, - просто совершенно разные изделия непонятно что делающие. Тайна еще больше интриговала и возникли невероятные по продуманности, изяществу и убедительности научные теории.
Все не могут думать одинаково, всегда кому-то чуть больше других повезет в том, что в его мозговом выступе шестеренки удачно сомкнутся на истине и он окажется правее всех других. Как только выскочка запалится, остальные, огорченные несправедливой неудачей, особо люто проявляют гражданский скептицизм.
И вот, один такой везунчик, решительно сметя из разума уже до предела истолченные скорлупки проблемы, вдруг увидел оставшееся: а, может быть, все дело лишь в том, что у всех штукенций за какое-то время что-то меняется при работе, иногда очень заметно на глаз, а что-то почти не заметно. Не веря в столь призрачные шансы, он собрал данные изменений во времени и ввел их в суперкомпьютер чтобы выявились похожести этих параметров. Получилось две очень выраженные, вездесущие корреляции изменений движений (для минут и часов): одна более слабая выпадала из общей картины (у тех штуковин, где не было секундной индикации) и остальные совсем слабые (недели, месяцы, годы).
Ученый тут же опубликовал свою работу, но изнеможенные неудачами коллеги его обсмеяли из-за неочевидности каких-то выводов и надуманности выбора параметра времени движений сопоставлений.
И тут в ложно-мозговой выступ марсианина тонкой струйкой закапала идея соотнести данные по параметру временной повторяемости. Тут же оказалось, что все виды корреляций с убедительной точностью не только относятся к одним и тем же интервалам времени, но и более длительные кратны целым числам более быстрых.
- Да это же просто как наше чувство времени! только примитивные и странные какие-то... - с отвращением проскрипел жевалами марсианин, протестующе не веря в такую простоту, но великая тайна растаяла бесследно. Потом он задумался о том, как написать итоговую работу так, чтобы его опять не высмеяли за столь вульгарную интерпретацию великой проблемы.
Те, кто не абстрагирует наиболее общее в явлениях, кто не имеет такого навыка, не может создать общую теорию, даже если им повезет с самовольными шестеренками в мозгах. Это тем сложнее, чем больше цепочка взаимосвязей таких общих составляющих в явлении.
С таким умением и даже с пониманием такой необходимости сегодня далеко не все хорошо. Когда-нибудь системология (и, в частности, системный анализ) обретет формы математического описания и методы, на которые каждый сможет опереться.
Для МВАП были сформулированы такие критерии: Критерии полноты и верности теории.
На этом занятии – очень много ключевых представлений, от которых зависит основа понимания всего остального. Эти представления были получены не просто при обобщении существующего массива фактических данных исследований, но, главное, из более общей картины каркаса модели совершенно бесспорных (эмпирически воспроизводимых) явлений, которые выделяются исследователями еще с середины прошлого века: безусловные, условные рефлексы, детали гомеостаза и т.п. (по тексту это будет видно).
Каждая конкретная природная реализация демонстрирует огромное разнообразие воплощений и механизмов одного и того же явления, причем это разнообразие не только по видам животных. но и индивидуальное (так, у человека >50 видов гемоглобина с одной системной функцией переносчика кислорода, вообще белки одного человека не совместимы с белками другого).
Буквально на любом уровне изучения организма физиологи озадачиваются этим разнообразием, не говоря про то, как странно и сложно бывает организованы сами механизмы. Видов синапсов – очень много и они очень разнообразны, а мы на занятиях говорили только про принцип синаптической передачи тока сигнала. Видов нейронов – очень много, а мы построили одну общую модель, которая отражает наиболее общую функциональность любых видов нейронов. Есть несинаптические случаи передачи возбуждения, которые мы вообще не рассматривали. Да и сам нейрон – невероятно неопределенная конструкция, в которой даже отдельные его дендриты способны начинать самостоятельно генерировать потенциалы действия, что, впрочем, на них и заканчивается если только не будет достигнуто условие генерации самого нейрона.
Природа – великий и бездумный рукожоп.
Исследователи строения глаза недоумевают о том, насколько нелогично, странно и нерационально устроена как оптика глаза, так и его чувствительная часть. Вот в фотоаппаратах все логично и правильно, там идеально и нет проблем, которые нейросети приходится решать, приспосабливаясь к столь капризному и неточному инструменту восприятия. Все можно сделать гораздо проще, точнее и лучше даже если использовать те же биоматериалы. И так – буквально во всех творениях природы. Мало того, все это постоянно кишит разными вариациями в каждой отдельной реализации. Зато глаз в воде продолжает функционировать, а фотоаппарат требует гидробокс. Если в глаз попала соринка, он проморгается, а соринку на матрице может вытащить только специалист и то не всегда без последствий. Видеокамеру связали шнурком с компьютером, тот запоминает все происходящее, но само по себе это - совершенно бессмысленно и только человек может разобрать этот смысл, а вот кошка уже – нет, если только по экрану не начнут бегать мышки.
