Психофизиология под редакцией Ю.И.Александрова
Психофизиология под редакцией Ю.И.Александрова
3-е издание 2007г.
избранные фрагменты из книги.
1.2.4. Пейсмекер
Один из удивительных видов электрической активности нейронов, регистрируемой внутриклеточным микроэлектродом, — это пейсмекерные потенциалы. А. Арванитаки и И. Халазопитис (1955) впервые описали ритмические потенциалы нервной клетки, не связанные с поступлением к ней синаитических влияний. Эти колебания в ряде случаев приобретают такой размах, что превышают критический уровень потенциала, необходимый для активации механизма электрической
возбудимости. Наличие в соме клетки таких моли мембранного потенциала было обнаружено на нейронах моллюсков. Они были расценены как проявление спонтанной, или ауторитмичсской, активности, имеющей эндогенное происхождение. Аналогичные ритмические колебания были затем описаны во многих типах нейронов. Способность к длительной ритмической активности сохраняется у некоторых клеток в течение длительного времени после полного их выделения. Следовательно, в ее основе действительно лежат эндогенные процессы, приводящие к периодическому изменению ионной проницаемости поверхностной мембраны. Важную роль играют изменения ионной проницаемости мембраны под действием некоторых цитоплазматнчеекпх факторов, например системы обмена циклических пуклеотидов. Изменения активности этой системы при действии на соматическую мембрану некоторых гормонов или других внесинаити-ческих химических влиянии могут модулировать ритмическую активность клетки (эндогенная модуляция).
Запускать генерацию колебаний мембранного потенциала могут сипаптические и внесипаитические влияния. Л. Тауц и Г. М. Гершепфельд 11960] обнаружили, что соматическая мембрана нейронов моллюсков, не имеющая на своей поверхности сипаптических окончаний, обладает высокой чувствительностью к мелиаторпым веществам и, следовательно, имеет молекулярные хемоуправляемые структуры, свойственные иостсинаптической мембране. Наличие внесинантической реценции показывает возможность модуляции псисмексриоп активности диффузным действием выделяющихся ме-диаторных веществ.
Сложившаяся концепция о двух типах мембранных структур — электровозбудимой и электроиевозбудимой, но химически возбудимой, заложила основу представлений о нейроне как о пороговом устройстве, обладающем свойством суммации возбуждающих и тормозных сипаитических потенциалов. Принципиально новое, что вносит эндогенный пейсмекерный потенциал в функционирование нейрона, заключается в следующем: пейсмекериый потенциал превращает нейрон из сумматора сипаптических потенциалов в генератор. Представление о нейроне как об управляемом генераторе заставляет по-новому взглянуть на организацию многих функций нейрона.
Пейсмекериыми потенциалами в собственном смысле этого слова называют близкие к синусоидальным колебания частотой 0,1-10 Гц и амплитудой 5-10 мВ. Именно эта категория эндогенных потенциалов, связанных с активным транспортом ионов, образует механизм внутреннего генератора нейрона, обеспечивающего периодическое достижение порога генерации ПД в отсутствие внешнего источника возбуждения. В самом общем виде нейрон состоит из электровозбудимой мембраны, химически возбудимой мембраны и локуса генерации пейсмекерной активности. Именно пейсмекерный потенциал, взаимодействующий с хемовозбудимой и электровозбудимой мембраной, делает нейрон устройством со «встроенным» управляемым генератором.
Если локальный потенциал является частным случаем механизма генерации ИД. то пейсмекерный потенциал принадлежит к особому классу потенциалов эдектрогеипому эффекту активного транспорта ионов, особенности ионных механизмов электрической возбудимости соматической мембраны лежат в основе важных свойств нервной клетки, в первую очередь ее способности генерировать ритмические разряды нервных импульсов. Электрогепиый эффект активного транспорта возникает в результате несбалансированного переноса ионов в разных направлениях. Например, в результате активного вывода ионов натрия возникает гииерполяризациоииый постоянный потенциал. Дополнительное включение активного насоса ионов натрия создает фазические медленные волны гиперполяризации (негативные отклонения от уровня мембранного потенциала покоя), обычно возникающие вслед за высокочастотной группой ПД, которая приводит к избыточному накоплению натрия в нейроне.
Несомненно, что некоторые из компонентов механизма электрической возбудимости соматической мембраны, а именно электроуиравляемые кальциевые каналы, вместе с тем являются фактором, сопрягающим мембранную активность с цитоплазматическими процессами, в частности с процессами иротонлазматиче-ского транспорта и нервной трофики. Детальное выяснение этого важного вопроса требует дальнейшего экспериментального изучения.
Пейсмекерпый механизм, будучи эндогенным но происхождению, может активироваться и ииактивироваться па длительное время в результате афферентных воздействий на нейрон. Пластические реакции нейрона могут обеспечиваться изменениями эффективности сииаптической передачи и возбудимости иейсмекер-ного механизма.
Пейсмекерпый потенциал является компактным способом передачи внутри-нейронной генетической информации. Приводя к генерации ПД, он обеспечивает возможность выхода эндогенных сигналов на другие нейроны, в том числе и эф-фекториые, обеспечивающие реакцию. Тот факт, что генетическая программа включает звено управления иейсмекериым потенциалом, позволяет нейрону реа-лизовывать последовательность своих генетических программ. Наконец, пейсмекерпый потенциал в той или иной степени может подвергаться сииаптическим влияниям. Этот путь позволяет интегрировать генетические программы с текущей активностью, обеспечивая гибкое управление последовательными программами. Пластические изменения иейсмекериого потенциала еще больше расширяют возможность приспособления наследственно фиксированных форм к потребностям организма. Пластические изменения развиваются в этом случае не в геноме а на пути выхода наследственной программы па реализацию (на уровне генерации ПД).
...
3.4. Детектирование сигналов
Детектированием называют избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего поведенческое значение. Осуществляют такой анализ нейроны-детекторы, избирательно реагирующие лишь на определенные свойства стимула. Так, типичный нейрон зрительной коры отвечает разрядом лишь на один из наклонов (ориентацию) светлой или темной полоски, расположенной в определенной части ноля зрения [Хьюбел, 1990]. При других наклонах той же полоски ответят другие нейроны. Такие нейроны называют детекторами первого порядка, так как они выделяют наиболее простые признаки сигнала. В высших отделах сенсорной системы сконцентрированы детекторы высших порядков, ответственные за выделение сложных признаков и целых образов. Примером могут служить детекторы лиц, найденные в нижневисочной коре обезьян (предсказанные много лет назад Ю. Коиорским, они были названы «сверхдетекторами»). Многие детекторы формируются в раннем детстве под влиянием опыта, а у части из них детекторные свойства заданы генетически (см. также 23.1.1).
....
5.4. Двигательные программы
Управление движениями немыслимо без согласования активности большого количества мышц. Характер этого согласования зависит от двигательной задачи. Так, если нужно взять стакан воды, то ЦНС должна располагать информацией о положении стакана относительно тела и об исходном положении руки. Однако чтобы движение было успешным, необходимо, чтобы кисть заранее раскрылась на
величину, соответствующую размеру стакана, чтобы сгибатели пальцев сжимали стакан с силой, достаточной для предотвращения проскальзывания, чтобы приложенная сила была достаточной для плавного подъема, но не вызывала резкого отрыва, чтобы ориентация стакана в кисти после захвата все время была вертикальной. Таким образом, чтобы реализация движения соответствовала двигательной задаче, необходимы не только данные о пространственных соотношениях, но и сведения о свойствах объекта манипулирования. Многие из этих сведений не могут быть получены в ходе самого движения посредством обратных связей, а должны быть предусмотрены на этапе планирования. Следовательно, для осуществления движения должна быть сформирована двигательная программа. Двигательную, или центральную, программу рассматривают как заготовленный набор базовых двигательных команд, а также набор готовых корректирующих подпрограмм, обеспечивающих реализацию движения с учетом текущих афферентных сигналов и информации, поступающей от других частей ЦНС.
Зарождение побуждения к движению связано с активностью подкорковых и корковых мотивациопиых зон. Замысел движения формируется в ассоциативных зонах коры. Далее происходит формирование программы движения с участием базальиых ганглиев и мозжечка, действующих па двигательную кору через ядра таламуса. За реализацию программы отвечает двигательная кора и нижележащие стволовые и спецальные двигательные центры.
Предполагается, что двигательная память содержит обобщенные классы двигательных программ, из числа которых в соответствии с двигательной задачей выбирается нужная. Программа модифицируется применительно к ситуации: однотипные движения могут выполняться быстрее или медленнее, с большей или меньшей амплитудой. Интересно, что одна и та же программа может быть реализована разными наборами мышц. Так, почерк человека сохраняет характерные черты при письме правой и левой рукой и даже карандашом, зажатым в зубах ил! прикрепленным к носку ботинка. Такой перенос навыка возможен потому, чт< система управления движениями является многоуровневой (уровень илаиирова пня движения и уровень его исполнения в ней не совпадают). Действительно произвольное движение планируется в терминах трехмерного евклидового про странства. Для исполнения этого плана необходимо перевести линейные пере мещсния в соответствующие угловые переменные (изменения суставных углов) определить, какие мышечные моменты необходимы для этих угловых перемещений, и, наконец, сформировать двигательные команды, которые вызовут активацию мышц, дающую необходимые значения моментов.
