Проблема научения и памяти
Научение и память: системная перспектива
|
Ю.И. Александров Лаборатория нейрофизиологических основ психики им. В.Б. Швыркова, Институт психологии РАН, Москва,
|
|
|
|
|
|
Saturday, 01 May 2004 |
|
Введение
Проблема научения и памяти принадлежит к числу наиболее интенсивно разрабатываемых. Являясь главной для множества исследователей самого разного профиля: в молекулярной биологии, нейрофизиологии, психологии, она, однако, важна не только для них, но, пожалуй, для всех исследователей упомянутых и ряда других областей
знания. Это подчеркивал и П.В. Симонов, который называл проблему «механизмов замыкания условных связей» «центральной» [30, с.35].
Чему бы ни была посвящена конкретная работа, она имеет дело с индивидом, у которого есть память. Пока индивид жив, ее содержание модифицируется за счет приобретения новых форм поведения. Но не только. Хотя особенно глубокая реорганизация памяти имеет место, как правило, при формировании новых поведенческих актов, но и, как будет специально сказано ниже, реализация приобретенной ранее памяти есть также «повторение без повторения» [15]. Таким образом, само существование индивида есть непрерывная модификация, а точнее - непрерывное созидание памяти.
Чрезвычайно важно замечание, напрямую иллюстрирующее идею теорем Геделя о неполноте. В течение жизни происходит непрерывное возникновение новых и уточнение старых функций распознавания, активные элементы которых, сочетаясь в общем ансамбле дают текущий образ восприятия, интегрирующий в границах, выделенных внимаем, внешние в внутренние признаки восприятия, включая личное отношение. Поэтому сама личность, само личное отношение постоянно меняется с каждым соприкосновением с реальностью, выделенным вниманием.
Как это следует из названия, в настоящем сообщении проблема научения и памяти будет рассматриваться с позиций системного подхода, развиваемого в течение многих лет в системной психофизиологии, базирующейся на теории функциональных систем. Позиции системной психофизиологии, будучи оригинальными, не остаются, однако, островком, изолированным от остальной науки. Содержание целого ряда теоретических и экспериментальных статей, появившихся за последнее время в авторитетных научных журналах, приводит к заключению, что налицо новая фаза движения науки от «стимульного» к «целевому» и «холистическому» детерминизму, к утверждению системности и подчеркиванию активности индивида. Это движение еще не является главным направлением (mainstream) развития науки, но получает все большую поддержку, «официализируется» [79,80,84,100,155,157].
Несмотря на отмеченную тенденцию, и в настоящее время сохраняются существенные различия между системной и традиционной парадигмами. Естественно, подход к экспериментальному изучению закономерностей научения и памяти и к трактовке полученного материала в значительной мере зависит от методологических установок исследователя. Эмпирическое явление превращается в факт, будучи интерпретированным в терминах теории [25,26,42]. (К сожалению, часто имплицитной и потому эклектичной.) Поэтому одно и то же явление может предстать для авторов, придерживающихся разных теорий, в качестве разных фактов.
Хочется уточнить, что если говорить об объективности фактов, обеспечиваемой Методологией науки то они достаточно однозначны на уровне возможности формализации, хотя, конечно, вызывают различное понимание.
Вот почему мне представляется важным показать, как выглядят закономерности научения и памяти с позиций системного подхода, и сравнить это взгляд с традиционным.
Из только что сказанного вытекает, что, прежде чем изложить взгляд на проблему научения и памяти с системных позиций, следует сначала описать тот концептуальный аппарат, который используется для интерпретации экспериментального материала в системной парадигме. Однако, принимая во внимание ограничения объема настоящего сообщения, а также то, что на протяжении ряда лет такие описания многократно приводились (см., например, [3,4,6,7,12,37,38]), я остановлюсь лишь на том, что считаю совершенно необходимым для последующего изложения логики системного анализа названной проблемы. Это - понимание нейрона не как проводника возбуждения, а как «организма» в организме.
Детерминация активности нейрона
От концепции нейрона сумматора и проводника возбуждения к концепции его интегративной деятельности
Традиционно нейрон рассматривается как элемент, входящий в проводящую структуру большей или меньшей сложности, например, в рефлекторную дугу. Тогда функцией нейрона оказывается обеспечение проведения возбуждения. При этом в качестве стимула рассматривается пресинаптическая импульсация, а в качестве реакции –импульсация постсинаптического нейрона. То есть нейрон, как и организм, реагирует на стимулы.
Замечание относится и к ранее выделенному фрагменту. Недопустимо утрировано потому, что функционально организмы, обладающие адаптивностью поведения, не просто алгоритмически реагируют на стимулы. Думаю, что аналогия с организмами легко может завести на неверные направления понимания.
Важнейшим событием в развитии теории функциональных систем стало определение системообразующего фактора - результата системы, под которым понимался полезный приспособительный эффект в соотношении организма и среды, достигаемый при реализации системы. Таким образом, в качестве детерминанты поведения в теории функциональных систем рассматривается не прошлое по отношению к поведению событие - стимул, а будущее – результат [12]. Однако теория функциональных систем включала наряду с понятием «результат», и понятие «пусковой стимул». Это сочетание отражало, как я полагаю, эклектичность классического варианта теории. Кажущаяся необходимость использования понятия «пусковой стимул» (чем-то ведь должен «запускаться» поведенческий акт, даже если его организация определяется будущим – результатом, на который этот акт направлен!) отпала при рассмотрении поведенческого акта не изолировано, а как компонента поведенческого континуума: последовательности поведенческих актов, совершаемых индивидом на протяжении его жизни. При этом оказывается, что следующий акт в континууме реализуется после достижения и оценки результата предыдущего акта. Есть серьезные возражения против отказа от участия пускового стимула, формализованные в статье Мотивация. Упомянутая оценка - необходимая часть процессов организации следующего акта, которые, таким образом, могут быть рассмотрены как трансформационные или процессы перехода от одного акта к другому. Места для стимула, в том числе и пускового, в континууме нет [37].
Решающий шаг на пути формирования системного подхода к пониманию активности нейрона был сделан П.К.Анохиным [12] . В концепции интегративной деятельности нейрона, предложенной им вместо традиционной «суммационной» концепции, рассматривающей нейрон как сумматор и проводник возбуждения в рефлекторной дуге, генерация потенциалов действия нейроном рассматривалась как обеспечение достижения результата системы и следствие внутринейронных химических процессов. Однако, хотя в этой новой концепции и подчеркивалась роль межклеточных контактов не в проведении возбуждения, а в обмене метаболическими субстратами между клетками, активность нейрона в ней все еще рассматривалась как реакция, следующая в ответ на стимул - импульсацию пресинаптических нейронов. При этом как-то забывалось все то, что собственно привело к рассмотрению нейрона как сумматора-порогового элемента, хотя он демонстрирует именно такие свойства в эксперименте (Нейроны под контролем полупроводниковой электроники ). Кроме того, на самом деле функционального обмена метаболическими субстратами между клетками через синоптические щели не происходит.
Нейрон как «организм» в организме
Следующим шагом в развитии системных представлений о детерминации активности нейрона была формулировка В.Б. Швырковым [38] представления о том, что нейрон, как и любая живая клетка, реализует генетическую программу, нуждаясь в метаболитах, поступающих к нему от других клеток. Активность нейрона, как и поведение организма, является не реакцией на прошлое событие, а средством изменения соотношения со средой, «действием», которое обусловливает достижение результата. Последовательность событий в деятельности нейрона становится аналогичной той, которая характеризует активный целенаправленный организм, а его импульсация - аналогичной действию индивида. Опять есть неопределенность в понимании абстракции "организм", усиленная упоминанием "активной целенаправленности" без определения в чем заключается суть и механизмы, обеспечивающие активную целенаправленность. Это - серьезное возражение потому, что вообще упускаются свойства личности, ее оценки значимости совершаемого и ее мотивации, отличающие организмам с адаптивным поведением.
Иначе говоря, активность нейрона, как и поведение организма, рассматривается не как реакция, а как средство изменения соотношения со средой, «действие», которое обусловливает устранение несоответствия между «потребностями» и микросредой, в частности, за счет изменения синаптического притока. Это изменение, если оно соответствует текущим метаболическим «потребностям» нейрона, приводит к достижению им «результата» и прекращению активности.
Предполагается, что рассогласование между «потребностями», определяемыми генетически, и реально поступающими метаболитами может иметь место как при генетически обусловленных изменениях метаболизма клетки, так и при изменении притока метаболитов от других клеток.
Таким образом, в рамках данных представлений, нейрон - не «кодирующий элемент», «проводник» или «сумматор», пусть даже очень «умный», а организм в организме, обеспечивающий свои «потребности» за счет метаболитов, поступающих от других элементов.
Представление о клетке как об организме в организме не является новой идеей, сформулированной впервые в системной психофизиологии. Так, более пятидесяти лет назад Ч. Шеррингтон [141, p.73] писал: «Утверждение, что из клеток, составляющих нас, каждая является индивидуальной эгоцентричной жизнью – не просто фраза. Это не просто удобный способ описания. Клетка как компонент тела – не только визуально ограниченный модуль, это отдельная жизнь, сосредоточенная на себе. Она живет собственной жизнью… Клетка – это отдельная жизнь, и наша жизнь, которая, в свою очередь, является отдельной жизнью, всецело состоит из жизней-клеток». Что действительно нового добавлено в описываемой концепции нейрона к этому утверждению – это приведение данного общетеоретического представления об «эгоистичной» клетке, в частности, о нейроне, в соответствие с представлением о системной детерминации ее активности, связываемой с рассогласованием между «потребностями» клетки и метаболическим притоком.
Т.е. импульсация нейрона - лишь проявление его эгоцентрической индивидуальности (и не важно, что подразумевается под словом индивидуальность), потребности. А "рефлектроные дуги" - лишь системная организация такой совокупной активности. Вообще-то, конечно, не важно какими словами все называть, но важно какие механизмы при этом имеются в виду. В данном же случае эти механизмы никак не обеспечат понимание, а что же такое сознание, что такое мотивация личности, что такое оценка личностью в категориях "хорошо-плохо" происходящего с ней. Чем далее погружаемся в идею организма в организме, так более мистической она представляется.
Роль большинства химических соединений, поступающих в «микросреду» клетки, сводится к изменению свойств и скорости синтеза имеющихся в ней белков или к инициации синтеза новых белков. На метаболизм нейрона влияют и нейромедиаторы, выделяемые из терминалей контактирующих с ним нейронов. Соединяясь со «своим» рецептором, медиатор не только изменяет проницаемость ионных каналов, но и оказывает влияние на внутриклеточные процессы, выступая в качестве индуктора внутриклеточных метаболических превращений (см., например, [34]). Однако, и изменение проницаемости ионных каналов, обычно связываемое лишь с модуляцией электрического потенциала нейрона, в свою очередь оказывает существенное влияние и на метаболизм клетки: трансмембранный транспорт метаболитов, поддержание уровня pH внутри клетки [136]. Только вот не объясняется, а зачем вообще тогда изменять проницаемость этих межнейронных каналов, так, всего лишь для некоторого влияния на метаболизм клетки...
Объединение нейронов в систему как способ достижения результата на уровне целого организма и отдельной клетки
Рассмотрение нейрона как организма в организме соответствует представлениям о значительном сходстве между закономерностями обеспечения жизнедеятельности нейрона и одноклеточного организма [105]. Однако между ними существует и серьезное различие. Одноклеточный организм (как, впрочем, и многоклеточный) может обеспечить свои метаболические потребности за счет лишь собственной активности. Причем эта активность не обязательно вступает в отношения взамосогласования с активностями других организмов. Нейрон же обеспечивает «потребности» своего метаболизма объединяясь с другими элементами организма в функциональную систему. Их взаимосодействие, совместная активность приводит к достижению результата, новому соотношению целостного индивида и среды.
«Извне», на уровне целостного индивида, при анализе его внешнего поведения, результат описывается как определенное соотношение организма и внешней среды, которое прекращает действие, направленное на его достижение, и делает возможной реализацию следующего поведенческого акта. «Изнутри», на уровне отдельных нейронов, достижение результата выступает как удовлетворение метаболических «потребностей» нейронов и прекращает их импульсную активность. Интересно то, что, в принципе, удовлетворить "метаболическую потребность" можно было бы сразу для всех нейронов разом. Для этого вовсе не нужно сложнейшее и динамическое распределение их активности. Почему именно рецепторы инициируют такие потребности, а в условиях сенсорной депривации эти потребности начинают замокать по схеме затухания сновидений?
Такой подход к пониманию активности нейронов предполагает применение для ее анализа «предрезультатных» гистограмм. Построение «предрезультатных» гистограмм позволяет обнаружить нейроны, активность которых закономерно увеличивается при реализации поведения, направленного на получение данного результата, и прекращается при его достижении.
В литературе имеются данные, которые позволяют связать частоту «предрезультатной» активности с величиной потребности, с одной стороны, и с появлением поведения, направленного на удовлетворение этой потребности, с другой. Показано, что у зависимых от кокаина животных, обученных нажимать на педаль для ведения себе кокаина (через канюлю, вживленную в яремную вену), частота активности нейронов, вовлекающихся в обеспечение инструментального кокаиндобывательного поведения, тем выше, чем ниже концентрация кокаина в организме [121]. Когда частота активности, постепенно нарастая, достигает определенного уровня, развертывается инструментальное поведение, достижение результата которого - введение кокаина, - проявляется в подавлении активности этих нейронов [63,121]. Затем, по прошествии некоторого времени, концентрация кокаина опять понижается, частота активности повышается и реализуется поведение добывания кокаина - цикл повторяется.
«Действие» нейрона
«Действие» нейрона, его импульсная активность, не только влияет на микросреду, но изменяет и сам импульсирующий нейрон. Уже давно было известно, что «следовые», постспайковые процессы (такие как изменение поляризации, ионной проницаемости) играют существенную роль в регуляции чувствительности нейрона к последующему притоку. Позже как на препаратах, так и на бодрствующих животных было показано, что потенциал действия, генерируемый нейроном, распространяется не только в «обычном» направлении - по аксону к другим клеткам, но и в обратном направлении - к дендритам данного нейрона (феномен «обратного распространения», «backpropagation») [60,85]. При этом его чувствительность к синаптическому притоку существенно модифицируется. При сочетании эффектов «обратного распространения» с пресинаптической импульсацией возникают изменения в дистальных дендритах, а также и в теле нейрона. Нужно уточнить - временные изменения. Частота отклика отдельного нейрона имеет предел, связанный с нехваткой времени на подготовку новой релаксации и процессами привыкания. Но отдельный нейрон не определяет поведение организма, а лишь является одним из составляющих детекторов признаков в общем ансамбле распознавания ситуации, обеспечивая реверберацию этого ансамбля и удержания активности необходимое время. Функционально время релаксации не играет никакой роли, хотя, конечно, влияет на механизмы в реальной организации нейронной сети. Выполненные на других элементах, не обладающих таким свойством (неких нейристорах, а вместо синапсов - синоптических коммутаторах), нейронная сеть будет обеспечивать функции адаптивного поведения (см. Совершенствуем природную реализацию принципа адаптивного поведения).
Имея в виду только что изложенное, активность нейрона, как и поведение индивида, можно рассмотреть как со стороны влияния на окружающую среду (микросреду), так и со стороны модификации активного агента, модификации, соответствующей ожидаемым параметрам эффекта этих влияний и являющейся непременной характеристикой активности. Тогда модификация нейрона вследствие его собственной активности может быть рассмотрена как показатель подготовки нейрона к будущему притоку, связанному с его активностью. Иначе говоря, эти данные свидетельствуют в пользу того, что, давая спайки, нейрон не только обеспечивает необходимый ему метаболический приток, но и подготавливается к его «утилизации» [4].
Множественность «нейротрансмиттеров»
В рамках традиционного подхода к пониманию нейрона как проводника возбуждения закономерно возникает вопрос, сформулированный Kandel E.: «Почему имеются разные нейротрансмиттеры, если лишь одного достаточно для того, чтобы опосредовать передачу всех электрических сигналов?» (см. в [27]). Сложность теоретических построений, необходимых для того, чтобы ответить на этот простой вопрос, впечатляет ([27] и другие статьи в том же номере журнала).