Чтобы построить системную модель принципов усложнения механизмов адаптивности просто необходимо суметь абстрагироваться от всех вариантов воплощения этих механизмов. И такая картина может быть не понимаема теми, кто досконально изучает тонкости природной реализации. Они будут читать и скептически пожимать плечами: слишком все вульгарно описано, ну нельзя же все так упрощать. Таких поделок навалом в интернете и уже приелось на них реагировать. Но ведь и среди этих специалистов однозначно всеми принятых мнений по интерпретации данных исследований нет, а есть только многообразие представлений и растерянность в неподдающихся их пониманию теориях мозга.
Системную модель понимания строить нужно и, главное, возможно, другого пути нет если мы не хотим утонуть в неисчислимых загадках воплощения очередного примера природного рукожопства. Нам важна ясная система, но не выдуманная из ничего, а в которую четко и непротиворечиво укладываются все известные фактические явления организации адаптивных механизмов. А проявления таких механизмов можно подсмотреть только в природной реализации.
Пример: природный персептрон поражает своей прямолинейной и грубой логикой. Он никак не может соревноваться с точностью распознавания искусственного многослойного персептрона. Но логика идеального персептрона приводит к тупику и получению хотя и очень классного распознавателя хоть чего, но этим все и ограничивается.
Мы в системной модели понимания принимаем эту обобщенную природную логику, но не во всем ее разнообразии воплощений, а лишь о самом общем принципе реализации, который обеспечивает все последующие принципы своей основой. Этот принцип выверен из сопоставлений всех известных примеров природной реализации. Он не способен на эволюцию для каких-то невероятно новых условий, как все еще способен природный, но нам этого и не нужно.
Другой пример. Когда природа “изобрела” нейромедиаторы в простых животных, то еще не было задачи определять ими стили поведения, они просто тыкались случайно и бессистемно, но в результате очень хорошо отделились реакции разного вида, не мешая одна другой в общей схеме. Потом это привело к очень естественному способу переключения стилей поведения и к сопутствующей гормональной регуляции этих стилей.
Скрупулезному исследователю роли нейромедиаторов очень непросто сделать вывод о такой их функциональности потому, что он видит множество примеров совершенно иного использования, не укладывающего в такие представления, это навевает свои интересные выводы, требует учесть это в общей картине, и с этим нужно что-то делать, как-то объяснять.
Есть такой мысленный эксперимент. Если бы у бесконечно большой стаи обезьян были бы пишущие машинки и эти обезьяны все время бы клацали по буквам, печатая бессмысленный и случайный текст, то у большинства бы получалась просто галиматья, но у единиц бы выходили произведения великих писателей. Это – слишком сильный эксперимент, хотя и корректный. В природе хоть и очень много времени, но нет таких бесконечностей для того, чтобы случайно создать самолет или человека. Поэтому эксперимент стоит сделать более реальным. Пусть после того, как каждая обезьяна напечатает слово, оно бы проверялось на осмысленность богом, и если оно осмысленно, то можно печатать дальше, если нет - последнее удаляется. И тогда да, за какое-то не бесконечное время точно будут напечатаны все произведения прозы и даже совершенно новые, неизвестные.
У природы таким богом выступает смерть, которая не пропускает нежизнеспособное. И уже совсем не нужно бесконечности чтобы создать человека. Вот так природа и работает: как тупые бессмысленные обезьяны. И это стоит постоянно иметь в виду.
Вот почему обращаю особое внимание: при чтении нужно стараться выделять именно такие обобщенные принципы и механизмы, и в таком смысле рассматривать все огромное разнообразие особенностей реализации в публикациях фактических данных исследований. Я это выделяю всякий раз и у вас есть возможность верифицировать верность и обоснованность таких выделений. Это делать необходимо потому, что конечная модель должна возникать именно в вашей голове и она может оказаться в чем-то отличной от той, что сложилась в моей.
Пожалуйста, проникнитесь принципами системологии в статье об этом и специально следите за тем, чтобы вовремя видеть несистемные факторы в явлениях, которые мы рассматриваем, когда опираемся на фактические данные природной реализации. Это – самое важное, это – очень непросто, но без этого не возникнет целостный и непротиворечивый каркас собственной модели понимания.
Итак, новый уровень, поехали.
В общей картине представлений о принципах развития нейросети важнейшим является роль рецепторов – датчиков информации для работы регулирующей нейронной схемы управления эффекторами, что, в конечном счете, позволяет поддерживать организм в жизнеспособном состоянии. Важнейшим потому, что если рецепторы молчат, то и остальному делать нечего, что убедительно демонстрируют опыты по депривации восприятия, см. 1, 2, 3.
Рецепторные датчики не только обеспечивают восприятие внешнего мира, они возникли буквально во всем теле и обеспечивают жизненно-важные механизмы регуляции в организме. На прошлом занятии упоминалась метасимпатическая нервная система, но этим далеко не ограничивается управляющие контуры регуляции. И от самых простых взаимосвязей развиваются все более сложные, затрагивая уже имеющиеся связи головного мозга.
