Биосенсоры органов чувств Ф.Г. Грибакин
Феликс Гурьевич Грибакин, доктор биологических наук, профессор, заведующий лабораторией эволюции органов чувств Института эволюционной физиологии и биохимии им. И.М.Сеченова РАН. Научные интересы связаны с сенсорной физиологией, главным образом с физиологией органов чувств насекомых. Автор 120 печатных работ, в том числе книги «Механизмы фоторецепции насекомых» (Л., 1981). Лауреат Государственной премии СССР (присуждена в 1987 г. авторскому коллективу за цикл работ «Физиология органов чувств насекомых»).
Как известно, знаменитую янтарную комнату в Царскосельском дворце украшали флорентийские мозаики, изображавшие в аллегорической форме пять человеческих чувств: зрение, слух, вкус, обоняние и осязание. Из четырех мозаик пока нашли только одну — «Осязание и обоняние»... Но цель автора — рассказ не о дальнейших поисках пропавших мозаик, а об изучении тех чувств, которые отображены в аллегориях. Речь пойдет в основном только о естественных биосенсорах — входных устройствах сенсорных систем, т.е. тех отделах нервной системы, которые обеспечивают организм информацией об изменениях во внешней среде.Сенсорные рецепторы и системы
Критически настроенный читатель несомненно упрекнет автора в терминологическом эклектизме. Действительно, словосочетание «биосенсоры органов чувств», интуитивно воспринимаемое неспециалистом как тавтология, режет слух, колет глаз, и раздражает чутье физиолога, привычно использующего латинизированный термин «сенсорные системы», как-то незаметно пришедший на смену «анализаторам» И.П.Павлова.
Как же соотносятся понятия «органы чувств», «сенсорные системы» и «биосенсоры»? Несомненно, у человека, да и всех позвоночных животных, глаз — это орган зрения, ухо — слуха, нос — обоняния, а язык — вкуса. Однако человека по его сенсорному оснащению вовсе не следует считать венцом творения: он не видит ультрафиолетового излучения и поляризованного света, как насекомые; не слышит ультразвук и не пользуется им для эхолокации, как летучие мыши и дельфины; а как «выглядит», например, образ океанской электромагнитной погоды (!) для рыбы, обладающей системой электрорецепторов, человек даже и представить себе не может, так как не имеет нужных биосенсоров. Да и наши антропоморфные обозначения органов чувств часто несостоятельны по отношению к беспозвоночным животным, явно превосходящим в биосфере по числу видов. Насекомые, например, лишены носа в нашем понимании, хотя чувствительность их хемосенсорных систем с обонятельными рецепторами на антеннах не уступает остроте обоняния наиболее способных к этому позвоночных. Однако миниатюрный мозг насекомого не в состоянии различать и идентифицировать такое громадное количество запахов, как, скажем, на много порядков более мощный мозг («нос») собаки или квалифицированного парфюмера.
Эти простые рассуждения уже позволяют нам понимать под биосенсорами те датчики, которые организм выносит на периферию нервной системы, чтобы получать информацию о процессах, происходящих во внешней среде. Термин «биосенсоры» — скорее технический, под ним подразумеваются прежде всего создаваемые современной биотехнологией искусственные датчики, или сенсоры, на основе биомолекул. Для физиологических наук естественные биосенсоры — это просто рецепторные, или сенсорные, клетки, располагающиеся в органах чувств и предназначенные для решения сложной диалектической задачи: изменяться под влиянием внешнего сигнала (иначе он не дойдет до нервной системы), оставаясь, однако, самими собой даже при очень сильных воздействиях. В качестве семантического курьеза стоит отметить, что термин «рецепторы», которым обозначали рецепторные клетки органов чувств в 60—70-е годы, постепенно взяли на вооружение специалисты по молекулярной биологии. В настоящее время под словом «рецептор» они однозначно подразумевают белковую молекулу, избирательно взаимодействующую с гормоном, нейромедиатором или феромоном, т.е. с сигнальным веществом. В сенсорной физиологии принято говорить о «рецепторных клетках», или, что то же самое, о «сенсорных рецепторах».