Точно так же как особенности внешней среды формируют функции распознавания ситуации, зависящие от особенностей ситуации, чтобы задать общий контекст понимания-поведения, так же это осуществляется когда сам организм переходит на тот или иной режим своего функционирования и с помощью неспецифического влияния обеспечивает выделение оптимизируемых только в этих условиях функций распознавания (см. Воздействие эмоций на организм). При этом преимущество получают те связи, в которых повышается концентрация присущих им медиаторов. Но это можно было бы организовать в искусственных системах, не прибегая к такому способу.
Во всяком случае, очень стоит допускать естественнее дополнение специфичной функциональности нейронной сети как общей системы, обеспечивающей адаптивное поведение, еще и неспецифической функциональностью, которая различием нейромедиаторов не ограничивается. Хотя бы потому, что такому допущению ничего не противоречит.
С позиций представления о системной детерминации активности нейронов нейротрансмиттер больше не рассматривается как стимул, действующий на нейрон (или отдельный его локус) а он и не является стимулом, а является условием, а нейрон – не рассматривается как передатчик электрических сигналов однако он выполняет роль порогового элемента, срабатывающего именно от суммарного воздействия через синапсы входных импульсов и более никак. И игнорирование этого в данной работе вызывает недоумение. Вместо этого нейротрансмиттеры рассматриваются как вещества необходимые для метаболизма клетки, которые вносят вклад в удовлетворение ее «потребностей». тогда возвращается вопрос: а зачем такое их разнообразие? Многообразие и сложность этих потребностей таково, что разнообразие медиаторов само по себе не кажется удивительным. Очень притянутое "объяснение". Проблема же «множественности нейротрансмиттеров» [27], т.е. множественности передатчиков гомогенных электрических сигналов, превращается при этом в проблему определения специфики метаболических паттернов, связанных с обеспечением тех или иных «потребностей», и в проблему модификации паттернов при формировании специализации нейронов в процессе системогенеза (см. ниже).
Заметим, что в рамках представлений, отрицающих подход к нейрону как к проводнику возбуждения почему именно как к проводнику? Но при этом - сумматору? Одно с другим не вяжется..., сам термин «медиатор» или «нейротрансмиттер» (используемый в значении «биологически активное вещество, являющееся посредником в процессе передачи возбуждения через осуществление синаптического влияния») представляется неадекватным. Замечу, что неадекватным - в контексте представлений автора, но никак не в контексте теории нейронной сети, дееспособно описывающей адаптивное поведение.
Научение как образование «следов» и как системогенез
Системно-селекционная концепция научения
Идея развития, наряду с идеей системности, относится к фундаментальным положениям теории функциональных систем. Обе они были воплощены в концепции системогенеза замечу, что в этом словосочетании так же скрывается мистическая недоопределенность и вот почему: системы, объекты и т.п. абстракции не существуют сами по себе объективно и поэтому что-то не может и порождать их, см. еще раз Абстракции, которая сформулирована с привлечением обширного экспериментального материала, накопленного при исследовании формирования нервной системы и поведения. В этих исследованиях было обнаружено, что в процессе раннего онтогенеза избирательно и ускоренно созревают именно те элементы организма, имеющие самую разную локализацию, которые необходимы для достижения результатов систем, обеспечивающих выживание организма на самом раннем этапе индивидуального развития [12].
В настоящее время становится общепризнанным, что многие закономерности модификации функциональных и морфологических свойств нейронов, а также регуляции экспрессии генов, лежащие в основе научения у взрослых, сходны с теми, которые определяют процессы созревания, характеризующие ранние этапы онтогенеза [9,10]. Это дает авторам основание рассматривать научение как реактивацию процессов созревания, имеющих место в раннем онтогенезе.
В теории функциональных систем, наряду с признанием специфических характеристик ранних этапов индивидуального развития по сравнению с поздними [3,41], уже довольно давно [32,36] было обосновано представление о том, что системогенез имеет место не только в раннем онтогенезе, но и у взрослых. Формирование нового поведенческого акта в любом возрасте есть формирование новой системы - системогенез. Ни в коем случае не могу с этим согласиться (не говоря о некорректности использования явно популистского словосочетания системогенез) потому, что поведенческий акт не формируется как нечто раз и навсегда неизменное, как уникальная система, а постоянно модифицируется при каждом актуальном воплощении элементов этого поведения, он корректирируется каждым текущим отношением к его результатам и он модифицируется каждым изменением условий, в которых осуществляется (а неизменных условий не бывает) уточнением проводимостей синапсов связей, которые составляли активный в этом поведении ансамбль возбуждения нейронной сети.
Позднее был сделан вывод о том, что принципиальным для понимания различий роли отдельных нейронов в обеспечении поведения является учет истории формирования поведения, т.е. истории последовательных системогенезов [3,5], а затем разработана системно-селекционная концепция научения ([147], см. также в [4,6,7,38]).
Системно-селекционной концепции созвучны современные идеи о «функциональной специализации», пришедшие на смену идеям «функциональной локализации», и о селективном (отбор из множества клеток мозга нейронов с определенными свойствами), а не инструктивном (изменение свойств, «инструктирование» клеток соответствующими сигналами) принципе, лежащем в основе формирования нейронных объединений на ранних и поздних стадиях онтогенеза [77]. Представление, сложившееся как в когнитивных науках, так и в биологии, о том, что индивид снабжен «многоцелевыми» врожденными элементарными «кирпичиками» - «примитивами», из которых в условиях инструктирующего влияния среды может быть сконструировано что угодно в процессе «ассоциативного обучения», подвергается разносторонней критике [126]. Так, например, Дж. Эдельманом доказательно обосновываются преимущества представления, согласно которому научение есть процесс селекции и прогрессивной специализации [77].
Принцип селекции по Эдельману может быть сведен к следующим положениям. В мозгу формируются группы нейронов, каждая из которых по-своему активируется при определенных изменениях внешней среды. Что заставляет их активизироваться? Если активность нейронов – выражение их собственной потребности? Специфика группы обусловлена как генетическими, так и эпигенетическими модификациями, происшедшими независимо от упомянутых изменений. А что делать с данными о развитии и специализации детекторов во входных “анализаторах”? Если лишать восприятие некоторых признаков во время развития таких слоев детекторов, то в дальнейшем они уже не распознаются. При этом вместо импульсов от рецепторов можно непосредственно электродом возбуждать эти детекторы, вызывая зрительные образы. А позднее научение имеет тот же механизм? Селекция имеет место уже при созревании мозга в раннем онтогенезе, в процессе которого множество нейронов гибнет. Отобранные же клетки составляют первичный ассортимент. Вторичный ассортимент, формируется в результате селекции, происходящей при научении в процессе поведенческого взаимодействия со средой. Принятие положения о селекции как основе развития на всех его этапах устраняет дихотомию между созреванием и научением [134]. Т.е. научение – только созревание? А во время запоминания стиха актером у него резко начинается созревание? Типы "потребностей" нейронов даются с генетическим кодом и если что-то в рамках адаптивного поведения потребует нового типа, то получится сбой адаптации? Не все новое может быть освоено?
В рамках системно-селекционной концепции научения формирование новой системы рассматривается как формирование нового элемента индивидуального опыта в процессе научения. В основе формирования новых функциональных систем при научении лежит селекция нейронов из «резерва» (предположительно низко активных или «молчащих» клеток). Эти нейроны могут быть сопоставлены с первичным ассортиментом и обозначены как преспециализированные клетки.
Селекция нейронов зависит от их индивидуальных свойств, т.е. от особенностей их метаболических «потребностей». Отобранные клетки становятся специализированными относительно вновь формируемой системы. Эти нейроны могут быть сопоставлены с вторичным ассортиментом по Эдельману. Во всех экспериментах по привязке нейронов к электронной схеме их свойства совершенно не завесили от того откуда и какой нейрон взят. Независимо от особенностей строения их тела они проявляли совершенно идентичные свойства, реагируя на деполяризацию мембраны своим разрядом. Это свойство, в дополнение с синоптическими коммутаторами является необходимым и достаточным для образования функциональных распознавателей любого уровня сложности, что строго было строго доказано моделями персептронов. Лишь эти однотипные функциональные элементы и разнообразие рецепторов и эффекторов (в том числе “внутренние” рецепторы значимости-отношения, самыми базовыми из которых являются “хорошо” и “плохо”) позволяют организовать адаптивное поведение с личностным отношением. См. статью Совершенствуем природную реализацию принципа адаптивного поведения.
Специализация нейронов относительно вновь формируемых систем — системная специализация — постоянна. (Интересно, что сформированная «память» иммунных клеток также постоянна, они никогда не «забывают» [93].) Таким образом, новая система оказывается «добавкой» к ранее сформированным, «наслаиваясь» на них. Не верно потому, что в дополнение к существующим, уже сформированным связям между нейронами, которые и определяют специализацию нейрона, возможно формирование все новых за счет еще не эффективных синапсов, оказывая как возбуждающее, как и тормозное влияние. Таким образом может корректироваться, уточняться специализация. Что и происходит на уровне, например, “зрительного анализатора“ когда корректируется изменение оптических свойств глаза с возрастом.
Положения о наличии в мозге животных разных видов большого числа «молчащих» клеток, об увеличении количества активных клеток при обучении, а также о том, что вновь сформированные специализации нейронов остаются постоянными (в эксперименте - в течение всего периода хронической регистрации: недели и даже месяцы), и что при научении происходит скорее вовлечение новых нейронов, чем переобучение старых, согласуются с данными, полученными в работах ряда лабораторий [59,63,99,152,156,160,162]. возражение – выше. При обучении свойства самих нейронов не меняются, а меняется проводимость их синапсов.
Актуализация памяти при реализации поведения
Как используется материал памяти, сформированный при научении, для реализации поведения? В экспериментах с регистрацией активности нейронов, специализированных относительно систем разного «возраста», обнаружено, что осуществление поведения обеспечивается не только посредством реализации новых систем, сформированных при обучении актам, которые составляют это поведение, но и посредством одновременной реализации множества более старых систем, сформированных на предыдущих этапах индивидуального развития. Последние могут вовлекаться в обеспечение многих поведений, т.е. относиться к системам, которые являются общими для разных актов. Замечу, что если один и тот же нейрон, принадлежащий к такой «общей» системе, вовлекается в разные акты, то характеристики его активации в этих актах различаются, так как в них он должен согласовывать свою активность с активностью разных наборов клеток [3]. Скорее можно говорить только о степени активности нейрона, вовлеченного в текущую общую активность нейронной сети, но не изменение каких-то характеристик этой активности, - выходная импульсация остается стандартной.
Таким образом, системы, реализация которых обеспечивает достижение результата поведенческого акта, формируются на последовательных стадиях индивидуального развития. Системная структура поведения отражает историю его формирования. Иначе говоря, реализация поведения есть реализация истории формирования поведения, т.е. множества систем, каждая из которых фиксирует этап становления данного поведения. Отсюда логически следует и экспериментально подтверждается [6,17] следующее положение. Нейрональное обеспечение внешне одного и того же поведения различается, если история формирования этого поведения различна. По сути утверждение верно, а вкладываемый смысл вызывает интересный вопрос: поведение всегда контекстно зависимо от условий и, мало того, от текущего эмоционального состояния организма. В различных эмоциональных состояниях в разном стиле выполняются внешне одни и те же поведенческие реакции. Но если поведенческие программы формуются как окончательно готовая "система", а для несколько отличной требуется уже заново формировать новую систему, а градаций условий бесконечное множество, то не кажется ли такое явное расточительство порочной моделью? Тем боле, что существуют модели, свободные от такого вот расточительства? А, следовательно, различается и структура памяти, актуализируемая (активируемая) при реализации этого поведения. Итак, поскольку история формирования поведения лежит в основе закономерностей его реализации, ясно, что «исторический нигилизм» отрицательно сказывается на понимании получаемого экспериментального материала.
Замечу, что «исторический нигилизм» может сказываться на понимании результатов исследования не только в тех случаях, когда анализируется нейронное обеспечение поведения. Содержание памяти всегда индивидуально. Даже в тех случаях, когда речь идет о видоспецифических актах. Это содержание определяет характер любого взаимодействия индивида со средой. Влияние особенностей индивидуального развития на активность мозга, свойства рецептивных полей нейронов, в том числе клеток первичных сенсорных областей коры, обнаруживается при решении самых разных исследовательских задач (см., например, [70,78,87,89,115,133,144]). Имея в виду методы экспериментирования аналитической физиологии, важно подчеркнуть, что эти особенности проявляется не только у бодрствующих индивидов. Они обнаруживает себя, например, в измененной чувствительности синапсов при исследовании срезов мозга [48] или при анализе «настройки» нейронов наркотизированных животных на те или иные параметры стимуляции [88,153].
Из сказанного следует, что описать механизмы поведения с системных позиций – значит выяснить, каковы «паттерны нейронных специализаций» [3,6] разных структур мозга, т.е. относительно каких конкретно систем специализированы нейроны этих структуры, как формируются системные специализации. Это значит также изучить динамику согласования активности нейронов, принадлежащих к одной и той же и к разным системам, т.е. динамику «внутрисистемных» и «межсистемных» отношений при реализации поведения. Я считаю, что рассмотрение упомянутых вопросов в качестве ключевых хорошо согласуется с тем набором проблем, которые выделял как центральные для системного подхода П.В. Симонов. Он подчеркивал, что «системный подход продуктивен только в том случае, если мы более или менее определенно указываем на конкретные элементы, из которых состоит данная система, на функциональное значение каждого из этих элементов и на правила взаимодействия элементов друг с другом при функционировании системы в целом» [30, с.95].
Неонейрогенез
К настоящему времени получены убедительные доказательства наличия неонейрогенеза у взрослых млекопитающих, в том числе у людей [82,129]. Получены также данные о том, что, хотя синаптическая активность появляется у этих новых нейронов довольно быстро после завершения миграции, но спайковая активность у них отсутствует; из всех свойств нейрона она оказывается последней по времени появления [61]. Следовательно, в динамике вовлечения вновь появившихся клеток в совместную деятельность наблюдаются переход от «молчания» к импульсацции, как это, возможно, происходит при вовлечении «резервных» клеток в процесс специализации Другими словами, для появления специализации данного молодого нейрона требуется его созревание до уровня спонтанной активности..Это - важный вывод. Упомянутые данные, наряду с результатами, полученными G.Kempermann et al. [103], показывающими, что число выживших нейронов, вновь появившихся в процессе нейрогенеза у взрослых животных, увеличивается при содержании последних в обогащенной среде (принцип «используй или потеряешь» [129]), а также данные о том, что искусственное угнетение нейрогенеза нарушает формирование памяти [146] думаю, что такой вывод нуждается в корректной и тщательной проверке, позволяют высказать следующее предположение: неонейрогенез может вносить вклад в процессы системогенеза (рис. 2). Наряду с рекрутированием клеток «резерва» и вновь появившиеся нейроны специализируются относительно новых систем. Как мне представляется, дополнительным сильным аргументом в пользу высказанного предположения являются данные, полученные в лаборатории F.N. Nottebohm [124]. Было обнаружено, что у канареек нейроны вентрального гиперстриатума, специализированные относительно новой песни (их метили пероксидазой хрена), имеют и тимидиновые метки, относясь к вновь появившимся нейронам.
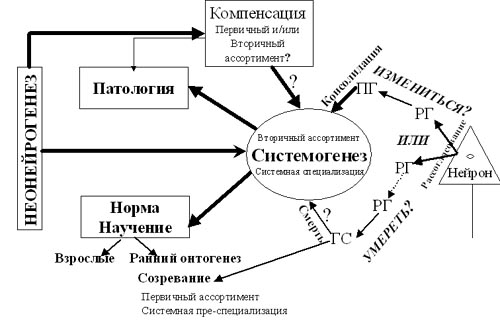
Рис. 2. Умереть или измениться? Теоретическая схема.
РГ – «ранние» гены, ПГ – «поздние» гены, ГС – гены «смерти». Подробности – см. текст.