Везде, где только можно, после включения в геном такого варианта развития клетки как нейрон, он используется в качестве промежуточных распознавателей состояний предшествующих рецепторов (в том числе и предшествующих сформировавших специализацию нейронов) в организме. При этом последние для данного организма дополнения в геноме постоянно пытаются модифицироваться многими механизмами (мутация, половое смешение и др.) так что в популяции возникает довольно широкий спектр генетического разнообразия. Последние эволюционные участки генома, способные к модификации генетическими механизмами разнообразия, – это примерно 1-10%, и чем сложнее организм, тем этот процент меньше. Более древняя, хорошо обкатанная жизнью часть генома остается неизменной, если только ее не меняют паразиты или поломки (например, радиация или мутагенные химические вещества). Если затрагивается древняя часть, то получаются монстры, как правило, мало приспособленные к жизни уроды. Вот почему из кота не может вдруг родиться удав, а виды, у которых геном не совпадает на достаточно раннем этапе формирования тела, не способны к появлению потомства при спаривании.
Это напоминает то, как не модифицируются уже специализированные слои развивающегося мозга. И здесь регулятор – смерть неудачного.
В результате во всем организме возникают более молодые варианты нейронной регуляции на основе уже устоявшихся и мало подверженных изменениям. Так продолжается эволюция, способная адаптировать организм к меняющимся условиям и при этом все более усложняющая общую систему регуляции.
Гомеостаз
Для определения границы допустимого в отклонении различных параметров жизнеспособности существуют внутренние рецепторы значений таких параметров, в отличие от рецепторов, параметров внешней среды. Таких параметров очень много.
Так, общие для всего организма параметры, которые поддерживаются в оптимальном состоянии (меняющимся в зависимости от условий):
кислотность (крови, лимфы, межклеточной жидкости, желудка),
концентрация углекислого газа в крови,
концентрация кислорода в крови,
концентрация глюкозы в крови,
концентрация ионов натрия в крови,
концентрация ионов калия в крови,
концентрация ионов кальция в крови,
давление в гидравлике кровообращения,
осмотическое давление крови,
частота сердечных сокращений,
температура крови,
и др.
Изменение любого из этих параметров в любую сторону резко влияет на функциональность организма. Это прямо означает необходимость эффекторов, которые регулируют эти параметры с одной стороны и необходимость наличия распознавания оптимального значения параметров и направления выхода их из оптимума (превышение или недостаток). Эти распознаватели должны учитывать, что в разных режимах работы организма нужен свой оптимум. А такие режимы переключаются в зависимости от стиля реагирования организма (пассивный, высокая физическая нагрузка, половое поведение, защитная пассивность, сон и т.п.), а также в зависимости от внешних условий: холод, жара, обезвоживание, голод, проблемы с воздухом и т.п.
Обкатанные жизнью системы регуляции основных параметров оказываются тем сложнее, чем для большего числа возможных стилей поведения и числа различных внешних условий и ситуаций они наработаны.
Понятно, что самые простейшие связи типа понизилась температура – дать больший сигнал на ее повышение, как в термостатах, это – лишь первый уровень такой регуляции, на который наслаиваются более сложные, учитывающие ту или иную специфику. И таких слоев оказывается немало даже еще до условно-рефлекторного уровня регуляции, до уровня развития ассоциативной теменной коры мозга, где формируются условные рефлексы. Эти модификации относятся ко вторичным зонам мозга (первичны развиваются сами, без модификации) и такая локализация очень важна для дальнейшего развития механизмов.
Точно так же, как возникают все более сложные слои распознавателей разных видов внешней рецепции, каждый из которых имеет свой уровень связей с эффекторами, обеспечивающий свою сложность реагирования, так же происходит и с внутренними рецепторами отклонения параметров гомеостаза и возврата их в норму.
Принципы поддержания оптимального уровня параметра
У простых термостатов (устройств для поддержания постоянно температуры в среде) есть некоторый гистерезис: включение нагревателя вызывает перегрев пока на это отреагирует датчик потому как у нагревателя есть инерция (он не может сразу нагреться и сразу остыть). Поэтому температура в таких термостатах постоянно колеблется, т.е. возникает генерация колебаний температуры. И чем больше инерция нагревателя или других элементов отрицательной обратной связи, тем больше будет амплитуда колебаний из-за того, что отрицательная связь все меньше попадает в нужное время и вместо противодействия возникает содействие. Т.е. при сдвиге фазы (момента начала) выходного сигнала из-за задержки по отношению ко входному отрицательная обратная связь становится все более положительной. Если задержка окажется настолько большой, что вместо отрицательной обратной связи возникнет положительная (т.е. момент воздействия окажется противоположным тому, что нужно бы для компенсации), то колебания станут максимальными по амплитуде вплоть до достижения предельного режима переключений: с одного предельного состояния в другое.
Но если сделать некий сглаживающий фильтр (распространенное схемотехническое понятие), например на выходе термометра, который не позволяет быстро меняться показаниям, а отслеживает некий средний уровень по времени, заведомо больший, чем период колебаний температуры (это можно сделать простейшей цепочкой из резистора и подключенного к нему заземленного конденсатора, и сглаживать он будет все, что имеет время изменения меньше, чем произведение сопротивления на емкость = постоянная время цепочки), то колебания прекратятся и установится равновесное состояние на заданной температуре.
Было описано наиболее общее условие генерации колебаний в системах с обратной связью, в независимости от природы колебаний.