Двигательная программа может быть реализована различными способами. В простейшем случае ЦНС посылает к мышцам заранее сформированную последовательность команд, не подвергающуюся во время реализации никакой коррекции. В этом случае говорят о разомкнутой системе управления. Подобное управление используется при осуществлении быстрых, так называемых «баллистических» движений. Чаще всего ход осуществления движения сравнивается с его планом на основе сигналов от многочисленных рецепторов, и в реализуемую программу вносятся нужные коррекции. Это замкнутая система управления с обратными связями. Однако и такое управление имеет недостатки. В связи с относительно малыми скоростями проведения сигналов, не превышающими 100-120 м/с, задержками в центральном звене обратной связи, составляющими порядка 1 мс па каждое си-нантическое переключение, и временем порядка 50-80 мс, необходимым для развития усилия мышцей, коррекция движения но сигналу обратной связи может запаздывать. Поэтому во многих случаях целесообразно реагировать не на отклонение от плана движения, а на само внешнее возмущение еще до того, как оно успело вызвать отклонение. Такое управление называют управлением по возмущению.
Другим способом уменьшения влияния задержек является антиципация, т. е. упреждение, или предвосхищение. Во многих случаях ЦНС способна предусмотреть в двигательной программе появление возмущений еще до их возникновения, особенно если эти возмущения связаны с собственными действиями человека. Примечательно, что эта упреждающая «познал» активность (антиципация) осуществляется автоматически с очень короткими центральными задержками. Роль упреждающей активности в стабилизации положения звеньев тела иллюстрирует простой пример. Если официант удерживает на ладони вытянутой руки поднос с бутылкой шампанского и рюмками, а другой человек внезапно снимет бутылку с подноса, то рука резко подпрыгнет вверх с соответствующими последствиями. Если же он сам снимет бутылку свободной рукой, то рука с подносом останется па прежнем уровне.
...
6.1. Концепция временной организации памяти
В опытах ио запоминанию слогов психологи Дж. Мюллер и А. Иильцекер еще в 1911 г. обнаружили, что заучивание второго ряда ухудшает запоминание первого. Такое тормозящее влияние одного обучения на другое объясняли нарушением реверберации процессов, развившихся после первого заучивания. Дж. Мюллер и А.Пильцекер применили эту идею для объяснения ретроактивного торможения, а позже она стала основой гипотезы консолидации следа памяти (Д. О, Хебб). Временная концепция памяти возникла па основе этой гипотезы.
Временная организация следа памяти подразумевает последовательность развития во времени качественно разных процессов, приводящих к фиксации приобретенного опыта. Основные понятия, которыми оперируют теория консолидации и другие теории, созданные в рамках концепции временной организации памяти, следующие: консолидация — процесс, приводящий к физическому закреплению энграммм, и реверберация — механизм консолидации, основанный на многократном пробега!ши нервных импульсов но замкнутым цепям нейронов. Длительность консолидации — интервал времени, необходимый для перехода следа иамяти из кратковременного храпения, в котором он находится н виде реверберирующей импульсной активности, в долговременное, обеспечивающее длительное существование эпграммы.
Основными в концепции временной организации являются понятия о кратковременной и долговременной памяти. Подразумевалось, что при фиксации происходит смена одной формы существования эпграммы на другую. Представления о кратковременной и долговременной форме существования следа базируется на предположении о разных нейрофизиологических, молекулярных,
биохимических и морфофупкцнональиых основах диграммы па разных стадиях се жизни. Предполагалось, что след памяти в своем становлении проходит два этапа — этап кратковременной памяти и затем этап долговременной памяти. В кратковременной памяти след находится в течение ограниченного отрезка времени (предполагалось, что не более нескольких минут), а в долговременной — в течение времени, соизмеримого с продолжительностью жизни данного живого существа. Физиологическим механизмом следа в кратковременной памяти является реверберация электрической активности по замкнутым цепям нейронов. Согласно Хеббу, в обучение вовлекаются определенные нейроны. При многократном действии одного и того же стимула происходит формирование замкнутого «клеточного ансамбля». По цепи нейронов многократно проходит — реверберируетп — электрическая активность. Продолжаясь определенное время, реверберация приводит к морфофункцпональным и биохимическим изменениям в синапсах. Это и есть консолидация, которая составляет основу долговременной памяти. Многократное использование одних и тех же синоптических контактов улучшает проведение импульсов и приводит к формированию специфических белков.
Существование следа памяти в виде реверберирующих электрических импульсов — необходимое условие для перехода эпграммы в долговременную память. Прерывание или предотвращение реверберации должно приводить к физическому разрушению следа памяти и его исчезновению.
Чтобы проверить правильность этой гипотезы, проводили опыты с использованием методов экспериментальной ретроградной амнезии. В первой фазе (кратковременной памяти) эпграмма была неустойчива, ее можно было разрушить действием амиестического агента (например, электрошока). Эта ее неустойчивость и уязвимость объясняется тем, что реверберирующую активность легко прервать. Но тогда не произойдет и перехода следа в долговременную память. Предполагали, что переход следа из кратковременного в долговременное хранение осуществляется за несколько секунд или минут. Полученные результаты в целом подтвердили генеральную идею: действительно, ампестический агент (например, электрошок), примененный через короткий интервал времени после обучения, вызывал нарушение памяти в форме ретроградной амнезии. Однако появились факты, которые не могли быть объяснены в рамках временной концепции. Самыми важными являются данные о восстановлении памяти, потому что они резко противоречат тезису о физическом уничтожении следа, если он до амнестического воздействия не прошел стадию консолидации.
...
7.2. Развитие базисных эмоций в раннем онтогенезе
У младенцев, особенно в возрасте до года, невозможно выделить все три известных у взрослых компонента эмоций. Отсутствие речи в младенчестве затрудняет выяснение того, осознает ли ребенок изменения, происходящие в организме (например, изменение сердцебиения). Поэтому детские психологи сосредоточивают усилия на изучении изменений, происходящих в мозге и организме младенца в ситуации, которая бы очевидным образом вызвала известное эмоциональное состояние у взрослого. К примеру, речь может идти о ситуации, связанной с болью (в ходе прививки) или с радостью (при социальном взаимодействии). Исследователи также фиксируют особенности выражения лица младенца в конкретных «эмоциональных условиях». Изменения выражения лица в ответ на эмоционально значимые события универсальны и сопровождаются закономерными изменениями в деятельности мозга и автономной нервной системы. Поэтому некоторые ученые, делая выводы относительно наличия у младенца определенного эмоционального состояния, основываются на физиологических показателях активности центральной и периферической нервной системы.
Первая эмоция, которую можно явно идентифицировать но поведению и мимике ребенка, — это эмоция удовольствия. Улыбка - самое приятное выражение лица младенца. Однако необходимо учитывать, что улыбка новорожденного младенца — это реакция рефлекторного характера и может вызываться простым поглаживанием губ или щек.
К двухмесячному возрасту улыбка возникает в ответ на разнообразные раздражители, по особенно часто — на вид человеческого лица и звуки голоса. В первые два месяца жизни младенец гораздо лучше реагирует на голос матери, чем на высокий мужской голос, а в последующие шесть недель более восприимчив к лицу, чем к голосу. Младенец может улыбаться, увидев, что объект имеет сходство с ранее усвоенной «зрительной схемой». Так, в возрасте 3 месяцев дети обычно улыбаются при виде человеческого лица, схожего с лицами родителей. Трехмесячные младенцы обычно улыбаются, когда взрослый смотрит на них и разговаривает с ними. Примерно в возрасте 4-5 месяцев младенцы начинают смеяться, и чаше всего — при социальном взаимодействии, неожиданном появлении в поле зрения объекта или же во время «игры в щекотки». На протяжении первого года жизни дети часто смеются, когда кто-то вводит их в игровую ситуацию — например, когда мать играет с ними в прятки или щекочет. Но в годовалом возрасте младенцы уже улыбаются и смеются, если сами становятся активными участниками игры.
В исследованиях на взрослых было выделено два вида улыбки: искренняя и неискренняя. При улыбке первого типа включаются мышцы вокруг глаз, а также вокруг рта. Только улыбка первого тина сопровождается самоотчетом о состоянии радости. Результаты исследований десятимесячных детей подтверждают правомерность этого разграничения. При искренней улыбке у них чаще наблюдался паттерн мозговой активности, который связывают с положительными эмоциями: В левой лобной доле регистрируется большая активность, чем в правой. И если монет появления первой улыбки и смеха связан с развитием когнитивных Процессов у ребенка, то частота улыбок зависит от разнообразных случайных обстоятельств и подкрепляющих событий.
В Израиле изучались особенности улыбок детей, воспитывающихся в разных условиях Дети, проживавшие в государственных учреждениях (интернаты, детские дома), редко видели своих родителей п получали типичный для подобных условий уход. Младенцы, воспитывавшиеся в кибуцах (израильских сельских об-шннах), росли в больших домах с воспитателями, однако в первый год жизни о чих заботились матери. В третью группу входили дети, которые воспитывались матерями и жили с ними в собственных квартирах).
Анализ частоты улыбок младенцев из этих трех групп при виде незнакомого женского лица показал, что она была максимальна в возрасте около 4 месяцев. Однако в течение следующего года дети из государственных учреждений улыбались все реже и реже, тогда как у детей из семей сохранилась высокая частота улыбок. Данное различие объясняется, вероятно, тем фактом, что дети, воспитывавшиеся в семье, получали социальную (положительную) обратную связь чаще, чем дети, выросшие в казенных учреждениях. Сходство эмоционального развития в первые месяцы жизни у младенцев, воспиты-вающихся в разных условиях, означает, что первоначально наиболее существенную роль играют факторы созревания нервной системы, и только на более поздних этапах начинает проявляться вклад научения. В первые недели жизни у младенцев наблюдаются разнообразные выражения лица, многие из которых внешне схожи с мимикой взрослых. К. Изард разработал схему кодирования для измерения кратковременных изменений в выражении лица младенца. Используя эту схему, он обнаружил, что: 1) выражения, соответствующие эмоциям «удивления» и «грусти», появляются к 4-му месяцу жизни; 2) выражения четраха» и «гнева» возникают не раньше 5-7-го месяца, а 3) выражения стыда и робости — не ранее 6 -8-го месяца. Лицо 8-месячного ребенка уже явно выражает гнев, когда взрослый отбирает у него сладости, которые он хотел съесть. Выражения презрения и вицы можно наблюдать у детей не младше 2 лет.