Неонейрогенез может иметь значение и для замены нейронов первичного и/или вторичного ассортиментов, гибнущих как в условиях нормы, так и при патологии (см. ниже). Что касается патологии, в литературе уже имеются данные об интенсификации неонейрогенеза при локальных поражениях мозга взрослых индивидов и о миграции вновь появившихся нейронов в область повреждения [163]. Знаки вопроса на рис. 2 (внутри фрагмента Компенсация и у стрелки, идущей от него) означают, что в настоящее время трудно сказать, какой именно ассортимент (или оба) подлежит восполнению, и каким образом мог бы меняться состав ранее специализировавшихся нейронов в том случае, если компенсация вторичного ассортимента имеет место.
С пополнением набора преспециализированных клеток (первичного ассортимента) ситуация кажется более ясной. Если он имеет место, то, по-видимому, «встраивание» новых нейронов должно происходить по общим для всех преспециализированных клеток «резерва» правилам – в процессе системогенеза. Если таковой не происходит в течение определенного времени, вновь появившиеся нейроны гибнут. Я считаю, что в пользу этого говорят многочисленные данные, подобные приведенным выше данным G.Kempermann.
Говоря о пополнении набора преспециализированных клеток (первичного ассортимента) за счет нейрогенеза в норме, можно отметить, что в настоящее время имеются веские основания думать, что научение ведет не только к выживанию уже появившихся нейронов, но и интенсификации пролиферации. Предполагается, что эта интенсификация имеет значение для будущего научения [129].
Что касается компенсации специализированных клеток (вторичного ассортимента), трудно допустить, чтобы и она могла происходить вне системных процессов, требующих активации клеток, принадлежащих к системе, и проб, направленных на достижение ее результата. Возможно такие пробные согласования новых и «старых» клеток могут развертываться во время реконсолидационных процессов (см. ниже).
Консолидация памяти: от формирования и упрочения «следов» посредством повышения эффективности синапсов к системогенезу
Альтернативно нейрогенез можно и нужно рассматривать и как средство реставрации утерянных элементов распознавания (причин таких утерь много) и как дополнительное средство, позволяющее формировать новые функции распознавания в данной зоне мозга.
Проблема формирования и консолидации памяти решается с применением самых современных методов и базируется на современных концептуальных схемах. Несмотря на это, в основе большинства этих схем и исследований – идея Декарта, сформулированная им более трехсот лет назад: «следы» существуют потому, что поры в мозгу, через которые дух проходил раньше, становятся все более проходимыми, когда дух проходит через них снова. И дух легче проходит именно через эти поры. В соответствии с этой идеей важно выяснить, каковы механизмы увеличения проходимости пор, в каких мозговых структурах больше подобных пор, все ли поры обладают свойством изменчивости, сколь долго сохраняется повышенная проходимость пор и т.д., и т.п. Соответственно, при всем разнообразии подходов к пониманию консолидации, в качестве наиболее базовой ее закономерности рассматривается долгосрочное усиление синаптического проведения в дуге (ах) рефлекса, сетях и пр.
Иначе выглядит процесс формирования и упрочения памяти с системных позиций. С этих позиций, как уже было отмечено, нейрон не является проводником возбуждения. такое упорное повторение намеренно упрощенного описания удручает, хотя автор прекрасно осведомлен о более полной концепции функциональности нейронной сети. Поэтому проблема увеличения эффективности проведения не возникает. Формирование нового поведения рассматривается не как проторение путей и образование «следов», осуществляемое за счет увеличения синаптической эффективности. Нет, конечно! происходит обычно не изменение ранее взвешенной проводимости (как взвешивается описывается в статьях по приведенным ссылкам), а формирование новых проводимостей, как правило, в других синапсах потому, что изменение условий приводит к изменению и узора активности предшествующих нейронов, что и закрепляет связи уже между ними, а не прежними. В этом - суть изменения функции распознавания. Интересно, почему так настойчиво автор не желает воспринимать в рамках функциональности изменение проводимости синапса, ведь совершенно не возможно отрицать его влияние на возбуждение нейрона. И это влияние неизмеримо более действенно и очевидно, чем предположение о внутренне обусловленном поведении нейрона. Не как связывание структур, последовательное активирование которых обеспечивает реализацию вновь сформированного поведения. Оно представляет собой формирование новой системы совместно активирующихся клеток организма, в том числе и нейронов, расположенных в самых разных структурах мозга, вовсе не обязательно связанных прямыми влияниями. Да, возможно неспецифическая, общая тенденция к преимущественно тем или иным активностям, что определяют концентрации медиаторов, но только не прямое согласованное взаимовлияние, переносчиком которого должно быть нечто вполне материальное. Возможность такого неспецифического фона реализована в дополнение, а не вместо взаимовлияний, обусловленных электрической активностью в нейронной сети.
Переход от первой ко второй, системной позиции осуществляется в настоящее время уже не только на основе теоретических соображений, но, дополнительно, исходя из результатов экспериментов. В недавней обзорно-теоретической статье G. Horn отмечает, что результаты кросскорреляционного анализа активности нейронов IMHV не подтверждают представления о том, что при научении происходит усиление связей между нейронами, как это должно было бы быть при образовании «хеббовских ансамблей». «Скорее, - заключает автор – нейроны формируют набор параллельно [организованных], в значительной степени прямо не связанных элементов, что обеспечивает большую эффективность в сохранении [памяти], чем система непосредственно связанных элементов» [95, p.121].
Полагаю, что подобная позиция приходит в серьезное противоречие с базовыми положениями традиционного подхода. И хотя обоснованные призывы «пересмотреть имеющиеся модели [консолидации] и не быть при этом догматичными в отношении взглядов, не совпадающих с нашими предвзятыми представлениями о работе мозга» [116, p.467] имеют место, но все же внутри традиционного подхода позиция G. Horn пока может рассматриваться как оригинальная. В том же журнале, в котором появилась цитированная выше статья G. Horn, тремя месяцами раньше, была опубликована дискуссия, участник которой J. Lisman писал: «Как считал Хебб, ансамбли формируются, когда синаптические связи между активными клетками избирательно усиливаются. Я не слышал ни о каких обоснованных альтернативах, объясняющих сохранение ассоциативной памяти» [110, p.929].
В 2002 году присуждена нобелевская премия в области физиологии и медицины за 2000 г. троим ученым — Арвиду Карлссону, Полу Грингарду и Эрику Канделу за открытия, касающиеся преобразования сигналов в нервной системе .
Какие же закономерности можно выделить в процессах формирования и сохранения памяти, если посмотреть на них с системной точки зрения? И если не рассматривать в качестве основы таких процессов механизм увеличения синаптической проводимости в цепях связанных нейронов, как быть с огромным материалом, полученным при изучении таких механизмов? К такому материалу относятся, в частности, многочисленные работы, демонстрирующие активацию генетического аппарата клетки, изменение возбудимости и морфологические перестройки нейронов при научении. Для ответа на первый вопрос необходимо сначала сказать несколько слов о реконсолидации.
Реконсолидация при реактивации памяти и при научении
Еще F. Bartlett [54] предлагал полностью отбросить взгляды, в соответствии с которыми «воспроизведение из памяти» рассматривается как «повторное возбуждение неизменных «следов». В настоящее время обнаружены молекулярно-биологические закономерности реконсолидации памяти, лежащие в основе ее модификации после повторной актуализации (см., например, [137]). При формировании нового материала памяти необходим процесс синтеза белков, лежащий в основе процессов ее консолидации. Активация памяти, как и ее формирование, требует синтеза белка для реконсолидационных процессов. В связи с этим в последнее время предлагается связывать протеин-зависимые консолидационные процессы не с «новой», а шире - с «активной» памятью [117].
Представление о реконсолидационных модификациях не вступают в противоречие с приведенным выше положением о постоянстве системной специализации нейронов, которая формируется при научении. оно просто для этого положения как коту второй хвост: ничего не дает, а вот для углубления представлений о механизмах коммутации сигналов нейронной сети дает очень немало.. Анализ накопленного экспериментального материала позволяет K.Nader заключить, что, конечно, данные о реконсолидации «противоречит взгляду на память как лабильную только один раз». Однако он специально подчеркивает, что «реактивация консолидированной памяти не отменяет модификаций, обусловивших формирование долговременной памяти (вне зависимости от того, какие они - структурные или химические)» [116, р.467].
Реконсолидационные процессы представляют для нас особый интерес в связи с тем системным представлением о формировании памяти, которое было изложено выше. С точки зрения системно-селекционной концепции, оно представляет собой специализацию новой группы нейронов относительно вновь формируемой системы и «добавление» последней к ранее сформированным системам. Это добавление, видимо, требует взаимного согласования нового элемента и ранее сформированных и приводит к модификации последних.
Слишком много слишком слабых предположений, в целом образующих слишком зыбкую конструкцию.
Поведенческие данные, полученные в лаборатории И.П.Павлова [24], позволили ему прийти к заключению о том, что прибавление новых условных рефлексов сейчас же отзывается на состоянии прежних. В последнее время на основании данных, полученных в экспериментах с определением системной специализации нейронов при последовательном формировании разных поведенческих актов, также был сделан вывод об изменении ранее сформированной системы поведенческого акта после обучения следующему акту. В экспериментах с регистрацией нейронной активности у животных, первоначально обученных инструментальному пищедобывательному поведению, а затем в той же экспериментальной клетке алкогольдобывательному поведению, было обнаружено, что нейроны, специализированные относительно систем первого поведения, претерпевают при формировании второго модификацию и начинают вовлекаться также и в обеспечение алкогольдобывательного поведения вместе с нейронами, вновь специализировавшимися относительно этого поведения. Эта реконсолидационная модификация, претерпеваемая предсуществующей, «старой» системой при появлении связанной с ней новой системы, была названа «аккомодационной» реконсолидацией [47].
Не сами нейроны претерпевают модификацию, а изменяется конфигурация проводимостей их синапсов. Только и всего. И не нужно думать о том, что произошло с “потребностью” самого нейрона.
Естественно, такая позиция предполагает постановку вопроса о сходстве и различиях между реактивационной и аккомодационной реконсолидациями. Экспериментальная разработка этой проблемы имела бы большое значение для формулировки более дифференцированных представлений о формировании и функционировании памяти.
Консолидация с системной точки зрения
Сказанное выше, как мне представляется, означает, что при анализе формирования памяти следует учитывать не только появление ее новых элементов, но и модификацию ранее сформированных. При учете этого фактора можно предложить системное описание процесса консолидации. Консолидация с этой точки зрения включает две группы неразрывно связанных процессов.
1) Процессы системной специализации: морфологическая и функциональная модификация нейронов, связанная с их вовлечением в обеспечение вновь формируемой системы.
2) Процессы аккомодационной реконсолидации, обусловленные включением этой системы в существующую структуру памяти индивида: морфологическая и функциональная модификация нейронов, принадлежащих к ранее сформированным системам.
Обнаруживаемые во множестве нейрофизиологических, морфологических, молекулярно-биологических и других исследований модификации нейронов, сопутствующие научению, могут быть связаны как с первой, так и со второй группой процессов. Так, например, появление активаций на условный сигнал у тех нейронов, которые до сочетания условного и безусловного сигналов отвечали только на безусловный, связаны скорее всего именно с аккомодационной реконсолидацией, а не с системной специализацией (см. также [3]). То же можно сказать относительно модификаций в первичной моторной коре, в которой после обучения разным поведенческим актам выявляется преобладание нейронов, специализированных относительно «старых» систем, сформированных на ранних этапах индивидуального развития [1,6]. Именно с реорганизацией этих нейронов, видимо, связано увеличение уровня экспрессии «ранних» генов (подробнее см. ниже) при обучении крыс инструментальному нажатию на педаль лапой, обнаруживаемое в зоне проекции этой лапы в первичной моторной коре [62].
Дифференцированный подход к модификациям первой и второй группы явится существенным шагом вперед на пути к пониманию закономерностей формирования памяти.
В начале формирования памяти: экспрессия «ранних» генов как показатель рассогласования
Консолидация, с какой бы точки зрения ее ни рассматривать, включает морфологические изменения нейронов (см., однако, [50]), такие, например, как изменение размера синапсов, изменение их числа [53]. Начальным звеном каскада молекулярно-биологических процессов, обусловливающих морфологические модификации нейронов как в процессе морфогенеза (ранний онтогенез), так и при консолидации памяти, формируемой у взрослых индивидов, является экспрессия «ранних» генов. Активация «ранних» генов довольно кратковременный процесс (занимает около 2 часов), сменяемый второй волной экспрессии - «поздних» генов; в составе второй волны активируются морфорегуляторные молекулы, имеющие непосредственное отношение к морфологическим модификациям нейрона [9,10].
В настоящее время связь экспрессии «ранних» генов с процессами научения, которая отмечалась уже давно (см. в [9,10]), становится все более очевидной [95]. В системно-селекционной концепции научение связывается с формированием специализаций нейронов относительно формируемой системы. Логично было бы тогда ожидать, что экспрессия «ранних» генов и формирование специализаций связаны между собой. Как-то все, безусловно, связано. И любые изменения не остаются бесследными и для тела нейрона, не только вот его основная функциональность как суммирующего порогового элемента, не меняется. В исследованиях, проведенных нашей лабораторией совместно с отделом системогенеза НИИ нормальной физиологии им. П.К. Анохина [29], были получены данные, указывающие на то, что экспрессия «ранних» генов лежит в основе формирования специализаций нейронов в отношении вновь формируемых инструментальных поведенческих актов. “В основе” – не обоснованное утверждение. В тех мозговых структурах, в которых после обучения обнаруживалось достоверно больше вновь специализированных нейронов, была соответственно достоверно более выражена и экспрессия (рис. 3; данные экспериментов О.Е. Сварник и др. [29]).
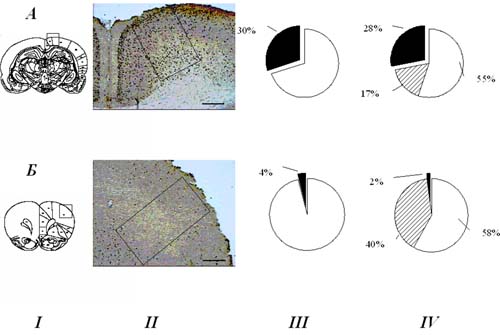
Рис. 3. Сопоставление относительного числа Fos-положительных клеток (III) с паттернами поведенческих специализаций нейронов (IV) в цингулятной (А) и антеролатеральной (Б) коре.
I - диаграммы фронтальных срезов, обозначающие исследуемые области. II - микрофотографии фронтальных срезов, показывающие Fos-окрашенные ядра нейронов крысы, сформировавшей новое поведение; калибровка: 500 мкм. III - темный сектор – процент нейронов, экспрессирующих c-Fos; светлый сектор – не экспрессирующих c-Fos. IV - темный сектор – процент нейронов, принадлежащих к «новым» системам, сформированным при обучении крыс инструментальному акту нажатия на педаль в пищедобывательном поведении; сектор со штриховкой – процент нейронов, принадлежащих к «старым» системам, сформированным на этапах индивидуального развития, предшествующих обучению инструментальному пищедобывательному поведению; светлый сектор – нейроны, не вовлекающиеся в обеспечение данного поведения. Видно, что в цингулятной коре значительно выше, чем в антеролатеральной, как число клеток, экспрессирующих c-Fos, так и число нейронов, специализированных относительно вновь сформированной системы инструментального поведенческого акта.
Активация «ранних» генов у взрослого индивида имеет место не только при научении, но и при голоде, стрессе, поражениях нервной системы или ишемии мозга [74,123,150]. Показано, что изменение микросреды нейронов обусловливает появление активности у ранее молчавших клеток [40] и экспрессию ранних генов [57,150]. Поэтому, имея в виду сказанное выше о детерминации активности нейрона рассогласованием между его «потребностями» и притоком метаболитов, а также аргументированную возможность рассмотреть экспрессию ранних генов как специфическое проявление активности клетки [65], возникающей в ситуации новизны [11], логично предполагать, что общим для всех перечисленных выше ситуаций, включая научение, является рассогласование. - опять очень слабое утверждение, как бы не замечающее другие возможные варианты “логичных” предположений. Оно возникает вследствие того, что ранее сформированные способы согласования метаболических «потребностей» нейронов оказываются неэффективными в условиях стойкого изменения микросреды нейронов.