Что же касается понятия «сенсорные системы», то оно включает в себя не только периферически расположенные биосенсоры, но и всю систему обработки передаваемых ими сигналов, т.е. мозг. Например, таким образом оптические биосенсоры, или фоторецепторы, превращают оптическое изображение в нейроизображение. Чтобы почувствовать, как наш собственный мозг видит и обрабатывает информацию, заключенную в нейроизображении, поставим простой эксперимент с последовательными образами (этот опыт много лет назад продемонстрировали сотрудники Института проблем передачи информации РАН Г.М.Зенкин и А.П.Петров). Войдем в хорошо затемненную комнату и адаптируемся к темноте в течение 10—15 мин. Затем возьмем в правую руку заранее приготовленную импульсную фотовспышку, направим ее на ближайший предмет — вначале на собственную свободную руку — и нажмем спусковую кнопку. Вспышка длительностью около миллисекунды давно кончилась, свет уже не действует на нашу сетчатку, но мы четко видим... нейроизображение собственной руки. Уберем ее за спину: где же она на самом деле? Глаз (точнее зрительная система!) говорит, что рука перед нашим лицом, а соматосенсорная система утверждает, что рука за спиной... Примерно в течение 10 с, пока мы еще видим так называемый последовательный образ, попробуем сделать легкое и плавное движение головой или телом — слегка повернем голову, чуть-чуть отклонимся вперед или отступим на полшага назад — и сможем изучать работу собственного мозга, удивляясь тому, как другие сенсорные системы влияют на характер нейроизображения. Мы смотрим глазом, а видим мозгом. И не удивляемся тому, что это давным-давно известное человечеству свойство сенсорных систем нашло свое отражение в языке: «смотреть» и «видеть», если речь идет о зрении, «слушать» и «слышать» — о слухе, «нюхать» и «чуять» — это уже об обонянии. Таким образом, в основе восприятия информации извне лежат сенсорные рецепторы, которые понимают язык внешней среды. За ними располагаются более или менее совершенные звенья центрального анализа, но они общаются и друг с другом, и с сенсорными рецепторами исключительно на языке нервных сигналов.
Естественно, что в зависимости от физической природы воспринимаемых стимулов, или, как говорят, от их модальности, существуют фоторецепторы (зрительная система), хеморецепторы (обонятельная и вкусовая системы), механорецепторы (фонорецепторы в слуховой системе, рецепторы органов равновесия, рецепторы наружных покровов тела), терморецепторы (в соматосенсорной системе), гигро- и электрорецепторы. Соответственно, рецепторами работают сенсорные системы: со светом — зрительная, с механическими колебаниями в среде — слуховая и акустико-латеральная (органы боковой линии у некоторых водных животных), с идентификацией пищевых веществ — вкусовая, с пахучими сигналами — обонятельная; положение организма в пространстве и ориентацию тела в нем определяет система органов равновесия; механические, температурные и ряд иных характеристик внешней среды оценивает соматосенсорная система (осязание), а электрическую «погоду» в водоемах — электросенсорная. Как уже говорилось, этими системами в той или иной комбинации снабжены все организмы, но не все системы представлены у человека, да и сам перечень сенсорных рецепторов и систем, возможно, пока не полон.
Морфология, физиология, биохимия...
Любому путешественнику нужна карта, указывающая расположение и характеристики населенных пунктов, водоемов, естественных препятствий и, конечно же, сеть связей между этими объектами — от тропинок до магистралей. Так и исследователю сенсорных систем необходимо описание конструкции органов чувств и их связей, т.е. их структурной организации. Без этой основы любое физиологическое или биохимическое исследование не стоит даже предпринимать. В отношении конструкции морфологи всегда выступают в роли путешественников-первопроходцев, и решительный прорыв в изучении нервной системы в целом и сенсорных систем, в частности, стал возможным только после работ классиков-нейроморфологов, нобелевских лауреатов 1906 г. — С. Рамон-и-Кахаля (Испания) и К. Гольджи (Италия). Из отечественных специалистов наиболее заметен вклад А.А. Заварзина (1886—1945). Физиологи часто заблуждаются, рассматривая морфологию как вспомогательную, а потому и второстепенную область. Развитие клеточной и молекулярной биологии поставило все на свои места. Стало ясно, что на этом уровне в любой процесс (физиология) вовлекаются клеточные структуры (морфология), в которых и взаимодействуют биомолекулы (биохимия). Сегодня, чтобы добиться успеха в работе, биолог должен в равной степени владеть представлениями морфологии, физиологии, биохимии, генетики, иммунохимии и иммуноцитологии, как и молекулярной биологии в целом.