В организме оно так же работает. Например, нужно удержать руку в каком-то положении, делается усилие поднять ее. Это усилие зафиксируют рецепторы положения руки (или зрительные рецепторы через все системы мозга, распознающие это положение) и когда рука достигнет нужной высоты, дается команда на уменьшение усилий до уровня сохранения нужной высоты. Но если есть какие-то задержки в реагировании, то момент нужного уровня будет пропущен и нужно будет опускать руку. Возникнет дрожание (тремор). При сильном волнении, когда на слежение за плавностью движения привлекается максимум внимания и движение контролируется буквально на всех стадиях, часто возникает тремор потому, что осмысление требует времени и возникает задержка. Это может закрепиться в автоматизмах. Или тремор может возникнуть из-за патологии каких-то промежуточных стадий, например, прекращения действия некоторых рецепторов положения руки и тогда момент нужного положения будет пропущен.
В быту люди с повышенным давлением склонны принимать сбивающий давление препарат, когда давление поднимается. А это бывает при стрессе, физической нагрузке и т.п. ситуациях, когда организму нужно поднять давление для обеспечения функциональности. В результате давление довольно быстро снижается и даже намного ниже, чем нужно. Возникает болезненная вялость, сонливость, нарушения координации, в общем, давления организму очень не хватает. Тогда гипертоник принимает препарат для повышения давления (пьет родиолу, крепкий чай и т.п.). Давление подскакивает. И т.п.
Если же не торопиться с решением противодействовать, а выждать время, то вообще бы не потребовалось принимать препараты потому, как последствия стресса довольно скоро бы закончились.
Бывает при сильной жажде так хочется пить, что выпивается огромное количество воды. Сигналы о восстановлении водного дефицита возникают намного позднее, когда на борт заливается слишком много воды. Это может нанести определенный вред (как с лекарствами от давления) и это стоит учитывать, не торопясь выпить побольше до того, как жажда не исчезнет.
Во всех случаях подобного восполнения жизненных параметров стоит помнить об этом и не переедать до вредных последствий, не перепивать, не переохлаждаться в жару и т.д.
Пожалуйста, прочтите внимательно статью: Гомеостаз и система значимости.
Гомеостаз органов
Кроме поддержания стабильности внутренней среды, физиологи различают две системы регуляции параметров отдельных органов и систем: симпатическая и парасимпатическая системы с взаимно противоположными видами воздействия на параметр, обеспечивающими его необходимый уровень поддержания:
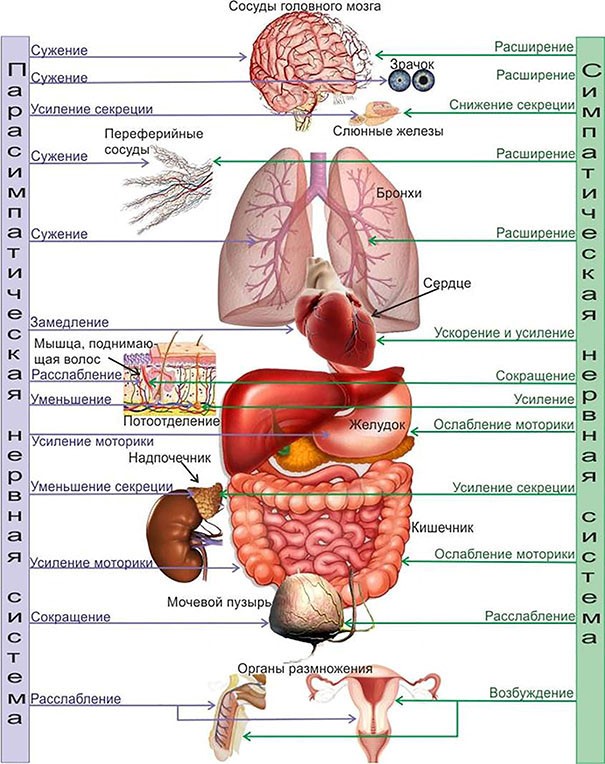
Такой вид регуляции на основе обратной связи с эффекторами противоположного действия применятся везде в организме, где необходимо иметь возможность быстро менять значение параметра. Это – мышцы-антагонисты, регуляция состояния органов и внутренней среды. К этому вынуждает необходимость адаптации. Если что-то не вызывает фатальное отставание, то оно может регулироваться просто включением и ожиданием естественного восстановления после отключения.
Отклонение жизненных параметров организма и отдельных органов опасно, и распознавание таких ситуаций важно для того, чтобы их избегать. Поэтому кроме распознавателей отклонения жизненных параметров, которые используются в контурах регулирования параметров, эти же распознаватели на последующем слое развития нейронов в этой области, могут использоваться как рецепторы опасного состояния, а также рецепторы восстановления после опасности.
Здесь так же, как в гомеостатических эффекторах-антагонистах, нужно при возникновении опасности активировать защиту от нее, но только до тех пор, пока опасность не минует чтобы прекратить эту защитную реакцию. Восстановление параметров сигнализирует о таком моменте.
Понятно, что распознаватели опасных состояний организма важнее, чем распознаватели восстановления (они первичны в логике происходящего) и они представительствуют в мозге гораздо больше потому, что во многих случаях обратной связи в такой регуляции не требуется и после возврата в норму ничего не требуется совершать.
Это также касается всех случав реакций избегания опасности, когда распознаются ситуации, могущие навредить на основе предыдущего опыта. Для таких реакций рецепторами являются слои распознавателей опасности, которые, в свою очередь, используют в качестве рецепторов слои распознавателей отклонения гомеостаза организмов и органов.