Рассогласование является, в известном смысле, межпарадигмальной концепцией, важнейшим компонентом не только теории функциональных систем, но и других теорий. Так, П.В.Симонов отмечал «генетическое родство» биологической и информационной теории эмоций и подчеркивал, что обе они восходят к одному «идейному источнику – к концепции Павлова о несовпадении (рассогласовании) ранее сложившегося внутреннего и изменившегося внешнего динамического стереотипа» [30, c.60]. а это – просто неимоверно притянутый за уши довод, учитывая то, насколько одиозна и надумана концепция эмоций Симонова: в отдельной статье. Почему-то постоянно упускается такой важный критерий, как личное отношение к происходящему: отрицательное или положительное, чаще неосознанное, что, по-сути и корректирует жизненный опыт (модифицирует память).
Поиск новых путей устранения рассогласования включает как модификации на уровне поведенческих адаптаций, так и молекулярно-генетические и морфологические перестройки. Очевидно, что подобные модификации имеют место как в норме, так и в патологии.
Хотя в «обыденной науке» (folk science; основанное на здравом смысле, не требующее точных определений понимание) норма и патология мыслятся как принципиально различные состояния, причем патология рассматривается как слом, разрушение нормальных отлаженных механизмов, но в собственно науке уже давно обосновано представление о том, что процессы, называемые патологическими, не являются дезорганизацией или хаосом, а лишь своеобразным, выработанным в процессе эволюции приспособлением к условиям существования; нет ни одного патологического процесса, который не имел бы своего прототипа в норме [14,19,90].
О формировании новой системы, направленной на достижение конкретного результата, как о важнейшем звене механизмов научения уже шла речь выше. Адаптационные изменения организации внутренней среды и соотношения индивида со средой внешней, возникающие в условиях патологии, могут быть также рассмотрены, как системный процесс, захватывающий весь организм и направленный на обеспечение возможности достижения положительных результатов [13,90]. Многочисленными исследованиями динамики активности мозга после локальных его повреждений показано, что в интактных структурах мозга развиваются процессы реорганизации, обусловливающие восстановление поведения, и что эти процессы сопоставимы с пластическими перестройками, имеющими место при научении в норме [46,68,158].
Так, в настоящее время накапливается все больше данных, позволяющих полагать, что между нейронными механизмами, лежащими в основе формирования долговременной памяти при обучении и в основе «долгосрочных адаптаций», возникающих при хроническом воздействии аддиктивных веществ, имеется существенное сходство [120,135]. Результаты наших исследований позволили предположить, что определенный вид «долгосрочных адаптаций», имеющих место при хроническом употреблении алкоголя, не просто сходен, но идентичен модификациям, лежащим в основе формирования нового опыта. Это означает, что введение любых психоделиков модифицирует долговременную память в соответствии с вызванными этими психоделиками неестественными состояниями. К ним относятся перестройки нейронов, связанные с формированием новых специализаций относительно алкогольдобывательного поведения, а также с процессами аккомодационной реконсолидации преморбидных (сформированных до возникновения потребности в алкоголе) специализаций [47].
В связи с только что отмеченным сходством, а также при учете сходства молекулярно-биологических механизмов, лежащих в основе созревания и научения, не удивительно, что реювенилизация - активация у взрослого процессов, характерных для созревания мозга в раннем онтогенезе, - имеет место не только при научении в норме, но и при восстановлении после поражений мозга [69]. Для целей дальнейшего обсуждения важно подчеркнуть, что к механизмам, реактивируемым в условиях патологии у взрослого, относят и апоптотическую гибель клеток: физиологический апоптоз в развивающемся мозге и патологический апоптоз во взрослом мозге имеют сходные молекулярные механизмы [164].
Нейрон, как отмечалось, может обеспечить «потребности» своего метаболизма, объединяясь с другими элементами организма в функциональную систему, извлекаемую из памяти. Что это за память, из которой можно извлечь функциональную систему??? Достижение результата системы устраняет рассогласование между «потребностями» и микросредой нейронов. Подобная динамика характеризует ситуацию дефинитивного поведения. Ситуация научения в норме и восстановления в патологии (например, после инсультов, травматических и т.п. поражений мозга) специфична тем, что проблему согласования «потребностей» нельзя решить с использованием имеющихся у индивида способов согласования (т.е. в рамках имеющегося у индивида опыта). Рассогласование в данной ситуации отличается от того, которое имеет место в дефинитивном поведении: оно устраняется поиском и фиксацией в памяти новых вариантов объединения элементов, развертыванием процессов системогенеза (рис. 2).
Таким образом, имея в виду научение и слегка перефразируя утверждение Сократа, можно резюмировать содержание данного раздела следующим образом: рассогласование есть начало всей мудрости. – очень поэтично, но очень притянуто за уши…
От рассогласования через согласование к консолидации
В случае если процессы системогенеза протекают успешно, как уже отмечалось, формируются новые системы, устанавливаются и консолидируются новые межнейронные отношения, обеспечиваемые, в том числе, морфологическими перестройками (в основе которых - активация генетического аппарата) и обеспечивающие удовлетворение метаболических «потребностей» нейронов, а, следовательно, и их выживание.
Как выглядит поиск путей согласования метаболизмов клеток в процессе обучения при анализе «внешнего» поведения или электрической (суммарной и импульсной) активности мозга? На уровне поведения, если это не особый случай обучения с одной реализации, могут быть отмечены ориентировочно-исследовательское поведение, пробы, не приводящие к достижению полезного результата, а затем завершающиеся его достижением. Однако даже достижения результата не означают, что процессы научения завершены. Анализ актограмм, числа ошибок и пр. демонстрирует динамику. Через некоторое время, разное для разных видов поведения, показатели поведения стабилизируется. Следует заметить, что впечатление стабилизации пропадает, если рассматривать поведение под большим «увеличением». Каждый новый акт – не просто повторное извлечение из памяти, а созидание. Поведенческие акты обнаруживают направленную динамику (совершенствуются) в течение тысяч реализаций [92]. Изменчивость поведения при повторных реализациях может быть исследована и на нейронном уровне [8].
Что касается суммарной активности мозга, регистрируемой у человека, показано, что она претерпевает изменения не только в процессе обучения навыку, но и через часы (и дни) после того, как по поведенческому критерию испытуемый уже обучился [102,142]. Иначе говоря, скажем, через 30 минут и через 5 часов с момента начала реализации только что выученного поведения, активность мозга существенно различается, несмотря на то, что результативность поведения в процессе реализации существенно не меняется. Это говорит о продолжающихся процессах поддержания активности воспринятых образов. В экспериментах на животных также было показано, что как характеристики активаций нейронов, так и число активирующихся клеток меняются от первых реализаций поведения в течение последующих часов и дней [39,64,81,86,95,96,99,111,119,131,140]. Число клеток изменяется нелинейно. Направленность изменения зависит, по-видимому, от характера формируемого поведения, структуры, в которой регистрируется активность нейронов, и этапа, на котором регистрация производится.
Данные, полученные О.Е. Сварник [28], показывают, что число клеток мозга, в которых наблюдается экспрессия ранних генов, во много раз превышает число нейронов данной области, у которых обнаруживается специализация относительно системы формируемого поведения. Мы предполагаем, что часть из множества генетически активированных клеток – это нейроны, специализированные относительно систем ранее сформированных актов, и экспрессия в них отражает начало процесса аккомодационной реконсолидации. Большинство же этих клеток являются преспециализированными, и их генетическая активация является предпосылкой перехода клеток в состояние готовности для селекции в процессе проб. Именно в процессе проб из активированных (как генетически, так, вероятно, и импульсно) клеток отбираются те, которые перейдут далее в разряд специализированных относительно формируемой системы.
Отражением протекания описанной селекции и изменений, происходящих в нейронном обеспечении вновь сформированного поведения, является обнаруженное в наших пилотажных экспериментах (Е.А. Кузина и др.) изменение вариативности связи нейронов цингулятной коры крыс с новым поведением. Отмечена тенденция к увеличению числа нейронов, у которых активация появлялась в 100% случаев (т.е. при каждой реализации специфического для данной клетки акта) при сравнении совокупностей нейронов, активирующихся в инструментальном пищедобывательном поведении в течение первых дней и через 7-15 дней после завершения обучения. По-видимому, это изменение активности, уменьшение ее вариативности по мере консолидации памяти, может быть связано с завершением селекции и стабилизацией состава нейронов, вовлекающихся во вновь сформированное поведение. Полученные результаты мы рассматриваем как предварительные, но они согласуются с результатами экспериментов других авторов. Ranganath C. и Rainer G. [131] описывают данные, полученные при регистрации активности нейронов префронтальной коры у обезьян, которым предъявляли новые и знакомые объекты. Хотя предъявление нового объекта вызывает активации у большего числа нейронов, чем предъявление знакомого объекта, но в последнем случае активации существенно более четкие.
Однако данные литературы не позволяют свести объяснение изменения состава активирующихся нейронов только к описанной селекции. Например, не исключено, что некоторые нейроны могут активироваться на начальной стадии обучения, затем прекратить активность, а затем вновь начать разряжаться [95]. В то же время показано, что часть клеток начинают активироваться на сравнительно поздних этапах обучения, лишь в период стабилизации поведения (см., например, [161]), в то время как другие клетки, и это особенно важно для настоящего анализа, активируются лишь на начальных стадиях обучения, а когда поведение стабилизируется, их активность прекращается и больше не возникает [161,144]. Одно из возможных и, как мне кажется, вполне правдоподобное объяснение этого феномена сводится к следующему.
Более 30 лет назад J. B. Ranck [130] описал в гиппокампе группу клеток, которые активировались лишь в ситуации, когда в среде, к которой животное было адаптировано, происходили изменения: появление нового объекта или исчезновение привычного. В других областях мозга также были описаны нейроны, активации которых оказывались специфически связанными с ситуацией новизны. К структурам, особенно чувствительными к упомянутой ситуации, кроме уже названного гиппокампа, относят зрительную, энторинальную, нижнюю височную, префронтальную, цингулятную, моторную и премоторную области коры, миндалину и др. [131].
Сопоставляя эти наблюдения с только что упомянутыми данными о существовании нейронов, активирующихся лишь в начале обучения (ситуация новизны), можно предположить, что первые пробные акты осуществляются за счет коактивации переменных составов преспециализированных нейронов и нейронов, которые пока условно назовем нейронами «новизны». - детекторы нового по Соколову, фактически реализующие функцию распознавания новизны ситуации в восприятии, что имеет чрезвычайное значение для оценки ее значимости. И эта важность упускается почти повсеместно. Именно их коактивация обеспечивает как совершение проб, так и достижение при обучении первых положительных результатов. это – опрометчивое утверждение. мотивация существенно более сложно организована. После стабилизации поведения нейроны «новизны» прекращают активность. Это означает серьезную реорганизацию структуры актуализируемой памяти и, возможно, инициирует следующую, дополнительную волну селекции.
В рамках традиционного подхода можно было бы говорить о том, что роль этих нейронов – обеспечить внимание, внимание никак не обходится без них повысить уровень бодрствования и пр. и тем самым воздействовать на обучающиеся нейронные сети. Однако с позиций последовательного системного подхода такие и им подобные объяснения не могут быть приняты.
Как было отмечено выше, поведение осуществляется за счет реализации множества систем. При этом активация нейрона – показатель того, что в данном поведении актуализирована система, к которой он принадлежит. С этих позиций «новизна» - это не описание специализации нейронов, а лишь феноменологическое описание свойств их активности (ничем, правда, не худшее, чем огромное множество других: сенсорные нейроны, моторные нейроны, нейроны места, нейроны сознания ??? и пр.). Системное же описание требует указания систем, сформированных для достижения определенного результата, к которым данные нейроны принадлежат.
П.В. Симонов считал окончательно доказанным существование у животных исследовательского поведения, которое побуждается «самостоятельной потребностью» в обеспечении контакта с объектами, «прагматическое значение» которых не выяснено [31]. Это – очень характерно для популистов типа Симонова, объявлять нечто окончательно доказанным в пользу своей теории. О механизмах исследовательского поведения есть альтернативные представления: Механизм ориентировочного рефлекса.
Думаю, что суть возражений концепции автора вполне обозначена и продолжать комментировать то, что далее развивает эту явно слабо обоснованную концепцию, не стоит.
Для достижения каких результатов формируются системы упомянутого поведения?
Память может быть разделена на домены. В.Б. Швырков [37] подчеркивал, что то, элементы какого именно домена памяти будут актуализированы индивидом в поведении, определяется как мотивацией, так и тем, в какой среде он находится. Наиболее глобальное разделение доменов подразумевает противопоставление памяти, сформированной для осуществления поведений «приближения» и «избегания» («approach» и «withdrawal» [138]; см. подробнее в [44,56]).
Когда индивид попадает в незнакомую ситуацию, ситуацию с элементами неопределенности, первоочередной задачей является достижение следующего результата: отнесение ситуации к тем, которые позволяют совершать одно из поведений группы «приближения», или предполагают поведение «избегания». Это означает возможность или необходимость актуализации элементов памяти того или другого домена. Достижение упомянутого результата позволяет перейти к осуществлению поведения, направленного на достижение следующего результата в поведенческом континууме. Каков будет этот следующий результат, определяется и тем, какова мотивация индивида, и тем, каков результат исследовательского поведения, т.е. тем, элементы какого домена могут быть актуализированы. Можно полагать, что необходимость решения подобных задач существует и при устранении неопределенности, осуществляемой как выбор «внутри» упомянутых больших доменов, т.е. выбор между субдоменами. Нельзя исключить, что авторы классифицируют как «исследовательское» также и то поведение, реализуемое индивидом, которое в сходной, с точки зрения индивида, ситуации (first person perspective), приводило ранее к достижению полезного результата. Каков был этот результат, может быть неизвестно наблюдателю. Вероятно, по крайней мере, часть из нейронов, активирующихся лишь на начальных этапах процесса формирования памяти, принадлежат к системам исследовательского поведения, актуализируемым в процессе обучения вместе с создаваемыми системами нового поведения.
По-видимому, динамику описанных в настоящем разделе процессов возникновения рассогласования и его устранения за счет установления новых межклеточных отношений и нового соотношения организма и среды характеризует и феномен долговременной потенциации (ДП; LTP). Однако для того, чтобы обосновать возможную системную трактовку этого феномена, необходимо прежде остановиться на описании ситуации затянутого рассогласования.
Элиминация нейронов как «альтруистичный суицид»
Если рассогласование между «потребностями» нейронов и их микросредой не устранено, нейроны гиперактивны, экспрессия «ранних» генов затягивается: одна волна экспрессии сменяет другую. В этих случаях в нейронах могут экспрессироваться так называемые гены «смерти», активация которых ведет к гибели нервных клеток (рис. 2). Существующие экспериментальные данные рассматриваются как серьезный аргумент в пользу утверждения о связи между затянутой экспрессией «ранних» генов и программируемой смертью нейронов. Напротив, сравнительно краткосрочная экспрессия возникает у нейронов, которые выживают [141].
Итак, при наличии у организма опыта удовлетворения данных «потребностей» в данной ситуации, избирательно активируется память, имеющая отношение к удовлетворению данного набора «потребностей», и возникает импульсная активность нейронов, специализированных относительно актуализируемых элементов памяти - систем. Эта активность и есть нейронные основы реализации поведения. Достижение результата поведения на уровне соотношения целостного индивида и среды прекращает поведение, направленное на достижение этого результата, а на уровне отдельного нейрона оно выступает как устранение рассогласования между «потребностями» нейрона и микросредой.
В случае же отсутствия подобного опыта, когда повторные импульсации коактивированных нейронов не приводят к достижению результата, возникает экспрессия «ранних» генов. Эта экспрессия может быть рассмотрена как предпосылка для активации других транскрипционных компонентов – основы принимаемого клеткой «решения жить или умирать» [108, p.2736].
Следовательно, в случае возникновения рассогласования между «потребностями» нейрона и его микросредой и при невозможности устранить рассогласование в рамках имеющегося опыта, как в норме (в раннем онтогенезе и у взрослого), так и в патологии у него имеется, образно говоря, следующая альтернатива: измениться, вовлекаясь в системогенез (формирование новой системы, затем консолидируемой) или умереть (рис. 2). Вовлечение может носить характер системной специализации или аккомодационной реконсолидации (см. выше).