Чувствительность и динамический диапазон сенсорных систем
Сегодня известно, что в ходе эволюции у многих животных чувствительность сенсорных рецепторов достигла физического предела. Так, фоторецепторы способны регистрировать поглощение единичных световых квантов и сигнализировать о них зрительным центрам. Именно в таком режиме работает зрение человека при очень слабом освещении, в сумерках или ночью, когда «все кошки серы». Наша слуховая система также находится на высшей грани чувствительности — будь ухо на порядок чувствительнее, оно воспринимало бы тепловой шум окружающего воздуха. Электросенсорные системы скатов (их электрорецепторы сосредоточены в так называемых ампулах Лоренцини) замечают изменения напряженности электрических полей всего 0.01—0.10 мкВ/см, а обонятельные системы «обнаруживают» 10—100 молекул сигнального химического вещества, скажем, полового феромона, примерно среди 2.7•1019 молекул, составляющих 1см3 воздуха.
Диапазон естественных стимулов (его называют динамическим), в котором работают сенсорные системы, чрезвычайно широк. Так, человек пользуется зрением в диапазоне девяти порядков освещенности: от 2•10–4лк (свет ночного безлунного неба) до 105лк (солнечные пляжи или снега горнолыжных курортов). Наша слуховая система, воспринимая акустические стимулы между слуховым порогом (примерно 10–11Вт/м2) и болевым ощущением (10 Вт/м2), перекрывает 12 порядков на шкале звукового давления. Естественно, для использования сенсорных систем в столь широких пределах организмам пришлось изобрести способы сжатия динамического диапазона и множество приспособлений, которые позволяют приводить внешний сигнал к уровню, доступному для обработки. Одни приспособления (например, линзовая система глаза, ушная раковина) используются для усиления, концентрации сигнала, чтобы выделить его из шума, создаваемого средой и самой нервной системой, другие (зрачок глаза позвоночных и головоногих моллюсков), наоборот, — для его ослабления. Однако помимо этих довольно очевидных устройств сенсорные системы снабжены весьма изощренными способами фильтрации биологически важной информации, которые работают и на уровне одиночной сенсорной клетки, и на уровне высших сенсорных и даже межсенсорных центров. Именно изучением принципов детектирования внешних сигналов и обработки информации о них в организме и занимается сенсорная физиология. Замечу, сейчас она привлекает ученых во многих странах и, кстати, весьма обильно финансируется за рубежом.
Устройство сенсорных рецепторов
Необходимость воспринимать сигналы из внешней среды предопределяет и положение сенсорных клеток в организме: как правило, они располагаются на границе с внешней средой, что для весьма чувствительных клеток далеко не безопасно. В наиболее трудных условиях оказываются хеморецепторные клетки — вкусовые и обонятельные. Правда, высшие животные предпочитают периодически отбирать для анализа небольшие отдельные пробы (например, слегка принюхиваясь), а также защищать сенсорную поверхность с помощью специальной среды (обонятельной слизи у позвоночных, рецепторной лимфы у насекомых), которая проницаема для внешних молекул. Другие рецепторы защищены иначе. Фоторецепторы, например, изолированы от воздействия химических и механических стимулов прозрачным только для света барьером (светопреломляющим аппаратом). Рецепторы же органов равновесия и слуха непосредственно со средой не сообщаются, так как размещены внутри специальных, сложно устроенных органов. Электрорецепторы ампул Лоренцини тоже не имеют прямого «механохимического» контакта со средой. Они соединяются с ней специальными каналами, открывающимися наружу и заполненными желе, которое хорошо проводит электрический ток, а стенки выстланы слоем клеток-изоляторов. Вся эта система напоминает хорошо известный портативный тестер, состоящий из измерительного прибора и двух проводов со щупами.
Обычно у сенсорной клетки, как в электрической машине, различают входное устройство, преобразователь и выходной механизм. На «входе» — специализированном аппарате — избирательно воспринимаются стимулы извне; задача «преобразователя», т.е. цепи структурно и пространственно распределенных молекулярных процессов, — усиливать поступивший сигнал и переводить его на язык внутриклеточной сигнализации. И наконец, на «выходе» через синаптический контакт осуществляется передача уже закодированного внутриклеточного сигнала на второе звено сенсорной системы — афферентный интернейрон. В некоторых сенсорных системах рецепторные клетки имеют свой собственный выходной «проводник» — центральный отросток, или аксон, по которому они и передают сигнал к интернейрону.
Входные устройства.