Система значимости
Так возникают немало все более усложняющихся уровней распознавателей образов негативного и позитивного для организма или распознавателей того, что данное сочетание входных признаков значит для жизни организма.
Такой функцией распознавания значимости не обладают сами по себе распознаватели всех других сенсорных систем: никакие образы восприятия сами по себе ничего не означают для организма в последствиях их появления. Но, сочетаясь в общий образ, где присутствуют рецепторы как различных видов рецепции и рецепторы значимости, так начальные эффекторы цепочек ответных реакций, возникают связанные между собой (ассоциированные) образы, обладающие значимостью того, что данный образ в данных условиях означает для сопутствующего состояния организма. Это происходит, начиная со вторичных зон и, особенно конкретно (специфично к какому-то распознанному образу) - в области формирования условных рефлексов, в теменной ассоциативной коре мозга, где сходятся все виды рецепции, включая рецепцию состояния гомеостаза.
Мы будем называть такие образы в ассоциативной теменной коре образами восприятия-значимости-действия (или просто образами), а систему распознавателей опасного отклонения параметров гомеостаза и восстановления их – системой гомеостатической значимости. В дальнейшем она будет дополняться более высокоуровневыми механизмами распознавания значимости происходящего для организма, образуя, то, что называют смыслом происходящего для субъекта.
Стоит заметить, что система значимости и система регуляции гомеостаза, хотя и основывается на одних базовых распознавателях и первичных рецепторах состояния параметров гомеостаза, но это – две разные системы с разными функциями. Это должно быть очевидным из уже сказанного.
Значимость образов позволяет организовывать реагирование на внешние условия и раздражители с учетом того, как это отражается на жизненном состоянии организма.
Именно это в ассоциативных зонах играет роль “учителя” в том, как оценивается верность распознавания ситуации в данных условиях, но здесь не происходит формирование распознавателей с коррекцией от учителя, а возникает лишь возможность такой коррекции в более сложных механизмах (самые простые из которых будут описываться через занятие).
Эта роль значимости организуется совсем не так, как в искусственных нейросетях, хотя там уже и начали использовать искусственную систему подкрепления, но она не отражает собственное состояние сети и параметры ее существования и поэтому не является для нее адаптивной в этом смысле.
Дополним таблицу различий природной и искусственной нейросетей:
|
Искусственная нейросеть |
Природная нейросеть мозга |
|
Критерием верности функции распознавания является привнесенные оператором условия, а не состояние самой сети. Во всей сети все ее элементы корректируются в соответствии с этим конечными условиями. |
Критерием верности распознавания образов восприятия-значимости-действия является сигналы специализированных рецепторов состояния гомеостаза и их интерпретация более высокоуровневыми рецепторами системы значимости. До ассоциативной зоны распознаватели формируются без учителя. |
Это – последнее такое различие потому, что далее, в сторону усложнения механизмов адаптации, уже не будет вообще никаких возможных сопоставлений.
Так, для организации обратных связей, обеспечивающих точное управление мышечными реакциями, используются специальные рецепторы положения и состояния мышечных волокон, называемые пропреорецепторами (они же дают сигналы о состоянии кожи, связок и суставных сумок).
С их помощью последовательность мышечных действий организуется поочередным срабатыванием управляющих звеньев цепочек программы движения, что в схемотехнике моделируется сдвиговым регистром, но, в отличие от него, дополненным синхронизирующими сигналами проприорецепторов.
С момента использования распознавателей значимости для возможностей определения направленности реагирования схема приобретает новое качество индивидуальной адаптивности по сравнению с адаптивностью, определяемой генетическим кодом потому, что начинает учитываться текущее состояние организма, что позволяет организовывать связи между теми условиями, которые оказывают влияние на это состояние и реакциями, нормализующими это состояние.
Конечно, уже и генетический код дает возможность учитывать условия и формировать нейросеть в зависимости от того, были ли нет условия, вызывающие ту или иную экспрессию генов, но теперь изменения реагирования происходят не из поколения в поколение, регулируемые смертью неудачных форм, а с учетом текущей ситуации.
Гомеостатическая значимость формируется так же, как и другие первичные и вторичные зоны мозга, образуя все более сложные примитивы того, что для организма хорошо, а что – плохо.
В третичной (ассоциативной) зоне, где сходятся все выходы разных сенсорных каналов распознавателей, возникают уже сложные образы, составленные из многих видов рецепторного восприятия.
Образы оказываются ассоциированы, в первую очередь, с отрицательным состоянием организма (опасный выход каких-то параметров гомеостаза из нормы), или ассоциированы с положительным состоянием возврата из аварийного состояния жизненных параметров в норму. По логике ясно, что не может быть возврата в норму до того, как параметр вышел из нормы. Так что состояние негативной значимости всегда первично.