Излагаемые здесь соображения, в основе которых – представление об активности нейрона как детерминированной «потребностями» в метаболитах, согласуются с данным о том, что программируемая клеточная смерть запускается в условиях отсутствия соответствующих «факторов выживания» [132,164]. В то же время упомянутые представления не предполагают, что какое-то вещество всегда и при любых условиях производит одинаковый эффект. Напротив, ясно, что этот эффект должен зависеть от текущих «потребностей» клетки. Одно и то же вещество может как удовлетворить их в одном состоянии, так и привести к рассогласованию, если «потребности» изменились. Неудивительно поэтому, что трофические факторы, рассматриваемые как сигнал выживания, в определенном состоянии клетки могут превращаться в сигнал рассогласования и запускать программу клеточной смерти [164].
В рамках излагаемых представлений множественные повторные волны экспрессии «ранних» генов на начальных стадиях онтогенеза [118] возможно связать как с интенсивным морфогенезом и формированием все новых поведенческих актов (у многих животных за первые недели постнатального онтогенеза формируется больше половины актов всего поведенческого репертуара; см. в. [3]), так и с гибелью в этот период множества нервных клеток [118] (см. рис. 2).
Второй вариант развития событий, составляющих альтернативу «измениться или умереть», - смерть клеток - часто имеет место при созревании, а также в условиях патологии, при кардинальных изменениях микросреды клеток, в условиях, в которых оказывается невозможно использовать имеющийся у индивида опыт согласования метаболизмов клеток организма. Но не только в патологии и при созревании. Имеются данные, свидетельствующие в пользу апоптоза в мозгу здоровых взрослых индивидов, а также в пользу большого значения апоптоза нейронов нервной системы взрослого для функционирования целого организма [66,75,97,118,109,148].
Эти данные при учете представления о том, что системогенетические закономерности являются общим принципом реализации процессов созревания, научения на любом этапе онтогенеза, адаптации и восстановления в патологии, позволяют предположить, что упомянутая альтернатива существует и в норме. И что элиминация нейронов как один из исходов нейроселекции в раннем онтогенезе, значение которой для формирования поведенческого репертуара не вызывает сомнений, вносит вклад и в системогенез у взрослого (рис. 2; Знак вопроса на схеме у слова «смерть» обозначает гипотетичность представления о смерти нейронов как факторе системогенеза.).
Следовательно, формулируемая позиция сводится не к альтернативе «системогенез или смерть», а, коротко говоря, к двум взаимосвязанным путям обеспечения системогенеза: модификация нейрона или его гибель. Блокирование любого из них нарушает системогенетические процессы. Таким образом, здесь подчеркивается именно позитивный, в общеорганизменном плане, аспект гибели нейронов.
Фатальный для отдельных клеток исход - гибель – можно представить себе в качестве неизбежной платы за возможность осуществления успешного системогенеза на протяжении всего индивидуального развития. Неизбежной по крайней мере, в тех случаях, когда метаболические «потребности» каких-либо клеток вступают в неустранимое противоречие с новыми способами согласования «потребностей» клеток индивида. Формирование этих способов диктуется необходимостью соответствовать изменившимся условиям внешней и/или внутренней среды и выражается в образовании новых систем и изменении межсистемных отношений.
Можно предположить также, что особенно выраженное нарастание частоты смерти нейронов обнаружится при таком обучении, когда индивид долго не способен решить сравнительно сложную проблему, в частности, когда индивид оказывается в «неизбегательной» ситуации, что в субъективном плане может выражаться в депрессивном состоянии.
Выше представление об активном нейроне было противопоставлено представлению о нейроне реактивном. Здесь я хотел бы подчеркнуть, что принцип активности распространяется на весь период и на все аспекты существования нейрона, включая и процессы, связанные с реализацией альтернативы: измениться или умереть. Данная позиция находится в соответствии с точкой зрения о том, что каждый из этапов элиминации клетки является активным [132] и что, по существу, элиминация является суицидом [109,164].
Можно добавить, что этот суицид альтруистичен. В том смысле, что клетка включает программу самоэлиминации для того, чтобы таким образом устранить метаболическое противоречие, которое неустранимо другим путем, и обеспечить успешную адаптацию индивида к изменившимся условиям. А значит, обеспечить выживание других клеток, принадлежащих к тому же клону.
Ранее другими авторами уже приводились аргументы в пользу существования «альтруистичного клеточного суицида» в нервной системе: при инфицировании нейротропным вирусом. Альтруистичный суицид рассматривается как стратегия мультиклеточного организма, ограничивающего размножение вируса путем самоуничтожения инфицированных клеток [49].
У альтруизма клеток многоклеточного организма имеются эволюционные предпосылки. Описана альтруистическая гибель у одноклеточных (амебы Dictyostelium discoideum), которые приносят себя в жертву другим клеткам своего клона (чужого – менее охотно), обеспечивая существование временно формирующегося многоклеточного образования за счет формирования нежизнеспособного стержня, вокруг которого организуется колония. Остальные (около 80 %) клетки превращаются в жизнеспособные споры, составляющие это образование [151].
Долговременная потенциация: полезный артефакт?
Начну этот раздел с формулировки, отражающей мое понимание феномена ДП, а затем приведу аргументы в пользу этого понимания, рассмотрев ДП с позиций системного подхода: ДП – артефакт, который по не предусмотренным его исследователями причинам может иметь отношение к механизмам научения и памяти.
Долговременная потенциация синаптической эффективности (LTP) в результате тетанического раздражения проводящих путей в гиппокампе считается претендентом на роль физиологического механизма долговременной памяти и рассматривается в качестве экспериментальной модели пластичности, зависимой от активности. В ряде исследований, (см., например, [17,91,104,107,114]), показано, что ДП возникает не только в гиппокампе, но и в корковых структурах, не только при исследовании препаратов, но и у бодрствующих свободно подвижных животных.
В той области знания, которая является предметом анализа в настоящем сообщении, исследования ДП рассматриваются в течение ряда лет как наиболее важные и приоритетные. И это не только потому, что данный феномен хорошо укладывается в традиционную схему, описывающую формирование памяти как повышение синаптической эффективности в рефлекторных дугах. J. W. Lichtman и J.R. Sanes [110] отмечают существование также и факторов, которые они называют «социологическими». «Главной наградой для многих ученых является публикация в журналах с наиболее высоким [импакт фактором], – пишут авторы - [и ДП] как раз та область исследований, которая используется этими журналами, чтобы протрубить о новых данных». Однако далее они отмечают, что «пока будет существовать именно такая награда, до тех пор она будет порождать скорее поддержание дискуссий, но не решение проблемы», инициируя бесконечное обсуждение того, что « [природа] ДП – пресинаптическая; [природа] ДП – постсинаптическая; нет, пресинаптическая; нет, постсинаптическая; эта киназа вовлечена; та киназа вовлечена ...» [110, p.928]. К упомянутым факторам можно добавить еще и следующий: «поддержание интереса к ДП, несмотря на раздражающую критику со стороны некоторых исследователей, которую изучающие ДП обычно приписывают зависти, [объясняется] отсутствием другой [лучшей] игрушки» [112, p.923]
С системной позиции феномен ДП может быть рассмотрен как электрофизиологическое описание процесса рассогласования. Действительно, исходя из системной концепции детерминации активности нейрона (см. выше), он не реагирует на входную импульсацию, а активируется, подготавливая будущий приток и подготавливаясь к нему. Можно полагать поэтому, что «незапланированный», «неожиданный» приток вызовет рассогласование и, следовательно, активность нейрона, направленную на устранение этого рассогласования. Если так, то, по-видимому, искусственная тетаническая электростимуляция служит мощным фактором рассогласования. И повышенная активность клетки, сохраняющаяся при тестировании – отражение данного рассогласования. Поэтому логично думать, что ДП, полученная в подобных условиях, имеет артефактную природу.
В пользу связывания ДП с процессом рассогласования свидетельствуют, кроме теоретических соображений, и экспериментальные данные. Обнаружено явное соответствие ДП тем процессам, которые имеют место в патологии при стойких отклонениях свойств метаболической среды. С использованием метода регистрации отдельных нейронов задних рогов спинного мозга крыс с экспериментально вызванным воспалением (моноартритом) показано, что стимуляция С-волокон у них значительно легче вызывает ДП, чем у здоровых животных. Авторы считают, что «механизмы, лежащие в основе ДП, сходны с теми, которые обусловливают сенситизацию при периферическом воспалении» [159, p.51].
Итак, хотя тетанизирующие экспериментаторы, не планируют вызвать рассогласование, именно его они вызывают. А рассогласование есть, как аргументировалось выше, первый этап научения, формирования новой памяти. Именно в этом смысле я и рассматриваю ДП как феномен, имеющий, возможно, отношение к механизмам научения и памяти. Имеющий вовсе не потому, что моделирует «повышение эффективности синаптической проводимости». А имеющий «возможно» потому, что неизвестно, обладает ли получаемое при индукции ДП рассогласование теми свойствами, которые присущи естественному рассогласованию при научении. Поэтому в названии настоящего раздела присутствует вопрос.
Shors T.J. и Matzel L.D [145] на основе подробного анализа экспериментальных данных о свойствах ДП пришли к заключению о несоответствии свойств ДП тем, которые должны были бы быть, если бы ДП обеспечивала сохранение долговременной памяти. В качестве наиболее серьезного сомнения при рассмотрении ДП в качестве механизма долговременной памяти авторы рассматривают ее недостаточную длительность. Да, ДП длится часы и дни, но достаточно ли это долго, чтобы обеспечить поддержание долговременной памяти? - спрашивают авторы. (К сомнениям авторов в достаточности длительности ДП для рассмотрения ее в качестве механизма долговременной памяти добавлю, что и та длительность, которая получается в подавляющем большинстве экспериментов с тетанической стимуляцией, с подведением БАВ и последующим тестирующим раздражением и которую обсуждают критики, возможно больше периода повышенной возбудимости, имеющего место в натуральных условиях (см. далее о работе [43]). Длительность может быть повышена потому, что у индивида не существует фило- или онтогенетически приобретенной памяти об устранении рассогласования, возникающего при электрической стимуляции мозга или инъекции значительных объемов БАВ.) Авторы приходят к выводу о том, что ДП - механизм, имеющий отношение не к поддержанию долговременной памяти, но к начальному периоду ее формирования и связан с механизмами пробуждения и внимания. Shors T.J. и Matzel L.D решают проблему не так, как предлагается здесь, но и они связывают ДП именно с инициацией процессов научения.
Для потока исследований ДП более шести лет, прошедшие с момента опубликования обзорно-теоретической статьи Shors T.J. и Matzel L.D, - большой срок. Однако и в настоящее время можно считать неустраненными противоречия между постулируемым механизмом долговременной памяти и свойствами ДП. Подтверждения того, что ДП моделирует механизм, лежащий в основе сохранения памяти за счет изменения эффективности синаптической проводимости, «отсутствуют сегодня», как и 10 лет назад, - констатируют J. W. Lichtman и J.R. Sanes [110, p.928]. Но «когда люди ищут что-нибудь, они часто это находят, даже если этого нет» [110, p. 929], - замечают они.
На фоне множества работ, проанализированнах Shors T.J. и Matzel L.D, как успешную находку можно оценить исследование W.C. Abraham et al. [43]. Авторы смогли подобрать условия, при которых ДП в гиппокампе крыс поддерживалась в течение нескольких месяцев. С традиционных позиций эти данные можно рассмотреть как подтверждение того, что ДП-подобный механизм может участвовать в поддержании долговременной памяти. (Правда, при этом возникают опредленные противоречия между тем, что феномен особенно длительной ДП обнаружен именно в гиппокампе, и представлением об участии гиппокампа лишь в начальных этапах поддержания памяти.)
Но с системных позиций можно рассмотреть данные W.C. Abraham et al. так, что они, в отличие от множества других авторов, лишь смоделировали такую ситуацию, в которой рассогласование, индуцированное тетанизацией, не устраняется сравнительно долго. Если принять последнюю точку зрения, то следует думать, что авторы получили рассогласование, длительность которого существенно превышает ту, которая имеет место в натуральных условиях. Это бы означало, иначе говоря, что с длительностью соответствующих процессов научения более сопоставимы данные авторов, демонстрирующих сравнительно короткоживущую ДП, чем результаты W.C. Abraham et al. Интересно, что W.C. Abraham et al. сами высказывают вполне обоснованное, с моей точки зрения, сомнение в том, что «механизмы памяти в реальном мире требуют такой стабильной пластичности»[43, p.9632].
В данной работе показано также, что помещение животных в обогащенную среду после индуцирования ДП, приводит к редукции последней. (Можно предположить, что процессы, индуцируемые помещением в обогащенную среду, накладываются на уже развернутое рассогласование, вызванное индукцией ДП, и облегчают поиск новых способов согласования клеток. То есть этот поиск происходит в контексте устранения «естественного» рассогласования.) Имея в виду упомянутую редукцию, W.C. Abraham et al. заключают, что « в норме при воздействии реальной и изменяющейся среды ДП в гиппокампе … может быть существенно менее длительным» (short-lasting) [43, p.9632] .
Исходя из предположения о ДП как электрофизиологическом описании процесса рассогласования, можно думать, что даже в тех случаях, когда речь идет о длительности, измеряемой не месяцами, а днями, то, хотя такая длительность недостаточна для рассмотрения ДП в качестве феномена, отражающего механизмы долговременной памяти, она в части случаев достаточна для того, чтобы рассмотреть ее как отражение затянутого рассогласования, ведущего к гибели нейронов. Проще говоря, логика предлагаемых здесь представлений допускает и даже предполагает возможную связь ДП с гибелью нейронов. В литературе существуют данные, свидетельствующие в пользу этого предположения.
Рассогласование, как уже отмечалось выше, может быть вызвано не только электрической стимуляцией, но и введением БАВ. Введение агониста метаботропных рецепторов mGluR: ACPD [1S,3R]-1-aminocyclopentane-1,3-dicarboxylate in vitro и in vivo индуцирует развитие медленно начинающейся (slow-onset) ДП в CA1, выражающейся при внутриклеточной регистрации в увеличении амплитуды ВПСП [58]. В лаборатории K.G. Reymann [113] выясняли, «каковы последствия этого феномена для клеток интактного гиппокампа». Авторы пришли к выводу о существовании явной связи « этой синаптической пластичности с патологическими процессами в гиппокампе in vivo» [113, pp.487, 493]. ДП, которая длилась более 4 часов и индуцировалась превышающей пороговую концентрацией ACPD (20 nmol) , влекла за собой выраженную гибель клеток CA1. Концентрации, которые не индуцировали ДП, не оказывали (или оказывали незначительный) нейротоксический эффект. Авторы отмечают, что гибель клеток может иметь место и в экспериментах других авторов, в которых индуцируется ДП.
Выдвижение и селекция гипотез при научении
Выше говорилось о том, что рассогласование устраняется путем проб и поиска новых путей достижения результата поведения. Может ли произойти селекция нейронов, их специализация и фиксация новой системы в памяти без реализации внешне наблюдаемого («внешнего») поведения? Если да, то как быть с положением о том, что новая интеграция фиксируется достигнутым результатом?
Описание соотношения организма со средой в новой ситуации как процесса, включающего выдвижение и селекцию гипотез, было в яркой форме представлено К.R. Popper [127]. D. Dennett [73], рассматривая гипотетическое «скиннеровское» существо, осуществляющее отбор одного удачного акта из ряда путем реализации последовательных проб «вслепую», отмечает, что каждая из таких проб может привести к гибели существа. Автор справедливо считает более эффективным способом формирования нового опыта соотношения организма со средой предварительную, «внутреннюю» селекцию актов, которую осуществляет «попперовское» существо. «Внутренняя» селекция делает возможной ситуацию, в которой, по словам К.Поппера, вместо нас гибнут наши гипотезы. Плодотворность идей К. Поппера о формировании и селекции гипотез для разработки представлений о механизмах научения отмечал П.В. Симонов [30]. В настоящее время эти идеи эффективно используется при формулировке новых концептуальных схем в когнитивных науках.