По расположению в ткани рецепторная клетка напоминает айсберг и, подобно ему, имеет надводную часть и подводную. Если говорить языком физиологов, то сенсорный домен, обращенный во внешнюю среду, — надводная часть «айсберга», а гомеостатический домен — подводная часть. Он соседствует с другими сенсорными или вспомогательными клетками и обеспечивает гомеостаз и жизнедеятельность своей хозяйки, а также передачу сигнала в нервную систему. Сенсорная клетка, как и любая другая, ограничена плазматической мембраной, и естественно, что любое возмущение во внешней среде воздействует прежде всего на мембрану надводной части «айсберга». В ходе эволюции именно плазматическая мембрана сенсорного домена рецепторной клетки и превратилась в молекулярный биосенсор той или иной системы модальности. Таким образом, можно сказать, что сенсорная клетка морфологически поляризована.
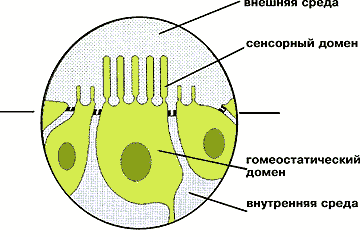
Сенсорная клетка с двумя доменами — сенсорным, обращенным во внешнюю среду,
и гомеостатическим, погруженным во внутреннюю среду организма.Детальное изучение этого явления связано с выполненными еще в 60—70-х годах пионерными работами доктора биологических наук Я.А.Винникова (1910—1997) и коллектива, который он возглавлял в нашем институте. Именно в этих работах показано, что сенсорный домен, как правило, оснащен специализированным органоидом — жгутиком (лат. cilium), который, как известно читателю, обеспечивает движение сперматозоидов, да, впрочем, и многих простейших — обитателей водной стихии. Оказалось, что сенсорные жгутики, часто видоизмененные, поистине вездесущи: они найдены в фоторецепторных клетках позвоночных животных и моллюсков; в обонятельных и слуховых клетках позвоночных и насекомых; в клетках органов равновесия позвоночных, ракообразных, моллюсков; в электрорецепторах ампул Лоренцини. Бывает, что к одному или многим жгутикам сенсорного домена клетки добавляются еще и микровиллы (микроворсинки, или микровыросты ее плазматической мембраны). Эти исследования, признанные во всем мире (а Винников — автор семи монографий, четыре из которых переведены на английский язык, член Академии наук Леопольдина, почетный доктор университета в Турине, лауреат медали Гольджи... — увы, не был избран даже членом-корреспондентом нашей Академии), показали, что молекулярные механизмы сенсорной рецепции следует искать в плазматической мембране жгутиков и микровилл. Сегодня мы знаем, что это именно так.
У позвоночных животных фоторецепция начинается с поглощения кванта света молекулой зрительного пигмента — родопсина. Локализован он в мембранах дисков наружного сегмента палочек и колбочек сетчатки, а сам сегмент, по конструкции похожий на пачку печенья, не что иное как сохранивший ножку видоизмененный жгутик: в палочках он имеет форму аккуратного цилиндра, а в колбочках — конуса. Внутри жгутика плазматическая мембрана образует многочисленные и регулярно расположенные диски, поставленные перпендикулярно ходу светового луча. Чем длиннее наружный сегмент, тем больше дисков должен пересечь луч и тем выше вероятность поглощения кванта (или, что то же самое, — коэффициент поглощения фоторецептора). Фоторецепторы членистоногих животных (насекомых, ракообразных, пауков) и головоногих моллюсков устроены не менее замечательно. Их наружный сегмент, называемый рабдомером, образован плотно упакованными микровиллами, плазматическая мембрана которых тоже содержит зрительный пигмент. Рабдомер же представляет собой оптический световод, «изобретенный» животными задолго до того, как появилось слово «оптика» и даже сам человек.