Однако, возбудить распознаватель возврата в норму (т.е. детектора, что с организмом все хорошо) бывает возможно возбудить разными химическими и физическими воздействиями, и тогда организм получает ложный сигнал “хорошо” в то время, как нет никаких таких причин и обстоятельств, которым нужно было бы противодействовать. Именно так были первоначально открыты точки мозга, где сконцентрированы распознаватели “хорошо” и “плохо”. НО стоит иметь в виду, что гомеостатическая значимость вовсе не означает субъективные переживания, что возврат жизненных параметров в норму не является переживанием “хорошо”, каким-то кайфом при раздражении таких распознавателей. Кайф возникает только при осознании этого состояния, и для переживания необходимо иметь дополнительные механизмы осознания, до которых мы еще не добрались в занятиях.
Эти участки были названы “эмоциональными центрами” потому, что их возбуждение приводит к переключению стиля поведения животного (в том числе человека), характерного для проявления какой-то эмоции. Но это – всего лишь тупо – распознаватели отклонения и возврата в норму, а не “чувства”, и понимание этого – чрезвычайно важно.
Стоит ознакомится с подборкой фактических данных исследований по теме системы значимости.
В плане эмоций и в самом деле, возбуждение распознавателей значимости приводят к осознанному переживанию тех или иных градаций состояния хорошо и плохо. Но такие переживания возможны только у животных, обладающих способностью осознавать и субъективно переживать, а что это такое – тема последующих занятий. У сложно организованных насекомых есть распознаватели состояния параметров гомеостаза, но нет механизмов субъективного осмысления, представленных совершенно определенными анатомическими образованиями, обеспечивающими высшие механизмы (по сравнению с рефлексами) индивидуальной адаптации к новому. Сейчас об этом преждевременно сказано для того, чтобы задать контекст понимания, что первичные связи и даже условные рефлексы (тема следующего занятия) – далеко не исчерпывают механизмы индивидуальной адаптации, а лишь представляют собой их базовую половину.
Самый важный принцип использования распознавателей значимости – возможность переключения вида реагирования в зависимости от условий и обстоятельств и формирование ответных реакций именно в этом контексте.
Можно выделить такие основные виды реагирования как пищевое поведение, половое поведение, защитное поведение (активная и пассивная защита), исследовательское поведение, сон. У каждого такого стиля должен быть инициирующий момент необходимости его активации (выход параметров гомеостаза из нормы) и стадия возврата гомеостаза в норму, что характеризует успешность варианта поведения (или неуспешность, если параметр еще более отклонится от нормы).
Половое поведение инициируется самим организмом, это - особый случай, когда выход параметров из нормы организован как механизм выживания вида. Поэтому с половым поведением связана возможность его возбуждения даже если гомеостаз в полном порядке и нет аварийного состояния, т.е. возможна его самоактивация (еще одно исключение – альтруизм, который так же есть у многих видов животных). Впрочем, есть возможность искусственной активации и любой другой потребности, скажем, воздержание от воды или еды чтобы потом получить удовольствие от восполнения дефицита. Есть анекдот, где медведь вдруг увидел, как заяц бьет себя кувалдой по яйцам и вопит на весь лес, а на вопрос медведя нашел силы ответить, что когда промахнется это – неимоверный кайф.
Каждый базовый стиль имеет более сложные градации его особенностей, обеспечиваемых последующими слоями распознавателей значимости, конкретизирующими распознанную ситуацию. Это позволяет разделять контексты реагирования.
Контексты значимости
Базовые стили поведения переключаются очень быстро с помощью изменения баланса нейромедиаторов в мозге так, что те цепи, которые не рассчитаны на использование синапсов, работающих через такие нейромедиаторы, перестают работать, а действуют только специфичные к имеющимся нейромедиаторам связи (анимация). Так выделяются целые программы ответных действий (анимация).
В теле при этом одновременно выделяются специфичные вещества – гормоны, которые изменяют режимы работы органов, оптимально приспосабливая их к данному стилю поведения: если нужны предельные усилия, то повышается частота сердечных сокращений и их сила, расширятся сосуды, становится интенсивнее дыхание, высвобождаются энергетические вещества и т.п.
Нормы гомеостаза соответственно меняются, что учитывается распознавателями значимости.
Таким образом формируется общий контекст восприятия и реакций организма, в котором оказывается возможным только присущий ему стиль поведения.
Все это – механизмы, предопределяемые наследственно. Но число вариантов реальных ситуаций таково, что они не могут быть обеспечены ограниченным набором нейромедиаторов и требуют более оперативного изменения поведенческих реакций в условиях, не предусмотренных наследственно. Т.е. для большей гибкости реагирования необходим механизм, который бы мог переключать более детализовано контексты реагирования.
По схемотехнической логике это реализовать просто: вместо того, чтобы отключать питание у схем, обеспечивающих все стили реагирования кроме нужного, можно просто ввести блокировочные сигналы или, наоборот, сигналы необходимости возбуждения данного контекста для распознавателей ситуации.
В природе реализованы оба варианта: активация распознавателя данного контекста со стороны иерархии слоев значимости и латеральное торможение между конкурирующими соседними контекстами.
Получается такая картина: распознаватели жизненных потребностей (при отклонении параметров гомеостаза) активируют специализированный контекст, а сигналы рецепторов внешней ситуации определяют стимулы для активации тех или иных реакций в этом контексте. Каждый образ активируется во вполне определенных условиях окружения при наличии “пускового стимула”.
Новые связи могут образовываться только в активном контексте, но не в других. Научившись танцевать у себя дома, не получится сразу так же хорошо продемонстрировать это на балу.