Нарастание способности совершения проб и ошибок «в уме», без реализации их во «внешнем» поведении, рассматривалось Л.В. Крушинским [23] в качестве показателя развития поведения в филогенезе. В области практической деятельности человека (спортивной, музыкальной и т.д.) реальность этой стадии формирования памяти уже давно была осознана. Анализ процесса освоения нового музыкального произведения или обучения новому акробатическому движению приводит, соответственно, к заключениям о том, что каждый пассаж должен быть «готов психически», прежде чем он будет испробован на рояле [18], и что чем лучше занимающийся представит изучаемое акробатическое движение [22], тем быстрее будет происходить процесс обучения.
Следует полагать, что выдвижение акта-гипотезы и его тестирование во «внутреннем плане», т.е. проверка гипотезы на соответствие сложившейся к данному моменту структуре памяти индивида (пробная организация совместной активности новой совокупности нейронов), занимают определенный временной интервал. Величина этого интервала зависит от многих обстоятельств: степень новизны гипотезы, сложность уже имеющегося у индивида опыта и др. Основываясь на многочисленных данных, показывающих значительное сходство паттернов мозговой активности при воображении или наблюдении действия и при реальном его осуществлении [83,98], можно допустить, что составы активированных нейронов при тестировании гипотезы во «внутреннем» и затем во «внешнем» планах существенно, хотя, конечно, не целиком, перекрываются.
В результате тестирования фиксируется новая интеграция и инициируется изменение структуры памяти. В литературе имеются данные, которые свидетельствуют в пользу возможности подобной фиксации [128]. В экспериментах с регистрацией активности нейронов у обезьян показано, что характеристики импульсации существенно изменяются уже на этапе, когда животное нашло правильное решение (в следующих после этого изменения реализациях поведения животное с вероятностью, равной единице, начинает действовать безошибочно), но еще ни разу не проверило его реализацией «внешнего» поведения, завершающегося подачей пищи. Сходные закономерности, естественно, могут быть выявлены и у людей, формирующих новых поведенческий акт. Так в экспериментах с испытуемыми, участвующими в стратегической игре (крестики-нолики), до первой реализации нового для игрока хода обнаруживаются реорганизации поведения: время выбора ходов и, как предполагается, число актуализируемых в игре связанных элементов памяти изменяется [2].
Таким образом, можно считать, что фиксация новой интеграции не обязательно требует достижения результата внешне наблюдаемого поведения. Но это, однако, не означает, что нарушается одно из основных положений теории функциональных систем: о результате как системообразующем факторе. В качестве системообразующеого фактора в этом случае выступает результат тестирования гипотезы во «внутреннем» плане.
Нет оснований думать, что консолидация памяти, лежащая в основе описанной фиксации интеграции, происходит мгновенно. В связи с этим возникает вопрос, отличаются ли стадии консолидации памяти в этом случае и, если отличаются, то как, от стадий консолидации памяти, формирующейся после реализации результативного «внешнего» поведения. Получение ответа на этот вопрос важно для понимания закономерностей формирования памяти.
В части случаев вновь сформированная интеграция как таковая не предполагает реализацию специального поведения для своего тестирования во «внешнем» плане; многие «внутренние действия» не «подлежат последующей экстериоризации» [20, с.171]. Подобные зафиксированные интеграции могут входить, по-видимому, в состав «семантической памяти».
В других случаях прошедший «внутреннее» тестирование акт-гипотеза формируется для того, чтобы совершить то или иное конкретное «внешнее» поведение (внешнее тестирование). Здесь после внутреннего тестирования могут иметь место минимум два исхода: такое поведение будет реализовано или нет. В первом случае, как и в ситуации с «внутренним» тестированием, временной интервал между последним и «внешним» тестированием может варьировать, причем очень сильно: от миллисекунд до лет. В течение этого интервала в памяти хранится «латентная» система.
Будет ли вновь сформированный акт реализован во внешне наблюдаемом поведении или нет, в результате тестирования фиксируется новая интеграция, т.е., как было сказано выше, происходит изменение структуры памяти, включающее аккомодационную реконсолидацию (см. выше). Логично предположить, что такое изменение структуры не может не сказаться на характеристиках реализации ранее сформированного поведения.
Поведение может измениться как до тестирования гипотезы во «внешнем» плане, так и до осознания результатов этого тестирования. Результаты экспериментов с участием испытуемых, выбирающих стратегию поведения в сложной среде, позволяют считать, что модификация поведения может происходить не только в интервале между проверкой гипотезы во «внутреннем» и «внешнем» планах, но и между проверкой гипотезы во «внешнем» плане и осознанием результатов этой проверки. Показано, что испытуемые после ряда тестирующих проб начинают адекватную модификацию своего поведения, соответствующую закономерностям организации среды, еще не будучи способными дать отчет об этих закономерностях [55]. Осознанию, «интеллектуальному» решению, как это было показано около тридцати лет назад О.К. Тихомировым и его сотрудниками[33], непосредственно предшествует стадия «эмоционального предрешения», когда у человека возникает ощущение, что им найден принцип решения задачи, но он еще не может его сформулировать. Эта стадия характеризуется «эмоциональным напряжением» и соотносима с первой «эмоциональной» стадией научения [31].
Вероятно, характер процессов консолидации памяти должен зависеть от величины временного интервала между «внутренним» и «внешним» тестированием. Если консолидация после внутреннего тестирования уже завершилась, ее тестирование во «внешнем» плане связано с модификациями, сопоставимыми с теми изменениями, которые имеют место при реконсолидации вследствие реактивации памяти. По-видимому, результирующая структура памяти, состояние её нейронного и молекулярно-биологического обеспечения будут разными в зависимости от того, произошло ли «внешнее» тестирование и, если да, то с какой задержкой после «внутреннего». Это предположение может быть проверено экспериментально.
Индивидуальное развитие как последовательность системогенезов
Наконец, последняя проблема, на которой мне хотелось бы остановиться: сравнить особенности системогенеза на последовательных этапах индивидуального развития, затронув вопрос о видоспецифическом и индивидуальноспецифическом поведении.
В основе процесса специализации нейрона при научении находится экспрессия «ранних» и затем «поздних» генов, ведущая к изменению структуры нейрона и его метаболических «потребностей» (см. выше). Если принять, что при формировании новой специализации нейронов в процессе научения используется очередной, новый вариант реализации индивидуального генома, то с позиций высказанных соображений индивидуальное развитие может быть представлено как последовательность системогенезов и «актуализация» генома, связанная с системогенезами.
Выше становление системной специализации нейрона в процессе научения было сопоставлено с формированием вторичного ассортимента, а формирование первичного ассортимента рассмотрено как образование преспециализаций нейронов в процессе раннего онтогенеза. Тогда научение – это «расходование» сформированных преспециализаций путем перевода их в конкретные специализации относительно вновь формируемых систем. Возможно, это «расходование» является одним из факторов, определяющих экспериментально выявляемый феномен понижения уровня экспрессии «ранних» генов с возрастом [108].
Преспециализация нейронов, предназначенных для систем видоспецифических актов, сравнительно жестко детерминирует, относительно системы какого конкретного акта они будут специализированы при научении. Однако эта детерминация не однозначна. Формирование видоспецифических актов зависит от особенностей ситуаций, в которых протекает развитие индивида. Так показано, что если крысят сразу после рождения лишить контакта с соском матери или искусственным соском и давать им молоко только вылитое на дно контейнера, то вместо поведения сосания, являющегося в норме исходным для дифференциации всего последующего пищевого поведения [139], у них формируется пищедобывательное поведение, сильно отличающееся от сосания [94]. Известно, что в норме у детей первичная вокализация - лепет - является доречевой стадией формирования речи. У немых детей, которые с рождения подвергаются воздействию жестовой речи, вместо вокализационного наблюдается «мануальный лепет»: они продуцируют отдельные «речевые жесты» и их бессмысленные последовательности [125]. С помощью картирования активности мозга взрослых обнаружено, что структуры мозга, активные в норме при голосовой речепродукции, активируются и у немых при жестовой речи [67]. Вероятно, в значительной степени перекрывающиеся, хотя и не идентичные, группы нейронов специализируются в отношении «систем речепродукции», которые оказываются «вокализационными» в норме или «жестовыми» у немых.
Менее ясно, на каком «языке» «написана» преспециализация нейронов, предназначенных для формирования индивидуальноспецифического поведения у взрослого индивида. Предположим, если один экспериментатор учит животное нажимать на педаль для получения пищи, он обнаруживает нейроны, специализированные относительно системы этого акта. Другие экспериментаторы после обучения животного инструментальному пищедобывательному акту потягиванию зубами кольца находят нейроны, специализированные относительно системы этого акта. Алфавит, используемый для «наименования» преспециализаций, конечно, задан (ограничен) филогенетическим опытом. Данный фактор учитывается экспериментатором в той форме, что он подбирает задачи, выполнению которых индивид данного вида может обучиться. Это, однако, не означает, что упомянутые выше инструментальные акты, а также и все другие акты, которые может придумать экспериментатор, заранее предопределены соответствующей преспециализацией. Скорее, говоря об индивидуальноспецифических актах, следует полагать, что отдельные группы преспециализаций предназначаются для последовательных стадий индивидуального развития на протяжении всей жизни. И язык преспециализаций, предназнчаенных для индивидуальноспецифических актов, есть язык стадий, отвлеченный от их конкретного индивидуальноспецифического содержания. Это содержание может быть описано через уникальный набор актов, формируемых данным индивидом в специфических условиях его существования. Язык преспециализаций становится языком конкретного акта в результате обучения этому акту и формирования специализаций нейронов в отношении системы этого акта.
Сказанное не следует понимать так, что какой бы акт ни сформировался на данной стадии, относительно системы этого акта окажется специализированной одна и та же группа нейронов. Выше уже говорилось о разных доменах памяти. Набор специализирующихся клеток, по-видимому, зависит и от того, какой именно домен пополняется за счет нового системогенеза. Ситуация усложняется тем, что обогащение разных доменов происходит неравномерно, т.е., упрощенно говоря, внутри разных доменов индивид находится на разных стадиях развития. Кроме того, можно предположить, что и при формировании систем разных поведенческих актов, относящихся к одному домену, но по-разному вовлекающих тело индивида, например, актов нажатия на педаль и потягивания зубами кольца, формируются специализации не одного и того же набора клеток.
Косвенным аргументом в пользу первого предположения служат данные о том, что смещение одного и того же рычага для получения пищи и для избегания электрокожного раздражения обеспечивается разными наборами активирующихся нейронов у обезьян [106,122]. В пользу второго – полученные у нас в лаборатории данные о том, что наборы клеток, активирующихся при нажатии на педаль и потягивании кольца для получения пищи, существенно перекрываются, но не являются одинаковыми [52].
Следует, однако, заметить, что упомянутые выше «специфические условия его существования», оставаясь уникальными, имеют в то же время много сходного с условиями, в которых развиваются другие особи данного вида. Биологическая и культурная эволюция, могут быть рассмотрены как аспекты единого процесса ген-культурной коэволюции. С одной стороны, индивиды данного вида вносят вклад в конструирование ниши а, с другой - ниша детерминирует давление отбора на них и их потомков [71], так что набор преспециализаций у особей данного вида оказывается в определенной степени нише- (культуро)зависимым [45]. Упрощая, ниша зависит, в том числе и от специализаций нейронов, а паттерн специализаций и от ниши.
Итак, набор и последовательность формируемых актов у особей, принадлежащих к данной популяции, оказываются во многом сходными. Поэтому, хотя язык преспециализаций – не язык конкретных актов, но, тем не менее, на соответствующей стадии индивидуального развития упомянутых особей, эти преспециализации трансформируются не в любые, а в довольно определенные специализации.
Весьма вероятно, что преспециализации нейронов, сформированные в раннем онтогенезе, не остаются неизменными в течение всей жизни до мемента формирования специализации этих нейронов. То есть до перехода из первичного во вторичный ассортимент. Логично полагать, что описанная выше процедура селекции при научении, затрагивающая множество клеток (существенно большее число, чем окажется специализированным после этого научения), не проходит бесследно и для тех нейронов, которые не отбираются для формирования специализации в отношении системы, образуемой в процессе данного научения. Иначе говоря, возможно, процедура научения модифицирует и нейроны «резерва», приводя их характеристики в соответствие с произошедшими изменениями структуры памяти. Если это так, то подобная модификация могла бы быть рассмотрена в качестве одного из факторов, обусловливающих феномен «переноса».
В литературе имеются попытки периодизации развития структуры личности, в том числе личности взрослого человека. Так, Э. Эриксон выделяет универсальные для человечества периоды (с 20 лет до смерти): ранней, средней и поздней зрелости, полагая, что эти периоды представляют собой эпигенетическое развертывание наследуемого генетически «плана личности» [35]. Каждый из указанных периодов включает, конечно, множество стадий, подчеркивая их некоторую общую характеристику. Выделить и описать все эти стадии пока невозможно. В то же время представляется логичным думать, что эти стадии могут быть описаны через число пройденных системогенезов, общее число сформированных систем и характер отношений (оппонентность, синергия и т.п.), сложившихся между системами и их группами и характеризующих структуру памяти на данной стадии.
По-видимому, имеется определенное соответствие «номера» стадии и возраста индивида. Но оно - неоднозначное: у разных индивидов скорости развертывания стадий, темпы индивидуального развития различаются [21].
«Вписать» вновь формируемую систему в структуру, содержащую на данном этапе индивидуального развития N взаимосвязанных систем, и в более сложную структуру, содержащую на более позднем этапе 100xN систем – разные задачи. Скорее всего, для этого нужны нейроны с разными свойствами (разными преспециализациями), с разными морфологическими связями. Эти различия свойств и связей могут быть одним из ключевых факторов, обусловливающих различия мозгового обеспечения «одного и того же» поведения, формируемого на ранних и поздних стадиях индивидуального развития, что выявляется как у животных при регистрации активности нейронов[154], так и у людей в экспериментах с картированием мозга. Так, при изучении с помощью ПЭТ активности отдельных структур мозга, вовлекающихся в обеспечение решения вновь освоенной экспериментальной задачи зрительной дискриминации, обнаруживается, что у молодых и пожилых испытуемых при решении данной задачи с одинаковой эффективностью согласованно активируются разные наборы структур [72].
Изучение и сравнение (в том числе и межвидовое) языков видоспецифических и индивидуальноспецифических преспециализаций представляется перспективной задачей.
Заключение
Проведенное выше обсуждение, как мне представляется, свидетельствует в пользу того, что из двух предположений, описывающих формирование и консолидацию памяти как постепенный, по «кускам» сбор мозаики вплоть до появления целостной «картины» памяти или как последовательность качественно различных фаз научения [76], второе описание соответствует реальности в большей степени. Это обсуждение позволяет представить следующую гипотетическую последовательность процессов формирования и функционирования памяти.
Научение начинается с рассогласования между потребностями индивида и теми возможностями, которые предоставляет ему сформированная к данному моменту структура памяти. Это рассогласование проявляется на клеточном уровне в несоответствии метаболических «потребностей» клетки и метаболического притока, ею получаемого. Рассогласование, которое в дефинитивном поведении устраняется совместной активностью клеток, объединенных в систему, представляющую собой элемент памяти, не может быть таким образом устранено в новой ситуации. То есть имеющийся у индивида опыт совершения поведения в новой ситуации оказывается непригодным.
Рассогласование, которое не может быть устранено посредством актуализации имеющейся памяти, приводит к экспрессии «ранних» генов. Экспрессия затрагивает, возможно, как преспециализированные клетки, так и некоторые клетки, ранее сформировавшие свою специализацию. Электрофизиологическим проявлением этих процессов является повышение чувствительности синапсов к притоку, которое в искусственных условиях моделируется как долговременная потенциация (ДП).
Рассогласование подобного рода может быть устранено формированием новой системы, т.е. нового элемента памяти. Формирование новой интеграции, которой предшествует «внутреннее» тестирование и отбор гипотез, выражается в пробах. На клеточном уровне эти пробы означают пробные объединения активирующихся клеток. В процессе проб осуществляется селекция: из множества активировавшихся преспециализированных клеток, в том числе и появившихся вследствие неонейрогенеза, отбирается соответствующая совокупность. Это такая совокупность, коактивация которой с ранее специализированными клетками, в том числе клетками исследовательского поведения, приводит к достижению необходимого поведенческого результата. На клеточном уровне достижение этого результата выступает как удовлетворение метаболических «потребностей» клеток и устранение рассогласования.
При развертывании селекции нейрон оказывается перед выбором одного из двух путей участия в системогенетических процессах: измениться, вовлекаясь в формирование новой системы, затем консолидируемой, или умереть. Элиминация клеток может рассматриваться как «альтруистичный суицид». Он совершается для преодоления вновь возникших и неустранимых другим путем метаболических противоречий между клетками и обеспечивает выживание других клеток, принадлежащих к тому же клону.
После достижения первых результатов клетки, специализированные относительно исследовательского поведения, постепенно уменьшают и прекращают активность. Это, возможно, проявляется как во временном изменении внешнего поведения, казалось бы, уже сформированного, так и в новом изменении состава активирующихся преспециализированных клеток. Постепенная стабилизация состава активирующихся нейронов выражается в более стабильном соотношении активаций нейронов и поведения.
Экспрессия «поздних» генов обеспечивает реорганизацию отобранных клеток, превращение их в специализированные относительно вновь сформированной системы. Она же в процессе аккомодационной реконсолидации модифицирует ранее специализированные клетки. Эта модификация ранее специализированных клеток обусловлена включением вновь сформированной системы в существующую структуру памяти индивида. Их специализация не изменяется, но теперь эта система, наряду со сформированной при обучении новому поведению дополнительно вовлекается и в обеспечение этого поведения.
Пожизненность сформированных специализаций означает пожизненность памяти. Однако не неизменность. Кроме изменений, обусловленных аккомодационной реконсолидацией при научении, память претерпевает модификации, связанные с ее реактивацией.
Благодарность
Благодарю за полезные комментарии К.В. Анохина, Л.И. Александрова, О.Е. Сварник, А.К. Крылова и Е.А. Кузину.
Поддержано РФФИ [№ 02-04-49443], РГНФ [№ 02-06-00011] и Советом по грантам Президента РФ ведущим научным школам РФ [№ НШ-1989.2003.6].
Литература
1. Аверкин Р.Г., Александров Ю.И., Гринченко Ю.В., Мац В.Н., Созинов А.А. Активность нейронов антеролатеральной области моторной коры мозга кролика при захвате пищевых и непищевых объектов в инструментальном поведении//Журнал высш. нерв. деят. 2001. Т. 51. N 6. С. 652-657.
2. Александров И.О., Максимова Н.Е. О виртуальности компонентов индивидуального знания на ранних стадиях их формирования//Виртуальная реальность. М. Российская Академия Искусственного Интеллекта. 1998. С. 61 - 82.
3. Александров Ю.И. Психофизиологическое значение активности центральных и периферических нейронов в поведении. М. Наука. 1989.
4. Александров Ю.И. Введение в системную психофизиологию //Психология XXI века. Дружинин В.Н. ред. М. Пер Се. 2003. С. 39-85.
5. Александров Ю.И., Александров И.О. Активность нейронов зрительной и моторной областей коры мозга при осуществлении поведенческого акта с открытыми и закрытыми глазами //Журн. высш. нервн. деят. 1980. Т. 31. N 6. С. 1179-1189.
6. Александров Ю.И., Греченко Т.Н., Гаврилов В.В., Горкин А.Г., Шевченко Д.Г., Гринченко Ю.В., Александров И.О., Максимова Н.Е., Безденежных Б.Н., Бодунов М.В. Закономерности формирования и реализации индивидуального опыта//Журнал высшей нервной деятельности. 1997. Т.47. N 2. С. 243-260.
7. Александров Ю.И., Брушлинский А.В., Судаков К.В., Умрюхин Е.А. Системные аспекты психической деятельности. М. Эдиториал УРСС. 1999.
8. Александров Ю.И., Шевченко Д.Г., Горкин А.Г., Гринченко Ю.В. Динамика системной организации поведения в его последовательных реализациях//Психол. журн. 1999а. Т. 20. N 2. С. 82-89.
9. Анохин К.В. Обучение и память в молекулярно-генетической перспективе //Двенадцатые сеченовские чтения. 1996. М. Диалог-МГУ. С. 23-47.
10. Анохин К.В. Молекулярные сценарии консолидации долговременной памяти // Журн.высш.нервн.деят. 1997. Т.47. N 2. С. 261-280.
11. Анохин К.В., Судаков К.В. Системная организация поведения: Новизна как ведущий фактор экспрессии ранних генов в мозге при обучении //Успехи Физиологических Наук. 1993. Т. 24. N.3. С. 53-70.
12. Анохин П.К. Очерки по физиологии функциональных систем. М. Медицина. 1975.
13. Анохин П.К. Проблема компенсации нарушенных функций и ее значение для клинической медицины. Сообщение I. //Хирургия. 1954. N. 10. С. 758-769.
14. Бернар К. Лекции по экспериментальной патологии. М.-Л. Биомедгиз. 1871/1937.
15. Бернштейн Н.А. Очерки по физиологии движений и физиологии активности М. Медицина. 1966.
16. Горкин А. Г., Шевченко Д.Г. Различия в активности нейронов лимбической коры кроликов при разных стратегиях обучения //Журн. высш.нервн.деят. 1995. Т.45. N 1. С. 90 - 100.
17. Горкин А.Г., Рейманн К.Г., Александров Ю.И. Долговременная потенциация и вызванные спайковые ответы в цингулятной коре свободноподвижных крыс//Журн. высш.нервн.деят. 2002. T. 52. N 6. C. 675-685.
18. Гофман И. Фортепианная игра. М. 1911 [цит. по Дымерский В.Я. О применении воображаемых действий в процессе восстановления и сохранения навыков//Вопросы психологии. 1955. N 6. С. 50].
19. Давыдовский И.В. Общая патология человека. М. Медицина. 1969.
20. Дубровский Д.И. Психические явления и мозг. М. Наука. 1971.
21. Емельянов С.В. Темп индивидуального развития животных и его роль в эволюции//Зоологический журнал. 1966. Т. XLV. N 3. С. 321-333.
22. Игнашенко А.М. Акробатика. М. 1951 [цит. по Дымерский В.Я. О применении воображаемых действий в процессе восстановления и сохранения навыков//Вопросы психологии. 1955. N 6. С. 50].
23. Крушинский Л.В. Биологические основы рассудочной деятельности. М. МГУ. 1986.
24. Павлов И.П. Лекции о работе больших полушарий головного мозга. М. Изд-во АМН СССР. 1952.
25. Полани М. Личностное знание. На пути к посткритической философии. Благовещенск. БГК им. И.А. Бодуэна де Куртенэ.1998.
26. Поппер К. Логика и рост научного знания. М. Прогресс. 1983.
27. Сахаров Д.А. Множественность нейротрансмиттеров: функциональное значение //Журн. эвол. биох. и физиол. 1990. Т. 26. N 5. С. 733-745.
28. Сварник О.Е. Формирование индивидуального опыта и его нейрогенетическое обеспечение: экспрессия гена c-fos. Дисс. канд. психол. н. М. Инстиут психологии РАН. 2003.
29. Сварник О.Е., Анохин К.В., Александров Ю.И. Распределение поведенчески специализированных нейронов и экспрессия трнаскрипционного фактора c-Fos в коре головного мозга крыс при научении //Журн.Высш.Нервн.Деят. 2001. Т. 51. N 6. С. 758-761.
30. Симонов П.В. Эмоциональный мозг. М. Наука. 1981.
31. Симонов П.В. Лекции о работе головного мозга. М. Институт психологии РАН.1998.
32. Судаков К.В. Системогенез поведенческого акта //Механизмы деятельности мозга. М. Госнаучтехиздат. 1979. С. 88-89.
33. Тихомиров О.К. отв. ред. Психологические исследования творческой деятельности. М. Наука. 1975.
34. Третьяк Т.М., Архипова Л.В. Внутриклеточная активность нейромедиаторов //Усп. Совр. Биол. 1992. Т. 112. N 2. С. 265-272.
35. Хьелл Л., Зиглер Д. Теории личности. Санкт-Петербург. Питер. 1997.
36. Швырков В.Б. Нейрональные механизмы обучения как формирование функциональной системы поведенческого акта //Механизмы системной деятельности мозга. Горький. 1978а. С. 147-149.
37. Швырков В.Б. Нейрофизиологическое изучение системных механизмов поведения. М. Наука. 1978б.
38. Швырков В.Б. Введение в объективную психологию. Нейрональные основы психики. М. Институт психологии РАН. 1995.
39. Швыркова Н.А., Швырков В.Б. Корковые нейроны в поисковом
поведении и
обучении//Поисковая активность и сон. Баку. Элм. 1986. С. 65-73.
40. Шерстнев В.В. Нейрохимическая характеристика “молчащих” нейронов коры мозга // Доклады Академии наук СССР. 1972. Т.202 N. 6. С. 1473-1476.
41. Шулейкина К.В., Хаютин С.Н. Развитие теории системогенеза на современном этапе //Журн.высш.нервн.деят. 1989. Т. 39. N1. С. 3-19.
42. Юревич А.В. Социальная психология науки. СПб. Изд-во РХГИ. 2001.
43. Abraham W.C., Logan B., Greenwod J.M., Dragunow M. Induction and experince-dependent consolidation of stable long-term potentiation lasting months in the hippocampus//The J. of Neurosci. 2002. V. 22. P. 9626-9634.
44. Alexandrov Yu. I. Comparative description of consciousness and emotions in the framework of systemic understanding of behavioral continuum and individual development//Neuronal bases and psychological aspects of consciousness. C. Teddei-Ferretti & C. Musio (Eds.). Singapour, N.Y., London, Hong-Kong. World Scientific. 1999. P. 220-235.
45. Alexandrov Yu.I. On the way towards neuroculturology: From the neuronal specializations through the structure of subjective world to the structure of culture and back again// Proceeding of the International simposium «Perils and prospects of the new brain sciences». Stockholm. 2001. P. 36-38.
46. Alexandrov Yu.I., Grinchenko Yu.V., Jarvilehto T. Change in the pattern of behavioral specialization of neurons in the motor cortex of the rabbit following lesion of the visual cortex // Acta Physiol. Scand. 1990. V. 139. P. 371-385.
47. Alexandrov Yu. I., Grinchenko Yu. V., Shevchenko D. G., Averkin R. G., Matz V. N., Laukka S., Korpusova A. V. A subset of cingulate cortical neurons is specifically activated during alcohol-acquisition behaviour // Acta Physiol. Scand. 2001. V. 171. P. 87-97.
48. Allen C. B., Celikel T., Feldman D. E. Long-term depression induced by sensory deprivation during cortical map plasticity in vivo// Nature Neurosci. 2003. V. 6. P. 291 –299.
49. Allsopp T.E., Fazakerley J.K. Altruistic cell suicide and the specialized case of the virus-infected nervous system //TINS. 2000. V. 23. P. 284-290.
50. Arshavsky Yu.I. Long-term memory: does it have a structural or chemical basis// TINS. 2003. V. 26. P. 465-466.
51. Averkin R.G., Grinchenko Yu.V., Sozinov A.A., Kuzina E.A., Alexandrov Yu.I. Anterolateral motor cortex unit activity in rabbits during realization of food-acquisition behavior in a single way or in two alternative ways// FENS abstr. 2002. V. 1. A040.1. P.105.
52. Averkin R.G., Sozinov A.A., Grinchenko Yu.V., Alexandrov Yu.I. Retrosplenial unit activity in rabbits during food-acquisition behavior performed by two sequentially learned alternative ways// IBRO World Congress of Neuroscience. 2003. Abstr. A1240.
53. Baily C.H., Kandel E.R. Structural changes accompanying memory storage //Ann.Rev.Physiol. 1993. V. 53. P. 397-426.
54. Bartlett F. Remembering. Cambridge. Cambridge Univ. Press. 1932.
55. Bechara A., Damasio H., Tranel D., Damasio A.R. Deciding advantageously before knowing the advantageous strategy. Science. 1997. V. 275. P. 1293-1295.
56. Berntson G.G., Boysen S.T., Cacioppo J.H. Neurobehavioral organisation and the cardinal principle of evaluative bivalence//Annals of the New York Acad. of Sci. 1993. V. 702. P. 75-102.
57. Berretta, S., Parthasarathy, H.B., & Graybiel, A.M. Local release of GABAergic inhibition in the motor cortex induces immediate-early gene expression in indirect pathway neurons of the striatum // J. Neurosci. 1997. V. 17. P. 4752-4763.
58. Bortolotto Z.A., Collongridge G.L. On the mechanism of long-term potentiation induced by [1S,3R]-1-aminocyclopentane-1,3-dicarboxylic acid [ACPD] in rat hippocampal slices//Neuropharmacology. 1995. V. 34. P. 1003-114.
59. Bradley, P.M., Burns, B.D., King, T.M., Webb, A.C. Electrophysiological correlates of prior training: an in vitro study of an area of the avian brain which is essential for early learning// Brain Research. 1996. V. 708. P. 100-108.
60. Buzsaki G., Kandel A. Somadendritic backpropagation of action potentials in cortical pyramidal cells of the awake rat//J. Neurophysiol. 1998 V. 79. P. 1587-1591.
61. Carleton A., Petreanu L.T., Lansford L., Lledo P.-M. Becoming a new neuron in the adult olfactory bulb//Nature Neurosci. 2003. V. 6. P. 507- 518.
62. Castro-Alamancos M.A., Borrell J., Garcia-Segura L.M. Performance in an escape task induces Fos-like immunoreactivity in a specific area of the motor cortex of the rat // Neuroscience. 1992. V. 49. P. 157-162.
63. Chang J.-Yu., Sawyer S.F., Lee R.-S., Woodward D.J. Electrophysiological and pharmacological evidence for the role of the nucleus accumbence in cocaine self-administration in freely moving rats//The J. of Neurosci. 1994. V. 14. P. 1224-1244.
64. Chen L.L., Wise S.P. Evolution of directional preferences in the supplementary eye field during acquisition of conditional oculomotor associations//J. of Neurosci. 1996. V. 16. P. 3067-3081.
65. Clayton D.F. The genomic action potential //Neurobiology of Learning and Memory. 2000. V. 74. P. 185-216.
66. Conti A.C., Raghupathi R., Trojanowski J.Q., McIntosh T.M. Experimental brain injury induces regionally distinct apoptosis during the acute and delayed post-traumatic period //The J. of Neurosci. 1998. V. 18. P. 5663-5672.
67. Corina D.P., Vaid J. , Bellugi U. The linguistic basis of left hemisphere specialization// Science. 1992. V. 255. P. 1258-1260.
68. Cotman C.W, Hailer N.P., Pfister K.K., Soltesz I., Schachner M. Cell adhesion molecules in neural plasticity and pathology: similar mechanisms, distinct organizations? //Prog Neurobiol. 1998. V. 55. P. 659-669.
69. Cramer S.C., Chopp M. Recovery recapitulates ontogeny //TINS. 2000. V. 23. P. 265-271.
70. Crist R.E., Li W., Gilbert C.D. Learning to see: experience and attention in primary visual cortex//Nature Neurosci. 2001. V. 4. P. 519 – 525.
71. Day R.L., Laland K.N., Odling-Smee J. Rethinking adaptation. The niche-construction perspective//Persp. in Biol. and Med. 2003. V. 46. P. 80-95.
72. Della-Magiorell V., Sekuler A.B., Gradyl L.G., Bennett P.J., Sekuler R., McIntosh R. Corticolimbic interactions associated with performance on a short-term memory task modified by age//The J. of Neurosci. 2000. V. 20. P. 8410-8416.
73. Dennett C.D. Darwin's dangerous idea. N.Y. Simon & Schucter. 1995.
74. Dragunow M., de Castro D., Faull R.L.M. Induction of Fos in glia-like cells after focal brain injury but not during wallerian degeneration//Brain Res. 1990. V. 527. P. 41-54.
75. Du C., Hu R., Csernansky C.A., Hsu C.Y., Choi D.W. Du C., Hu R., Csernansky C.A., Hsu C.Y., Choi D.W. Very delayed infarction after mild focal cerebral ischemia: a role for apoptosis? // J. Cereb. Blood Flow Metab. 1996. V. 16. P. 195-201.
76. Dudai Y. Caught in the act//Nature. 2003. V. 424. P. 377-378.
77. Edelman G.M. Neural Darwinism: The theory of neuronal group selection. N.Y. Basic. 1987.
78. Elbert T., Pantev C., Wienbruch C., Rockstroh B., Taub E. Increased cortical representation of the fingers of the left hand in string players//Science. 1995. V. 270. P. 305-307.
79. Ellis R.D. Dynamical systems as an approach to consciousness: emotion, self- organization and the mind-body problem//New Ideas in Psychology. 1999. V. 17. P. 237-250.
80. Engel K.A., Fries P., Singer W. Dynamic predictions: oscillations and synchrony in top-down processing//Nature Rev. Neurosci. 2001. V. 2. P. 704-716.
81. Erickson C. A., Desimone R. Responses of macaque perirhinal neurons during and after visual stimulus association learning//J. of Neurosci. 1999. V.19. P. 10404- 10416.
82. Eriksson P.S., Perfilieva E., Bjork-Eriksson T., Alborn A-M., Nordbrog C., Peterson D.A., Gage F.H. Neurogenesis in the adult human hippocampus//Nature Medicine. 1998. V. 4. P. 1313-1317.
83. Ferrari P.F., Gallese V., Rizzolatti G., Fogassi L. Mirror neurons responding to the observation of ingestive and communicative mouth actions in the monkey ventral premotor cortex//Eur. J. of Neurosci. 2003. V. 17. P. 1703-1714.
84. Freeman W.J. Three centuries of category errors in studies of the neural basis of consciousness and intentionality//Neural Networks. 1997. V. 10. P. 1175-1183.
85. Fregnac Y. A tale of two spikes//Nature Neurosci. 1999. V. 2. P. 299-301.
86. Gandolfo F., Li C.-S. R., Benda B. J., Schioppa C. P., Bizzi E. Cortical correlates of learning in monkeys adapting to a new dynamical environment// Proc. Natl. Acad. Sci. U S A. 2000. V. 97. P. 2259-2263.
87. Gauthier I., Skudlarski P., Gore J. C., Anderson A. W. Expertise for cars and birds recruits brain areas involved in face recognition//Nature Neurosci. 2000. V. 3. P. 191 –197
88. Gentner T.Q., Margoliash D. Neuronal populations and single cells representing learned auditory objects//Nature. 2003. V. 424. P. 669-674.
89. Gilbert C. Adult cortical dynamics//Physiol. Rev. 1998. V. 78. P. 467 – 485.
90. Goldstein K. The organism. N.Y. American Book Company. 1933.
91. Gorkin A.G., Alexandrov Yu.I., Reynmann K.G. Long-term potentiation in cingulate cortex of freely moving rats// Neuroscience Res. Commun. 1997. V. 21. P. 119-124.
92. Gottlieb G.L., Corcos D.M., Jaric S. Practice improves even the simplest movements//Exp. Brain. Res. 1988. V. 73. P. 436 -440.
93. Hagmann М. Memory T cells don't need practice //Science. 1999. V. 286. P. 1266 – 1267.
94. Hall W.G., Bryan T.T. The ontogeny feeding in rats. II. Independent ingestive behavior//J. Comp. Physiol. Psychol. 1980. V. 94. P. 746 – 736.
95. Horn G. Pathways of the past: The imprint of memory// Nature Rev. Neurosci. 2004. V. 5. P. 108-121.
96. Horn G., Nicol A.U., Brown M.W. Tracking memory’s trace// Proc. Natl. Acad. Sci. U S A. 2001. V. 98. P. 5282-5287.
97. Developmental expression of Bcl-2 protein in human cortex //Brain Res. Dev. Brain Res. 2000. V. 119. P. 225-230.
98. Jeannerod M. The 25th Bartlett lecture. To act or not to act: Perspectives on the representation of actions//The Quarterly Journal of Experimental Psychology.1999. V.52A. P. 1-29.
99. Jog M.S., Kubota K, Connolly C.I., Hillegaart V., Graybiel A.M. Bulding neural representations of habits// Science. 1999. V. 286. P. 1745-1749.
100. Jordan J. S. Recasting Dewey’s critique of the reflex-arc concept via a theory of anticipatory consciousness: implications for theories of perception//New Ideas in Psychol. 1998. V. 16. P. 165-187.
101. Kaczmarek L., Chaudhuri A. Sensory regulation of immediate–early gene expression in mammalian visual cortex: implications for functional mapping and neural plasticity // Brain Res. Rev. 1997. V. 23. P. 237-256.
102. Karni A., Meyer G., Jezzard P., Adams M.M., Turner R., Ungerleider L.G. Functional MRI evidences for adult motor cortex plasticity during motor skill learning//Nature. 1995. V. 377. P. 155-158
103. Kempermann G., Kuhn G.H., Gage F.H. Experience-induced neurogenesis in the senescent dentate gyrus//The J. of Neurosci. 1998. V. 18. P. 3206-3212.
104. Komatsu Y., Fujii K., Maeda J., Sakaguchi H., Toyama K. Long-term potentiation of synaptic transmission in kitten visual cortex//J. Neurophysiol. 1988. V. 59. P. 124-141.
105. Koshland D.E. The bacterium as a model neuron//TINS.1983.V.6. P.133-137
106. Koyama T., Kato K., Tanaka Z., Mikami T. Anterior cingulate activity during pain-avoidance and reward tasks in monkeys//Neurosci. Res. 2001. V. 39. P. 421-430.
107. Krug M., Lossner B., Ott T. Anisomicin blocks the late phase of long-term potentiation in the dentate gyrus of freely moving rats//Brain Res. Bull. 1984. V.13. P. 39-42.
108. Lee Y., Park K.H., Baik S.H., Cha Ch.I. Lee Y., Park K.H., Baik S.H., Cha Ch.I. Attenuation of c-Fos basal expression in the cerebral cortex of aged rat // NeuroReport. 1998. V. 9. P. 2733-2736.
109. Leist M., Jaattela M. Four deaths and a funeral: from caspases to alternative mechanisms//Nature Rev. 2001. V. 2. P. 1-10.
110. Lisman J., Lichtman J.W., Sanes J.R. LTP: perils and progress// Nature Rev. Neurosci. 2003. V. 4. P. 926-929.
111. Ljungberg T., Apicella P., Schultz W. Responses of monkey dopamine neurons during learning of behavioral reactions//J. of Neurophysiol. 1992. V. 67. P. 145-163
112. Malenka R.C. The long-term potential of LTP// Nature Rev. Neurosci. 2003. V. 4. P. 923-926.
113. Manahan-Vaughan D., Behnish T., Reymann K.G. ACPD-mediated slow-onset potentiation is associated with cell death in the rat CA1 region in vivo// Neuropharmacology. 1999. V. 38. P. 487-494.
114. Matthies H., Ruethrich H., Ott T., Matthies H.K., Matthies R. Low frequency perforant path stimultaion as a conditioned stimulus demonstrates correlations between long-term synaptic potentiation and learning//Physiol. Behav. 1986. V. 36. P. 811-821.
115. Merzenich M.M., Kaas J.H., Sur M., Lin C.S. Double representation of the body surface within cytoarchitectonic areas 3b and 1 in «SI» in the owl monkey [Aotus trivirgatus]// J. Comp. Neurol. 1978. V. 181. P. 41-73.
116. Nader K., Schafe G.E., Le Doux J.E. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval//Nature. 2000. V. 406. P. 722-726.
117. Nader K. Response to Arshavsky: Challenging the old views//TINS. 2003. V. 26. P. 466-468.
118. Najbauer J., Leon M. Olfactory experience modulates apoptosis in the developing olfactory bulb //Brain Res. 1995. V. 674. P. 245-251.
119. Nakamura A.E., Sakai K., Hikosaka O. Neuronal activity in medial frontal cortex during learning of sequential procedures//J. of Neurophysiol. 1998. V. 80. P. 2671-2687.
120. Nestler E.J., Aghajanian G.K. Molecular and cellular basis of addiction // Science. 1997. V. 278. P. 58 ‑ 63.
121. Nicola S.M., Deadwyler S.A. Firing rate of nucleus accumbens neurons is dopamine –dependent and reflects the timing of cocaine-seeking behavior in rats on a progressive ratio schedule of reinforcement// The J. of Neurosci. 2000. V. 20. P. 5526-5537.
122. Nishijo H., Yamamoto Y., Ono T., Uwano T., Yamashita J., Yamashima T. Single neuron responses in the monkey anterior cingulate cortex during visual discrimination//Neurosci. Let. 1997. V. 227. P. 79-82.
123. Onodera H., Kogure K., Ono Y., Igarashi K., Kiyota Y., Nagaoka A. Proto-oncogene c-fos is transiently induced in the rat cerebral cortex after forebrain ischemia // Neurosci lett. 1989. V. 98. P. 101-104.
124. Paton J.A., Nottebohm F.N. Neurons generated in the adult brain are recruited into functional circuits//Science. 1984. V. 225. P. 1046-1048.
125. Petitto L.A., Marentette P.F. Babbling in the manual mode: evidence for the ontogeny of language//Science. 1991. V. 251. P. 1493 – 1496.
126. Piatelli-Palmarini M. Evolution, selection and cognition: From «learning» to parameter setting in biology and in the study of language//Cognition. 1989. V. 31. P.1-44.
127. Popper K.R., Eccles J .C. The self and its brain. Berlin. Springer. 1977.
128. Procyk E., Tanaka J.L., Joseph J.-P. Anterior cingulate activity during routine and non-routine sequential behaviors in macaques//Nature Neurosci. 2000. V. 3. P. 502-508.
129. Prickaerts J., Koopmans G., Blokland A., Scheepens A. Learning and adult neurogenesis: Survival with or without proliferation?// V. 81. P. 1-11.
130. Ranck J. B. Studies on single neurones in dorsal hippocampal formation and septum in unrestrained rats: I. Behavioral correlates and firing repertoires//Experimental Neurology. 1973. V. 41. P. 461-555.
131. Ranganath C., Rainer G. Neural mechanisms for detecting and remembering novel events// Nature Rev. Neurosci. 2003. V. 4. P. 193-202.
132. Raoul C., Pettmann B. , Henderson C.E. Active killing of neurons during development and following stress: a role for p75NTR and Fas? // Current Opin. in Neurobiol. 2000. V. 10. P. 111-117.
133. Recanzone G.H., Schreiner C.E., Merzenich M.M. Plasticity in the frequency representation of primary auditory cortex following discrimination training in adult owl monkeys//J. of Neurosci. 1993. V. 13. P. 87 – 103.
134. Reed E.S. Neural regulation of adaptive behavior. An essay review of neural darwinism by Gerald M. Edelman //Ecological Psychol. 1989. V. 1. P. 97-117.
135. Robbins T.W., Everitt B.J. Drug addiction: bad habits add up // Nature. 1999. V. 398. P. 567 ‑ 570.
136. Rose C.R., Intracellular Na+ regulation in neurons and glia: functional implications//The Neuroscientist. 1997. N 3. P. 85-88.
137. Sara S.J. Retrieval and reconsolidation: toward a neurobiology of remembering//Learn. and Mem. 2000. V. 7. P. 73-84.
138. Schneirla T.C. A theoretical consideration of the basis for approach-withdrawal adjustments in behavior//Psychol. Bull. 1939. V. 37. P. 501-502.
139. Schneirla T.C., Rosenblatt J.C. Behavioral organization and genesis of the social bond in insects and mammals//Animal behavior. N.Y. Holt, Rinehart and Winston. 1965. p. 247 – 270.
140. Schoenbaum G., Chiba A. A.., Gallagher M. Neural encoding in orbitofrontal cortex and basolateral amygdala during olfactory discrimination learning//J of Neurosci. 1999. V. 19. P. 1876-1884.
141. Schreiber S.S., Baudry M. Selective neuronal vulnerability in the hippocampus –– a role for gene expression? //TINS. 1995. V. 18. P. 446- 451.
142. Shadmehr R., Holcomb H.H. Neural correlates of motor memory consolidation//Science. 1997. V. 277. P. 821-825
143. Sherrington Ch. Man on his Nature. 1951. Pelican Books. A 322.
144. Shima K, Mushiake H, Saito N, Tanji J. Role for cells in the presupplementary motor area in updating motor plans//Proc. Natl. Acad. Sci. U S A. 1996. V. 93 P. 8694-8698.
145. Shors T.J., Matzel L.D. Long-term potentiation [LTP]: what's learning got to do with it?// Behav. and Brain Sci. 1997. V. 20. P. 597-655.
146. Shors T.J., Miesegaes G., Beylin A., Zhao M., Rydel T., Gould E. Neurogenesis in the adult is involved in the formation of trace memories //Nature. 2001. V. 410. P. 372-376.
147. Shvyrkov V.B. Behavioral specialization of neurons and the system-selection hypothesis of learning // Human memory and cognitive capabilities. Amsterdam. Elsevier. 1986. P. 599-611.
148. Smale G., Nichols N.R., Brady D.R., Finch C.E., Horton W.E. Jr. Evidence for apoptotic cell death in alzheimer's disease //Exp. Neurol. 1995. V.133. P. 225-230.
149. Spinelli D.N. Neural correlates of visual experience in single units of cats visual and somatosensory cortex//Frontiers in visual science. N.Y. Springer. 1978. P. 674-688.
150. Stone E.A., Zhang Y., John S., Filer D., Bing G. Effect of locus coeruleus lesion on c-fos expression in the cerebral cortex caused by yohimbine injection or stress //Brain Res. 1993. V. 603. P. 181-185.
151. Strassmann J.E., Zhu Y., Queller D.C. Altruism and social cheating in the social amoeba Dictyostelium discoideum // Nature. 2000. V. 408. P. 965-967.
152. Swadlow H.A., Hicks T.P. Subthreshold receptive fields and baseline excitability of «silent» S1 callosal neurons in awake rabbits: contributions of AMPA/kainate and NMDA receptors// Exper. Brain Res. 1997. V. 115. P. 403-409.
153. Tanaka K. Neuronal mechanisms of object recognition//Science. 1993. V. 262. P. 685-688.
154. Tanila H., Shapiro M., Gallagher M., Eichenbaum H. Brain aging: changes in the nature of information coding by the hippocampus// The J. of Neurosci. 1997. V. 17. P. 5155-5166.
155. Thompson E., Varela F. J. Radical embodiment: neural dynamics and consciousness//Trends in Cognit. Sci. 2001. V. 5. P. 418-425.
156. Thompson L.T., Best P.J. Long-term stability of the place-field activity of single units recorded from the dorsal hippocampus of freely behaving rats//Brain Res. 1990. V. 509. P. 299-308.
157. Vandervert L.R. Consciousness: a preliminary multidisciplinary mapping of concepts//New Ideas in Psychol. 1998. V. 16. P. 159 – 164.
158. Le Vere T.E. Recovery of function after brain damage: a theory of the behavioral deficit.//Physiol. Psychol. 1980. V. 8. P. 297-308.
159. Vikman K.S., Duggan A.W., Siddall P.J. Increased ability to induce long-term potentiation of spinal dorsal horn neurons in monoarthritic rats//Brain Res. 2003. V. 990. P. 51-57.
160. Wilson M.A., McNaughton B.L. Dynamics of the hippocampal ensemble code for space// Science. 1993. V. 261. P. 1055-1058.
161. Wirth S., Yanike M., Frank L.M., Smith A.C., Brown E.N., Wendy A.S. Single neurons in the monkey hippocampus and learning new associations//Science. 2003. V. 300. P. 1578-1581.
162. Woodward D.J., Janak P.H., Chang J.-Yu. Ethanol action on neuronal networks studied with multineuron recording in freely moving animals//Alcohol: Clin. and Exper. Res. 1998. V. 22. P. 10-22.
163. Xue Z.M. The studies on neurogenesis induced by brain injury in adult ring dove//Cell Res. 1998. V.8. P.151-162.
164. Yuan J., Yankner B.A. Apoptosis in the nervous system//Nature. V. 2000. V. 407. P. 802- 809.