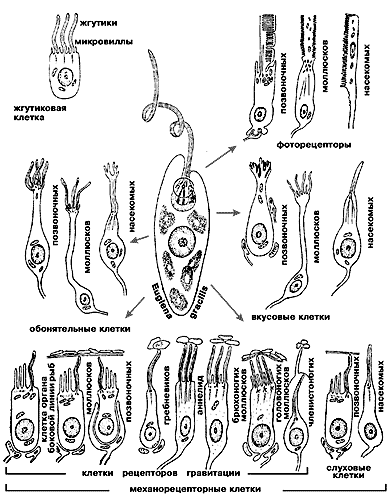
Схема эволюционного разнообразия сенсорных клеток
(Винников Я.А. // Эволюция рецепторов. Л., 1979).Оптические свойства фоторецепторов оказались настолько интересными, что возникла и успешно развивается такая область сенсорной физиологии, как оптика фоторецепторов. А молекулярной основой фоторецепции остается мембранный белок, точнее, окрашенный хромопротеид, родопсин. Его белковый компонент — опсин — не поглощает свет, эту способность и окраску он приобретает при взаимодействии с хромофором — 11-цис-изомером ретиналя (альдегида витамина А — ретинола). Максимумы спектров поглощения родопсина у разных животных могут располагаться как в видимой, так и в ультрафиолетовой части спектра. В отдельных случаях молекулы родопсина ориентированы в фоторецепторной мембране в одном предпочтительном направлении, и тогда она проявляет дихроизм. Некоторые группы животных (членистоногие, высшие моллюски) используют это свойство для анализа естественного поляризованного света (голубое небо, отражение от водных поверхностей). Что же касается опсинов, то это — видоспецифичные белки. Сейчас уже известен их аминокислотный состав для животных многих видов, оценено сходство молекул, т.е. степень гомологии, и на этой основе развиваются представления об эволюции фоторецепторов.
У животных самых разных таксономических групп плазматическая мембрана жгутиков и микровилл обонятельных клеток содержит рецептивные белки, способные реагировать с летучими (и даже малолетучими!) пахучими веществами — одорантами. Как правило, в каждой клетке обонятельного эпителия синтезируется набор из нескольких белков, причем их наборы в разных клетках неодинаковы. В результате адсорбции того или иного одоранта на мембранах жгутиков клетки хемосенсорного эпителия возбуждаются в разной степени, и на его поверхности «рисуется» обонятельное нейроизображение, свое — для каждого одоранта. Клетки, настроенные на восприятие феромонов — сигнальных веществ, испускаемых животными своего вида, — синтезируют специальный белок, молекулярный рецептор феромона.
В механорецепторах — так называемых волосковых клетках внутреннего уха и органов равновесия позвоночных животных — механическое воздействие, вызванное звуковой волной или инерциальными силами, отклоняет от вертикального положения пучок тесно прилегающих друг к другу «микровилл» (стереоцилий) плазматической мембраны, связанных между собой особыми мостиками. При отклонении пучка стереоцилии скользят одна вдоль другой, мостики натягиваются и прямо открывают ионные каналы в плазматических мембранах стереоцилий.
Таким образом, сенсорный домен плазматической мембраны рецепторной клетки настроен на восприятие определенных физических изменений во внешней среде, а их первичное детектирование происходят на молекулярном уровне.
Преобразующие и «усилительные» каскады. Внутриклеточным сигналом, передаваемым на выходное устройство сенсорного рецептора, т.е. на синапс, служит изменение разности потенциалов на плазматической мембране клетки. Это изменение принято называть рецепторным потенциалом, если сенсорный рецептор не имеет механизма импульсного кодирования, или генераторным, если рецепторный сигнал передается к синапсу импульсами. Потенциал покоя клетки отрицателен (обычно в пределах от –40 до –60 мВ). Под воздействием стимула он может или увеличиваться по абсолютной величине, становясь более отрицательным, тогда клетка гиперполяризуется, или уменьшаться, и клетка деполяризуется. На феноменологическом уровне эти события обусловлены изменением проницаемости клеточной мембраны для катионов (натрия, кальция и калия) или анионов (обычно хлора) за счет активации (открывания) или инактивации (закрывания) ионных каналов.
Итак, в сенсорной клетке существует механизм преобразования, с помощью которого активность ионных каналов меняется в зависимости от состояния собственно сенсоров — рецептивных белков. Этот механизм переводит внешний сигнал с языка физического воздействия на универсальный язык, понятный нервной системе. Поскольку рецепторный потенциал возникает как результат сложения индивидуальных токов многих ионных каналов, то иногда говорят об «усилении» входного сигнала. Но такой термин вряд ли применим к сенсорным рецепторам, поскольку усиление подразумевает операции с сигналами одной природы: это может быть электрический ток, напряжение или мощность, существуют пневматические усилители, работающие на потоках газа, и т.д. В сенсорных же рецепторах осуществляется, как мы видим, процесс, который преобразует единичное физическое «микроскопическое» воздействие (квант света, молекула одоранта) на сенсор в макроскопический рецепторный потенциал. Поэтому такое преобразование получило общее название сенсорной трансдукции (частные случаи — фото-, хемо-, механотрансдукция).
Изучение сенсорной трансдукции стало в наши дни поистине одной из самых горячих точек современной биологии. Оказалось, что в сенсорной трансдукции действуют механизмы, во многом сходные с механизмами восприятия гормональных сигналов и нейромедиаторов. В известном смысле сенсорные рецепторы можно рассматривать как удобную модель для изучения общих принципов внутриклеточной и межклеточной сигнализации, которая играет важнейшую роль в обеспечении целостности многоклеточных организмов.
Конкретным механизмам сенсорной трансдукции уже посвящена обширная специальная литература, поток ее стремительно нарастает. Но мы не будем останавливаться на этом, а наметим лишь основные вехи, которыми, как правило, отмечен путь сенсорного преобразования внешнего сигнала.
Рецептивный белок при взаимодействии с сигнальной молекулой или квантом света изменяет свою конформацию, «активируется» и становится доступным для взаимодействия с G-белком (от англ. gain — усиление). За время своей активации рецептивный белок переводит в активное состояние несколько десятков молекул G-белка, а каждая из них в свою очередь успевает «возбудить» несколько десятков молекул специального фермента. Этот фермент либо продуцирует, либо разрушает молекулы внутриклеточного медиатора (его называют также мессенджером, внутриклеточным посредником), который непосредственно регулирует состояние ионных каналов. Таким образом, единичное микрособытие на входе сенсорного рецептора приводит к массовому изменению состояния ионных каналов. Это изменение на выходе регистрируется нервной системой в виде тоже единичного, но уже макрособытия — рецепторного сигнала.
В центре внимания исследователей находятся сегодня два механизма сенсорной трансдукции. Один из них связан с использованием клеткой циклических нуклеотидов, открывающих катионные каналы — циклического аденозинмонофосфата (цАМФ) или циклического гуанозинмонофосфата (цГМФ). В первом случае G-белок активирует фермент аденилатциклазу, она продуцирует цАМФ, благодаря чему увеличивается число открытых каналов, и входящие катионы деполяризуют клетку. Если же G-белок активирует фосфодиэстеразу, которая разрушает цГМФ и уменьшает его концентрацию, ранее открытые каналы закрываются, и клетка гиперполяризуется. Известно, что в обонятельных рецепторах ведущую роль играет аденилатциклазный механизм, а в фоторецепторах позвоночных — фосфодиэстеразный. Первый механизм, хотя и несколько усложненный, работает во вкусовых рецепторах позвоночных при восприятии сладких веществ: аденилатциклаза активирует фермент протеинкиназу А, и уже этот фермент закрывает калиевые ионные каналы в базолатеральной части клетки (подводной части «айсберга») — вкусовая клетка деполяризуется. Заметим, что, когда калиевые каналы открываются, калий выходит из клетки, и она гиперполяризуется.
По второму механизму сигнал передается с помощью инозитолтрифосфата (ИТФ). Во этом случае G-белок активирует фермент фосфолипазу С, которая отщепляет от липидов плазматической мембраны диацилглицерол и инозитолтрифосфат, а он также служит вторичным посредником, открывающим катионные каналы. Наряду с аденилатциклазным механизмом инозитольный работает, в частности, в обонятельных рецепторах позвоночных и насекомых, а также во вкусовых рецепторах позвоночных при восприятии горьких веществ. В этом году сделано любопытное открытие. Оказалось, что в фоторецепторах дрозофилы, где G-белок активирует фосфолипазу С, в качестве внутриклеточного медиатора работает не ИТФ, а жирные кислоты — арахидоновая и линоленовая — производные диацилглицерольного пути.
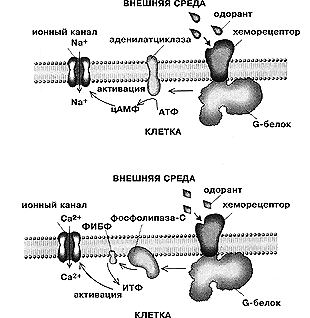
Два типичных молекулярных механизма сенсорной трансдукции —
аденилатциклазный и инозитольный — в обонятельных рецепторах.
ФИБФ — фосфоинозитолбифосфат.Сильно укороченный без вторичных посредников процесс трансдукции характерен для вкусовых рецепторов позвоночных при восприятии соленых и кислых стимулов. Когда воздействует соленый стимул, через специфические (амилорид-чувствительные) натриевые каналы во вкусовую клетку входят ионы натрия и прямо деполяризуют ее. Кислые стимулы воспринимаются иначе: образующиеся ионы водорода блокируют или натриевые каналы (у млекопитающих), или калиевые (у некоторых земноводных), и в результате электрическое состояние вкусовой клетки опять-таки меняется без помощи вторичных мессенджеров.
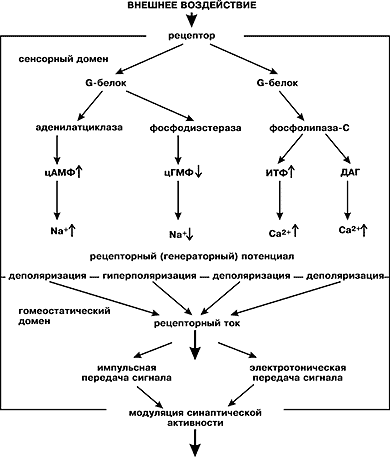
Схема основных путей сенсорной трансдукции
с участием внутриклеточных посредников.
ДАГ — диацилглицерол.Приведенные здесь механизмы даны весьма примитивно. В них многое не упомянуто: ни обратные связи, ни модулирующие влияния со стороны других ферментных систем, ни очень важные (и во многом неясные) процессы адаптации и «выключения» трансдукции, не учтена роль таких первостепенных регулирующих ее факторов, как ионы кальция.
Исследования механизмов трансдукции основаны на передовых технологиях и тонких методах молекулярной биологии: выделении генов, клонировании белков, получении трансгенных животных, химер и т.д. Стоимость таких работ чрезвычайно высока. Понятно, что эта область почти не доступна отечественным специалистам, и у нас нет крупных достижений. Правда, в 1985 г. в лаборатории Е.Е.Фесенко Института биологической физики (Пущино) были впервые обнаружены цГМФ-чувствительные каналы плазматической мембраны наружных сегментов фоторецепторов позвоночных, и на эту работу, опубликованную в журнале «Nature», исследователи ссылаются до сих пор. В лаборатории академика В.Е.Соколова (Институт проблем экологии и эволюции им.А.Н.Северцова РАН), где изучались феромоны и хемокоммуникация животных, Э.П.Зинкевич и Е.В.Аронов впервые синтезировали фрагменты молекулы феромона, которые вызывали у животного такую же поведенческую реакцию, как и сам феромон. У нас есть способные исследователи, но многие из них, не имея средств для выполнения своих проектов по сенсорной трансдукции, вынуждены работать за рубежом. В частности, роль цГМФ как посредника фототрансдукции в зрительных клетках позвоночных, изучается под руководством В.И.Говардовского в рамках российско-американского проекта по программе Американского фонда гражданских исследований и развития US Civilian Research and Development Foundation — CRDF.
Выходные устройства.
В обонятельных рецепторах позвоночных и многих беспозвоночных, механорецепторах насекомых, электрорецепторах имеется специальный механизм, кодирующий интенсивность возбуждения сенсорных клеток в виде цепочки регенеративных нервных импульсов, которые распространяются по аксону. Чем сильнее внешнее воздействие на сенсорный рецептор, тем больше степень его возбуждения, или генераторный потенциал, и тем выше частота следования импульсов. В некоторых сенсорных клетках такого механизма нет, аксон играет роль обычного электрического провода, и сигнал передается без импульсов, постоянным электрическим током, или, как говорят, электротонически. Такая передача на большие расстояния неэффективна из-за сильного затухания и сглаживания формы сигнала, поскольку удельное сопротивление цитоплазмы аксона велико (около 100 Ом•см), а его изоляция неидеальна. Этот недостаток особенно сильно сказывается при необходимости передавать «быстрые» сигналы, т.е. короткие импульсы с крутыми фронтами. Улучшить эффективность передачи можно укорочением аксона, вплоть до полной его редукции, а также путем увеличения его диаметра и/или обертывания дополнительными слоями «изоляции».
В сенсорных системах можно найти все варианты таких «линий передачи». Кажется парадоксальным, что для всех исследованных к настоящему времени фоторецепторных клеток характерна безымпульсная передача сигнала. Хотя ее предельная дальность достигает 10 мм (глаза некоторых видов «неторопливого» животного балянуса), многие животные располагают все звенья безымпульсной передачи и первичной обработки сигнала как можно ближе к фоторецепторам, вероятно, чтобы избежать его затухания и искажения, неизбежных при передаче постоянным током. Так устроена, например, сетчатка позвоночных животных. У них импульсная передача начинается только на уровне третьего звена обработки — в ганглиозных клетках — и далее идет по зрительному нерву от глаза в мозг. Подобно позвоночным, насекомые, т.е. высшие животные совершенно другой филетической ветви, тоже не используют импульсную передачу в двух первых звеньях обработки зрительного сигнала. Вспомним, однако, что фоторецепторы высших животных умеют считать кванты света, генерируя короткий одиночный электротонический сигнал. Быть может, такие дискретные одноквантовые ответы играют в зрительной системе роль нервных импульсов? Так это или нет, но в любом случае фоторецепторы давным-давно «знают», что свет имеет дискретную, квантовую природу.
Итак, в центральные отделы сенсорных систем поступает импульсная информация. На старый вопрос — одинаков ли импульсный язык в сенсорных системах разной модальности? — пока точного ответа нет. Кодируется ли модальность входного сигнала сенсорного элемента по принципу «меченой линии» или по принципу «структуры ответа» — тоже неизвестно. Есть определенные факты в пользу того, что, например, во вкусовой системе позвоночных кинетика рецепторного потенциала (а значит, и структура ответа) зависит от качества стимулирующего раствора — соленый он или кислый.
Мы хорошо знаем, что энергия передается по высоковольтным линиям, а по телеграфным и телефонным проводам идут только сообщения. Передача сенсорного сигнала — сообщение. Информационная ценность телеграммы и телефонного сообщения не зависит от размера и начертания шрифта или тембра и громкости голоса собеседника. Нервной системе энергия как таковая нужна только для поддержания ее элементов и механизмов в рабочем состоянии. Энергия же посланного сообщения необходима лишь для его надежного приема на следующем уровне. Эти азы теории информации хорошо известны не только нам, но и нашим биосенсорам. Так, если в процессе фотосинтеза у растений энергия потока световых квантов тщательно утилизируется и используется для обеспечения всех жизненно важных функций своего хозяина, то энергия кванта, поглощенного фоторецепторной клеткой, в конечном итоге рассеивается в виде тепла. Для фоторецептора важен только сам факт прибытия кванта, а его энергия практически не используется. Повторимся: в биосенсорах происходит не усиление сигнала, а его преобразование.
В век бурного расцвета и поголовного увлечения молекулярной биологией нельзя не вспомнить имена двух академиков-классиков А.А.Заварзина и Л.А.Орбели (1882—1958). Заварзин в свое время обратил внимание на удивительное анатомическое сходство в построении цепей передачи информации в зрительной системе позвоночных, насекомых и головоногих моллюсков — высших животных из далеко отстоящих таксонов. Такое сходство, по мнению Заварзина, трудно приписать происхождению от общего предка, поскольку в эволюции эти линии разошлись на довольно низком уровне и развивались параллельно. Замеченный им параллелизм Заварзин считал следствием «какого-то принципа, мало еще учтенного современной биологией».
Уже в наше время обнаружено столь же удивительное сходство в «схемотехнике» обонятельных систем позвоночных, насекомых и моллюсков. По-видимому, каждая модальность внешнего воздействия — свет, звук, запах — может анализироваться с помощью весьма ограниченного набора основных принципов схем их осуществления. В этом отношении идея Орбели о функциональной эволюции систем представляет полезную методологическую основу для выяснения, почему конкретная система, в том числе и сенсорная, развивается именно по данному пути, а не по какому-нибудь иному. Сегодня трудно сказать, как идет развитие сенсорных систем: по сходному плану, под давлением естественного отбора в соответствии с физическими свойствами внешнего мира и общими принципами переработки информации, или оно в основном запрограммировано в давно существующих генах и только развертывается в более или менее совершенной форме у животных разного уровня? Такой вопрос сейчас не выглядит крамольным и имеет под собой некоторые молекулярнобиологические основания. Так, совсем недавно обнаружено удивительное сходство стуктуры генов pax-6 позвоночных и eyeless насекомых. Оба гена определяют развитие зрительной системы, но — у неродственных животных!
Исследования сенсорных систем в России имеют крепкие корни, хорошие традиции и признанные достижения. Но, глядя в прошлое — сними шляпу, глядя в будущее — засучи рукава, как сказал кто-то. В России есть еще люди, способные засучить рукава, но современную сенсорную биологию голыми руками уже не возьмешь!