Это приводит к тому, что образ одних и тех же признаков восприятия приобретает совершенно разную значимость (ассоциируясь с признаками значимости) в разных контекстах. Соответственно, реакции в разных контекстах оказываются неповторимо специфического стиля так, что животное как бы приобретает разные свойства, подчас противоположные, в разных своих состояниях. Люди могут быть неузнаваемо разными в разных ситуациях.
Любая деятельность и ее навыки развиваются только для определенного контекста, в котором возникает понимание ситуации, восприятие ее особенностей и выбор действий, прогнозирующих желательный результат. Поэтому понадобились механизмы, отзеркаливающие навыки одних контекстов для использования в других и не только своих контекстов, но и внешне наблюдаемые проявления чужих и вообще потребовалось наблюдение с формированием модели причин и следствий наблюдаемого. Об этом – позже.
Рефлексы, приобретенные в одних условиях, оказываются неприменимыми для других. И тут не поможет механизм, когда, скажем, линии разной толщины и наклона все равно идентифицируются как линия потому, что простейшие примитивы достаточно схожи и детектируются рядом соседних распознавателей, так что свести их все по ИЛИ в общий распознаватель механически просто. Но распознать линию в карандаше, ребре бумаги, кривую линию на рисунке и т.п. на этом уровне принципиально невозможно. Для этого нужен совершенно иной принцип организации контекстов: по смыслу воспринимаемого, а не по его форме и других сенсорных характеристиках в контексте значимости. Этот механизм реализован в иных по локализации слоях мозга по отношению к тем, где формируются условные рефлексы, до которых еще нужно добраться в наших занятиях.
Понятие контекстов значимости – базовое для всех последующих занятий. В общем смысле контекст – ограничение восприятия и возможности создавать новые реакции механизмом, игнорирующим все в восприятии, что не относится к данному контексту значимости состояния организма. Наиболее широкий, общий контекст – состояние аварийное отклонение какого-либо параметра гомеостаза (аварийное состояние) – контекст общей негативной значимости и состояние возврата к норме – контекст позитивной значимости.
Более частные, вложенные контексты ограничивают все вне пищевого поведения, оборонительного поведения, полового поведения и т.п.
Еще более частные контексты – уточнение стиля поведения базовых контекстов. Наиболее узкий частный контекст – условия формирования конкретной условно-рефлекторной реакции.
И здесь – особенно много вариабельности природной реализации. Так, кроме иерархии базовых контекстов значимости есть фоновый механизм “сторожевых реакций” (“старт-реакции”), который работает вне контекстов значимости. Мы его не будем рассматривать.
В теме 4 был вопрос: “Нарисуйте в свободном виде нейросеть управления втягиванием тела улитки в раковину при прикосновении к усикам (на схеме изобразить один рецептор касания усика), при попадании острого вещества на кожу (тоже один рецептор), но не втягивать при ползании (рецептор чувствительности на основании улитки)”.
Во всех присланных ответах верно было показано, что при движении прикосновения игнорируются и для этого было предложено тормозить их в таких условиях. Можно рассматривать движение как контекст значимости движения (у него есть какие-то гомеостатические причины, вплоть до поиска еды). И тогда можно не тормозить сигналы от рецепторов прикосновения, а просто игнорировать отсутствием достаточного подвозбуждения или даже отсутствием нужного для оборонительной реакции баланса медиаторов.
Прогноз
То, что многие образы восприятия несут с собой уже имеющуюся ассоциацию с позитивным и негативным результатом появления такого образа, позволяет использовать их как прогноз, активируя ту или иную значимость, связанную с ним. И так – со всеми образами, составляющими последовательность цепочек программы реагирования. Поэтому если подсмотреть, какая значимость связана с последним звеном, можно узнать заранее, чем может закончится такая реакция и предпринять меры, если конец плох. Правда, для этого нужен механизм произвольности подсмотра.
Для этого цепочки реакций оборудованы средствами отсечения тормозных связей, блокирующих активность последующего звена, пока не сработает предыдущее и будет сигнал о законченной фазе движения (которые унаследовали эту возможность из торможения во время сна всего из восприятия и реакций). Это соответствует вопросу в вопроснике: “Что произойдет в нейронной схеме организации цепочки действий если во всех элементах после первого убрать тормозные нейроны, а первый элемент сработал?”. Произойдет мгновенный доступ к значимости конечного звена, то, что И.Павлов назвал “опережающим возбуждением” анимация и еще.
Хотя на уровне рефлекторных реакций такой прогноз не используется, но уже есть задел.
Активность возбужденных образов, совпадающая с активностью базовых элементов системы значимости, позволяет укреплять связи от ассоциативных зон к первичным зонам системы значимости. Такая связь позволяет устанавливать более общий контекст в зависимости от воспринятого образа. Увидел льва далеко – прятаться, близко – бежать, совсем близко – паника от отсутствия заготовленных реакций или бешеная агрессия. Все это пока еще - не осознанная оценка и выбор поведения, а тупые рефлекторные реакции, возникшие в условиях данных обстоятельств. Т.е. связь локализуется около нейронов образов в ассоциативной коре. И эта связь должна иметь возможность обрываться, что и поделывают переключатели специализированного органа мозга – гиппокампа.