Эмоции, которые вызывает определенная ситуация, могут быть разными в разном возрасте, так как младенец начинает по-разному интерпретировать происходящее. Детей ц возрасте от 2 до 19 месяцев снимали на видеопленку в момент, когда ИМ делали болезненную (но необходимую) прививку, и дети обычно начинали плакать. При этом лица младенцев в возрасте от 2 до 8 месяцев выражали сильное эмоциональное напряжение (стресс), что проявлялось в округлении i-уб и закрытии [113 во время плача. А на лицах 8-месячных детей был «написан* уже гнев, что выражалось положением губ в виде квадрата и открытыми глазами но время плача.
Еще один вид страха, возникающий во второй половине первого года жизни, — страх упасть с высоты. Интенсивность этого страха измеряется по стремлению младенца избегать высоты в экспериментальной ситуации «визуального обрыва», разработанной Элеонор Гибсоп с целью изучения восприятия глубины у маленьких детей. Суть методики в следующем. Младенца помещают на узкую дорожку на широкой стеклянной поверхности, прямо под которую подложена ткань в клетку. За пределами дорожки ткань опускают на один-два фута (около полуметра) ниже уровня стекла, создавая с этой стороны видимость глубины (отсюда и название «визуальный обрыв»). Большинство детей в возрасте до 7-8 месяцев, т. е. до момента появления в репертуаре их эмоциональных реакций эмоции тревоги, не боятся «глубокой стороны» стеклянной поверхности. Они перебираются через нее, если с противоположной стороны их зовет мать. Однако после 8 месяцев младенцы, как правило, начинают избегать «обрыва» и плачут, если их посадят на «глубокой стороне». Страх перед мнимой глубиной «визуального обрыва» обусловлен освоением способности к восприятию глубины.
...
8.3.4. Модулирующие нейроны
В нервной системе выделена особая группа клеток — модулирующих нейронов, которые сами не вызывают реакции, по регулируют активность других нейронов. Они образуют контакты с другими нейронами тина «синапс на синапсе». Модулирующие нейроны иричастиы к регуляции болевой чувствительности. Схема взаимодействия модулирующего нейрона с сенсорным, проводящим сигналы о болевом раздражении, можно представить следующим образом. Передача болевых сигналов в головной мозг на уровне синапса осуществляется с помощью вещества, которое находится под контролем иейропептидов, подавляющих болевые сигналы. В задних рогах спинного мозга имеются вставочные модулирующие нейроны, выделяющие пейроиептид — энкефалин, которые образуют синапсы на аксонных окончаниях болевых нейронов. Энкефалин тормозит выход вещества Р, что уменьшает возбуждение постсинантического нейрона, посылающего в головной мозг сигналы о боли.
Функции модулирующих нейронов наиболее хорошо изучены па моллюсках. Показана причастность модулирующих серотоиипергических нейронов виноградной улитки к регуляции состояния организма. Улитку, которая ползала но шару, плавающему в воде, в течение 8 дие»1 подвергали сильному электрическому раздражению без сочетания с тестирующим стимулом — слабым тактильным раздражением. Это увеличивало величину реакции улитки па тактильный стимул — втягивание глазных щупалец. Эффект сохранялся в течение многих дней. Однако он проявлялся только в той обстановке, в которой применялось болевое раздражение (на шаре в воде) и отсутствовал, когда улитка тестировалась в других условиях — па стеклянной крышке аквариума. Применение нейротоксина, избирательно нарушающего работу серотоиипергических нейронов, устраняло его проявление. Это одна из форм долговременной сенситизации, проявляющая себя
как обстановочный условный рефлекс и требующая участия модулирующих серотопипсргических нейронов. Предполагается, что данный обстановочный условный рефлекс реализуется на входах модулирующих нейро-иов, которые являются нейронами «состояния».
Модулирующие нейроны участвуют в процессе научения (см. главу 15), изменяя проводимость синапса на нресинаитическом уровне. Эффект пресинаптической пластичности, создаваемый модулирующими нейронами, может быть начальным звеном в цепи событий, приводящих к пластическом модификации поведения, связанной с выработкой условных рефлексов. Безусловное подкрепление как очень сильный раздражитель нейрона также возбуждает модулирующие нейроны и тем самым инициирует процесс иресинаптических изменений. За начальным этапом пластических изменений следует этан клеточных и молекулярных преобразований нейронов, характеризующий ассоциативное обучение.
Однако прссииаптическое облегчение, создаваемое модулирующими нейронами, может возникать и вне ситуации ассоциативного обучения только за счет активации модулирующих нейронов сильными сенсорными раздражителями. Группа американских ученых Колумбийского университета, руководимая Эриком Кэиделом, рассматривает его как результат «нремодулирующего совпадения», которое на иресипаптическом уровне повышает проводимость, а само это повышение зависит от активности. Другое название этого явления — сенситизация, или неассоциативиое обучение. Оно отличается от ассоциативного обучения, основанного на «преностсииаптическом совпадении» (рис. 8.1). В 2000 г. Эрик Кэидел стал лауреатом Нобелевской премии за открытия, касающиеся передачи сигнала в нервной системе.
Эффект иресинаптического облегчения с участием модулирующих нейронов хорошо изучен при формировании оборонительного рефлекса у виноградной улитки. Внутриклеточная стимуляция ее модулирующих нейронов, которые представлены пулом серотонипергических клеток педального ганглия, вызывает в командном нейроне оборонительной реакции появление деиоляризациоппой волны и увеличение амплитуды суммарного ВПСП. Эти изменения превращают сенсорное подпороговое раздражение командного нейрона в пороговое, что выражается в появлении у него спайковой активности.
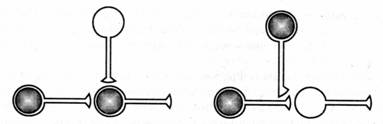 Рис. 8.1. Два
механизма изменения силы связи между нейронами. Один требует совпадения активности
пресинаптического и постсинаптическо-го нейронов во времени (два нейрона, расположенные
горизонтально). Другой механизм основан на совпадении активности пресинаптической
клетки и модулирующей (левый горизонтально и вертикально расположенные нейроны).
Нейроны, участвующие в научении, заштрихованы (по Э. Кэнделу и Р. Хокинсу,
1992]
Рис. 8.1. Два
механизма изменения силы связи между нейронами. Один требует совпадения активности
пресинаптического и постсинаптическо-го нейронов во времени (два нейрона, расположенные
горизонтально). Другой механизм основан на совпадении активности пресинаптической
клетки и модулирующей (левый горизонтально и вертикально расположенные нейроны).
Нейроны, участвующие в научении, заштрихованы (по Э. Кэнделу и Р. Хокинсу,
1992]
Описано два механизма длительного пресинаитического облегчения (фасили-тации) под влиянием серотонипа за счет фосфорилирования белков двух типов ионных каналов: кальциевых и калиевых (рис. 8.2).
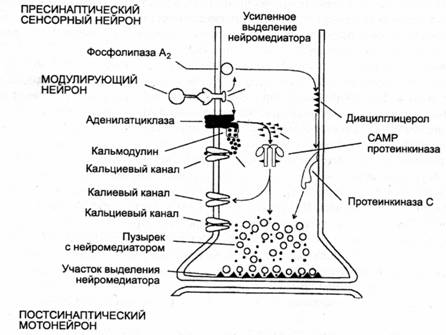 Рис. 8.2. Схема молекулярных
реакиий в пресинаптическом нейроне под влиянием возбуждения модулирующего серотонинергического
нейрона — научение, зависимое от активности |по Э. Кэнделу и Р. Хокинсу, 19921
Рис. 8.2. Схема молекулярных
реакиий в пресинаптическом нейроне под влиянием возбуждения модулирующего серотонинергического
нейрона — научение, зависимое от активности |по Э. Кэнделу и Р. Хокинсу, 19921
Участие кальциевых каналов в механизме повышения проводимости через синапс выглядит следующим образом. Во время потенциала действия ионы кальция и натрия входят в клетку, а ионы калия выходят из нее. Когда нейрон находится в активном состоянии, в нем увеличивается уровень содержания ионов кальция. Кальций, вошедший в клетку, действует на нее после того, как связывается с белком — кальмодулииом. Этот комплекс так же, как и серотоиин, активирует адени-латциклазу. За счет двойного воздействия на нее резко увеличивается образование САМР из аденозинтрифосфата (АТФ), который служит в клетке универсальным источником энергии для биохимических процессов; САМР действует как вторичный посредник и активирует протеиикииазу. Протеипкиназа фосфорили-рует рецепторные белки кальциевых каналов (присоединяет к мим фосфор). Это переводит кальциевые каналы в активное состояние — в клетку входит больше кальция, что увеличивает выделение из нее медиатора. Одновременно протеипкиназа фосфорилирует белки калиевых каналов, что ведет к уменьшению проводимости для ионов калия, т. е. к уменьшению калиевого тока, что ограничивает калиевую гипериоляризацию, следующую за ВПСП и потенциалом действия. Таким образом, потенциал действия продлевается, а это, в свою очередь, увеличивает продолжительность активного состояния кальциевых каналов.
Особую функцию в модуляции возбудимости нервной системы выполняют пейсмекерные нейроны (см. главу 1). Будучи активированными, они своими длительными, ритмическими разрядами могут повышать проводимость сигнала через синапс. Открыт специальный тин пейсмекерпых нейронов в таламусе, генерирующих разряды на частоте гамма-колебаний, которые отражают колебания кальциевого тока на деидритах клеток таламуса и которые, по-видимому, обеспечивают синхронизацию активности у групп нейронов, объединенных общими резонансными свойствами.
...
Изучение нейронных коррелятов ориентировочного рефлекса выявило существование особого класса нейронов — нейронов новизны, которые были обнаружены в разных структурах мозга. Впервые нейроны новизны были найдены в зрительной покрышке лягушки. Эти клетки переставали отвечать па повторяющееся движение зрительного объекта, совершаемое в одном и том же направлении. Изменение траектории движения восстанавливало ответ нейрона в виде спайковых разрядов. Нейроны новизны были эфферентными, а их длинные аксоны оканчивались в сетчатке. Эти нейроны сходны с «нейронами внимания», выделенными в слуховой коре кошки. Нейроны со сходными характеристиками были также найдены в ретикулярной формации ствола мозга кошки. Изучение неспецифического таламуса кролика показало существование особой формы нейронной реакции
активации, отличной от простого учащения спайковых разрядов. Новые стимулы вызывали десипхроиизацию снайковой активности нейрона, которая состояла из разрушения его пачечной активности, коррелирующей с волнами ЭЭГ, и замену ее на раидоминизнроваипые одиночные спайки, появление которых совпадало с блокадой альфа-подобной активности (см. главу 8). Реакция де-синхропизации нейронов таламуса демонстрирует все свойства ориентировочной реакции.
Совместное действие «нейронов новизны» и «нейронов тождества» подчеркивает новизну раздражителей и ослабляет действие привычных стимулов за счет их противоположного влияния на активирующую и инак-тивируюшую части модулирующей системы мозга.
Сначала формирование нервной модели стимула Е. Н. Соколов связывал с функцией корковых нейронов. Однако после открытия О. С. Виноградовой в ноле САЗ пшпокампадвух групп нейронов, реакции которых обнаружили характеристики ОР (В-псйропы, реагирующие возбуждением, и Т -нейроны, отвечающие на новый стимул торможением;, он связал нервную модель стимула с нейронами гиппокампа.
Е. Н. Соколов рассматривает эти две группы нейронов гиппокампа как «нейроны новизны» (В-иейроны) и «нейроны тождества» (Т-нейроны). Их совместное действие позволяет подчеркивать новые раздражители и ослаблять действие привычных стимулов за счет противоположного влияния па активирующую и ииак-тивирующую части модулирующей системы мозга. Формирование нервной модели определяется тем, что множество нейронов-детекторов (см. также 23.1.1) конвергируют на нейроны поля САЗ гиппокампа (нейроны новизны и тождества). Их аксоны образуют множество синапсов на этих нейронах гиппокампа. Они создают па каждом из них универсальное рецептивное поле, охватывающее всю рецепторную поверхность животного. При действии стимула происходит селективное включение детекторов. С повторением раздражителя возбуждение детекторов сохраняется прежним. Однако меняются их сипаитические контакты на нейронах гиппокампа. Веса задействованных пластических синапсов па гиппо-кампальпых нейронах уменьшаются пропорционально силе приходящего возбуждеиия. В результате возбудительный ответ нейрона новизны на повторяющийся с I пмул подавляется, и от пего регистрируется лишь фоновая активность. С повторением стимула параллельно меняется и реакция нейронов тождества, с которыми детекторы связаны не возбуждающими, а тормозными пластическими синапсами. Их тормозная реакция па новый стимул постепенно уменьшается, и нейрон сохраняет свою фоновую активность. Е. Н. Соколов предположил, что реципрок-ВЫесигналы от нейронов новизны и тождества противоположно воздействуют на активирующую и синхронизирующую системы мозга. Новый раздражитель вызывает реакцию активации, так как возбуждает активирующую и тормозит синхронизирующую (ииактивирующую) системы мозга. Привычный стимул перестает возбуждать активирующую неспецифическую систему и при этом через нейроны тождества стимулирует неспецифические тормозные влияния.
Таким образом, нервная модель стимула представлена на нейронах новизны и тождества матрицами потен-циированных синапсов, связанных со свойствами раздражителя и отражающими его конфигурацию. Новый стимул активирует новые синапсы нейронов новизны и тождества, еще не подвергнувшихся пластическим перестройкам. Это определяет появление ориентировочного рефлекса за счет возбуждения активирующей и подавления ипактивирующей систем мозга. Величина ОР увеличивается пропорционально величине различия (рассогласования) между новым стимулом и сформированной нервной моделью.
Дальнейший прогресс в понимании механизма ОР связан с применением компьютерной техники усреднения при изучении потенциалов мозга, связанных с событиями (ССП). Значительный вклад в идентификацию и классификацию различных компонентов ССП и в поиски мозговых аналогов сигналов рассогласования внесли работы Р. Наатапена и его коллег. Им выявлена специфическая мозговая волна, названная негативностью рассогласования (НР). Она получена в опытах, в которых в случайном порядке и с короткими межстимульными интервалами (не более 10-14 с) следовали стандартный (часто повторяющийся, 80-90%) идевиантные (несколько отклоняющиеся от стандарта и редко предъявляемые, 20-10%) стимулы. При этом внимание испытуемого было направлено на чтение интересной книги. Р. Наатанен для извлечения НР предложил процедуру вычитания ССП на стандартный стимул из ССП на девиантиый стимул. Им представлено большое количество экспериментальных доказательств связи НР с операцией сличения действующего стимула с короткоживущим (не более 20 с) следом стандартного стимула в памяти. Чем сильнее они отличаются, тем больше амплитуда и короче латенция НР.
Исследователями выявлено два компонента НР: модально-специфический и фронтальный. Определяя локусы генерации модалыю-специфической НР для звуковых стимулов с помощью МЭГ (см. главу 2), они нашли их в специфической слуховой коре, где, как предполагает Р. Наатанен, хранятся короткоживушие сенсорные следы. Для разных параметров звуковых стимулов (частоты, интенсивности и пространственного расположения) обнаружены разные НР с несовпадающими локусами их генерации. Предполагают, что сенсорно-специфическая НР трансформируется во фронтальную НР.
Процесс сличения стимула с короткоживущим следом памяти, который представлен в НР, протекает автоматически и не требует сознания. В частности, это подтверждается тем, что НР может возникать и па поднороговые девиации. При этом чем больше амплитуда иоднороговой НР, тем ниже субъективный порог различения испытуемого. НР рассматривается как начальная фаза обработки информации о сигнале, за которой может следовать фаза непроизвольного внимания (безусловный ориентировочный рефлекс) к девиаптному стимулу. Р. Наатанен связывает его с волной Н1.
НР как выражение сигнала рассогласования отличается от классического ОР, который, как уже упоминалось, также возникает в результате рассогласования нервной модели стимула и наличного возбуждения от раздражителя, иднако в последнем случае время жизни периной модели значительно больше, что позволяет при угашении ОР и формировании нервной модели использовать длительные межстимульные интервалы порядка нескольких десятков секунд.
Сравнивая НР с классическим ОР, Р. Наатаиен отмечает, что процесс генерации сенсорно-специфической НР имеет очевидное сходство с процессом рассогласования, постулируемым Е. Н. Соколовым для возникновения ОР, и в тоже время между ними имеется существенное различие. НР — первое событие, возникающее в активности мозга, которое направлено па инициацию ОР в отиет на физическое изменение стимула, когда стимулы предъявляются с достаточно короткими межстимульными интервалами, чтобы вызвать НР. Появление ОР в начале стимулыюй последовательности и в ответ на изменение стимула при длительных межстимульпых интервалах имеет другой механизм, отличный от вызова ОР на основе НР.
Применение вместо метода усреднения ССП процедуры удаления шумового компонента из ЭЭГ позволило исследовать динамику одиночных ССП на стимулы, предъявляемые с большими интервалами, и выявить другой сигнал рассогласования, который более тесно связан с классическим ОР. Исследователи нашли, что при угашении ОР (его компонента в виде увеличения кожной проводимости пальцев) параллельно идет процесс уменьшения в одиночном ССП несиецифиче-ской части его негативной волны (Н1). Новый стимул восстанавливает неспецифический компонент Н1, и реакцию повышения кожной проводимости. Выделенная часть компонента Н1, демонстрирует неспецифические свойства ОР и зависимость от долгоживущего следа памяти (десятки секунд).
Таким образом, сигналы рассогласования, участвовавшие в генерации непроизвольного ОР, возникают по крайней мере па двух разных типах следовых процессов: короткоживущей и долгоживущей моделях стимула.
Следует отметить, что теория ориентировочного рефлекса Е. Н. Соколова, несмотря на ее большую популярность, все же подвергалась критике. Так, отмечалось, что с ее позиций трудно объяснить появление ОР в случаях когнитивного рассогласования, т. е. когда не подтверждается ожидание относительно последовательности стимулов. Например, ориентировочная реакция возникает па цифру 9 в последовательности 1, 2, 3,4,5,6, 7,9, так как перед ней пропущена цифра 8. Другое направление критики связано с утверждением о том, что теория ОР Е. Н. Соколова не объясняет появления ОР на значимые стимулы. Сторонники концепции значимости считают, что решающее значение для появления ОР имеют корковые процессы интерпретации и усиления. С этих позиций теория ОР, возникающего па любое физическое изменение стимула, рассматривается как мало значимая для объяснения реального поведения и деятельности.
 Рис.
10.1. Усиление КГР — компонента ориентировочного
рефлекса на звук, на который испытуемый должен был реагировать движением руки
(10-24)
Рис.
10.1. Усиление КГР — компонента ориентировочного
рефлекса на звук, на который испытуемый должен был реагировать движением руки
(10-24)
1-8 — индифферентные звуковые стимулы до инструкиии; 9 — инструкция. Видно, что усиление фазических КГР-реакиий сопровождается тоническим снижением сопротивления кожи, которое сохраняется до 24-го применения стимула. По оси ординат — сопротивление в кОм |по Е. Н. Соколову, 19581
Однако участники полемики не учитывают, что наряду с безусловным ОР Е. Н. Соколов вводит понятие условного ориентировочного рефлекса, возникающего на значимый сигнальный стимул. Он показал, что придание раздражителю сигнального значения (обычное помощью инструкции или речевого подкрепления, побуждающих человека отвечать определенной реакцией на определенный стимул) восстанавливает и усиливает ОР па условный сигнал. Эффект усиления ОР сохраняется до тех нор, пока условный рефлекс не достигнет стадии автоматизации. На рис. 10.1 представлено усиление КГР, измеряемого как падение сопротивления кожи, но Фере, при придании звуковому стимулу сигнального значения. По инструкции испытуемый должен был сжимать руку в кулак при предъявлении звука после предварительного угашепия па него безусловного ОР. Выработку условного ориентировочного рефлекса можно увидеть при формировании ассоциации: звук 70 дБ — звук 90 дБ (рис. 10.2). После угашения КГР на индифферентный звук 70 дБ присоединение к нему второго стимула — 90 дБ восстанавливает реакцию, которая усиливается но мере сочетаний стимулов. Постепенно КГР-реакция па комплекс уменьшается и исчезает. Однако изолированное применение стимулов вызывает усиленный ОР, выражающий рассогласование стимула со сформированной в памяти моделью комплекса.
Сравнивая форму ССП па редко предъявляемые девиаитпые стимулы, когда они были релевантными п испытуемый должен был па них как-то реагировать и когда те же стимулы им игнорировались (испытуемый читал книгу), Р. Наата-иен выделил среди компонентов ССП особый потенциал, который коррелировал с произвольным вниманием. Эта волна получила название процессной негативности (ПН). Под влиянием инструкции быть внимательным к определенным стимулам ПН усиливается на все стимулы, но особенно на релевантные. Чем короче межстимульиый интервал, тем раньше возникает ПН (в среднем ее ЛП около 150 мс, но она может появляться и через 50 мс). ПН не тождественна волне Н.хотя и совпадает с ней по латенции. Критическим признаком, отличающим волну Н от ПН, является его инверсия при смещении активного электрода но слуховой коре относительно мостадоидов (сосцевидных отростков).
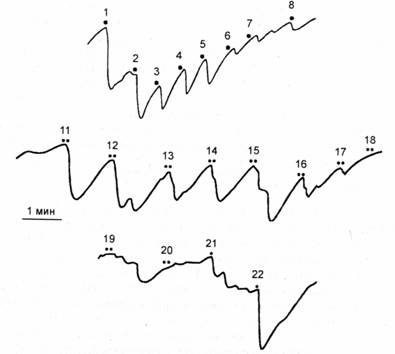 Рис.
10.2. Выработка условного ориентировочного
рефлекса при формировании ассоциации двух звуковых стимулов
Рис.
10.2. Выработка условного ориентировочного
рефлекса при формировании ассоциации двух звуковых стимулов
1-Я _ звук 1000 Гц, 70 дБ, 5 с; 11-20 — комплекс: 70 дБ-90 дБ; 21 — 70 дБ, 22 — 90 дБ. По оси ординат — сопротивление в кОм [поЕ. Н.Соколову, 1958]
относительно мостадоидов (сосцевидных отростков). Объясняя ПН, Р. Наатансп вводит понятие «следа селективного внимания», который связан с памятью. Он полагает, что произвольное внимание актуализирует в сенсорной памяти (по-видимому, во вторичной ассоциативной коре) след релевантного стимула. «След внимания» понимается им как механизм опознания стимула, так как входной сигнал всякий раз сличается со следом релевантного стимула, который непрерывно поддерживает в активном состоянии произвольное внимание. Чем ближе стимул к релевантному, тем больше амплитуда ПН. Р. Наатанеи выделяет раннюю ПН с локусом в ассоциативной коре, выражающую завершение процесса сличения, и позднюю ПН с фронтальной локализацией. Последняя отражает произвольное усилие, направленное на дальнейшую обработку стимула и включающее произвольный ОР па значимый стимул.
Суммируя результаты собственных исследований с данными Р. Наатанена и его коллег, Е. Н. Соколов [1997] выделяет два типа энграмм, с которыми связано возникновение непроизвольного и произвольного ОР. Когда единицы кратковременной памяти подключены к нейронам новизны и тождества прямо в виде потенциированных синапсов от детекторов-признаков, сигнал рассогласования, возникающий при несовпадении стимула с энграммой, определяет генерацию непроизвольного ОР. Его сигнал рассогласования представлен в HP и неспецифическом компоненте Нг Если же к нейронам новизны и тождества подключены единицы долговременной памяти и они обеспечивают произвольное удержание шаблона, то совпадение стимула с шаблоном порождает сигнал согласования или совпадения. В терминах нейронных популяций он представлен ПН. Сигнал совпадения достигает нейронов активирующей системы и вызывает условный ОР, направленный на значимый стимул. В случае несовпадения стимула с шаблоном сигнал совпадения и ПН не возникают и ОР на значимый стимул не появляется. При этом стимулы, отличающиеся от релевантного, вызывают безусловный ОР, непроизвольное внимание, которое представлено негативностью рассогласования и неспецифическим компонентом Н1.
10.2. Ориентировочно-исследовательская деятельность
Ориентировочную реакцию (как настройку анализаторов на лучшее восприятие нового стимула) следует отличать от исследовательских реакций и ориентиро вочно-исследовательского поведения. Последние могут следовать за ОР — они направлены па более детальное ознакомление с вызвавшим их объектом.
Сегодня можно считать доказанным, что ориентировочно-исследовательская деятельность животного и человека побуждается особой потребностью в получении новой информации. П. В. Симонов относит ее к группе идеальных потребностей, создающих основу для само развития индивида. На основе ориентировочно-исследовательского подкрепления (получения новой информации) возможна выработка самых различных условных инструментальных рефлексов у животных. Крысы обучаются различать цвета слайдов, чтобы попасть в более сложную часть лабиринта. Обезьяны выучивают сложные действия, чтобы иметь возможность выглядывать через окошко камеры в помещение лаборатории. Подкрепляющая функция ориентировочно-исследовательской активности обусловлена активацией эндогенных опиатов. Подтверждена связь ОР с положительными эмоциями и активацией бета-эн-дорфииов, снижающих болевую чувствительность и тормозящих оборонительные реакции. Во время исследовательского поведения активируются нейроны удовольствия центров самораздражения.
Согласно Д. Берлайиу, существуют две группы детерминант ориентировочно-исследовательского поведения. С одной стороны, это дефицит активации в организме, который побуждает к поиску стимулов, удовлетворяющих эту потребность. Берлайн называет этот тип поведения, который направлен на поиск стимуляции, разнонаправленным исследованием. Примером может служить поведение крысы, которая начинает нажимать на педаль, чтобы получить кратковременное освещение в лабиринте. Обезьяна, содержащаяся в клетке и часами открывающая окно, чтобы увидеть, что происходит за ее пределами, также стремится поднять свой уровень активации. С другой стороны, исследовательская деятельность побуждается неопределенностью ранее полученной информации. Это стимулирует особую форму активности в виде целенаправленной любознательности. Ее цель — получить недостающие уточняющие сведения и тем самым снизить неопределенность ситуации.
...
Сознание
Концепция «светлого пятна» Павлова получила свое развитие в новейших гипотезах в виде «теории прожектора». Данная концепция была сформулирована Ф. Криком — соавтором теории двойной спирали и нобелевским лауреатом. Здесь хотелось бы сделать небольшое отступление: можно сказать, что проблемы сознания как бы притягивают к себе внимание нобелевских лауреатов, хотя никто еще Нобелевскую премию за исследования сознания пока не получил. И. П. Павлов был только первым, но не последним в этом ряду, ведь проблема сознания — одна из важнейших и интереснейших в науке, и лучшие умы, воодушевленные высоким признанием, тянутся к ее разрешению.
Основные предпосылки и положения «теории прожектора» сводятся к следующему. Вся информация поступает в кору по сенсорным путям через переключательные ядра в дорзальном таламусс (включая и примыкающие к нему коленчатые тела). Возбудимость этих переключательных ядер может быть избирательно изменена за счет коллатералей от нейронов ретикулярного комплекса таламуса, входящего в его вентральный отдел. Взаимоотношения между этими частями таламуса построены таким образом, что в каждый данный момент одна из нейронных групп дорзалыюго таламуса оказывается в состоянии высокой возбудимости, что значительно усиливает импульсный ноток к коре, в то время как другие группы оказываются, наоборот, заторможенными. Период такой высокой возбудимости длится около 100 мс, а затем усиленный приток поступает к другому отделу коры. Ф. Крик предполагает па этом основании, что область наиболее высокой имнульсации представляет в данный момент как бы центр внимания, а благодаря перемещению «прожектора» в другие участки становится возможным их объединение в единую систему. В этом процессе большую роль играет синхронизация активности нейронных групп па одной частоте. В последние годы особое значение в процессах интеграции придается высокочастотному ритму — частотой около 40 Гц.
Важно, что в число вовлеченных в совместную деятельность нейронных групп входят нейронные ансамбли в различных областях коры. Подобная интеграция, по мнению автора, и обеспечивает осуществление высших психических функций. Предиолагаегся также, что одновременно могут действовать несколько «прожекторов». Данная концепция была разработана, в первую очередь, для обработки зрительных сигналов, но автор предполагает, что описанная схема является общей для всех анализаторов.
11.3. Повторный вход возбуждения
и информационный синтез
Концепция «светлого пятна» исходит из того, что сознание определяется некоторым уровнем возбудимости мозговых структур. Однако можно предположить, что лого недостаточно и в действительности возникновение психического связано с некоторым принципом в организации процессов мозга, предполагающим их определенное усложнение но сравнению с более простыми функциями мозга.
Одна из гипотез, привлекающих в последние годы внимание исследователей, — представление о возврате возбуждения в проекционные зоны коры после дополнительного анализа в других структурах мозга и о возникающих на этой основе сопоставлении и синтезе имевшейся ранее и вновь поступившей ипформации. Таким образом, схема рефлекса дополняется здесь еще одним звеном, что превращает рефлекторную дугу в кольцо. Впервые идея об информационном синтезе как мозговой основе возникновения субъективных переживаний была выдвинута в середине 70-х гг. прошлого века А. М. Иваницким [1999] на основе исследований физиологических механизмов ощущений.
11.3.1. Мозговая основа ощущений
Уже в 1920-х гг. психологам было известно, что ощущение возникает довольно поздно — после 100 мс от момента предъявления стимула (т. е. значительно позже прихода сенсорных импульсов в кору). В проведенной А. М. Иваницким и его со трудниками работе Оыла поставлена задача понять, что происходит за это время и какой стадии мозговых процессов соответствует возникновение субъективного образа. В исследовании при предъявлении одних и тех же стимулов записывали объективные показатели мозговой деятельности в виде вызванных потенциалов и определяли количественные показатели восприятия. Для получения последних использовались методы теории обиаруже мия сигнала, которая описывает процесс восприятия с помощью двух независимых переменных: показателя сенсорной чувствительности (I и критерия решения, определяемого мотивациоиными факторами. Основные выводы построены на вычислении корреляций между показателями физиологии и психологии, а именно амплитудой волн вызванного потенциала и психофизическими индексами. Исследования проведены на соматосеисорном и на зрительном анализаторах, причем полученные на обеих модальностях результаты были принципиально сходными.
Было установлено, что амплитуда ранних волн вызванного потенциала (ВП) обнаруживала статистически достоверную корреляцию с показателем еГ, а поздних волн — с критерием решения. Промежуточные волны с латентиостью 140 мс для соматосеисориого и 180 мс для зрительного анализатора коррелировали с обоими перцептивными индексами, причем такая двойная корреляция обнаруживалась только для волн проекционной коры. Амплитуда этих волн определялась, таким образом, как сенсорными свойствами стимула, так и его значимостью. На основании имеющихся данных о гепезе отдельных волн вызванного потенциала был описан механизм, обеспечивающий указанную двойную корреляцию.
В основе этого механизма лежало кольцевое движение нервных импульсов с «центральной станцией» в проекционной коре. Сначала возбуждение поступало от проекционной к ассоциативной коре. Для зрительных стимулов это были отделы височной коры, играющей важную роль в опознании стимулов, а для кожных раздражений — вторичные и третичные зоны соматосенсорной коры. Затем нервные импульсы приходили к структурам лимбико-гиппокампалыюго комплекса, играющего важную роль в процессах памяти, и подкорковым центрам эмоций и мотиваций. До этой стадии поступательное движение возбуждения от рецептивных зон к исполнительным центрам полностью укладывалось в схему рефлекса. Однако процесс на этом не заканчивался: он включал еще возврат возбуждения в кору, в том числе и в ее проекционные отделы. Благодаря возврату возбуждения нервные импульсы, приходящие из мотивациоииых центров, накладывались в нейронах проекционной коры на следы сенсорного возбуждения. На этом этапе или несколько раньше в процесс включалась и лооная кора, что проявлялось в синхронизации рисунка ВП в интервале от 100 до 200 мс после стимула. Было высказано предположение, что указанные промежуточные компоненты ВП отражали синтез на корковых нейронах двух видов информации: наличной информации о физических характеристиках стимула и извлекаемых из памяти сведений о его значимости.
Самое интересное, однако, заключалось в том, что латентиость этих волн ВП точно совпала со временем появления ощущения, измеренным ранее в психологических экспериментах. Следовательно, можно было предположить, что синтез двух видов информации — наличной и извлекаемой из памяти — и составляет тот ключевой механизм, который лежит в основе ощущения как феномена уже не физиологического, а психического уровня (рис. 11.1).
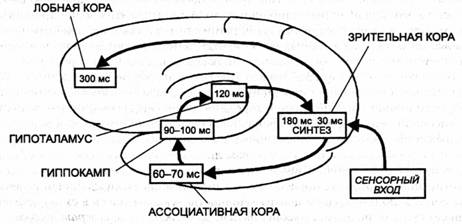 Рис. 11.1. Схема
кольиевого движения возбуждения при возникновении зрительных ошушений. Ведушее
звено в этом процессе — синтез информации о физических и сигнальных свойствах стимула
на нейронах проекционной коры
Рис. 11.1. Схема
кольиевого движения возбуждения при возникновении зрительных ошушений. Ведушее
звено в этом процессе — синтез информации о физических и сигнальных свойствах стимула
на нейронах проекционной коры
Другими словами, был сделан шаг к преодолению барьера между двумя уровнями организации мозговых процессов, один из которых не сопровождается, а другой сопровождается субъективными переживаниями. Согласно предложенной концепции, последовательное поступление информации от рецепторов приводит к повторному движению возбуждения но указанному кольцу, обеспечивая постоянное сопоставление сигналов, приходящих из внешней и внутренней среды, что и составляет психическии мониторинг происходящих изменении. сггот процесс осуществляется с периодом приблизительно в 150 мс, составляющим минимальную длительность ощущений. Позднее сходное кольцо возбуждения было описано М. Мишкиным у обезьян при анализе стимулыюй информации, образовании и извлечении памятных следов.
Идея о повторном входе возбуждения в нервные структуры как базисном механизме возникновения субъективного опыта высказывалась в последние годы многимн авторами, паиоолее полно она оыла развита в трудах нобелевского лауреата (за исследование антител) Дж. Эделмана (Edelman, Tonony, 2000), теория сознания которого получила достаточно широкую известность. По мнению этого автора, в основе возникновения субъективных феноменов лежит механизм повторного входа возбуждения в те же нейронные группы после дополнительной обработки информации в других группах или поступления сигналов из внешней среды, причем эти обратные проекции могут соединять как анатомически близкие, так и удаленные структуры. Этот повторный вход (гееп1епп&) дает возможность сравнивать полученные ранее сведения с изменениями, происшедшими за период времени от предыдущего повторного входа. Сходные концепции сознания были высказаны и другими авторами.
Для оценки предложенных концепций важное значение может иметь соотнесение временной шкалы мозговых процессов со временем субъективно переживаемых событий. По Дж. Эдельману, время одного цикла прохождения возбуждения составляет 100-150 мс. Если прибавить к этому время, необходимое для прихода сенсорных импульсов в кору, то в сумме получается время, достаточно близкое к тому, которое было найдено в наших экспериментах. Другие авторы считают, что в процессах сознания ведущую роль играют процессы, связанные с тета-ритмом, что также дает время около 150 мс.
В исследованиях Б. Либета и др. с регистрацией вызванных потенциалов на электрокожиые стимулы с поверхности коры во время нейрохирургических операций было показано, что на слабые, ноднороговые стимулы в коре регистрируются только ранние волны ответа с латентностыо до 100 мс. При усилении раздражения в ВП появляются и более поздние колебания с латентностью 150 мс, что сопровождается возникновением субъективных ощущений и практически совпадает с латентностыо волн ВП, обнаруживших в наших исследованиях двойную корреляцию с показателями восприятия.
Итак, идея повторного входа возбуждения и информационного синтеза в течение последних 20 лет высказывалась независимо друг от друга рядом авторов, относящихся к разным школам и строившим свои заключения на основе разных данных. Очевидно, подобное совпадение не может рассматриваться как случайное. Скорее, оно свидетельствует о приближении к истинному пониманию тех принципов организации мозговых процессов, которые лежат в основе субъективных переживаний.
Приведенные выше данные касались происхождения одного из наиболее простых психических явлений — ощущений. В связи с этим могут возникнуть вопросы: насколько данные закономерности универсальны и может ли принцип информационного синтеза быть распространен и на более сложные психические проявления, например на процесс мышления?
11.3.2. Механизмы мышления
Ответам на эти вопросы были посвящены работы А. М. Иваницкого и его сотрудников, проводивших исследования в течение последних 15 лет [Ивапицкий, 1999]. В работе ставилась задача изучить структуру корковых связей при различных мыслительных операциях. При этом авторы исходили из фундаментальных идей российской нейрофизиологической ШКОЛЫ Н. Е. Введенского — А. А. Ухтомского о том, что связь между нервными структурами возникает на основе уравнивания ритмов их деятельности. М. Н. Ливановым и В. С. Русииовым было показано, что эти ритмы находят отражение в частотных параметрах электроэнцефалограммы (ЭЭГ), из чего последовал решающий вывод о том, что синхронизация частотных характеристик биопотенциалов мозга может быть условием и индикатором внут-рикорковой связи.
В результате развития этих идей был создан метод, получивший название картирования внутрикоркового взаимодействия. В основе его лежат представления о том, что присутствие в спектрах ЭЭГ разных областей коры точно совпадающих частотных пиков является указанием на наличие в этих областях нейронных групп, работающих в одном ритме и, следовательно, функционально связанных друг с другом.
Испытуемым на экране монитора предъявлялись задачи на образное, пространственное и абстрактно-вербальное мышление. В нервом случае субъект должен был опознать эмоцию на фотографии лица (использовались четыре базовые эмоции: радости, страха, гнева и горя, а также смешанные состояния), во втором — сравнить две геометрические фигуры с целью определения их идентичности или зеркальной симметрии. В качестве вербальных задач использовались решение анаграмм или выбор из четырех слов одного, относящегося к другой смысловой группе
Было установлено, что простой и достаточно симметричный рисунок связей, характерный для состояния покоя, изменялся при умственной работе. Связи начинали сходиться к определенным областям коры, образуя как бы узлы или центры связей, названные фокусами взаимодействия. При этом топография фокусов взаимодействия оказалась специфичной для мыслительных операций разного знака. Так, при образном мышлении фокусы локализовались преимущественно в теменио-височпых областях, а при абстрактно-вербальном — в лобных отделах коры. Пространственные задачи, включавшие элементы обоих видов мышления, характеризовались образованием фокусов на более ранних этапах в задних, а затем и в передних отделах коры (рис. 11.2).
Кроме того, в исследованиях, в которых испытуемому давалось задание мысленно построить зрительный образ из ограниченного набора простых элементов, было показано, что у лиц с преобладанием первой сигнальной системы, но И. П. Павлову, фокусы взаимодействия локализовались преимущественно в правом, а у лиц с преобладанием второй, речевой системы — в левом полушарии. При этом на этапе нахождения образа фокусы были расположены в затылочных и височных (зона опознания) отделах полушарии, а на этапе конструирования образа — в лобной коре. Следует отметить, что нахождение решения при всех типах задач, даже если речевой ответ не требовался, сопровождалось включением в функцию вербальной левой височной зоны.
Обобщая эти данные, можно сделать вывод о том, что важным признаком организации корковых связей при мышлении является их конвергенция к определенным центрам — фокусам взаимодействия. При этом связи, подходящие к фокусу, устанавливаются на разных частотах; собственно, именно это обстоятельство и лежит в основе образования фокуса, так как связи на одной частоте образовывали бы однородную сеть, не имеющую центров. Можно предполагать, что каждая из связей приносит к центру из определенной области коры или подкорковых образований свою информацию. В фокусе эта информация может быть сопоставлена и перекомбииирована определенным образом. Основную функцию фокуса взаимодействия составляет, таким образом, информационный синтез, т. е. процесс, сходный с тем, который мы наблюдали в проекционной коре при возникновении ощущений.
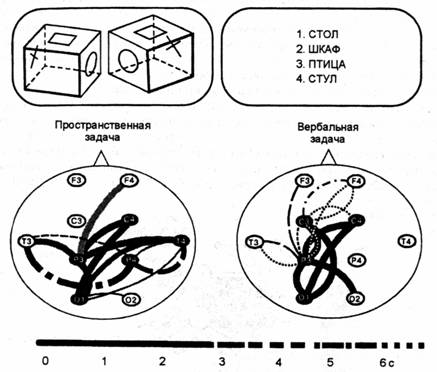 Рис.
11.2. Корковые связи в диапазоне частот бета^ритма при двух мыслительных операциях
Рис.
11.2. Корковые связи в диапазоне частот бета^ритма при двух мыслительных операциях
Вверху представлены примеры задач; слева — связи при задаче на пространственное мышление, которая заключалась в сравнении двух геометрических фигур, предъявляемых на экране монитора, с иелью определения их идентичности или зеркальной симметрии, что решалось путем мысленной ротаиии; справа — связи при задаче на вербальное мышление, которая состояла в том, что из четырех слов на экране монитора субъект должен был выбрать одно, относящееся к другой смысловой группе.
На рисунке представлены статистически значимые связи при сравнении со зрительно-моторным контролем, включавшим предъявление изображений и двигательный ответ, но не требовавшим мыслительных операиий. Толстые линии — связи на частоте 16-19 Ги, тонкие линии — связи на частоте 13-16 Гц. Шкала — время появления связи после предъявления задачи. Средние данные по группе из 43 человек
Основное различие состоит в том, что место сенсорного сигнала здесь могла занять информация, хранящаяся в оперативной памяти (например, об условиях решаемой задачи), а ведущая роль в процессах информационного синтеза принадлежала не проекционной, как при возникновении ощущении, а ассоциативной коре. Предполагается, что на основе происходящего в фокусе сопоставления и достигается конечная цель мыслительного процесса в виде нахождения решения. Субъективно все это переживается как процесс думания и нахождения ответа.
Гипотетическая структура фокуса взаимодействия состоит из групп нейронов с различными частотными характеристиками, настроенных на одинаковые с ними по частоте группы па периферии. Эти связи но своей природе должны быть двусторонними, т. е. как прямыми, так и обратными: если две группы имеют одну частоту, то любая из них в равной степени сносоона и воспринимать, и передавать информацию связанной с ней группе (в зависимости от соотношения фаз колебаний). Одно кольцо при ощущении заменяется здесь, таким образом, как бы системой колец, замыкающихся на один центр.
Внутри фокуса группы нейронов должны быть объединены связями, образованными на ином принципе: так как они работают па разных частотах, принцип изолабильности, т. е. одинаковой частоты смены циклов возбуждения, здесь не применим. Очевидно, это должны быть жесткие связи, основанные на структурных изменениях в синапсах. Данные связи эффективны в любой фазе цикла возбудимости нейрона или группы нейронов (рис. 11.3). Идея об обеспечении психической функции за счет сочетания жестких и гибких звеньев была впервые высказана Н. П. Бехтеревой [1980].
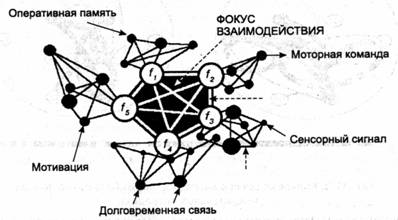 Рис.
11.3. Схема фокуса взаимодействия
Рис.
11.3. Схема фокуса взаимодействия
Фокус состоит из групп нервных клеток, отличающихся различными частотными параметрами и входяшими в состав нейросетей, где нейронные ансамбли объединены гибкими связями на основе идентичности частотных характеристик. Группы внутри фокуса соединены жесткими связями, основанными на структурных изменениях в синапсах. Благодаря такой структуре фокус получает информацию, циркулирующую в различных нейронных сетях, включая: сенсорную информацию, сведения, хранящиеся в оперативной и долговременной памяти, а также сигналы из центров мотивации. На основе сравнения и синтеза различной информации достигается решение, которое реализуется в моторных командах
...
Добавим, что последовательно развивая эти представления, можно сделать и следующий таг. Внутренняя логика психических событий такова, что она допускает возможность действительного выбора поведенческого ответа на основе субъективной, по реально существующей оценки значения того или иного фактора или мотива поведения. При этом осознание всей неоднозначности этих оценок определяет и возможность альтернативных решений.
...
12.1. Вилы и формы бессознательного
Понятие бессознательного в широком смысле включает в себя все психические явления вне сферы сознания, т. е. те содержания психической жизни, о наличии которых человек либо не подозревает в данный момент, либо не знает о них в течение длительного времени, либо вообше никогда не знал. В качестве примера можно привести факт неосознаваемости сигналов, непрерывно поступающих в головной мозг из самого организма, его внутренних органов мышц, суставов. Они начинают осознаваться в основном при патологии соответствующего органа, когда нарушается его функция. Однако в норме они создают определенный положительный чувственный фон, обеспечивающий нормальное самочувствие человека.
Следует подчеркнуть одно важное обстоятельство, общее для всех видов бессознательного: отсутствие осознания не есть чисто негативный факт, в смысле психическое — минус сознание, а результат активных процессов, происходящих в центральной нервной системе. Это хорошо видно на такой форме бессознательного, как автоматизированное поведение, когда с упрочением навыка все большая часть поступающей извне информации не доходит до сознания, а обрабатывается и переключается па эффекторную (двигательную) систему на более низких уровнях центральной нервной системы. Каждый человек знает, что в процессе обучения, особенно па его начальных стадиях, сознание принимает непосредственное участие в произвольной двигательной деятельности, например, при обучении игре па рояле, ходьбе, письму и т. д.
Приведем еще пример подобной бессознательной психической деятельности. В памяти человека хранятся тысячи слов. При разговоре из кладовой памяти извлекаются нужные и к месту слова для выражения определенной мысли автоматически, неосознанно. Сознание не участвует и в грамматическом построении речи. В последние годы показано, что эта речевая деятельность осуществляется на основе особой формы бессознательной (имплицитной) памяти (см. главу 6). Человек может включать по своему усмотрению сознание в эту деятельность, например, подыскивать наиболее подходящие, но его мнению, слова, но речь в этих случаях значительно замедляется, нарушается ее плавность.
Увлечение гипнозом, начавшееся более 100 лет назад, объясняется тем, что он как прием изучения бессознательного имеет несомненное преимущество перед другими методами, так как позволяет исследовать бессознательные психические явления в ходе эксперимента.
...
Еще одна форма бессознательного связана с загадочным психическим явлением, с так называемым озарением в творчестве. Бытует иллюзорное представление, будто творческий акт совершается в неведомых глубинах личности, результат его — решение научной или художественной задачи — выглядит как «случайный» успех, озарение (ипсайт), интуитивная догадка («ага»-реакция), т. е. как результат работы подсознания, особенно во сне («проснулся с готовым решением»). Объяснительная сила подобных мифов об иррациональном происхождении творчества ничтожна, поскольку они не содержат никакой перспективы для причинного объяснения открытия. В частности, неясно, каким образом рациональное знание, присущее науке, возникает непостижимым для логического созвательного анализа путем.
Решение давнего спора о том, следует ли у человека все психическое отождествлять с сознательным опытом или необходимо допустить существование бессознательных психических явлений, кроме естественнонаучного значения имеет несомненный методологический, философский аспект. Отрицание бессознательного с неизбежностью закрывает естествоиспытателю путь для выявления причинных связей и причинных отношений между отдельными явлениями психической жизни человека. Важнейшее свойство сознания — прерывистость, пали чие перерывов между отдельными элементами, нередко отсутствие между ними видимых связей. Понятие бессознательного заполняет пробелы между сознательными психическими явлениями. Это дает возможность изучать с позиций детерминизма (а это основной и обязательный принцип любого научного исследования) все психические функции у человека вплоть до самых высших ее форм.
Таким образом, бессознательное — это гносеологически необходимая категория. Если исходить из этого принципиального положения, а его справедливость подтверждается множеством фактов, полученных в экспериментально-психологических и психофизиологических исследованиях, то следует считать, что бессознательное — это такая же психическая реальность, как и сознательная психическая жизнь. Более того, бессознательное составляет значительную и весьма важную часть психической жизни человека, его познавательной, эмоциональной и поведенческой сфер деятельности.
В современной психофизиологии все больше употребляется термин неосознаваемый. Он, как и понятие о бессознательном, обозначает ряд неоднородных явлений. Некоторые ученые считают эти термины синонимами, другие — в основном психофизиологи — термин «неосознаваемый» относят к более узкому кругу явлений. К ним относят феномен, обозначаемый иногда также как предсознательный, — это психические процессы на начальных стадиях восприятия и обработки поступающей извне информации или же явления, которые в данный момент неосознаваемы, так как находятся вне сферы избирательного внимания субъекта, но они могут легко стать осознаваемыми при переключении на них внимания. Самый обычный пример такого феномена: человек перестает слышать тиканье настенных часов, если он какое-то время находится в комнате, где они висят, и снова начинает их слышать, когда но какому-то поводу обращает на них внимание.
Широкий круг психических явлений у человека, с которыми имеют дело психологи, психофизиологи и психиатры в норме и патологии, связан с неосознаваемым, в смысле подпороговым по отношению к сознанию, восприятием эмоционально или мотивационно значимых, но физически слабых сигналов, которые не достигают уровня сознания и не осознаются субъектом, однако вызывают вегетативные, биоэлектрические и эмоциональные реакции и могут влиять на поведение.
...
Воздействие неосознаваемых слов на способность к фантазированию исследовали у нормальных взрослых людей. Авторы такого рода работ исходили из предпосылки о том, что на содержание фантазий наиболее сильно влияют неосознаваемые раздражители. По предварительной инструкции испытуемые должны сочинить и написать историю о художественной картине, которая демонстрируется в течение 20 с. В этом же промежутке времени на экране мелькают слова, которые не осознаются. Треть испытуемых получает слова положительного эмоционального тона, другая треть — негативного, а остальные — нейтрального содержания. Рассказ испытуемого но поводу содержания и смысла картины анализируют два психолога, которые не знают о характере иодпороговых слов, предъявленных данному субъекту. Если для интерпретации предъявлялась картина с недвусмысленным содержанием, то не отмечалось какого-либо эффекта иодпороговых слов на характер свободного рассказа о картине. Достоверное влияние неосознаваемых слов на эмоциональную окраску рассказа наблюдалось в случаях, если картина была написана нечетко или же она была двусмысленного, неопределенного содержания.
Необходимо отметить, что неосознаваемые слова не вызывают больших модификаций фантазий. Как правило, они лишь усиливают наличный фон эмоционального состояния, если совпадают с ним. Например, слова положительного эмоционального тона действуют сильнее в случаях, когда человек должен описать картину с положительным эмоциональным содержанием.
Определяющая роль эмоционального состояния или мотивации в эффекте неосознаваемых слоев была показана в серии исследований американского психолога Д. Сиене. В ее опытах применялись физически слабые словесные стимулы, которые предъявлялись на экране на очень короткое время, так что только в 5% проб испытуемые были в состоянии отличить слайд со стимулом от «пустого» слайда и в 1% из этих проб — опознать слово. Таким образом, неосознаваемые словесные стимулы находились как бы у нижней границы пороговой зоны их действия. Основное наблюдение заключалось в пятикратной экспозиции неосознаваемого слова, а вслед за этим — списка из ясно видимых слов, вперемежку ассоциативно связанных и не связанных с неосознаваемым словом (список слов составляли на основании норм словесных ассоциаций, разработанных в Миниесотском университете). Список показывали испытуемому один раз, затем он должен был вспомнить приведенные в списке слова. За подиороговый эффект принимали разницу между количеством воспроизведенных слов, ассоциативно связанных и не связанных с неосознаваемым словом. Эффект неосознаваемого слова наблюдался только при определенных состояниях человека. Например, спустя 3-4 часа после приема пищи испытуемого просили оценить свое чувство голода в данный момент или же давали прочесть текст, в котором упоминалась вкусная еда. Этими способами оживляли пищевую мотивацию. Затем в качестве неосознаваемых стимулов применяли «пищевые» слова. В этих случаях подиороговый эффект был значительным. Если испытуемый находился в сытом состоянии, эффекта почти не было.
Таким образом, неосознаваемые словесные стимулы оказывают влияние на функцию запоминания и воспроизведения слов только при определенном, достаточно высоком уровне мотивации субъекта. При этом неосознаваемые стимул и функция, на которую он влияет, должны быть связаны с преобладающей в данный момент мотивацией или же с определенным эмоциональным состоянием. Только в этих случаях даже очень слабые неосознаваемые стимулы могут вызвать соответствующий эффект.
Следовательно, для проявления эффекта неосознаваемых слов необходимо, во-первых, чтобы они были эмоционально значимы в данный период времени для субъекта, во-вторых, чтобы уровень мотивации или эмоционального напряжения личности был достаточно высок. Наибольший иодиороговый эффект наблюдается при совпадении этих двух переменных факторов. Они не всегда принимаются во внимание при попытках выявить эффект неосознаваемых слов на те или иные физиологические и психологические функции. Это относится в значительной мере к проблеме пресловутого «25 кадра».
...
(тормозное влияние отрицательной значимости, блокировка нежелательного)
12.3. Феномен психологической зашиты
Экспериментальные исследования влияния на психические и физиологические функции человека слов, которые им не осознаются, были начаты в середине прошлого столетия группой американских психологов Иетю Ьоок. При предъявлениях отдельных слов на светодиодном экране было обнаружено, что у нормальных людей, студентов, порог опознания бранных слов («неприличные», табуированиые слова) явно повышен по сравнению с порогом нейтральных, т. е. обычных слов. На «табу» слова регистрировались КГР во время таких коротких экспозиций, при которых исследуемые еще не могли правильно назвать эти слова. Если исследуемые высказывали догадки, то такие догадки обычно не были связаны с этим словом. Было удивительно наблюдать, пишет психолог, как нормальная молодая девушка с ненарушенным зрением в отдельных случаях была не в состоянии прочесть «неприятное» слово, в то время как оно было ясно видно другому наблюдателю. Повышение порога опознания слова стали объяснять как «защиту восприятия», «эмоциональную защиту», а чаще как психологическую защиту, играющую роль «фильтра» в зрительном восприятии, который, насколько это возможно, предохраняет субъекта от осознания эмоционально неприятных раздражителей внешней среды. Феномен повышения порога осознания эмоционально неприятных вербальных стимулов был получен также экспериментальным путем: у здоровых взрослых людей вырабатывалась оборонительная условная реакция на отдель иые слоги при электрокожном подкреплении. После этого измеряли пороги зрительного опознания различных слогов. Оказалось, что порог опознания слога, па который была выработана оборонительная реакция, явно выше, чем на другие «нейтральные» слоги, хотя какой-либо разницы в степени употребления исследуемыми этих слогов в прошлом не было и нельзя было думать, что они подавляют свою реакцию.
В многочисленных психофизиологических исследованиях показано, что в случаях, когда порог восприятия «конфликтных», связанных с отрицательным эмоциональным переживанием слов повышен, регистрируется их подпороговый эффект, т. е. биоэлектрические и вегетативные реакции возникают раньше, чем исследуемый опознает слово, может его прочесть. Подпороговая зона — это разница между порогом опознания (осознания) слова и порогом биоэлектрических и вегетативных реакций па пего.
Величина подиороговой зоны у разных людей или же у одного и того же человека колеблется в широких пределах, она зависит от множества факторов (степень эмоционального переживания, уровень мотивации, значимость слова, индивидуальные особенности личности и т. д.). Подпороговый эффект в виде биоэлектрических и вегетативных реакций нередко наблюдается при относительно коротких экспозициях эмоционально неприятных слов, соответствующих нижней половине подпороговой зоны. С увеличением времени предъявления слова этот аффект может даже исчезнуть и вновь появиться уже при околопороговых параметрах стимула, что является, по-видимому, результатом частично осознанного различения слова.
Наличие неэффективной зоны, когда влияние слов не усиливается с увеличением их экспозиции в пределах иод-пороговой зоны, а нередко, наоборот, пропадает, служит еще одним экспериментальным доказательством возможности семантического дифференцирования на неосознаваемом уровне. Против предположения о том, что подпороговый эффект эмоциональных слов может быть обусловлен тем, что они «смутно осознаются», говорят результаты специальных опытов, которые показали, что догадки испытуемых о нейтральных словах были больше похожи на действительно предъявляемый стимул, чем в случаях применения эмоциональных слов.
Возникновение биоэлектрических и вегетативных реакций на эмоциональные слова при экспозициях, значительно меньших, чем пороги их опознания, наличие «неэффективной зоны» убеждают в том, что центральная нервная система человека при определенных условиях в состоянии осуществлять семантическое дифференцирование на неосознаваемом уровне, т. е. различать слова, которые не осознаются. Это происходит на основе эмоциональной памяти без оживления в сознании словесных символов в случаях, когда повышен порог осознания эмоционального слова.
...
В настоящее время не вызывает сомнения положение о том, что изменения порога осознания эмоционально значимых слов обусловлены дополнительной нервной имиульсацией из структур лимбической системы, в которых интегрируются механизмы эмоциональных реакций. Если это так, то надо думать, что восходящее «эмоциональное» воздействие па кору происходит до того, как слово осознается, что и приводит к изменению порога осознания. Это важный пункт для выявления нейрофизиологических механизмов изменений восприятия эмоционально значимых слов, в частности повышения порога их осознания, а следовательно, и для понимания сущности безотчетных эмоций.