Это – очень важная особенность – возможность влияния от более поздних структур к более ранним, реализуется во всех зонах мозга из-за наследственно предопределенного принципа образования двунаправленных связей (т.е. посылка роста аксонов в обратном направлении). Эта связь не участвует в функции распознавания примитива восприятия, а только коммутирует его возможность работать или молчать, обеспечивая контекст восприятия. Т.е. наличие активности такой связи оказывает подвозбуждающее воздействие, на фоне которого специализируется данный распознаватель. Без него профиль рецепторов не возбудит распознаватель потому, что у него не хватит возбуждающего потенциала. А с ним все так же хорошо контактируется тормозным влиянием соседей в четкий образ. В тех зонах мозга, где организовано это, распознаватели подвозбуждаются текущим контекстом системы значимости (а не последующим образом, которого при формировании распознавателя еще нет и в помине).
Так называемые активирующие и тормозное влияние – базовая фишка, точнее, фича организации мозга.
Это происходит частично в первичных зонах и, в основном, в последующих вторичных зонах с влиянием распознавателей системы значимости, формирующей контекст (см. 0. 1. 2).
Эти связи помогают выделять стимулы в текущем контексте значимости и удерживать их. Нейрофизиологи из Университета Карнеги — Меллона (США) сумели подсмотреть, как мозг это делает:
“Информация из зрительной коры поступает в теменную, и если там какой-то объект будет признан особо важным, то соответствующий участок теменной коры усилит подведомственный участок коры зрительной — и зрительное внимание сосредоточится на этом объекте.”.
В третичной зоне рецепторы гомеостаза (в ипостаси нейронов-распознавателей особенностей сочетания признаков отклонения параметров) не образует контекст, а непосредственно придает определенную значимость данному образу, точнее это есть предельно узкий, пока еще вырожденный контекст для его расширения в случае ветвления от этого образа других реакций.
Как уже говорилось, после развития слоев мозга, они утрачивают возможность образования там новых связей. Поэтому в первичных и вторичных зонах остаются только те связи с другими областями мозга, которые были активны во время их специализации. Понятно, что третичных зон еще не было и связи от них поэтому быть не может. А вот подкорка уже была активна.
Я приношу извинения за некоторую сумбурность последних абзацев, затрудняющую понимание, но дело в том, что вынужден втиснуть сюда сразу несколько принципов, которые еще толком не рассматривались. Надеюсь, при итерации все будет пониматься лучше. Альтернатива – очень занудное и долгое рассмотрение этих принципов и потом сведение в общую картину как было сейчас сделано. Ни я, ни участники, итак напряженные сложностью раскрывающейся картины, к этому пока не готовы.
Если есть возможность связи от образа в ассоциативной коре к базовым (и не только базовым) распознавателям значимости, от которых образует контекст восприятия, выделяющий предшествующим примитивы восприятия на уровне вторичной коры, то при замыкании такой связи образуется кольцо: активный образ поддерживает активность распознавателей значимости, которые обеспечивают активность этого образа и такая циркуляция возбуждения будет продолжаться пока ее не прервет что-то.
Возникает вопрос: а как может активность всего лишь подвозбуждающей силы значимости активировать образ во вторичной зоне мозга? Схемотехнически это реализовать довольно просто, переключая силу подвозбуждения от фоновой до активирующей. Как именно это проделывается в природе точно не ясно. Есть свойство нейрона временно облегчать свое возбуждение, если только что он был возбужден. Но есть совершенно достоверные факты исследований, что именно такие зацикленные связи удерживают стимул в активном состоянии. А.Иваницкий несколько преждевременно полагал, что это – и есть субъективное переживание, элемент мысли, но это – всего лишь одна из основ организации субъективности.
Если бы взгляд был постоянно прикован к изображению, то нейрон активного распознавателя быстро бы истощался постоянной генерацией. Но зрачок постоянно скачет на небольшие расстояния (стаккады), расширяя зону наиболее четкой своей оптики, а распознаватели генерируют не постоянную серию импульсов, а короткие сигналы детекции образа, которые удерживаются на уровне вторичной коры и поддерживающего контекста локального внимания от системы значимости. Можно рассматривать все вокруг, а следы распознавания образов будут продолжать воздействовать в данном контексте на реакции. При циркуляции активности по кольцу нейроны вполне успевают восстановиться и не происходит их истощение (анимация). Но с накоплением числа таких закольцовок общая локальная область мозга начинает испытывать проблемы регенерации энергетики и продуктов метаболизма. Кроме того, при этом возникают множественные активности, мешающие новым более слабым за счет бокового торможения (острота и концентрация внимания снижается) вплоть до настолько полного забивания всех каналов, что становится невозможной нормальная гомеостатическая регуляция. При длительном лишении сна происходит ухудшение возможности реагирования вплоть до смерти из-за множественной разрегуляции параметров при, казалось бы, полной исправности регулирующих структур мозга.
В искусственной зрительной системе не нужны были бы стаккады глазных яблок т.к. ничто бы не истощалось, но поле зрения все равно ограничено и так же возникает проблема удержания образов в активном контексте.
Дополнительно:
От простых схем управления телом - к контекстным
Видео: