МОЗГ Д. Хьюбел
Содержание
Предисловие редактора перевода
В 1979 году американский междисциплинарный (это определение мне кажется более точным, чем «научно-популярный») журнал «Scientific American» почти целиком посвятил свой сентябрьский номер описанию наиболее значительных достижений в области изучения строения и деятельности мозга. В качестве авторов выступили ученые, непосредственно причастные к этим достижениям, внесшие в развитие науки о мозге свой ощутимый вклад. Такие имена, как Д. Хьюбел, Т. Визель, Э. Кэндел, Э. Эвартс, У. Наута и другие, хорошо известны специалистам всего мира, без этих имен трудно представить себе современную нейрофизиологию.
В данном случае они предстали перед читателем в качестве увлекательных рассказчиков, весьма осторожных в оценке полученных результатов, зато ясно отдающих себе отчет в том, как неполны и противоречивы наши реальные знания о деятельности мозга — сложнейшего и загадочного порождения Природы. Доступная широким кругам читателей информация «из первых рук» всегда остается идеалом популяризации любой области знания. В лежащей перед нами книге это условие соблюдено максимально.
Прогресс науки о мозге неоспорим и очевиден. По справедливому замечанию Д. Хьюбела, нейрофизиолог 50-х годов был бы поражен, если бы ему сказали, как много нового станет нам известно тридцать лет спустя. Непрерывное накопление информации о биологически активных веществах-передатчиках и модуляторах нервного возбуждения — чем - то напоминает ситуацию в области ядерной физики, где с каждый годом увеличивается семейство открытых элементарных частиц. Постепенно проясняющееся значение нарушений химизма мозга в генезе психических заболеваний коренным образом меняет облик описательной психиатрии. Поистине революционизирующее влияние на представления о физиологических основах человеческой психики-сознания и мышления оказало открытие функциональной специализации двух полушарий. Эти и другие успехи нейрофизиологии подробно освещены в настоящем издании.
Вместе с тем содержащиеся в нем материалы отнюдь не исчерпывают того, что достигнуто современной наукой. Например, в статьях американских коллег совершенно отсутствуют результаты, полученные советскими исследователями мозга. Это касается прежде всего таких разделов, как изучение фундаментальных закономерностей физиологии нервной клетки (П. Г. Костюк, А. И. Шаповалов), физиологии зрения (А. Л. Бызов, В. Д. Глезер, И. А. Шевелев) и слуха (Г. В. Гершуни), нейрофизиологических механизмов организации движения (В. С. Гурфинкель, Г. Н. Орловский, М. Л. Шик). Весьма значителен вклад советских исследователей в анализ клеточных механизмов обучения-формирования условных рефлексов (Н.Н.Василевский, Л.Л.Воронин, У. Г. Гасанов, А. Б. Коган, М. Н. Ливанов, М. Я. Рабинович, В. С. Русинов, Е. Н. Соколов, В. Б. Швырков, Г. И. Шульгина, М. М. Хананашвили и многие другие). Принципиальное значение для понимания закономерностей высших форм интегративной деятельности мозга имеют теоретические обобщения П.К.Анохина, Э.А. Асратяна, И. С. Бериташвили, Н.А.Берн штейна, Л. А. Орбели и других отечественных ученых.
Из перечня охваченных тем составителями исключены сведения, касающиеся строения и функций тех мозговых структур, которые непосредственно вовлечены в процесс актуализации важнейших потребностей организма - голода, жажды, полового влечения, в генез эмоциональных реакций — ярости, страха, удовольствия и т. п. В результате становится непонятным, как связаны столь тщательно изученные сенсорные системы (зрение, слух и т.п.) с не менее глубоко проанализированными механизмами организации движений. Иными словами, где локализованы структуры мозга, решающие вопрос о том, на какие воздействия внешней среды и каким образом должен отвечать живой организм?
Здесь-то и возникает та проблема «гомункулуса», о которой пишет в своей заключительной статье Ф. Крик. С редкой проницательностью и остротой один из первооткрывателей генетического кода ставит этот вопрос перед умом естествоиспытателя. Как часто самые разные авторы, вне зависимости от того, оперируют ли они понятием «информации», «нейропсихического кода» или «субъективного образа», забывают о существовании проблемы «получателя информации, смотрящего на экран маленького телевизора в мозгу», или заменяют гомункулуса таким психологическим понятием, как «личность».
Ф. Крик не предлагает своего решения проблемы, но совершенно справедливо требует раз и навсегда расстаться с идеей гомункулуса в любых ее вариантах, поскольку отказ от ложного пути сам по себе представляет немалый успех ищущей мысли. В отличие от всех других объектов естествознания изучение мозга человека отягощено возможностью интроспекции, т.е. возможностью взгляда «изнутри». Очевидность и наглядность представлений о своем собственном внутреннем мире затрудняют постижение законов деятельности мозга в несопоставимо большей степени, чем наглядность движения Солнца вокруг Земли столетиями искажала картину Солнечной системы. Отсюда и возникает вопрос, прозвучавший в самом начале: может ли мозг понять мозг, может ли мозг понять разум?
Лучшим ответом на этот вопрос являются работы, помещенные в настоящем выпуске, реальные достижения современной науки. Быть может, одно из важных достоинств этих статей как раз и состоит не только в информировании читателя о том, что уже познано, но и в уточнении формулировки нерешенных проблем, в определении стратегических задач дальнейших исканий.
П. В. Симонов
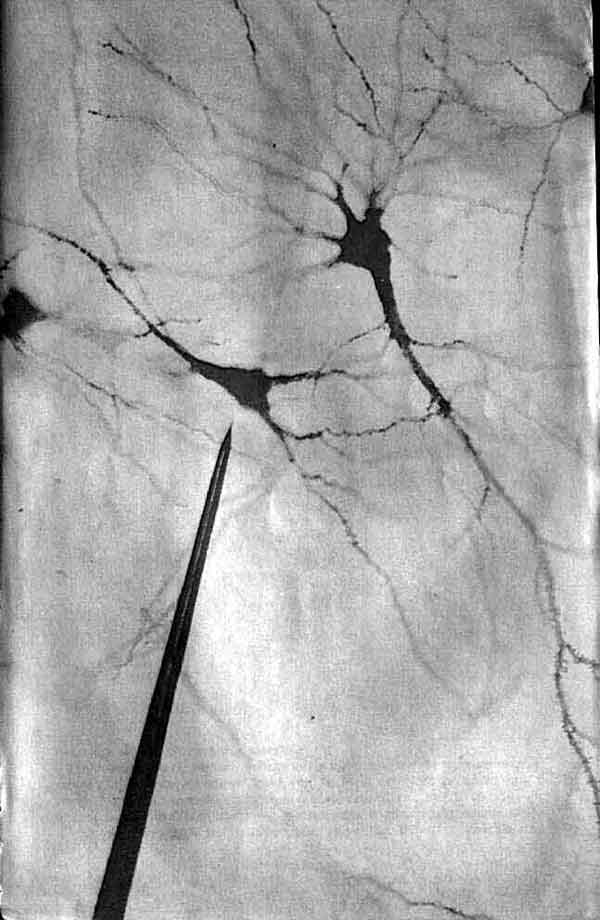
Три основных орудия нейробиолога (символическое изображение)-это микро-скоп, избирательное окрашивание нервной ткани и микроэлектрод. На рисунке приведена световая микрофотография нескольких окрашенных по Гольджи нейронов в срезе ткани мозга из зрительной коры обезьяны; длинная игла - кончик микроэлектрода, расположенного как бы для записи электрических импульсов, которые генерирует одна из клеток. Фотография получена Ф. Горо (F. Goro).
Д. ХЬЮБЕЛ
Мозг
Вступление к выпуску журнала, посвященного нейробиологии и ее центральной проблеме: как работает человеческий мозг? Несмотря на значительные достижения, этот вопрос остается одним из самых трудных в современной науке
Может ли мозг понять мозг? Может ли он понять разум? Что он такое - гигантский компьютер, или какая-либо иная гигантская машина, или же нечто большее? Эти вопросы задаются постоянно, и было бы полезно избавиться от них. Утверждение, что мозг не может быть понят мозгом, аналогично афоризму, что человек не в состоянии поднять себя сам за ушки собственных сапог. Но такая аналогия не бесспорна. Несомненно, даже беглый взгляд на то, что достигнуто на пути к пониманию мозга, убедит любого читателя этого выпуска «Scientific American», что со времен френологии мы ушли далеко вперед. Темп этого движения сейчас очень быстрый. Таким образом, с учетом всех практических задач нейробиологи работают в ожидании того, что смогут понять мозг, и в настоящее время они в этом преуспевают.
Думаю, что трудности, связанные с вопросами подобного рода, носят семантический характер. Они несут на себе груз таких слов, как «понимать» и «разум» - слов для многих целей полезных, но с расплывчатыми очертаниями и неуместных в применении к этим вопросам, которые из-за них становятся бессмысленными или неразрешимыми.
Мозг представляет собой ткань - запутанную, сложно сотканную, сложнее всего, что нам известно во Вселенной, однако, как любая живая ткань, она состоит из клеток. Вместе с тем, хотя это высокоспециализированные клетки, их активность подчинена законам, которые управляют любыми другими клетками. Их электрические и химические сигналы можно обнаружить, зарегистрировать и истолковать; вещества, из которых они состоят, можно определить; связи, образующие густую сеть, можно проследить. Короче говоря, мозг можно изучать совершенно так же, как изучают любой другой орган, скажем почку.
Проблема возникает тогда, когда мы ставим вопрос о понимании, потому что такое слово привносит сложность внезапного откровения или прояснения, предполагает наличие такой минуты, когда можно сказать, что из темного туннеля мы выходим на свет. Но мне не ясно, может ли в данном случае наступить такая минута и узнаем ли мы о ее наступлении.
Исследование мозга - это древняя область науки, а область спекуляций о мозге — еще древнее. Его изучение сильно ускорилось к концу XIX в.; новые методики, разработанные после второй мировой войны, привели к значительным успехам, и в последнее десятилетие нейробиология стала одной из самых активных отраслей науки. Следствием этого недавно явился подлинный взрыв открытий и прозрений. Но тем не менее изучение мозга только начинается. Констатация невероятной сложности мозга стала штампом, однако же это факт.
Проблема понимания работы мозга в чем-то сходна с проблемой понимания структуры и функции белков. Каждый организм содержит миллионы сложных изощренных молекулярных комбинаций, причем один человек совершенно отличен от другого. Для того чтобы детально изучить структуру хотя бы одного белка, по-видимому, потребуются годы, не говоря уже о том, чтобы узнать точно, как он работает. Если понять белки — это значит узнать, как все они работают, то перспективы здесь, пожалуй, отнюдь не радужные. Точно так же мозг состоит из очень большого числа (хотя и не миллионов) функциональных подразделений, из которых каждое обладает своей особой архитектоникой и своей сетевой схемой; а дать описание одного из них вовсе не значит описать их все. Поэтому понимание пойдет медленно (хотя бы по практическим причинам — в зависимости от времени и числа занятых этим людей), неуклонно (надо надеяться) и будет развиваться по асимптоте, безусловно с прорывами, но вряд ли достигнет конечной точки.
Разум — это тоже полезное слово, но (увы!) еще более расплывчатое. Поскольку оно поддается лишь нечеткому определению, говорить о его понимании (не слова, а того, к чему оно относится) - это значит говорить об упражнении в мыслительной гимнастике, которая, по-видимому, лежит за пределами естественных наук. Математик Г. Харди (G. Hardy) будто бы сказал, что математик — это тот, кто не только не знает, о чем говорит, но и не интересуется этим. Тех, кто углубляется в такие вопросы, как физиология разума, вероятно, это занимает, но я не представляю себе, как они когда-либо смогут ответить на эти вопросы.
Число нервных клеток, или нейронов, из которых состоит примерно полтора килограмма человеческого мозга, достигает порядка 1011 (сто миллиардов) или же на порядок больше или меньше. Вокруг нейронов лежат, их поддерживают и питают глиальные клетки, которых тоже очень много. Типичный нейрон имеет тело (диаметром от 5 до 100 микрометров), от которого отходит одно крупное волокно, аксон, и множество мелких ветвей, дендритов. Аксон отдает веточки в начале и часто сильно ветвится в конце. Говоря в общем, дендриты и тело клетки принимают поступающие сигналы; тело клетки комбинирует и интегрирует их (грубо говоря, усредняет) и выдает выходные сигналы; оно также поддерживает общее состояние клетки; аксон проводит выходные сигналы к своим окончаниям, которые передают информацию следующей группе нейронов.
Сигналы бывают двоякими - электрическими и химическими. Сигнал, генерируемый нейроном и проводимый по его аксону, представляет собой электрический импульс, но от клетки к клетке он передается молекулами передатчиков, медиаторов - веществ, которые перемещаются через особый контакт - синапс - между структурой, доставляющей информацию (окончанием аксона или, в отдельных случаях, дендритом), и структурой, воспринимающей ее (дендритом, телом клетки, или, в отдельных случаях, окончанием аксона). К одному нейрону обычно подходят отростки сотни или тысячи других нейронов, и он в свою очередь отдает веточки к сотням или тысячам других нейронов.
Этого, пожалуй, достаточно, чтобы сделать возможной попытку сравнить мозг с компьютером. Большинство нейробиологов при обсуждении этого вопроса согласятся с тем, что мозг можно рассматривать как машину, которая не обладает свойствами, лежащими за пределами возможностей научного исследования. Но верно и то, что с этим согласятся не все нейробиологи. В то же время несомненно все будут согласны с тем, что компьютер — это машина и ничего больше. И таким образом, в зависимости от вкусов и убеждений, мозг и компьютер в определенном смысле или сходны по своим основным качествам, или даже совершенно различны. По моему мнению, нельзя решить этот вопрос рассуждениями.
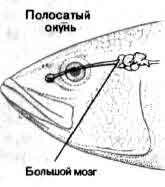
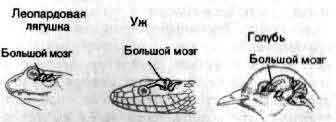
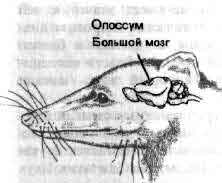
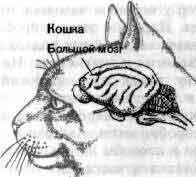
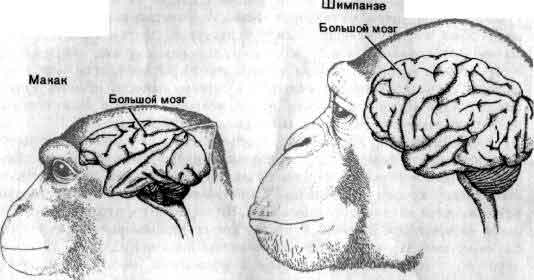
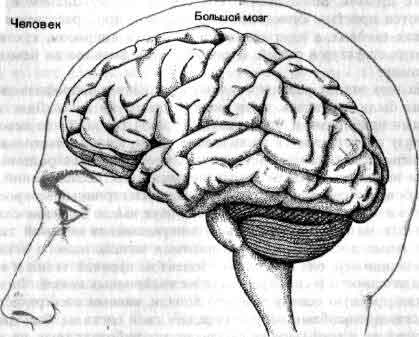
Увеличение размеров большого мозга у позвоночных. В одном масштабе показан мозг некоторых различных классов. У рыб, амфибий, рептилий и птиц большой мозг невелик. У хищных млекопитающих и особенно у приматов величина и сложность мозга резко возрастают.
Если считать машинами и мозг и компьютер, то в таком случае как их сравнивать? Такое упражнение представляет интерес. Компьютеры изобретены человеком и поэтому совершенно понятны, если принять, что человеческие существа вообще что-нибудь понимают; но чего люди не знают, это какими окажутся будущие компьютеры. Мозг создан эволюцией и во многих важных отношениях остается непонятным. Обе машины перерабатывают информацию и обе имеют дело с сигналами, грубо говоря, электрическими. В самых крупных вариантах и та и другая машина содержит множество элементов. Однако в этом между ними имеется интересное различие. Биологическим путем клетки производятся достаточно просто, и нейроны создаются в поистине огромных количествах. Между тем, умножить число элементов в компьютере, по-видимому, не так легко, даже несмотря на то, что это число быстро растет. Но если принять, что элементами нервной системы являются не нейроны, а синапсы, то я не могу себе представить, что компьютеры могли бы сравняться с ней. Никто не станет гадать, сколько в мозгу синапсов, но число 1014 (100 триллионов) не кажется невероятным. Еще большее значение имеет одно качественное различие. Мозг не подчиняется чему-либо подобному линейной программе, во всяком случае, те его части, о которых нам хоть что-нибудь известно. Он скорее сходен с цепью в радиоприемнике или телевизоре, а может быть, с сотнями или тысячами таких цепей, соединенных последовательно и параллельно, со множеством перекрестных соединений. Мозг, по-видимому, основан на принципе относительно жестко запаянного блока из сложных цепей, элементы которого работают на низких скоростях, измеряемых тысячными долями секунды; компьютер подчиняется программам, содержит гораздо меньше элементов и работает на скоростях, при которых имеют значение миллионные доли секунды. В числе мозговых сетей должно быть много таких, которые служат для поддержания эволюции, связанной с борьбой за существование и половыми инстинктами. Компьютер пока что свободен от всего этого; он эволюционирует другими способами.
Как надо изучать такой орган, как мозг? Основной подход состоит, разумеется, в том, чтобы изучить его составные части, а затем попытаться узнать, как они действуют сообща. Это осуществляется главным образом на животных, а не на человеке. Принципы нейронной функции удивительно сходны у столь далеких друг от друга животных, как улитка и человек; большая часть того, что известно о нервном импульсе, изучена на кальмаре. Даже основные структуры головного мозга так схожи. например у кошки и человека, что нередко не имеет значения, чей мозг изучать. Наряду с этим нейробиология отличается широким диапазоном привнесенных в нее подходов и методик, от физики и биохимии до психологии и психиатрии. Ни в одной другой области исследования не так важен этот широкий подход, который здесь начали применять в последние годы.
Нейроанатомия и нейрофизиология — две традиционные смыкающиеся ветви нейробиологии. Анатомия стремится описать различные элементы мозга и способы их объединения; физиология занимается изучением того, как эти части работают вместе. Исследователи этих двух областей пошли было разными путями и даже обосновались на разных университетских кафедрах, но в действительности они тесно связаны друг с другом. Большинство современных нейроанатомов не довольствуются простым описанием структуры и пространственных отношений как таковых, а идут дальше и задаются вопросом, каково назначение этих структур и связей. В свою очередь физиология невозможна без анатомии.
На всех этапах развития и нейроанатомия и нейрофизиология вынуждены были выжидать, пока физические науки не снабдят их необходимыми приборами и методиками. Нейрон так мал, что невооруженному глазу представляется лишь в виде точки, и также настолько мал, что его сигналы нельзя зарегистрировать простыми электродами. Для того чтобы выйти за пределы самых примитивных исследований, анатомии потребовался сначала световой, а затем электронный микроскоп, а физиологии — микроэлектрод. Для обеих наук имело значение создание специальных методик избирательного окрашивания нервной ткани.
Главные достижения нейроанатомов начала нашего века состояли в признании нейрона основным элементом нервной ткани и в открытии упорядоченности и специфичности межнейронных связей. Физиологи заложили прочную основу тем, что поняли, какими электрическими и химическими способами нейрон передает свои сигналы. Эти два вида достижений ни в коей мере не раскрыли, как работает мозг, но они служат совершенно необходимым фундаментом для этого. Один из способов увидеть, как далеко вперед ушла нейробиология (и, безусловно, как невероятно далеко ей еще надо идти), состоит в том, чтобы рассмотреть некоторые исторические этапы на пути к современному пониманию мозга и дать краткий обзор теперешнего состояния исследований в некоторых разделах этой области.
Почему, прежде всего, так трудно было установить, что отдельный нейрон служит основным элементом нервной ткани? Главными препятствиями являлись ничтожные размеры, фантастические формы и огромное разнообразие форм этих клеток, а также тот факт, что ветви, принадлежащие соседним клеткам, тесно переплетаются между собой. Слово «клетка» вызывает в воображении представление о кирпичике или кусочке студня; на самом же деле нейрон выглядит как миниатюрное деревце с разветвленной кроной, ствол которого имеет толщину от 10 до 20 мкм (тысячных долей миллиметра) и длину от 0,1 мм иногда до метра. Для того чтобы увидеть отдельные нейроны, требуется не только микроскоп, но и краситель, выделяющий их из окружения. Обычно нейроны упакованы так плотно, что в любом участке сотни их системами своих ветвей переплетены в густую чащу, в которой соседние веточки разделены пленками жидкости толщиной всего лишь около 0,02 мкм, и таким образом фактически все пространство занято клетками и их разнообразными отростками; а если на данном участке окрашены все клетки, то в световой микроскоп виден лишь плотный мазок.
Поэтому самым важным достижением нейроанатомии (после самого микроскопа) было открытие, сделанное примерно в 1875 г. итальянским анатомом К. Гольджи. Он изобрел метод, при котором одновременно окрашивается, по-видимому, в случайном порядке, лишь очень малая доля всех клеток данного участка, но зато эти клетки окрашиваются целиком. При хорошо выполненном окрашивании по Гольджи вместо невразумительной мазни на препарате видны лишь несколько нейронов, но каждый из них полностью, со всеми своими ветвями. Просмотрев много срезов мозга, окрашенных по Гольджи, анатом может дать перечень разных клеток в этой ткани. Доныне никто не знает, как и почему срабатывает метод Гольджи, окрашивая полностью одну из 100 клеток и совершенно не затрагивая все остальные.
Современник Гольджи, испанец С. Рамон-и-Кахал посвятил всю свою поразительно плодотворную жизнь приложению нового метода практически ко всем частям нервной системы. Его гигантская «Histologie du systeme nerveux de l'homme et des vertebres» («Гистология нервной системы человека и позвоночных животных»), впервые опубликованная в 1904 г. на испанском языке, до сих пор остается самой фундаментальной монографией по нейробиологии. Во времена Кахала шел спор о степени непрерывности между клетками. Отделены ли клетки одна от другой полностью, или же они соединены от аксона к дендриту в непрерывную сеть? Если бы существовала непрерывность протоплазмы, то сигналы, генерируемые одной клеткой, могли бы переходить в соседнюю, не прерываясь; если же непрерывности нет, то тогда должен существовать специальный процесс генерации сигналов заново в каждой клетке.
На препаратах Кахала, окрашенных по Гольджи, выявляется множество обособленных, полностью окрашенных клеток и никогда не было видно ничего похожего на сеть. Таким образом, его первым большим вкладом явилось представление о нервной системе как о совокупности отдельных, обособленных клеток, которые сообщаются друг с другом с помощью синапсов.

К. Гольджи (1844-1926) дал ключ к микроскопическому исследованию нервной системы, разработав (1875 г.) метод избирательного окрашивания нервной ткани, при котором в данном участке одновременно окрашивается лишь небольшая доля клеток, но зато полностью. Фотография сделана в начале восьмидесятых годов, в бытность Гольджи профессором гистологии и общей патологии в Университете в Павии.
С. Рамон-и-Кахал (1852-1934) посвятил свою жизнь тщательному изучению при помощи метода Гольджи буквально всех частей нервной системы множества разных животных. Признанием трудов Гольджи и Рамон-и-Кахала по изучению строения нервной системы явилось присуждение им в 1906 г. Нобелевской премии по физиологии и медицине. Рамон-и-Кахал преподавал в нескольких испанских университетах и дольше всего в Мадридском. Эта фотография сделана самим Рамон-и-Кахалом в двадцатых годах.
Кахал внес второй вклад, пожалуй, еще более значительный. Он собрал множество данных о том, что невероятно сложные связи между нейронами не случайны, как иногда думали, а представляют полную противоположность случайности, т.е. высоко структурированы и специфичны. Он дал исчерпывающее описание архитектоники десятков различных структур мозга и в каждом случае идентифицировал и классифицировал разные клетки, а иногда показывал, насколько позволяли его методы, как эти клетки связаны между собой. Со времени его трудов стало ясно, что если нейробиолог хочет понять мозг, он должен не только изучить, как построены разные его части, но и раскрыть их назначение и детально исследовать их работу как отдельных структур и в совокупности. Но сначала нужно узнать, как отдельный нейрон генерирует сигналы и передает их следующей клетке. Хотя Кахал, быть может, никогда не формулировал проблемы, связанные с пониманием мозга, с полной четкостью именно в этих выражениях, но знакомясь с его трудами, нельзя не извлечь из них такого вывода.
Долгое время нейроанатомам приходилось довольствоваться все более подробными описаниями, основанными на световой микроскопии с окрашиванием по Гольджи и по Нисслю (Nissl) (последнее выделяет тела отдельных клеток без дендритов и аксонов). Первым действенным орудием прослеживания связей между разными мозговыми структурами — например, между разными областями коры большого мозга или между корой и стволом мозга и мозжечком - явился метод окрашивания, который предложил в начале пятидесятых годов нашего века в Голландии У. Наута (W. Nauta), который теперь работает в Массачусетском технологическом институте. Этот метод основан на том, что при разрушении нейрона (механическим, электрическим или тепловым воздействием) отходящее от него нервное волокно дегенерирует и, пока оно еще не совсем исчезло, окрашивается иначе, чем соседние нормальные волокна. Если разрушить определенную часть мозга и через несколько дней окрасить мозг методом Науты, а затем исследовать под микроскопом, то наличие избирательно окрашенных волокон в какой-либо другой и, возможно, даже отдаленной его части будет означать, что эта часть получает волокна от разрушенного участка. Такой метод привел к необычайному расширению и детализации карты мозга.
За последнее десятилетие благодаря новейшим эффективным методам нейроанатомия продвинулась вперед больше, чем за предыдущие 50 лет (см. У. Наута и М. Фейртаг «Организация мозга»). Успехи достигнуты отчасти благодаря усовершенствованным химическим методикам и лучшему пониманию того, как различные вещества воспринимаются нейронами и передаются в обоих направлениях вдоль нервных волокон. Типичным примером может служить радиоавтография. Радиоактивное вещество вводится в ту или иную структуру мозга; тела клеток поглощают его, пересылают по своим аксонам, и оно накапливается в их окончаниях. Если затем приготовить срез ткани мозга, наложить его на фотоэмульсию и исследовать под микроскопом расположение проявленных зерен серебра, удается выявить «места назначения» аксонов. Можно вводить другие вещества, которые, наоборот, воспринимаются нервными окончаниями и передаются по аксонам в обратном направлении - к телу клетки, выявляя место возникновения аксона.
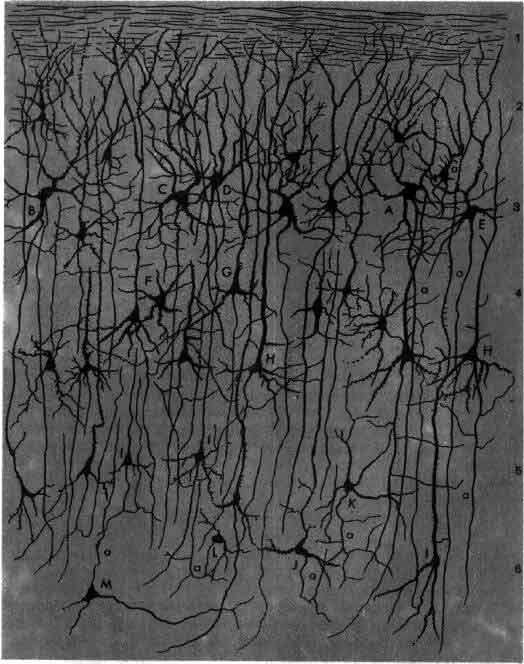
Окрашенная по Гольджи нервная ткань из зрительной коры крысы зарисована Рамон-и-Кахалом в 1888 г. Цифры по правому краю обозначают слои клеток; заглавными буквами помечены отдельные нейроны. Одним из самых важных вкладов Рамон-и-Кахала в нейробиологию явилось доказательство того факта, что нейрон представляет собой отдельную, обособленную клетку, а не элемент непрерывной сети.
Последним в этом ряду достижений явилась методика, разработанная несколько лет назад Л. Соколовым (L. Sokoloff) в Национальном институте охраны психического здоровья. Глюкоза служит «топливом» для нейронов и в активном состоянии клетки потребляют больше глюкозы, чем в покое. Меченая дезоксиглюкоза усваивается клетками, как если бы это была глюкоза. Она расщепляется, как глюкоза, но продукт первого этапа ее метаболизма не подвергается дальнейшим превращениям. Не имея поэтому возможности выйти из клетки, этот продукт скапливается в ней, и степень радиоактивности в определенных клетках указывает на их функциональную активность. Можно поставить, например, такой опыт: ввести это вещество внутривенно лабораторному животному, а затем предъявить звуковой раздражитель; микроскопическое исследование мозга позволит выявить те его области, которые связаны со слухом. Совсем недавно разработана новая методика-позитронно-эмиссионная томография, которая позволяет обнаруживать с помощью наружных датчиков присутствие дезоксиглюкозы или других веществ, меченных радиоактивными изотопами, испускающими позитроны. Эта перспективная методика делает возможным картирование активных структур мозга in vivo у лабораторного животного или у человека.
Применение всех существующих методик для выявления в первом приближении, без деталей, связей в одной только структуре — скажем, в части коры больших полушарий или в мозжечке — может занять у одного-двух анатомов пять или десять лет. Законченных нейроанатомов, людей особой породы, часто одержимых, а порой даже полупараноиков, во всем мире насчитывается всего несколько десятков. А поскольку мозг состоит из сотен разных структур, становится ясно, что одного только понимания связей в головном мозгу придется ждать еще много лет.
Кроме того, изучить связи данной структуры в мозгу — это еще не значит понять ее физиологию. Для этого надо прежде всего узнать, как работают отдельные нейроны. За последние три-четыре десятилетия стало более или менее понятно, как один нейрон генерирует электрические сигналы и передает информацию другим клеткам. Над выяснением этого механизма трудились многие ученые, в их числе: Г. Дэйл (Н. Dale), О. Леви (О. Loewi), А. Ходжкин (A. Hodgkin), А. Хаксли (A. Huxley), Б. Катц (В. Katz), Дж. Экклс (J. Eccles) и С. Куффлер (St. Kuffler). Одно из удивительных открытий состояло в том, что все нейроны, несмотря на различия по величине и форме, используют два одинаковых вида электрических сигналов: градуальные потенциалы и потенциалы действия.
Весь нейрон — его тело, длинный аксон и ветвящиеся дендриты — поляризован таким образом, что внутри он заряжен отрицательно приблизительно на 70 мВ по отношению к наружной поверхности. Этот «потенциал покоя» создается двумя свойствами клеточной мембраны. Во-первых, мембрана активно переносит ионы, выводя из клетки положительно заряженные ионы натрия и пропуская внутрь положительно заряженные ионы калия, вследствие чего концентрации этих двух видов ионов внутри клетки и снаружи совершенно различны. Во-вторых, способность проникать через мембрану у натрия и калия также различна.
Изменения разности потенциалов, которая создается таким образом между наружной и внутренней сторонами клетки в состоянии покоя, -это электрические сигналы нервов. Изменение трансмембранного напряжения на любом участке клетки или ее отростков быстро распространяется по мембране во всех направлениях, постепенно затухая; уже на расстоянии нескольких миллиметров сигнал вряд ли удается обнаружить. Это электрический сигнал первого рода, т.е. градуальный потенциал. Его основная функция состоит в передаче сигналов на очень короткие расстояния.
Сигнал второго типа, потенциал действия, передает информацию на большие расстояния. Если мембрана деполяризована (ее потенциал снижен) до критического уровня — от уровня покоя, равного 70 мВ, до приблизительно 50 мВ, - то наступает внезапное, резкое изменение: на время снимаются существующие препятствия току ионов калия и натрия и возникает локальный поток ионов, достаточный для того, чтобы изменить знак мембранного потенциала, который становится положительным внутри и достигает 50 мВ, а затем полярность снова меняется и восстанавливается нормальный потенциал покоя. Все это занимает около тысячной доли секунды. За это время первое изменение знака (на положительный внутри) создает мощный градуальный потенциал, который распространяется и доводит соседний участок мембраны до критического уровня; это вызывает изменение полярности в следующем участке мембраны, что в свою очередь создает изменение знака в соседнем участке. В результате временное изменение полярности быстро распространяется по нервному волокну.
Этот распространяющийся потенциал действия, «бегущий» по всей длине волокна, не ослабевая, и является нервным импульсом. Любая сигнализация в нервной системе на расстоянии миллиметра и более происходит в форме импульсов. Независимо от типа волокна и от того, связано ли оно с движением, зрением или мышлением, сигналы везде практически одинаковы. Меняется же в данном нервном волокне при определенных условиях просто число импульсов в секунду.
Приход импульса к окончанию аксона сказывается на следующем нейроне в цепи: изменяется его склонность генерировать импульсы. На пресинаптической мембране окончания выделяется медиатор - вещество, которое диффундирует через щель между двумя клетками и оказывает на постсинаптическую мембрану по ту сторону щели одно из двух воздействий. В возбудительном синапсе медиатор вызывает понижение постсинаптического мембранного потенциала, и в результате постсинаптическая клетка генерирует импульсы с большей частотой. В тормозном синапсе эффект медиатора состоит в стабилизации постсинаптического мембранного потенциала, из-за чего возбудительным синапсам труднее деполяризовать постсинаптическую клетку, и вследствие этого генерация импульсов либо прекращается, либо идет с меньшей частотой.
Является ли данный синапс возбудительным или тормозным, зависит от того, какой медиатор выделяет пресинаптическая клетка, и от химизма мембраны постсинаптической клетки. Почти к каждому нейрону приходят входы от множества окончаний, обычно от многих сотен, а иногда и тысяч, причем одни из них возбудительные, а другие тормозные. В каждый данный момент одни входы активны, а другие бездействуют, и от суммы возбудительных и тормозных эффектов зависит, будет ли клетка генерировать импульсы, и если будет, то с какой частотой. Иными словами, нейрон — это нечто гораздо большее, чем простой прибор для пересылки импульсов с одного места на другое. Каждый нейрон непрерывно оценивает все сигналы, приходящие к нему от других клеток, и выражает результаты этой оценки частотой своих собственных сигналов.
Таким образом, распространение обоих типов сигналов по мембране нейрона и химические процессы на уровне синапсов в общих чертах изучены. Однако далеко не ясна связь между формой нейрона (форма дерева с разветвленной кроной) и способом суммирования и оценки получаемых сигналов. Два входных сигнала, из которых каждый может быть возбудительным или тормозным, несомненно суммируются совсем по-разному, в зависимости от того, расположены ли синапсы по соседству (например, на одной и той же дендритной веточке) или один синапс находится на одной веточке, а другой далеко от него (возможно, на веточке другого дендрита) или же один синапс образуется на веточке, а другой на теле клетки. Форма, весьма различная у разных классов нейронов, должна играть важную роль в их функции, но пока это все, что можно сказать с уверенностью.
Близкая к этому группа вопросов касается значения некоторых синапсов (обычного вида синапсов с пресинаптическим и постсинаптическим компонентами), которые соединяют два дендрита или два аксона, а не как обычно — аксон с дендритом или с телом клетки. Мягко выражаясь, никто толком не знает, что о них думать. Наконец, положение еще больше осложняется тем, что некоторые синапсы совершенно отличны от обычного химического типа, так как в них не происходит диффузия медиатора, а течет электрический ток. Их открыли в 50-х годах Э. Фершпен (Е. Furshpan) и Д. Поттер (D. Potter) из Лондонского университетского колледжа. Почему в одних синапсах природа применяет передачу химическую, а в других - электрическую, пока что остается загадкой.
Если глубже рассматривать многие еще не получившие ответа вопросы относительно нервных сигналов, то они оказываются связанными с тонкой структурой и функцией нейронной мембраны, потому что на молекулярном уровне точно еще не известно, как ионы проходят через мембраны или как влияют изменения потенциала и медиаторы на проницаемость для отдельных ионов (см. Ч. Стивене «Нейрон»). Особый интерес представляет химизм синаптической передачи, при которой обнаружено уже больше 20 медиаторных веществ, а также те способы, какими нейроны синтезируют, выделяют, поглощают и разрушают эти уже достаточно хорошо известные вещества (см. Л. Иверсен «Химия мозга»).
Еще не полное, но в значительной мере уже достигнутое понимание химии синапсов нашло свое отражение в психиатрии и фармакологии. Многие заболевания - от паркинсонизма до депрессии, - по-видимому, связаны с нарушением синаптической передачи, и действие многих лекарств сводится к усилению или ослаблению этой передачи (см. ).
Лет через десять основные черты активности отдельных нейронов, надо полагать, будут детально изучены. Теперь же, когда положено известное начало пониманию структуры и работы отдельных клеток, нейробиологи находятся в положении человека, который, немного разбираясь в физических свойствах сопротивлений, конденсаторов и транзисторов, пытается заглянуть в телевизор. Он не поймет, как работает этот прибор в целом, пока не узнает, как соединены его элементы, и пока не получит хоть некоторое представление о назначении прибора, о его блоках и их взаимодействии. В изучении мозга первую ступень после нейрона и его работы составляет знакомство с тем, как взаимосвязаны более крупные части мозга и как устроена каждая из них. Следующей ступенью явится попытка установить, как нейроны взаимодействуют, и выяснить значение посылаемых ими сигналов.
Один из способов разобраться в общей организации мозга состоит в том, чтобы рассмотреть ее в форме приблизительной схемы. На входе лежат группы рецепторов — видоизмененные нервные клетки, специализированные для преобразования в электрические сигналы разных форм информации, которая приходит к ним из внешнего мира. Одни рецепторы реагируют на свет (зрение), другие на химические воздействия (вкус и обоняние), третьи на механическую деформацию (осязание и слух). Рецепторы образуют контакты с первой группой нейронов, те в свою очередь со следующими, и так далее. На каждом этапе аксоны ветвятся и подходят к следующему нейрону цепи, на котором конвергирует множество аксонов. Каждая воспринимающая клетка интегрирует возбудительные и тормозные импульсы, конвергирующие на ней от клеток низшего порядка. Раньше или позже, после ряда этапов нервные аксоны оканчиваются на клетках железы или мышцы — выходах нервной системы.
Коротко говоря, имеется вход — единственный для человека способ узнавать о внешнем мире. Имеется выход — единственный для человека способ реагировать на внешний мир и воздействовать на него. А между входом и выходом находится все остальное, что должно включать восприятие, эмоции, память, мысль и все прочее, что делает человека человеком.
Вряд ли нужно говорить, что описанный путь от входа к выходу очень упрощен. Хотя главное движение происходит на схеме слева направо, но на любом этапе между клетками часто существуют боковые связи; нередко имеются также связи, идущие в обратном направлении — от выхода к входу, совершенно так же, как имеется обратная связь во многих электронных цепях. От входа к выходу ведет не один только путь; множество разных наборов рецепторов специализированы для разных видов чувствительности и для определенных форм энергии, действующих на каждый орган чувств, и при этом имеются бесчисленные шунты, переключения и обходы. Синапсов между рецепторами и мышцами может быть или очень много, или всего лишь два или три. (Когда число их невелико, цепь обычно называют рефлексом; сужение зрачка в ответ на освещение сетчатки представляет собой рефлекс с участием четырех или пяти синапсов.) При этом следует снова указать, что синапс бывает или возбудительный или тормозный; если в определенный момент клетка испытывает оба этих воздействия, в результате они могут полностью погасить друг друга.
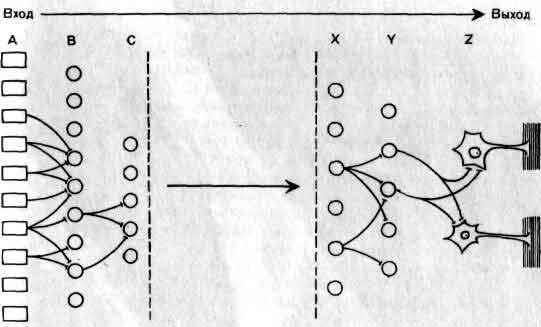
Общая организация головного мозга показана в виде примерной схемы, на которой изображен поток информации от сенсорных сигналов на входе рецепторных клеток (А) до конечного выхода мотонейронов (Z) на мышечные клетки. На своем выходе рецепторы и нейроны обычно ветвятся и посылают к следующему уровню дивергирующие сигналы. Большинство нейронов получает на входе от нижних уровней конвергирующие сигналы как возбудительные, так и тормозные. Кое-что известно о значении связей близ входа (В, С) и близ выхода (X, Y) головного мозга. Гораздо меньше известно о работе областей лежащих между ними, которые составляют большую часть мозга.
Теперь физиологи имеют некоторое представление о том, какого рода операции нервная система производит на входе и на выходе. На входе система, очевидно, занята главным образом извлечением из внешнего мира информации, которая представляет биологический интерес. Обычно рецепторы реагируют лучше всего на начало или на конец стимула, например давления на кожу. Нам важнее всего узнавать об изменениях; никому не хочется и не нужно, чтобы 16 часов в день ему напоминали, что на ногах у него обувь.
Если в качестве примера взять зрительную систему, то здесь важны контрасты и движения, и на первых двух-трех этапах значительная часть цепей занята усилением эффектов контраста и движения. На последующих уровнях зрительной системы поведение клеток становится сложнее, но оно всегда упорядочено и, к счастью, имеет смысл в применении к восприятию. В настоящее время путь информации, идущей от светочувствительных клеток сетчатки, прослежен в головном мозгу до шестого-седьмого этапа, до роли коры больших полушарий, связанной со зрением (см. ).
Хотя в настоящее время зрительная система является одной из лучше всего изученных частей мозга, нейробиологам еще далеко до понимания того, как происходит восприятие или опознание предметов. Правда, объем сведений, приобретенных за несколько лет, прошедших с момента создания микроэлектродов, позволяет думать, что такой отдел головного мозга, как кора больших полушарий, со временем будет доступен — во всяком случае в принципе — сравнительно простому объяснению.
На выходе нервной системы механизм, каким нейрон посылает импульс к мышечному волокну, в последнее время стал в основном понятен, и уже более 50 лет нам известны те части головного мозга, которые управляют движением. Однако, что именно делают в действительности эти структуры, когда человек движется или собирается двигаться, все еще во многом неизвестно. Изучение идет медленно главным образом потому, что исследовать произвольные движения надо у бодрствующих и хорошо обученных животных, тогда как изучать сенсорные системы можно у наркотизированных животных. Одно из основных усилий направлено сейчас на то, чтобы проследить за ходом моторного импульса от мотонейрона обратно к таким структурам, как моторная кора и мозжечок, с целью узнать, как на решение совершить движение влияют разные сигналы, идущие от входа нервной системы (см. ).
На выходе важно не сокращение отдельного волокна, а координированное сокращение и одновременное расслабление многих мышц; например, при сжимании кисти в кулак или схватывании предмета недостаточно просто согнуть пальцы, сократив сгибатели в предплечье, - надо также сократить разгибатели, чтобы мышцы-сгибатели пальцев не согнули кисть. Это противодействующее разгибание кисти происходит автоматически, без размышлений (это можно проверить, сжав руку в кулак и почувствовав при этом, как сокращаются мышцы-разгибатели на наружной стороне предплечья).
Очевидно, что команда сжать кисть в кулак, идущая от головного мозга, включает в себя импульсацию клеток, удаленных на несколько переключений от выхода нервной системы, — клеток, аксоны которых распределяются по разным мотонейронам и тормозным нейронам, снабжающим в свою очередь все мышцы, участвующие в выполнении команды. При других движениях действуют другие цепи, в которые могут быть включены те же мышцы, но в других комбинациях.
Таким образом, не трудно представить себе ряд такого рода функций, осуществляемых нервной системой, особенно близ ее сенсорного и моторного кондов. Недостает сведений главным образом относительно обширной промежуточной области — лобная и теменная доли, лимбическая система, мозжечок и так далее, и так далее, — хотя об их анатомии известно немало. В некоторых случаях известна своего рода физиология основной схемы связей, например установлено, какими нейронами возбуждаются или тормозятся другие нейроны. Для мозжечка не только описана в деталях схема связей, но и ясно, какие синапсы возбудительные, а какие тормозные; для некоторых видов синапсов с достаточной степенью надежности химически идентифицированы медиаторы. Но как работает мозжечок, известно только самым неопределенным образом. Он несомненно участвует в регуляции движений, мышечного тонуса, равновесия, но как такие функции выполняются этой великолепно построенной, упорядоченной и фантастически сложной машиной, совершенно неизвестно.
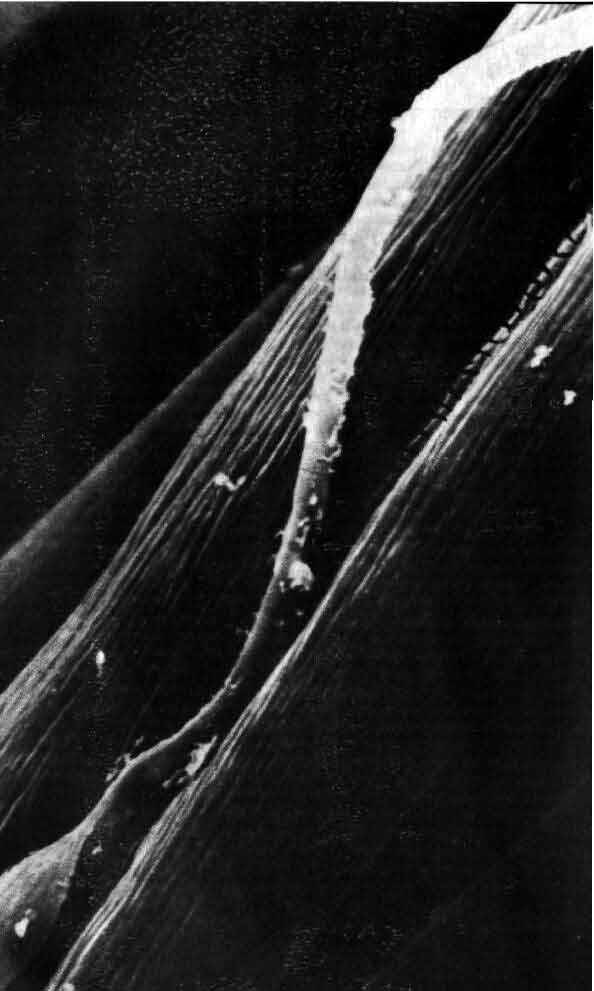
На выходе нервная система активирует мышечное волокно по концевым аксонным веточкам мотонейрона. Эта микрофотография получена с помощью сканирующего электронного микроскопа Б. Риз и Т. Ризом из Национального института инсульта, неврологических и коммуникативных нарушений. Одна концевая веточка лежит (в некоторых местах она отодвинута) в бороздке мышечного волокна. Нервное волокно в значительной части окружено оболочкой, образуемой шванновской клеткой, тело которой видно слева внизу. Бороздку пересекают узкие складки постсинаптической мышечной мембраны. Каждая складка примыкает к «активной зоне» нервного волокна; когда к синапсу приходит импульс из этой зоны через «окна» в оболочке, образованной шванновской клеткой, выделяется химический медиатор ацетилхолин. Молекулы ацетилхолина активируют рецепторы на мышечной мембране, вызывая таким образом мышечное сокращение.
По общему признанию, работать с мозжечком трудно; он образует водораздел между сенсорными и моторными процессами в совершенно незаполненной части приведенной на рисунке схемы. Виды сигналов, поступающих на его входы — специфическое значение импульсов, приходящих в мозжечок из коры больших полушарий, спинного мозга и так далее, — известны недостаточно; нейронные структуры, к которым направлены его выходы и которые в свою очередь связаны с мышцами, тоже мало изучены. По тем же причинам и другие части мозга в большинстве своем все еще мало понятны. Несмотря на методические успехи последних лет, крайне необходимы новые революционные методы. В качестве одного лишь примера можно указать, что сейчас не существует способа изучать сигналы отдельных клеток у человека, не вскрывая его черепа на операционном столе, последнее же совершенно недопустимо. Тем не менее достигнуты большие успехи в понимании некоторых высших функций человеческого мозга (см. Н. Гешвинд «Специализация человеческого мозга»), но чтобы действительно понять такую специально человеческую функцию, как речь, нужно разработать способы экстракраниальной регистрации активности отдельных нейронов.
Указать, как соединены между собой нейроны и как они работают в каждый данный момент — это лишь одна из конечных целей нейрофизиологии. Некоторые важные аспекты деятельности мозга лежат за этой гранью. Например, память и обучение безусловно представляют собой кумулятивные процессы, связанные с изменением во времени, а о механизмах, лежащих в их основе, известно еще очень мало.
Пожалуй, нейробиология особенно склонна к увлечениям, из-за которых порой она чуть не сходит с рельсов. Несколько лет тому назад возникла мысль, что следы памяти могут откладываться в форме крупных молекул, причем информация кодируется последовательностью мелких молекул подобно тому, как генетическая информация закодирована в ДНК. Мало кто, знакомый с высоко упорядоченной специфичностью связей в головном мозгу, принял эту идею всерьез, и тем не менее немало времени было потрачено во многих лабораториях на то, что животных обучали выполнению какой-либо задачи, затем растирали их мозг, а далее находили или химические отличия в их мозге, или «статистически значимое» усиление способности обучаться тем же задачам у животных, которым был введен экстракт мозга обученных животных. Увлечение это угасло, но факт тот, что не всегда нейробиология двигалась вперед или даже стояла на месте — временами она даже откатывалась назад.
В конечном счете изучение памяти, вероятно, будет касаться двух совсем разных ее компонентов. Первый компонент составляют изменения, которые вероятнее всего происходят в синапсах в результате многократного использования нейронных цепей. Например, возможно повышение эффективности одного синапса за счет других на той же клетке. Определенные комбинации стимулов, повторяясь, могут таким образом усиливать один из многих проводящих путей данной нейронной структуры.
Исследования такого плана трудно осуществимы на высших животных; их гораздо легче производить на малых системах нейронов, которые составляют полностью или частично нервную систему некоторых низших животных. В отдельные клетки этих животных удается легко ввести микроэлектрод, и, что еще важнее, такие клетки часто обладают индивидуальностью; говоря, например, о клетке №56 в определенном ганглии у рака, можно быть уверенным, что у всех других раков она занимает буквально то же положение и обладает теми же связями. (В этом глубокое различие между мозгом многих беспозвоночных и мозгом человека. Дать номер какому-нибудь нейрону в человеческом мозгу невозможно так же, как невозможно дать номер волосу на голове или поре на коже.)
Изящные опыты проводились на уровне отдельных нейронов у беспозвоночных при их обучении (см. ). Например, показано даже, что при выработке реакции или ее забывании у животного происходят доступные определению изменения в передаче сигналов через те или иные синапсы. Разумеется, обучение здесь простое, но, по-видимому, истинное. Снова и снова на беспозвоночных делались открытия, которые потом были распространены высшие формы. Поэтому маловероятно, что исследователи откажутся работать на таких животных, убоявшись насмешек начетчиков над невероятным звучанием таких тем, как «Решение задачи пиявкой».
Второй компонент, которым придется овладевать при изучении памяти, будет гораздо труднее. Все то, что человек помнит (в любом смысле этого слова, кроме самого рудиментарного), связано с восприятием или с движениями, или с жизненным опытом. Для проникновения в механизм памяти в любом истинном смысле нужно знать, что происходит, когда человек воспринимает, действует, думает и испытывает, для того чтобы понять, что из всего этого воспроизводится, когда он вспоминает что-то или чему-то научается. Из этих двух компонентов первый — синаптический — представляется мне сравнительно легким, а второй неимоверно трудным.
Понять работу мозга у взрослого животного — задача достаточно трудная; понять, как мозг становится мозгом, пожалуй, по меньшей мере так же трудно. Как развивается нервная система до и после рождения? Центральная задача состоит в том, чтобы раскрыть, как информация, закодированная в молекулах ДНК, трансформируется в связи между клетками внутри структур, в пространственные соотношения этих структур и связи между ними. Зрительный нерв, например, содержит около миллиона волокон, каждое из которых начинается в определенной точке маленького участка сетчатки. В свою очередь каждое волокно соединено в определенном порядке с плоским латеральным коленчатым телом в головном мозгу, и таким образом на коленчатом теле создается в известном смысле карта сетчатки. Каким образом во время развития волокна выходят из сетчатки, доходят до коленчатого тела и распределяются там с абсолютной топографической точностью? Такие же наборы связей, подобных проводам, смонтированные по своим особым схемам и соединяющие между собой топографически организованные области, имеются по всей нервной системе, а как собираются эти точные схемы, остается одной из важных неразрешенных проблем (см. ).
Изучение развития мозга потенциально важно не только потому, что оно проливает свет на работу мозга, но и потому, что о многих нервных болезнях известно точно, а о многих предположительно, что они возникают в период развития. Сюда относится большинство врожденных аномалий, синдром Дауна, некоторые виды мышечной дистрофии, вероятно, некоторые обычные формы эпилепсии и множество более редких заболеваний.
Остается только гадать о том, сколько пройдет времени, пока можно будет сказать, что мозг — или разум — в общем и целом понят (снова эти расплывчатые слова). Еще недавно, в 1950 г., того, кто предсказал бы, что через 10 лет процессы, лежащие в основе жизни, будут разгаданы, — сочли бы оптимистом, если не чудаком, а между тем, так и случилось. Думаю, что понадобится гораздо более 10 лет, чтобы понять мозг, просто потому, что он так многогранен, — это шкатулка, до краев наполненная остроумными решениями огромного множества задач. Весьма возможно, что человеческие существа никогда не разгадают всех тех отдельных задач, которые задает им мозг. Но можно надеяться, что по мере того, как будут рассмотрены одна за другой все области мозга, станет все яснее и яснее, что функции мозга упорядочены и доступны уразумению в понятиях физики и химии, без привлечения непознаваемых, сверхестественных процессов (см. ).
На этом пути будут отдельные главные вехи. Например, может быть раскрыт некий единый механизм, посредством которого работает память (ее синаптический компонент), или некий единый процесс, который объясняет, как в процессе развития нервные волокна находят места свoero назначения. Однако это не значит, что в будущем в какой-то момент будет сделано открытие или ряд открытий, в результате которых мозг станет совершенно понятным. Исследования мозга прогрессируют медленно. Методические усовершенствования нескольких последних десятилетий заметно ускорили этот процесс, но, безусловно, не привели К каким-либо внезапным переворотам, подобно тем, какие были совершены Коперником, Ньютоном, Дарвином, Эйнштейном или Уотсоном и Криком.
Каждая из таких революций отличалась тем, что переводила какой-нибудь очень важный раздел проводимых человеком исследований природы в область рационального и экспериментального анализа, уводя его от сверхестественного. Если Коперник показал, что Земля не является центром мироздания, а Галилей увидел в небе не ангелов, а звезды и планеты; если Дарвин установил, что человек состоит в родстве со всеми остальными живыми существами; если Эйнштейн ввел новые представления о времени и пространстве, о массе и энергии; если Уотсон и Крик показали, что биологическая наследственность объяснима в физических и химических понятиях, то наряду с этими открытиями, сужающими границы сверхъестественного, главное, что еще остается науке решать, — это, очевидно, проблема мозга и то, представляет ли он собой нечто большее, чем чрезвычайно сложную великолепную машину.
Этот вопрос затрагивает само существо человека, и поэтому принципиальные изменения в наших взглядах на человеческий мозг не могут не оказывать глубокого влияния на наши взгляды на нас самих и на окружающий мир. Разумеется, такие достижения произведут значительное воздействие и на другие области исследования. Будут до некоторой степени захвачены те разделы философии, которые занимаются природой мышления и восприятия, а также, я полагаю, некоторые части психологии, которые пытаются получить ответы на подобные вопросы косвенным путем. Если будут открыты механизмы обучения и памяти, это отразится на всей области воспитания.
В нейробиологии революция истинно коперниковских или дарвиновских масштабов во всяком случае не совершится одним ударом, возможно, никогда не совершится, а если произойдет, то постепенно, в течение многих десятилетий. С каждым ее этапом человеческие существа, несомненно, будут все ближе к пониманию самих себя.
Ч. СТИВЕНС
Нейрон
Это отдельная нервная клетка, строительный блок мозга. Она передает нервные импульсы по единственному длинному волокну (аксону) и получает их по многочисленным коротким волокнам (дендритам)
Нейроны, или нервные клетки, являются строительными блоками мозга. Хотя они имеют те же самые гены, то же самое общее строение и тот же самый биохимический аппарат, что и другие клетки, они обладают и уникальными особенностями, которые делают функцию мозга совершенно отличной от функции, скажем печени. Важными особенностями нейронов являются характерная форма, способность наружной мембраны генерировать нервные импульсы и наличие уникальной структуры, синапса, служащего для передачи информации от одного нейрона другому.
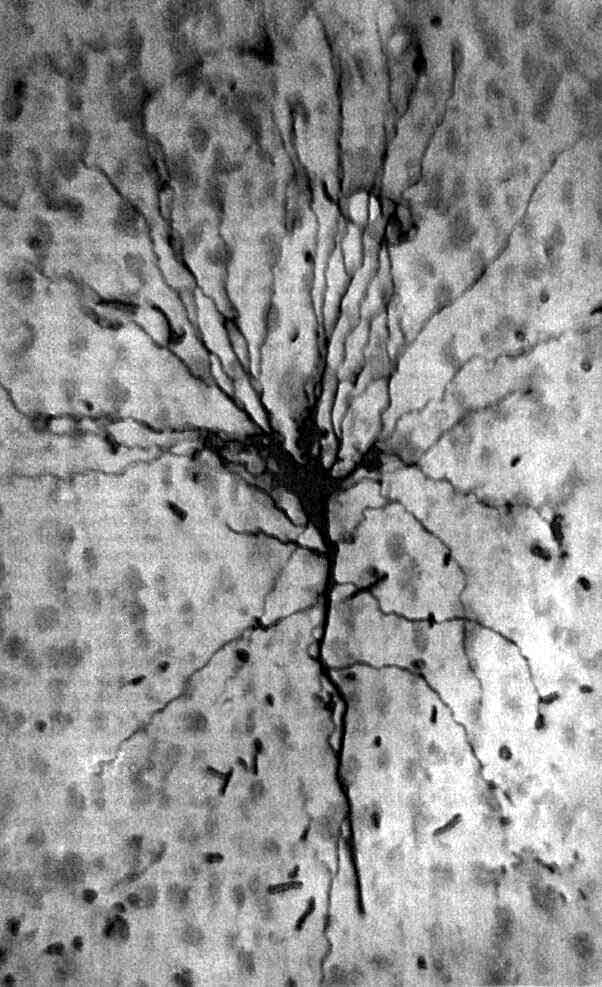
Нейрон зрительной коры кошки, представленный на микрофотографии, был окрашен путем инъекции фермента — пероксидазы хрена. Тела нейронов, образующие фон, докрашены фуксином. Все волокна, отходящие от тела клетки, являются дендритами, получающими информацию от других нейронов. Передающее информацию волокно, аксон, намного тоньше и его нелегко увидеть при таком увеличении. Самое толстое волокно, направленное вертикально вверх, называют апикальным дендритом; в данный срез попала лишь небольшая его часть. При данном увеличении полная длина апикального дендрита должна составлять около 75 см. (Его ход можно проследить по соседним срезам.) При регистрации активности этой клетки у живой кошки обнаружено, что она реагирует на границу светлое-темное, идущую под углом 60° к вертикали. Благодаря его форме этот нейрон называют пирамидной клеткой. Это один из двух наиболее многочисленных типов нейронов в коре млекопитающих. Микрофотография получена Ч. Джильбертом и Т. Визелем из Гарвардской медицинской школы.
Полагают, что мозг человека состоит из 1011 нейронов: это приблизительно столько же, сколько звезд в нашей Галактике. Не найдется и двух нейронов, одинаковых по виду. Несмотря на это, их формы обычно укладываются в небольшое число широких категорий, и большинству нейронов присущи определенные структурные особенности, позволяющие выделить три области клетки: клеточное тело, дендриты и аксон. Тело содержит ядро и биохимический аппарат синтеза ферментов и других молекул, необходимых для жизнедеятельности клетки. Обычно тело нейрона имеет приблизительно сферическую или пирамидальную форму. Дендриты представляют собой тонкие трубчатые выросты, которые многократно делятся и образуют ветвистое дерево вокруг тела клетки. Они создают ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксон тянется далеко от тела клетки и служит той линией связи, по которой сигналы, генерируемые в теле данной клетки, могут передаваться на большие расстояния в другие части мозга и остальной нервной системы. Аксон отличается от дендритов как по строению, так и по свойствам своей наружной мембраны. Большинство аксонов длиннее и тоньше дендритов и имеет отличный от них характер ветвления: если отростки дендритов в основном группируются вокруг клеточного тела, то отростки аксонов располагаются на конце волокна, в том месте, где аксон взаимодействует с другими нейронами.
Функционирование мозга связано с движением потоков информации по сложным цепям, состоящим из нейронных сетей. Информация передается от одной клетки к другой в специализированных местах контакта — синапсах. Типичный нейрон может иметь от 1000 до 10000 синапсов и получать информацию от 1000 других нейронов. Хотя в своем большинстве синапсы образуются между аксонами одной клетки и дендритами другой, существуют и иные типы синаптических контактов: между аксоном и аксоном, между дендритом и дендритом и между аксоном телом клетки.

Типичный нейрон позвоночного способен передавать нервные импульсы на значительные расстояния. У изображенного здесь нейрона все части увеличены пропорционально. Импульсы возникают в теле клетки и распространяются вдоль аксона, который имеет одну или несколько ветвей. Этот аксон, который для удобства изображен в виде гармошки, в действительности имеет в длину 1 см. Бывают аксоны длиной более 1 м. Конечные ветви аксона образуют синапсы более чем на 1000 других нейронов. Большинство синапсов связывает аксонные окончания одного нейрона с дендритами, образующими «дерево» вокруг клеточного тела другого нейрона. Таким образом, дендриты, окружающие нейрон на этой схеме, могли бы получать входные сигналы от десятков, сотен и даже тысяч других нейронов. Многие аксоны, такие, как этот, снабжены изолирующей миелиновой оболочкой, прерывающейся через определенные интервалы участками, называемыми перехватами Ранвье.
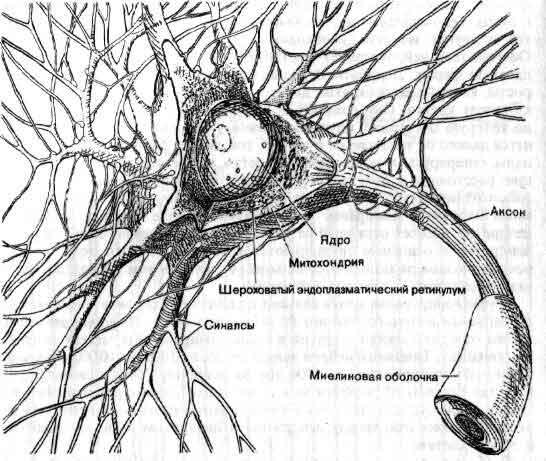
Тело нейрона содержит общий для всех клеток генетический материал и сложный метаболический аппарат. Однако в отличие от большинства других клеток нейроны после завершения эмбрионального периода не делятся; исходный их запас должен служить в течение всей жизни организма. От тела клетки отходит несколько дендритов и один аксон. Тело клетки и дендриты покрыты синапсами — бляшкообразными структурами, через которые поступает информация от других нейронов. Митохондрии снабжают клетку энергией. Белки синтезируются на эндоплазматическом ретикулуме. Транспортная система перемещает белки и другие вещества от тела клетки к тем местам, где они требуются.
В области синапса аксон обычно расширяется, образуя на конце пре-синаптическую бляшку, которая является передающей информацию поверхностью контакта. Концевая бляшка содержит мелкие сферические образования, называемые синаптическими пузырьками, каждый из которых содержит несколько тысяч молекул химического медиатора. По прибытии в пресинаптическое окончание нервного импульса некоторые из пузырьков выбрасывают свое содержимое в узкую щель, отделяющую бляшку от мембраны дендрита другой клетки, предназначенного для приема таких химических сигналов. Таким образом, информация передается от одного нейрона другому с помощью некоторого посредника, или медиатора. Импульсация нейрона отражает активацию воздействующими нейронами сотен синапсов. Некоторые синапсы являются возбуждающими, т. е. они способствуют генерации импульсов, тогда как другие — тормозные — способны аннулировать действие сигналов, которые в их отсутствие могли бы возбудить разряд нейрона.
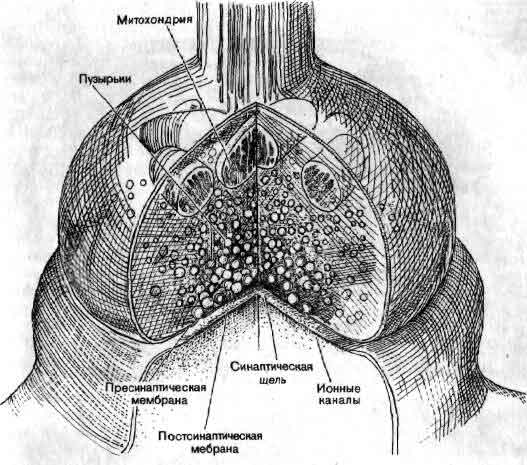
Синапс — это место переключения, в котором происходит передача информации от одного нейрона к другому с помощью химических медиаторов. Синапс состоит из двух частей: бляшкообразного утолщения, принадлежащего окончанию аксона, и рецепторной области на поверхности другого нейрона. Мембраны разделены синаптической щелью шириной около 200 нм. Молекулы химического медиатора, запасенные в пузырьках аксонного окончания, выделяются в щель под действием приходящих нервных импульсов. Медиатор изменяет электрическое состояние воспринимающего нейрона, увеличивая или уменьшая вероятность генерации этим нейроном импульса.
Хотя нейроны и являются строительными блоками мозга, это не единственные клетки, которые в нем имеются. Так, кислород и питательные вещества поставляются плотной сетью кровеносных сосудов. Существует потребность и в соединительной ткани, особенно на поверхности мозга. Один из важных классов клеток центральной нервной системы составляют глиальные клетки, или глия. Глия занимает в нервной системе практически все пространство, которое не занято самими нейронами. Хотя функция глии пока не вполне изучена, по-видимому, она обеспечивает структурную и метаболическую опору для сети нейронов.
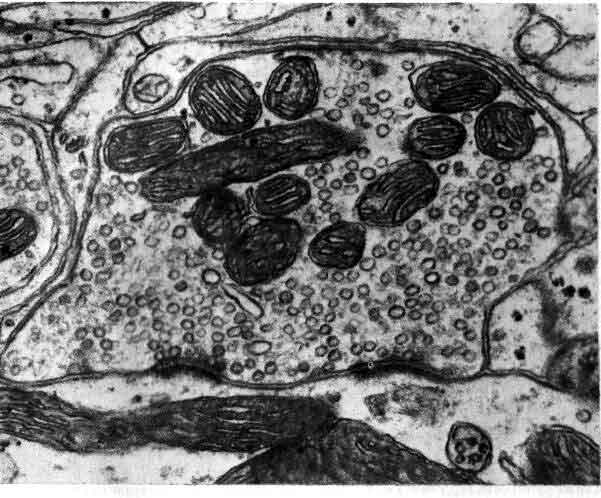
Синаптическое окончание занимает большую часть этой электронно-микроскопической фотографии, полученной Дж. Хойзером из Университета калифорнийской медицинской школы в Сан-Франциско и Т. Ризом из Национальных институтов здравоохранения. Щель, отделяющая пресинаптическую мембрану от постсинаптической, проходит вдоль нижней части фотографии. Крупные, темные структуры — это митохондрии; многочисленные округлые тела — пузырьки, содержащие медиатор; расплывчатые темные участки, расположенные вдоль щели, предположительно являются основными местами выделения медиатора.
Еще одним типом клеток, повсеместно встречающихся в нервной системе, являются шванновские клетки. Оказывается, все аксоны заключены в оболочку из шванновских клеток. В некоторых случаях шванновские клетки просто окутывают аксон тонким слоем. Во многих же случаях в ходе эмбриогенеза шванновская клетка закручивается вокруг аксона, образуя несколько плотных слоев изоляции, называемой миелином. Миелиновая оболочка прерывается примерно через каждый миллиметр по длине аксона узкими щелями — так называемыми перехватами Ранвье. В аксонах, имеющих оболочку такого типа, распространение нервного импульса происходит путем его перескакивания от перехвата к перехвату, где внеклеточная жидкость оказывается в непосредственном контакте с клеточной мембраной. Эволюционный смысл миелиновой оболочки, по-видимому, состоит в экономии метаболической энергии нейрона. Как правило, миелинизированные нервные волокна проводят нервные импульсы быстрее, чем немиелинизированные.
Нейроны способны выполнять свою функцию только благодаря тому, что их наружная мембрана обладает особыми свойствами. Мембрана аксона по всей его длине специализирована для проведения электрического импульса. Мембрана аксонных окончаний способна выделять медиатор, а мембрана дендритов реагирует на медиатор. Кроме того, мембрана обеспечивает узнавание других клеток в процессе эмбрионального развития, так что каждая клетка отыскивает предназначенное ей место в сети, состоящей из 1011 клеток. В связи с этим многие современные исследования сосредоточены на изучении всех тех свойств мембраны, которые ответственны за нервный импульс, за синаптическую передачу, за узнавание клеток и за установление контактов между клетками.
Мембрана нейрона, как и наружная мембрана любой клетки, имеет в толщину около 5 нм и состоит из двух слоев липидных молекул, упорядоченных таким образом, что их гидрофильные концы обращены в сторону водной фазы, находящейся внутри и снаружи клетки, а гидрофобные концы повернуты в сторону от водной фазы и образуют внутреннюю часть мембраны. Липидная часть мембраны приблизительно одинакова у клеток всех типов. Что делает одну мембрану отличной от другой, так это специфические белки, которые связаны с мембраной тем или иным способом. Белки, которые фактически встроены в двойной липидный слой, называются внутренними белками. Другие белки, периферические мембранные белки прикреплены к мембранной поверхности, но не являются неотъемлемой частью ее структуры. В связи с тем, что мембранные липиды — жидкости, даже внутренние белки часто могут свободно перемещаться с места на место путем диффузии. Однако в некоторых случаях белки жестко закрепляются с помощью вспомогательных структур.
Мембранные белки всех клеток распадаются на пять классов: насосы, каналы, рецепторы, ферменты и структурные белки. Насосы расходуют метаболическую энергию для перемещения ионов и молекул против концентрационных градиентов и поддерживают необходимые концентрации этих молекул в клетке. Поскольку заряженные молекулы не могут пройти через сам двойной липидный слой, клетки приобрели в процессе эволюции белковые каналы, обеспечивающие избирательные пути для диффузии специфических ионов. Клеточные мембраны должны узнавать и прикреплять многие типы молекул. Эти функции выполняют рецепторные белки, которые представляют собой центры связывания, обладающие высокой специфичностью и сродством. Ферменты размещаются внутри мембраны или на ней, чем облегчается протекание химических реакций у мембранной поверхности. Наконец, структурные белки обеспечивают соединение клеток в органы и поддержание субклеточной структуры. Эти пять классов мембранных белков не обязательно взаимно исключают друг друга. Так, например, тот или иной белок может быть одновременно и рецептором, и ферментом, и насосом. Мембранные белки — это ключ к пониманию функций нейрона, а следовательно, и функций мозга. Поскольку они занимают такое центральное место в современных представлениях о нейроне, я сконцентрирую свое обсуждение вокруг описания ионного насоса, различных типов каналов и ряда других белков, которые в совокупности наделяют нейроны их уникальными свойствами. Общая идея состоит в том, чтобы суммировать важные характеристики мембранных белков и показать, как эти характеристики определяют нервный импульс и другие сложные особенности функций нейрона.
Подобно всем другим клеткам нейрон способен поддерживать постоянство своей внутренней среды, заметно отличающейся по составу от окружающей его жидкости. Особенно поразительны различия в концентрациях ионов натрия и калия. Наружная среда приблизительно в 10 раз богаче натрием, чем внутренняя, а внутренняя среда примерно в 10 раз богаче калием, чем наружная. Как калий, так и натрий способны проникать через поры в клеточной мембране, поэтому некоторый насос должен непрерывно производить обмен вошедших в клетку ионов натрия на ионы калия из наружной среды. Такое выкачивание натрия осуществляется внутренним мембранным белком, называемым Na-K-аденозинтрифосфатазным насосом, или, как его чаще называют, натриевым насосом.
Белковая молекула натриевого насоса (или комплекс белковых субъединиц) имеет молекулярный вес около 275000 и размеры порядка 6x8 нанометров, что несколько больше толщины клеточной мембраны. Каждый натриевый насос может использовать энергию, запасенную в форме фосфатной связи в аденозинтрифосфате (АТФ), для того, чтобы обменять три иона натрия внутренней среды клетки на два иона калия наружной среды. Работая с максимальной скоростью, каждый насос способен транспортировать через мембрану около 200 ионов натрия и 130 ионов калия в секунду. Однако фактическая скорость регулируется в соответствии с потребностями клетки. У большинства нейронов имеется от 100 до 200 натриевых насосов на квадратный микрон мембранной поверхности, но в некоторых участках этой поверхности их плотность почти в 10 раз выше. Типичный мелкий нейрон имеет, по-видимому порядка миллиона натриевых насосов, способных перемещать около 200 миллионов ионов натрия в секунду. Именно трансмембранные градиенты натрия и калия обеспечивают возможность проведения по нейрону нервного импульса.
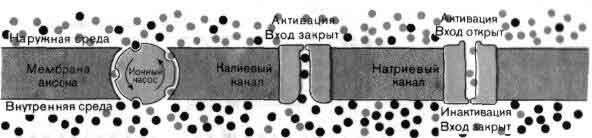
Мембрана аксона разделяет жидкости, сильно отличающиеся по содержанию ионов натрия (цветные кружки) и ионов калия (черные кружки). Наружная среда приблизительно в 10 раз богаче ионами натрия, чем калия; во внутриклеточной среде соотношение ионов обратное. Мембрана пронизана белками, которые действуют как избирательные каналы, предпочтительно пропускающие либо ионы натрия, либо ионы калия. В состоянии покоя, когда импульсы не передаются, каналы обоих типов закрыты, и ионный нанос поддерживает ионные градиенты, выкачивая ионы натрия в обмен на ионы калия. Внутренность аксона в норме имеет отрицательный потенциал в 70 мВ по отношению к наружному раствору. Если эта разность потенциалов уменьшается вследствие прихода нервного импульса, натриевый канал открывается и позволяет ионам натрия входить внутрь аксона. Мгновением позже натриевый канал закрывается, а открывается калиевый, позволяя ионам калия выходить из клетки. Последовательное открывание и закрывание каналов двух типов приводит к распространению нервного импульса; этот процесс представлен на следующей иллюстрации.
Мембранные белки, которые служат каналами, существенны для многих сторон деятельности нейрона и в особенности для генерации нервного импульса и синаптической передачи. Чтобы представить значение каналов для электрической активности мозга, я коротко опишу механизм нервного импульса, а затем опять вернусь к более систематическому описанию свойств каналов.
Поскольку концентрации ионов натрия и калия по ту и другую сторону мембраны различаются, внутренность аксона имеет отрицательный потенциал примерно в 70 мВ по отношению к наружной среде. Четверть века тому назад в своих классических работах по изучению передачи нервного импульса вдоль гигантского аксона кальмара английские исследователи А. Ходжкин, А. Хаксли и Б. Катц показали, что распространение нервного импульса сопровождается резкими изменениями проницаемости мембраны аксона для ионов натрия и калия. Когда нервный импульс возникает в основании аксона (в большинстве случаев он генерируется клеточным телом в ответ на активацию дендритных синапсов), трансмембранная разность потенциалов в этом месте локально понижается. Непосредственно впереди области с измененным потенциалом (по направлению распространения нервного импульса) открываются мембранные каналы, пропускающие в клетку ионы натрия.
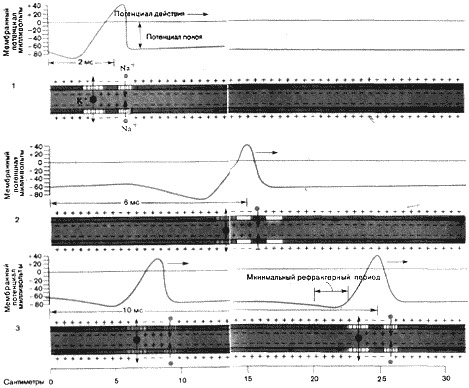
Распространение нервного импульса по аксону сопряжено с появлением локальных потоков ионов натрия (Na + ) внутрь, сменяемых потоками ионов калия (К+) наружу через каналы, которые регулируются изменениями напряжения на мембране аксона. Электрический процесс, приводящий к распространению нервного импульса вдоль аксона, обычно развивается в клеточном теле. Генерация импульса начинается со слабой деполяризации, или уменьшения отрицательного потенциала внутренней поверхности мембраны, в том месте, где аксон отходит от клеточного тела. Этот небольшой сдвиг потенциала открывает некоторые из натриевых каналов, вызывая тем самым дальнейшее уменьшение потенциала.
Поток ионов натрия внутрь будет ускоряться до тех пор, пока внутренняя поверхность мембраны не станет локально положительной. Изменение знака потенциала приведет к закрыванию натриевых каналов и открыванию калиевых. Поток ионов калия наружу быстро восстановит отрицательный потенциал. Кратковременная реверсия потенциала, получившая название потенциала действия, сама распространяется по аксону (1, 2). После короткого рефрактерного периода за первым импульсом может следовать второй (3). Скорость распространения нервного импульса на схеме соответствует таковой в гигантском аксоне кальмара.
Этот процесс является самоусиливающимся: поток ионов натрия через мембрану способствует открыванию большего числа каналов и облегчает другим ионам возможность следовать за ними. Проникшие в клетку ионы натрия изменяют отрицательный внутренний потенциал мембраны на положительный. Вскоре после открывания натриевые каналы закрываются, но теперь открывается другая группа каналов, которая позволяет ионам калия выходить наружу. Этот поток восстанавливает потенциал внутри аксона до величины его потенциала покоя, т.е. до —70 мВ. Резкий скачок потенциала сначала в положительную, а затем в отрицательную сторону, который выглядит на экране осциллографа как пик («спайк»), известен под названием потенциала действия и является электрическим выражением нервного импульса. Волна изменения потенциала стремительно проносится по аксону до самого его конца во многом подобно тому, как бежит пламя по бикфордову шнуру.
Это краткое описание нервного импульса иллюстрирует важность каналов для электрической активности нейронов и подчеркивает два фундаментальных свойства каналов: избирательность и наличие воротных механизмов. Каналы проницаемы избирательно, и степень избирательности варьирует в широких пределах. Так, каналы одного типа позволяют проходить ионам натрия, но сильно препятствуют прохождению ионов калия, тогда как каналы другого типа делают обратное. Однако избирательность редко бывает абсолютной. Канал одного типа, который практически не обладает избирательностью, позволяет проходить примерно 85 ионам натрия на каждые 100 ионов калия; другой канал, с большей избирательностью, пропускает только около 7 ионов натрия на каждые 100 ионов калия. Канал первого типа, известный как активируемый ацетилхолином, имеет пору диаметром около 0,8 нм, которая заполнена водой. У канала второго типа, известного как калиевый канал, пора значительно меньше и содержит меньше воды.
Ион натрия приблизительно на 30% меньше иона калия. Точная молекулярная структура, позволяющая более крупным ионам проходить через клеточную мембрану легче, чем более мелким, неизвестна. Однако общие принципы, лежащие в основе такой дискриминации, понятны. Они включают взаимодействия между ионами и участками канальной структуры, сочетающиеся со специфическим упорядочением молекул воды внутри поры.
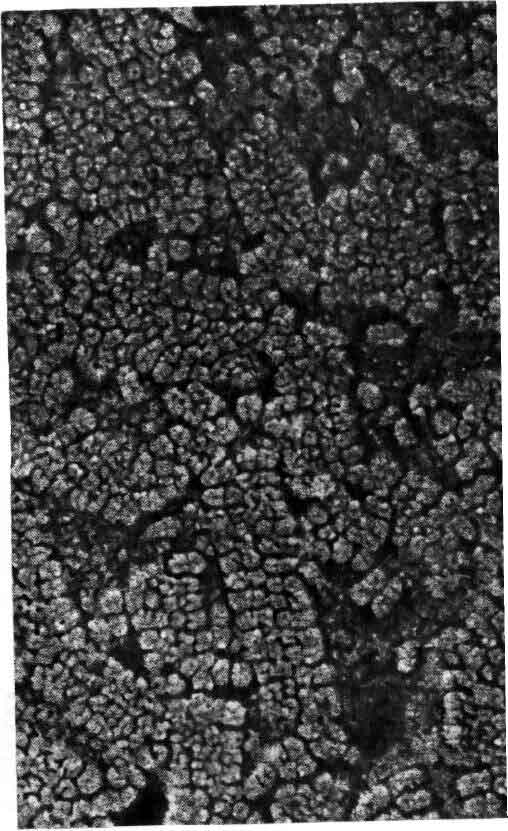
Активируемые ацетилхолином каналы плотно упакованы в постсинаптической мембране клетки электрического органа ската — рыбы, которая может наносить электрический удар. На этой микрофотографии показана покрытая платиной реплика мембраны, которая была заморожена и вытравлена. Размеры частиц платины не позволяют разрешать детали мельче 2 нм. Согласно последним данным, белковая молекула канала, размеры которой составляют 8,5 нм в поперечнике, состоит из пяти субъединиц, окружающих канал, наименьший размер которого составляет 0,8 нм. Микрофотография получена Хойзером и С. Салпетером (Heuser, S. Salpeter).
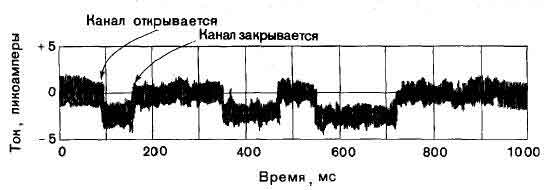
Ответ одиночного мембранного канала на медиатор ацетилхолин был зарегистрирован с помощью созданного недавно метода, который применили Э. Нехер и Дж. Стейнбах (медицинский факультет Йельского университета). Активируемые ацетилхолином каналы, имеющиеся в постсинаптических мембранах, пропускают приблизительно равные количества ионов натрия и калия. На записи показан ток через одиночный канал постсинаптической мембраны мышцы лягушки, возникающий при активации этого канала субэрилдихолином — веществом, имитирующим действие ацетилхолина, но открывающим каналы на более длительное время. Эксперимент показал, что процесс открывания каналов подчиняется закону «все или ничего» и время их пребывания в открытом состоянии варьирует случайным образом.
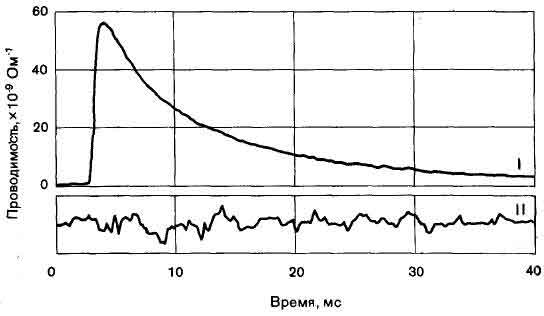
Натриевые каналы аксона также работают по принципу «все или ничего» и при этом независимо друг от друга, что было установлено исследованиями, проведенными Ф. Сигуорсом (медицинский факультет Йельского университета). В немиелинизированной области мембраны аксона, названной перехватом Ранвье, во время распространения нервного импульса обычно открывается около 10000 каналов, I-изменения проницаемости для натрия во времени; II-получена при 12-кратном усилении по сравнению с верхней; показаны флуктуации проницаемости вокруг среднего значения, обусловленные вероятностным характером процессов открывания и закрывания каналов.
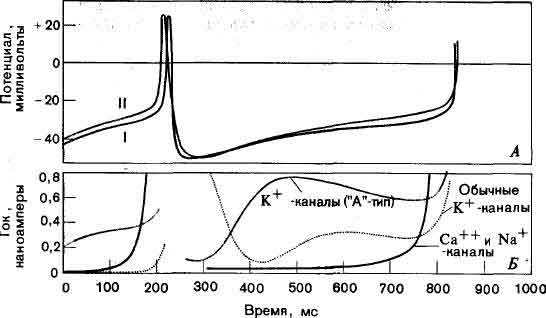
Развитие нервных импульсов в телах нейронов требует координированного открывания и закрывания каналов пяти типов, пропускающих разные виды ионов (натрия, калия или кальция). Вклад различных каналов в нервный импульс можно оценить, решая систему нелинейных дифференциальных уравнений. А. Зависимость от времени фактически зарегистрированных (I) и вычисленных на основании уравнений (II) изменений потенциала внутри тела нейрона. Б. Изменения во времени всех токов, протекающих через основные типы каналов. Для возникновения серии нервных импульсов необходимо сложное взаимодействие каналов разных типов. Исследования, на основании которых построены данные кривые, были проведены Дж. Коннором в Иллинойском университете и автором статьи на медицинском факультете Йельского университета.
Воротные механизмы, регулирующие открывание и закрывание мембранных каналов, представлены двумя основными типами. Канал одного типа, упоминавшийся выше при описании нервного импульса, открывается и закрывается в ответ на изменения потенциала клеточной мембраны, поэтому говорят, что он управляется электрически. Второй тип каналов управляется химически. Такие каналы реагируют лишь слабо, если вообще реагируют, на изменения потенциала, но открываются, когда особая молекула — медиатор — связывается с некоторой рецептор ной областью на белке канала. Химически управляемые каналы обнаружены в рецептивной мембране синапсов: они ответственны за перевод химических сигналов, посылаемых окончаниями аксона в процессе синаптической передачи, в изменения ионной проницаемости. Химически управляемые каналы обычно именуют в соответствии с их специфическим медиатором. Так, например, говорят об АХ-активируемых каналах или о ГАМК-активируемых каналах (АХ — ацетилхолин, ГАМК — гамма-аминомасляная кислота). Электрически управляемые каналы принято называть по иону, наиболее легко проходящему через данный канал.
Функционируя, белки обычно изменяют свою форму. Такие изменения формы, называемые конформационными, особенно ярко выражены у сократимых белков, ответственных за движение клеток, но они не менее важны и для многих ферментов и других белков. Конформационные изменения канальных белков составляют основу воротных механизмов, поскольку они обеспечивают открывание и закрывание канала за счет малых перемещений частей молекулы, расположенных в критическом месте и позволяющих блокировать или освобождать пору.
Когда электрически или химически управляемые каналы открываются и пропускают ионы, возникает электрический ток, который можно измерить. Совсем недавно в нескольких случаях удалось зарегистрировать ток, проходящий через одиночный канал, так что его открывание и закрывание можно было исследовать непосредственно. Обнаружилось, что время, на протяжении которого канал остается открытым, варьирует случайным образом, так как открывание и закрывание канала есть результат некоторых конформационных изменений белковой молекулы, встроенной в мембрану. Наличие случайности в воротных процессах проистекает из случайных столкновений молекул воды и других молекул со структурными элементами канала.
Кроме ионных насосов и каналов для выполнения основных функций нервной системы нейронам требуются и другие мембранные белки.
Одним из таких необходимых белков является фермент аденилатциклаза, который регулирует внутриклеточную концентрацию циклического аденозинмонофосфата (циклического АМФ). Циклические нуклеотиды, такие, как циклический АМФ, существенны для ряда клеточных функций, механизмы которых в деталях еще не изучены. Мембранный фермент аденилатциклаза, по-видимому, состоит из двух основных субъединиц — каталитической и регуляторной. Каталитическая субъединица способствует образованию циклического АМФ. Различные регуляторные субъединицы, которые, как полагают, физически обособлены от каталитических, могут связывать специфические молекулы (включая медиаторы, открывающие и закрывающие каналы) и тем самым контролировать содержание циклического АМФ в клетке. Регуляторные субъединицы разных типов называются в соответствии с теми молекулами, которые в физиологических условиях с ними связываются; одна из них, например, названа серотонинактивируемой аденилатциклазой. Известно, что аденилатциклаза и родственные ей мембранные ферменты выполняют в нейронах ряд регуляторных функций, и точный механизм их действия является сейчас объектом интенсивного исследования.
В процессе эмбриогенеза нервной системы клетка должна уметь узнавать другие клетки, чтобы рост каждой из них происходил в «правильном» направлении и заканчивался образованием «правильных» связей. Процесс узнавания клетки клеткой и формирования на основе этого соответствующей структуры определяется мембранными белками специального класса, связанными с особыми углеводами. Изучение белково-углеводных комплексов, ответственных за узнавание клеток, находится пока на ранней стадии.
Внутренние мембранные белки, о которых я здесь рассказываю, не распределяются по всей клеточной мембране однородно и не присутствуют в равных количествах во всех нейронах. Плотность и тип белка определяются потребностями клетки и различны для разных нейронов и для разных частей одного и того же нейрона. Так, плотность каналов определенного типа варьирует от 0 до 10000 на квадратный микрон. Аксоны обычно не имеют химически управляемых каналов, тогда как в постсинаптических мембранах дендритов плотность таких каналов лимитируется лишь упаковкой канальных молекул. В то же время мембраны дендритов обычно имеют мало электрически управляемых каналов, тогда как в мембранах аксонов их плотность может доходить в некоторых местах до 1000 каналов на квадратный микрон.
Внутренние мембранные белки синтезируются первоначально в теле нейрона и хранятся в мембране в небольших пузырьках. Для перемещения таких пузырьков от места их синтеза к месту их функционирования нейроны имеют специальную транспортную систему. Эта система, по-видимому, перемещает пузырьки небольшими скачками с помощью сократимых белков. Достигнув места своего назначения, белки встраиваются в поверхностную мембрану, где и функционируют до тех пор, пока не будут удалены оттуда и не распадутся внутри клетки. Точно не известно, каким образом клетки решают, куда какой мембранный белок поместить. Также неизвестен и механизм, который регулирует синтез, встраивание и разрушение мембранных белков. Метаболизм мембранных белков составляет одну из центральных проблем биологии клетки.
Каким образом свойства различных мембранных белков, которые я здесь обсуждаю, связаны с функцией нейрона? Чтобы ответить на этот вопрос, вернемся к рассмотрению нервного импульса и проанализируем более детально те молекулярные процессы, которые лежат в основе его генерации и распространения. Как мы видели, внутренность нейрона имеет отрицательный потенциал в 70 мВ относительно наружной среды. Этот «потенциал покоя» является следствием ионных градиентов, создаваемых натриевым насосом, и присутствием в клеточной мембране некоторого класса постоянно открытых каналов, избирательно проницаемых для ионов калия. Насос выталкивает наружу ионы натрия, обменивая их на ионы калия, и делает внутриклеточную среду в 10 раз богаче ионами калия по сравнению с наружной средой. Калиевые каналы мембраны позволяют ионам калия, находящимся в непосредственной близости от мембраны, выходить из клетки совершенно свободно. В состоянии покоя проницаемость мембраны для ионов натрия низка, так что не существует почти никакого встречного потока ионов натрия из внешней среды во внутреннюю, несмотря на то что внешняя среда в 10 раз богаче ионами натрия, чем внутренняя. В связи с этим поток калия создает дефицит положительных зарядов на внутренней поверхности клеточной мембраны и избыток положительных зарядов на ее наружной поверхности. В результате возникает разность потенциалов в 70 мВ, причем внутренность клетки имеет отрицательный потенциал по отношению к наружной среде.
Распространение нервного импульса определяется присутствием в мембране нейрона электрически управляемых натриевых каналов, открывание и закрывание которых ответственно за потенциал действия. Каковы характеристики этих важных канальных молекул? Хотя с химической точки зрения натриевый канал еще не достаточно хорошо изучен, известно, что он является белком с молекулярным весом в диапазоне от 250000 до 300000. Диаметр поры этого канала составляет 0,4-0,6 нм; через такую пору могут проходить ионы натрия, связанные с молекулами воды. На поверхности канала имеется много заряженных групп, размещенных в критических точках. Эти заряды обусловливают наличие большого электрического дипольного момента, который меняется по направлению и по величине в соответствии с конформационными изменениями канала, сопровождающими переход из закрытого состояния в открытое.
Поскольку поверхностная мембрана клетки очень тонка, трансмембранная разность потенциалов в 70 мВ создает внутри покоящейся мембраны сильное электрическое поле порядка 100 кВ/см. Подобно тому как магнитные диполи имеют тенденцию ориентироваться вдоль силовых линий магнитного поля, электрические диполи белка натриевого канала стремятся встать параллельно линиям электрического поля мембраны. Изменения напряженности электрического поля могут переводить канал из закрытого состояния в открытое. По мере того как под влиянием входящих ионов натрия внутренняя поверхность мембраны становится все более положительной, натриевые каналы все дольше находятся в открытом состоянии. Процесс открывания натриевых каналов под влиянием изменения потенциала мембраны называют активацией натриевых каналов.
Этот процесс останавливается благодаря развитию другого процесса, названного натриевой инактивацией. Трансмембранная разность потенциалов, явившаяся причиной открывания натриевых каналов, затем переводит их в особое закрытое конформационное состояние, отличное от состояния, характерного для канала в покое. Второе закрытое состояние, названное состоянием инактивации, развивается медленнее, чем процесс активации, так что до того, как каналы закроются под влиянием инактивации, они остаются короткое время открытыми. В состоянии инактивации каналы пребывают несколько миллисекунд, а затем возвращаются в нормальное состояние покоя.
Полный цикл активации и инактивации в норме включает в себя открывание и закрывание тысяч натриевых каналов. Как можно узнать, с чем связано увеличение общей мембранной проницаемости: с открыванием и закрыванием некоторого числа каналов по закону «все или ничего» или с работой каналов, у каждого из которых проницаемость может меняться градуально? Частичный ответ на этот вопрос был получен с помощью новой методики, которая соотносит флуктуации мембранной проницаемости с вероятностным характером конформационных изменений канальных белков. Можно много раз вызывать открывание канала и вычислить среднюю проницаемость за какое-то время, а также точные ее значения в каждом испытании. Флуктуации точных значений проницаемости относительно среднего значения укладываются в 10% или около того. Анализ этих флуктуации показывает, что натриевые каналы работают по закону «все или ничего» и что открывание каждого канала увеличивает проводимость мембраны на 8-10-12 Ом-1. Одним из принципиальных моментов для понимания работы нейрона является необходимость развития сколько-нибудь полной теории, которая опишет поведение натриевых каналов и свяжет его с молекулярной структурой канального белка.
Как я уже кратко отметил выше, аксоны также имеют электрически управляемые калиевые каналы, которые помогают прекращать нервный импульс, позволяя ионам калия выходить из аксона, противодействуя тем самым входящему потоку ионов натрия. В теле нейрона ситуация еще более сложная, поскольку мембрана там пронизана каналами пяти типов. Различные каналы открываются с различными скоростями, остаются открытыми на протяжении разных интервалов времени и являются избирательно проницаемыми для разных ионов (натрия, калия и кальция).
Наличие в теле нейрона каналов пяти типов (в аксоне их только два) приводит к более сложным законам генерации нервных импульсов. Если на аксон подается некоторый постоянный стимул, аксон генерирует только одиночный импульс в ответ на начало стимуляции. Тело же клетки генерирует в таком случае целый ряд импульсов, частота которых определяется интенсивностью стимула.
Нейроны способны генерировать нервные импульсы в широком диапазоне частот: от одного или менее до нескольких сотен в секунду. Все нервные импульсы имеют одну и ту же амплитуду, так что информация, которую они несут, представлена числом импульсов, генерируемых в единицу времени: такой способ кодирования известен под названием частотного кодирования. Чем больше величина сигнала, который должен быть передан, тем выше частота разряда.
Когда нервный импульс проходит по всей длине аксона и прибывает к его окончанию, из пресинаптической мембраны высвобождается один из видов медиаторов. Этот медиатор диффундирует к постсинаптической мембране, где индуцирует открывание химически управляемых каналов. Ионы, проходящие через открытые каналы, вызывают изменения потенциала, известные под названием синаптических потенциалов.
Большая часть того, что известно о синаптических механизмах, получена в экспериментах на определенном синапсе: нервно-мышечном соединении, управляющем сокращением мышц лягушки. Аксон мотонейрона лягушки проходит на протяжении нескольких сотен микрон вдоль поверхности мышечной клетки, образуя несколько сотен синаптических контактов на расстояниях порядка микрона друг от друга. В каждой пресинаптической области легко обнаружить характерные синаптические пузырьки.
В синаптическом пузырьке содержится около 10000 молекул медиатора — ацетилхолина. Когда нервный импульс достигает синапса, запускается цепь событий, кульминацией которых являются слияние пузырька с пресинаптической мембраной и происходящее благодаря этому высвобождение ацетилхолина в щель между пресинаптической и постсинаптической мембранами; этот процесс называют экзоцитозом. Слившийся с мембраной пузырек в дальнейшем отделяется от нее и быстро вновь наполняется ацетилхолином, чтобы затем вновь опорожниться.
В последнее время были вскрыты многие детали событий, приводящих к экзоцитозу. Выяснилось, что слияние пузырьков с пресинаптической мембраной, по всей видимости, запускается быстрым, но кратковременным увеличением концентрации кальция в окончании аксона. Прибытие в окончание нервного импульса приводит к открыванию химически управляемых кальциевых каналов и появлению потока кальция внутрь окончания. Однако наблюдающееся вследствие этого увеличение концентрации кальция бывает лишь кратковременным, поскольку в окончании содержится специальный механизм, который быстро устраняет свободный кальций и восстанавливает его концентрацию до нормального, очень низкого уровня. Кратковременный резкий подъем уровня свободного кальция приводит к слиянию заполненных медиатором пузырьков с пресинаптической мембраной, но точный механизм этого важного процесса еще не совсем понятен.
Интересные детали структуры пресинаптической мембраны были выявлены методом криоскалывания, который позволяет разъединить слои двуслойной мембраны и делает внутренние мембранные белки доступными для исследования методом электронной микроскопии. В нервно-мышечном соединении лягушки на ширине каждого синапса тянется двойной ряд крупных мембранных белков. К этим белкам или вблизи них прикрепляются синаптические пузырьки. Только такие пузырьки могут сливаться с мембраной и выделять медиатор; другие пузырьки, по-видимому, остаются в резерве на некотором расстоянии от мембраны. Слияние пузырька с мембраной является случайным процессом, и каждый пузырек ведет себя при этом независимо от других.
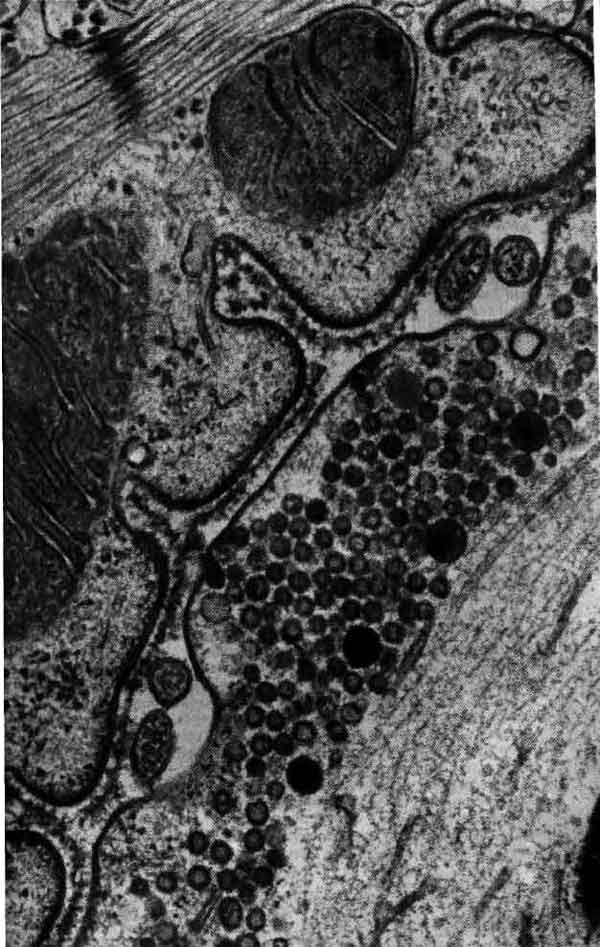
Нервно-мышечное соединение лягушки; электронная микрофотография получена Хойзером. Аксон отделяется от мышечной клетки синаптической щелью. Синаптические пузырьки группируются вдоль пресинаптической мембраны; ближе к центру видны два синаптических контакта. Постсинаптическая мембрана мышечной клетки имеет одну особенность, не обнаруживаемую в других синапсах: против каждого контакта мембрана образует складки.
Реплики пресинаптической мембраны нервно-мышечного соединения лягушки после криоскалывания. А. Состояние мембраны через 3 мс после раздражения мышцы. Через мембрану аксона тянется двойной ряд частиц — мембранных белков, которые могут быть либо кальциевыми каналами, либо структурными белками, присоединяющими к себе пузырьки. Б. Состояние мембраны через 5 мс после стимуляции. Стимуляция привела к слиянию синаптических пузырьков с пресинаптической мембраной и образованию углублений.
Менее чем за 100 микросекунд ацетилхолин высвобождается из слившишхся с мембраной пузырьков, пересекает синаптическую щель и связывается с ацетилхолиновым рецептором — внутренним мембранным белком, встроенным в постсинаптическую мембрану. Рецептор одновременно является канальным белком, химически управляемым ацетилхолином. Когда к каналу прикрепляются две молекулы ацетилхолина, они снижают энергетический уровень молекулы белка в конформации, соответствующей открытому состоянию, и тем самым увеличивают вероятность того, что канал будет открыт. Переход канала в открытое состояние является случайным событием; среднее время пребывания в этом состоянии около миллисекунды. Каждый пакет из 10000 ацеталхолиновых молекул приводит к открыванию примерно 2000 каналов.
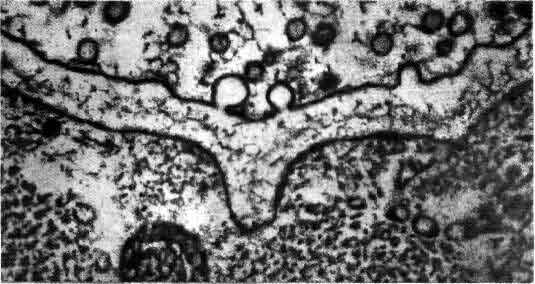
Медиатор выделяется в щель синаптического контакта между нейронами из пузырьков, которые сливаются с пресинаптической мембраной аксона и раскрываются; этот процесс назван экзоцитозом. На данной электронной микрофотографии нервно-мышечного синапса лягушки пузырьки аксонного окончания запечатлены в момент высвобождения ацетилхолина; микрофотография получена Хойзером.
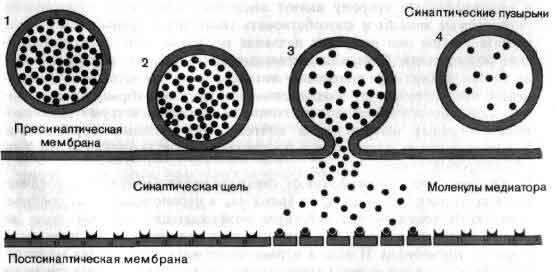
Синаптические пузырьки группируются вблизи пресинаптической мембраны. На схеме показаны предположительные стадии экзоцитоза. Заполненные пузырьки движутся к синаптической щели, сливаются с мембраной, выделяют содержимое, а затем вновь отделяются от мембраны, восстанавливают свою форму и заполняются медиатором.
За тот короткий период, в течение которого канал остается открытым, через него проходит около 20000 ионов натрия и приблизительно столько же ионов калия. В результате этих ионных потоков трансмембранная разность потенциалов уменьшается почти до нуля. Насколько близко она подходит к нулю, зависит от того, как много каналов было открыто и как долго они оставались открытыми. Ацетилхолин, высвобожденный типичным нервным импульсом, приводит к возникновению постсинаптического потенциала, или изменению напряжения, длящегося всего около пяти миллисекунд. Поскольку постсинаптические потенциалы обусловлены работой каналов, управляемых химически, а не электрически, их параметры сильно отличаются от параметров нервного импульса. Они обычно меньше по амплитуде, имеют большую длительность и могут плавно меняться по величине в зависимости от количества выделенного медиатора и, следовательно, от числа открытых каналов.
Различные типы химически управляемых каналов демонстрируют разную избирательность. Некоторые из них сходны с ацетилхолиновым каналом, пропускающим ионы натрия и калия почти без предпочтения. Другие каналы высоко избирательны. Изменение потенциала, возникающее на данном синапсе, зависит от избирательности открывающихся каналов. Если в клетку входят положительные ионы, происходит изменение потенциала в положительном направлении. Сдвиги потенциала в положительную сторону имеют тенденцию открывать электрически управляемые каналы и способствовать генерации нервных импульсов; в связи с этим они получили название возбуждающих постсинаптических потенциалов. Если положительные ионы (обычно калий) выходят из клетки, происходит изменение потенциала в отрицательном направлении, что способствует закрыванию электрически управляемых каналов. Такие постсинаптические потенциалы противодействуют возникновению нервных импульсов, и поэтому они названы тормозными. И возбуждающие, и тормозные постсинаптические потенциалы обычны для нейронов мозга.
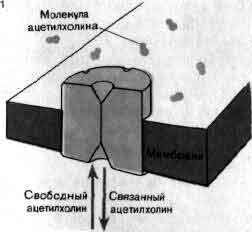
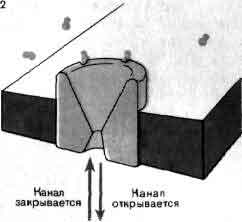
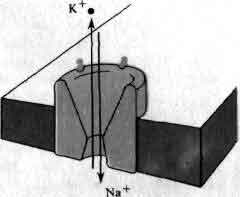
Ацетилхолиновый канал в постсинаптической мембране открывается молекулами ацетилхолина, выделяемыми в синаптическую щель. Рисунок изображает ацетилхолиновый рецептор в нервно-мышечном соединении лягушки. Две молекулы ацетилхолина быстро связываются с закрытым каналом в покое и формируют комплекс из рецептора и ацетилхолина (1, 2). Этот комплекс претерпевает конформационные изменения, в результате которых канал открывается для прохождения ионов натрия и калия (3). Время, необходимое для конформационных изменений комплекса, лимитирует скорость реакции. Канал остается открытым в среднем около 1 мс и затем вновь превращается в рецептор-ацетилхолиновый комплекс. Пока канал открыт, через него проходит около 20000 ионов натрия и равное количество ионов калия. Ацетилхолин быстро отделяется и разрушается ферментом ацетилхолинэстеразой.
Синапсы мозга отличаются от синапсов нервно-мышечного соединения в нескольких аспектах. В то время как в нервно-мышечных соединениях ацетилхолин всегда действует возбуждающе, действие того же самого вещества в мозгу в одних синапсах является возбуждающим, а в других тормозным. И если в нервно-мышечных соединениях медиатором обычно служит именно ацетилхолин, то каналы мозговых синапсов регулируются самыми различными медиаторами. Однако каждое данное синаптическое окончание выделяет только один тип медиатора, и в соответствующей постсинаптической мембране имеются каналы, управляемые этим медиатором. В противоположность активируемым ацетилхолином каналам нервно-мышечного синапса, всегда открывающимся примерно на одну миллисекунду, в некоторых типах мозговых синапсов имеются каналы, открывающиеся на доли миллисекунды, а в некоторых других каналы могут оставаться открытыми сотни миллисекунд. Последнее большое различие состоит в том, что в области нервно-мышечного соединения лягушки аксон образует сотни синаптических контактов с мышечной клеткой, а в мозгу аксоны обычно устанавливают только один-два синаптических контакта с данным нейроном. Как и можно было ожидать, такие различия в функциональных свойствах коррелируют со значительными различиями в структуре.
Как мы видели, интенсивность стимула кодируется частотой импульсов. В синапсе декодирование производится с помощью двух процессов: временной суммации и пространственной суммации. В процессе временной суммации каждый постсинаптический потенциал добавляется к суммарному потенциалу предшественников, вызывая таким образом изменение потенциала, средняя амплитуда которого отражает частоту поступающих импульсов. Другими словами, нейрон, который разряжается с высокой частотой, выделяет из своих синаптических окончаний больше молекул медиатора, чем нейрон, разряжающийся с меньшей частотой. А чем больше молекул медиатора выделяется за данное время, тем больше каналов открывается в постсинаптической мембране и, следовательно, тем выше постсинаптический потенциал. Пространственная суммация — это в некотором смысле эквивалентный процесс, только он отражает интеграцию нервных импульсов, прибывающих от всех нейронов, находящихся в синаптическом контакте с данным нейроном. Итоговое изменение потенциала, получившееся в результате временной и пространственной суммации, кодируется частотой нервных импульсов для передачи в другие клетки, расположенные в нейронной сети «ниже по течению».
Я описал здесь то, что обычно понимают под нормальной передачей информации в нервных сетях, при которой изменения постсинаптического потенциала кодируются частотой нервных импульсов и передаются по аксону другим нервным клеткам. Однако в последние годы было обнаружено, что в некоторых случаях постсинаптический потенциал не трансформируется в нервный импульс. Так, изменение напряжения, связанное с синаптическим потенциалом, может непосредственно вызвать выделение медиатора из соседней зоны без возникновения импульса. Полагают, что такое непосредственное воздействие может играть роль в синаптических контактах между дендритами, а также в некоторых цепях с обратной связью, где один дендрит вступает как пресинаптический в контакт с другим дендритом, а тот в свою очередь, тоже как пресинаптический, вступает в контакт с первым. Такие короткие цепи обратной связи, по-видимому, обычны для мозга, но их роль в переработке информации пока еще не выяснена.
Большинство современных исследований нейрона сосредоточено на мембранных белках, наделяющих двухслойную клеточную мембрану, которая сама по себе лишена каких-нибудь особенностей, специфическими свойствами, определяющими функционирование мозга. Что касается канальных белков, то имеется еще много нерешенных вопросов, касающихся механизма открывания, избирательности и регуляции. В последующие 5—10 лет, возможно, удастся связать физические процессы открывания и избирательность с молекулярной структурой каналов. Основы регуляции каналов меньше всего изучены, но сейчас начинают интенсивно исследоваться. Как теперь становится понятно, в регуляции каналов определенную роль могут играть гормоны и другие вещества. К центральным проблемам синаптического взаимодействия нужно отнести экзоцитоз и другие процессы, связанные с метаболизмом и выделением медиаторов. Можно ожидать, что усиленное внимание будет сконцентрировано на роли поверхностной мембраны в процессе роста и развития нейронов и образования синаптических связей, т.е. на тех удивительных процессах, которые закладывают фундамент интеграции нервной системы.
Э. КЭНДЕЛ
Малые системы нейронов
Такие системы представляют собой элементарные единицы мозговой деятельности. Изучение простых животных, например крупного брюхоногого моллюска аплизии, показывает, что Малые системы нейронов способны к некоторым формам обучения и памяти
По убеждению многих нейробиологов в конце концов будет доказано, что уникальные свойства каждого человека — способность чувствовать, думать, обучаться и помнить — заключены в строго организованных сетях синаптических взаимосвязей между нейронами головного мозга. Поскольку в человеческом мозгу исследовать эти сети трудно, важная задача нейробиологии состояла в том, чтобы создать на животных модели, пригодные для изучения того, как взаимодействующие системы нейронов формируют поведение. Нейронные сети, осуществляющие завершенные поведенческие акты, позволяют исследовать иерархию взаимосвязанных вопросов. В какой мере варьируют свойства разных нейронов? Чем определяется организация взаимосвязей между нейронами? Как разная организация взаимосвязей создает разные формы поведения? Может ли обучение модифицировать взаимосвязанные нейроны, управляющие определенным видом поведения, а если может, то посредством каких механизмов происходит запоминание?
Среди многих функций, осуществляемых благодаря взаимодействию нейронов, самые интересные те, которые связаны с обучением (способностью изменять поведение под влиянием опыта) и с памятью (способностью сохранять эти изменения в течение некоторого времени). Обучение и память — это, пожалуй, наиболее отличительные черты умственной деятельности высших животных, достигающие наивысшей формы у человека. В самом деле, человек является тем, чем он есть, в значительной мере благодаря тому, чему он обучился. Поэтому чтобы понять обучение и изучить эволюцию поведения, теоретически важно определить, на каком филогенетическом уровне нейронной и поведенческой организации можно распознать начальные проявления процессов обучения и памяти, характерных для человеческого поведения. Такое определение важно также для практики. Исследовать клеточные механизмы памяти в мозгу человека или других млекопитающих трудно потому, что у них мозг невероятно сложен. Кроме того, таким исследованиям на мозге человека препятствуют этические соображения. Поэтому для науки важно было бы эффективно исследовать эти процессы в простых системах.
Группа нейронов на микрофотографии дорсальной поверхности абдоминального ганглия морского моллюска аплизии. Справа виден особенно крупный более темный нейрон. На карте абдоминального ганглия аплизии эта клетка идентифицирована как нейрон R2.
Могут возразить, что нельзя успешно изучать память и обучение человека на простых нейронных системах. Организация человеческого мозга представляется столь сложной, что попытка изучить обучение человека в упрощенной форме на простых системах обречена на неудачу. Человек обладает интеллектом, весьма развитой речью и абстрактным мышлением, которых нет у низших животных и которые, возможно, требуют качественно иных типов нейронной организации. Хотя такие возражения существенны, решающим не является вопрос о том, есть ли нечто специфическое в человеческом мозгу. Несомненно, есть. Но вопрос скорее состоит в том, есть ли у человеческого мозга и человеческого поведения что-нибудь общее с мозгом и поведением низших животных. Там, где есть элементы сходства, они могут свидетельствовать об общих принципах организации мозга, которые доступны успешному изучению в простых нервных системах.
Ответ на вопрос о сходстве очевиден. Этологи К. Лоренц (К. Lorenz), H. Тинберген (N. Tinbergen) и К. Фриш (К. von Frisch) показали, что у людей с низшими животными много общих форм поведения, в том числе элементарное восприятие и координация движений. В особенности широко распространена способность к обучению; она развилась у многих беспозвоночных и у всех позвоночных. Сходство некоторых процессов обучения позволяет думать, что нейронные механизмы данного процесса могут обладать общими свойствами на всем протяжении филогенеза. Например, нет, по-видимому, принципиальной разницы в структуре, химизме или функции между нейронами и синапсами у человека, кальмара, улитки и пиявки. Следовательно, полный и тщательный анализ обучения у такого беспозвоночного, вероятно, способен выявить механизмы, имеющие общее значение.
Примитивные беспозвоночные привлекательны для таких исследований тем, что их нервная система содержит от 10000 до 100000 клеток в отличие от многих миллиардов у более сложных животных. Клетки собраны в отдельные группы, называемые ганглиями, и каждый ганглий обычно содержит от 500 до 1500 нейронов. Такое количественное упрощение позволило связать функцию отдельных нейронов непосредственно с поведением. В результате получен ряд важных фактов, которые приводят к новым представлениям о связи между мозгом и поведением.
Первый важный вопрос, который следует рассмотреть исследователям простых нейронных систем, состоит в том, отличны ли друг от друга разные нейроны одной области. Этот вопрос — центральный для понимания того, как поведение осуществляется нервной системой, был до последнего времени предметом обсуждения. Некоторые нейробиологи считали, что по своим свойствам нейроны мозга достаточно сходны, чтобы можно было считать их идентичными элементами, связи между которыми обладают приблизительно одинаковым весом.
Против этого теперь выдвигаются веские возражения, особенно на основании изучения беспозвоночных, показавшего, что многие нейроны доступны индивидуальной идентификации и инвариантны у каждого члена вида. Представление об индивидуальных свойствах нейронов предложено еще в 1912 г. немецким биологом Р. Гольдшмидтом (R. Goldschmidt) на основании исследования нервной системы у примитивного червя, кишечного паразита аскариды. Мозг этого червя состоит из нескольких ганглиев. Изучая эти ганглии, Гольдшмидт обнаружил, что они содержат точно 162 клетки. Число это никогда не варьировало от животного к животному, и каждая клетка всегда занимала свое характерное положение. Несмотря на такие четкие результаты, работа Гольдшмидта осталась практически незамеченной.
Более 50 лет спустя к этой проблеме вернулись независимо друг от друга две группы исследователей в Гарвардской медицинской школе. М. Оцука (М. Otsuka), Э. Кравиц (Е. Kravitz) и Д. Поттер (D. Potter), исследуя омара, и У. Фрезье (W. Frazier), И. Купферман (I. Kupfermann), Р. Вазири (R. Waziri) и Р. Коггсхолл (R. Coggeshall), исследуя крупного морского моллюска аплизию, нашли подобную же, хотя и менее полную инвариантность в более сложных нервных системах этих более развитых беспозвоночных. Сходная инвариантность вскоре была обнаружена у многих беспозвоночных, в том числе у пиявки, рака, саранчи, сверчка и ряда моллюсков. Я ограничусь здесь рассказом об исследованиях аплизии, преимущественно исследованиях отдельного ганглия, а именно абдоминального. Сходные данные получены также на других беспозвоночных.
В абдоминальном ганглии аплизии нейроны варьируют по величине, положению, форме, пигментации, по характеру импульсации и химическим веществам, посредством которых они передают информацию другим клеткам. Такие различия позволяют распознавать и называть индивидуальные клетки (Rl, L1, R151 и т.д.). Некоторые из этих различий проявляются в характере импульсации. Одни клетки обычно «молчат», другие спонтанно активны. Среди активных одни генерируют регулярные потенциалы действия, или нервные импульсы, другие выдают повторные краткие залпы или серии импульсов. Теперь известно, что различия в импульсации объясняются разными типами ионных токов, генерируемых мембраной тела нейронов. Мембрана тела нервной клетки совершенно отлична от мембраны ее длинного отростка, аксона. Когда мембрана аксона активна, она, как правило, создает только приток ионов натрия и отставленный по времени выход ионов калия, тогда как мембрана тела клетки может продуцировать шесть или семь разных ионных токов в различных комбинациях.
1 R, Right — правый; L, Left — левый.— Прим. ред.
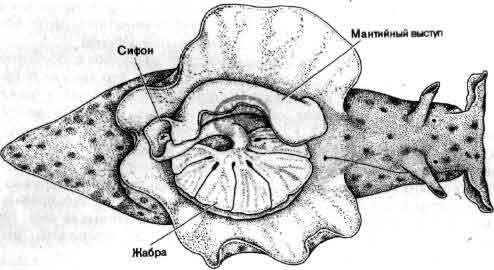
Рефлекс втягивания жабры возникает у аплизии в ответ на стимуляцию сифона или мантийного выступа. При этом жабра принимает положение, обозначенное цветной линией.
Пока еще неизвестно, обладает ли большинство клеток в нервной системе млекопитающих такой индивидуальностью. Впрочем, исследования сенсорных систем млекопитающих, описанные Д. Хьюбелом и Т. Визелем в настоящем выпуске, выявили поразительные важные различия между соседними нейронами (см. ). Работы по изучению развития головного мозга у позвоночных, описанные У. Коуэном, приводят к такому же выводу (см. ),
В связи с тем фактом, что нейроны инвариантны, возникают новые вопросы. Инвариантны ли также синаптические связи между клетками? Всегда ли данная идентифицированная клетка соединена точно с тем же следующим нейроном, а не с другими? Многие физиологи изучали эти вопросы на беспозвоночных и нашли, что действительно клетки всегда образуют одни и те же виды соединений с другими клетками. Инвариантны не только связи, но и «знак», или функциональное выражение, этих связей, т.е. их тормозное или возбудительное действие.
Поэтому в, дальнейшем Фрезье, Дж. Бленкеншип (J. Blankenship), Г. Вахтель (Н. Wachtel) и я пользовались идентифицированными клетками, чтобы выявить правила, определяющие функциональные свойства связей между клетками. Отдельный нейрон обладает множеством веточек и образует множество соединений. Мы задались рядом вопросов: все ли связи нейрона специализированы для торможения или возбуждения, или же его импульсация может производить разное действие в его разных ветвях? Чем определяется возбудительный или тормозный эффект связи? В чем причина того, что связь является возбудительной или тормозной? Определяется ли знак синаптического действия химической структурой медиатора, выделяемого пресинаптическим нейроном, или же определяющим фактором служит природа постсинаптического рецептора? Выделяет ли нейрон на всех своих окончаниях один и тот же медиатор?
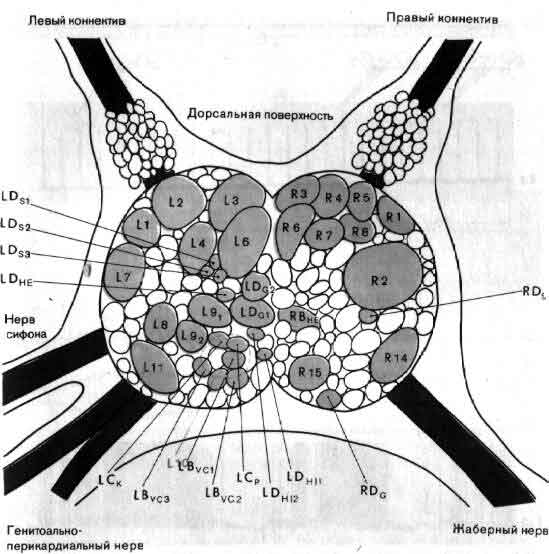
Карта абдоминального ганглия Aplysia californica, на которой показано положение идентифицированных нейронов; они обозначены L или R (от Left - левая и Right — правая половины ганглия) и пронумерованы. Эти нейроны являются частями скопления, образованного клетками с одинаковыми свойствами; они, кроме того, помечены буквой, обозначающей скопление (LD), и индексом, указывающим на поведенческую функцию нейрона, например, НЕ для возбудителя сердца (от Heart excitator) и G1 и G2 для двух мотонейронов жабры (от Gill — жабра).
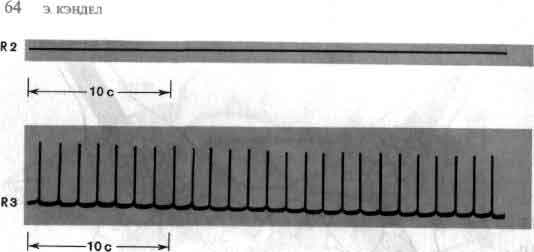


Характер импульсации идентифицированных нейронов в абдоминальном ганглии аплизии. R2 обычно «молчит», R3 выдает регулярную импульсацию, R15 — регулярные пачки импульсов и L10 — нерегулярные пачки. L10 является командной клеткой, управляющей остальными клетками в системе.
Один из путей исследования этих вопросов состоит в том, чтобы проследить за разными связями клетки. Первая же рассмотренная нами клетка дала ясный ответ: по разным своим связям она оказывала разное действие. Она возбуждала одни следующие за ней клетки, тормозила другие и (пожалуй, совершенно неожиданно) образовывала двойственную связь, как возбудительную, так и тормозную, с третьей клеткой. Кроме того, она всегда возбуждала точно одни и те же клетки, всегда тормозила другую определенную группу клеток и всегда имела двойственную связь с третьей группой. Ее синаптическое действие создавалось одним и тем же медиатором ацетилхолином. Будет ли это действие возбудительным или тормозным, зависело от реакции медиатора с разными типами рецепторов на постсинаптических клетках. Рецепторы определяли знак синаптического действия, управляя разными ионными каналами в мембране, в основном натриевыми, для возбуждения и хлорными для торможения. Клетки, к которым приходили связи двойного действия, обладали для одного и того же медиатора рецепторами двух типов: один управлял натриевым каналом, другой — хлорным. Таким образом, функциональное свойство химической синаптической передачи определяется типом рецептора, который находится на данном постсинаптическом участке следующей клетки. [Сходные результаты получила Ж. Кихоу в Эколь Нормаль в Париже, детально проанализировав свойства разных ацетилхолиновых рецепторов.]
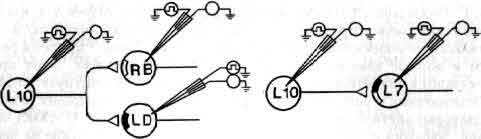
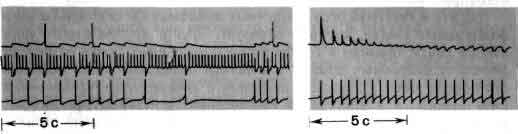

Инвариантность связей между клеткой L10 и некоторыми из следующих за ней клеток. А. Опыт, в котором двуствольные микроэлектроды для регистрации и для проведения тока были введены в L10, пресинаптический нейрон, и три следующие за ним клетки: L10 вызывает возбуждение (обозначено белым) в RB, торможение (черным) в LD и возбуждение и торможение в L7. Б. Соответствующие записи импульсации. В. Несколько наложенных записей, на которых виден короткий, но постоянный латентный период между импульсом в пресинаптическом нейроне и ответом двух следующих за ним клеток. Г. Наложения записей, полученных от L10 и L7, показывают, что эффект бывает возбудительным, когда L10 действует первым, как показывают высокие и узкие импульсы, и тормозным, когда он действует вторым, как показывают низкие и широкие импульсы.
Следовательно, как и предположили в свое время Л. Тауц (L. Tauc) и Г. Гершенфельд (Н. Gerschenfeld) из Института Марея в Париже, химический медиатор является лишь разрешающим агентом, а директивным компонентом синаптической передачи служат природа рецептора и ионные каналы, с которыми он взаимодействует. Этот принцип оказался в значительной мере универсальным. Он действует в нейронах позвоночных и беспозвоночных и в нейронах, использующих разные медиаторы: ацетилхолин, гамма-аминомасляную кислоту (ГАМК), серотонин, дофамин и гистамин. (Ему подчиняется также действие на нейроны некоторых пептидных гормонов, о чем пойдет речь ниже.)
То, что в ганглиях беспозвоночных животных были открыты идентифицируемые клетки, которые образуют друг с другом строго определенные связи, привело к составлению «монтажных схем» разных поведенческих нейронных цепей и тем самым позволило точно исследовать причинную связь отдельных нейронов с поведением. Термин «поведение» относится к доступным наблюдению действиям организма, начиная от таких сложных актов, как речь или ходьба, и до таких простых, как движение части тела или изменение ритма сердца. К типам поведения, хотя бы отчасти исследованным у пиявок, раков и брюхоногих моллюсков, относятся питание, разные локомоторные действия и различные реакции избегания и защиты.
Первый факт, полученный в этих исследованиях, состоит в том, что отдельные клетки осуществляют над поведением специфичный и часто поразительно мощный контроль. Это можно проиллюстрировать сравнением нервного контроля работы сердца у аплизии и у человека.
Человеческое сердце бьется спонтанно. Присущий ему ритм модулируется тормозным действием холинергических нейронов (медиатором служит ацетилхолин), аксоны которых идут в составе блуждающего нерва, и возбуждающим действием норадренергических нейронов, аксоны которых образуют ускоряющий нерв. В модуляции участвует несколько тысяч нейронов. У аплизии сердце тоже бьется спонтанно; его ритм модулируется тормозным действием холинергических нейронов и возбуждающим действием серотонинергических нейронов, но в модуляции участвуют всего лишь четыре нейрона! Две клетки возбуждают сердце (по существу, важную роль играет только «главная возбудительная» клетка), и две клетки тормозят его. Три другие клетки вызывают сокращение кровеносных сосудов и тем самым регулируют кровяное давление. Поскольку отдельные клетки неизменно связаны с одними и теми же следующими за ними клетками и могут производить эффекты разного знака, некоторые из них, расположенные в критических точках нервной системы, способны управлять целой последовательностью поведенческих актов. Еще в 1938 г. К. Вирсма (С. Wiersma) из Калифорнийского технологического института в опытах на раках установил важное значение отдельных клеток для поведения и назвал их «командными клетками». Такие нейроны найдены у многих животных. Некоторые из них оказались нейронами двойного действия. Впоследствии Дж. Кестер (J. Koester), И. Мейери (Е. Mayeri) и я, работая на аплизии в Медицинской школе Ньюйоркского университета, установили, что описанный нейрон двойного действия является командной клеткой для нейронной цепи, управляющей кровообращением. Одна эта клетка учащает ритм сердца и увеличивает объем выталкиваемой им крови, возбуждая главную клетку, возбуждающую сердце, и одновременно тормозя клетки, тормозящие сердце, а также клетки, вызывающие сужение крупных кровеносных сосудов. В результате усиленной активности одного этого нейрона сердце бьется быстрее и накачивает больше крови.
Таков лишь один простой пример поведенческих функций командной клетки. У рака и даже у более сложного животного — золотой рыбки — один импульс одного командного нейрона заставляет животное спасаться от грозящей ему опасности. Недавно В. Маунткасл (V. Моuntcastle) из Медицинской школы Университета Дж. Гопкинса высказал в этой связи предположение, что в головном мозгу приматов небольшие группы клеток, возможно, выполняют подобные командные функции по управлению произвольными движениями.
Следовательно, функциональное назначение клеток двойного действия состоит в том, что они вызывают совокупность разных физиологических эффектов. Подобная же совокупность действий может быть произведена нейроэндокринными клетками — нейронами, которые выделяют гормоны (химические вещества, обычно переносимые кровотоком для действия на больших расстояниях). Абдоминальный ганглий аплизии содержит два скопления нейроэндокринных клеток, которые называются пазушными клетками. Купферман (Kupfermann), работающий в нашем отделе Медико-хирургического колледжа Колумбийского университета, Ст. Арч (St. Arch) из Колледжа Рида, а также Ф. Штрумвассер (F. Strumwasser) с сотрудниками из Калифорнийского технологического института показали, что пазушные клетки выделяют полипептидный гормон, регулирующий яйцекладку. Мейери нашел, что этот гормон оказывает длительное действие на разные клетки абдоминального ганглия, возбуждая одни и тормозя другие.
Одной из клеток, возбуждаемых этим гормоном, является командная клетка двойного действия, управляющая ритмом сердца. При этом ритм ускоряется и приток крови к тканям усиливается, что необходимо во время яйцекладки. Таким образом, на точное расположение связей, которые служат взаимодействию на близких расстояниях, накладывается столь же точно организованное взаимодействие на больших расстояниях, которое осуществляется гормонами, выделяемыми нейроэндокринными клетками. Строго определенный эффект каждого гормона, по-видимому, зависит, как и синаптические эффекты, от природы рецепторов на клетках-мишенях.
На основании того факта, что поведение управляется инвариантными клетками, связанными друг с другом точно и инвариантно, можно было бы думать, что простые животные отличаются от более сложных стереотипными и фиксированными репертуарами активности. Но это не так. Изучение разных беспозвоночных показало, что поведение простых животных вполне способно видоизменяться посредством обучения.
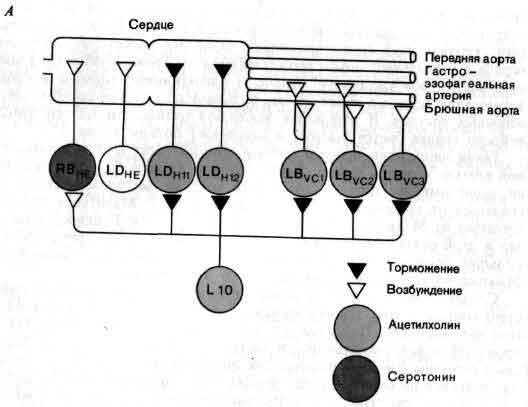
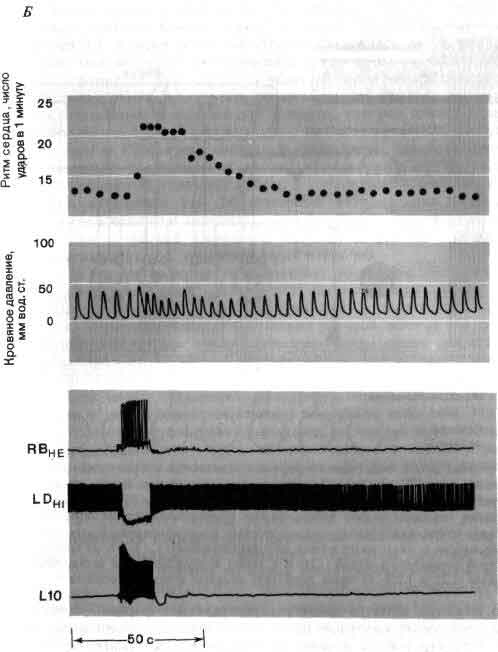
Регуляция поведения аплизии, осуществляемая одним нейроном (L10) и выражающаяся в его воздействии на сердечно-сосудистые мотонейроны. Известно, что L10 образует синаптические связи (А) с шестью клетками (наличие такой синаптической связи у LDHE еще не исследовано); цвет каждой клетки показывает, какой медиатор она использует. Можно видеть (Б), что активность L10, возбуждая RBHE и тормозя LDHI, ускоряет сокращение сердца и повышает кровяное давление.
Мы исследовали этот вопрос весьма детально на примере одного из самых простых видов поведения аплизии — защитном рефлекторном акте, при котором стимул вызывает втягивание жабры. Жабра находится в мантийной полости. Полость покрыта мантийным выступом который заканчивается мясистым желобом — сифоном. При слабой или умеренной стимуляции сифона жабра сокращается и втягивается в мантийную полость. Этот рефлекс аналогичен реакциям избавления, присущим почти всем высшим животным, например отдергиванию руки от горячего предмета у человека. У аплизии и других животных при таких рефлексах наблюдаются две формы обучения: привыкание (габитуация) и сенситизация.
Привыкание означает ослабление поведенческой реакции при многократном повторении стимула, который вначале был новым. Когда на животное действует новый стимул, оно сначала отвечает комбинацией ориентировочного и защитного рефлексов. При повторении сигнала животное быстро обучается узнавать его. Если за ним не следует награда или он оказывается безвредным, животное ослабляет и в конце концов подавляет свои реакции на него. Хотя привыкание удивительно просто, оно, пожалуй, является самой распространенной формой обучения. Путем привыкания животные, включая человека, научаются игнорировать стимулы, утратившие новизну или значение. Привыкание освобождает их для реакций на стимулы, которые влекут за собой награду или имеют значение для выживания. Считается, что привыкание является первым процессом обучения, возникающим у детей; оно обычно используется для изучения развития таких интеллектуальных процессов, как внимание, восприятие и память.
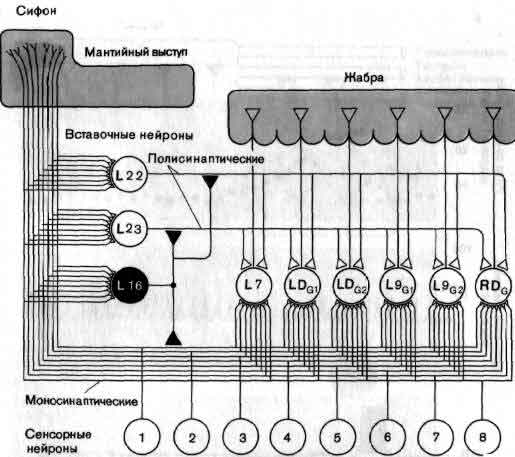
Нейронная цепь поведенческого рефлекса втягивания жабры у аплизии показана в виде схемы. Животное рефлекторно втягивает жабру, когда на сифон действует какой-нибудь стимул. Кожу сифона иннервируют 24 сенсорных нейрона; на схеме показаны только восемь из них. Сенсорные нейроны образуют моносинаптические, или прямые, связи с шестью идентифицированными мотонейронами жабры, показанными в ряду, который начинается с L7, и по меньшей мере с одной тормозной клеткой (L16) и с двумя возбуждающими вставочными нейронами (L22 и L23), которые образуют синапсы с мотонейронами.
В привыкании у позвоночных интересно то, что оно порождает и кратковременную и долговременную память и поэтому используется для исследования связи между ними. Мы с Т. Кэрью (Т. Carew) и Г. Пинскером (Н. Pinsker) нашли подобную связь у аплизии. После одного тренировочного опыта, состоящего из 10-15 тактильных раздражений сифона, наступает привыкание рефлекса втягивания. Память на этот стимул кратковременная; через час можно обнаружить частичное восстановление, а через день оно обычно становится почти полным. При таком типе обучения восстановление равнозначно забыванию. Однако при повторении более сложных задач обучения четыре повторных тренировочных опыта, всего лишь по 10 стимулов каждый, вызывают глубокое привыкание и запоминание стимула, которое длится недели.
Первый вопрос, каким задались мы с В. Кастеллучи (V. Castellucci), Купферманом и Пинскером, состоял в следующем: где локализуется кратковременное привыкание и каковы его механизмы? Нейронная цепь, управляющая втягиванием жабры, совсем проста. Стимуляция кожи сифона активирует там 24 сенсорных нейрона; они образуют прямые связи с четырьмя мотонейронами в жабре, а мотонейроны прямо соединены с мышцей. Сенсорные нейроны возбуждают также несколько вставочных, т.е. промежуточных нейронов.
Исследуя эти клетки во время привыкания, мы нашли, что при кратковременном привыкании изменяется сила связи сенсорных нейронов со следующими за ними центральными клетками — вставочными и мотонейронами. Такая локализация была весьма удачной, потому что позволяла исследовать, что происходит при привыкании, путем анализа изменений в обеих клетках — пресинаптическом сенсорном нейроне и постсинаптическом мотонейроне — и в единственной группе связей между ними.
Силу связи можно определять, регистрируя синаптическое действие, вызываемое в мотонейронах отдельным сенсорным нейроном. Можно имитировать опыт по тренировке привыкания, состоящей из 10—15 стимулов, непосредственно стимулируя сенсорный нейрон в той же строгой временной последовательности, какая применяется для целого животного. Стимул можно отрегулировать таким образом, чтобы он вызывал один потенциал действия. Когда нейрон отвечает потенциалом действия впервые, он производит весьма эффективное синаптическое действие, которое выражается крупным возбудительным постсинаптическим потенциалом в мотонейроне. Последующие потенциалы действия, создаваемые в сенсорном нейроне во время тренировочного опыта, вызывают все меньшие возбудительные постсинаптические потенциалы. Эта депрессия эффективности связи идет параллельно поведенческому привыканию и определяет его. Как и поведение, синаптическая депрессия, создаваемая одним тренировочным опытом, сохраняется больше часа. После второго такого опыта депрессия синаптического потенциала выражена сильнее, а последующие тренировочные опыты могут подавить синаптический потенциал полностью.
В чем причина изменений силы синаптических связей? Происходит ли изменение в пресинаптическом сенсорном нейроне, которое выражается в пониженном выходе медиатора, или же изменяется постсинаптическая клетка, что выражается в снижении чувствительности рецепторов к медиатору? Ответить на эти вопросы можно, проанализировав изменения амплитуды синаптического потенциала, проявляющиеся в его «квантовых» компонентах.
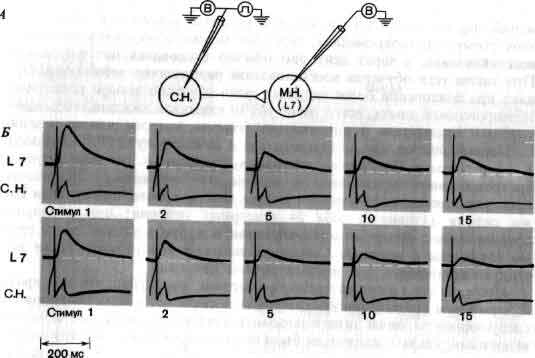
Процесс привыкания, когда реакция животного на стимул постепенно ослабевает, если он оказывается несущественным, является элементарной формой обучения и памяти, которая обнаруживается на уровне одиночного мотонейрона (М. Н.). Здесь показан такой опыт (А), в котором сенсорный нейрон (С. Н) аплизии образующий синапс на мотонейроне L7, стимулируется каждые 10 секунд Б. Отдельные кадры из двух последовательных тренировочных серий по 15 стимулов каждая, разделенных интервалами по 15 минут, показывают как ответ L7 они жается и исчезает.
Как впервые показали X. дель Кастильо (J. del Castillo) и Б. Катц (В. Katz) в Лондонском университетском колледже, медиатор выделяется не в виде отдельных молекул, а «квантами», или мультимолекулярными пакетами. Все пакеты содержат приблизительно одинаковое количество медиатора (по нескольку тысяч молекул). Как полагают кванты хранятся в субклеточных органеллах, называемых синаптическими пузырьками, которые во множестве видны в синаптических окончаниях на электронных микрофотографиях. Поскольку число молекул медиатора в каждом кванте обычно не меняется, количество квантов высвобождаемых каждым потенциалом действия, служит достаточно надежным показателем общего количества выделившегося медиатора Каждый квант в свою очередь вызывает в постсинаптической клетке миниатюрный постсинаптический потенциал характерной величины Величина эта служит показателем того, насколько чувствительны постсинаптические рецепторы к нескольким тысячам молекул медиатора выделяемым каждым пакетом.
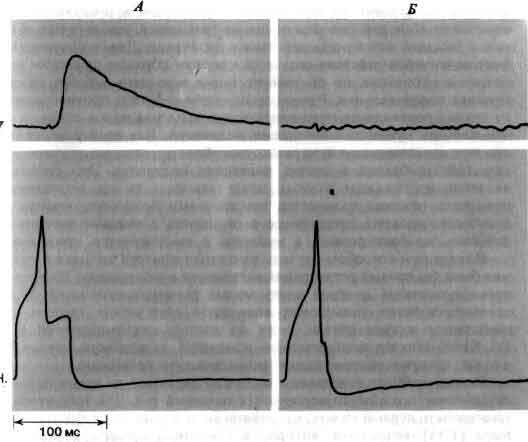
Долговременное привыкание выявляется при сравнении синаптических связей между сенсорным нейроном (С. Н.) и мотонейроном L7 у нетренированной аплизии (контроль, А) и у аплизии, тренированной на долговременное привыкание (Б). У контрольных животных за импульсом в сенсорном нейроне следует крупный возбудительный синаптический ответ мотонейрона. У тренированных животных синаптическую связь почти невозможно обнаружить.
Исследуя аплизию, мы с Кастеллучи, нашли, что снижение амплитуды синаптического потенциала действия по мере привыкания идет параллелъно уменьшению числа выделяющихся химических квантов. Напротив, величина миниатюрного постсинаптического потенциала не менялась, что указывало на неизменность чувствительности постсинаптического рецептора. Эти данные говорят о том, что кратковременное привыкание локализуется в пресинаптических окончаниях сенсорных нейронов и что механизм привыкания состоит в постепенном снижении количества медиатора, высвобождаемого окончаниями сенсорного нейрона на центральных клетках-мишенях. Опыты на раке, проведенные Р. Цукером (R. Zucker) из Калифорнийского университета в Беркли и Ф. Красне (F. Krasne) из Калифорнийского университета в Лос-Анджелесе, а также опыты на кошке, поставленные П. Фейрелом (P. Farel) и Р. Томпсоном (R. Thompson) из Калифорнийского университета в Эр-вине, показывают, что этот механизм может быть универсальным. От чего зависит уменьшение числа квантов, высвобождаемых каждым потенциалом действия? Это число в значительной степени определяется концентрацией свободного кальция в пресинаптическом окончании. Кальций является одним из трех ионов, участвующих в генерации каждого потенциала действия в окончании. Деполяризующий эффект потенциала действия создается главным образом притоком ионов натрия в окончание, но он требует также меньшего и более позднего притока ионов кальция. Реполяризационное действие противоположного знака производится преимущественно выходом ионов калия, а приток кальция нужен для выделения медиатора. Как полагают, кальций придает синаптическим пузырькам способность связываться с пресинаптической мембраной в местах выделения медиатора. Это связывание является критическим этапом, предваряющим выход медиатора из пузырьков (процесс называется экзоцитозом). Возможно поэтому, что количество кальция, приходящее в окончания с каждым потенциалом действия, не фиксировано, а меняется и модулируется привыканием.
Наилучшим способом изучать изменения притока кальция в окончания была бы прямая регистрация активности в окончаниях. Нам не удалось осуществить ее из-за очень малых размеров окончаний. Но поскольку свойства кальциевых каналов в окончаниях сходны с их свойствами в теле клетки, один из наших дипломников М. Клейн (М. Klein) занялся исследованием изменений кальциевого тока в теле клетки, которые сопровождают синаптическую депрессию.
Кальциевый ток возникает и медленно нарастает во время потенциала действия; его обычно перекрывает калиевый ток. Для того чтобы демаскировать первый из них, мы действовали на ганглий тетраэтиламмонием (ТЭА) - веществом, которое в некоторой степени избирательно блокирует поздний калиевый ток. Блокируя реполяризующее действие калиевого тока, ТЭА незначительно удлиняет потенциал действия. Удлинение это в большой мере обусловлено беспрепятственным действием кальциевого тока. Вызываемое ТЭА удлинение потенциала действия служит хорошим критерием изменения кальциевого тока.
Затем мы определяли выделение медиатора из окончаний сенсорных нейронов по величине синаптического потенциала в мотонейроне и измеряли одновременно регистрируемые изменения кальциевого тока по критерию длительности потенциала действия. Мы нашли, что многократная стимуляция сенсорного нейрона с частотой, вызывающей привыкание, приводила к постепенному укорочению кальциевого компонента потенциала действия, что происходило параллельно уменьшению выхода медиатора. Спонтанное восстановление синаптического потенциала и поведения сопровождалось усилением кальциевого тока.
То, что мы до сих пор узнали о механизмах кратковременного привыкания, показывает, что этот тип обучения связан с модуляцией силы ранее сформированной синаптической связи. Сила связи определяется количеством выделяемого медиатора, которое в свою очередь зависит от того, насколько потенциал действия в пресинаптическом окончании активирует кальциевый ток. Поэтому сохранение кратковременного привыкания основано на том, что депрессия кальциевого тока в пресинаптическом окончании не прекращается в течение минут и часов.
Каковы пределы этого изменения? Насколько может измениться эффективность данного синапса в результате обучения, и как долго сохраняются такие изменения? Выше я указывал, что многократные тренировочные опыты могут привести к полной депрессии синаптических связей между сенсорными и моторными клетками. Удерживается ли такое состояние? Может ли долговременное привыкание привести к полной и длительной инактивации ранее активного синапса?
В связи с этими вопросами исследователи обучения давно ведут споры о соотношении кратковременной и долговременной памяти. Наиболее принято представление, что эти два вида памяти связаны с разными процессами. Однако оно основано на косвенных данных.
Совместно с Кастеллучи и Кэрью мы предприняли более прямую проверку этой гипотезы путем сравнения эффективности связей, образоманных популяцией сенсорных нейронов с идентифицированным мотонейроном жабры L7, в четырех группах аплизий: у нетренированных животных, служивших контролем, и в группах, исследованных соответственно через сутки, неделю и три недели после выработки долговременного привыкания. Было обнаружено, что у контрольных животных около 90% сенсорных нейронов имели чрезвычайно эффективные связи с L7, тогда как у животных, исследованных через сутки и через неделю после долговременного привыкания, их число составляло 30%. Даже через три недели только у 60% клеток можно было обнаружить связи с L7. Таким образом, в результате простого обучения ранее эффективные синаптические связи становятся неактивными и остаются в таком состоянии более недели.
Следовательно, в то время как кратковременное привыкание сопровождается кратким ослаблением эффективности синапса, долговременное привыкание вызывает более продолжительное и глубокое изменение, которое приводит к функциональному нарушению большинства ранее эффективных связей. Эти данные интересны по трем причинам: 1) они служат прямым доказательством того, что специальный случай долговременной памяти может быть объяснен продолжительным изменением эффективности синапса; 2) они показывают, что достаточно поразительно малой тренировки, чтобы вызвать глубокое изменение синаптической передачи в синапсах, играющих решающую роль при обучении, и 3) они ясно показывают, что кратковременное и долговременное привыкания локализуются в одной и той же части нейрона, а именно в синапсах, образованных сенсорными нейронами на мотонейронах. Кратковременное и долговременное привыкания связаны также с одним и тем же клеточным механизмом — депрессией передачи возбуждения. Теперь нужно определить, является ли долговременная синаптическая депрессия пресинаптической и связана ли она с инактивацией натриевого тока. Если окажется, что это так, то тем самым на более основательном уровне подтвердится представление о едином механизме кратковременной и долговременной памяти.
Сенситизация представляет собой несколько более сложную форму обучения; ее можно обнаружить на рефлексе втягивания жабры. Сенситизация представляет собой длительное усиление ранее существовавшего ответа животного на стимул в результате нанесения другого, ноцицептивного (болевого, повреждающего) стимула. В то время как привыкание требует от животного, чтобы оно научилось итерировать определенный стимул, потому что его последствия несущественны, сенситизация требует, чтобы животное научилось обращать внимание на стимул, так как он сопровождается потенциально болезненным эффектом или опасными последствиями. Поэтому когда на голову аплизии наносится вредоносный стимул, рефлекс втягивания жабры па многократную стимуляцию сифона очень усиливается. Как и привыкание, сенситизация может длиться от минут до дней и недель в зависимости от длительности тренировки. Здесь будет рассмотрена только ее кратковременная форма.
Мы с Кастеллучи установили, что сенситизация влечет за собой изменение синаптической передачи в том же самом пункте, который участвует в привыкании, а именно в синапсах, образованных сенсорными нейронами на центральных клетках-мишенях. Наши физиологические исследования и последующие морфологические исследования К. Бейли (С. Baily), М. Чен (М. Chen) и Р. Хоукинса (R. Hawkins) показывают, что нейроны, осуществляющие сенситизацию, оканчиваются близ синаптических терминалей сенсорных нейронов и усиливают выход медиатора, увеличивая число квантов, высвобождаемых каждым потенциалом действия в сенсорном нейроне. Поэтому такой процесс называется пресинаптическим облегчением. Эти данные представляют интерес, так как они показывают [как и более ранние данные о пресинаптическом торможении в другой системе, полученные Дж. Дьюделом (J. Dudel) и С. Куфлером (S. Kuffler) из Гарвардской медицинской школы], что нейроны обладают рецепторами для медиаторов в двух совершенно разных участках. От рецепторов на теле клетки и на дендритах зависит, возникнет ли в клетке потенциал действия, а от рецепторов на синаптических окончаниях зависит, сколько медиатора высвободится при каждом потенциале действия.
Поэтому на одни и те же пункты — пресинаптические окончания сенсорных нейронов — могут воздействовать противоположным образом две противоположные формы обучения. Во время привыкания они угнетаются в результате возникающей собственной активности нейрона, а при сенситизации в них наступает облегчение вследствие активности других нейронов, которые образуют на них синапсы. Эти данные, полученные на уровне отдельной клетки, подтверждают наблюдение, сделанное на поведенческом уровне, что привыкание и сенситизация представляют собой независимые друг от друга противоположные формы обучения.
В связи с этими данными возникает интересный вопрос. Сенситизация способна усилить нормальный рефлекс, но может ли она противодействовать глубокой депрессии рефлекса, вызванной долговременным привыканием? Если может, то восстанавливает ли она синаптические связи, полностью инактивированные долговременным привыканием?
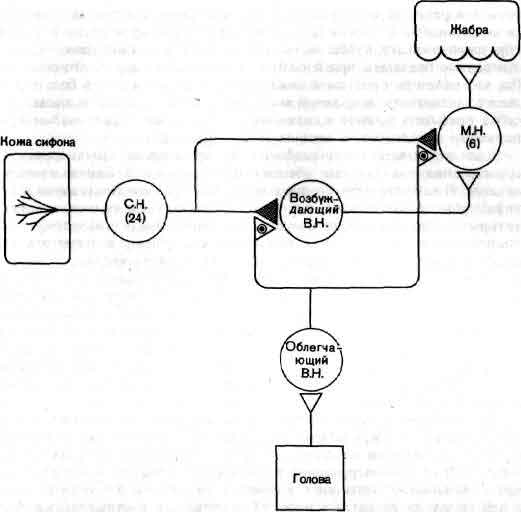
Сенситизация является такой формой обучения и памяти, при которой ответ на стимул усиливается под влиянием другого, более сильного стимула. Здесь рефлекс втягивания жабры у аплизии усиливается в результате ноцицептивного раздражения области головы. Этот стимул активирует нейроны, которые возбуждают облегчающие вставочные нейроны (В. Н.); последние оканчиваются на синаптических окончаниях сенсорных нейронов. Эти нейроны пластичны, т. е. способны изменять эффективность своего синапса. Медиатор облегчающих вставочных нейронов, предположительно серотонин (точки в кружке), модулирует выделение медиатора сенсорного нейрона на возбуждающие вставочные нейроны и мотонейроны.
Кэрью, Кастеллучи и я изучили этот вопрос и нашли, что сенситизация устраняла депрессию в поведении. Кроме того, за один час сенситизирующая стимуляция головы аплизии восстанавливала функционально инактивированные синапсы (которые оставались бы в таком состоянии в течение недель).
Следовательно, в головном мозгу имеются синаптические пути, которые детерминированы процессами развития, но которые, будучи предрасположены к обучению, могут быть функционально инактивированы или реактивированы опытом. В самом деле, для этих поддающихся модификации синапсов достаточно небольшой тренировки или приобретенного опыта, чтобы вызвать в них глубокие изменения. Если бы эти данные оказались применимыми к мозгу человека, то это означало бы, что даже при простом социальном акте, например при беседе двух людей, активность нейронной механики в мозгу одного человека способна оказывать прямой и, возможно, длительный эффект на модифицируемые синаптические связи в мозгу другого.
Для экспериментатора особенно привлекательна кратковременная сенситизация, так как она обещает быть доступной биохимическому анализу. В качестве первого шага мы с Хоукинсом и Кастеллучи идентифицировали специальные клетки в абдоминальном ганглии аплизии, которые создают пресинаптическое облегчение. Вводя животному электроноплотную метку, чтобы наполнить ею нейрон и пометить его синаптические окончания, мы нашли, что последние содержат пузырьки, сходные с обнаруженными у аплизии Л. Школьник (L. Shkolnik) и Дж. Шварцем (J. Schwartz) в нейроне, медиатором которого, как ранее было установлено, является серотонин. В соответствии с возможной серотонинергической природой этой клетки, М. Брунелли (М. Brunelli), Кастеллучи, Т. Томоски-Сайкс (Т. Tomosky-Sykes) и я нашли, что серотонин усиливал моносинаптическую связь между сенсорным нейроном и мотонейроном L7, тогда как другие вероятные медиаторы подобного действия не оказывали.
Далее мы обнаружили интересную связь между серотонином и внутриклеточным посредником — циклическим аденозинмонофосфатом (цАМФ). Со времени классической работы И. Сьюзерленда-мл. (Е. Sutherland, Jr.) и его сотрудников в Университете Вандербилта известно, что большинство пептидных гормонов не входят в клетку-мишень, а действуют на рецептор клеточной поверхности и стимулируют фермент, называемый аденилатциклазой; этот фермент катализирует в клетке превращение аденозинтрифосфата (АТФ) в цАМФ, который затем действует как «второй посредник» («первым посредником» является гормон) в нескольких точках внутри клетки и вызывает ряд соответствующих изменений функции.
X. Седар (Н. Cedar), Шварц и я нашли, что сильная и длительная стимуляция проводящего пути, который идет у аплизии от головы и по которому осуществляется сенситизация, через синапс повышала содержание цАМФ во всем ганглии. Седар и Шварц, а также И. Левитан (I. Levitan) и С. Барондес (S. Barondes) показали, что длительное повышение содержания цАМФ можно вызвать также инкубацией ганглия с серотонином. Для выяснения связи между серотонином и цАМФ Брунелли, Кастеллучи и я вводили цАМФ в тело сенсорного нейрона и установили, что он тоже вызывает пресинаптическое облегчение, тогда как инъекция 5'-АМФ (продукта распада цАМФ) или другого второго посредника — циклического ГМФ — не давала такого эффекта.
Поскольку привыкание связано с уменьшением кальциевого тока, заманчиво было думать, что цАМФ производит свое облегчающее действие, усиливая кальциевый ток. Как было сказано, кальциевый ток обычно маскируется калиевым током. Поэтому мы с Клейном проследили за потенциалами действия в сенсорных нейронах, в которых калиевый ток был ослаблен посредством ТЭА. Стимуляция пути, который идет от головы и по которому осуществляется сенситизация, или же стимуляция одиночного облегчающего нейрона, усиливала кальциевый ток, что было видно по удлинению потенциала действия на фоне ТЭА, и это усиление сохранялось 15 минут или дольше. Оно шло параллельно с увеличением выхода медиатора, и параллельно этим двум синаптическим изменениям возрастал рефлекс на сенситизирующий стимул.
Усиление кальциевого тока, выраженное в продлении кальциевого компонента потенциала действия после стимуляции сенситизирующего пути, можно было создать внеклеточным нанесением серотонина или двух веществ, которые повышают внутриклеточный уровень цАМФ, ингибируя фосфодиэстеразу — фермент, катализирующий расщепление цАМФ. Такие же эффекты наблюдались после прямой внутриклеточной инъекции цАМФ, но не 5'-АМФ.
На основании этих данных Клейн и я предположили, что стимуляция облегчающих нейронов сенситизирующего пути ведет к выходу серотонина, который активирует серотонин-чувствительный фермент (аденилатциклазу) в мембране окончания сенсорного нейрона. Возникающее при этом в окончании увеличение количества цАМФ ведет к усиленной активации кальциевого тока либо непосредственно путем активации кальциевого канала, либо косвенно, снижением противодействующего калиевого тока. С каждым потенциалом действия растет приток кальция и выход медиатора.
Главное, что вначале привлекло к использованию аплизии для изучения поведения, было наличие крупных клеток, электрические свойства и связи которых доступны детальному исследованию. А теперь размеры этих клеток могут оказаться еще более выгодными для изучения субклеточных и биохимических механизмов обучения, с одной стороны, и возможных изменений мембранной структуры — с другой. Так, например, интересно узнать поточнее, как повышение уровня цАМФ при сенситизации связано с активацией кальциевого тока, поскольку установление механизма этой связи может послужить первым шагом к пониманию на молекулярном уровне этой простой формы кратковременного обучения.
На память приходит целый ряд механизмов. Как полагают, каналы, по которым ионы проходят через мембрану нейрона, состоят из белковых молекул. Поэтому очевидная возможность состоит в том, что цАМФ активирует одну или более протеинкиназ — ферментов, которые, по предположению П. Грингарда (P. Greengard) из Медицинской школы Йельского университета, могли бы служить общим молекулярным механизмом различных эффектов цАМФ внутри клетки. Протеинкиназы — это ферменты, которые фосфорилируют белки, иначе говоря, присоединяют фосфорильную группу к боковой цепи аминокислот серина или треонина в белковой молекуле и этим придают белкам иной заряд и иную конфигурацию, что меняет их функцию, активируя одни и инактивируя другие. Фосфорилирование могло бы служить эффективным механизмом регуляции памяти. Один из способов, каким могла бы действовать сенситизация, состоит в том, что белок кальциевого канала активируется (или же белок антагонистического калиевого канала инактивируется) при фосфорилировании его протеинкиназой, зависимой от цАМФ.
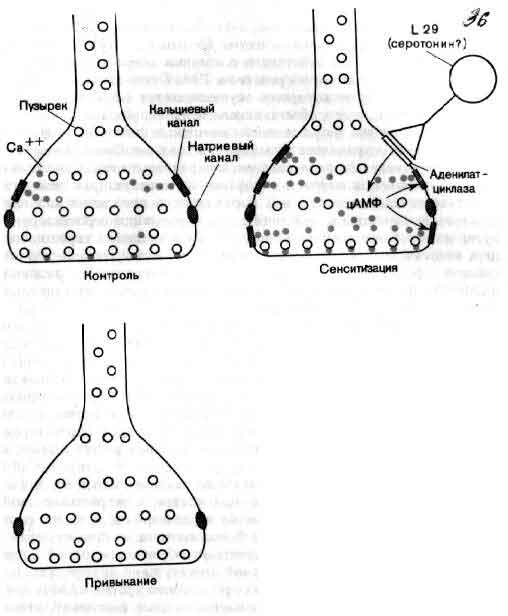
Модель кратковременной сенситизации и привыкания на уровне одиночного сенсорного нейрона, начиная с контрольной ситуации, когда клетка генерирует импульсы до наступления сенситизации или привыкания. Нервный импульс в концевой мембране нейрона открывает параллельно натриевым каналам (Na + ) ряд каналов для ионов кальция (Са++). Сенситизация вызывается группой клеток L29 (возможно, и другими), которые, как полагают, выделяют медиатор серотонин. Он действует на аденилатциклазу, фермент, катализирующий синтез циклического аденозинмонофосфата (цАМФ) в нейронных окончаниях. цАМФ повышает приток ионов кальция, возможно, создавая больше кальциевых каналов. Кальций усиливает слияние содержащих медиатор пузырьков с мембраной в местах его выделения, увеличивая вероятность последнего. При привыкании многократная импульсация в окончаниях, возможно, уменьшает число открытых кальциевых каналов, понижая приток кальция и инактивируя синапс.
Сенситизация занимает интересное место в иерархии обучения. Ее часто считают предшественницей классического условного рефлекса. В обеих этих формах рефлекс на стимул усиливается в результате активации другого пути. Сенситизация отличается от условного рефлекса тем, что она не ассоциативна; сенситизирующий стимул повышает рефлекторную реактивность независимо от того, сочетается ли он во времени со стимулом, вызывающим рефлекс. Теперь Э. Гелперин (A. Gelperin) из Принстонского университета, Дж. Мпитсос (G. Mpitsos) и С. Коллин (S. Collins) из Университета Кейса Западной резервной территории, а также Т. Кроу (Т. Crow) и Д. Олкон (D. Alkon) из Национальных институтов здравоохранения к настоящему времени обнаружили у моллюсков несколько типов ассоциативного обучения. Недавно мы с Т. Уолтерсом (Т. Walters) и Кэрью получили доказательства ассоциативного обучения у аплизии. Поэтому скоро мы будем в состоянии точно проанализировать, как механизмы сенситизации соотносятся с механизмами ассоциативного обучения.
Другое возможное теперь направление исследования состоит в рассмотрении связи между исходным развитием нейронной цепи у эмбриона и ее последующей модификацией обучением. И при развитии, и при обучении в нервной системе происходят функциональные изменения — меняются эффективность синапсов и другие свойства нейронов. Как соотносятся такие изменения? Основаны ли механизмы обучения на механизмах пластичности, проявляющейся во время развития или же позднее формируются совершенно новые специализированные для обучения процессы?
Какими бы ни оказались ответы на эти интригующие вопросы, при исследовании беспозвоночных животных возникло поразительное и обнадеживающее обстоятельство: теперь стало возможным точно локализовать и наблюдать на клеточном - а в конце концов, возможно, и на молекулярном - уровне простые свойства памяти и обучения. Хотя для сложного головного мозга высших животных характерны некоторые высшие проявления умственной деятельности, теперь стало ясно, что элементарные свойства того, что считается мыслительными процессами, можно обнаружить в активности лишь очень малого числа нейронов. Поэтому и с философской, и с методической точек зрения интересно будет узнать, в какой степени высшие формы мышления можно объяснить действием более простых компонентов и механизмов. В той мере, в какой возможны такие редукционистские объяснения, важно также установить, как единицы этой элементарной азбуки комбинируются для создания языка гораздо более сложных мыслительных процессов.
У. НАУТА, М. ФЕЙРТАГ
Организация мозга
Головной и спинной мозг млекопитающих, включая человека, состоит из нескольких миллиардов нервных клеток, причем отдельные нейроны могут быть связаны с тысячами других. Как же организована эта огромная трехмерная сеть?
Мы видим два общих подхода к представлению о нейроанатомии. Первый — высокопарный: утверждается, что мозг — вместилище ума, чувств и желаний, памяти и способности учиться, и того любопытного ощущения, которое свойственно людям, — ощущения будущего. Затем мы начинаем внимательно рассматривать, как этот таинственный орган выглядит, так сказать, «во плоти». Определенные части мозга, в особенности кора больших полушарий, удивительным образом организованы; другие поражают своей кажущейся неупорядоченностью. Но даже и самые высокоупорядоченные структуры, в которых организация нейронов и разнообразных связей между ними напоминает электронные схемы, сопротивляются нашим сегодняшним попыткам понять их.
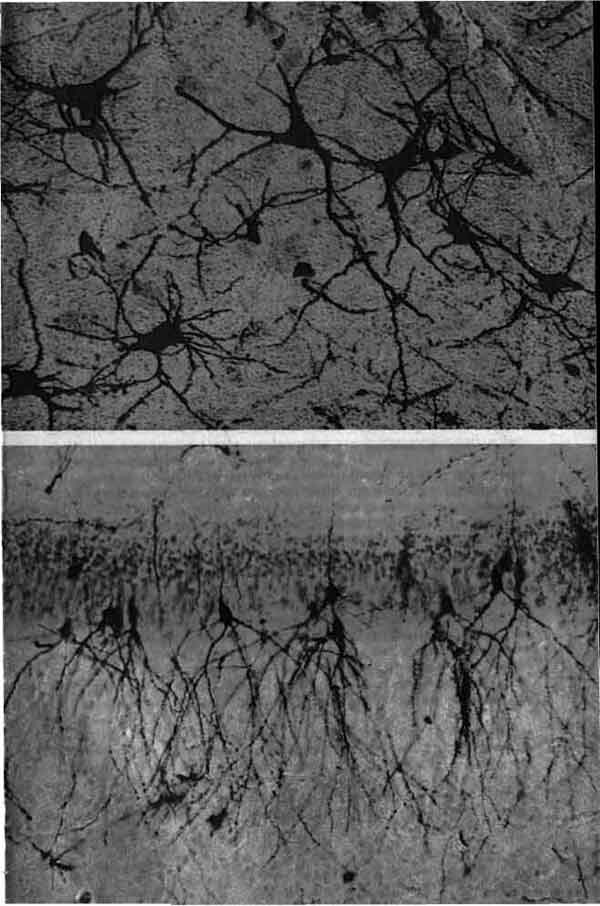
О порядке и беспорядке в клеточной организации мозговых структур можно судить по этим двум микрофотографиям. Обе они представляют тонкие срезы мозга кошки, которые подверглись двойному окрашиванию: по методу Гольджи, позволяющему выявить некоторые из нейронов со всеми их отростками в виде черных силуэтов, и по методу Ниссля, приводящему к окрашиванию всех клеточных тел в синий цвет. Метод Гольджи обеспечивает прокрашивание только 5 или менее процентов всех нейронов, по-видимому, по случайному выбору; если бы прокрашивались все нейроны, данная ткань выглядела бы однородно черной. А. Зубчатая извилина гиппокампа; тела нейронов в форме продолговатых пирамид располагаются в ряд, отсылая свои отростки почти параллельным пучком. Б. Микрофотография относится к крупноклеточной ретикулярной формации. Здесь нейроны образуют неупорядоченную сеть.
Второй подход более прозаический. Мозг просто подразделяется на части, поскольку подходящие методы окрашивания выявляют в одних местах скопления нейронов, опутанных густой сетью их же собственных нитевидных отростков, в других же ткань состоит в основном из длинных Волокон нервных клеток — аксонов, которые обслуживают связи на далекие расстояния в нервной системе. Ткань первого рода — это серое вещество, второго — белое вещество.
Конечно, есть искушение приписать каждой области какую-нибудь функцию, как если бы мозг в целом был чем-то вроде радиоприемника. Но существо работы центральной нервной системы - головного и спинного мозга — это проведение входящей сенсорной информации к множеству структур и конвергенция на нейронах, которые возбуждают эффекторные органы: мышцы и железы. Следовательно, система в целом обладает свойствами, выходящими за пределы тех, которые можно обнаружить у простого набора модулей.
Возьмем, например, такую структуру мозга как субталамическое ядро. Его разрушение у человека ведет к моторной дисфункции, известной под названием гемибаллизма, при которой больной делает непроизвольные движения, как бы бросая мяч. Следует ли отсюда, что нормальной функцией субталамического ядра должно быть подавление движений, напоминающих бросание мяча? Конечно, нет; данное состояние больного только дает представление о работе центральной нервной системы, выведенной из равновесия отсутствием субталамического ядра.
Мы упоминаем эти вещи, чтобы установить ограничения для любых объяснений анатомии мозга. Мы дадим здесь какое-то объяснение, но оно по необходимости будет несколько расплывчатым. Делать иного рода предположения было бы просто неправомерно.
Полезно сделать некоторые предварительные замечания. В первые десятилетия этого века Дж. Паркер (G. Parker) из Йельского университета искал самую примитивную рефлекторную дугу. Такие дуги были идентифицированы у позвоночных животных; это состоящие из одного или нескольких нейронов пути, с помощью которых возбуждение, порождаемое сенсорным стимулом, действующим на некоторую часть тела, может быть проведено к эффекторной ткани и, соответственно, вызвать движение. Во времена Паркера рефлекторные дуги рассматривались как простейшие схемы, посредством которых природа объединяет клетки в нервную систему; в соответствии с этим было широко распространено мнение, что нервная система возникает тогда, когда некоторый организм приходит к необходимости иметь какую-нибудь клетку или цепочку клеток для посредничества между внешним стимулом и ответным движением животного. Со временем предполагалось установить, что у более развитых организмов эволюция нервной системы продвигалась вперед путем увеличения числа и сложности таких цепочек.
Сначала внимание Паркера привлекли эпителиальные слои некоторых морских гидроидных полипов и актиний, потому что иногда они содержали клетки, которые выглядели (если были подходящим образом окрашены) как нейроны. У основания такой клетки Паркеру удалось разглядеть начало нити, весьма похожей на аксон, которая, подходя к мышечному волокну, разветвлялась. Паркер не был уверен в том, что тут образовывался контакт, но предположил, что между аксоном и мышечным волокном устанавливается какая-то связь. Конечно, он был прав, но это устройство весьма примитивно; его схему можно было бы назвать однонейронной нервной системой, поскольку вся линия проведения состоит всего лишь из одной клетки. Что такая нервная система будет делать в ответ на стимул, столь же предсказуемо, как и то, что будет делать дверной звонок, если нажать на кнопку. В то же время относительно нервной системы человека ясно, что поведение, на которое она делает способными человеческие существа, самое что ни на есть непредсказуемое.
Очевидно, что-то должно вмешиваться в работу механизма дверного звонка, и потому Паркер исследовал ситуацию у несколько более сложных организмов. В эпителиальном слое у некоторых полипов и медуз он обнаружил нейронные структуры, сходные с найденными им ранее. Однако под эпителием он теперь нашел дополнительные нейроны, которые вместе образовали широко разветвляющуюся сеть. Таким образом, нервная система этой второй группы организмов оказалась усложненной : нейроны эпителиального слоя образовывали контакты с субэпителиальной сетью, а клетки этой сети в свою очередь образовывали контакты с сократимыми тканями в глубине организма. Следовательно, уже можно было говорить о двухнейронной нервной системе, в которой сенсорные нейроны (у этих простых созданий нейроны лежат вблизи поверхности и находятся в непосредственном контакте с окружающей средой) связаны с мотонейронами (нейронами, которые образуют контакты с эффектерными клетками, в данном случае — сократимыми клетками, и, значит, по существу, с мышечными волокнами).
Остается ли эта сеть в высшей степени предсказуемой? По-видимому, нет. Предположим, что мотонейроны связаны друг с другом и на вход каждого из них поступают не только сообщения, идущие из окружающей среды через посредство сенсорных нейронов, но и сообщения от соседних мотонейронов. Вообразим далее, что некоторые из этих сообщений могут быть возбуждающими, т.е. заставляют данный мотонейрон с большей готовностью генерировать и передавать свои собственные сигналы в ответ на поступление других сигналов, и что другие сообщения могут быть тормозными. В таких условиях приходится решать вот какую задачу: чтобы предсказать, что будет делать некоторый нейрон в ответ на различные входные сигналы, нужно алгебраически просуммировать возбуждающие и тормозные сообщения, которые на нем конвергируют.
Возможно, что такое двухнейронное устройство позволяет облагодетельствованной природой медузе быть более непредсказуемой в своем поведении, чем актинии и другие организмы с однонейронной нервной системой. Однако затем следует дальнейшее достижение, и оно тоже обнаружено у очень примитивных организмов — некоторых других меду:». В каком-то смысле это высшее достижение, так как и нервная система таких медуз, и нервная система чело века — обе состоят по существу только из трех классов нейронов. У этих медуз, как и у человека, сенсорные нейроны, как правило, больше не связаны непосредственно с мотонейронами. Между этими двумя элементами развивается барьер из нейронов, которые имеют взаимосвязи не только с мотонейронами, но и друг с другом.
Правда, этот третий и последний шаг, возможно, уже был сделан всеми организмами, которые имеют субэпителиальную сеть нервных клеток. Выше, при рассмотрении двухнейронной нервной системы, предполагалось, что все клетки, составляющие этот слой, — мотонейроны, т.е. клетки, иннервирующие эффекторные ткани. Однако в действительности такие связи могут устанавливать только некоторые из многих субэпителиальных клеток. Остальные могут располагаться в сплетении таким образом, что получают на вход сингалы от сенсорных нейронов эпителия, а сообщаются лишь с другими нейронами того же типа или с мотонейронами, но не с эффекторными структурами. Не являясь ни сенсорными, ни моторными, они служат посредниками при проведении сигнала по сенсомоторному пути.
Короче, здесь тоже имеются промежуточные (вставочные) нейроны. Хотя трехнейронную организацию трудно вычленить в диффузной нейронной сети, она широко представлена на более поздних стадиях эволюции; у животных, более высоко развитых, чем медузы, диффузная субэпителиальная нервная сеть концентрируется либо в последовательность сегментарных ганглиев (скоплений нейронов), либо в единую несегментированную центральную нервную систему. Таким образом, критическим моментом явилось изобретение «великой промежуточной сети» — барьера из промежуточных нейронов, которые вклинились между сенсорными нейронами и мотонейронами, на раннем этапе эволюции животных.
Насколько далеко зашел прогресс в развитии этой промежуточной сети на сегодняшний день, легче всего показать с помощью некоторых цифр. Начнем с вопроса о том, сколько нейронов в центральной нервной системе человека. В качестве ответа часто можно услышать: порядка 1010. Это результат подсчета вставочных нейронов и мотонейронов, ибо случилось так, что настоящие сенсорные нейроны лежат не в центральной нервной системе, а в ганглиях, которые расположены вне головного и спинного мозга. Это весьма привлекательная цифра, которую легко запоминать и использовать. И все же имеются еще классы нейронов, которые столь малы и столь плотно упакованы, что оценить их число трудно или невозможно. Один из таких классов — это клетки-зерна. Как раз в одной из частей головного мозга — мозжечке — находится так много клеток-зерен, что величина 1010 нейронов для всей центральной нервной системы становится сомнительной. Общее число вполне может быть на порядок, а возможно, и на два порядка больше.
Все же примем на минуту, что общее число действительно 1010. Сколько же из этих клеток мотонейроны? Ответ таков, что их, оказывается, не может быть больше двух или трех миллионов. Эта цифра представляется смущающе малой в свете того факта, что только через мотонейроны работа нервной системы может выразиться в движении. Более того, такой ответ заставляет предполагать, что на мотонейроны должно конвергировать неправдоподобно большое число влияний; другими словами, это заставляет предполагать, что типичный мотонейрон должен образовывать синаптические связи с огромным числом аксонов, исходящих от равно огромного числа нейронов великой промежуточной сети.
Полагают, что на поверхности типичного мотонейрона в спинном мозгу человека имеется что-то около 10000 синаптических контактов, из которых около 2000 - на клеточном теле и 8000 - на дендритах - отростках, ветвящихся локально, в отличие от единственного аксона. Это не означает, что на данный нейрон посягают 10000 вставочных нейронов: когда такие нейроны устанавливают связи с какой-то клеткой, они имеют тенденцию образовывать множественные синаптические контакты. Однако все равно получается, что средний мотонейрон должен подвергаться массированному воздействию: число 1010 для нейронов центральной нервной системы подразумевает, что на каждый мотонейрон приходится от 3000 до 5000 нейронов великой промежуточной сети.
Вот какое последнее заключение должно быть выведено из цифр, которые мы процитировали: весь головной и спинной мозг человека - это великая промежуточная сеть, за исключением явно немногих миллионов мотонейронов. А когда великая промежуточная сеть начинает включать 99,98 процента всех нейронов, составляющих центральную нервную систему, этот термин теряет большую часть своего смысла: он начинает отражать саму ту сложность, с которой каждый должен столкнуться при попытках постигнуть нервную систему. Этот термин остается полезным только как напоминание о том, что большая часть нейронов мозга не принадлежит ни к сенсорным, ни к моторным. Строго говоря, они вставлены между чисто сенсорной частью организма и чисто моторной его частью. Они являются компонентами вычислительной сети.
Вторая группа предварительных замечаний касается общей анатомии центральной нервной системы. Отметим, в частности, что у всех видов позвоночных головной и спинной мозг сначала появляются у зародыша всего лишь в виде трубки толщиной в один клеточный слой. В передней части этой нервной трубки, которая в конечном итоге будет заключена в череп, вскоре появляются три утолщения — первичные мозговые пузыри. Это ромбэнцефалон, или задний мозг; мезэнцефалон, или средний мозг; и прозэнцефалон, или передний мозг. («Энцефалон» происходит от греческого «внутри головы».)
Из этих трех первичных пузырей передний пузырь является самым продуктивным по числу образующихся отделов и дальнейшей дифференцировке. Основным событием его эмбрионального развития является образование камер с левой и правой стороны. Они превращаются в полушария большого мозга; эту часть мозга часто называют также конечным мозгом (телэнцефалон); у некоторых видов он имеет скромные размеры, у других - чрезвычайно большие. Между полушариями лежит непарный центральный отдел переднего мозга, от которого отпочковались полушария. Он имеет название промежуточный мозг, или диэнцефалон.
Эти два среза фиксированного препарата мозга человека иллюстрируют сложность внутреннего строения мозга. Срезы были обработаны краской, которая избирательно окрашивает в черный цвет жировую миелиновую оболочку нервных волокон; в результате белое вещество предстает как черное, а серое (состоящее преимущественно из клеточных тел) остается неокрашенным. Пустые пространства на этих срезах указывают положение желудочков—заполненных жидкостью полостей внутри мозга. Верхний рисунок. Срез проходит примерно посередине мозга и захватывает кору больших полушарий, гиппокамп и таламус. Нижний рисунок. Срез сдвинут кзади и проходит через ствол мозга. Как сделаны срезы и где располагаются упомянутые структуры, поясняет следующий рисунок. (Препарат из коллекции профессора П. Яковлева из Гарвардской медицинской школы.)
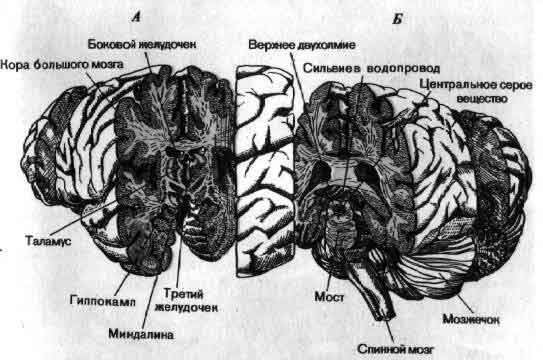
Мозг человека разрезан на пять частей и раскрыт как книжка, что позволяет проследить связи между наружным и внутренним строением. Два сечения, представленные на предыдущем рисунке, обозначены буквами А и Б.
В процессе развития из переднего мозга выделяется еще одна пара камер-глазные пузыри. Их имеют даже незрячие животные, а у животных, способных видеть, они удлиняются в направлении к поверхности головы и в конечном счете превращаются в две сетчатки, соединенные с основанием переднего мозга своими стеблями-зрительными нервами. Наконец, из внутренней поверхности первичного переднего мозга развивается непарная срединная камера, которая дифференцируется и образует заднюю долю гипофизарного комплекса.
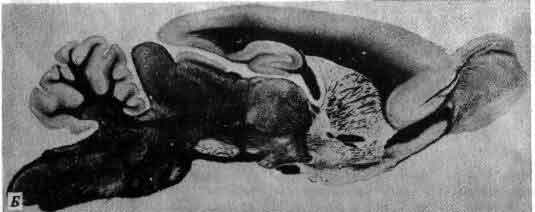
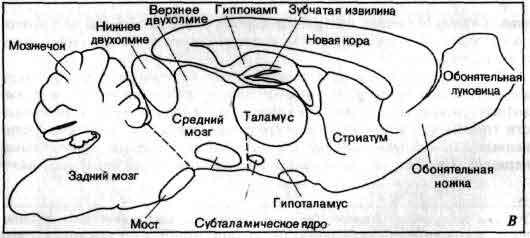
Два метода окрашивания дают возможность получить взаимно дополняющие друг друга изображения внутреннего строения мозга крысы. А. Показано сечение, проходящее чуть в стороне от срединной плоскости параллельно ей; срез был обработан по методу Ниссля, который позволяет избирательно окрашивать клеточные тела. Следовательно, каждая точка на микрофотографии соответствует какой-то отдельной клетке. Б. Срез обработан по методу Лойе (Loyez), который позволяет избирательно окрашивать миелинизированные волокна, оставляя тела клеток непрокрашенными, благодаря чему выделяются приводящие пути. В. Схема, на которой показаны различные структуры мозга.
показан результат этого эмбрионального роста. Такая схема в общем выдерживается для всех млекопитающих; она изображает полностью сформированную центральную нервную систему млекопитающего, разбитую на несколько отделов. Слева - спинной мозг (на рисунке — сильно укороченный). Вправо от него, без какой-либо резкой границы, отходит задний мозг — самый нижний отдел головного мозга. Его дорсальная часть (часть, наиболее близкая к спине животного) - придаток, называемый мозжечком.
За задним мозгом идет средний мозг, который у млекопитающих включает две пары структур, область из четырех бугров, известную под названием четверохолмия, крыши среднего мозга, или тектума («тектум» в переводе — «крыша»). Нижняя пара структур называется нижним двухолмием, верхняя пара — верхним двухолмием. Кроме этого, средний мозг дает мало оснований для подразделения, по крайней мере в продольном направлении. В сущности это довольно короткий участок человеческого мозга.
Далее идет центральный непарный отдел переднего мозга — промежуточный мозг. Его дорсальные две трети — это таламус. Остальная часть — гипоталамус. (Несколько в стороне от гипоталамуса есть третий отдел промежуточного мозга — субталамус, самую удивительную группу клеток которого — субталамическое ядро — мы упоминали вначале. Включение его усложнило бы рисунок.) Гипоталамус характеризуется наличием железистого придатка, называемого гипофизарным комплексом. Он также продолжается в переднем направлении, переходя в перегородку — структуру, которую, несмотря на ее расположение, лучше всего классифицировать как принадлежащую к промежуточному мозгу. Остальная часть переднего мозга — конечный мозг, мозговые полушария. В мозгу млекопитающих это наибольшая часть, намного превосходящая другие, и у многих видов млекопитающих оболочка полушарий — мозговой плащ, или кора большого мозга, — испещрена извилинами (gyri) и бороздами (sulci). У основания каждого полушария имеется простирающаяся вперед структура, которая полностью состоит из серого вещества (коры), хотя это и кора с очень примитивной клеточной структурой. Ее вздутый передний конец — обонятельная луковица, а ее ножка — обонятельный стебель; только та часть коры, которая лежит непосредственно под остальной частью полушария, является настоящей обонятельной корой. Вторая большая часть коры мозга у млекопитающих находится там, где мозговой плащ подворачивается и образует сложные складки, которые в поперечном сечении напоминают орнамент в стиле рококо. Эта замечательная структура известна под названием гиппокампа.
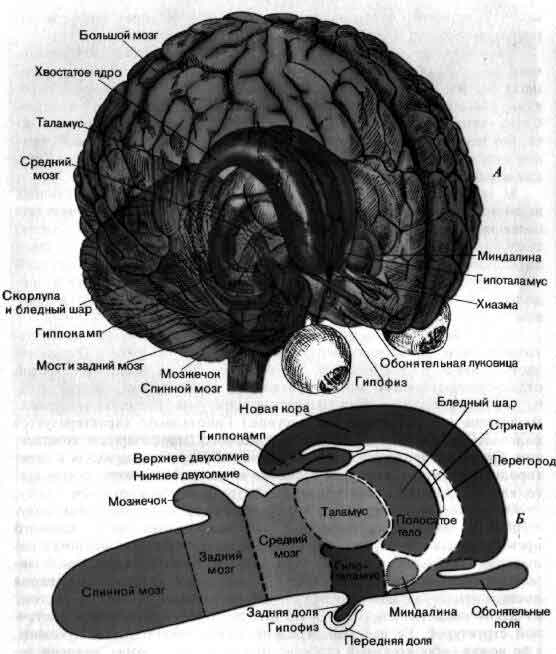
Головной и спинной мозг человека и других млекопитающих можно подразделить на отделы, исходя из данных анатомии, эмбриологии и клеточной организации. А. Мозг человека изображен таким образом, что его внутренние структуры просматриваются сквозь «прозрачные» наружные слои мозжечка. Б. Обобщенная схема мозга млекопитающего; эта схема условно принята для всех остальных рисунков данной статьи. Соответствующие структуры на обоих рисунках окрашены одинаково. Обычно принято делить мозг на передний, средний и задний. Задний мозг включает мозжечок. К среднему мозгу относятся бугорки четверохолмия - нижнее и верхнее двухолмия. Строение переднего мозга более сложно. Его наружная часть - полушарие, поверхность которого - складчатый пласт мозговой коры, включающей гиппокамп, новую кору и обонятельные поля. Внутри полушария располагаются миндалина и полосатое тело; последнее подразделяется на бледный шар и стриатум, включающий хвостатое ядро и чечевицеобразное ядро. Остальная часть переднего мозга — промежуточный мозг: верхние его две трети составляет таламус (который имеет множество подразделении), а нижнюю треть - гипоталамус (который соединяется с гипофизарным комплексом).
После выделения перечисленных частей остается еще одна часть коры большого мозга млекопитающих, которая имеет большую протяженность и чрезвычайно сложное строение; у человека и других приматов она, согласно оценкам, содержит не менее 70 процентов всех нейронов центральной нервной системы. Это новая кора — самая поздняя кора в процессе эволюции. Мы приобрели ее благодаря расхождению путей эволюции: после рептилий одна линия животных развивалась, усовершенствуя прежний тип, и дошла до птиц, а другая — более «авантюрная» - приобрела новую кору и дошла до млекопитающих. Таким образом, с чисто филогенетической точки зрения птицы воплощают в себе логическое завершение традиционного развития мозга, а млекопитающие представляют собой отклонение, поскольку они не имеют в своей родословной птиц. В результате одного из многочисленных разветвлений эволюции млекопитающих появились приматы - отряд, в котором новая кора достигает своего максимального развития. Мы, человеческие существа, пожинаем все плоды этого развития, возможно, включая и психопатологию.
В глубине каждого полушария большого мозга млекопитающих имеется несколько скоплений серого вещества. Одно из них — это миндалина, которая лежит под обонятельной корой. Другое - полосатое тело, находящееся в самой середине полушария. Это тело в свою очередь подразделяется на две части, которые различаются по клеточному составу. Первая из них — это внутренняя зона, называемая палеостриатумом, или бледным шаром. Вторая зона - наружная; она известна под названием стриатум.
Вернемся теперь к схеме связей в центральной нервной системе млекопитающих. Начнем с идентификации сенсорных нейронов, типа тех, которые Паркер нашел в эпителиальном слое медуз. Однако у позвоночных расположение сенсорных нейронов совсем иное. Известен всего один пример, где сенсорный нейрон является также рецептором на поверхности тела: только обонятельные эпителиальные клетки в слизистой крыши носа контактируют с внешней средой. Все другие сенсорные нейроны в теле позвоночных надежно упрятаны под поверхность и находятся в ганглиях, располагающихся вдоль спинного мозга по всей его длине, или в аналогичных ганглиях, лежащих в стороне от мозга. (У позвоночных термин «ганглий» сохранен для скоплений нейронов вне центральной нервной системы.) У каждого сенсорного нейрона есть аксон, который делится на две части: одна часть идет в центральную нервную систему, а другая иннервирует периферические структуры.
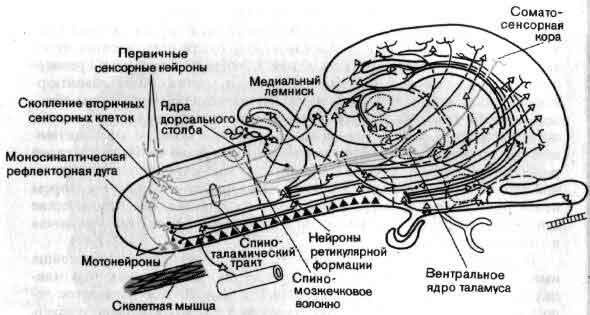
Соматосенсорная информация, например сообщения о кожных ощущениях, передается по нескольким путям спинного мозга. В левой части схемы показана пара репрезентативных сенсорных нейронов, доставляющих сигналы от периферических сенсорных рецепторов в спинной мозг. Отсюда один путь сразу же ответвляется к мотонейронам (сплошные треугольники), волокна которых идут за пределы мозга к скелетным мышцам. Все другие пути сначала идут в скопления вторичных сенсорных нейронов, располагающиеся либо в той же самой области спинного мозга, либо в его верхней части, в ядрах дорсального столба. Путь, называемый медиальным лемниском, поднимается вверх от ядер дорсального столба к вентральному ядру таламуса, которое в свою очередь посылает волокна в соматосенсорную зону коры. Второй путь, называемый спино-таламическим трактом, поднимается к переднему мозгу из скоплений вторичных сенсорных клеток по всей длине спинного мозга, по ходу раздавая в стороны часть своих волокон. Малая доля волокон спино-таламического тракта в конечном счете достигает вентрального ядра. Скопления вторичных сенсорных клеток также посылают свои волокна в мозжечок.
На приведенном здесь рисунке, одна из таких клеток — назовем ее первичным сенсорным нейроном — посылает свой аксон в спинной мозг, доставляя туда сенсорные сообщения о таких событиях, как прикосновение к коже, движение сустава или сокращение мышцы. Эти сообщения не сразу поступают на мотонейроны; свои первые синаптические контакты сенсорный нейрон устанавливает с так называемыми вставочными нейронами.
Однако имеется одно исключение. Это моносинаптическая рефлекторная дуга, образуемая боковой ветвью первичного сенсорного волокна, идущей «напролом» и устанавливающей прямой синаптический контакт с мотонейроном. Сначала это кажется обескураживающим: всего лишь несколькими абзацами выше мы отмечали, что после самых ранних ступеней эволюции нервной системы мотонейроны больше не утруждают себя обработкой сырых данных. Мы предположили, что вместо этого они теперь получают краткий итог обработки информации («дайджест») нейронами великой промежуточной сети. Следовательно, моносинаптическая дуга может рассматриваться как очень примитивный тип нервной сети; с другой стороны, ее можно считать относительно новой: по-видимому, ее вновь открыли только наземные животные. В конце концов, воздух и земля — самые суровые среды; для горного козла один неверный шаг может оказаться роковым. В противоположность этому рыба может без всякого риска для себя сделать любое количество аналогичных ошибочных движений. Рыбы прекрасно уравновешены в воде, и сила тяжести для них далеко не так обременительна и враждебна. Таким образом, именно наземное, а не водное существование, по-видимому, требует наличия высоконадежных рефлекторных систем для удержания равновесия и, в частности, наличия пути, по которому мышца может сигнализировать соответствующим мотонейронам (и только им), что она чрезмерно растянута силой тяжести.
Моносинаптические рефлекторные дуги никогда не обнаруживаются за пределами таксах корректирующих механизмов. Так что короткие цепи между сенсорным входом и моторным выходом оказываются представленными в ничтожно малом числе. Подавляющее большинство первичных сенсорных волокон млекопитающих входит в великую промежуточную сеть и синаптически контактирует с клетками, составляющими группу, которую мы будем называть группой вторичных сенсорных клеток: это нейроны, стоящие первыми в цепи приема первичных сенсорных сигналов. Оттуда многие пути более или менее прямо направляются к мотонейронам. Все они в совокупности могут быть названы локальным рефлекторным путем, если не упускать из виду, что слово «локальный» может ввести в заблуждение, так как есть несколько рефлексов, которые вовлекают в работу весь спинной мозг по всей его длине, но тем не менее относятся к локальным, поскольку не выходят за его пределы. Первое звено в локальном рефлекторном пути — это некоторый нейрон из группы вторичных сенсорных клеток. Многие из таких клеток сами не образуют контактов с мотонейронами; вместо этого они синаптически переключаются на некоторые другие нейроны великой промежуточной сети, и только уже эти последние нейроны, наконец, замыкают дугу.
Другие пути состоят из аксонов, не идущих к мотонейронам. Возьмем мозжечковые каналы: из скоплений вторичных сенсорных нейронов заднего и спинного мозга многие аксоны восходят прямо к мозжечку. Показанный на рисунке на стр. 94 аксон, который идет таким образом, берет свое начало в группе вторичных сенсорных клеток спинного мозга и потому называется спино-мозжечковым волокном. (Слова «аксон» и «волокно» в нейроанатомии используются как синонимы.)
Третий путь — это лемнисковый путь. Слово «лемниск» в переводе с латинского — лента (петля); здесь им обозначаются пучки волокон, которые берут начало в группах вторичных сенсорных клеток и поднимаются к переднему мозгу, в частности к таламусу. На рисунке (стр. 94) показано, как один такой пучок идет по центру спинного мозга. В действительности он проходит латеральнее: упрощенную иллюстративную схему трудно сделать топографически точной. Этот пучок назван спино-таламическим трактом, хотя только одно из трех его репрезентативных волокон изображено как оканчивающееся в таламусе. Два других волокна сопровождают его на некотором расстоянии, а затем, так сказать, «совершают непредвиденную посадку»: на рисунке показано, что оба оканчиваются на нейронах заднего мозга, хотя с таким же успехом они могли бы окончиться несколько далее по ходу — в среднем мозгу. Важно то, что из волокон спино-таламического тракта лишь небольшая доля действительно достигает таламуса. Тем не менее тракт назван в честь этого преуспевшего меньшинства, которое оканчивается в специфической части таламуса — вентральном ядре. Здесь волокна образуют синапсы с таламическими нейронами, аксоны которых идут, не прерываясь, в специализированное поле новой коры, называемое соматосенсорной корой.
Заметим, что на пути от первичного сенсорного нейрона до новой коры в данном случае насчитывается только два синаптических переключения. Первое — в спинном мозгу между первичным сенсорным волокном и нейроном из группы вторичных сенсорных клеток. Второе переключение — в промежуточном мозгу, между волокнами лемниска и нейронами вентрального ядра таламуса. Однако то, что происходит в новой коре, можно назвать синаптическим катаклизмом. В новой коре в реакцию на пришедший сигнал уже на первом этапе вовлекаются сотни, а то и тысячи нейронов. А действуя через синаптические связи, эти первые нейроны, возбужденные сигналом, вовлекают бесчисленное множество следующих.
Двухсинаптическая сенсорная линия проведения, идущая в новую кору, может быть названа «сквозной линией», так как два синапса — это, по-видимому, минимум для такого рода систем. Она также может быть названа закрытой, или маркированной линией, поскольку, как правило, сенсорные пути с минимальным числом переключений сохраняют топографию той сенсорной периферии, от которой они идут. Так, кончик пальца способен различить два отдельных стимула, когда к нему прикасаются ножками измерительного циркуля в точках, находящихся всего лишь в двух или трех миллиметрах друг от друга. Это называется разрешающей способностью. Наличие такой способности означает, что каждая ножка циркуля стимулирует проводящие пути, которые достаточно независимы для того, чтобы обеспечить это, если можно так сказать, сенсорное разрешение. Какая-нибудь клетка соматосенсорной коры, если ее «допросить» с помощью микроэлектрода, может «признаться» в том, что ее единственный интерес составляет квадратный миллиметр кожи на указательном пальце. Одна из ее ближайших соседок может контролировать смежный квадратный миллиметр, и так далее. Благодаря этому топография поверхности тела будет верно воспроизведена.
Проводящие пути, диаметрально противоположные маркированным,- это такие, по которым проводятся топографически перепуганные сообщения от данного сенсориума или даже от нескольких различных сенсориумов. Такая любопытная организация действительно существует: один из спино-таламических «отщепенцев», показанных на рисунке (стр. 94), синаптически переключается на нейрон заднего мозга, аксон которого продолжает путь в таламус. Однако на уровне этого «лишнего» переключения линия принимает сообщения не только от спино-таламического волокна, но и от слуховой системы.
Как же таламус может понять, что случилось, когда импульсы поступают по этой системе путей? Нейроны заднего мозга называют мультимодальными, или неспецифическими, а рассмотренный выше проводящий путь можно назвать открытым: везде, где есть синаптические переключения, линия открыта для входа сигналов от других нейронов. Огромное большинство нейронов в глубине заднего и среднего мозга относится к нейронам именно такой любопытной неспецифической природы. Кажется, что они сидят, раскинув свои дендриты, словно длинные руки, на расстояния в несколько миллиметров, как бы стараясь поймать сигналы любого рода. Эти нейроны типичны для так называемой ретикулярной формации, в которой лишь сравнительна малое число клеточных скоплений получает гомогенные входные сигналы.
Если бы эту ситуацию описали инженеру-электронщику, он, вероятно, отнесся бы к ней неодобрительно; он сказал бы, что от такой схемы нечего надеяться получить что-либо, кроме шума. Однако такая ситуация преобладает в мозгу всех позвоночных, включая человека. Так что ее существование, по-видимому, вызвано необходимостью удовлетворять какие-то специфические потребности. На данный момент можно сказать, что в функции ретикулярной формации входит обеспечение фона общей активации в центральной нервной системе и что ретикулярная формация воплощает в себе некий механизм регуляции состояний активности по всей центральной нервной системе. Некоторые из этих состояний подвержены суточным колебаниям — скажем сон и бодрствование. В каждом из этих состояний можно выделить большое количество «полутонов» возбуждения и расслабления. Все они связаны с тем или иным характером активности ретикулярной формации.
Электронщика наверняка больше бы устроил второй соматосенсорный лемниск, выходящий из спинного мозга. Это медиальный лемниск. Он намного более компактно организован: почти все его волокна - маркированные линии, восходящие прямо к центральному ядру таламуса из двух скоплений вторичных сенсорных клеток на переходе между спинным и задним мозгом, которые называются ядрами дорсального столба. Наш инженер не удивился бы, узнав, что медиальный лемниск обеспечивает лучшее разрешение двух точек, чем спино-таламический тракт.
А что сказать о других сенсориумах? Маленький орган, который для упрощения чертежа изображен на рисунке (стр. 99) возле мозжечка, - это орган слуха. Внутри него, в высоко специализированном эпителиальном комплексе, называемом кортиевым органом, обнаружены клетки, которые имеют по одной-единственной ресничке. Они иннервируются первичными сенсорными нейронами, у которых направляющиеся в центр отростки оканчиваются на нейронах кохлеарных ядер - скоплений вторичных сенсорных нейронов в заднем мозге, специализированных для приема и обработки сигналов исключительно от слухового сенсориума. На рисунке показаны только два нейрона; в действительности их десятки тысяч. В кохлеарных ядрах берут свое начало волокна латерального лемниска, восходящие к таламусу. Ни одно из них не выходит за пределы нижнего двухолмия. На этом пути при переключении в среднем мозгу начинаются аксоны, которые добираются до таламуса, где они заканчиваются в медиальном коленчатом теле. (На рисунке не показано несколько других слуховых «станций», по-видимому, менее обязательных и связанных с самим латеральным лемниском.) Нейроны медиального коленчатого тела в свою очередь проецируются (посылают свои аксоны) в специальную область новой коры, называемую слуховой корой.
Сравним это со зрительной системой. Множество нейронов сетчатки обрабатывает выходные сигналы фоторецепторного аппарата глаза. Аксоны определенной части этих клеток сначала объединяются в зрительный нерв. Затем происходит перераспределение аксонов по двум путям, в процессе которого аксоны, вышедшие из назальной (носовой) половины сетчатки одного глаза, пересекают срединную плоскость головы, чтобы объединиться с аксонами, которые вышли из темпоральной (височной) половины сетчатки другого глаза. В результате образуются зрительные тракты. Аксоны, составляющие зрительный тракт, распределяются между двумя большими конечными областями. Одна из них — это верхнее двухолмие; однако у всех приматов более важной областью, по крайней мере если судить по количеству аксонов, является латеральное коленчатое тело таламуса. Нейроны этого ядра проецируются в свою очередь в новую кору, а именно - в определенную зону у заднего полюса полушария мозга, известную под названием зрительной коры.
Отметим, что в противоположность слуховым волокнам, ни одно из которых не может достичь своего таламического ядра без синаптических переключений, большая часть зрительных волокон (у приматов) это делает. Следовало бы добавить, однако, что многие из нейронов верхнего двухолмия, получающих зрительные волокна, посылают свои собственные аксоны в таламус, но не в латеральное коленчатое тело, а в латеральное заднее ядро. Нейроны этого последнего клеточного скопления в свою очередь проецируются в новую кору, однако не в ту область, в которую идут аксоны из латерального коленчатого тела, а в соседнюю, которая отличается от первичной зрительной коры. По-видимому, зрительная система имеет два пути, восходящих к зрительной коре.
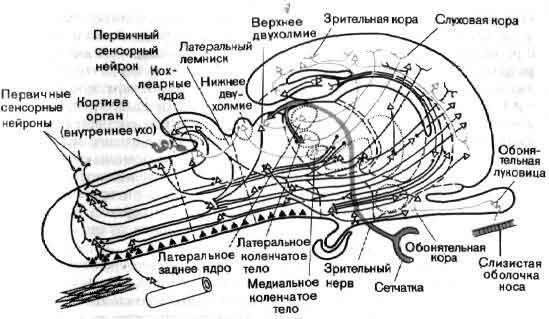
Три сенсориума (слуховой, зрительный и обонятельный) посылают свою информацию в кору различными путями. Слуховые пути, прежде чем они достигнут слуховой зоны коры больших полушарий, последовательно проходят через кохлеарные ядра заднего мозга, нижнее двухолмие среднего мозга и медиальное коленчатое тело таламуса. Зрительные пути начинаются в сетчатке (которая в действительности является частью мозга) и затем расходятся по двум различным каналам: один идет через латеральное коленчатое тело таламуса в зрительную кору, другой направляется через верхнее двухолмие среднего мозга в таламус, а затем — в область коры, расположенную около первичной зрительной зоны. В обонятельной системе рецепторные нейроны слизистой оболочки носа проецируются, минуя таламус, в обонятельную луковицу, которая является частью коры больших полушарий, а обонятельная луковица в свою очередь проецируется в обонятельную кору.
Обонятельная система нарушает все законы, которым, казалось бы, подчиняется структура других сенсорных механизмов. Это, как мы отмечали, единственная из известных систем, в которой сенсорные нейроны лежат на поверхности. Там нет посредника - передающего элемента, как, скажем, в кортиевом органе; обонятельные эпителиальные клетки сами воспринимают внешние воздействия. Отходящие от таких нейронов очень тонкие аксоны проецируются в обонятельную луковицу, нейроны которой дают начало аксонам, заканчивающимся синаптическими контактами на клетках обонятельной коры.
Мы только что проследили волокна четырех сенсориумов - соматического, слухового, зрительного и обонятельного. Теперь уже начинают выявляться некоторые существенные моменты. Во-первых, таламус предстает как узловая станция, конечный контрольно-пропускной пункт, дающий разрешение на вход в высшие инстанции мозга сигналам от всех сенсориумов (за исключением, по-видимому, обонятельного). Каждое такое переключение хочется сравнить с передачей эстафеты или с ретрансляционной станцией, однако то, что происходит в местах разрыва нейронной цепи, может быть намного сложнее того, что происходит в эстафетном беге, где каждый бегун передает эстафетную палочку следующему и на конечный пункт маршрута эта палочка прибывает в неизменном виде. В центральной нервной системе «эстафета» совершенно иного рода. На каждом синаптическом переключении сенсорного пути входные сигналы трансформируются: код, с помощью которого записано прибывшее сообщение, коренным образом меняется. Можно предположить, что исходные данные не могут быть интерпретированы на высших уровнях; по-видимому, необходим их перевод на другой язык, и о синаптических ретрансляционных станциях лучше говорить как о станциях переработки данных.
Затем можно сделать заключение, что кора является конечной станцией сенсорных проводящих путей. Нейроанатом бывает в высшей степени удовлетворен, когда ему удается проследить зрительные пути, скажем, от сетчатки до латерального коленчатого тела и оттуда — до зрительной коры. Трудность любого дальнейшего прослеживания заключается в сложности коры мозга, содержащей в себе 70 процентов всех нейронов центральной нервной системы человека. Что они делают со своими входными сигналами? Тут уместно привести следующие два наблюдения.
Во-первых, таламо-кортикальные проекции имеют реципрокный характер: зрительная кора проецируется обратно на латеральное коленчатое тело, из которого в нее поступают входные сигналы; слуховая кора проецируется обратно на медиальное коленчатое тело, а соматосенсорная кора - на вентральное ядро. Такие возвратные связи, без сомнения, говорят о том, что функциональное состояние коры может влиять на ту процедуру, с помощью которой сенсорные станции таламуса «просеивают» направляемый в кору поток информации.
Во-вторых, зрительная, слуховая и соматосенсорная зоны коры воплощают в себе только первые этапы обработки сенсорной информации. Из этих первичных сенсорных полей выходят волокна, синаптически воздействующие на смежные области, которые без оговорки нельзя назвать сенсорными: они представляют, так сказать, следующий этап обработки входных сигналов. А из этих областей выходят волокна, которые оканчиваются в областях, еще более далеких от первичных сенсорных полей. Области новой коры, более или менее удаленные от первичных полей, называются ассоциативными зонами, и у человека они занимают подавляющую часть поверхности коры; зрительная, слуховая и соматосенсорная зоны вместе занимают только около четверти всей ее площади. По всей вероятности, в ассоциативных зонах информация подвергается дальнейшей, более сложной переработке. Так, есть места, где слуховые и зрительные сигналы сходятся. Теперь известно, что в процесс обработки нервных сигналов в новой коре обычно последовательно вовлекается целый ряд ассоциативных областей и что местом назначения выходящей информации, по-видимому, неизменно являются либо гиппокамп, либо миндалина, либо и то и другое.
В 1870 г. Г. Фрич (G. Fritsch) и Э. Гитциг (Е. Hitzig) опубликовали сообщение о том, что электрический ток минимальной силы, пропускаемый через область новой коры, расположенную фронтально по отношению к центральной борозде, может вызвать подергивание скелетных (но не висцеральных) мышц на стороне тела, контралатеральной месту стимуляции. Так, часто приходит в движение рука или нога. Это открытие, возможно, являющееся первым указанием на наличие в коре большого мозга функциональных подразделений, возбудило стойкий интерес к организации тех частей мозга, которые связаны с эффекторными (или моторными) функциями. В конце концов была открыта моторная кора — ограниченная область на самом высоком уровне мозга, которая явно причастна к движениям тела. Казалось, что теперь чисто моторную функцию можно будет, так сказать, вычленить из всего головного и спинного мозга.
Отсюда начинаются поиски «моторной системы». Этот неопределенный термин включает не только мотонейроны, управляющие скелетной мускулатурой, но и нервные пути, которые сходятся на мотонейронах. Поиски эти длятся и по сей день, и резонно спросить, могут ли они когда-нибудь быть завершены? Рассмотрим поле 19 — полоску новой коры, отличающуюся по цитоархитектонике от соседних зон и расположенную недалеко от зрительной коры. Когда поле 19 стимулируют электрически у какого-нибудь экспериментального животного, оба глаза этого животного синхронно поворачиваются к противоположной стороне, т.е. взор отворачивается от раздражаемой током половины. В связи с этим появляется искушение назвать поле 19 моторной зоной. Однако сделать это — значит поступить произвольно, поскольку с другой точки зрения поле 19 — сенсорное: известно, что оно повторно обрабатывает информацию, которая прошла через зрительную кору. Сходный пример можно подобрать и для слухового сенсориума: вблизи соответствующей слуховой коры имеется область, называемая полем 22, электрическая стимуляция которой тоже заставляет животное поворачивать глаза к контралатеральной стороне. И при этом поле 22 состоит со слуховой корой в синаптических отношениях, во многом напоминающих отношения между полем 19 и зрительной корой.
Урок из всего этого такой: между сенсорной и моторной структурами мозга нельзя провести разделяющую грань. Представляя ситуацию с другой стороны, можно сказать, что все нервные структуры занимаются программированием и регулированием поведения организма. По существу, это и есть функция нервной системы и эволюция разума содействовала ее развитию. Правда, некоторые структуры имеют в пределах великой промежуточной сети такое расположение, которое склоняет к определению их как сенсорных; примером тому служит латеральное коленчатое тело таламуса. А к другим структурам, расположенным на расстоянии не слишком большого числа синаптических переключений от мотонейронов, есть искушение приклеить ярлык «моторные». Это, впрочем, единственный способ разумного использования данных терминов. Соответственно при исследовании моторных функций центральной нервной системы, может быть, лучше всего начинать с уровня мотонейронов, которые однозначно являются частью моторной системы согласно любому определению, а затем пытаться проследить в мозгу те пути, которые воздействуют на них. Нужно только отдавать себе отчет в том, что применение этой стратегии означает движение вверх по течению, против превалирующего направления нервных потоков.
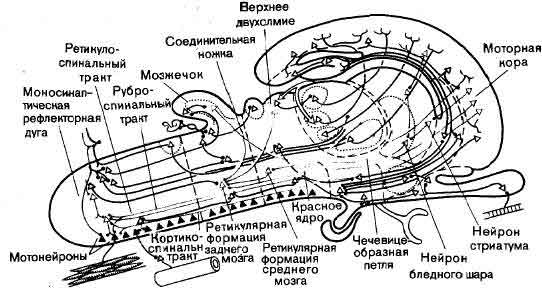
Мотонейроны головного и спинного мозга (сплошные треугольники) получают информацию из многих конвергирующих к ним каналов. Здесь показаны мотонейроны, на вход которых поступают сигналы от первичных сенсорных нейронов, из скоплений вторичных сенсорных нейронов в спинном мозгу, из ретикулярной формации ствола мозга, из красного ядра среднего мозга и из моторной коры переднего мозга. Красное ядро и ретикулярная формация сами получают сигналы из различных источников. Один из примеров конвергенции сигналов на мотонейронах является особенно впечатляющим: вся новая кора, включая слуховую, зрительную, соматосенсорную, а также моторные и другие поля, направляет пучки волокон в полосатое тело. Это ядро в свою очередь посылает массивные пучки волокон в ретикулярную формацию, которая, в конце концов, воздействует на мотонейроны. Второй путь, идущий из полосатого тела, играет роль петли обратной связи: составляющие его волокна направляются в область таламуса, которая проецируется обратно, опять же в моторную кору.
Первый шаг «вверх» от мотонейронов, как правило, короток, поскольку самые сильные воздействия на типичный мотонейрон чаще всего исходят от ансамблей клеток, которые обычно меньше по размеру и располагаются поблизости. Давайте назовем совокупность всех мотонейронов и управляющих ими нейронных ансамблей «низшей моторной системой» и расчленим эту систему на функциональные субъединицы, назвав каждую «локальным моторным аппаратом», соответствующим определенной части тела: руке, ноге, глазу и т.д. Каждый локальный моторный аппарат есть, по-видимому, своего рода картотека, в которой хранятся программы, представляющие возможные движения определенной части тела. Мозг посылает сигнал в эту картотеку, используя свою систему нисходящих волокон, и выбирает подходящую программу.
Но что в таком случае служит источником сигналов, посылаемых по нисходящим волокнам? Что именно оказывает влияние на локальный моторный аппарат? Мотонейроны располагаются внутри спинного, заднего и среднего мозга; в переднем мозгу нет ни одного. Здесь мы можем рассмотреть только проекции, которые сходятся на спинном мозгу. Они начинаются на всех уровнях центральной нервной системы. Внутри самого спинного мозга многие из них берут свое начало в группах вторичных сенсорных клеток, или даже, как в случае моносинаптических рефлекторных дуг, образуются коллатералями определенных первичных сенсорных волокон. В пределах заднего мозга пути исходят главным образом от внутренних двух третей заднемоэговой ретикулярной формации — области, которую называют крупноклеточной ретикулярной формацией, так как ее содержимое составляют крупные и очень крупные тела нейронов. В среднем мозгу пути исходят из верхнего двухолмия, а также из большой клеточной массы, называемой красным ядром. Вообще говоря, все эти три системы нисходящих волокон, идущих в спинной мозг (а именно: ретикуло-спинальная, текто-спинальная и рубро-спинальная), должны рассматриваться как несущие сообщения, или, если хотите, команды, которые могли зарождаться в обширных областях мозга. Верхнее двухолмие получает входные сигналы не только но зрительному нерву, но также из обширных областей коры, включая зрительную и многие другие. В красное ядро пути идут главным образом из мозжечка и моторной коры.
Что же касается ретикулярной формации, то она особенно замечательна тем, что является местом, куда сходится информация самого различного происхождения. Мы уже рассуждали об этом, когда говорили о восходящих системах; при рассмотрении нисходящих систем это тоже существенно. Нейрон, представляющий данное свойство, показан на ; за основу этой модели взяты нейроны, электрическую активность которых регистрировал Дж. Моруцци (G. Moruzzi) из Пизанского университета, а также некоторые другие исследователи. Данный нейрон лежит в ретикулярной формации заднего мозга, и, по-видимому, он будет реагировать на поступление сигналов от группы вторичных сенсорных клеток спинного мозга. Однако вспышка света тоже способна активировать этот нейрон, поскольку сообщение об этом событии может успешно достичь ретикулярной формации по нисходящему пути из верхнего двухолмия. Кроме того, эта клетка реагирует и на сигналы из мозжечка, новой коры или ретикулярной формации среднего мозга.
Короче, на этой клетке конвергирует большое число гетерогенных сигналов. Очевидно, ретикулярная формация должна интегрировать эти весьма разнообразные нервные сигналы, поступающие к ней по восходящим и нисходящим путям, а затем она может отправлять импульсы по ретикуло-спинальным волокнам, которые оканчиваются на промежуточных нейронах спинного мозга или даже, хотя и не часто, прямо на мотонейронах. Возможно, тут читатель опять услышит вопль электронщика: с инженерной точки зрения ретикулярная формация — бессмысленное образование.
Теперь нам остается наложить на энцефало-спинальные системы заднего и среднего мозга те нисходящие системы, которые берут свое начало в переднем мозгу. Прежде всего практически все области новой коры проецируются в стриатум — наружную часть полосатого тела. В общей схеме связей сохраняется топографическое соответствие: соматосенсорная кора проецируется в область стриатума, отличную от той, куда приходятся зрительная проекция, слуховая проекция или проекции ассоциативных областей и моторной коры. Из стриатума масса волокон идет в бледный шар — внутренний отдел полосатого тела. В бледном шаре намного меньше нейронов, чем в стриатуме, так что эту систему можно рассматривать как своего рода воронку.
Из бледного шара путь продолжается вниз, образуя пучок волокон, называемый чечевицеобразной петлей; здесь, однако, следует учесть любопытное исключение: большая часть волокон этой петли поворачивает назад в составе того же пучка и затем входит в верхнюю часть вентрального ядра. Мы отмечали, что в это клеточное ядро таламуса входят два больших соматосенсорных пути — медиальный лемниск и спино-таламический тракт и что оно проецируется на соматосенсорную кору. Однако только задняя часть вентрального ядра является промежуточной соматосенсорной станцией. В переднюю часть этого же клеточного скопления входят две мощные системы волокон: чечевицеобразная петля и восходящая проекция мозжечка — соединительная ножка. Эта часть вентрального ядра тоже проецируется в новую кору, но не в сенсорную, а в моторную зону.
Патологические процессы, сопровождающиеся разрушением этой любопытной цепи, которая характеризуется наличием петель, вызывают серьезные двигательные расстройства. Одна из таких патологий касается пути в стриатум, который идет не из коры больших полушарий. Он идет из клеточной массы среднего мозга, нейроны которой пигментированы; у человека они черные даже на неокрашенных препаратах. По этой причине данная клеточная группа стала известна с конце XVIII в. как substantia nigra, т.е. черное вещество. Существенная потеря этих пигментированных нейронов вызывает двигательное расстройство, известное под названием паркинсонизма. Это расстройство характеризуется ригидностью мышц, которая сильно затрудняет движения, и диагностируется по ряду симптомов, в том числе по маскообразному лицу. Кроме того, наблюдается также специфический (низкочастотный и почти вращательный) тремор рук. Однако первая жалоба больного обычно состоит в том, что ему трудно начать движение, которое он хочет сделать. Скажем, он собирается поправить какую-то деталь своего костюма, но ему почему-то трудно начать.
Таким образом, можно считать, что полосатое тело существенно влияет на двигательную сферу. В более широком плане его можно рассматривать как принадлежащее к большому числу тех мозговых структур, выходные пути которых, по всей видимости, направлены к мотонейронам. Любопытно, что при этом полосатое тело не может непосредственно воздействовать на такие нейроны или хотя бы прямым путем влиять на те нейронные ансамбли, которые играют при мотонейронах роль привратников. Мы только что видели, что некоторая часть его выходного тракта, чечевицеобразной петли, поворачивает вверх и входит в вентральное ядро таламуса. Оставшаяся часть этой петли продолжает после этого поворота идти вниз, но далее каудальной границы среднего мозга не идет; на приведенном рисунке () один нейрон символизирует группу из нескольких тысяч нейронов, составляющих ядро ножки моста. Это часть ретикулярной формации среднего мозга. Начиная отсюда, нисходящие пути становятся неопределенными. Ретикулярная формация - это область, представляющая почти непреодолимые трудности для анатомического и функционального анализа.
Волокна, проецирующие новую кору на стриатум, ни в коем случае не являются единственными кортикофугальными волокнами. Как мы отмечали, некоторые из выходных путей новой коры заканчиваются и различных ядрах таламуса, которые в свою очередь проецируются обратно на новую кору. Некоторые кортикофугальные волокна проникают в средний мозг, чтобы закончиться там в верхнем двухолмии, красном ядре и ретикулярной формации среднего мозга. Еще один контингент составляют волокна, идущие от всех частей новой коры и образующие синапсы на клетках варолиева моста - отдела заднего мозга, который в свою очередь проецируется на мозжечок. Остальные кортикофугальные волокна - те, которые проходят за пределами моста,- берут свое начало главным образом в моторной коре. Некоторые из них идут не далее чем до ретикулярной формации заднего мозга; другие достигают всех уровней спинного мозга.
Эти последние волокна, которые составляют кортико-спинальный тракт, заслуживают особого упоминания. Само по себе замечательно уже то, что они проделывают путь от коры до спинного мозга, поскольку волокна, нисходящие из полосатого тела, доходят лишь до среднего мозга. Замечательно также то, что, согласно оценкам, 5 процентов кортико-спинальных волокон синаптически переключается прямо на мотонейроны. Это почти невероятный обходной путь: данные волокна не только входят в спинной мозг, но избегают нейронных ансамблей локальных моторных механизмов. Оказалось, что они «предпочитают» устанавливать синаптические связи с мотонейронами, которые заведуют мышцами конечностей. Несомненное наличие кортико-спинального тракта позволяет объяснить тот факт, что из всех областей коры больших полушарий моторная кора требует самой слабой электрической стимуляции для возбуждения в эксперименте движений тела. Объяснение состоит в том, что из всех областей коры больших полушарий мозга моторная кора удалена от мотонейронов на наименьшее число синаптических переключений.
Моторная кора почти не поддается исследованию с точки зрения противопоставления произвольного и непроизвольного поведения. Рассмотрим обидную ситуацию, знакомую теннисистам. Игрок блестяще отбивает мяч и чувствует себя окрыленным. Но потом он приходит к выводу, что это была просто счастливая случайность: в следующий раз, когда теннисный мяч будет лететь к нему по сходной траектории, он может отбить его плохо. Это правда, что трудное произвольное движение было выполнено успешно, но есть ли в этом заслуга лица, которое его выполнило?
Несмотря на загадку произвольного контроля, субъективный опыт с волевыми усилиями дал название моторной системе, которая иннервирует скелетную мускулатуру: это произвольная, или соматическая, нервная система, в отличие от непроизвольной, или автономной, нервной системы, которая иннервирует железы и гладкие мышцы внутренних органов. Однако последняя терминология не лучше застрахована от неправильного толкования, чем первая. Она имеет дело с термином «автономный», что значит «самоуправляемый». Автономная нервная система вовсе не является самоуправляемой. Ее функции увязаны с произвольными движениями не менее, чем с побуждениями и эмоциями. Короче, ее корни находятся в мозгу: каждому удается время от времени управлять не только сокращениями скелетных мышц, но и отправлением функций некоторыми внутренними органами. Тем не менее термин «автономная» победил во всем мире, говорящем по-английски. В других языках используются другие термины. На немецком говорят о висцеральной нервной системе (das viszerale Nervensystem), на французском — о вегетативной нервной системе (le systeme nerveux vegetatif).
Автономная периферия условно изображена на рисунке () в виде трубчатого полого органа, под которым можно подразумевать кишечный тракт, мочевой пузырь, бронх или артерию; все они, в сущности, трубчатые структуры, толщина которых определяется числом слоев гладких мышц. Моторная иннервация такой мышечной ткани (или железы) включает два нейрона. Первый расположен в центральной нервной системе. Из него выходит очень тонкий аксон, который на периферии синаптически переключается на второй нейрон, часто располагающийся в ганглии. Второй нейрон в свою очередь посылает аксон для образования конечного соединения в висцеральную эффекторную ткань.
В мозгу нейроны, которые специфически воздействуют на активность автономной нервной системы, сконцентрированы в гипоталамусе.
Цепи обратной связи замыкаются на гипоталамус, который регулирует активность желез и гладких мышц (таких, как не поддающиеся произвольному контролю мышцы внутренностей) через автономную нервную систему и гипофизарный комплекс. В гипоталамус приходят пути от гиппокампа и миндалины, которые являются главными компонентами так называемой лимбической системы. Кроме того, в гипоталамус поступают также волокна, исходящие из ретикупярной формации. Другим источником его входных сигналов является лобная кора, располагающаяся в самой передней части полушарий большого мозга. Лимбическая система-место назначения путей, исходящих из коры большого мозга, включая прямые проекционные волокна обонятельной коры.
Доказательства этого очевидны; когда гипоталамус любого животного, а тем паче человека, внезапно разрушается, его обладатель умирает из-за сдвигов в том, что Клод Бернар назвал внутренней средой - этот термин охватывает те тканевые жидкости и функции органов, которые определяют кровяное давление, частоту пульса, частоту дыхания и т.д. В связи с этим нейрохирурги, которые вынуждены оперировать в области таламуса, всегда озабочены тем, чтобы не затронуть эту структуру. И все-таки больные умирают иногда от гипертермии (резкого повышения температуры тела) даже после успешной во всех других отношениях операции на мозге, в которой, казалось бы, были приняты все меры предосторожности, дабы не повредить гипоталамус. С другой стороны, когда массивное поражение гипоталамуса развивается постепенно, например в результате медленно растущей опухоли, острых тяжелых симптомов может вообще не быть. Создается впечатление, что в автономной системе существует командная цепь, или, как Клод Бернар определил это, автоматизм разных уровней: когда гипоталамус медленно теряет свои способности, области мозга ниже гипоталамуса сами могут обеспечить поддержание стабильности внутренней среды, хотя и в узких пределах.
Все это хорошо согласуется с тем, что известно о путях автономной системы. Недавно были обнаружены волокна, идущие без переключений от гипоталамуса до мотонейронов автономной системы, расположенных в сером веществе спинного мозга, но они, по-видимому, составляют ничтожную часть выходящих из гипоталамуса волокон; для проведения нисходящих сигналов гипоталамус не имеет ничего похожего на кортико-спинальный тракт. Напротив, оказывается, что значительная часть волокон гипоталамуса идет не далее среднего мозга, где эстафету принимает ретикулярная формация. В действительности, для путей, нисходящих к автономным мотонейронам, типично прерывание на многих уровнях. При каждом таком прерывании в нисходящую линию проведения могут входить разнообразные дополнительные инструкции. Было бы разумно, если бы так было на самом деле. Жизнь зависит от иннервации внутренних органов; в каком-то смысле все остальное - биологическая роскошь. И жизненно важные системы должны быть построены по такому принципу, что никакое одиночное влияние не должно сильно сказываться на их работе. В действительности, конвергенция информации на мотонейронах для автономной нервной системы может быть столь же характерной, сколь и для соматической.
Пока мы говорили о нисходящих нервных влияниях, которые в конечном счете достигают эффекторных тканей внутренних органов. А чем вызываются эти влияния? Или конкретнее: откуда и какие пути идут в гипоталамус? На рисунке (стр. 107) показан входной канал, который начинается в клетке ретикулярной формации среднего мозга — клетке, получающей свои входные сигналы от волокон спино-таламического тракта. Можно предположить, что при помощи этого пути таламус способен контролировать состояние внутренней среды. За пределами этого поиски входов в гипоталамус заводят далеко в область тканей мозга, причастных к эмоциям и побуждениям, область, в которой, скажем, эпилептические разряды могут, среди прочих симптомов, вызывать изменения в настроении, иногда до душевной боли или беспричинного страха. Это не должно нас удивлять. В конце концов эмоции и побуждения находят явное выражение в висцеральных и эндокринных изменениях.
Значит, можно почти не сомневаться в том, что основные влияния на гипоталамус со стороны полушарий большого мозга исходят из гиппокампа и миндалины. Они делят свою «власть» с малым числом других частей полушарий. По этой причине общая ссылка на гиппокамп и миндалину оправдана: они являются главными компонентами того, что называют лимбической системой. Обратите внимание на показанную на рисунке двухканальную систему волокон, которая закругляется по краю новой коры и идет из гиппокампа в гипоталамус. Этот пучок образует свод. Он проходит вдоль свободного края мозгового плаща. И у кошки, и у обезьяны примерно две трети волокон этого пучка при выходе из гиппокампа направляются прямым путем в гипоталамус. Оставшаяся треть устанавливает синаптические связи в перегородке, из которой, как показано на рисунке, пути направляются также в гипоталамус.
Мы уже указывали, что гиппокамп является станцией назначения для многоступенчатых проекций, которые размещены на обширной поверхности новой коры. Таким образом, при прослеживании путей управления висцеральной моторикой, как и в случае соматической моторики, оказывается, что по ходу прослеживания, если оно производится против течения, т. е. против направления передачи импульсов, приходится подключать к делу все большую часть великой промежуточной сети. Конечно, тут есть и отличие. Из данной области новой коры, скажем из первичной зрительной зоны, путь в гиппокамп может проходить с переключениями через ряд промежуточных неокортикальных полей. Концом неокортикального пути является энторинальная область коры большого мозга, смежная с гиппокампом и промежуточная по структуре между ним и новой корой. Выходящий отсюда последний отрезок завершает путь волокон в гиппокамп. В противоположность этому, путь в стриатум из любого поля новой коры является непрерывающимся.
Рассмотрим далее миндалину. Хотя по цитоархитектонике она сильно отличается от гиппокампа, значительная часть ее выходных волокон тоже направляется в гипоталамус. Для гипоталамуса миндалина - это место, куда сходятся волокна из областей новой коры, синаптически удаленных от каких-либо первичных сенсорных полей. Однако она также является и адресатом волокон, которые начинаются в обонятельной коре; действительно, сюда проецируется энторинальная область. Кроме того, часть миндалины получает волокна от обонятельной луковицы. Таким образом, в случае обоняния передача сенсорных сигналов в лимбическую систему осуществляется поразительно прямым путем. Почему бы это могло быть? Почему обонятельная система находится в привилегированном положении по отношению к другим сенсорным системам?
Один из возможных ответов заключается в следующем: вполне вероятно, что обонятельный сенсориум представляет собой самое раннее образование в эволюции способности к сенсорному восприятию на расстоянии; быть может, это самая ранняя система, с помощью которой свободно странствующий организм мог находить следы источников пищи и узнавать членов своего собственного вида и представителей других. Возможно, что обонятельная система, появившись раньше других, установила ряд относительно прямых связей. Второй предположительный ответ, не отвергающий первый, заключается в том, что зрительное опознание объекта (если взять только один пример) требует сложной переработки: в своей наивысшей форме оно требует, чтобы поступающие сенсорные данные обеспечивали представление, не зависящее от угла зрения, расстояния и освещения. Иначе как можно узнать объект, т.е. связать его с прошлым опытом? Даже в самых простых случаях, скажем при узнавании полос (например, когда одна рыба рассматривает бок другой) или движущейся точки (например, когда лягушка глядит на муху), оно требует хорошего представления в нервных сигналах топологических соотношений между входными сигналами, действующими на сенсорную поверхность — сетчатку. В противоположность этому обонятельная система функционирует просто как распознаватель градиентов интенсивности. Короче, обоняние как средство обеспечения необходимого для поддержания жизни поведения избавлено от многих вычислительных трудностей, которые внутренне присущи зрению и другим сенсориумам.
Конечно, аргументы такого рода могут показаться не очень убедительными. Если лимбическая система требует от зрительного, слухового и соматического сенсориумов многократных переключений в новой коре и, следовательно, каскада повторных представлений данных, которые первоначально были сенсорными, то почему это в той же степени не справедливо для стриатума? И почему, далее, в стриатум входят пути и из первичных сенсорных полей, и из различных ассоциативных областей, в которых первичные кортикальные данные претерпели последовательные трансформации? Возможно, основная трудность ответа на эти вопросы связана с непостижимым хаосом, творящимся в любой мозговой структуре, в которую тем или иным путем поступают сигналы со всей (или почти всей) площади новой коры. Охарактеризовать сколько-нибудь систематически поток информации в такую структуру кажется невозможным, и потому наше сознание отказывается вообразить, что же такая структура может делать с этим потоком. Тем не менее в центральной нервной системе представлено несколько таких структур: лимбическая система, стриатум, мост (и через его посредство — мозжечок) и верхнее двухолмие.
Наш обзор центральной нервной системы млекопитающих на этом подходит к концу. У него много недостатков, которых трудно избежать. Во-первых, он дает слабое представление об истинной сложности нервной сети; если бы в нем были упомянуты все известные системы проведения, рисунки к статье могли бы стать безнадежно запутанными. Так, мы не рассматривали варолиев мост, хотя этот отдел получает массивные пучки волокон от всех частей новой коры и посылает массивные пучки в мозжечок. Линии, представляющие эти пучки, пересекались бы с несколькими восходящими и нисходящими системами волокон заднего мозга.
Во-вторых, мы смогли лишь слабо отразить различия между путями, состоящими из миллионов волокон, и путями, в которых число волокон составляет лишь часть этого числа. Так, у приматов спино-таламический пучок, входящий в вентральное ядро таламуса, может насчитывать не более нескольких сотен волокон, тогда как медиальный лемниск содержит миллион волокон, если не больше. Кроме того, мы не имели возможности указать, какие из путей пересекают срединную плоскость и проходят на противоположную сторону центральной нервной системы, а какие имеют места назначения на той же стороне, из которой исходят. Центральная нервная система, как и остальная часть тела, билатерально симметрична. Различение прямых и перекрестных путей играет важную роль в клинической диагностике.
Более важно то, что, обратившись только к связям внутри мозга, а именно — только к происхождению и назначению различных систем волокон, мы можем создать лишь грубый набросок нейроанатомии и не более того, т.е. показать мозг как трехмерный тканевый комплекс, заключенный в черепе и спускающийся вниз по позвоночному каналу. Тем самым мы совсем не касаемся того, над чем человек мучительно размышляет в течение тысячелетий — самой таинственной сущности мозга.
У. КОУЭН
Развитие мозга
В период внутриутробного развития нейроны мозга человека образуются со скоростью сотен тысяч в минуту. Одна из проблем нейробиологии состоит в том, каким образом нейроны находят свое место и формируют надлежащие связи
Общие изменения мозга в процессе развития эмбриона и плода были описаны уже в прошлом столетии, однако до сих пор еще относительно мало известно о лежащих в их основе клеточных процессах, тех процессах, которые обеспечивают формирование отдельных частей мозга и их связей друг с другом. Ясно одно — это, что нервная система берет начало от пласта клеток на дорсальной поверхности развивающегося эмбриона (от нервной пластинки), что эта ткань складывается затем в удлиненную полую структуру (нервную трубку) и что на головном конце трубки выделяются три выпуклости, соответствующие трем главным частям мозга — переднему, среднему и заднему мозгу.
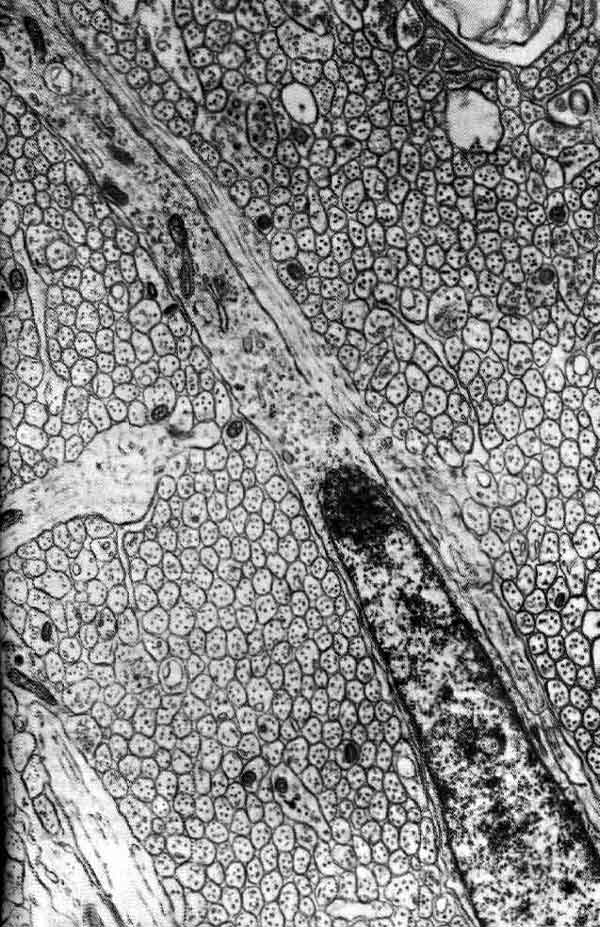
На электронной микрофотографии, полученной П. Ракичем из Медицинской школы Йельского университета, запечатлен момент миграции молодого нейрона у плода обезьяны от места появления в глубине мозжечка к месту назначения вблизи наружной поверхности развивающегося мозга. Мигрирующий нейрон - это более широкая из двух диагональных полос, проходящих через всю микрофотографию от левого верхнего угла до правого нижнего; вытянутое темное образование в верхней части полосы-ядро нервной клетки. Более светлая, узкая полоса, тянущаяся вдоль нижней границы нейрона,-это удлиненный отросток глиальной клетки, которая служит опорной структурой и ориентиром для мигрирующего нейрона. Нейрон движется через плотный нейропиль, или «войлок» из нервных волокон, расходящихся в разных направлениях. (Большинство округлых структур на фотографии являются поперечными срезами аксонов, расположенных более или менее перпендикулярно к плоскости снимка.) Хотя мигрирующий нейрон таким образом контактирует с тысячами других клеточных отростков, он остается тесно связанным именно с глиальной клеткой на протяжении всей ее длины.
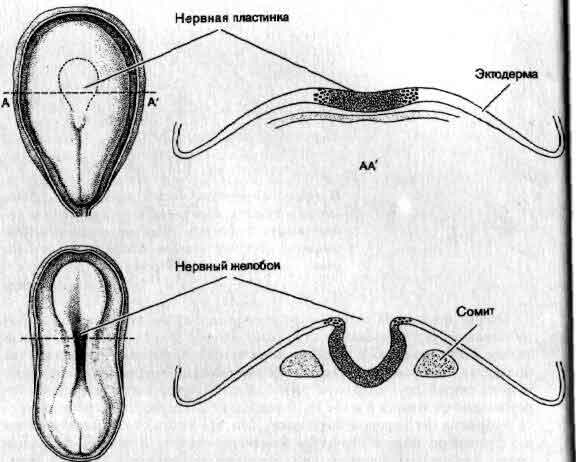
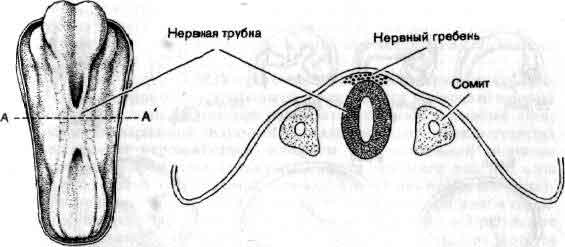
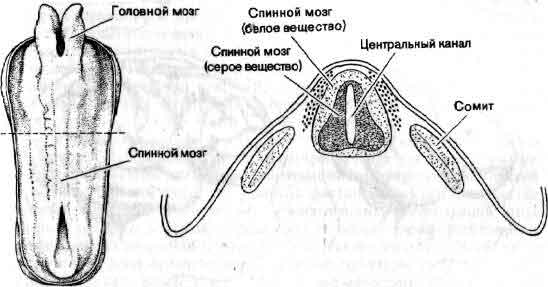
Четырьмя парами рисунков, которые показывают внешний вид развивающегося зародыша (вверху) и соответствующие поперечные срезы на уровне середины будущего спинного мозга (внизу), иллюстрируется происхождение нервной системы из эктодермы, или наружного слоя клеток, зародыша человека на 3-й и 4-й неделе после зачатия. Центральная нервная система берет начало от нервной пластинки - плоского слоя эктодермальных клеток на дорсальной поверхности зародыша. Пластинка впоследствии замыкается в полую структуру, называемую нервной трубкой. Головной конец центрального канала расширяется, образуя желудочки, или полости, головного мозга. Периферическая нервная система формируется в основном из клеток нервного гребня и из волокон двигательных нервов, выходящих из нижних участков каждого сегмента будущего спинного мозга.
Однако не на этих изменениях внешней формы развивающегося мозга сконцентрировано внимание нейробиологов развития в последние годы. Ставятся более интересные вопросы. Как, к примеру, возникают различные составные элементы крупных частей нервной системы, как они занимают определенное положение внутри мозга, каким образом происходит дифференцировка нейронов и окружающих их глиальных клеток, как устанавливают связи друг с другом нейроны различных участков мозга? Несмотря на многочисленные попытки исследований в этих направлениях, еще невозможно дать полное описание развития какой-либо части мозга, и тем более мозга в целом. Определяя, однако, какие события являются главными в развитии нервной системы, можно рассчитывать на успешное решение этих кардинальных вопросов.
В развитии любой части мозга можно выделить восемь основных стадий.
Последовательно это:
1) индукция нервной пластинки,
2) локализованное деление клеток в различных участках,
3) миграция клеток из зоны, в которой они возникли, к местам, где они останутся
окончательно,
4) агрегация клеток, приводящая к формированию идентифицируемых участков мозга,
5) дифференцировка незрелых нейронов,
6) формирование связей с другими нейронами,
7) избирательная гибель некоторых клеток и
8) ликвидация одних ранее сформированных связей и стабилизация других.
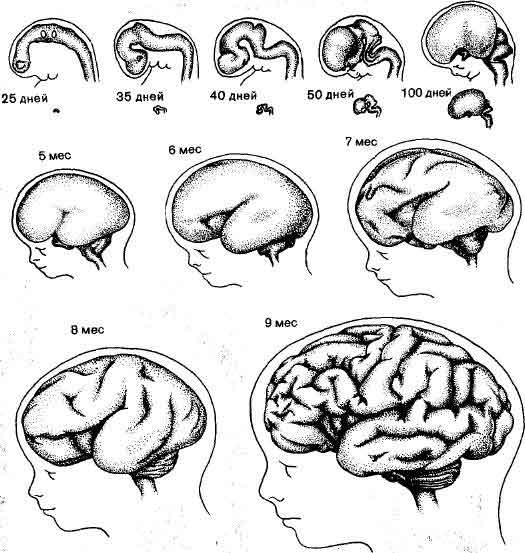
В этой серии рисунков показано развитие мозга человека (вид сбоку) на эмбриональных стадиях и у плода. Рисунки основной серии (внизу) все приведены в одном масштабе. Первые пять эмбриональных стадий (вверху) проиллюстрированы увеличенными рисунками, чтобы яснее показать детали. Три основные части головного мозга (передний, средний и задний мозг) образуются как выпуклости головного конца нервной трубки. У людей полушария головного мозга со временем нарастают на средний и задний мозг, а также частично закрывают мозжечок. Характерные для головного мозга извилины и борозды не обнаруживаются до середины периода внутриутробного развития. Если предположить, что полностью развитый головной мозг содержит порядка 100 миллиардов нейронов и что после рождения фактически не добавляется новых нейронов, то можно вычислить, что в период развития нейроны должны образовываться со скоростью более чем 250000 в минуту.
Процесс, посредством которого часть клеток эктодермы, или наружного слоя развивающегося зародыша, превращается в специализированную ткань, в ткань, из которой развивается головной и спинной мозг, называется нейральной индукцией. С двадцатых годов нашего столетия известно, что определяющим событием для нейральной индукции является взаимодействие эктодермы и части лежащего под ней слоя ткани, называемого мезодермой. Природа этого взаимодействия остается неясной, но есть основания полагать, что оно включает в себя специфический перенос веществ из мезодермы в эктодерму и что в результате этого переноса недифференцированная ткань эктодермы оказывается необратимо вовлеченной в формирование нервной ткани. Ясно также, что последующее взаимодействие различных участков эктодермы и мезодермы приводит к пространственному детерминированию главных частей будущего головного и спинного мозга. Передняя часть мезодермы, входя в контакт с эктодермой, специфически индуцирует структуры переднего мозга, следующая за ней часть обеспечивает формирование структур среднего и заднего мозга, и, наконец, задняя ее часть, расположенная под эктодермой, ответственна за происходящее позднее формирование спинного мозга.
Многие детали процесса пространственной детерминации остаются неясными. Эксперименты с дезагрегированными эктодермальными и мезодермальными клетками из зародышей соответствующих стадий позволили предположить, что важным фактором для этого процесса является создание определенных относительных концентраций двух агентов, которые, вероятно, представляют собою низкомолекулярные белки. Один из них, нейрализующий агент, как бы подготавливает эктодерму к будущему превращению в нервные структуры, а другой — мезодермализующий агент, — присутствуя в варьирующих концентрациях, определяет региональные различия внутри эктодермы.
В 30-е и 40-е годы были сделаны многочисленные попытки выделить предполагаемые индуцирующие агенты, однако сегодня стало ясно, что эта работа была преждевременной. Только в последние два десятилетия существенно обогатились наши знания о природе генной индукции в целом, но и сейчас осталось далеко не ясным, являются ли механизмы индукции, обнаруженные у микроорганизмов, теми же, что и в животных клетках. Существует и другая причина того, почему проблема нейральной индукции трудно поддается решению. Единственной экспериментальной системой, доступной для изучения нейральной индукции, является эктодерма, выделенная у зародыша соответствующей стадии, при этом,, поскольку эктодерма способна отвечать на индуктивный стимул только в течение короткого периода развития, исследователям приходится работать с предельно малым количеством ткани. Нужно отдать дань изобретательности и экспериментальному искусству тех, кто взялся за эту проблему и благодаря кому уже достигнут такой большой прогресс.
По мере того как основные части нервной системы детерминируются, их потенции становятся все более и более ограниченными. К примеру, головной конец нервной пластинки представляет собой изначально переднемозговое - глазное поле, из которого впоследствии разовьются как передний мозг, так и нервные структуры глаза. Если на этой стадии удалить небольшой кусочек эктодермальной ткани, дефект будет быстро устранен за счет размножения соседних клеток, и развитие как переднего мозга, так и глаза пройдет совершенно нормально. Если ту же самую операцию произвести на несколько более поздней стадии, останется неустранимый дефект либо переднего мозга, либо глаза, в зависимости от локализации удаленного кусочка. Иными словами, на более поздней стадии удается с определенностью выделить поле переднего мозга, которое образует дефинитивные структуры переднего мозга, и глазное поле, которое образует только нервные структуры глаза.
На еще более поздних стадиях разграничиваются специализированные области внутри самого переднемозгового поля. С применением различных приемов маркирования клеток стало возможно начертить «карты судьбы» клеток, которые с достаточной точностью предсказывают окончательную локализацию клеток каждого участка раннего переднемозгового поля (см. ). Факторы, приводящие к прогрессивному выделению все более и более мелких единиц, дающих начало специализированным частям мозга, неизвестны, но есть основания предполагать, что, когда мы будем больше знать о клеточной дифференцировке вообще, проблема эта прояснится.
Исследованиями зародышей амфибий установлено, что число клеток в их нервной пластинке относительно мало (порядка 125000), и оно почти не меняется в период формирования нервной трубки. Однако после того, как нервная трубка замыкается, размножение клеток возобновляется в быстром темпе, в результате чего пласт эпителиальных клеток, образующих, нервную пластинку, вскоре превращается в довольно толстый слой клеток, в котором клеточные ядра располагаются на различных уровнях. Микроскопическое исследование клеток, дополненное в ряде случаев введением радиоактивно меченного тимидина, предшественника ДНК, показало, что все клетки в стенке нервной трубки способны размножаться и что характерный вид «псевдонаслоения» эпителия создается благодаря тому, что ядра клеток лежат на разных уровнях. Ядра клеток синтезируют ДНК, когда они лежат в глубине эпителия, а перед делением они мигрируют к вентрикулярной поверхности и убирают свои периферические отростки. После митоза (клеточного деления) дочерние клетки вновь образуют отростки и их ядра возвращаются до возобновления митотического цикла в более глубокие области эпителиального слоя. Миграция ядер размножающихся нейронов весьма характерна для эпителиальных клеток этого типа.
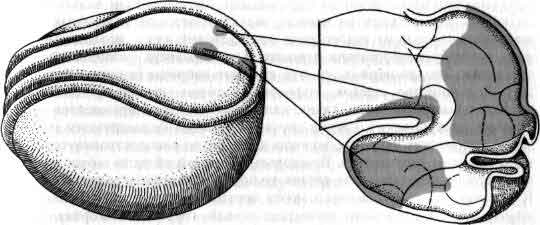
По расположению меток в различных областях нервной пластинки подопытного животного, поставленных на очень ранних эмбриональных стадиях при использовании разных способов маркирования клеток, можно выяснить происхождение каждой из больших областей головного мозга. Здесь показано, как составляются «карты судьбы»; метки вводились в три области нервной пластинки раннего зародыша аксолотля - крупной амфибии (А). Окончательное местоположение маркированных клеток определялось по сагиттальным срезам головного мозга на более поздних стадиях эмбрионального развития (В). (Рисунок сделан по работе Джекобсона из Университета Упсалы.)
После того как клетки проходят ряд таких циклов (число их варьирует от области к области и от популяции к популяции внутри одной и той же области), они теряют способность синтезировать ДНК и мигрируют из эпителия, образуя второй клеточный слой по соседству с вентрикулярной зоной. Клетки, составляющие этот покровный, или промежуточный, слой, являются либо молодыми нейронами, которые уже больше не делятся, либо предшественниками глиальных клеток, сохраняющих способность к делению на протяжении всей своей жизни.
Хотя до сих пор неизвестно, чем включается и выключается механизм размножения в каждой данной области нервной системы, ясно, что сроки, по истечении которых различные популяции клеток перестают делиться, жестко детерминированы, и, более того, имеются веские основания полагать, что периоды эти являются критическими в жизни всех нейронов. Выход клеток из митотического цикла, по-видимому, не только влечет за собой последующую миграцию клеток в промежуточный слой, но и обеспечивает клеткам окончательный их «адрес», в том смысле, что если известна «дата их рождения» (определяемая по времени, когда клетки теряют способность синтезировать ДНК), то можно предсказать и будущее их местоположение. Более того, в некоторых случаях оказывается, что именно в это время определяется характер связей, в конечном счете формируемых нейроном.
На основании экспериментов, в которых небольшие количества меченого радиоактивного тимидина вводились в зародыш (или в случае млекопитающих в организм беременной), исследователи знают теперь даты рождения клеток во многих частях мозга для различных видов животных. Исходя из такого рода исследований, стало возможным сделать некоторые обобщения относительно характера размножения клеток мозга. Это, во-первых, то, что крупные нейроны (а к ним принадлежит большинство клеток, отростки которых распространяются на значительные расстояния, таких, например, как проецирующиеся в зрительные центры мозга клетки сетчатки) обычно формируются раньше, чем более мелкие нейроны, волокна которых не распространяются далеко за пределы тела клетки. Во-вторых, в каждой области мозга имеется характерное распределение клеток ко времени завершения размножения. К примеру, в коре головного мозга первые клетки, прекратившие размножение, впоследствии занимают самый глубинный кортикальный слой, а клетки, образовавшиеся в значительно более позднее время, соответственно создают все более поверхностные слои коры.
С другой стороны, в нейральном компоненте сетчатки (являющейся своеобразным продолжением мозга) пространственное распределение клеток разных генераций прямо противоположное: первое поколение образовавшихся клеток (ганглиозные) мигрируют в самый поверхностный слой сетчатки, а следующие популяции занимают все более глубокие слои. В других областях мозга расположение клеток более сложное, но и для подобных случаев можно считать доказанным, что клетки, локализующиеся в пределах одной общей зоны, образованы одновременно, и наоборот, клетки, образованные в разное время, располагаются, как правило, в разных зонах. Третье обобщение, которое может быть сделано, состоит в том, что в большей части мозга первые опорные клетки появляются примерно в то же самое время, что первые нейроны, но чаще всего размножение глиальных клеток продолжается значительно дольше.
Число нейронов, первоначально образованных в любой части мозга, определяется тремя факторами. Первым фактором является длительность пролиферативного периода: по последним данным она может варьировать от нескольких дней до нескольких недель. Второй фактор — это длительность клеточного цикла: у ранних зародышей она составляет несколько часов, однако далее по мере развития она может увеличиться до 4-5 дней. Третьим фактором является число клеток-предшественников, из которых образуется популяция нейронов.
В настоящее время существует ряд методов для определения длительности пролиферативного периода и длительности клеточного цикла, но лишь в малом числе случаев удается выявить пул клеток-предшественников. Одна из причин такого рода затруднения состоит в том, что пока еще нет способа проследить за судьбой отдельных клеток развивающегося мозга млекопитающих, как это было сделано при изучении значительно более простых форм нервной системы беспозвоночных. Зародыши этих организмов часто бывают совершенно прозрачными, и поэтому отдельные клетки можно наблюдать на протяжении нескольких митотических циклов с помощью светового микроскопа, оснащенного дифференциально-интерференционной оптикой. Иногда клетки-предшественники у таких организмов бывают настолько крупными, что могут быть без труда помечены с помощью внутриклеточных инъекций молекул-маркеров, таких, как пероксидаза хрена; если маркер не разрушается, то он обнаружится во всех потомках меченой клетки, по крайней мере в нескольких поколениях.
Поскольку большинство нейронов образуется в вентрикулярной зоне нервной трубки или в непосредственной близости от нее, а окончательно располагается на некотором расстоянии от этого слоя, следовательно, они должны пройти хотя бы одну стадию миграции после выхода из цикла. Только в небольшом числе случаев клетки, мигрируя из вентрикулярной зоны, продолжают делиться. Деление это обычно наблюдается в особой области между вентрикулярной и промежуточной зонами, названной субвентрикулярной зоной. Этот слой, наиболее отчетливо выраженный в переднем мозгу, дает начало многим более мелким нейронам глубинных структур больших полушарий (базальных ганглиев), некоторым мелким кортикальным нейронам и многим глиальным клеткам коры головного мозга и подлежащего белого вещества. В заднем мозгу ряд клеток из субвентрикулярной зоны вторично мигрирует под поверхность развивающегося мозжечка, где они дают начало специальной пролиферативной зоне, известной под названием наружного зернистого слоя. У человека размножение клеток в этом слое мозга продолжается в течение нескольких недель; здесь образуется большинство вставочных нейронов коры мозжечка, включая миллиарды специфических клеток мозжечка — так называемых клеток-зерен. За этим и некоторыми другими исключениями, в миграции участвуют постмитотические клетки.
В большинстве случаев движение нейронов при миграции носит амебоидный характер. Мигрирующая клетка вначале выбрасывает ведущий отросток, который прикрепляется к подходящему субстрату; ядро перетекает или втягивается в отросток, после чего подтягивается задний отросток. В целом это довольно медленный процесс: средняя скорость миграции клетки составляет около одной десятой миллиметра в день. В некоторых случаях клетка как целое не мигрирует. Вместо этого вначале на ранней стадии развития она выпускает несколько отростков, а позднее тело клетки перемещается постепенно все дальше и дальше от первых отростков, которые остаются при миграции тела на прежнем месте.
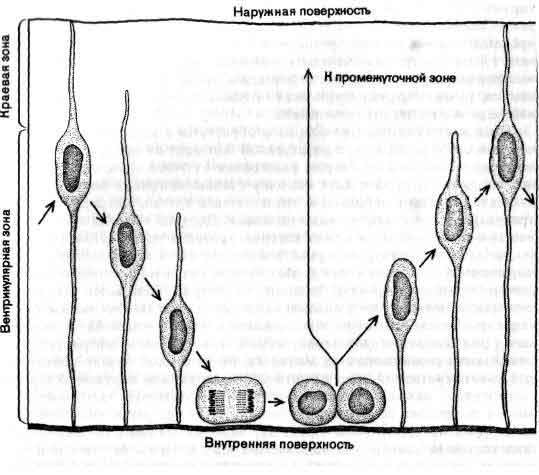
Ядра нервных клеток мигрируют в слое эпителиальной ткани, образующей стенку нервной трубки развивающегося зародыша. В период, когда в клетках, расположенных в этом слое, называемом нейроэпителием, или вентрикулярной зоной, реплицируется ДНК, их ядра движутся по направлению к внутренней поверхности эпителия, периферические отростки отделяются от наружного слоя, и клетки перед делением округляются. После митоза дочерние клетки либо выпускают новый отросток, по которому их ядра могут мигрировать обратно в средний слой эпителия, либо (если клетки прекращают делиться) удаляются из эпителия, принимая участие в образовании промежуточного слоя стенки мозга.
Поскольку нейроны часто мигрируют на значительные расстояния, интересно знать, на какого типа направляющие сигналы они реагируют. В частности, возникает вопрос, «откуда они знают», в какой момент времени следует сделать остановку и начать агрегацию с другими подобными нейронами. Уже на протяжении некоторого времени известно, что в развивающемся мозге существуют специализированные глиальные клетки, тела которых расположены в вентрикулярной зоне, а отростки вытянуты радиально к поверхности. Поскольку эти клетки появляются на ранних стадиях развития и продолжают существовать еще некоторое время после того, как нейроны прекратят миграцию, предполагается, что они могут служить удобными направляющими, вдоль которых нейроны могут двигаться. На электронных микрофотографиях многих частей развивающегося мозга мигрирующие клетки почти всегда обнаруживаются в тесном контакте с соседствующими отростками глиальных клеток. Этот факт позволил П. Ракичу (P. Rakic) из Медицинской школы Йельского университета постулировать, что мигрирующие клетки направляются к своему постоянному местоположению отростками глиальных клеток. В пользу этой точки зрения говорят наблюдения Ракича и Р. Сидмана (R. Sidman) из детской больницы Медицинского центра в Бостоне, заметивших, что в результате одной из наиболее поразительных генетических мутаций, затрагивающих мозжечок мыши, радиальные отростки глиальных клеток дегенерируют на сравнительно ранней стадии, и миграция большинства клеток-зерен полностью нарушается, видимо, как результат этой дегенерации.
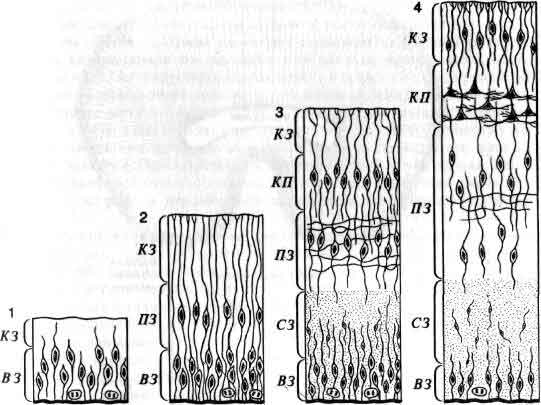
Здесь показано постепенное утолщение стенки развивающегося мозга. На самой ранней стадии (1) стенка содержит только «псевдомногослойный» эпителий, в вентрикулярной зоне (ВЗ) которого находятся тела клеток, а в краевой зоне (КЗ) только вытянутые наружу отростки. Когда некоторые из клеток теряют способность синтезировать ДНК и выходят из митотического цикла (2), они образуют второй слой - промежуточную зону (ПЗ). В переднем мозгу клетки, проходящие через эту зону, агрегируют с образованием корковой пластинки (КП) - области, в которой развиваются различные слои коры головного мозга (3). На самой поздней стадии (4) исходная вентрикулярная зона остается в виде эпендимы - выстилки желудочков мозга, а относительно свободная от клеток область между этой выстилкой и корой становится подкорковым белым веществом, сквозь которое нервные волокна входят в кору и выходят из нее. Субвентрикулярная зона (СЗ) является вторичной зоной размножения, где образуются многие глиальные клетки и некоторые нейроны переднего мозга.
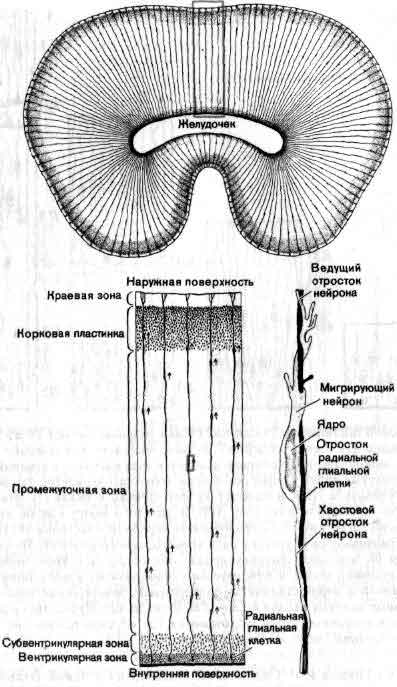
Специализированные опорные клетки - радиальные глиальные клетки - возникают на ранних стадиях развития нервной системы. Для этих клеток характерны необычайно длинные отростки, которые целиком «пробуравливают» стенку нервной трубки и производных структур. На рисунке вверху показано, как выглядят радиальные глиальные клетки на окрашенном по Гольджи толстом поперечном срезе препарата полушария головного мозга у плода обезьяны. Тела клеток лежат в вентрикулярной зоне, а их отростки протягиваются до наружной поверхности окружающих слоев, где они, по-видимому, прикрепляются с помощью разветвленного конца. Внизу слева показан увеличенный сегмент этого поперечного среза. Небольшой кусочек ткани в рамке показан внизу справа крупнее в виде объемной картинки, сделанной на основании микроскопического исследования Ракича. Рисунок показывает тесную связь между отростками радиальных глиальных клеток и мигрирующими нейронами; связь эта наблюдается при развитии большинства отделов мозга.
Если учесть расстояние, на которое многие нейроны перемещаются за период развития, то не приходится удивляться тому, что во время своей миграции некоторые клетки отклоняются от правильного пути и оказываются в итоге не на своем месте. Долгое время патологи считали такое смещение положения нейронов (названное эктопией) сопутствующим обстоятельством некоторых серьезных нарушений развития мозга и не принимали во внимание, что даже во время нормального развития часть мигрирующих клеток может неадекватно реагировать на обычный направляющий стимул и оказаться в неверном положении. Современные методы исследования в ряде случаев дали возможность выявить клетки такого рода; при этом оказалось, что большинство подобных смещенных нейронов не удается обнаружить на более поздних стадиях развития. При тщательном изучении с этой точки зрения одной из популяций нейронов оказалось, что неточно мигрирует около 3% клеток, и за редким исключением все смещенные нейроны дегенерируют.
Когда мигрирующие нейроны достигают своего окончательного местоположения, они агрегируют с другими аналогичными клетками, образуя либо корковые слои, либо ядерную массу. Способность развивающихся клеток одинакового происхождения избирательно слипаться с другими аналогичными клетками была впервые обнаружена более 50 лет назад, однако только в последнее десятилетие этот факт привлек заслуженное внимание нейроэмбриологов. Отправным стимулом для современных исследований послужил поиск молекулярных механизмов, лежащих в основе формирования специфических связей между родственными группами нейронов. К сожалению, задача эта оказалась трудно поддающейся решению, хотя большая часть проделанной при этом работы непосредственно подводит к важным выводам в отношении того, как образуются в развивающемся мозге дискретные популяции нейронов.
Возможно, наиболее важным в этой серии исследований явилось следующее наблюдение: если клетки из двух или трех областей развивающейся нервной системы диспергировать (механически или же с помощью мягкой химической обработки), смешать вместе, а затем оставить реагрегировать в подходящей среде, то они проявляют тенденцию сортироваться таким образом, что клетки каждой данной области агрегируют преимущественно с клетками той же области. Избирательная адгезивность, по-видимому, является общим свойством всех живых клеток и обеспечивается наличием на их поверхности особого класса больших молекул, которые служат как для «узнавания» клеток того же сорта, так и для связывания клеток друг с другом. Эти молекулы, функционирующие как межклеточные связки (лиганды), высоко специфичны для каждого крупного типа клеток. В процессе развития они, вероятно, изменяются либо в количественном отношении, либо по характеру распределения. В последнее время сотрудники нескольких лабораторий предпринимают попытки выделить и охарактеризовать те или иные поверхностные лиганды, и, быть может, проблема эта окажется первой среди других проблем биологии развития нервной системы, которые будут успешно решены на молекулярном уровне.
Еще одна особенность агрегации клеток в развивающейся нервной системе состоит в том, что в большинстве областей мозга клетки не только агрегируют друг с другом, но и приобретают некоторую предпочтительную ориентацию. К примеру, в коре головного мозга большинство крупных пирамидных нейронов согласованно выстраиваются в ряд таким образом, что их выступающие апикальные дендриты оказываются направленными к поверхности, а их аксоны располагаются в направлении подлежащего белого вещества. Непонятно, что помогает клеткам выстраиваться в ряд, однако представляется вероятным, что этому способствует либо наличие на поверхности клетки молекул разных классов, задающих ориентацию, либо избирательное перераспределение поверхностных молекул, определяющее исходную клеточную агрегацию.
Одной из наиболее ярких особенностей развивающихся нейронов является постепенное развитие их отростков. Одинаково важными чертами являются также приобретение нейронами одного из способов проведения сигналов (большинство нейронов генерирует потенциал действия, а некоторые обеспечивают только проведение с декрементом) и выбор одного из двух способов взаимодействия клеток друг с другом (либо посредством формирования традиционных синапсов, обеспечивающих выделение химического медиатора, либо посредством формирования щелевых контактов, обеспечивающих электрическое взаимодействие клеток). Для нейробиологов только теперь начинают проясняться некоторые из этих наиболее глубоких аспектов дифференцировки нейронов, и становится понятно, что нейроны могут оказаться значительно более сложно организованными клетками, чем ранее представлялось. Например, недавно было показано, что некоторые нейроны могут переключаться с одного типа медиатора на другой (в частности, с норадреналина на ацетилхолин) под влиянием факторов среды, тогда как другие могут на разных стадиях развития менять тип иона, используемого для распространения нервного импульса (к примеру, кальций - на натрий).
Значительно больше известно о процессе формирования нервных отростков. Нейроны мозга млекопитающих в большинстве своем являются мультиполярами — с несколькими конусообразными дендритами, функционирующими как рецептивные отростки, и одним аксоном, который служит главным эффекторным отростком клетки. Хотя некоторые клетки, как известно, образуют отростки до того, как начинают мигрировать, большинство образует их после того, как займет окончательное местоположение. Точно не ясно, что стимулирует образование отростков. Исследованиями, в которых изолированные незрелые нейроны поддерживались в культуре, выяснено, что отростки образуются только на клетках, прикрепляющихся к подходящему субстрату, и что при этом условии клетки часто способны образовывать совершенно нормальный набор дендритов и аксонов. В некоторых случаях, несмотря на пребывание в искусственных условиях, общий вид дендритов, образованных в культуре, точно соответствует характерному для интактного мозга, хотя при этом клетки лишены всех контактов с другими нейронами и даже с глиальными клетками. Наблюдения такого рода дают основания предполагать, что информация, необходимая для формирования характерного ветвления дендритов, детерминирована генетически.
Доказано также, что во время нормального развития большинство нейронов испытывает различного рода локальные механические влияния, которые модифицируют их форму. Несомненно, что количество и распределение получаемых клеткой импульсов способно коренным образом влиять на окончательную форму нейронов. Яркий пример такого рода влияния представляет мозжечок. Дендриты наиболее характерного класса нейронов коры мозжечка - клеток Пуркинье - в норме располагаются в плоскости, ориентированной под прямым углом к аксонам клеток-зерен, представляющих их основной вход; если по какой-либо причине нарушится обычное регулярное расположение этих аксонов, то соответственно изменится и характер расположения дендритов клеток Пуркинье.
Теперь стало достаточно ясно, каким образом вытягиваются отростки нейронов. У большинства отростков на их растущих концах находятся особые структуры, названные конусами роста. Эти развернутые, весьма подвижные структуры в живом виде выглядят как постоянно и активно исследующие свое непосредственное окружение. Они являются местами, в которых происходит добавление большей части нового материала к растущему отростку. Когда отросток разветвляется, он почти всегда образует новый конус роста. Хотя и по косвенным данным, можно, однако, полагать, что внутри или на поверхности конуса роста на молекулярном уровне заложена способность обнаруживать субстрат, вдоль которого следует расти, а также идентифицировать соответствующие мишени. Эксперименты, в которых нейронам создавались условия для роста на различных искусственных субстратах, показали, что большинство отростков растет преимущественно вдоль поверхности повышенной адгезивности.
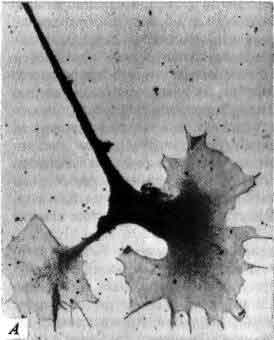
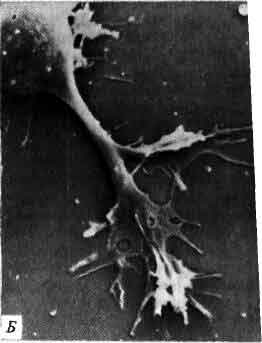
На этих двух электронных микрофотографиях видны конусы роста (растянутые, очень подвижные структуры на концах растущих отростков нервных клеток). А. Микрофотография, полученная с помощью обычного электронного микроскопа, показывает два конуса роста на конце аксоноподобного отростка клетки из симпатического ганглия крысы. Диссоциированные клетки росли в культуре, и показанный отросток разветвился за несколько минут до того, как клетка была фиксирована и обработана (без приготовления среза) для микроскопии. Тонкие пальцевидные выросты являются филоподиями, а распластанные поверхности между ними, похожие на тонкие пленочки, - ламеллоподиями. Б. На микрофотографии, полученной с помощью сканирующего электронного микроскопа, показан растущий дендрит нейрона из гиппокампа плода крысы. Конусы роста в этом месте образовались уже через 2 часа после того, как диссоциированные клетки были перенесены в культуру, Оба снимка были сделаны в Медицинской школе Вашингтонского университета Дж. Кочраном, М. Банг (А) и Ст. Ротменом (Б).
Одна из наименее понятных проблем нейробиологии развития состоит в том, как аксоны определяют правильный путь. Это особенно трудно понять, когда аксонам приходится протягиваться на значительные расстояния сквозь структуры мозга, и в одной или нескольких точках своего пути отклоняться либо влево, либо вправо, продвигаясь к противоположной стороне мозга и образуя одно или несколько ответвлений до того, как они окончательно достигнут предопределенного местоположения. В некоторых системах это выглядит так, как если бы аксоны росли просто под влиянием каких-то градиентов, направленных вдоль больших осей головного и спинного мозга; в других системах больше похоже на то, что направление определяется взаимоотношениями с ближайшими соседями. Во многих случаях, однако, кажется, что внутри растущего аксона закодирован тонкий молекулярный механизм, позволяющий аксону точно реагировать на химические или структурные указатели на его пути.
Пример такого направленного роста продемонстрировала недавно Р. Леви-Монтальчини (R. Levi-Montalcini) из Лаборатории биологии клетки Национального совета по научным исследованиям в Риме. Когда она и ее коллеги вводили в головной мозг молодым крысятам белок, известный под названием фактора роста нервов, наблюдался аномальный рост аксонов клеток симпатического ганглия (периферических нейронов, лежащих рядом с позвоночником и, как известно, чувствительных к фактору роста) в спинной мозг и вверх по направлению к головному мозгу, по-видимому, вдоль пути, по которому диффундировал инъецированный фактор роста. В этом случае фактор роста нервов действовал не столько как трофический, или стимулирующий, рост, сколько как тропный, или направляющий, рост, при этом аксоны симпатических нервов отвечали хемотропизмом на присутствие этого фактора. Имеются еще два аспекта проблемы роста нервных отростков, заслуживающие обсуждения. Во-первых, это то, что большая часть нейронов образует вначале значительно больше отростков, чем им необходимо или же чем они способны впоследствии сохранить. Многие молодые нейроны несут на себе множество коротких дендритоподобных отростков, из которых почти все по мере «взросления» клеток втягиваются назад. Аналогично большинство развивающихся аксонов образует значительно больше связей, чем их остается во взрослом состоянии, иными словами, существует процесс ликвидации связей, во время которого устраняются многие (а в некоторых случаях все, за исключением одной) первоначальные связи. Второй аспект состоит в том, что у аксонов имеется тенденция расти в тесном соседстве друг с другом. Этот феномен известен под названием фасцикуляции (расположения пучками). В современных работах предполагается, что фасцикуляция связана с появлением на поверхности большей части аксонов веществ-лигандов, которые делают их способными объединяться и расти совместно с другими родственными аксонами. Одним из преимуществ латерального объединения такого типа является, по-видимому, то, что только первому нейрону в группе необходим традиционный конус роста, другие же аксоны лишь следуют за лидером.
Несомненно, что наиболее важным из нерешенных вопросов в развитии мозга остается вопрос, каким образом нейроны формируют специфическую пространственную структуру связей. Существовавшее ранее представление о том, что большинство связей мозга функционально отбирается из совокупности случайно сформированных соединений, в настоящее время кажется неправомочным. Уже на ранних стадиях развития большинство связей устанавливается точно; существует много доказательств того, что образованные связи специфичны не только для данной области мозга, но и для данного нейрона (а в некоторых случаях и для данных частей нейрона) внутри этой области.
Выдвинуто несколько гипотез для объяснения того, как достигается такая замечательная точность. Некоторые исследователи считают возможным объяснить это тем, что растущие аксоны поддерживают такие же топографические взаимоотношения друг с другом, как и тела их клеток. Другие подчеркивают значение временного аспекта (в частности, критическим является момент, к которому различные группы волокон достигают области своего назначения). Одной из гипотез, в которой делается попытка объяснения всех наблюдаемых особенностей роста, может послужить гипотеза химического сродства, сформулированная Р. Сперри (R. Sperry) из Калифорнийского технологического института. Согласно его точке зрения, большинство нейронов (или, что более вероятно, большинство малых популяций нейронов) приобретают химические различия на ранней стадии своего развития в зависимости от занимаемого положения, и эта их дифференцированность выражается в наличии соответствующих меток, которые и позволяют аксонам узнавать либо аналогичную, либо комплементарную метку на поверхности нейронов-мишеней.


Одним из экспериментальных подходов к изучению того, как нейроны образуют специфическую пространственную картину связей в развивающемся мозге, является воздействие на проекцию сетчатки в зрительном тектуме среднего мозга. При таком подходе, впервые разработанном Р. Сперри из Калифорнийского технологического института, глазное яблоко взрослой лягушки (или головастика на разных стадиях развития) поворачивают или трансплантируют. Позднее, после регенерации зрительного нерва (или после того как головастик превратится в лягушку и аксоны ганглиозных клеток сетчатки, составляющие зрительный нерв, образуют связи со зрительным тектумом), можно видеть, какое воздействие оказала операция на поведение лягушки. Данная серия рисунков, основанная на работе Сперри, вначале демонстрирует поведение нормальной лягушки (А). В первом эксперименте (Б) правый глаз был повернут на 180°; некоторое время спустя после регенерации зрительного нерва оказывалось, что попытка лягушки напасть на приманку, помещенную в верхнем поле зрения, была ошибочной точно на 180°. В следующем эксперименте (В) левый глаз был заменен на правый таким образом, что инвертировалась только дорсовентральная ось (толстая стрелка); в этом случае лягушка делала прыжок вперед к приманке, но в сторону нижнего, а не верхнего поля зрения. Затем (эксперимент Г) была осуществлена сходная трансплантация, но на этот раз глаз был повернут в передне-заднем направлении (тонкая стрелка); лягушка ощущала, что приманка располагалась в верхнем поле зрения, но прыгала вперед, вместо того, чтобы прыгать назад. Вывод из этих экспериментов согласуется с предположением о том, что волокна зрительного нерва при регенерации всегда прорастают опять в ту же часть зрительного тектума, которую они первоначально иннервировали, и что во время нормального развития они «находят путь» к правильному местоположению в тектуме сходным образом. Эти результаты объясняются гипотезой, что как ганглиозные клетки сетчатки, так и нейроны-мишени в тектуме несут химические признаки, помогающие им идентифицировать друг друга.
Хотя эта проблема может быть распространена на все части нервной системы, наиболее интенсивно она исследуется на двух системах - системе иннервации мускулатуры конечностей мотонейронами спинного мозга и системе проекции ганглиозных клеток сетчатки к месту их назначения (у низших позвоночных) в зрительный тектум мозга. Изучение иннервации мышц показывает, что при нормальных обстоятельствах на ранней стадии развития небольшие популяции мотонейронов, называемые пулами, отделяются друг от друга: каждый пул мотонейронов иннервирует преимущественно отдельную мышцу конечности, и при этом случается очень мало ошибок. Хотя пространственное соответствие при иннервации в норме является точным, точность эта не абсолютна. Поэтому если заднюю конечность куриного эмбриона-донора трансплантировать рядом с ногой эмбриона-реципиента, то мышцы дополнительной конечности будут иннервироваться неизменно пулами мотонейронов, которые в норме иннервируют или мускулатуру туловища, или мускулатуру тазового пояса. Картина иннервации явно аномальная, однако тот факт, что мышцы трансплантированной конечности всегда иннервируются одними и теми же популяциями клеток, служит убедительным подтверждением того, что даже при этих необычных условиях мотонейроны руководствуются некоторым определенным (пока еще неустановленным) набором правил.
Ретино-тектальная система оказалась особенно подходящей для анализа обсуждаемой проблемы. На эмбриональных и личиночных стадиях развития амфибий удается производить различные экспериментальные манипуляции, такие, как поворот глазного яблока, составление глаза из различных сегментов двух или более сетчаток, удаление или поворот частей тектума. После окончательного развития системы оказывается возможным анатомически, электрофизиологически, а также в поведенческих опытах выявить связи, сформированные ганглиозными клетками сетчатки. Кроме того, у рыб и амфибий зрительный нерв (состоящий из аксонов ганглиозных клеток сетчатки) способен регенерировать после разрушения, что позволяет проводить аналогичные эксперименты на ювенильных и взрослых животных. В настоящее время на этой системе получено огромное количество данных, и мы суммируем здесь только некоторые основные достижения.
Пожалуй, наиболее важные результаты в исследованиях такого рода получены в двух основных группах экспериментов. В первой группе у лягушек и хвостатых амфибии перерезали зрительный нерв и поворачивали глаз на 180°. В другой группе у золотой рыбки и лягушки иссекали участки зрительного тектума и либо поворачивали их, либо пересаживали в другую часть тектума. В обеих группах экспериментов регенерирующие волокна зрительного нерва, как это могло быть зарегистрировано в электрофизиологических или поведенческих опытах, прорастали в те же самые участки тектума, которые они исходно иннервировали. Наиболее простое объяснение этих результатов состоит в том, что аксоны ганглиозных клеток и их нейроны-адресаты в зрительном тектуме маркированы одинаковым образом и что регенерирующие аксоны растут до тех пор, пока не «опознают» соответствующие метки на нужных нейронах тектума.
Трудно отрицать возможность того, что при этих обстоятельствах волокна различных участков сетчатки смогли оставить свой след на соответствующих группах клеток тектума и что аксоны или нейроны тектума сумели «запомнить» свое предыдущее положение. Однако есть основания полагать, что подобный механизм если и срабатывает, то только на начальных стадиях развития системы. Так, если развивающийся глаз лягушки перевернуть ранее некоторой критической стадии развития, то проекция сетчатки на гектум останется нормальной. Если, однако, поворот совершить после критического момента, то и сетчаточная проекция обязательно повернется на столько же градусов. Аналогичным образом, если перевернуть на 180° весь зрительный тектум зародыша в направлении от хвоста к голове (вместе с частью переднего мозга, лежащего впереди него), то и сетчаточная проекция также инвертируется.
Из этих экспериментов следует, что существует определенная стадия в развитии большинства нервных центров, во время которой центры становятся топографически поляризованными, в результате чего входящие в их состав нейроны приобретают определенные характеристики, которые задают пространственную организацию проекции в целом. М. Джекобсон (М. Jacobson) из Медицинской школы Университета Майами несколько лет назад показал, что сетчатка шпорцевой лягушки Xenopus laevis становится поляризованной примерно к тому времени, когда первые ганглиозные клетки прекращают делиться. Хотя на этой стадии имеется еще только около 1% ганглиозных клеток, будущая пространственная структура проекции сетчатки в тектум, по-видимому, уже детерминируется в это время. Совершенно непонятно, каким образом нейроны получают информацию о положении и как она реализуется в процессе роста их отростков. Однако представляется вероятным, что механизмы детерминации полярности не являются специфичными для нервной системы — они едины для всего организма. Р. Хант (R. Hunt) из Университета Джонса Гопкинса, а также Джекобсон обнаружили, что если развивающийся глаз трансплантировать головастику на бок до начала периода осевой спецификации и оставить в этом аномальном положении на весь критический период, то затем, после ретрансплантации в глазную впадину, ганглиозные клетки формируют связи со зрительным тектумом в соответствии с тем, как глаз был ориентирован в тот период, когда он находился на боку, а не в соответствии с его новым положением в глазнице.
Когда растущий аксон достигает своего адресата, будь то другая группа нейронов или ткань эффектора, например совокупность мышечных или железистых клеток, он образует с ними специализированные функциональные контакты — синапсы. Именно в этих местах информация передается от одной клетки к другой обычно с помощью малых количеств соответствующих медиаторов (см. Л. Иверсен «Химия мозга»). Многочисленные феноменологические данные позволяют предположить, что в синаптической области осуществляется двусторонний перенос веществ, необходимых для выживания и нормального функционирования как пресинаптической, так и постсинаптической клеток. Эти вещества, в комплексе отнесенные к трофическим факторам, являются в значительной мере гипотетическими; только одно (фактор роста нервов) было выявлено и охарактеризовано химически. Это вещество, впервые идентифицированное В. Хамбургером (V. Hamburger) и Р. Леви-Монтальчини из Вашингтонского университета еще в 50-х годах, оказалось белком, существующим в норме в виде двух идентичных аминокислотных цепей, каждая из которых имеет молекулярный вес несколько больше 13000.
Хотя способ действия фактора роста нервов пока неясен, известно, что он необходим для роста и выживания клеток симпатического ганглия и что в период развития он специфически содействует росту отростков этих клеток, а также некоторых клеток спинальных ганглиев. Кроме того, как я уже отмечал, он может в некоторых случаях оказывать влияние на направленный рост волокон симпатического нерва. Если ввести новорожденным мышатам антитела к фактору роста нервов, это приведет к поражению всей симпатической нервной системы. Даже у взрослых животных фактор роста нервов, по-видимому, постоянно доставляется симпатическим нейронам их тканями-мишенями; при этом белок, попавший в окончания аксонов, переносится в направлении тела клетки. Если доставку фактора прекратить посредством перерезки аксонов симпатических нейронов, их функциональная целостность в значительной мере нарушается и синапсы, которые оканчивались на клетках, быстро удаляются. Возможно, в ближайшие годы будут выделены другие аналогичные вещества и тогда можно будет утверждать, что большинство классов нейронов нуждается в специфическом агенте для обеспечения выживания и для направленного роста их отростков.
В настоящее время получены доказательства того, что развитие многих структур и тканей сопровождается строго запрограммированными фазами гибели клеток. Это справедливо и в отношении развивающегося мозга. Во многих его областях образуется значительно больше нейронов, чем выживает в последующий период развития. В каждой области, для которой были возможны количественные оценки, обнаружено, что число нейронов регулируется избирательной гибелью клеток, происходящей в предсказуемый период развития (обычно тогда, когда популяция нейронов как целое формирует синаптические связи с тканями-мишенями). Неизвестно, проявляется ли этот феномен в любой части мозга (он был изучен главным образом в небольших группах клеток), но там, где он был обнаружен, он затрагивает от 15 до 85% исходной популяции нейронов.
Поэтому кажется, что окончательные размеры популяции нейронов во многих частях мозга устанавливаются в две фазы: на ранней стадии, когда образуется сравнительно много клеток, и на более поздней, когда происходит регуляция числа нейронов в соответствии с размерами иннервируемого ими поля. Вообще же можно предположить, что лимитирующим фактором, определяющим окончательное число клеток, является количество функциональных контактов, имеющихся в распоряжении аксонов развивающихся нейронов. Ясно, что если в эксперименте уменьшить размеры проекционной зоны, то естественная гибель клеток усиливается в такой же степени. Для спинальных мотонейронов куриного зародыша, иннервирующих мускулатуру задних конечностей, удалось снизить гибель клеток экспериментально трансплантацией дополнительной конечности. По современным данным, однако, более вероятным кажется, что критическим является не столько само образование связей, сколько количество трофического фактора, достигающего клеток.
На более поздней стадии развития осуществляется дополнительная регуляция, но не столько численности популяции нейронов в целом, сколько количества сохраняемых клетками отростков. Феномен элиминации отростков (и синапсов) впервые наблюдали в иннервируемой мускулатуре конечностей у крысят. Можно показать, что в то время как у взрослых животных большинство мышечных клеток иннервируется единичным аксоном, на первой неделе постнатального периода с каждым мышечным волокном образуют синапсы 5-6 аксонов. На протяжении следующих двух-трех недель развития лишние аксоны последовательно исчезают и в результате остается только один. Аналогичная стадия процесса элиминации выявлена при образовании межнейронных связей в периферической нервной системе и в головном мозгу. Приведем только один пример: в мозжечке взрослого животного на каждую клетку Пуркинье приходится по одному нервному волокну, которое известно под названием лиановидного; в раннем же постнатальном периоде с каждой клеткой Пуркинье может контактировать несколько таких волокон. За исключением случаев отдельных генетических мутаций, затрагивающих мозжечок, все, кроме одного из этих волокон, впоследствии элиминируются.
Последовательное исчезновение возникших отростков заставляет задуматься над тем, чем определяется, какой отросток выживет, а какой будет элиминирован. В настоящее время можно лишь догадываться, что развивающиеся волокна каким-то образом конкурируют друг с другом. Имеются косвенные доказательства того, что одним из факторов, обеспечивающих подобное преимущество, может служить их функциональная активность. Во всяком случае, ясно, что для многих систем итоговая картина распределения той или иной популяции нейронов формируется постепенно из довольно аморфной структуры, и при этом часто оказывается возможным заметно изменить окончательный вид структуры и ее связей, воздействуя во время определенного критического периода развития на ее функцию. Для объяснения этого высказывания приведем два примера, касающихся соматосенсорной области коры большого мозга.
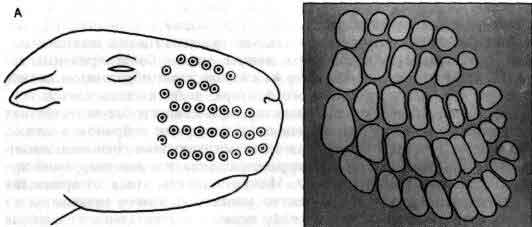
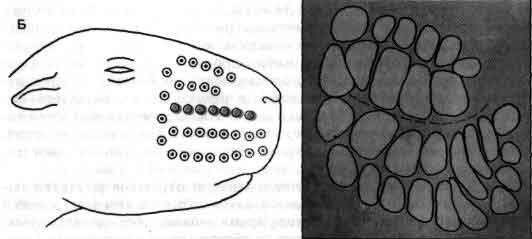
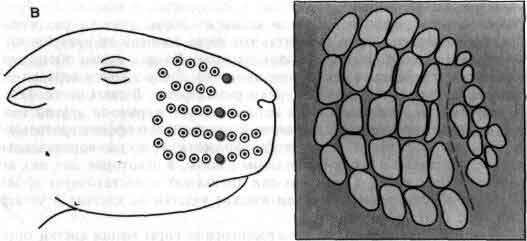
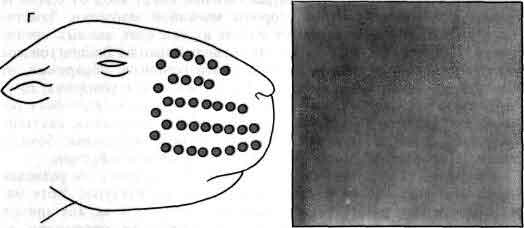
Вибриссы и бочонки соматосенсорной коры молодых мышей служат примером одной из многих систем, в которых обнаружена определяющая их зависимость топографии развивающейся нервной системы от ее входов. Вибриссы - это чувствительные волоски на мордочке мыши, а бочонки - это специализированные скопления нейронов в слое IV коры головного мозга. Входом для каждого бочонка служит одиночная вибрисса с контралатеральной стороны мордочки (А). Если вскоре после рождения животного разрушить один из рядов вибрисс, то позднее будет обнаружено, что соответствующий ряд бочонков в коре большого мозга отсутствует, а соседние бочонки увеличены (Б, В). Если разрушить все вибриссы, то бочонки вовсе исчезнут (Г). Должна иметь место значительная пластичность в развивающейся коре, поскольку волокна, иннервирующие вибриссы, не прямо контактируют с корой, а связаны с ней посредством по крайней мере двух синаптических переключений. Рисунок основан на работе Т. Вулси из Медицинской школы Вашингтонского университета.
У макака информация из сетчатки достигает четвертого слоя зрительной коры через посредство структуры, названной латеральным коленчатым телом. На этом уровне входы в кору от каждого глаза совершенно раздельны, что было прямыми методами показано на подопытных животных с помощью инъекций большого количества меченых аминокислот в одно глазное яблоко. Ганглиозные клетки сетчатки поглощают аминокислоту, включают ее в белок и транспортируют в латеральное коленчатое тело. Здесь некоторая часть метки освобождается и становится доступной для включения в клетки коленчатого тела, которые транспортируют ее по своим аксонам далее в зрительную кору. На соответствующим образом приготовленных радиоавтографах (которые позволяют выявить распределение меченых волокон, достигающих коры) видно, что первичная зрительная область организована в виде перемежающихся так называемых глазодоминантных полосок, причем каждая полоса имеет ширину около 400 мкм и получает входные сигналы либо от правого, либо от левого глаза. Д. Хьюбел, Т. Визель и С. Ле Вэй (D. Hubel, Т. Wiesel, S. Le Vay) из Гарвардской медицинской школы показали, что если сшить веки одного глаза у подопытного новорожденного животного (так чтобы сетчатка глаза не подвергалась воздействию структурированных зрительных стимулов), глазодоминантные полоски, связанные с закрытым глазом, будут заметно уже, чем в норме. В то же время полоски проекции открытого глаза соответственно расширятся (общая ширина двух соседних полосок, различающихся стороной стимуляции, при этом постоянна).
Результат этот частично вызван сжатием глазо-доминантных полосок «выключенного» глаза, сопровождающимся вторичным расширением полосок нормального глаза, и отчасти - сохранением неплотной группировки волокон из нормального глаза, имевшей место на более ранней стадии. Если исследовать входы от обоих глаз на различных стадиях развития, можно показать, что когда волокна из латерального коленчатого тела только достигают зрительной коры, входы обоих глаз существенно перекрываются в зоне проекции. Лишь к концу первого месяца жизни животного полоски четко разделяются. В свете этого факта (и результатов экспериментов, в которых затем закрывали другой глаз, и открывали первый) представляется вероятным, что эффект зрительной депривации состоит в том, чтобы привести геникуло-кортикальные клетки, связанные с нестимулируемым глазом, в некоторое для них невыгодное положение, в котором они проявляют недостаточную эффективность в конкуренции за синаптические участки на клетках в четвертом слое зрительной коры.
В соответствующем слое соматосенсорной коры мыши клетки организованы в четкие группы, названные бочонками. Физиологические исследования показывают, что каждый бочонок имеет вход от одной вибриссы с контралатеральной стороны мышиной мордочки. Заметим, что у мыши вибриссы являются одним из наиболее важных органов чувств. Т. Вулси (Th. Woolsey) из Медицинской школы Вашингтонского университета, первым открывший значение бочонков, обнаружил, что если удалить небольшую группу вибрисс сразу после рождения, то соответствующая группа бочонков в коре не разовьется. Этот факт особенно интересен потому, что между сенсорными нейронами, связанными с вибриссами, и нейронами, образующими кортикальные бочонки, лежат по крайней мере еще две группы вставочных нейронов.
Эти и многие другие наблюдения приводят к выводу, что развивающийся мозг является исключительно пластичной структурой. Хотя многие области могут быть «жестко запаяны», другие (такие, как зрительная кора) открыты для различных воздействий - как внутренних, так и внешних. Способность мозга к реорганизации в ответ на внешние воздействия или на локальное повреждение в настоящее время является наиболее активно изучаемым явлением нейробиологии не только потому, что очевидна его связь с такими феноменами, как обучение и память, и его отношение к способности мозга восстанавливаться после повреждения, но и потому, что они открывают тайны нормального развития мозга.
В заключение хочется подчеркнуть, что развитие мозга, как и развитие большинства других биологических структур, не обходится без ошибок. Ранее уже было упомянуто, что ошибки могут происходить во время миграции нейронов. Известны такие примеры возникновения ошибок во время формирования связей. В зрительной системе, как было замечено рядом исследователей, некоторые волокна зрительного нерва, которые должны были бы пересечь среднюю линию в составе перекреста, начинают в результате ошибки расти к ипсилатеральной стороне мозга. В некоторых таких ситуациях, если у животного на ранней стадии развития удалить один глаз, то число волокон, отклонившихся от правильного направления, может значительно возрасти. Поскольку аномально ориентированные волокна часто не обнаруживаются в зрелом мозгу, создается впечатление, что такие неправильно адресованные нейроны (и любые ими сформированные ошибочные связи) ликвидируются на более поздних стадиях развития. Остается загадкой, как выясняется, что они ошибочные, и каким образом они устраняются. Учитывая сложность механизмов развития, не приходится удивляться тому, что ошибки появляются. Удивительнее то, что они появляются редко и что чаще всего они эффективно исправляются.
Л. ИВЕРСЕН
Химия мозга
Сигналы передаются от нейрона к нейрону разными химическими медиаторами. Эти химические системы, наложенные на нейронные цепи головного мозга, добавляют к его функции еще одно измерение
Нейроны имеют биохимический аппарат, общий со всеми остальными живыми клетками, в том числе способность генерировать химическую энергию путем окисления пищевых веществ, а также восстанавливать и сохранять свою целостность. Нейроны обладают, кроме того, специфическими свойствами, которых лишены другие клетки и которые связаны с особой функцией нейронов как передатчиков нервных импульсов; сюда относятся необходимость в поддержании ионных градиентов, что требует большой затраты энергии, и свойства, связанные со способностью нейронов производить и выделять набор химических передатчиков, называемых нейромедиаторами. В синапсах - микроскопических участках, где тесно соприкасаются окончание одного нейрона и воспринимающая поверхность другого, приход импульса вызывает внезапное выделение молекул медиатора из, окончания. Затем эти молекулы диффундируют через заполненную жидкостью щель между двумя клетками и воздействуют на специфические рецепторы постсинаптической мембраны, изменяя при этом электрическую активность воспринимающего нейрона.
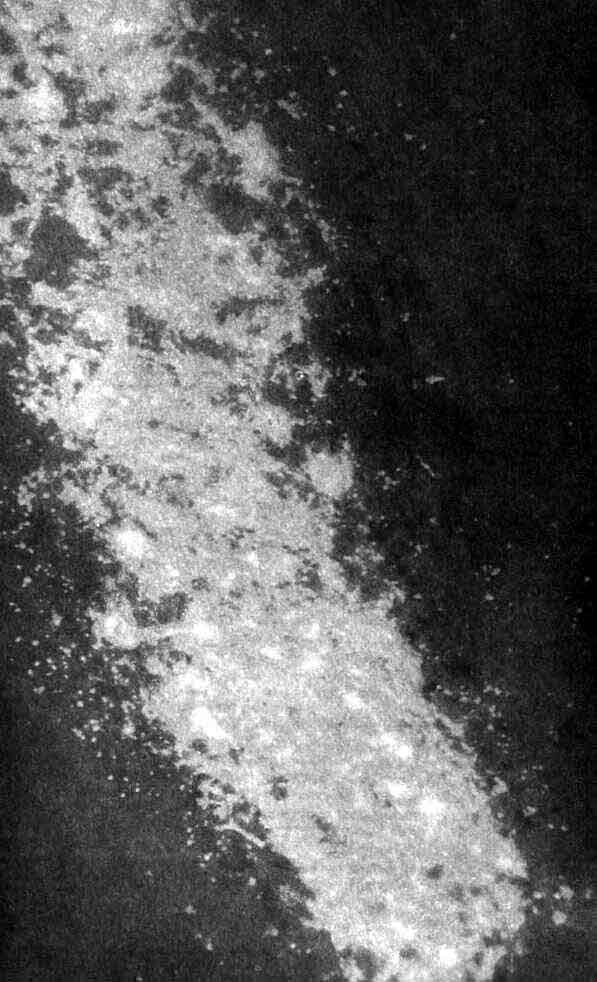
Нейроны, содержащие норадреналин - химический медиатор в головном мозгу, - ярко светятся на этом срезе мозга крысы под флуоресцентным микроскопом. Такие клетки, расположенные в участке мозга, именуемом locus coeruleus, стали видны под воздействием глиоксиловой кислоты, которая превращает норадреналин в его флуоресцирующее производное. В этом поле находятся еще тысячи других нейронов, но в них содержатся другие медиаторы и поэтому они не видны. Норадреналиновые нейроны в locus coeruleus посылают свои аксоны во многие отделы мозга, в том числе в мозжечок и передний мозг. Как полагают, они имеют отношение к регуляции сна, настроения, а также к системе поощрения. Микрофотография получена Ф. Блумом (F. Bloom), Г. Джонсом (G. Jones) и Ж. Мак-Джинти (J. McGinty) из Института Солка.
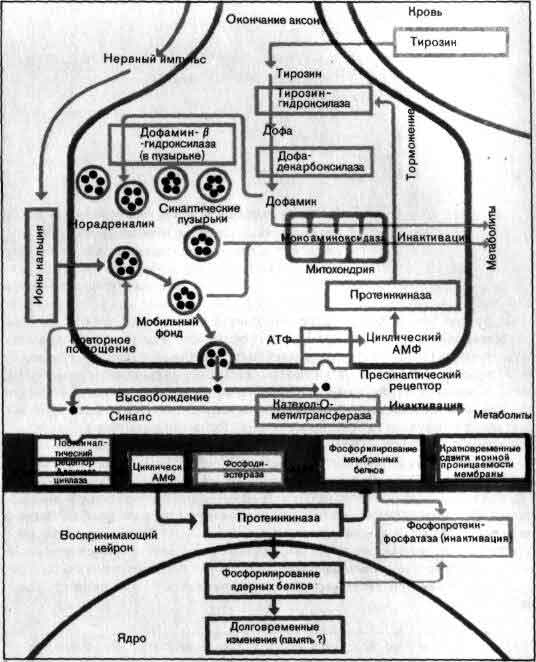
Химическая передача через синапс - узкую щель между двумя нейронами - в головном мозгу состоит из сложной последовательности молекулярных процессов. На рисунке дана схема процесса передачи в норадреналиновом синапсе. Сначала в три стадии происходит синтез норадреналина из аминокислоты тирозина, причем каждая стадия катализируется особым ферментом. Затем медиатор в сочетании с белками накапливается в пузырьках, примыкающих к мембране. Приходящий в аксонное окончание нервный импульс запускает приток ионов кальция, который вызывает высвобождение норадреналина из пузырьков в синаптическую щель. Молекулы медиатора связываются со специфическими рецепторными белками, включенными в постсинаптическую мембрану, запуская серию реакций, которая заканчивается кратковременными (электрическими) и долговременными воздействиями на воспринимающий нейрон. После этого действие норадреналина прекращается различными способами, в том числе быстрым возвращением медиатора в аксонное окончание и разрушением его ферментами. Выход некоторого количества норадреналина в синаптическую щель активирует пресинаптические рецепторы на аксонном окончании, вызывая выработку циклического АМФ, который активирует белковую киназу, стимулируя тем самым новую выработку норадреналина.
Известно около 30 разных веществ, относительно которых доказано или подозревается, что они играют роль медиаторов в головном мозгу, и каждое из них оказывает на нейроны характерный возбуждающий или тормозный эффект. Медиаторы распределены в мозгу не в случайном порядке, а локализованы в особых группах нейронов, аксоны которых идут к другим высокоспециализированным областям мозга. Наложение этих разнообразных химически закодированных систем на нейронные сети наделяет головной мозг еще одним измерением модуляции и специфичности.
За последние годы достигнуты значительные успехи в изучении различных медиаторных веществ (хотя многие из них, несомненно, еще не открыты), в составлении карт их распределения по мозгу и в выяснении молекулярных процессов синаптической передачи. Такими исследованиями установлено, что действие многих лекарственных веществ и нейротоксинов на поведение основано на их способности прерывать или модифицировать химическую передачу от нейрона к нейрону. В них есть также указания на то, что причинами психических болезней, возможно, окажутся в конечном счете нарушения функции специфических медиаторных систем мозга.
Что касается общего энергетического обмена, то из всех органов тела головной мозг является самым активным потребителем энергии, что отражается в его обильном кровоснабжении и интенсивном потреблении кислорода. Мозг настолько интенсивно использует кислород (50 миллилитров в минуту), что, составляя всего 2% общего веса тела, поглощает примерно 20% поступающего в организм кислорода.Такое огромное потребление энергии, как полагают, объясняется необходимостью поддерживать ионные градиенты по обе стороны нейронной мембраны, от чего зависит проведение импульсов в миллиардах нейронов мозга. Кроме того, это потребление энергии идет непрерывно: интенсивность метаболизма в мозгу относительно постоянна днем и ночью и иногда даже несколько возрастает во время фазы сна со сновидениями. Однако, чтобы не создалось ошибочного представления, следует сказать, что весь энергетический эквивалент метаболизма мозга составляет всего около 20 ватт.
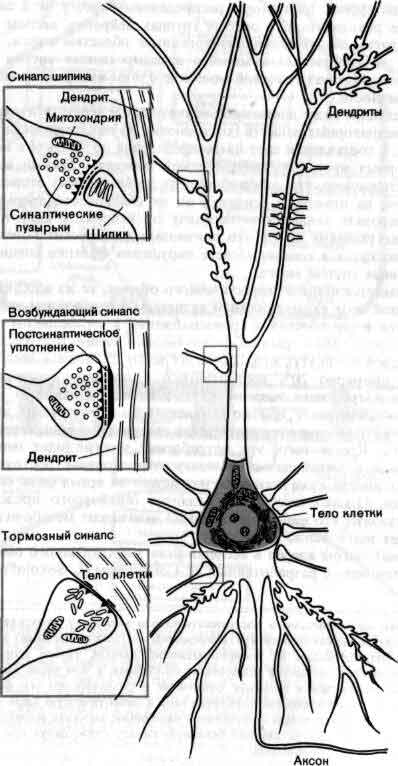
Синапсы на типичном нейроне в головном мозгу являются либо возбуждающими, либо тормозными, в зависимости от типа выделяющегося в них медиатора. Они различаются морфологически под электронным микроскопом: для возбуждающих синапсов характерны сферические пузырьки и сплошное утолщение постсинаптической мембраны, а для тормозных — уплощенные пузырьки и несплошное утолщение мембраны. Синапсы можно также классифицировать по их положению на поверхности воспринимающего нейрона — на теле клетки, на стволе или «шипике» дендрита, или на аксоне.
Важным шагом вперед в исследовании энергетического обмена мозга является метод, разработанный Л. Соколовым (L. Sokoloff) с сотрудниками в Национальном институте охраны психического здоровья. Этот метод позволяет визуально определять интенсивность энергетического обмена в клетках мозга. Нейроны приспосабливают потребление глюкозы к удовлетворению своих метаболических потребностей в данный момент. Следовательно, в активном состоянии они поглощают ее быстрее, чем в покое. Поглощенная глюкоза обычно претерпевает быстрые превращения; ее химический аналог, 2-дезоксиглюкоза, поглощается клетками точно так же, но не подвергается метаболизму. Если ввести в кровь дезоксиглюкозу с радиоактивной меткой, то она накопится в нейронах, и скорость накопления послужит показателем метаболической активности клетки. Накопление радиоактивной дезоксиглюкозы можно установить и измерить, накладывая тонкие срезы замороженного мозга на радиочувствительную пленку. На проявленной пленке выявляются участки, богатые меченым веществом. Эта методика открыла совершенно новую область исследований мозга, поскольку она позволяет установить, какие клетки в головном мозгу были активны во время данного эксперимента. Например, воздействуя световым стимулом (вспышка) на правый или левый глаз, можно определить, какие именно области мозга получают зрительную афферентацию от того или другого глаза.
В отличие от других органов тела, способных использовать разные виды «топлива» (сахара, жиры и аминокислоты), нейроны используют только глюкозу крови. Кроме того, в отличие от таких тканей, как мышцы, способных кратковременно функционировать в отсутствие кислорода, головной мозг полностью зависит от окислительного метаболизма. Если приток окисленной крови к мозгу прекратится, то через 10 секунд наступит потеря сознания, а затем появятся стойкие нарушения. Подобный же эффект вызывает любое состояние, сопровождающееся понижением содержания глюкозы в крови, например гипогликемия у больного диабетом, вызванная передозировкой инсулина. Хотя тонкие регуляционные механизмы обеспечивают постоянство кровяного давления и постоянный уровень кислорода и глюкозы в крови, очевидно, что чрезвычайная гибкость поведения, ставшая возможной благодаря большим размерам и емкости головного мозга млекопитающих, приобретена в процессе эволюции ценой высоких метаболических затрат.
Что касается клеток, то нейроны чрезвычайно чувствительны: их активность нарушается токсическими веществами, попавшими в кровоток, а также мелкими молекулами, обычно присутствующими в крови, например аминокислотами. Такая чувствительность, возможно, служит причиной того, почему головной мозг отделен от общего кровообращения избирательной фильтрационной системой, называемой гематоэнцефалическим барьером. Эффективность этого барьера объясняется относительной непроницаемостью кровеносных сосудов головного мозга и наличием плотного слоя глиальных клеток (опорных мозговых клеток) вокруг них. Хотя такие мелкие молекулы, как молекулы кислорода, легко проникают сквозь барьер, большинство более крупных молекул, необходимых клеткам мозга, например молекулы глюкозы, должны захватываться активно с помощью специальных транспортных механизмов. О гематоэнцефалическом барьере следует помнить при создании лекарственных средств, действующих непосредственно на мозг: для того чтобы такие вещества проходили через барьер, они должны состоять из очень малых молекул или быть легко растворимыми в жировых мембранах глиальных клеток. Несколько участков мозга не защищены тематоэнцефалическим барьером; к ним относятся такие структуры, которые специфически реактивны в отношении содержащихся в крови гормонов, и такие, функция которых состоит в регуляции химического состава крови.
Внутри отдельных нейронов существуют свои транспортные проблемы, связанные с тем, что клетка частично состоит из длинных, тонких волокон. Длина аксона, проводящего нервный импульс от тела нейрона, может исчисляться в миллиметрах или сантиметрах. Нейроны в мозгу взрослого животного незаменяемы, сохраняются на протяжении всей жизни; поэтому должны существовать механизмы возобновления всех их компонентов. Для выполнения этого требования клетка должна синтезировать ферменты и другие сложные молекулы, а такой синтез возможен только в районе клеточного ядра, т. е. в теле нейрона. Поэтому для возобновления своих компонентов аксону необходимо средство транспортировки их внутри клетки на значительные расстояния. И действительно, от тела клетки по всей длине аксона непрерывно движутся белки и другие составные части.
Феномен аксонного транспорта был открыт более 30 лет назад П. Вейсом (P. Weiss) и его сотрудниками из Чикагского университета. До этого все считали, что аксоплазма - желеобразная жидкость внутри аксона - служит лишь неподвижной механической опорой для возбудимой мембраны, по которой распространяется нервный импульс. Но когда Вейс пережал аксон в определенной точке, он отметил, что через несколько дней волокно разбухло около места сжатия со стороны тела клетки и сузилось с противоположной стороны. Когда же он устранил сжатие, задержанная аксоплазма возобновила свое течение.
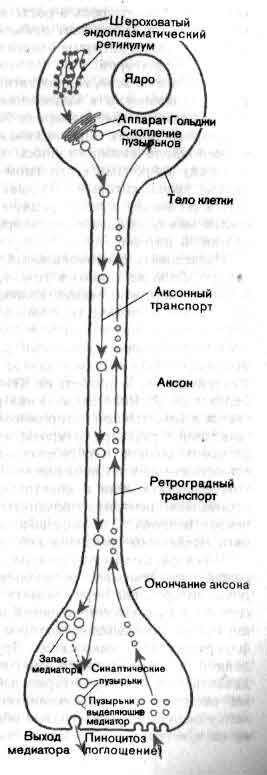
Аксонный транспорт обеспечивает передвижение компонентов клетки, таких, как пузырьки и ферменты, от места их образования в теле нейрона к окончанию аксона на расстояние в несколько миллиметров или сантиметров. В обратном направлении - от окончаний аксона к телу клетки-переносятся вещества, нужные для поддержания нормального состояния нейрона. Аксонный транспорт можно использовать, чтобы проследить ход проводящих путей, наблюдая за движением вдоль аксонов меченных изотопами молекул или таких ферментов, как пероксидаза хрена.
Известно, что аксоплазма служит артерией для оживленного движения молекул в обе стороны между телом нервной клетки и окончаниями ее аксона. В этом принимают участие несколько разных систем, в том числе система медленного транспорта, когда вещество течет от тела клетки со скоростью около одного миллиметра в сутки, и система быстрого транспорта, когда вещество движется в обе стороны со скоростью от 10 до 20 сантиметров в сутки. Система медленного транспорта - это течение основной массы аксоплазмы с переносом компонентов, играющих важную роль в росте и регенерации аксона; система быстрого транспорта охватывает движение более специализированных компонентов клетки, к которым относится ряд ферментов, участвующих в выработке медиаторов.
Пока еще не ясно, как достигаются эти разные скорости транспорта, но, по-видимому, и в медленном и в быстром механизмах участвуют многочисленные фибриллярные белки аксона, видимые на электронных микрофотографиях. Прослеживая транспорт белков с радиоактивной меткой вдоль аксона, удалось точно установить анатомические связи между нейронами в головном мозгу. При помощи фермента пероксидазы хрена, который обладает особым свойством быстро передвигаться по аксону в ретроградном направлении, можно составить карту связей между окончаниями нейрона в одном участке мозга и телом клетки — в другом.
Исследовать функциональный химизм головного мозга чрезвычайно трудно. Дело не только в том, что медиаторные вещества содержатся в нем в ничтожно малых количествах, но и в том, что ткань мозга структурно и химически очень сложна, и выделить для исследования определенную медиаторную систему не легко. Однако для преодоления этих устрашающих препятствий разработаны новые методики. Большой шаг вперед был сделан в начале 1960-х годов благодаря открытию В. Уиттейкера (V. Whittaker) из Кембриджского университета и Э. де Робертиса (Е. de Robertis) из Университета в Буэнос-Айресе. Оно заключается в том, что при осторожном разрушении ткани мозга путем гомогенизации в растворе сахарозы многие нервные окончания отрываются от своих аксонов и образуют особые замкнутые частицы, названные «синаптосомами». Синаптосомы содержат механизмы синтеза, хранения, высвобождения и инактивации медиатора, связанные с нервным окончанием; центрифугированием их можно очистить от других компонентов нейрона. Эта методика дала нейрохимикам возможность изучать механизмы синаптической передачи в пробирке.
Пожалуй, самым плодотворным методическим достижением явилась разработка приемов, позволяющих избирательно окрашивать нейроны, содержащие определенный медиатор. Один из подходов к этому состоит в том, что естественный медиатор преобразуется в его флуоресцирующее производное, которое светится в ультрафиолетовых лучах флуоресцентного микроскопа. Другой подход состоит в том, что в головной мозг подопытного животного вводят молекулы медиатора с радиоактивной меткой, которые избирательно захватываются нейронными окончаниями, выделяющими этот медиатор; такие содержащие метку окончания можно затем обнаружить, поместив тонкие срезы ткани на чувствительную пленку. Третий подход использует высокую специфичность антител. Фермент, участвующий в синтезе данного медиа-гора, выделяют из ткани мозга и вводят в организм животного, где он вызывает образование антител, способных специфически реагировать с данным ферментом. Затем антитела подвергают очистке, метят флуоресцирующим красителем или используют какую-либо другую метку и применяют для избирательного окрашивания нейронов, содержащих данный фермент.
Эти методики избирательного окрашивания позволили получить поток сведений о детальном анатомическом распределении отдельных медиаторов в сложных нейронных цепях головного мозга. Они показали, что медиаторы распределены не диффузно по всей ткани мозга, а в высшей степени локально в ограниченных центрах и путях. Из всех медиаторов наиболее полно составлены карты для моноаминов - норадреналина, дофамина и серотонина (названных так. потому что все они содержат по одной аминогруппе). Б. Фальк (В. Falck) из Лундского университета и Н. Хжлларп (N. Hillarp) из Каролинского института в Швеции впервые показали, что воздействуя формальдегидом или глиоксиловой кислотой, можно превратить эти медиаторы в их флуоресцирующие производные, и тогда нейроны, содержащие моноамины, дают зеленую или желтую флуоресценцию. Такие работы показали, что многие содержащие норадреналин клетки мозга сосредоточены в стволе и образуют скопление, известное как locus coeruleus. Аксоны этих нейронов сильно ветвятся и проецируются в различные области — гипоталамус, мозжечок и передний мозг. Норадреналиновые нейроны оказались причастными к поддержанию бодрствования, к системе поощрения (центр удовольствия), к сновидениям и к регуляции настроения.
Нейроны, содержащие моноамин дофамин, сосредоточены в областях среднего мозга, известных под названиями substantia nigra и вентральной покрышки. Многие содержащие дофамин нейроны посылают свои аксоны в передний мозг, где, как предполагают, участвуют в регуляции эмоциональных реакций. Другие дофаминовые волокна оканчиваются в головном мозгу, в области полосатого тела. Здесь дофамин, очевидно, играет решающую роль в регуляции сложных движений. Дегенерация приходящих в эту область дофаминовых волокон приводит к ригидности мышц и тремору, столь характерным для болезни Паркинсона.
Моноаминовый медиатор серотонин сосредоточен в области ствола мозга, где находятся так называемые «ядра шва». Нейроны этого центра проецируются в гипоталамус, таламус и многие другие области мозга. Как полагают, серотонин участвует в терморегуляции, сенсорном восприятии и засыпании.
Идентифицировано еще много других медиаторов, часть из них считаются «предполагаемыми», поскольку их причастность к синаптической передаче в мозгу еще не доказана. Так, например, некоторые аминокислоты - блоки, из которых построены белки, - по-видимому, действуют как медиаторы. Обычно содержащиеся в организме в большом количестве аминокислоты - глутаминовая и аспарагиновая - оказывают сильное возбуждающее действие на большинство нейронов, и весьма возможно, что они являются наиболее распространенными возбудительными медиаторами в синапсах головного мозга. Известно, что самая простая из всех аминокислот, глицин, служит тормозным медиатором в спинном мозгу.
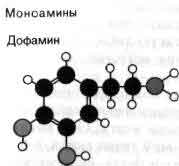
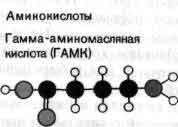

Медиаторы представляют собой вещества, молекулы которых имеют небольшой размер и положительно заряженный атом азота. Каждое такое вещество оказывает характерное либо возбуждающее, либо тормозное действие на нейроны; правда, есть медиаторы, которые действуют как возбуждающие в одной области мозга и как тормозные в другой. Принадлежность гистамина и таурина к медиаторам точно не доказана. Согласно принципу Дэйла, на всех окончаниях одного аксона выделяется один и тот же медиатор. Однако недавно обнаружены исключения из этого правила.
Дофаминовые проводящие пути показаны на рисунке схематически. Тела нейронов, содержащих дофамин, образуют скопления в двух небольших участках среднего мозга - substantia nigra и покрышке. От этих нейронов отходят широко ветвящиеся волокна, которые оканчиваются в полосатом теле, регулирующем двигательную активность, и в лимбической системе переднего мозга, связанной с эмоциями. Небольшая группа дофаминовых нейронов в гипоталамусе регулирует также секрецию гормонов гипофизом. Дофамин связывают с двумя заболеваниями: дефицит медиатора в полосатом теле создает ригидность и тремор, характерные для болезни Паркинсона, а избыток дофамина в лимбической системе переднего мозга, возможно, причастен к шизофрении.
К обычным тормозным медиаторам головного мозга относится гамма-аминомасляная кислота (ГАМК), которая не входит в состав белков. ГАМК занимает среди кислот особое место — она вырабатывается почти исключительно в головном и спинном мозгу. Вычислено, что не менее трети синапсов головного мозга используют в качестве медиатора ГАМК. Содержащие ее нейроны можно идентифицировать двумя способами: пометив их либо радиоактивной ГАМК, либо с помощью антител, специфичных в отношении глутаматдекарбоксилазы - фермента, который катализирует синтез ГАМК. Интересно отметить, что глутаминовая кислота считается кандидатом в возбудительные медиаторы, тогда как ГАМК, которая отличается от нее лишь на одну химическую группу, служит тормозным медиатором. Ясно, что небольшие различия в молекулярном строении медиаторов могут давать совершенно разные физиологические эффекты.
Исследование механизмов действия ГАМК в головном мозгу было стимулировано в последние годы открытием, сделанным недавно Т. Перри (Т. Perry) из Университета Британской Колумбии. Оно состоит в том, что для хореи Гентингтона - наследственного неврологического синдрома — характерен специфический дефицит ГАМК в мозгу. Возникающие при этой болезни непроизвольные движения связаны с наступающим в среднем возрасте постепенным разрушением полосатого тела. Посмертные исследования показали, что поражение мозга состоит в гибели тормозных нейронов, которые в норме содержат ГАМК; можно предположить, что в основе болезни лежит дефицит этого медиатора. К сожалению, попытка лечить больных заменой недостающей ГАМК пока неосуществима, поскольку еще не созданы аналоги ГАМК, способные проходить сквозь гематоэныефалический барьер.
Недавно было высказано предположение, что на ГАМК направлено действие таких транквилизаторов, как диазепам и другие производные бензодиазепина. Из всех психотропных препаратов врачи предпочитают прописывать бензодиазепины, но механизм их действия до сих пор неизвестен. Имеющиеся данные позволяют думать, что эти препараты повышают эффективность ГАМК на уровне ее рецепторов в головном мозгу. Хотя в мозгу идентифицированы специфические центры связывания диазепама, явно отличные от рецепторов ГАМК, оба типа рецепторов, по-видимому, взаимодействуют друг с другом. Заманчиво предположить, что в головном мозгу содержится некое еще не открытое вещество, которое в норме действует на рецепторы диазепама; возможно, это естественное соединение, вызывающее или ослабляющее состояние тревоги.
Нейрохимики не только выяснили молекулярную структуру и анатомическое распределение разных медиаторов, но и достигли больших успехов в понимании точной последовательности биохимических явлений, участвующих в синаптической передаче. Процесс химической передачи проходит через ряд этапов: синтез медиатора, его накопление, высвобождение, взаимодействие с рецептором и прекращение действия медиатора. Каждый из этих этапов детально охарактеризован, и найдены препараты, которые избирательно усиливают или блокируют конкретный этап. Эти исследования позволили проникнуть в механизм действия психотропных лекарственных средств, а также выявить связь некоторых нервных и психических болезней со специфическими нарушениями синаптических механизмов.
Первым этапом химической передачи является синтез молекул медиатора в нервных окончаниях. Каждый нейрон обычно обладает только таким биохимическим аппаратом, какой ему нужен для синтеза медиатора одного типа, который выделяется из всех окончаний его аксона. Молекулы медиатора не синтезируются de novo, а изготовляются путем переработки предшественника — обычно аминокислоты — в результате ряда ферментативных реакций.
Создание медиатора требует или одного этапа ферментативного катализа (как в случае ацетилхолина), или до трех этапов (как адреналина). При синтезе норадреналина исходным веществом служит аминокислота тирозин, которая поступает в нервное окончание из крови. Тирозин сначала превращается в промежуточное соединение L-ДОФА; затем второй фермент превращает L-ДОФА в дофамин («полноправный» медиатор); а третий фермент превращает дофамин в норадреналин.
После выработки молекул медиатора они хранятся в окончании аксона в маленьких связанных с мембраной мешочках, называемых синаптическими пузырьками. В одном окончании могут быть тысячи синаптических пузырьков, а каждый пузырек содержит от 10000 до 100000 молекул медиатора. Пузырьки защищают их от разрушения ферментами внутри окончания.
Приход нервного импульса в окончание аксона вызывает высвобождение множества молекул медиатора из окончания в синаптическую щель. Механизм такого выделения остается спорным: одни исследователи полагают, что синаптические пузырьки прямо сливаются с пресинаптической мембраной и выбрасывают свое содержимое в синаптическую щель; другие утверждают, что подвижное скопление молекул медиатора выходит через специальные каналы. Но в любом случае известно, что нервный импульс запускает выход медиатора, повышая проницаемость нервного окончания для ионов кальция, которые устремляются в него и активируют механизм высвобождения молекул.
Вышедшие молекулы медиатора быстро проходят через наполненную жидкостью щель между окончанием аксона и мембраной воспринимающего нейрона. Здесь они взаимодействуют со специфическими рецепторами постсинаптической мембраны. Рецепторы фактически представляют собой крупные белковые молекулы, погруженные в полужидкую матрицу клеточной мембраны: части их торчат над и под мембраной подобно айсбергам. Выходящий на поверхность участок рецепторного белка и молекула медиатора имеют одинаковые очертания; они соответствуют друг другу наподобие ключа и замка.
Взаимодействие медиатора с его рецептором меняет трехмерную форму рецепторного белка, инициируя этим определенную последовательность событий. Это взаимодействие может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, а также образование и выделение гормона клеткой железы. Во всех этих случаях рецептор переводит сообщение, закодированное в молекулярной структуре медиатора, в специфическую физиологическую реакцию. Одни реакции, например сокращение произвольной мышцы, происходят за долю секунды; другие, например секреция гормона, занимают минуты, а иногда часы.
Многие рецепторы медиаторов имеют два функциональных компонента: центр связывания молекулы медиатора и пору, пронизывающую мембрану, избирательно проницаемую для определенных ионов. Связываясь с рецептором, медиатор меняет его форму так, что пора открывается и ионы, находящиеся внутри и снаружи клеточной мембраны, перемещаются вдоль градиента концентрации, оказывая этим возбуждающий или тормозный эффект на частоту импульсации нейрона. Будет ли электрический потенциал, создаваемый медиатором, возбудительным или тормозным, зависит от того, какие именно ионы перемещаются, и от направления их движения. Ацетилхолин является возбуждающим медиатором в синапсе между нервом и мышцей, потому что он заставляет положительно заряженные ионы натрия входить в клетку и понижать ее отрицательный потенциал покоя. ГАМК, напротив, соответствует рецептор, у которого пора избирательно проницаема для отрицательно заряженных ионов хлора. Когда эти ионы входят через открытые поры в воспринимающую клетку, они повышают трансмембранный потенциал и на время инактивируют клетку.
Другие медиаторы, например дофамин и норадреналин, по-видимому, действуют посредством более тонкого механизма. В середине 50-х годов И. Сьюзерленд мл. (Е. Sutherland, Jr.) и его коллеги по Университету Кейса Западной резервной территории показали, что эти и другие медиаторы повышают или снижают концентрацию «второго посредника» в воспринимающих клетках. Затем второй посредник передает электрические или биохимические эффекты медиатора, или «первого посредника». Позднее в работе, которая в 1971 г. принесла Сьюзерленду Нобелевскую премию по физиологии и медицине, он установил, что второй посредник представляет собой небольшую молекулу циклического аденозинмонофосфата (цАМФ).
Согласно гипотезе Сьюзерленда, белковый рецептор норадреналина (и многих других медиаторов) соединяется в мембране клетки-мишени с ферментом аденилатциклазой, которая катализирует превращение в клетке богатой энергией молекулы аденозинтрифосфата (АТФ) в цАМФ. Аденилатциклаза обычно неактивна, но, когда норадреналин связывается с постсинаптическим рецептором, фермент автоматически включается и внутри клетки начинается быстрое превращение АТФ в цАМФ. Затем цАМФ действует на биохимический аппарат клетки, вызывая физиологическую реакцию, характерную для данного медиатора.
Система второго посредника сходна поэтому с эстафетой, в которой медиатор передает свое сообщение циклическому АМФ в мембране клетки. Разумеется, сигнал передается не одной, а многим тысячам молекул цАМФ, которые генерируются активированной аденилатциклазой, связанной с каждым занятым рецептором. В результате очень слабый сигнал, создаваемый взаимодействием медиатора с рецептором, усиливается внутри клетки в несколько тысяч раз благодаря интенсивному образованию цАМФ.
Приложение теории Сьюзерленда о втором посреднике к функции головного мозга является одной из самых увлекательных областей современной биохимии. В 1971 г. Ф. Блум (F. Bloom) с сотрудниками в Национальных институтах здравоохранения показали, что цАМФ способен влиять на генерацию сигналов в нейронах. Позднее П. Грингард (P. Greengard) и его группа в Медицинской школе Йельского университета пришли к заключению, что цАМФ участвует в синаптическом действии нескольких медиаторов, в том числе норадреналина, дофамина, серотонина и гистамина. Грингард предложил обобщающую гипотезу, согласно которой цАМФ активирует специфические ферменты в постсинаптической клетке, именуемые белковыми киназами; затем эти ферменты катализируют внедрение фосфатных групп в специальные белки в мембране нейрона, изменяя проницаемость мембраны для ионов и тем самым изменяя уровень возбудимости клетки-мишени. Поскольку система второго посредника работает сравнительно медленно по шкале времени нейронных событий, она больше всего пригодна для участия в более длительных эффектах медиаторов мозга, таких, как медленные сдвиги мембранного потенциала и, возможно, образовании следов долговременной памяти. Как только цАМФ передал свое сообщение дальше, он инактивируется в клетке под действием фермента фосфодиэстеразы. Поэтому препараты, ингибирующие этот фермент, повышают уровень цАМФ в постсинаптических клетках и усиливают действие медиатора.
Итак, по-видимому, существуют два основных типа медиаторных рецепторов: быстро действующие рецепторы, которые осуществляют передачу информации, регулируя проницаемость ионной поры, и медленно действующие рецепторы, которые вызывают образование второго посредника; последний в свою очередь опосредует эффекты, производимые медиатором в постсинаптическом нейроне. Для многих медиаторов имеется по два и более типов рецепторов. Например, реакция на ацетилхолин в синапсе между мотонейроном и мышечной клеткой осуществляется простым током ионов натрия через мембрану. Но в головном мозгу эффекты ацетилхолина в большинстве своем опосредуются, по-видимому, молекулой еще одного второго посредника, циклического гуанозинмонофосфата, или цГМФ. Точно так же недавно полученные данные позволяют думать, что дофамин действует на уровне двух разных типов рецепторов в головном мозгу: рецептора D1, который связан с системой второго посредника цАМФ, и рецептора D2, который с ней не связан.
Как только молекула медиатора свяжется со своим рецептором, она должна быть быстро инактивирована во избежание слишком длительного ее действия и нарушения точного контроля передачи. Нервные волокна способны проводить несколько сот импульсов в секунду только при условии, что постсинаптическая мембрана восстанавливает свой потенциал покоя за долю миллисекунды. Некоторые медиаторы инактивируются ферментами, находящимися в синаптической щели. Ацетилхолин, например, разрушается ферментом ацетилхолинэстеразой, которая за секунду расщепляет 25000 молекул медиатора. Норадреналин инактивируется в синапсе по совсем иному механизму.
Ю. Аксельрод (J. Axelrod) с сотрудниками из Национальных институтов здравоохранения установили, что выделившийся из аксонного окончания норадреналин снова быстро всасывается в окончание. Затем поглощенные молекулы норадреналина либо разрушаются ферментами - катехол-О-метилтрансферазой (КОМТ) и моноаминоксидазой (МАО), которые содержатся в нервном окончании, либо возвращаются обратно в синаптические пузырьки. Позднее такие же механизмы возврата были найдены и для других медиаторов — дофамина, серотонина и ГАМК. Возврат имеет то очевидное преимущество перед разрушением фермента, что молекулы медиатора сохраняются в течение нескольких циклов выделения и поглощения.
Выяснение этапов синаптической передачи пролило свет на способ действия психотропных препаратов. Некоторые из них действуют, либо усиливая, либо ослабляя высвобождение данного медиатора из аксонных окончаний. Например, под действием сильного стимулятора амфетамина в мозгу из нервных окончаний выделяется дофамин - медиатор, связанный с системами бодрствования и удовольствия. Чрезмерное применение амфетамина может привести к расстройствам мышления, галлюцинациям и мании преследования, т.е. к симптомам, очень сходным с теми, какие возникают при некоторых формах шизофрении. Эти и другие данные послужили основанием для предположения о том, что в основе симптомов шизофрении, возможно, лежит повышенная активность дофаминовых систем мозга.
Вызывает интерес также тот факт, что разнообразные лекарственные средства, созданные для лечения шизофрении, например хлорпромазин (торазин) и галоперидол (галдол), обладают общим свойством — они тесно связываются с дофаминовыми рецепторами головного мозга, тем самым мешая естественному медиатору активировать их. Это открытие оказалось одним из самых многообещающих в современном исследовании шизофрении. Последние данные позволяют думать, что шизофрения связана с избыточным образованием дофамина или с чрезмерной реактивностью в отношении медиатора в определенных участках головного мозга. Работами моей лаборатории в Отделе нейрохимической фармакологии Британского совета медицинских исследований и работами Т. Кроу (Т. Crow) в Центре клинических исследований Британского совета медицинских исследований в Лондоне, а также исследованиями Ф. Симена (Ph. Seeman) из Торонтского университета обнаружены необычно высокие концентрации дофамина в дофаминовых рецепторах в головном мозгу умерших больных шизофренией; это особенно относится к лимбической системе, т. е. тем областям мозга, которые ответственны за эмоциональное поведение. Поэтому возможно, что главной мишенью антипсихотических препаратов служат прежде всего пути дофамина в этих областях.
Многие психотропные препараты, возможно, действуют на уровне постсинаптических рецепторов, имитируя естественные медиаторы. Например, многие галлюциногены сходны по своей структуре с истинными медиаторами: мескалин похож на норадреналин и дофамин, a LSD и псилоцибин сходны с серотонином. Поэтому возможно, что перечисленные препараты действуют на моноаминовые механизмы, хотя способ их действия точно еще неизвестен. LSD выделяется своей необычайно высокой активностью: всего 75 микрограммов (едва видимой пылинки) достаточно чтобы вызвать галлюцинации.
Производные метилксантина кофеин и теофиллин, как полагают, действуют через систему второго посредника. Они специфически ингибируют фермент фосфодиэстеразу, который разрушает цАМФ, и таким образом в конечном счете увеличивают количество цАМФ, образующегося в ответ на действие медиатора. В результате эти препараты оказывают на головной мозг общее мягкое стимулирующее действие. Кофеин является главным действующим началом кофе и чая; более слабый стимулятор теофиллин содержится преимущественно в чае. Ежегодное потребление кофе и чая составляет миллиарды фунтов в год, так что метилксантины являются самыми широко потребляемыми препаратами.
Наконец, некоторые препараты потенцируют действие медиатора, блокируя его разрушение в синапсе. Одна из таких групп препаратов представлена ипрониазидом (марсилидом) и другими соединениями, ингибирующими фермент моноаминоксидазу, который разрушает норадреналин, дофамин и серотонин. Вследствие блокады этого фермента эффекты перечисленных моноаминов усиливаются, чем объясняется антидепрессивное действие таких лекарств. Вторая группа антидепрессантов - трициклические соединения - также усиливает действие норадреналина и серотонина в головном мозгу. Эти лекарственные средства, из которых лучше всего известны имипрамин (тофранил) и амитриптилин (элавил), блокируют обратный захват норадреналина и серотонина из синаптической щели; стимулятор кокаин, по-видимому, действует по тому же механизму. Указанные наблюдения говорят о том, что депрессия, по-видимому, связана с низким уровнем этих медиаторов.
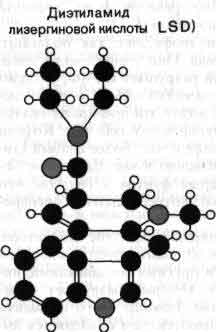
Галлюциногены по своей структуре очень похожи на моноаминовые медиаторы; это позволяет допустить, что они оказывают сильные воздействия на сознание, имитируя действие этих естественных медиаторов на синаптические рецепторы в головном мозгу. Мескалин содержит в своей структуре бензольное кольцо подобно дофамину и норадреналину. а псилоцибин и LSD - индольное кольцо серотонина.
За последние годы, после того как в мозгу был обнаружен новый класс химических соединений нейропептиды, число известных систем химических посредников в головном мозгу резко возросло. Эти молекулы представляют собой цепочки из аминокислотных остатков (от двух до 39); они обнаружены в нейронах и предположительно являются медиаторами. Некоторые из них были вначале идентифицированы как гормоны, выделяемые гипофизом (АКТГ, вазопрессин), как локальные гормоны кишечника (гастрин, холецистокинин) или как гормоны, выделяемые гипоталамусом для регуляции выделения других гормонов гипофизом — люлиберина (гормона, стимулирующего выделение лютеинизирующего гормона), соматостатина.
Самыми новыми и вызывающими наибольший интерес нейропептидами являются энкефалины и эндорфины — вещества, встречающиеся в головном мозгу в норме и удивительно сходные с морфием — наркотиком, извлекаемым из снотворного мака. Вслед за открытием этих пептидов последовало осознание того факта, что некоторые области головного мозга с высокой степенью сродства связывают препараты группы опия. Рецепторы опия обнаружены путем определения связывания меченых его соединений с частицами нейронных мембран. Такие методики нанесения радиоактивных меток на рецепторы разработаны тремя группами исследователей, руководимыми С. Снайдером (S. Snyder) и К. Пертом (С. Pert) в Медицинской школе университета Джонса Гопкинса, Э. Саймоном (Е. Simon) из Медицинской школы Ньюйоркского университета и Л. Терениусом (L. Terenius) из Упсальского университета. Эти исследователи установили, что рецепторы опия сосредоточены в тех участках головного и спинного мозга млекопитающих, которые имеют отношение к восприятию и интеграции боли и эмоций.
Затем в 1975 г. Дж. Хьюз (J. Hughes) и X. Костерлиц (Н. Kosterlitz) в Абердинском университете выделили из головного мозга два естественных пептида, которые тесно связываются с рецепторами опия, и назвали их энкефалинами. Оба энкефалина представляют собой цепочки, состоящие из пяти аминокислот в одинаковой последовательности, за исключением последней аминокислоты: в одном случае это метионин, а в другом — лейцин. Позднее из гипофиза были выделены другие морфиноподобные пептиды, названные эндорфинами. Недавно проведенные опыты показали, что некоторые процедуры, применяемые для устранения хронических болей,- иглоукалывание, прямая электрическая стимуляция головного мозга и даже гипноз, — возможно, действуют, вызывая выделение энкефалинов или эндорфинов в головном и спинном мозгу. Эта гипотеза основана на том, что эффективность всех этих процедур сильно снижается налоксаном (нарконом) — препаратом, который избирательно блокирует связывание морфина с рецептором опия.

Нейропептиды - короткие цепи аминокислот, обнаруженные в ткани мозга. Многие из них локализованы в аксонных окончаниях и высвобождаются из них под влиянием кальция, что дает основание считать их медиаторами. Однако нейропептиды отличаются от ранее идентифицированных медиаторов тем, что они организуют такие сложные явления, как жажда, память и половое поведение. Кроме того, они играют многообразную роль во многих других областях тела. Например, соматостатин подавляет выделение гормона роста гипофизом (у человека), регулирует секрецию инсулина и глюкагона поджелудочной железой и, по-видимому, функционирует в качестве медиатора в спинном и головном мозгу.
Показано, что многие нейропептиды, обнаруженные в головном мозгу млекопитающих, сосредоточены в окончаниях особых групп нейронов, и о некоторых из них известно, что выход их из аксонных окончаний зависит от кальция. Такие данные, а также наблюдение, что очень малые количества нейропептидов оказывают значительный эффект на активность нейронов или на поведение экспериментальных животных, в какой-то мере свидетельствуют в пользу того, что эти вещества вполне могут представлять собой новый класс медиаторов. Однако полученные данные еще не настолько убедительны, чтобы можно было считать это доказанным.
Пожалуй, самым первым кандидатом на статус медиатора среди нейропептидов является вещество Р - цепочка из 11 аминокислот. Оно присутствует во многих специфических нейронных путях в головном мозгу, а также в первичных сенсорных волокнах периферических нервов. Некоторые из этих сенсорных нейронов, тела которых лежат в сенсорных ганглиях по обе стороны спинного мозга, содержат вещество Р и выделяют его из своих аксонных окончаний в синапсах со спинальными нейронами. Поскольку вещество Р возбуждает те спинальные нейроны, которые легче всего реагируют на болевые стимулы, было высказано предположение, что оно служит сенсорным медиатором, специфически связанным с передачей информации о боли от периферических болевых рецепторов в центральную нервную систему.
Морфиноподобный пептид энкефалин тоже в изобилии содержится в мелких нейронах в той части спинного мозга, куда приходят волокна, содержащие вещество Р. Мы с Т. Джесселом (Т. Jessel) в Отделе нейрохимической фармакологии Британского совета медицинских исследований показали, что энкефалин и препараты опия способны подавлять выделение вещества Р из сенсорных волокон. Поэтому нейроны, содержащие энкефалин, могут регулировать поступление болевых сигналов в головной мозг, модулируя выделение вещества Р на уровне первого переключения в центральной нервной системе. Подобные же тормозные взаимодействия возможны и на более высоких уровнях мозга. Вещество Р - не единственный предполагаемый медиатор, локализующийся, как показано, в сенсорных нейронах; к таким же идентифицированным к настоящему времени веществам относятся ангиотензин, холецистокинин, соматостатин и глутаминовая кислота. Таким образом, по мере того как все больше становится известно о сенсорных медиаторах и их модуляционных механизмах в спинном мозгу, начинает возникать картина поразительной химической сложности.
Замечательное свойство нейропептидов мозга состоит в глобальном характере некоторых их эффектов. Введение очень малых количеств нейропептида (обычно прямо в головной мозг, чтобы обойти гематоэнцефалический барьер) вызывает у экспериментального животного сложное и весьма своеобразное поведение. Например, введение в мозг нескольких нанограммов нейропептида ангиотензина II приводит к тому, что животные интенсивно и долго пьют, хотя до этого не испытывали жажды. Другой пептид - гормон люлиберин, при введении его в головной мозг самке крысы создает характерное для самки половое поведение. Еще поразительнее, что, как показал Д. де Виед (D. de Wied) с сотрудниками из Утрехтского университета, введение малых количеств нейропептида вазопрессина лабораторным животным заметно улучшает запоминание действий, которым их обучили. Теперь проводятся предварительные клинические испытания этого препарата, чтобы установить, может ли он дать эффект в случаях потери памяти.
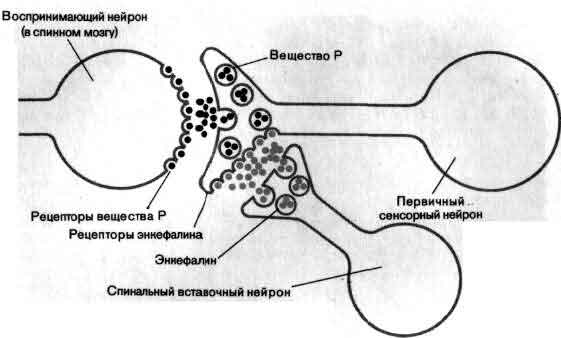
Гипотетический воротный механизм в первом синаптическом переключении, возможно, регулирует передачу информации о боли от периферических болевых рецепторов к головному мозгу. В задних рогах спинного мозга вставочные нейроны, содержащие пептидный медиатор энкефалин, образуют синапсы на аксонных окончаниях болевых нейронов, которые в качестве медиатора используют вещество Р. Выделяемый вставочными нейронами энкефалин тормозит выход этого вещества, из-за чего воспринимающий нейрон в спинном мозгу получает меньше возбуждающей стимуляции и поэтому посылает в головной мозг меньше связанных с болью импульсов. Такие опийные препараты, как морфий, по-видимому, связываются с незанятыми рецепторами энкефалина, имитируя подавление боли, производимое энкефалиновой системой.
Поэтому можно думать, что нейропептиды являются химическими посредниками, отличными от ранее идентифицированных медиаторов: по-видимому, они служат глобальным средством химического кодирования тех форм активности головного мозга, которые связаны с отдельными функциями поддержанием водно-солевого баланса, половым поведением, ощущениями боли или удовольствия. Неожиданным оказалось наблюдение, что биологически активные пептиды, первоначально обнаруженные в желудочно-кишечном тракте - гастрин, вещество Р, вазоактивный кишечный полипептид и холецистокинин - содержатся также и в центральной нервной системе. И наоборот, некоторые пептиды, сначала найденные в головном мозгу, позднее были обнаружены в кишечнике (соматостатин, нейротензин, энкефалины). Следовательно, можно думать, что эти пептиды выполняют множество ролей, действуя как локальные гормоны или медиаторы в желудочно-кишечном тракте и как глобальные медиаторы в головном мозгу. Р. Гиймен (R. Guillemin) из Института Солка высказал предположение, что разнообразные функции нейропептидов объясняются «оппортунизмом» эволюционного процесса, во время которого молекулу, обслуживающую одну функцию, в другое время и в другом месте можно приспособить для другой функции.
Локализация вещества Р в спинном мозгу обезьяны установлена путем обработки ткани специфическими антителами, меченными темным красителем. Краситель обнаруживается только в задних рогах спинного мозга, к которым подходят периферические болевые волокна. Микрофотография получена С. Хаитом (S. Hunt) из Кембриджского университета.
Ряд других веществ, по-видимому, играет роль модуляторов в межнейронных коммуникациях. На высоких уровнях головного мозга в его ткани найдены простагландины, которые состоят из пятичленного углеродного кольца и двух связанных с ним длинных углеродных цепей; простагландины оказывают разнообразное возбуждающее и тормозное действие на нейроны в зависимости от данной молекулярной структуры простагландина и характера клетки-мишени. В отличие от медиаторов с их быстрыми и кратковременными эффектами простагландины вызывают продолжительные сдвиги в поляризации нейронной мембраны; это говорит скорее об их модулирующей, а не о медиаторной функции. Возможно, что они действуют сообща с медиаторами, эффекты которых они мягко видоизменяют.
Еще одна группа веществ выполняет питательную функцию, а не функцию посредников. Считается, что эти «трофические» вещества выделяются нервными окончаниями и поддерживают жизнеспособность клетки-мишени; другие же трофические вещества захватываются нервными окончаниями, переносятся по аксону в ретрогралном направлении и питают этот же нейрон. Хорошо известный факт атрофии мышцы мосле нарушения ее иннервации, возможно, объясняется тем, что мышечные клетки перестают получать необходимые им трофические вещества. Некоторые дегенеративные заболевания мозга, возможно, возникают от того, что центральные нейроны перестают обмениваться трофическими веществами. В настоящее время из трофических веществ лучше всего описан фактор роста нервов — белок, который необходим для дифференцировки и выживания периферических сенсорных и симпатических нейронов и который, возможно, благоприятствует также поддержанию жизнеспособности центральных моноаминоэргических нейронов.
Наряду со все возрастающим числом химических медиаторов, становится также очевидным многообразие разных механизмов, посредством которых медиаторы производят свои эффекты. Например, вместо того чтобы прямо возбуждать или тормозить клетку-мишень, медиатор, выделяющийся из одного нервного окончания, может воздействовать на соседнее нервное окончание, усиливая или снижая выход медиатора из него. Ясно также, что для данного медиаторного вещества возможны Несколько типов рецепторов (на одни из них действуют, а на другие не действуют системы вторых посредников), чем объясняются неодинаковые возбудительные или тормозные эффекты данного медиатора в разных частях мозга. Даже утвердившееся представление [впервые высказанное Г. Дэйлом (Н. Dale)] о том, что из всех своих окончаний нейрон выделяет один и тот же химический медиатор, может утратить свою нерушимость: установлено, что с норадреналином или серотонином в том же нейроне сосуществует ряд нейропептидов. Функциональное значение таких двойных медиаторных систем пока еще не раскрыто. Кроме того, остаются во многом неизвестными те конкретные химические нарушения, которые лежат в основе таких обычных заболеваний, как эпилепсия, старческое слабоумие, алкоголизм, шизофрения и депрессия. Хотя исследование медиаторных систем мозга уже очень много раскрыло в химических механизмах обучения, памяти, сна и настроения, тем не менее ясно, что самые волнующие открытия еще впереди.
Д. ХЬЮБЕЛ, Т. ВИЗЕЛЬ
Центральные механизмы зрения
Путем изучения активности и пространственной организации нейронов первичной зрительной коры выявляется функциональная схема, которая может лежать в основе переработки сенсорной информации в коре
Рассматриваемая как продукт эволюции кора большого мозга должна считаться одним из самых больших достижений в истории всего живого. У позвоночных, стоящих ниже млекопитающих, кора мозга едва выражена, если вообще можно говорить о ее существовании. Внезапно приобретя внушительные размеры у низших млекопитающих, она начинает доминировать в мозгу хищников и взрывоподобным образом увеличивается у приматов; у человека она почти полностью охватывает остальную часть мозга, имея тенденцию затмевать другие отделы. Степень зависимости животного от какого-то органа является показателем важности последнего, причем показатель этот даже более убедителен, чем размер, а зависимость от коры быстро нарастала по мере эволюции млекопитающих. Мышь без коры ведет себя относительно нормально, по крайней мере при поверхностном исследовании; в то же время человек без коры - это почти растение, бессловесное, невидящее, бесчувственное.
Колонки глазодоминантности, одна из двух основных систем, характеризующих функциональную организацию первичной зрительной коры, видны на этом радиоавтографе как периодические светлые участки (срез коры обезьяны-макака, рассматриваемый в темном поле). Данные колонки (в действительности, искривленные подслои коры, видимые здесь в поперечном сечении на срезе мозга, перпендикулярном поверхности) - это участки, в которых все нейроны реагируют более активно на правый глаз, чем на левый; темные участки, разделяющие светлые области, - колонки предпочтения левого глаза. Для получения этого радиоавтографа в правый глаз анестезированного животного была введена меченая аминокислота. Поглощенная телами клеток сетчатки аминокислота доставлялась к клеткам хоры через коленчатое тело - промежуточную станцию мозга. Срез коры был покрыт фотографической эмульсией, которая экспонировалась в течение нескольких месяцев, а затем была проявлена. Засвеченные зерна серебра над радиоактивными участками образовали рассеивающие свет области, которые соответствуют колонкам глазодоминантности.
Колонки глазодоминантности, видимые в фас на радиоавтографе, полученном методом аксонного транспорта, для среза, параллельного, а не перпендикулярного поверхности первичной зрительной коры. Как можно видеть на предыдущем радиоавтографе, самые яркие метки находятся в одном слое складчатой коры - слое IV. Это уровень, на котором оканчиваются аксоны, несущие информацию в кору; соответственно здесь и накапливается метка. Данный срез был сделан в плоскости, тангенциальной колоколовидной поверхности коры, сразу под слоем IV, который в связи с этим выглядит как кольцо с текстурой из приблизительно параллельных ярких полос. Это выявленные радиоактивными метками области глазодоминантности, которые видны теперь сверху, а не сбоку. Действительная ширина областей глазодоминантности, как правило, составляет около 0,4 миллиметра.
Понимание значения этого большого и незаменимого органа пока еще находится в жалком состоянии. Частично это объясняется тем, что он очень сложен, не только структурно, но и функционально, а частично тем, что интуитивные предположения нейробиологов относительно его функций весьма часто оказывались неверными. Однако сейчас перспектива изменяется в связи с тем, что методы усовершенствовались, а исследователи научились тому, как обращаться с огромным числом охваченных сложнейшими связями нейронов, которые являются основными элементами коры, с импульсами, которые они проводят, и с синапсами, с помощью которых они связаны. В настоящей статье мы попытаемся обрисовать современное состояние знаний, касающихся одной из подобластей коры - первичной зрительной проекционной зоны (известной также под названием стриарной коры, или поля 17) - самой простой из тех кортикальных зон, которые связаны со зрением. Это неизбежно приведет нас к рассмотрению соответствующих вопросов зрительного восприятия, так как работу какого-либо органа нелегко отделить от его биологического назначения.
Кора большого мозга - сильно складчатая нервная ткань толщиной около двух миллиметров - представляет собой самую наружную оболочку, покрывающую верхнюю часть полушарий головного мозга и несколько заходящую на их нижнюю поверхность. У человека вся площадь коры, если ее разгладить, равна примерно полутора квадратным футам. (В 1963 г. в своей статье в «Scientific American» один из нас привел для этой площади цифру 20 квадратных футов и был тут же поправлен нашим другом нейроанатомом из Торонто, который сказал, что, по его мнению, эта площадь составляет около 1,5 квадратных фута, «по крайней мере у канадцев».) Складчатость, по-видимому, в основном является результатом того, что такую необычную структуру нужно было упаковать в объеме черепа.
При рассмотрении коры под микроскопом сразу же бросается в глаза огромное множество нейронов: около 105 (100000) на каждый квадратный миллиметр поверхности, откуда можно заключить, что вся кора в целом содержит что-то около 1010 (10 миллиардов) нейронов. Тела клеток образуют полдюжины слоев, причем слои с плотным и редким расположением клеток чередуются. В противоположность столь выраженным различиям в плотности клеток в последовательных слоях коры на различной глубине в плоскости каждого данного слоя распределение явно однородно во всех участках и по всем направлениям. Морфологически кора довольно однородна по двум своим измерениям.
Одно из первых великих прозрений, касающихся организации коры, относится к концу XIX в., когда постепенно осознали, что этот сравнительно однородный пласт ткани подразделяется на некоторое число отдельных областей, сильно различающихся по функции. Свидетельства в пользу этого поступали из клинических, физиологических и анатомических источников. Было замечено, что повреждения мозга, в зависимости от их локализации, могут вызывать либо паралич, либо слепоту, либо потерю тактильной чувствительности, либо потерю речи; при этом слепота бывает общей или проявляющейся только в половине или меньшей части поля зрения, а потеря чувствительности может затрагивать одну конечность или несколько пальцев. Устойчивость взаимосвязи между данным дефектом и локализацией данного поражения позволила постепенно составить карты наиболее очевидных из этих специализированных областей: зрительной, слуховой, соматосенсорной (ощущение тела), речевой и моторной.
Во многих случаях более внимательное рассмотрение под микроскопом коры, окрашенной для выявления клеточных тел, показывает, что, несмотря на относительную однородность, там имеются структурные вариации, особенно в характере слоистости, которые хорошо коррелируют с клинически выявленными подразделениями. Дополнительное подтверждение этого последовало из наблюдений, касающихся локализации (на поверхности мозга) электрических волн, возникающих при стимуляции животного прикосновением к телу, звуковыми щелчками или тональными сигналами, действующими на ухо, или вспышками света перед глазом. Сходным образом, моторные области могут быть картированы с помощью электрической стимуляции коры и регистрации того, какая часть тела животного при этом приходит в движение. Такое систематическое картирование коры вскоре привело к фундаментальному заключению: большинство сенсорных и моторных областей содержит закономерно построенные систематические двумерные карты того внешнего мира, который они представляют. Разрушение некоторого небольшого участка коры может привести к параличу одной руки; сходное повреждение в другой ограниченной области приводит к «онемению» одной кисти или верхней губы, или к слепоте, проявляющейся в небольшой части поля зрения; если на кору мозга животного поместить электроды, можно установить, что прикосновение к одной конечности вызывает серию соответственным образом локализованных электрических потенциалов. Несомненно, что тело представлено систематическим образом в соматосенсорной и моторной зонах, а зрительный мир представлен в виде карты в первичной зрительной коре-области затылочной доли, которая у человека и обезьян-макаков (того животного, на котором в основном проводились наши исследования) имеет площадь около 15 квадратных сантиметров.
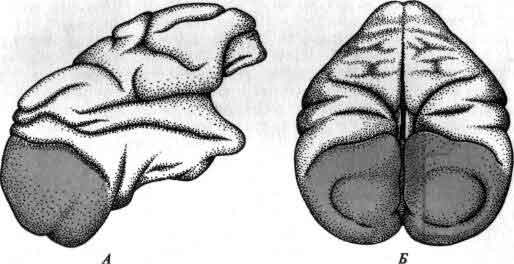
Первичная зрительная кора, известная также под названием стриaрной коры, или поля 17, - участок коры большого мозга, слоистой нейронной ткани, покрывающей мозг приматов. В мозгу макака, для которого здесь показаны вид сбоку (А) и сверху и сзади (Б), первичная зрительная кора (окрашенная область) занимает большую часть видимой поверхности обеих затылочных долей. Она также загибается на медиальную поверхность мозга. Кроме того, она продолжается в виде сложных складок и под выпуклой наружной поверхностью, как это показано на парасаггитальном срезе (см. следующий рисунок), который был сделан вдоль цветной линии и рассматривается со стороны, указанной стрелками.
В первичной зрительной коре карта не осложнена разрывами и смещениями; наблюдается лишь примечательное расщепление мира точно по его середине, причем левая его половина проецируется на правую часть мозговой коры, а правая половина — на левую. Карта тела более сложна и, по всей вероятности, пока еще не до конца выяснена. Тем не менее в ней есть своя система и она сходным образом перекрещена: правая часть тела проецируется в левое полушарие, а левая часть — в правое. (Следует заметить, что ни у кого нет даже самой отделенной идеи относительно того, какой смысл может заключаться в этой удивительной тенденции к перекрещиванию нервных путей.)
Важной особенностью кортикальных карт является наличие в них искажений. Масштабы этих карт варьируют наподобие проекций Меркатора, причем правило для коры такое: области наибольшего разрешения или тонкости функций занимают относительно большие области коры. Если взять поверхность тела, то миллиметр поверхности пальцев, губ или языка проецируется на большую часть коры, чем миллиметр туловища, ягодиц или спины; в зрении центральная часть сетчатки представлена примерно в 35 раз более детально, чем периферические области.
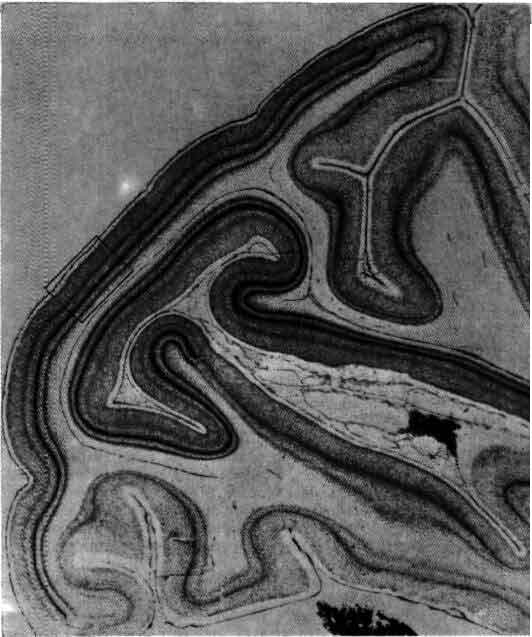
Срез зрительной коры, прошедший вдоль цветной линии, показанной на предыдущем рисунке, окрашен по методу Ниссля, выявляющему тела клеток, но не волокна. Видно, что зрительная кора представляет собой непрерывный слой нейронов толщиной около 2 мм. В рамке участок среза, показанный при большем увеличении на следующем рисунке.
Существенно, что достижения в составлении карт кортикальных проекций иногда уводили в сторону от главной проблемы: как мозг анализирует информацию. Получалось так, будто установление представительства само по себе может быть конечным итогом, а не служит более скромной цели, — как если бы задачей коры было снабжение информацией маленького гномика, который сидит внутри головы и рассматривает образы, возникающие на коре. В ходе этой статьи мы покажем, что по крайней мере в зрении мир представлен в гораздо более искаженном виде; любой маленький гномик, пытающийся собрать по частям информацию с кортикальных проекций, пришел бы в крайнее замешательство.
Поперечное сечение первичной зрительной коры макака (окраска по методу Ниссля) демонстрирует слоистую структуру и представляет общепринятые обозначения шести слоев. Белые пустоты - срезы кровеносных сосудов.
Тем не менее первым важным прозрением относительно организации коры было именно осознание этого разделения на зоны, имеющие сильно различающиеся функции и тенденцию к упорядоченной планировке. Сколько же точно имеется таких зон — этот вопрос был предметом широких спекуляций. Оценки анатомов в целом весьма высоки и доходят до нескольких сотен зон; конкретные цифры зависят от индивидуальной чувствительности исследователя к тонким различиям микроскопических структур, а иногда также и от его способности обманываться. Физиологи начали с более низких оценок, но позже, в результате использования более эффективных методов картирования, пересмотрели эти оценки в сторону их повышения. Важная основополагающая идея состоит в том, что информация любой данной модальности, например зрительная или слуховая, поступает сначала в первичную зону коры, а оттуда, либо прямым путем, либо через таламус, последовательно передается в ряд высших центров. Современные догадки относительно числа зон коры колеблются между 50 и 100.
Второе важное прозрение относительно организации коры связано с работами анатома С. Рамон-и-Кахала и его ученика Р. Лоренте де Но. Это было осознание того, что операции, которые кора осуществляет над получаемой информацией, локальны. Что это означает, легче всего понять, рассматривая схему связей, установленную с помощью метода Гольджи, которой пользовались Рамон-и-Кахал и Лоренте де Но. В сущности, связи эти просты. Пучки волокон несут информацию в кору; на пути они прерываются несколькими синаптическими переключениями и вводят информацию во все клеточные слои, пересекаемые по вертикали; наконец, по нескольким другим пучкам волокон модифицированные сообщения выходят из данной зоны коры. Детали связей между входами и выходами различаются от зоны к зоне, но внутри данного поля они кажутся довольно стереотипными. Общим для всех зон является локальный характер связен. Информация, поступающая в кору по одному волокну, может в принципе пройти сквозь всю толщину коры, примерно через три или четыре синапса, тогда как расползание в стороны, производимое ветвлениями аксонов и дендритов, практически во всех случаях ограничено несколькими миллиметрами - малой частью обширной площади коры.
Это имеет далеко идущие следствия. Что бы данная область коры ни делала, она делает это локально. На стадиях, где имеет место детальное систематическое топографическое картирование любого типа, анализ должен быть «дробным». Так, например, в соматосенсорной коре сигналы от одного пальца могут комбинироваться и сопоставляться со входами от этого же или от соседнего пальца, но они едва ли будут комбинироваться с влияниями от туловища или от ноги. То же самое применимо и к зрительному миру. Если взять детальную схему входов в первичную зрительную кору, то покажется невероятным, чтобы эта область могла каким-нибудь способом сопоставить информацию, поступающую из точек, расположенных намного выше и намного ниже горизонта или из правой и левой половин рассматриваемой картины. Какие бы операции эти кортикальные зоны ни выполняли, это должен быть какого-то рода локальный анализ сенсорного мира. Можно лишь предположить, что по мере того, как зрительная, тактильная или слуховая информация передается от одной зоны коры к следующей, карты становятся все более и более расплывчатыми, а передаваемые сообщения - все более и более абстрактными.
Хотя исследования с применением метода Гольджи еще в начале девятисотых годов прояснили, что кора должна выполнять локальный анализ, прошло полстолетия, прежде чем у физиологов появились хоть какие-то идеи относительно того, какого же именно типа должен быть этот анализ в разных зонах коры. Первые открытия были сделаны на первичной зрительной зоне, являющейся сейчас наиболее изученной и пока еще единственной, в которой анализ и последовательные преобразования информации известны сколько-нибудь детально. После описания основных преобразований, которые имеют место в первичной зрительной коре, мы покажем, как успехи в понимании этих кортикальных функций открыли целый мир схем организации, который иначе оказался бы недоступным наблюдению.
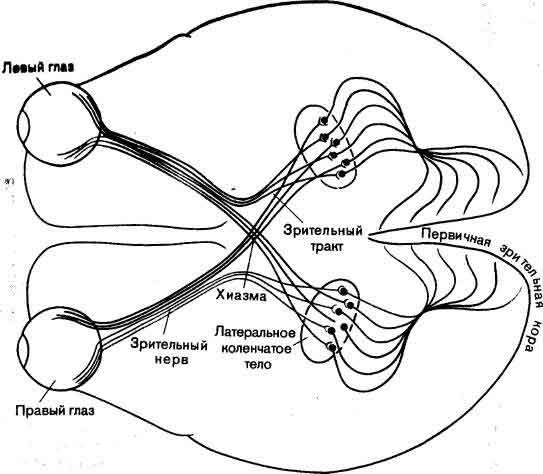
Схема зрительных путей мозга человека (вид снизу). По аксонам ганглиозных клеток, объединенным в зрительный нерв, выходные сигналы сетчатки передаются в латеральные коленчатые тела; примерно половина аксонов перекрещивается и идет на противоположную сторону мозга, так что представление каждой половины зрительной сцены проецируется на коленчатое тело противоположного полушария мозга. Нейроны коленчатых тел посылают свои аксоны в первичную зрительную кору.
Начать нам лучше всего с прослеживания зрительного пути у приматов от сетчатки до коры. Выходные сигналы каждого глаза идут в мозг примерно по миллиону нервных волокон, объединенных в зрительный нерв. Эти волокна — аксоны ганглиозных клеток сетчатки. Прежде чем сигналы светочувствительных элементов — палочек и колбочек — дошли до ганглиозных клеток, они уже преодолели от двух до четырех синапсов и вовлекли в работу четыре других типа клеток, так что довольно сложный анализ информации уже имел место.
Латеральное коленчатое тело обезьяны (А) - слоистая структура, в которой клетки слоев 1, 4 и 6 (если считать снизу вверх) получают на вход сигналы от контралатерального глаза, а слоев 2, 3 и 5 - от ипсилатерального. Карты расположены упорядоченно, так что нейроны, лежащие вдоль любого радиуса (черная линия), получают сигналы от одной и той же части зрительной картины. Послойное распределение входящих аксонов демонстрируется на двух латеральных коленчатых телах животного, у которого был зрячим только левый глаз (Б, В): в каждом коленчатом теле клетки трех слоев, имеющих входы от правого глаза, атрофированы.
Большая часть волокон зрительного нерва идет, не прерываясь, к двум клеточным ядрам, расположенным в глубине мозга; ядра эти называются латеральными коленчатыми телами; здесь волокна образуют синапсы. В свою очередь клетки латеральных коленчатых тел направляют свои аксоны прямо в первичную зрительную кору. Оттуда после нескольких синаптических переключений сообщения посылаются в ряд более далеких пунктов назначения: в соседние области коры, а также в несколько отделов в глубине мозга. Одна группа волокон даже направляется обратно в латеральные коленчатые тела; функция этой цепи обратной связи неизвестна. На данный момент самое главное то, что первичная зрительная кора ни в коей мере не является концом зрительного пути. Это только первая ступень, возможно, весьма ранняя в смысле степени абстрагирования информации, которую она обрабатывает.
Вследствие частичного перекреста зрительных нервов в хиазме коленчатое тело и кора левой стороны соединены с левыми половинами обеих сетчаток и, соответственно, имеют дело с правой половиной поля зрения, а для коленчатого тела и коры правой стороны справедливо обратное. Каждое коленчатое тело и кора получают сигналы от двух глаз и имеют дело с противоположной половиной зрительного мира.
Наша стратегия исследования работы этого зрительного пути с конца 50-х годов была в принципе простой. Начав, скажем, с волокон зрительного нерва, мы регистрировали микроэлектродами активность отдельных волокон и пытались найти способ наиболее эффективного влияния на их разряд путем стимуляции сетчатки светом. Для этой цели можно использовать световые стимулы любого мыслимого размера, формы и цвета, яркие на темном фоне и, наоборот, неподвижные и движущиеся. Хотя это порой отнимало много времени, но раньше или позже мы добивались того, что находили наилучший стимул для исследуемой клетки, в данном случае - ганглиозной клетки сетчатки. (Иногда мы ошибались!) Мы регистрировали результаты и переходили к другому волокну. После обследования нескольких сотен клеток могло обнаружиться, что новые типы попадаются редко. Удовлетворившись тем, что мы примерно знаем, как работают нейроны этой ступени, мы переходили к следующей ступени (в данном случае - коленчатому телу) и повторяли процедуру. Сравнение двух наборов результатов могло сказать нам кое-что о характере преобразований в коленчатом теле. Тогда мы шли далее к следующей ступени - первичной коре - и снова повторяли процедуру.
Действуя таким образом, удалось обнаружить, что ганглиозные клетки сетчатки и клетки коленчатого тела лучше всего реагируют на приблизительно круглые пятна некоторого конкретного размера в определенной части поля зрения. Размер является критическим параметром, поскольку рецептивное поле каждой клетки (совокупность рецепторов сетчатки, посылающих сигналы к данной клетке) целится на две части и имеет либо возбуждающий центр и тормозную периферию (клетки с «on-центром»), либо прямо противоположную организацию (клетки с «off-центром»). Такая концентрическая организация впервые была описана С. Куффлером (S. Kuffler) из Медицинской школы Университета Джонса Гопкинса в 1953 г. Пятно, в точности совпадающее с центром рецептивного поля, служит при этом более эффективным стимулом, чем пятно большего размера, заходящее на тормозные области, или чем диффузный свет. Линия (полоска света) эффективна, если она закрывает большую часть центральной области и только малую часть периферии. Поскольку поля этих клеток имеют радиальную симметрию, реакция на такие линии не зависит от их ориентации. Суммируя, можно сказать, что ганглиозные клетки сетчатки и клетки коленчатого тела, т. е. клетки, снабжающие входными сигналами зрительную кору, - это клетки с концентрическими рецептивными полями, центр и периферия которых антагонистичны. Они занимаются в первую очередь не оценкой уровней освещенности, а скорее сравнением уровня освещенности в некоторой небольшой области поля зрения со средней освещенностью ее непосредственного окружения.
Первое из двух основных преобразований, выполняемых зрительной корой, - это такое перераспределение входной информации, в результате которого большинство клеток начинает отвечать не на пятна, а на определенным образом ориентированные отрезки линий. В коре имеется много разнообразных типов клеток; по свойствам своих реакций некоторые из них просты, некоторые сложны, и при знакомстве с ними довольно скоро возникает впечатление, что имеется определенная иерархия, причем более простые клетки подают свои сигналы на вход более сложных. У обезьяны прежде всего выделяется группа клеток, которые ведут себя (насколько это известно) как клетки коленчатого тела: они имеют поля с радиальной симметрией. Все такие клетки располагаются в нижней части одного слоя, называемого слоем IV; он является как раз тем слоем, в который поступает львиная доля волокон коленчатого тела. Создается впечатление, что именно эти наименее сложно устроенные кортикальные клетки являются клетками, ближе всего стоящими ко входу.
Сравнение рецептивных полей различных клеток зрительного пути. Ганглиозные клетки сетчатки и нейроны латеральных коленчатых тел имеют концентрические поля либо с возбуждающим центром и тормозной периферией (А), либо наоборот. Пятно света, падающее на центр, вызывает реакцию такой клетки; реакцию вызывает и полоса света, падающая на поле в любой ориентации, если она проходит через центр. В зрительной коре существует иерархия нейронов со все усложняющимися характеристиками. Кортикальные клетки, которые получают сигналы прямо из коленчатых тел, имеют поля с радиальной симметрией. Однако кортикальные клетки, расположенные по зрительному пути дальше, реагируют только на линейные стимулы, имеющие определенную ориентацию. Простые клетки (Б) реагируют на такие линейные стимулы лишь в том случае, если они появляются в определенной части поля. Сложные клетки (В) реагируют на определенным образом ориентированные линии независимо от их расположения в поле, и особенно на линии, которые движутся в определенном направлении (стрелка).
За пределами слоя IV все клетки лучше всего реагируют на отрезки линий, ориентированные определенным образом. Типичная клетка реагирует только тогда, когда свет падает на определенную часть поля зрения, но освещение этой части диффузным светом дает малый или нулевой эффект, и небольшие пятна света тоже ненамного лучше. Самая сильная реакция получается, когда нужным образом ориентированная линия вспыхивает в этом месте или, в случае некоторых клеток, поступательно движется через этот участок. Наиболее эффективная ориентация варьирует от клетки к клетке и обычно определена довольно строго, так что отклонение на 10 или 20 градусов по или против часовой стрелки заметно уменьшает реакцию или уничтожает ее. (Остроту этой избирательности представить трудно. Если 10-20° звучит как огромный диапазон, вспомните, что угол на часах между 12 и 1 равен 30°.) Линия, ориентированная под 90° к оптимальному направлению, почти никогда не вызывает реакции.
Позиции рецептивных полей кортикальных нейронов (1-9), картированных при погружении электрода примерно под прямым углом к поверхности, очень близки (А), хотя поля имеют различные размеры и некоторый разброс. При наклонном погружении (5) было зарегистрировано по 2-4 клетки в каждой из четырех точек (1-4) на расстоянии 0,1 мм. В каждой группе попадаются разные размеры и имеется некоторый разброс, но теперь заметен также систематический сдвиг: поля последовательных групп клеток несколько смещены относительно друг друга.
В зависимости от конкретной клетки лучшим стимулом может быть яркая линия на темном фоне или наоборот: это может быть и граница между светлой и темной областями. Если это линия, ее толщина обычно существенна; увеличение ее свыше некоторого оптимального значения уменьшает реакцию точно так же. как увеличение диаметра пятна в случае ганглиозных клеток или клеток коленчатого тела. При этом для данной части поля зрения диаметры центральных зон рецептивных полей клеток коленчатого тела и оптимальные для коры толщины линий сравнимы.
Чувствительные к ориентации нейроны варьируют по своей сложности. Самые простые, которые мы назовем «простыми» клетками, ведут себя так, как будто к ним на вход поступают сигналы непосредственно от нескольких клеток с концентрическими полями, имеющими радиальную симметрию, т. е. от клеток того типа, которые обнаружены в слое IV. Свойства этих простых клеток, реагирующих на оптимально ориентированные линии, предъявляемые в строго определенном месте, легче всего объяснить, приняв, что все центры соответствующих полей входных клеток являются либо возбуждающими, либо тормозными и что они располагаются вдоль прямой линии. В настоящее время у нас нет непосредственных подтверждений этой схемы, но она привлекает своей простотой и тем, что согласуется с рядом косвенных свидетельств. Согласно работе Дж. Ланд (J. Lund) из Медицинской школы Вашингтонского университета, которая за последние несколько лет сделала больше, чем кто-либо другой для изучения этой области коры методом Гольджи, клетки слоя IV проецируются в слои, находящиеся прямо над ними, т. е. примерно туда, где обнаружены простые клетки.
Вторая группа чувствительных к ориентации нейронов - это гораздо более многочисленные «сложные» клетки. Они подразделяются на несколько категорий, но основная их особенность - это то, что местоположение линий для них не так существенно. Сложные клетки ведут себя так, как будто они получают сигналы от некоторого числа простых клеток, имеющих рецептивные поля с одинаковой оптимальной ориентацией, но несколько различающейся локализацией. Эта схема хорошо объясняет сильную стабильную импульсацию, возбуждаемую в сложных клетках, когда линию держат в оптимальной ориентации и проводят через рецептивное поле. При оптимальной ориентации линии многие клетки предпочитают одно из направлений движения другому, противоположному. Для объяснения такого поведения было предложено несколько возможных схем, но точный механизм пока неизвестен.
Хотя нет прямых доказательств того, что чувствительные к ориентации клетки имеют какое-либо отношение к зрительному восприятию, весьма соблазнительно думать, что они представляют некую раннюю ступень анализа зрительных образов. Имеет смысл спросить, от каких клеток этой ранней ступени можно ожидать реакции на какой-нибудь очень простой зрительный стимул, скажем темное пятно на светлом фоне. Любая клетка, рецептивное поле которой окажется полностью внутри или вне границ такого стимула, совершенно не будет затронута его присутствием, поскольку клетки коры эффективно игнорируют изменения диффузной засветки всего рецептивного поля.
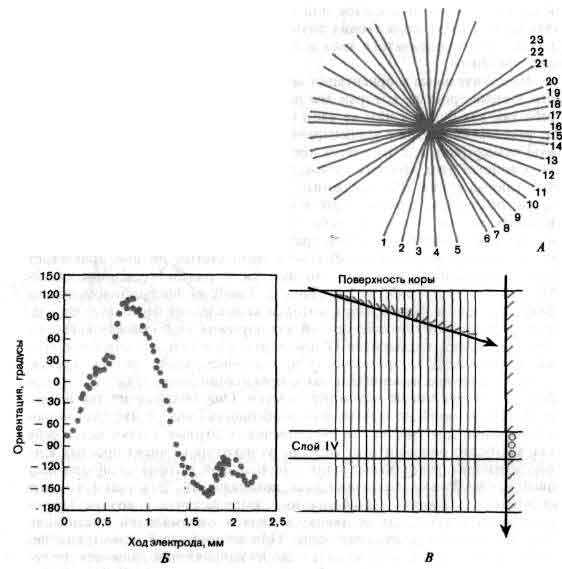
Показаны предпочтительные ориентации 23 нейронов, встретившихся при наклонном погружении микроэлектрода в кору (А); самый эффективный наклон стимула неуклонно сдвигался в направлении против часовой стрелки. Б. График сходного эксперимента, однако в этом случае несколько раз наблюдалась инверсия направления сдвига; результаты многих таких экспериментов в сочетании с тем фактом, что микроэлектрод, проникающий в кору под прямым углом, регистрирует только клетки, предпочитающие одну и ту же ориентацию (кроме клеток слоя IV, имеющих поля с радиальной симметрией и не имеющих предпочтительных направлений), заставляют предполагать, что кора подразделена на приблизительно параллельные подслои ткани, причем каждый подслой, называемый ориентационной колонкой, содержит нейроны со сходной ориентационной избирательностью (В).
Единственными клетками, которые должны прореагировать, будут те, через поля которых пройдет граница. Из клеток с радиальной симметрией сильнее всего возбудятся те, у которых граница коснется центра (так как тогда возбуждающая и тормозная зоны будут максимально неравно освещены). Из клеток, чувствительных к ориентации, должны активироваться только те, чьим оптимальным ориентациям случилось совпасть с превалирующим направлением границы. И среди таких клеток простые клетки более требовательны к стимулу, чем сложные, так как они реагируют оптимальным образом только тогда, когда граница пройдет вдоль линии, разделяющей возбуждающую и тормозную зоны. Важно представлять себе, что эта область коры производит свои операции только локально, на маленьких участках изображений; как анализируется или обрабатывается мозгом все изображение, т. е. как эта информация комбинируется и обобщается на более поздних ступенях, если действительно это имеет место, пока неизвестно.
Вторая важная функция зрительной коры обезьяны - комбинировать входы от двух глаз. В латеральном коленчатом теле любой нейрон может реагировать либо на стимуляцию левого глаза, либо на стимуляцию правого, но ни одна клетка не реагирует на стимуляцию обоих глаз. Это может показаться удивительным, поскольку в каждое коленчатое тело поступают сигналы от обоих глаз; однако коленчатые тела устроены таким образом, что пути от двух глаз проходят через них, не смешиваясь. Каждое коленчатое тело подразделяется на шесть слоев, причем три слоя, связанные с левым глазом, переплетаются, как пальцы, с тремя слоями, предназначенными для правого глаза. В каждом слое представлена карта контралатеральной половины зрительного мира (причем все шесть карт располагаются строго упорядоченно, так что на радиальном пути, пересекающем шесть слоев, рецептивные поля всех клеток, которые при этом встретятся, будут иметь практические идентичные позиции в поле зрения). Поскольку каждый данный слой получает входные сигналы только от одного глаза, отдельные клетки каждого слоя должны быть монокулярными.
Даже в зрительной коре нейроны, на которые клетки коленчатого тела переключаются непосредственно - клетки с радиальной симметрией из слоя IV, - все (насколько нам известно) строго монокулярны; то же самое справедливо для всех простых клеток. Только на уровне сложных клеток пути от двух глаз сходятся, но и там смешение информации неполное и принимает специфическую форму. Примерно половина всех сложных клеток монокулярна в том смысле, что любая такая клетка может быть активирована только стимуляцией какого-то одного глаза. На остальные клетки могут оказывать независимые влияния оба глаза.
Если построить карты рецептивных полей бинокулярной клетки для правого и левого глаза (посылая стимулы сначала в один глаз, а затем в другой) и сравнить эти два поля, то окажется, что они имеют одинаковые позиции, уровни сложности, предпочтительные ориентации и направления - т. е. все, что можно узнать о клетке, стимулируя один глаз, подтверждается при стимуляции второго. Есть только одно исключение: если сначала один, а затем второй глаз тестировать идентичными стимулами, две реакции не обязательно будут одинаковыми количественно; во многих случаях один глаз доминирует, устойчиво вызывая более частые разряды, чем второй.
Перебирая клетки, можно найти все степени доминирования глаз, от полной монополии одного глаза через равенство до исключительного влияния второго глаза. У обезьяны клетки с заметным предпочтением одного глаза встречаются несколько чаще, чем клетки, в реакции которых оба глаза вносят примерно равные вклады. По-видимому, связи типичной бинокулярной клетки первичной зрительной коры с двумя глазами практически одинаковы по организации, но могут различаться по числу соединительных волокон.
Весьма замечательно, что сложные схемы связей, обеспечивающих избирательность по отношению к ориентации и направлению движения, а также другие особые свойства, должны быть представлены в виде двух копий. И, пожалуй, еще более поразительно, что все это можно наблюдать у новорожденных животных. Данные схемы связей в основном врожденные и, по-видимому, детерминированы генетически. (Однако в одном специальном отношении некоторое дозревание бинокулярных связей осуществляется главным образом после рождения.)
Теперь мы обратимся к рассмотрению закономерностей группировки клеток в коре. Каким образом распределены там клетки со сходными характеристиками - близкие по степени сложности, локализации рецептивного поля и доминированию глаз - сгруппированы вместе или разбросаны в случайном порядке? Из приводимого описания будет очевидно, что сходные по сложности клетки имеют тенденцию группироваться в слои, причем клетки с радиальной симметрией лежат внизу слоя IV, простые клетки - прямо над ними, а сложные клетки - в слоях II, III, V и VI. Сложные клетки можно подразделить еще на несколько категорий, и те, которые находятся в одном слое, в ряде отношений сильно отличаются от клеток других слоев.
Эти различия между слоями приобретают особый интерес в аспекте следующего важного открытия, подтвержденного рядом физиологов и анатомов в последние несколько десятилетий: волокна, выходящие из разных слоев коры, имеют различные места назначения. Так, самый глубокий слой зрительной коры - слой VI - проецируется главным образом (а возможно, и исключительно) обратно в латеральное коленчатое тело; слой V проецируется в верхнее двухолмие, зрительный отдел среднего мозга; слои II и III направляют свои волокна в другие части коры. Такая определенность в локализации проекций каждого слоя, возможно, заслуживает того, чтобы рассматривать ее как третье важное прозрение относительно организации коры.

Ориентационные колонки удалось выявить в виде уловимых анатомических структур с помощью дезоксиглюкозных радиоавтографов, полученных авторами и М. Страйкером. Обезьяне вводили дезоксиглюкозу с радиоактивной меткой; она поглощалась преимущественно работающими клетками, и в них накапливались продукты первых этапов метаболизма. Сразу после инъекции животному предъявляли решетку из вертикальных полос, в связи с чем клетки, реагирующие предпочтительно на вертикальные линии, были самыми активными, а затем стали самыми радиоактивными. На этом срезе перпендикулярные к поверхности области активных клеток представляют собой узкие полосы, расположенные на расстоянии около 0,5 мм. Слой IV (не проявляющий избирательности в отношении ориентации), как и ожидалось, однороден по радиоактивности.
Следующий варьирующий параметр стимуляции, который нужно рассмотреть, - это положение рецептивного поля в поле зрения. Описывая латеральное коленчатое тело, мы указывали, что в каждом его слое образуется упорядоченная топографическая карта контралатеральной половины поля зрения. В проекции латерального коленчатого тела на зрительную кору этот порядок сохраняется, в результате чего создается кортикальная карта поля зрения. При наличии такой упорядоченной карты не удивительно, что соседние клетки в этой части коры всегда имеют рецептивные поля, располагающиеся в непосредственной близости; в действительности, они обычно перекрываются. Если в кору под прямым углом к поверхности погружать микроэлектрод и регистрировать активность клетки за клеткой, сделав до 100 или 200 отведений во все более глубоких слоях, то опять же рецептивные поля будут большей частью перекрываться, причем каждое новое поле будет накладываться на все предыдущие. Область, занимаемая всей «пачкой» полей, обычно в несколько раз превышает размер одного типичного поля.
Размеры этих рецептивных полей несколько варьируют. Некоторые вариации соотносятся со слоистостью: наибольшие поля при любом погружении имеют тенденцию обнаруживаться в слоях III, V и VI. Однако наиболее важные вариации коррелируют с эксцентриситетом, или расстоянием рецептивного поля клетки от центра взора. Размеры полей и величина соответствующего разброса в тех областях коры, где картирована область центра взора, выглядят крошечными по сравнению с размерами и разбросом в тех частях, где картирована далекая периферия. Пачку накладывающихся полей, которые картируются при погружении, начинающемся в любой точке коры, мы называем «агрегатным» полем этой точки. Очевидно, что величина агрегатного поля является функцией расстояния от центра взора.

Картина распределения предпочтительных ориентации, если ее рассматривать в фас, неожиданно оказывается весьма сложной. Этот радиоавтограф получен с помощью дезоксиглюкозы со среза, тангенциального к несколько изогнутым слоям коры. Более темные области соответствуют однородно радиоактивному слою IV. В других слоях ориентационные области выглядят как сложным образом изогнутые полосы, напоминающие стены рассматриваемого сверху лабиринта, однако расстояния между полосами примерно одинаковы.
Если электрод входит наклонно, почти параллельно поверхности, снова выявляется разброс в позиции полей от точки к точке, но теперь этот разброс накладывается на систематическое смещение позиции поля, направление которого диктуется топографической картой поля зрения. При этом обнаруживается интересная закономерность: оказывается, что продвижение электрода примерно на 1-2 миллиметра всегда приводит к такому смещению в поле зрения, которого достаточно для перехода в совершенно новую область. Короче, величина продвижения в поле зрения примерно соответствует при этом размеру агрегатного рецептивного поля. Для первичной зрительной коры это справедливо, где бы ни производилось погружение. В центре взора поля и их разброс малы, но таковы же и смещения, соответствующие продвижению на миллиметр вдоль коры. С увеличением эксцентриситета (по мере приближения к границам поля зрения) как поля и их разброс, так и смещения, становятся пропорционально больше. Получается, что повсюду кусочек коры протяженностью в один или два миллиметра является тем блоком, который обслуживает область поля зрения, эквивалентную величине агрегатного поля.
Бинокулярные клетки могут возбуждаться как каждым глазом независимо, так и более сильно - двумя глазами вместе. Здесь представлены относящиеся к левому и правому глазам рецептивные поля сложной клетки, контролирующей область в верхнем левом квадранте поля зрения. (Оси координат соответствуют горизонтальному и вертикальному меридианам поля, пересекающимся в точке фиксации.) Два данных рецептивных поля идентичны, но сила реакции может зависеть от того, какой глаз стимулируется: правый или левый. Глаз, оказывающий более сильное влияние, называется доминантным.
Из этого наблюдения следует предположение о способе, с помощью которого кора решает такую фундаментальную проблему: как сделать, чтобы зрительная картина анализировалась детально в центральной части и намного грубее на периферии. В сетчатке, перед которой стоит такая же проблема, по очевидным оптическим причинам число миллиметров, соответствующих градусу поля зрения, постоянно. Сетчатка обрабатывает центральные участки более детально благодаря тому, что имеет огромное количество ганглиозных клеток, каждая из которых обслуживает крошечную область в центре поля зрения; слой ганглиозных клеток в центральной части сетчатки относительно толст, тогда как в периферических частях сетчатки он очень тонок. В то же время, было, по-видимому, желательно, чтобы кора везде имела одинаковую толщину. Здесь нет никаких оптических ограничений типа накладываемых на сетчатку, и потому площади просто распределяются в соответствии с проблемами, которые должны решаться.
На каждом квадратном миллиметре коры предположительно действуют примерно такие же механизмы, как на любом другом. Несколько тысяч волокон из коленчатого тела входит в такую функциональную ячейку коры, и что-то около 50000 волокон выходит из нее, независимо от того, представлена ли здесь малая часть зрительного мира очень детально или большая по размеру часть соответственно менее детально. Как мы указывали вначале, предположение об однородности коры возникает при взгляде на окрашенные срезы. Оно убедительно подтверждается, когда мы исследуем архитектонику дальше, специально обращая внимание на чувствительность к ориентации или на доминирование глаз.
В физиологических исследованиях была выявлена группировка клеток в соответствии с доминированием глаз. При некотором произвольном погружении в кору под прямым углом к поверхности (1) микроэлектрод может встретить только клетки, которые оказывают предпочтение левому глазу (Lr), а в слое IV - клетки, которые возбуждаются только левым глазом (L); при другом вертикальном погружении (2) для всех клеток доминантным будет правый глаз (R,), а в слое IV клетки будут возбуждаться исключительно правым глазом (R). При наклонном погружении будет наблюдаться регулярное чередование доминирования глаз. Результаты многократных погружений наводят на мысль, что кора подразделена на области с поперечным сечением шириной около 0,4 мм и со стенками, перпендикулярными поверхности и слоям коры - колонки глазодоминантности.
Относительно ориентации мы исследовали группировки клеток точно так же, как это мы делали при рассмотрении локализации полей, т. е. сначала изучали пары клеток, расположенных в непосредственной близости друг от друга. Две такие клетки почти всегда имеют одинаковые оптимальные ориентации стимулов. Если электрод вводится в направлении, перпендикулярном к поверхности, все клетки вдоль пути его проникновения имеют одинаковые или почти одинаковые предпочтительные ориентации (кроме клеток в глубине слоя IV, которые совсем не имеют оптимальных ориентации). Однако при двух погружениях перпендикулярно поверхности, произведенных на расстоянии в один миллиметр или около того, обнаруженные две ориентации, как правило, различны. Следовательно, кора должна подразделяться на какие-то вертикальные ячейки, внутри которых оптимальная ориентация одинакова для рецептивных полей всех клеток. Когда мы натолкнулись на эту систему почти 20 лет назад, она заинтриговала нас, поскольку она так хорошо соответствовала иерархическим схемам, предложенным нами для объяснения того, каким образом сложные клетки снабжаются входными сигналами от простых: эти схемы включали связи с клетками, поля которых покрывают одну и ту же часть поля зрения и которые реагируют на одну и ту же ориентацию линий. Казалось совершенно разумным, что прочно взаимосвязанные клетки должны быть сгруппированы вместе.
Если кора разбита на области с постоянной для рецептивных полей оптимальной ориентацией, можно ли сказать что-нибудь большее о трехмерной форме этих областей, кроме того, что их стенки перпендикулярны поверхности? Соотносятся ли соседние области сколько-нибудь систематическим образом, или обслуживающие всевозможные направления колонки разбросаны по коре случайно? Мы начали изучать ни вопросы, просто вводя электрод в кору наклонно или параллельно поверхности. Когда мы первый раз поставили такой эксперимент в 1961 г., результат был столь удивителен, что мы с трудом в него поверили. Вместо случайного набора сменяющихся ориентации наблюдалась изумительная упорядоченность. Каждый раз, когда микроэлектрод продвигался всего на 25-50 мкм (тысячных долей миллиметра), оптимальная ориентация менялась небольшим скачком, в среднем примерно на 10°; скачки в одном направлении - по часовой стрелке или против нее - продолжали наблюдаться в довольно большом диапазоне углов, что-то от 90 до 270°.
С тех пор как было сделано это первое наблюдение, мы находим аналогичную упорядоченность почти у каждой обезьяны. Либо стабильно регистрируется поворот ориентации, либо, реже, встречаются участки, в пределах которых ориентация остается постоянной. Последовательные изменения ориентации достаточно малы, поэтому трудно быть уверенным в том, что области постоянной ориентации имеют конечные размеры; возможно, что по мере продвижения электрода вдоль коры оптимальные направления меняются в каком-то смысле непрерывно.
У нас все больше нарастал интерес к трехмерной форме этих подобластей. Уже из рассмотрения одной геометрии очевидна возможность существования малых или нулевых изменений в любом направлении при горизонтальном или тангенциальном проникновении к параллельным слоям ткани, содержащим клетки со сходной специфичностью, и таким, что каждый слой перпендикулярен поверхности. Слои не обязательно должны быть плоскими как ломти хлеба; при взгляде сверху некоторые из них имеют вид завихрений, с помощью которых легко объяснить инверсии в направлении поворота ориентации. Запись от множества клеток по ходу нескольких параллельных погружений электрода, по-видимому, подтверждает такое предположение, однако с помощью микроэлектрода мы могли исследовать не более чем крошечный участок мозга.
К счастью, как раз тогда, когда нам было нужно, был изобретен идеальный анатомический метод. Это был метод оценки активности мозга с помощью 2-дезоксиглюкозы, предложенный Л. Соколовым (L. Sokoloff) и его группой из Национального института охраны психического здоровья (метод этот описан в данном выпуске в статье Л. Ивер-сена). В основу метода положен тот факт, что клетки мозга используют в качестве источника метаболической энергии главным образом глюкозу и что близкородственное ей соединение, 2-дезоксиглюкоза, может в какой-то степени ее имитировать. Если в организм животного ввести дезоксиглюкозу, она будет так же интенсивно поглощаться нейронами, как и сама глюкоза, и чем выше активность нейрона, тем интенсивнее будет поглощение. Соединение начинает включаться в метаболический цикл, но по причинам, которые лучше известны биохимикам, процесс останавливается на метаболитах, не способных проникать через клеточные мембраны и потому накапливающихся в клетках.
Анатомическое подтверждение существования колонок глазодоминантности было получено с помощью различных методов окраски и методом аксонного транспорта с применением радиоавтографии. Данный составной радиоавтограф, проявляющий узор на площади шириной около 10 мм, был получен путем изготовления и совмещения срезов из областей, представляющих слой IV в некотором числе параллельных сечений.
Метод Соколова состоит в том, что животному вводят меченную радиоактивным изотопом (14С) дезоксиглюкозу, затем воздействуют раздражителем, предназначенным активировать определенные нейроны, и сразу же определяют радиоактивность ткани мозга; в результате обнаружатся активные области, где клетки поглотили больше дезоксиклюкозы, чем в областях, остававшихся в покое. Обычный способ исследования мозга для этой цели - изготовить тонкие срезы (как это делается для гистологического исследования) и прижать эти срезы к фотопластинке, чувствительной к радиактивному излучению. После проявления все участки, находившиеся в контакте с радиоактивным материалом, будут видны как темные массы осажденных зерен серебра. Вместе с М. Страйкером (М. Stryker) мы приспособили метод Соколова к нашим проблемам, вводя анестезированному животному дезоксиглюкозу и затем двигая изображения черно-белых вертикальных полос туда и обратно на расстоянии 1,5 м от животного в течение 45 минут. Затем мы готовили срезы мозга либо перпендикулярно поверхности коры, либо параллельно ей.
Реконструкция узора глазодоминантности по всей наружной части правой первичной зрительной коры была выполнена авторами и С. ЛеВэем по сериям срезов, окрашенных методом серебрения, разработанным ЛеВэем. Левая граница проходит по медиальному краю затылочной доли, где кора загибается внутрь.
Полученные радиоавтографы быстро подтвердили физиологические результаты. На срезах, перпендикулярных поверхности, выявились узкие полосы радиоактивности, встречающиеся примерно через каждые 570 мкм (грубо говоря, полмиллиметра) и простирающиеся по всей толще коры. Очевидно, это и были области, которые содержали клетки, реагирующие на вертикальные линии. Глубокая часть слоя IV была однородно радиоактивна, как мы и ожидали на основании того факта, что клетки в этом слое имеют рецептивные поля с радиальной симметрией и не проявляют избирательности по отношению к ориентации.
На срезах, параллельных поверхности, выявилась неожиданно сложная система чередующихся полос, часто образующих завихрения, неоднократно расходящихся и воссоединяющихся и только местами образующих регулярные параллельные переслаивания. Что тут было особенно удивительно, так это равенство расстояний между полосами по всей коре. Это прекрасно согласовалось с идеей об однородности коры. Кроме того, величина расстояний между полосами хорошо увязывалась с идеей о том, что кортикальные механизмы повторяются, по крайней мере через каждый миллиметр. Если бы расстояние, требующееся для поворота от вертикали через 180° назад к вертикали, было равно, скажем, 10 миллиметрам, значительная часть коры не содержала бы клеток, чувствительных к какой-то заданной ориентации, что вело бы к фрагментарному и чрезвычайно причудливому представлению зрительной картины.
Последняя переменная, вклад которой в архитектонику надо рассмотреть, - это предпочтение того или другого глаза. При микроэлектродных исследованиях почти неизменно оказывалось, что соседние клетки «предпочитают» один и тот же глаз. Если при вертикальном погружении первая клетка, на которую мы натыкались, предпочитала правый глаз, то также было и со всеми другими клетками вплоть до основания слоя VI; если же первая клетка предпочитала левый глаз, то же было верно и для остальных. Вероятности предпочтения того или другого глаза при произвольном отведении были равны. (Поскольку клетки слоя IV монокулярны, тут нужно говорить не о предпочтении, а о монополии глаз.) Если электрод вводили наклонно или горизонтально, области предпочтения левого и правого глаз чередовались, причем переключения происходили довольно резко - примерно через каждые полмиллиметра. Таким образом, в коре обнаружилась вторая система подобластей с вертикальными стенками, простирающимися через всю ее толщу. Система глазодоминантности, по-видимому, совершенно независима от ориентационной системы, так как при наклонном или тангенциальном введении электрода две последовательности не имеют очевидного отношения друг к другу.
Происхождение колонок глазодоминантности, как их потом назвали, представляется весьма простым. Окончания волокон коленчатого тела, часть которых обслуживает левый глаз, а часть - правый, группируются на входе в кору таким образом, что в слое IV перемешивания нет. Это приводит к появлению «лево»- и «правоглазых» участков, располагающихся с интервалами около полмиллиметра. Нейрон, находящийся ниже или выше слоя IV, образует связи в этом слое в радиусе примерно до миллиметра по всем направлениям. По-видимому, самые прочные связи образуются с областью слоя IV, наиболее близкой к данному нейрону, так что, по-видимому, на него будет оказывать доминирующее влияние тот глаз, который «обслуживает» эту область.
И снова нам было очень интересно выяснить, как эти «левоглазые» и «правоглазые» области могут выглядеть в трех измерениях. То чередование, которое обнаруживалось физиологически, можно было объяснить и геометрически. Ответ сначала последовал из работ, выполненных методом дегенерации с применением серебрения, предложенным для картирования связей У. Наута (W. Nauta) из Массачусетского технологического института. Потом мы нашли три других независимых анатомических метода для демонстрации этих колонок.
Участок коры площадью около одного квадратного миллиметра и высотой в два миллиметра (окрашенный столбик) можно рассматривать как элементарную ячейку первичной зрительной коры. Она содержит один набор ориентационных колонок, обслуживающих все направления, и один набор слоев глазодоминантности, обслуживающих оба глаза. Эта структура многократно повторяется по всей первичной зрительной зоне. Границы можно проводить произвольно (по правому или по левому глазу, по вертикальной, горизонтальной или наклонной ориентации); представление данных подслоев в виде плоских пластин, пересекающихся под прямыми углами, является крайним упрощением.
Один из особенно эффективных методов (поскольку он позволяет наблюдать на одном животном распределение колонок по всей первичной зрительной коре) основан на явлении аксонного транспорта. Процедура сводится к инъекции в нервную ткань меченой аминокислоты. Аминокислота поглощается телом клетки, по-видимому, включается в белок и затем транспортируется по аксону к его окончаниям. Когда мы инъецировали аминокислоту в один глаз обезьяны, ганглиозные клетки сетчатки поглощали ее и транспортировали по своим аксонам — волокнам зрительного нерва. После этого мы исследовали места назначения этих волокон в латеральном коленчатом теле, покрывая срезы тканей серебряной эмульсией и проявляя ее. Оказалось, что радиоактивная метка отчетливо выделяется в трех взаимно дополняющих друг друга слоях коленчатых тел.
Однако этот метод в обычном применении не дает возможности проследить путь от терминали одного аксона через синапс в следующий нейрон и к его окончаниям, а мы хотели проследить весь путь через кору. В 1971 г. Б. Графштейн (В. Grafstein) из Медицинского колледжа Корнеллского университета показала, что после инъекции достаточно большого количества радиоактивного материала в глаз мыши часть его выходит из терминалей зрительного нерва, поглощается клетками коленчатого тела и транспортируется по их аксонам в кору. Мы подумали, что сходная инъекция в комбинации с методом радиоавтографии позволит выявить в IV слое зрительной коры окончания волокон клеток коленчатого тела, принадлежащие одному глазу.
Наша первая попытка закончилась весьма плачевно: в слое IV были видны лишь слабые тени из нескольких зерен серебра. Только после нескольких недель мы осознали, что обратившись к наблюдению под микроскопом в условиях темного поля, можно извлечь выгоду из свойства зерен серебра рассеивать свет, благодаря чему чувствительность метода возрастет. Мы позаимствовали темнопольный конденсор, и когда взглянули на наш первый слайд в микроскоп, там в слое IV во всей своей красе сияли периодические структуры, выявленные метками (см. верхний рисунок на стр. 166).
Следующим нашим шагом была попытка увидеть картину, так сказать, «в лицо», делая срезы коры параллельно поверхности. Кора обезьяны куполообразна, так что на срезе, параллельном поверхности и тангенциальном слою IV, этот слой выглядит как круг или овал, а на срезе, сделанном ниже слоя IV, его сечение представлено в виде кольца. Монтируя вместе серии таких овалов и колец из набора срезов, можно реконструировать картину на большой площади коры.
Из такой реконструкции сразу стало очевидно, что общий план организации - это чередование параллельных полос, которые представляют окончания, принадлежащие глазу, подвергшемуся инъекции, и промежутков, которые представляют другой глаз. Полосы не так регулярны, как на обоях. (Время от времени мы напоминали себе, что это все-таки биология!) Тут и там полоса, представляющая один глаз, разветвляется на две полосы или кончается тупиком в точке, где ветвится полоса другого глаза. Нерегулярности наиболее обычны вблизи центра взора и вдоль линии, представляющей горизонт. Полосы по всей видимости всегда перпендикулярны к границе между первичной зрительной корой и ее соседом - полем 18, и здесь регулярность наибольшая. Такое общее правило, по-видимому, применимо к мозгу всех макаков, хотя узоры варьируют от одного индивидуума к другому и даже у одной и той же обезьяны от одного полушария к другому.
Гипотетическая картина корковой активности, которая может быть результатом стимуляции левого глаза одним коротким отрезком горизонтальной линии, помешенным в верхнем левом квадранте поля зрения, показана цветными штрихами на схеме участка правой коры, рассматриваемой в фас. Область, в которую поступают входные сигналы от объекта в поле зрения, обведена пунктирной черной линией. Если колонки глазодоминантности и ориентационные колонки расположены так, как это здесь изображено, из всех клеток активируются те, которые оптимально реагируют на приблизительно горизонтальные стимулы, предъявляемые левому глазу.
Ширина комплекта из двух полос постоянна, около 0,8 мм, по всей первичной зрительной коре, что еще раз подчеркивает однородность коры. Опять же эта ширина прекрасно согласуется с той идеей, что в пределах одного квадратного миллиметра коры должны содержаться все механизмы, необходимые для «присмотра» за областью, размером с агрегатное поле. Два описанных выше метода — применение меченой дезоксиглюкозы и транспорт аминокислоты, имеют огромное достоинство в том отношении, что они взаимно совместимы, в связи с чем мы можем применять их одновременно, один — для картирования ориентационных полос, а другой — для выявления колонок глазодоминантности. Число препаратов мозга, исследованных к настоящему времени, слишком мало, для того чтобы можно было сделать те или иные окончательные выводы, но пока эти две системы полос кажутся совершенно независимыми; они и не параллельны, и не перпендикулярны, а пересекаются случайным образом.
Функция, выполняемая колонками глазодоминантности, пока остается тайной. Мы знаем, что нейроны со всеми градациями предпочтения глаза имеются по всей бинокулярной части поля зрения, и возможно, что некоторая регулярная упорядоченная система конвергенции входов гарантирует однородность распределения, благодаря чему ни один глаз не окажется случайно выделенным ни в одном месте. Зачем нужны повсеместно все эти градации предпочтения глаза, само по себе неясно; мы можем только догадываться, что это имеет какое-то отношение к восприятию глубины.
Если собрать вместе все, что стало известно о первичной зрительной коре, будет ясно, что элементарным участком коры нужно считать блок площадью примерно в квадратный миллиметр и глубиной два миллиметра. Знать организацию такого кусочка ткани — это значит знать организацию всего поля 17; целое должно быть в значительной степени простым повторением этой элементарной единицы. Конечно, данную элементарную единицу не следует рассматривать как отдельный изолированный блок. С чего начинать отсчет ориентационных колонок — с колонки, представляющей вертикальную ориентацию, или наклонную или горизонтальную, - совершенно безразлично; точно так же все равно, с какой пары начинать последовательность полос глазодоминантности: левый глаз - правый глаз или правый глаз - левый глаз. Это же справедливо для любой ячейки кристалла хлористого натрия и для любого сложного повторяющегося узора типа тех, какие печатают на обоях.
На что же тогда становится похожей зрительная картина, когда она проецируется на зрительную кору? Предположим, что животное фиксирует свой взгляд на некоторой точке и что единственный объект в поле зрения - прямая линия выше и чуть левее той точки, к которой прикован взгляд. Если бы каждая активная клетка должна была загораться и если бы мы могли стоять над корой и смотреть на нее сверху, какую картину мы бы увидели? Чтобы сделать задачу более интересной, предположим, что картина рассматривается только одним глазом. Ввиду только что описанной архитектоники объект предстанет не в виде линии, а всего лишь в виде набора регулярно расположенных обрывков. Это рассуждение можно проверить непосредственно, если предъявлять обезьяне, у которой один глаз закрыт, набор вертикальных полос, а затем, применив дезоксиглюкозу, изготовить радиоавтограф. Полученная картина не будет большим сюрпризом: это будет набор регулярно расположенных пятен, который будет отражать пересечение двух систем колонок. Вообразите удивление и замешательство маленького гномика, если бы он увидел такую версию внешнего мира!
Реальный образец активности коры, которая была вызвана предъявлением одному левому глазу решетки из вертикальных полос. Полученный с помощью дезоксиглюкозы радиоавтограф тангенциального среза верхних слоев коры. Картина регулярно расположенных темных участков радиоактивности отражает пересечение систем колонок глазодоминантности и предпочтительной ориентации.
Почему эволюция пришла к тому, чтобы взять на себя труд изобрести столь сложную схему, - это вопрос, который продолжает волновать нас. Возможно, самое подходящее объяснение состоит в том, что системы колонок — это решение проблемы отображения более двух измерений на двумерной поверхности. Кора имеет дело по крайней мере с четырьмя наборами значений: двумя для х и у-координат в поле зрения, одним - для ориентации и одним - для различных степеней предпочтения глаза. Две координаты точек на поверхности коры используются для указания позиции поля; две другие переменные удалось разместить в коре благодаря ее подразделению на столь мелкие участки, что, пробежав весь набор ориентации или степеней предпочтения глаза, мы будем иметь лишь такой сдвиг позиций в поле зрения, который будет мал по сравнению с разрешающей силой в этой части зрительного мира. Стратегия членения коры на мелкие вертикальные подразделения явно не ограничена первичной зрительной зоной. Такие подразделения впервые были обнаружены в соматосенсорной области В. Маунткаслом (V. Mountcastle) из Медицинской школы Университета Джонса Гопкинса примерно за 10 лет до нашей работы на зрительной коре. В соматосенсорной области, как мы указывали выше, в основе топографии лежит картирование противоположной половины тела, но сверх этого имеется еще двойная система подразделений, так что есть области, где нейроны реагируют на движение суставов или давление на кожу, и другие области, где нейроны реагируют на прикосновение или отклонение волосков. Как и в случае зрительных колонок, полный комплект (набор нейронов всех типов) занимает здесь место протяженностью около миллиметра. Эти подразделения аналогичны колонкам глазодоминантности в том отношении, что они определяются в первую очередь распределением волокон при входе в кору (там — от левого и правого глаза, здесь — от глубоко расположенных рецепторов и от рецепторов верхних слоев кожи), а не связями внутри коры, типа тех, которые определяют предпочтение ориентации и связанную с этим систему ориентационных колонок. Смысл колонок, обнаруженных в первичной зрительной и соматосенсорной коре, истолкован лучше всего, однако имеются указания на наличие сходных вертикальных подразделений и в некоторых других зонах: ряде высших зрительных областей, сенсорных теменных областях, недавно изученных Маунткаслом, и в слуховой зоне, где Т. Имиг (Т. Imig), X. Эдриен (Н. Adrian) и Дж. Брадж (J. Brugge) из Медицинской школы Висконсинского университета нашли подразделения, в которых два уха представляются попеременно то суммирующими идущую от них информацию, то конкурирующими.
Для большинства из этих физиологически установленных систем (кроме зрительных) до настоящего времени неизвестно анатомических коррелятов. С другой стороны, в последние годы несколько анатомов, в особенности Э. Джонс (Е. Jones) из Медицинской школы Вашингтонского университета, а также Наута (Nauta) и П. Голдмен (P. Goldmen) из Массачусетского технологического института показали, что пути из одной зоны коры в другую (например, из соматосенсорной зоны одной стороны в соответствующую зону на другой стороне) оканчиваются в участках, имеющих правильные чередования с периодом около миллиметра. Здесь колонки видны морфологически, но нет никакой идеи относительно их физиологической интерпретации. Ясно, однако, что тонкие периодические подразделения — действительно общая черта коры мозга. Таким образом, можно сказать, что первое наблюдение Маунткаслом такого свойства способствовало четвертому глубокому прозрению относительно организации коры.
Конечно, было бы неверно считать, что данное рассмотрение зрительной коры в какой-то степени исчерпывает предмет. Кора, по-видимому, имеет дело и с цветом, и с движением, и со стереоскопической глубиной, но в какой степени и каким образом — пока неясно. Из наших работ, относящихся к восприятию глубины, и из работ по цветовому зрению, выполненных С. Зеки (S. Zeki) из Лондонского университетского колледжа, можно заключить, что высшие кортикальные зрительные зоны, в которые первичная кора проецируется непосредственно или обходным путем, могут быть специализированы для обработки соответствующих параметров, но мы еще очень далеки от понимания того, в чем заключается эта обработка.
Что делается за пределами первичной зрительной коры и как информация об ориентации используется на последующих стадиях? Нужно ли думать, что в конечном счете обнаружится клетка, специфически реагирующая лишь на некоторый очень определенный объект? (Обычно в качестве такого объекта выбирают чью-то бабушку по причинам, которые мы уяснить не можем.) Наш ответ состоит в том, что мы сомневаемся в существовании таких клеток, но мы не можем предложить взамен ничего хорошего. К счастью, широкие спекуляции на тему о том, каким образом мозг мог бы работать, это не единственный путь, открытый исследователями. Изучать мозг — это более увлекательное и, кажется, более полезное занятие.
Было время, и не так давно, когда, глядя на миллионы нейронов в различных слоях коры, можно было сомневаться в том, что у кого-нибудь когда-нибудь может возникнуть хоть какая-нибудь идея относительно их функции. Работают ли все они параллельно, как клетки печени или почки, выполняя свои функции сообща, или каждый из них делает что-то свое, особое? Для зрительной коры ответ представляется теперь в общем плане известным: нейроны возбуждаются или тормозятся специфическими стимулами; группы нейронов действительно выполняют специальные преобразования. Если окажется возможным разгадать секреты нескольких подобных областей, будет резонно полагать, что и другие области со временем также раскроют свои тайны.
Э. ЭВАРТС
Механизмы головного мозга, управляющие движением
Как головной и спинной мозг управляют движениями тела? Мозг не только посылает команды мышцам, но и получает по обратной связи сигналы, которые помогают ему согласовывать эти команды
Одно из первых сведений, полученных более ста лет назад, об управлении движением со стороны головного мозга состояло в том, что движения тела могут быть вызваны сигналами, приходящими в спинной мозг из специальной области головного мозга - моторной зоны коры больших полушарий. Движения имеют широкий диапазон - от мышечных координации, требуемых для грубой ручной работы или быстрого перемещения всего тела, до тонких движений пальцев при хирургических операциях, выполняемых под микроскопом.

Три белые «тени» на микрофотографии представляют собой метки, специально созданные, чтобы облегчить исследование важного аспекта связи между головным мозгом и движением, а именно химизма мышечного сокращения, следующего за импульсацией мотонейрона. На микрофотографии показан поперечный срез одной из мышц конечности кошки. «Тени» образованы отдельными мышечными волокнами в одной двигательной единице. Предварительное изучение этой единицы показало, что она относится к «медленной» мышце, т. е. такого рода мышце, которая развивает небольшую силу, но функционирует не утомляясь. Эти три волокна превратились в метки в результате длительной стимуляции мотонейрона, который управляет их сокращениями, что привело к истощению запаса гликогена - особой формы хранения глюкозы, которая служит источником энергии для работы мышцы. При окрашивании среза все мышечные волокна с нормальным содержанием гликогена стали розовыми. На следующих рисунках показаны другие срезы той же мышцы с теми же тремя метками. Они были окрашены, чтобы определить связь между химическими и механическими свойствами мышечных волокон. Микрофотографии получены Р. Берком (R. Burke) и П. Церисом (P. Tsairis) в Национальных институтах здравоохранения.
Эта выходная активность моторной коры сама является результатом сигналов, поступающих из других пунктов - не только от других областей коры, например тактильной, но и от подкорковых структур мозжечка и базальных ганглиев, которые посылают сигналы в моторную кору еще через одно подкорковое образование — таламус. Основная часть современных исследований мозговых механизмов движения направлена на лучшее понимание того, как сигналы, приходящие от различных корковых и подкорковых структур, объединяются в контроле над конечными выходами из моторной коры к спинному мозгу и оттуда к мышцам. В настоящей статье будет рассмотрен современный уровень наших знаний, которые имеют важное значение по двум причинам. Во-первых, они связаны с фундаментальными проблемами общей организации головного мозга. Во-вторых, они имеют отношение к лечению и, возможно, к предупреждению таких неврологических заболеваний, как болезнь Паркинсона и хорея Гентингтона (две болезни из числа тех, при которых затронуты базальные ганглии), различные проявления инсульта, рассеянного склероза, а также многих других нарушений, возникающих при повреждении мозжечка.
Каковы элементарные условия для выполнения движения? Первое — это мышца, второе — это сигнализирующая система, которая вызывает упорядоченное сокращение мышцы. Если начать с мышц, то надо сказать, что не все они работают одинаково. Рассмотрим мышцы глаза и руки у человека. Глазные мышцы должны работать с высокой скоростью и большой точностью, быстро ориентируя глазное яблоко в пределах нескольких дуговых минут. В то же время глазной мышце не приходится справляться с такими внешними задачами, как поднимание груза. Тонкое управление, требуемое при движении глаза, требует высокого иннервационного индекса — отношения числа нейронов, аксоны которых оканчиваются на наружной мембране мышечных клеток, к числу мышечных клеток.
Для глазной мышцы иннервационный индекс составляет 1:3; это значит, что аксонные окончания одного мотонейрона выделяют свой медиатор не более чем на три отдельные мышечные клетки. (Мотонейрон — это такой нейрон, тело которого лежит в спинном мозгу, а аксон оканчивается на мембране мышечной клетки.) По-иному обстоит дело с мышцами руки: аксонные окончания одного мотонейрона, например иннервирующего бицепс, могут действовать своим медиатором на сотни мышечных волокон, и поэтому у такой мышцы иннервационный индекс составляет всего 1:100. В результате действие одной двигательной единицы мышцы конечности — одно быстрое сокращение (twitch), возникающее под влиянием одного импульса, вызывающего выделение медиатора из окончаний одного мотонейрона, — соответственно оказывается грубым.
Двигательные единицы мышц различаются также по тому, насколько они подвержены утомлению. На одном конце шкалы лежат двигательные единицы медленного сокращения, способные длительно функционировать без утомления. Такие единицы могут быть активными в течение длительного времени, но развивают сравнительно небольшое мышечное напряжение. На другом конце шкалы находятся двигательные единицы быстрого сокращения; они могут создавать высокие пики мышечного напряжения, но быстро утомляются. Такие единицы обычно иннервируются мотонейронами с диаметром аксонов и скоростью проведения нервного импульса выше средних.
В одной и той же мышце содержатся волокна и быстрых и медленных двигательных единиц. В 1968 г. шведские исследователи Э. Кугельберг (Е. Kugelberg) и Л. Эдстром (L. Edstrom) нашли способ определять, какие отдельные моторные волокна относятся к данной двигательной единице. Длительной стимуляцией аксона одного мотонейрона эти исследователи вызывали продолжительное сокращение мышечных волокон одной двигательной единицы. Сокращение приводило к истощению в отдельных мышечных волокнах запаса гликогена, который является источником энергии. При специальном окрашивании ткани на гликоген волокна истощенной двигательной единицы принимали вид белых «теней», рассеянных среди розовых волокон с нормальным запасом гликогена.
Такой гистохимический эффект представляет собой демонстрацию биохимической реакции в живой микроанатомической структуре. Так, применив подход Кугельберга и Эдстрома, Р. Берк (R. Burke) с сотрудниками из Национальных институтов здравоохранения показал с помощью гистохимического окрашивания, что «быстрые» мышечные единицы, используя в качестве источника энергии аденозинтрифосфат (АТФ), расщепляют его ферментативным путем быстрее, чем это происходит в «медленных» двигательных единицах. Это ферментативное расщепление считается одним из важных факторов, определяющих присущую мышце скорость сокращения. Таким образом, гистохимические данные помогают объяснить различия в скорости сокращения. Равным образом, гистохимическое исследование других ферментов — тех, которые расщепляют сахара и жиры, — помогает объяснить весьма существенные различия в утомляемости между двумя видами двигательных единиц.
Каково значение этих противоположных свойств двигательных единиц для организации движения? Посмотрим, как двигательные единицы мышцы последовательно «вовлекаются» в процесс движения. В общем мышечное напряжение регулируется двумя путями. Первый состоит в контроле над числом двигательных единиц, вовлекаемых в активность. Второй — в регуляции частоты импульсации вовлеченных единиц. Первыми вовлекаются единицы медленного сокращения, не склонные к утомлению и развивающие сравнительно слабое напряжение. Последними вовлекаются двигательные единицы быстрого сокращения, т. е. те, которые дают высокие пики напряжения, но быстро утомляются.

Второй срез той же мышцы был окрашен для определения относительной способности мышечных белков расщеплять аденозинтрифосфат (АТФ). Темная окраска свидетельствует о более высокой расщепляющей активности. Три волокна-метки очень слабо окрашены (схема справа); такая низкая активность характерна для медленных мышечных волокон.
Третий срез окрашен, чтобы показать способность мышечных белков расщеплять АТФ после их предварительной обработки кислотой. Обратное соотношение интенсивности окраски по сравнению со вторым срезом дает дальнейшие сведения о химизме мышечного волокна.
Четвертый срез был окрашен, чтобы показать относительную способность мышечных волокон к окислительному метаболизму, определяемому по наличию ключевого фермента в митохондриях клетки. Три волокна медленной двигательной единицы (см. схему) находятся среди интенсивно окрашенных волокон; картина согласуется с представлением о меньшей утомляемости таких двигательных единиц.
Э. Хеннемен (Е. Henneman) из Гарвардской медицинской школы внес важный вклад в общее понимание порядка вовлечения в активность отдельных двигательных единиц. Он отметил, что напряжение мелких мышц создается и точно контролируется избирательной мобилизацией разного числа мелких двигательных единиц. Между мелкими и более крупными единицами существует немалая разница. Например, самая крупная двигательная единица в икроножной мышце человека развивает напряжение в 200 раз больше, чем самая мелкая. Когда требуется общее усиление работы мышц, то, как установил Хеннемен, в активность вовлекаются более крупные двигательные единицы, что создает большие элементарные приросты напряжения. Это означает, что по мере увеличения общего напряжения оно создается меньшим числом дополнительных единиц. Разумеется, когда обстоятельства требуют резкого повышения общего напряжения мышцы, двигательные единицы вовлекаются не последовательно, а активируются практически одновременно.
Так обстоит дело с разными видами мышц и их двигательными единицами. Теперь посмотрим, что заставляет эти единицы сокращаться. Мышечные сокращения совершаются благодаря тому, что медиатор ацетилхолин, который выделяется в нервно-мышечном соединении при каждом импульсе, исходящем от мотонейрона, способен вызвать импульс в мышечной клетке. Блокирование передачи на уровне соединения, например при помощи алкалоида кураре, препятствует мышечному сокращению. Такую блокаду воспроизводили и у людей. Испытуемым вводили кураре в условиях искусственного дыхания. Во время паралича, вызванного кураре, выделение ацетилхолина аксонами мотонейронов продолжается, но блокируется взаимодействие медиатора с соответствующими рецепторами на мембране мышечной клетки; в результате мышцы перестают реагировать на команды, поступающие от коры больших полушарий. Испытуемые продолжали мыслить и чувствовать, но внешние проявления деятельности мозга исчезали. Речь, выражение лица, способность направлять взор — все эти формы поведения зависят от сокращения мышц.
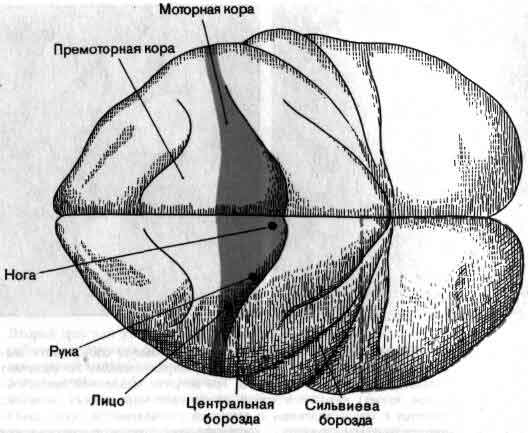
Головной мозг макака, вид сверху. Помечены разные участки моторной коры больших полушарий. Окрашенная полоса обозначает ту часть мозга, которая удалена, чтобы показать подкорковые структуры на соседнем рисунке.
Многие современные представления о механизмах движения возникли на основе трудов английского физиолога Шеррингтона, который в начале века занимался изучением функции мотонейронов в некоторых рефлекторных формах двигательной активности, таких, как чесание и ходьба. Сигналы, идущие от многих различных областей головного мозга, часто воздействуют на несколько одних и тех же мотонейронов спинного мозга. Установив этот факт, Шеррингтон охарактеризовал мотонейроны как «общий конечный путь», связывающий головной мозг с мышечным актом. Он изучал движение мышц у животных после перерезки спинного мозга, т.е. после нарушения связи мотонейронов с головным мозгом.
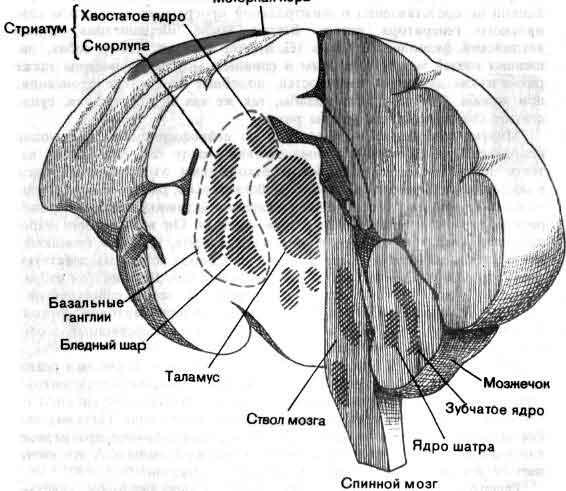
Головной мозг макака, вид сзади. Сектор, величиной 90°, удален, чтобы показать некоторые детали подкорковых структур. Цветная пунктирная линия окружает базальные ганглии левого полушария - части стриатума,скорлупу и хвостатое ядро, а также смежный с ними бледный шар. Ближе к средней линии лежит левый таламус, а по обе стороны от средней линии — правая половина мозжечка, так что видны его внутренние части - зубчатое ядро и ядро шатра. Моторная область коры больших полушарий (окрашена) - филогенетически более поздняя структура по сравнению с базальными ганглиями и мозжечком. Проводимые в настоящее время исследования показывают, что импульсация нейронов моторной коры вызывается сигналами, приходящими в нее через таламус от филогенетически более древних подкорковых структур.
Шеррингтон установил, что через несколько месяцев после такой перерезки у собаки удавались вызвать чесательный рефлекс механическими стимулами - щекотанием кожи или легким потягиванием за волосок где-нибудь на обширной поверхности спины. Описывая эти реакции, он указал, что такие движения «происходили без видимого нарушения направления или ритма». Работа Шеррингтона по чесательному рефлексу привела к современной концепции о «запускаемом движении», основанной на представлении о «центральной программе» с участием спинального генератора ритма. Вскоре после Шеррингтона другой английский физиолог Г. Браун (G. Brown) показал, что у собак, лишенных связей между головным и спинным мозгом, возможны также ритмические движения конечностей, подобные тем, какие происходят при ходьбе. Очевидно, для ходьбы, так же как и для чесания, существуют спинальные генераторы ритма.
Многие теперешние исследования по нейрофизиологии локомоции направлены на выяснение взаимодействия между тем, что можно назвать центральными программами, исходящими от головного мозга, и сенсорными обратными связями. Действительно, работа Шеррингтона особенно касалась того, какими способами активность мотонейронов регулируется сенсорными обратными связями. Он ввел термин «проприоцепция» для обозначения сенсорных входов, которые возникают в процессе вызываемых из центра движений, когда «стимулы, действующие на рецепторы, возникают в самом организме». Шеррингтон избрал приставку «проприо» (от латинского «proprius - «свой собственный»), так как он считал, что главная функция проприоцепторов состоит в том, чтобы по механизму обратной связи давать информацию о собственных движениях организма.
Мышечные проприоцепторы делятся на два вида. Один вид реагирует на удлинение, другой - на напряжение. Мышечные рецепторы длины посылают волокна в спинной мозг и образуют там синапсы на мотонейронах, которые оканчиваются на тех же мышцах. Поэтому любое усиление активности рецептора длины, происходящее при удлинении мышцы, активирует мотонейроны удлиненной мышцы. А это вызывает ее сокращение, противодействующее удлинению.
Рецепторы напряжения - второй вид проприоцепторов - чувствительны не к удлинению, а к силе; их активация ведет к торможению связанных с ними мотонейронов. Таким образом, когда прирост напряжения мышцы активирует эти рецепторы, в ответ они действуют на связанные с ними мотонейроны, и это приводит к снижению силы. Поэтому как рецепторы длины, так и рецепторы напряжения можно считать тем, что инженер назвал бы системой управления с помощью отрицательной обратной связи. Данная система поддерживает свою стабильность, противодействуя изменениям длины и напряжения мышцы.
Работа такого сервомеханизма с отрицательной обратной связью станет яснее, если рассмотреть в качестве примера какую-либо такую систему в действии. Представим себе человека, который пытается без наличия внешних помех удерживать свою руку неподвижно вытянутой в сторону. Разумеется, рука при этом непроизвольно слегка колеблется, особенно когда устанет. Например, случайное невольное ослабление напряжения в мышцах, сопротивляющихся силе тяжести, приведет к удлинению этих мышц. А вследствие этого усилится активность одних проприоцепторов - рецепторов длины - и в то же время (вследствие снижения напряжения в мышце) уменьшится активность других проприоцепторов - рецепторов напряжения.
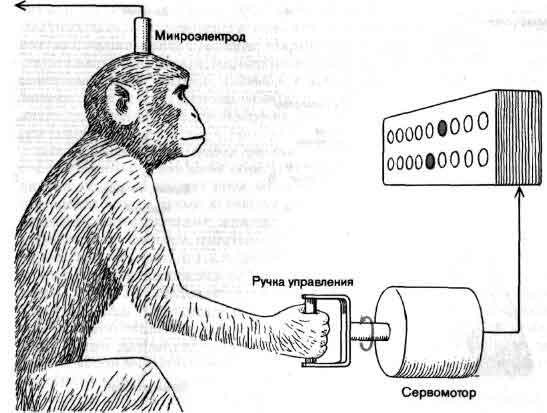
Подопытные обезьяны оказались способными обучаться точным мышечным реакциям на стимулы. В этом опыте экспериментатор зажигал одну из девяти ламп в верхнем ряду панели. Поворотом рукоятки обезьяна могла перемещать верхнюю лампу влево или вправо и получала вознаграждение, когда обе лампы оказывались на одной линии. Микроэлектрод, вживленный в область моторной коры, связанную с точными манипуляциями, регистрировал активность клеток, участвующих в мышечной реакции. Даже незначительное произвольное движение сопровождалось резким усилением активности моторной коры; доля разряжающихся клеток головного мозга была гораздо больше доли активных мотонейронов в спинном мозгу.
Несмотря на то что эти изменения идут в противоположных направлениях, их центральные эффекты не вычитаются один из другого, а суммируются: усиленная импульсация рецепторов длины возбуждает мотонейроны, действующие на мышцы, а ослабление импульсации рецепторов напряжения снимает торможение с тех же самых нейронов. Это синергическое действие двух видов проприоцепторов вступает в силу, когда изменения длины мышцы происходят вследствие непроизвольных изменений в ее напряжении, вызываемых внутренними причинами, но оно не возникнет, если изменения длины и напряжения произойдут вследствие приложения или удаления внешней силы. Например, удлинение мышцы, вызванное увеличением внешней нагрузки, приводит к усилению активности рецептора длины, связанному не с понижением, а с повышением активности рецепторов напряжения. Создавая термин «проприоцептор», Шеррингтон привлек внимание к существенным различиям в нейронной организации, лежащей в основе активного и пассивного движений. В данном контексте «активные» означает собственные движения испытуемого, а «пассивные» — перемещения, производимые внешними силами.
Опыт с «подготовкой к движению» требовал дополнительного обучения обезьян. Их приучали удерживать рукоятку в фиксированном «нейтральном» положении и быть готовыми толкнуть ее вперед через некоторое неопределенное время после вспыхивания зеленого светового сигнала (А) или притянуть назад через некоторое время после вспыхивания красного сигнала (Б). Активность одного нейрона в моторной коре регистрировалась в течение одной секунды до и после сигнальных вспышек; его импульсы записывались в виде горизонтальных рядов точек, где каждый ряд соответствует одному предъявлению стимула. Как показывают эти растровые записи, подготовка к толканию рукоятки вперед вызывала усиление нейронной активности, а подготовка к притягиванию ее вызывала ослабление активности.
Концепция Шеррингтона о связи между рецепторами длины в мышце и движением вообще получила яркое воплощение, когда шведский физиолог Л. Лекселл (L. Leksell) обнаружил, какую роль играют специальные нейроны, называемые гамма-мотонейронами. В отличие от обычных, или альфа-мотонейронов, которые действуют на мышечные волокна, производящие мощное сокращение, гамма-мотонейроны действуют на особые мелкие мышечные волокна, которые регулируют чувствительность рецепторов длины. Следовательно, мотонейроны также бывают двух видов. Один вид действует на мышцы, производящие движения тела (альфа), а второй служит для оптимизации работы рецепторов длины (гамма).
Итак, разные виды мышц, разные виды мотонейронов и связанные с ними системы управления образуют элементарные компоненты механизмов, осуществляющих произвольные и рефлекторные движения. Соотношения между этими компонентами можно изучать в лаборатории на самых разнообразных животных. Например, многие исследования управления движением производятся теперь на моллюсках, а также на членистоногих - насекомых и ракообразных. Простота нервной системы у этих беспозвоночных представляет для исследователя заметные выгоды. Однако эти животные лишены коры большого мозга и связанных с ней структур. Для того чтобы узнать, как головной мозг посылает сигналы мотонейронам и таким образом управляет движениями человека, нужны данные, получаемые на животных, которые обладают не только корой большого мозга, но и ее специализированным отделом — моторной корой, управляющей движением.
Моторная кора была открыта в 1870 г., когда обнаружили, что электрическим раздражением коры большого мозга можно вызвать движения тела. Опыты с такой стимуляцией подтвердили вывод, к которому уже до этого пришел английский невропатолог Дж. Джексон (J. Jackson) на основании клинических наблюдений. Он заметил, что раздражающий очаг повреждения в коре одного полушария большого мозга может вызывать эпилептические движения противоположной стороны тела. Результаты первых опытов по стимуляции, произведенных на собаках, были подтверждены в 1873 г. английским невропатологом Д. Ферье (D. Ferrier) на обезьянах.
Эти исследования моторной коры оказали большое влияние на неврологическое мышление. Чтобы оценить это полностью, надо учесть, что до 1870 г. думали, будто кора больших полушарий служит только для мышления. Джексон выразил тогдашнее общее мнение в следующих словах: «Идея о том, что большие полушария служат для движения, видимо, встречает непреодолимые возражения... Причина этого, как я думаю, лежит в том, что по существующему представлению, извилины [коры] существуют не для движений, а для мыслей».
Следующий шаг был сделан в 1874 г. Он состоял в открытии особой группы гигантских нейронов, которые образуют проводящий путь между моторной корой и спинным мозгом. В этом году русский анатом В. Бец обнаружил в двигательной коре обезьяны и человека необычайно крупные нейроны. Теперь их называют клетками Беца. Было установлено, что аксоны этих клеток спускаются по головному мозгу и образуют прямые связи со спинномозговыми мотонейронами, в частности с нейронами, управляющими теми мышцами, которые служат человеку для точных движений пальцев и речи. Дальнейшее исследование показало, что, казалось бы, непропорционально большая часть моторной коры управляет очень малой долей всей мускулатуры человеческого тела; это наглядно показано на знаменитых «гомункулусах» - картах моторной коры, составленных У. Пенфилдом (W. Penfield) и его сотрудниками из Монреальского неврологического института. Теперь найдены некоторые прямые связи между нейронами моторной коры и мотонейронами грудного отдела спинного мозга, где множество мотонейронов приводит в действие межреберные дыхательные мышцы.
На первый взгляд кажется странным, что нейроны моторной коры, управляющие главным образом точными движениями, оканчиваются на мотонейронах, которые контролируют такое автоматическое и примитивное действие, как дыхание. Но, как указывает английский физиолог Ч. Филиппе (Ch. Phillips), эти связи головного мозга со спинным, вероятно, имеют отношение не к дыханию, а к использованию дыхательных мышц в таких искусных актах, как речь и пение. Таким образом, проекция от моторной коры создает новую регуляцию мышц, предназначенных для старых рефлекторных актов. Как установил сто лет назад Джексон, сама по себе утрата кортико-спинальных связей не парализует мышцы, она препятствует использованию их в некоторых движениях. В частности, разрушение кортико-спинальных путей к грудным мотонейронам не сказывается на использовании дыхательных мышц для дыхания, хотя эти мышцы становятся непригодными для речи.
За последнее десятилетие многое стало известно о контроле над произвольным движением со стороны моторной коры. Новые данные получены в значительной степени благодаря хитроумным методикам, позволяющим вводить микроэлектроды в головной мозг подопытного животного (обычно обезьяны), способного выполнять искусные движения. Пользуясь этими методиками, мы с К. Фроммом (С. Fromm) из Дюссельдорфского университета провели в Национальном институте охраны психического здоровья исследование свойств двигательной коры, лежащих в основе ее решающей роли в точном управлении движениями кисти. Это того же рода точное управление, какое дает возможность хирургу, глядя в микроскоп, передвигать хирургический инструмент с точностью до малых долей миллиметра.
Нам с Фроммом казалось, что если точные мелкие движения управляются выходными сигналами из моторной коры, то тогда ее нейроны должны сильно модулироваться даже мельчайшими флуктуациями мышечной активности. Кроме того, точное управление кистью очень зависит от сенсорной обратной связи, и поэтому мы склонялись к той точке зрения, что активность моторной коры при точно контролируемых движениях кисти должна быть под непрерывным контролем отрицательной обратной связи по замкнутой петле.
Опыт со «стартом», продолжение опыта с подготовкой к движению, имел цель рассмотреть соотношение между произвольными и рефлекторными реакциями. «Стартовый сигнал», производимое мотором перемещение рукоятки из ее нейтрального положения, возникал через одну-пять секунд после включения светового сигнала к подготовке. Активность нейрона моторной коры регистрировали в течение секунды до и после стартового сигнала. Вызывало ли движение рукоятки реакцию толкания (А) или же притягивания к себе (Б), немедленная реакция обезьяны была чисто рефлекторной, о чем на растровых записях говорит усиление нейронной активности сразу же после стимула (через 0,5 секунды). Через 40 миллисекунд этот рефлекторный ответ сменялся произвольным «молчанием» нейрона, связанным с подготовкой к притягиванию рукоятки (белый уча» сток на нижнем растре). После короткого рефлекторного ответа и более продолжительной произвольной реакции (толкания), видных на верхнем растре, животное возвращало рукоятку в нейтральное положение перед следующей пробой; для этого нужно было потянуть ее, что создавало зону молчания, которая видна на верхнем растре (с правой стороны). Точно так же толкание рукоятки для возвращения ее в нейтральное положение вызывало возобновление нейронной активности, видимое на нижнем растре.
Для проверки наших предположений мы начали с того, что обучали обезьян точно передвигать рукоятку прибора. Вращение рукоятки управляло панелью со световыми сигналами; за точные движения обезьяна получала вознаграждение. При каждом тесте регистрировалась импульсация клеток Беца в моторной коре животного. Мы установили, что малейшее перемещение рукоятки сопровождалось поразительным усилением активности этих клеток. Так, доля нейронов моторной коры, которые разряжались при управлении этими тонкими движениями, была гораздо больше доли участвующих в них спинальных мотонейронов.
Была отмечена еще одна особенность в основе той роли, какую играет моторная кора в управлении точными движениями. Это система отрицательной обратной связи, автоматически регулирующая выходную активность моторной коры. Система была сосредоточена преимущественно на тех корковых нейронах, которые контролируют самые точные мелкие движения. Анатомические пути возврата в двигательную кору в системе отрицательной обратной связи детально еще не прослежены. Во всяком случае, одним источником сигналов является соматосенсорная область коры, лежащая непосредственно позади моторной и соединенная с ней множеством связей. Участки соматосенсорной коры, получающие сигналы от кисти, очевидно передают сигналы в моторную кору, замыкая петлю (хотя, вероятно, не единственную) в этой системе обратной связи.
Осуществляемый моторной корой контроль, о котором шла речь выше, имеет решающее значение для возникновения и стабилизации самых точных движений человека. Что происходит, когда человек хочет совершить движение, которое направлено против нормальной рефлекторной реакции? Для исследования этого вопроса мы с Дз. Тандзи (J. Tanji) из Университета Хоккайдо изучали активность нейронов двигательной коры у обезьян, которые были обучены реагировать на непроизвольное движение своей руки. Это обучение иногда требовало мышечной реакции, полностью противоположной нормальному рефлекторному движению. Воспользуемся аналогией с человеком и представим себе испытуемого, который стоит прямо. Он получил инструкцию: когда экспериментатор толкает его, он должен наклониться вперед, не переставляя ног. Испытуемый сначала подготовится к движению вперед, а затем будет ждать толчка. Если толчок последует сзади, то для сохранения равновесия испытуемый рефлекторно отогнется назад. Для осуществления центрально запрограммированного движения, направленного вперед в соответствии с инструкцией, он должен теперь выйти из способа реагирования по замкнутой петле, который сохраняет ему равновесие, и перейти к реакции по открытой петле, которая приведет к тому, что он качнется вперед (в данном случае упадет в сетку или на руки к тому, кто стоит наготове, чтобы поймать его).
Мы с Тандзи обучали обезьян отвечать на такую же последовательность событий. Животные начинали с того, что точно устанавливали рукоятку и удерживали ее в неподвижности в течение нескольких секунд. Во время этого короткого периода выход сигналов из их моторной коры к мышцам руки регулировался отрицательной обратной связью по замкнутой петле. Затем включалась цветная лампа. Ее цвет показывал обезьяне, как реагировать на предстоящее перемещение рукоятки извне. Если свет был красный, обезьяна должна была тянуть рукоятку назад, если зеленый - толкать вперед. Обезьяны получали вознаграждение за правильные ответы на движения рукояток после получения обоих сигналов. Подготовка к реакции на эти сигналы занимала около 200 миллисекунд.
Регистрируя импульсацию нейронов моторной коры, мы отметили, что после подготовки к движению животному нужно было всего 40 миллисекунд, чтобы правильно реагировать на движения рукоятки. В коротком интервале, следующем за движением, корковый контроль резко переходил от обратной связи по замкнутой петле (которая рефлекторно обеспечивает устойчивость позы) к связи по открытой петле, которая была нужна, чтобы совершить запрограммированное движение.
Итак, соматосенсорная область коры больших полушарий выполняет функцию передачи сигналов, контролирующих выходную активность моторной коры по принципу замкнутой петли. Но соматосенсорная область не посылает тех сигналов, которые лежат в основе запрограммированных движений, осуществляемых по принципу открытой петли, несмотря на (а не вследствие) рефлекторные эффекты соматосенсорной входной активности. Сигналы, связанные с запрограммированными движениями, приходят в двигательную кору из подкорковых структур, в особенности из мозжечка через таламус. П. Стрик (P. Strick) из Национальных институтов здравоохранения поставил опыты, которые показали, что в центрально запрограммированном управлении по открытой петле участвует путь, идущий от мозжечка через таламус в кору. Проводя опыты в основном по тому же плану, что и мы с Тандзи, Стрик регистрировал активность отдельных нейронов в определенных структурах мозжечка.
Он обучал своих обезьян двигать рукой в ответ на поданный знак, состоявший в перемещении руки посредством производимого извне передвижения рукоятки, которую держала обезьяна. Стрик обнаружил, что если предварительно у обезьян при помощи системы красных и зеленых сигналов было запрограммировано перемещение рукоятки в том или другом направлении, то эта программа оказывала сильное влияние па реакции определенных, так называемых зубчатых нейронов мозжечка: эти нейроны разряжались в пределах 30 миллисекунд после того, как был подан знак в виде перемещения руки. Тем самым у активности зубчатого нейрона было 10 миллисекунд на то, чтобы пройти через таламус и вызвать, центрально запрограммированную активность моторной коры (проявляющуюся через 40 миллисекунд после подачи знака).
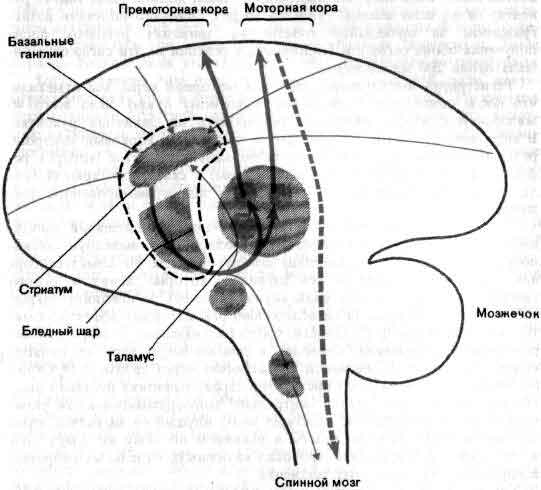
Проводящие пути между некоторыми областями коры больших полушарий и определенными подкорковыми структурами показаны на схеме продольного разреза большого мозга обезьяны. Тонкими стрелками обозначены входы в базальные ганглии (структуры внутри черных границ), которые проводят разнообразную информацию от коры больших полушарий. Один из компонентов, стриатум, является главным связующим звеном между ассоциативными областями коры и моторной областью. Путь, выходящий из стриатума, проходит к разделенному на две части бледному шару (толстая стрелка); это образование в свою очередь посылает связи в другую подкорковую структуру - таламус, в особенности к двум его ядрам: ventralis lateralis и ventralis anterior (толстая и менее толстая стрелки). Пути, выходящие из таламуса, идут главным образом в премоторную область коры (толстая стрелка); дополнительные пути (менее толстая стрелка) направляются в моторную кору, откуда сигналы идут к мотонейронам спинного мозга (пунктирная стрелка). Подкорковые входы играют важную роль в центральном программировании движений.
Факты, полученные Стриком, согласуются с данными У. Тэтча мл. (W. Thatch, Jr.) из Вашингтонского университета. Работая в Национальном институте охраны психического здоровья, Тэтч показал, что импульсация нейронов мозжечка намного опережала мышечную активность у обезьяны, обученной отвечать на световой стимул. Роль сигналов, идущих от мозжечка, в генерации активности моторной коры была также экспериментально показана В. Бруксом (V. Brooks) с сотрудниками из Университета Западного Онтарио. Они искусственно понижали температуру мозжечка у обезьян, а затем давали ей вернуться к норме. Активность нейронов двигательной коры измерялась до, во время и после охлаждения мозжечка. Было установлено, что во время охлаждения импульсация нейронов моторной коры и соответствующее запрограммированное движение запаздывали.
Кроме проведения сигналов от мозжечка в моторную кору таламус передает сигналы еще от одной подкорковой структуры, а именно от большой совокупности клеточных групп, объединяемых общим названием базальных ганглиев. Во время своей работы в Национальном институте охраны психического здоровья М. Де-Лонг (М. DeLong) из Университета Джонса Гопкинса показал, что клетки базальных ганглиев разряжаются задолго до произвольных движений, совершаемых животным в ответ на сигналы, Этот факт согласуется с результатами наблюдений, сделанных в неврологических отделениях; согласно этим наблюдениям, базальные ганглии имеют решающее значение для самых ранних стадий инициации движения - стадий, когда в результате еще не разгаданных процессов абстрактная мысль переводится в конкретный двигательный акт.
Рефлексы и произвольные движения не противоположны друг другу. Это признал еще сто лет назад Джексон, когда писал, что произвольные движения подчиняются законам, управляющим рефлекторными актами. Однако если произвольным движениям нельзя дать определение путем исключения, т. е. определить их как нечто такое, что не является рефлексом, то в таком случае как же их определить? Самое сжатое из известных мне определений дал шведский нейрофизиолог Р. Гранит (R. Granit) в своей недавно опубликованной книге «Целенаправленный мозг»: «Произвольным в произвольном движении является его цель». С такой точки зрения произвольные свойства моторного акта надо рассматривать в аспекте цели совершаемого действия. В то же время действительные события, лежащие в основе достижения цели, строятся из разнообразных рефлекторных процессов.
Недавно я обсуждал этот вопрос с русским кибернетиком Виктором Гурфинкелем. Он тоже определяет произвольное движение в связи с его целью. Описывая виды процессов, на которых основано произвольное движение, он рассказал мне о некоторых кинезиологических исследованиях, направленных на оценку характеристик управления движением у лучших стрелков-спортсменов. Гурфинкель указал, что важным свойством меткого стрелка является его способность удерживать пистолет неподвижным. Как показали исследования электромиографических и кинематических характеристик снайперов, при том что многие части их тела двигались, пистолет оставался практически неподвижным. Положение руки стрелка в пространстве стабилизировали всякого рода рефлекторные механизмы; вестибуло-окулярная система, вестибуло-спинальная система и другие. Приведенный Гурфинкелем пример со стрелком вызывает в памяти точку зрения У. Джеймса (W. James) относительно сущности произвольного движения. Почти сто лет назад он писал: «Меткий стрелок в конце концов думает только о точном положении цели, певец — только о совершенстве голоса, эквилибрист — только о конце шеста, колебаниям которого он должен противодействовать».
Здесь мы видим поразительное согласие между учеными самых разных школ, дисциплин и эпох. Гранит разделяет точку зрения Джеймса и Гурфинкеля. Гранит и Гурфинкель принимают представление Шеррингтона о том, что целенаправленные движения строятся на основе рефлекторных процессов. Так же думал английский невропатолог К. Уилсон (К. Wilson). Он писал в 1928 г., что «большая часть всякого произвольного движения непроизвольна и лежит вне сознания».
Каковы же в таком случае свойства, которые отличают целенаправленные движения от лишенных цели? Несомненно, существует много видов непроизвольных движений, например возникающие при некоторых нервных болезнях. Они наблюдаются при различных поражениях базальных ганглиев. Характерно заметное нарушение произвольных движений: они или не совершаются, когда нужны, или же возникают, когда не нужны. Как указал Уилсон, у больных хореей Гентингтона возникают невольные движения, которые похожи на движения, «выполняемые по велению воли. Кажется, что каждое новое движение направлено на определенную цель, которой оно никогда не достигает». Мышечная деятельность, связанная с невольными движениями, схожа с деятельностью, связанной с произвольными движениями здорового человека. Но при хорее движения лишены цели.
По существу ясно, что законы рефлекторной деятельности, регулируемой на уровне спинальных мотонейронов, действуют также на уровне моторной коры при произвольных движениях. В свою очередь на нейроны моторной коры влияют транскортикальные входы. Таким образом, двигательная кора млекопитающих - филогенетически новая часть головного мозга — подчиняется тем же законам рефлекторной деятельности, которые характерны для его более древних частей. Кроме того, на моторную кору действует вторая мощная группа входов. Эти входы лежат в основе генерируемых изнутри моторных программ, которые создаются в результате активности базальных ганглиев и мозжечка и доходят до моторной коры через таламус.
Таким образом, из двух основных классов входов, которые воздействуют на моторную кору головного мозга и порождают поток импульсов, идущий к спинному мозгу, наиболее понятным представляется класс входов, которые работают автоматически, т. е. транскортикальная петля: она работает в соответствии с шеррингтоновскими принципами рефлекторной деятельности. Второй класс входов, берущих начало в базальных ганглиях и мозжечке и идущих в моторную кору через таламус, составляет более сложную картину. Чтобы понять произвольное движение, нужно понять те виды информации, которые перерабатываются подкорковыми структурами, и установить, как выходы из мозжечка и базальных ганглиев взаимодействуют в таламусе. Для исследователей механизмов головного мозга, управляющих движениями, эти вопросы являются самыми главными.
Н. ГЕШВИНД
Специализация человеческого мозга
Некоторые высшие функции, такие, как речь, связаны со специализированными областями человеческого мозга. То же и в более крупном масштабе: два полушария большого мозга специализированы для различных видов умственной деятельности
Нервные системы всех животных имеют ряд общих фундаментальных функций, среди которых можно особо выделить управление движениями и анализ раздражителей. Отличительной чертой человеческого мозга является разнообразие высоко специализированных действий, которым он способен обучаться. Превосходнейшим примером является речь: никто не рождается знающим язык, но практически все научаются говорить и понимать произносимые слова, и людей любого уровня культуры можно научить писать и читать. Восприятие музыки также является универсальным свойством человека: люди, не проходившие специального обучения, способны узнавать и воспроизводить десятки мелодий. Добавим к этому, что почти каждый может рисовать простые фигуры, да и уменье хорошо рисовать тоже встречается не так уж редко.

Способности каждого из полушарий человеческого мозга проверялись на испытуемом, два полушария которого были разобщены хирургическим путем. Операция заключалась в перерезке двух главных пучков нервных волокон, соединяющих полушария, - мозолистого тела и передней комиссуры. В процессе тестирования испытуемому предъявляли каждый из узоров средней колонки и просили воспроизвести эти узоры, размещая нужным образом цветные блоки. Задание выполнялось либо одной правой рукой (которая связана главным образом с левым полушарием), либо одной левой рукой (которая управляется в основном правым полушарием). Частота ошибок не зависела от руки, но типичные для каждой руки ошибки были совершенно различными. Эти результаты заставляют предполагать, что каждая сторона мозга может использовать для выполнения такого рода задач свой определенный набор способностей; данное наблюдение согласуется с другими свидетельствами в пользу того, что два полушария специализированы для различных функций.
По крайней мере некоторые из этих высших функций человеческого мозга управляются специально предназначенными для них сетями нейронов. Уже более 100 лет известно, например, что по крайней мере две четко ограниченные области коры мозга существенны для лингвистических способностей; они кажутся созданными специально для обработки словесной информации. Определенные структуры на внутренней поверхности нижней части височной доли, в том числе гиппокамп, по-видимому, необходимы для долговременной памяти. В некоторых случаях функциональная специализация нервной системы оказывается очень узкой: так, на обеих сторонах коры большого мозга есть область, которая занимается в первую очередь узнаванием лиц. Похоже, что другие виды умственной деятельности также связаны с определенными нервными сетями. Например, можно думать, что музыкальные и артистические способности определяются специализирозанными системами в мозгу, хотя эти структуры пока еще не установлены.
Другой отличительной характеристикой человеческого мозга является распределение функций между двумя полушариями. О том, что человеческий мозг не полностью симметричен в своих функциях, можно догадаться хотя бы по такому наблюдению из повседневной практики: большинство людей отдает предпочтение правой руке, управляемой левой половиной мозга. Лингвистические способности также приурочены главным образом к левой стороне. По этой причине когда-то говорили, что левое полушарие мозга доминирует, и думали, что правая половина мозга играет подчиненную роль. В последние годы эта концепция была пересмотрена, так как стало очевидно, что каждое полушарие обладает своими «особыми талантами». Правое полушарие доминирует в таких свойствах, как музыкальность и распознавание сложных зрительных образов; оно также более важно для выражения и распознавания эмоций. В последние несколько лет эти функциональные асимметрии были соотнесены с анатомическими, и было положено начало исследованию их значения у других биологических видов, кроме человека.
У человека, как и у других видов млекопитающих, обширные области коры отведены под относительно элементарные сенсорные и моторные функции. Дуга, которая идет, грубо говоря, от уха до уха через крышу мозга, соответствует первичной моторной коре, осуществляющей произвольное управление мышцами. Параллельно этой дуге и прямо позади нее лежит соматосенсорная зона, которая получает сигналы от кожи, костей, суставов и мышц. Почти каждый участок тела представлен соответствующей областью как в первичной моторной, так и в соматосенсорной коре. В задней части мозга, и в частности на внутренней поверхности затылочных долей, располагается первичная зрительная кора. Первичные слуховые зоны находятся в височных долях; обоняние сосредоточено в определенной области на нижней части лобных долей.
Первичные сенсорные и моторные зоны специализированы в том смысле, что каждая предназначена для специфической функции, но сами эти функции имеют очень широкий спектр, так что данные зоны участвуют в самых разнообразных видах деятельности. Отметим, что у всех видов животных, имеющих хорошо развитую кору, найдены гомологичные области. В данной статье будут рассмотрены главным образом те зоны коры, которые управляют более узкой областью поведения. Некоторые из этих высоко специализированных зон являются общими для многих видов, тогда как другие присущи только человеку.
Серии экспериментов по обучению обезьян иллюстрируют, какими тонкими могут быть различия между двумя сетями нейронов. Обезьяну можно научить безошибочно выбирать один предмет или одну фигуру из пары. Задача делается несколько более трудной, если предметы предъявляются, а затем убираются, и обезьяне разрешается проявить свой выбор только после определенной задержки, в течение которой объекты спрятаны за экраном. Было обнаружено, что выполнение этой задачи заметно нарушается, если на обеих сторонах мозга разрушить небольшую область лобной доли. Затруднения в эксперимент можно также ввести, усложнив фигуры, но позволив делать выбор, не убирая тест-объекта. Повреждение совершенно другой области коры снижает способность выполнять эту задачу, но не влияет на решение задачи с задержкой.
Эти эксперименты иллюстрируют также один из основных способов получения информации о функциях мозга. Повреждение определенного участка в результате заболевания или травмы вызывает иногда вполне определенные нарушения поведения. Во многих случаях можно заключить, что некоторые аспекты затронутого поведения в норме зависят от той части мозга, которая была разрушена. У человека самая обычная причина повреждения мозга — это тромбоз мозговых сосудов; закупорка мозговой артерии приводит к гибели ткани, снабжаемой данным сосудом. К 1920 г. обследование больных со стойкими последствиями такого повреждения позволило идентифицировать ряд функциональных областей мозга, включая речевые зоны.
Изучение последствий поражения мозга все еще остается важным методом исследования функций мозга, но сейчас мы располагаем и другими методами. Один из наиболее важных был доведен до высокого уровня совершенства немецким нейрохирургом О. Ферстером (О. Foerster) и У. Пенфилдом (W. Penfield) из Неврологического института в Монреале. Во время хирургической операции на мозге производили электрическую стимуляцию различных его участков и изучали при этом реакцию больного, находившегося в сознании. Таким способом удалось составить карту зон, ответственных за ряд функций. Кроме важности этого метода для изучения мозга он имеет и клиническое значение, так как позволяет хирургу избежать областей, повреждение которых могло бы иметь пагубные последствия.
Ценные сведения были получены также при хирургических вмешательствах по поводу тяжелой эпилепсии. Один из методов избавления от частых эпилептических припадков (применяемый только тогда, когда все другие терапевтические методы не дают эффекта) состоит в удалении той области коры, где находится патологический очаг. Функциональные расстройства, которые возникают иногда в результате такого вмешательства, были подробно изучены Б. Милнер (В. Milner) из Неврологического института в Монреале.
Специализацию полушарий можно изучать у больных со стойкими повреждениями комиссур, соединяющих две половины мозга; наиболее важной из них является мозолистое тело. В первых таких случаях, изученных в конце XIX в. Ж. Дежрином (J. Dejerine) во Франции и Г. Липманом (Н. Lipmann) в Германии, повреждения были вызваны инсультом. Сравнительно недавно разделение полушарий путем перерезки комиссур стали применять при лечении эпилепсии. Обследования таких больных «с расщепленным мозгом», выполненные Р. Сперри (R. Sperry) из Калифорнийского технологического института и М. Газзанигой (М. Gazzaniga) из Медицинского колледжа Корнеллского университета, позволили получить более подробные сведения о функции разобщенных полушарий. Д. Кимура (D. Kimura), которая работает сейчас в Университете Западного Онтарио, была пионером разработки метода, названного дихотическим слушанием, который дает представление о специализации полушарий интактного человеческого мозга.
Наиболее детально были исследованы те специализированные области мозга, которые связаны с речью. В 1860-х годах французский исследователь П. Брока (Р. Вгоса) показал, что повреждение определенного участка коры неизменно вызывает афазию, или расстройство речи. Этот участок находится у края лобной доли, и сейчас его называют передней речевой зоной, или просто зоной Брока. Брока пошел дальше и сделал второе важное открытие. Он показал, что в то время, как повреждение этого участка на левой стороне мозга ведет к афазии, при сходном повреждении корреспондирующей области на правой стороне речь не страдает. С тех пор это открытие было многократно подтверждено: можно гарантировать, что более 95 процентов афазий, связанных с повреждением мозга, являются результатом поражения левого полушария.
Зона Брока граничит с лицевой областью моторной коры, управляющей мышцами лица, языка, челюстей и глотки. Когда при инсульте повреждается зона Брока, почти во всех случаях наблюдается также серьезное повреждение лицевой зоны левого полушария, и поэтому можно подумать, что расстройство речи вызвано частичным параличом мышц, необходимых для артикуляции. Легко показать несостоятельность такого объяснения. Во-первых, повреждение соответствующей области на правой стороне не приводит к афазии, хотя имеется частичный паралич лицевых мышц. Во-вторых, известно, что при афазии Брока мышцы, которые не выполняют своей речевой функции, в остальном действуют нормально. И далее, самое простое: больной с афазией Брока, которому очень трудно говорить, нередко прекрасно и легко поет. Наконец, речь больного с афазией Брока нарушена грамматически, что уже никак нельзя объяснить слабостью мышц.
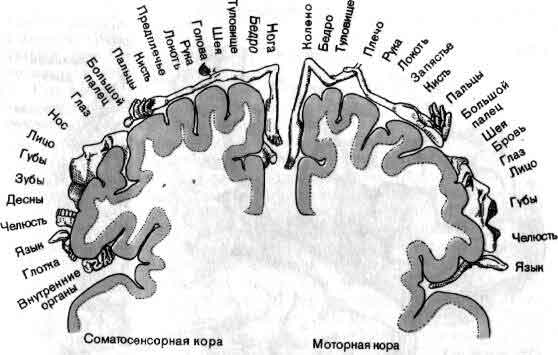
Специализация соматосенсорной и моторной зон коры мозга выражается в том, что каждый участок этих зон можно связать с определенной частью тела. Другими словами, большая часть тела может быть представлена на коре в виде схемы; результатом этого будут два искаженных гомункулуса. Искажения связаны с тем, что отведенная под данную часть тела площадь коры пропорциональна не величине этой части, а необходимой точности управления. У человека очень велики площади моторной и соматосенсорной зон, соответствующие лицу и рукам. Здесь показаны только половины каждой из таких корковых зон: левая соматосенсорная зона (которая получает сигналы преимущественно от правой половины тела) и правая моторная зона (которая осуществляет управление движениями левой половины тела).
Другой тип афазии описал в 1874 г. немецкий исследователь К. Вернике (С. Wernike). Этот тип афазии связан с повреждением другой области коры, находящейся также в левом полушарии, но не в лобной, а в височной доле. Эта область, именуемая теперь зоной Вернике, лежит между первичной слуховой корой и структурой, называемой угловой извилиной, которая, по-видимому, проходит между зрительным и слуховым центрами мозга. Позднее было установлено, что зона Вернике и зона Брока соединяются группой нервных волокон-дутообразным пучком.
Поражение каждой из этих зон — как зоны Брока, так и зоны Вернике — ведет к расстройствам речи, но характер этих расстройств совершенно различен. При афазии Брока речь становится затрудненной и медленной, с нарушенной артикуляцией. Ответ на вопрос часто имеет смысл, но он редко дается в виде грамматически правильно построенного предложения. Особые трудности возникают со спряжением глаголов, с произношением и со связующими словами, а также со сложными грамматическими конструкциями. В результате речь приобретает телеграфный стиль. Например, если больного спрашивают о назначении к зубному врачу, он отвечает невнятно и заикаясь: «Да... Понедельник... Папа и Дик... Среда девять часов... 10 часов... доктора... и... зубы». Такого же типа ошибки делаются и при письме.
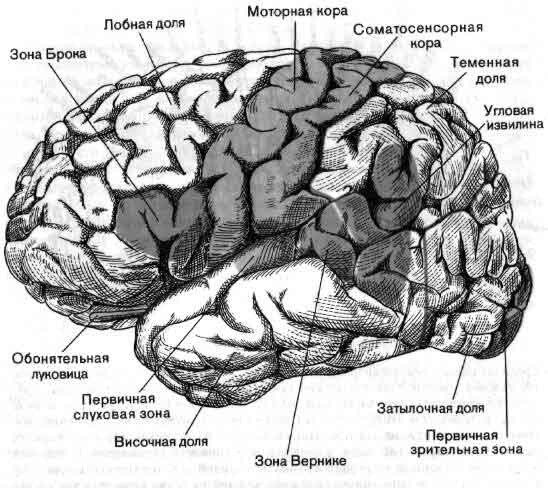
На карте коры человека показаны области, функциональная специализация которых установлена. Большая часть коры отведена под сравнительно элементарные функции: управление движениями и первичный анализ раздражителей. Эти области, включающие моторную и соматосенсорную зоны, а также первичные зрительные, слуховые и обонятельные области представлены у всех видов, которые имеют хорошо развитую кору, и вовлекаются в работу при многих родах деятельности. Некоторые другие области (темный цвет) более узко специализированы. Зоны Брока и Вернике участвуют в формировании и восприятии речи. Предполагается, что соотнесение зрительного и слухового представлений информации производит угловая извилина. Такие функциональные специализации обнаружены только на левой половине мозга; соответствующие области правого полушария не имеют аналогичной связи с лингвистическими способностями. Правое полушарие, которое здесь не показано, определяет свои собственные специфические способности, в частности касающиеся некоторых аспектов восприятия музыки и сложных зрительных образов. Однако анатомические зоны, ассоциирующиеся с этими способностями, определены не так хорошо, как речевые зоны. Даже в левом полушарии соотнесение функций с участками коры лишь приблизительное; некоторые зоны могут иметь иные функции, кроме указанных, а в осуществлении отдельных функций может принимать участие несколько зон.
При афазии Вернике речь фонетически и даже грамматически нормальна, но ее семантика нарушена. Слова обычно связываются без особого труда и имеют правильные окончания, так что высказывания носят характер правильно построенных предложений. Однако слова нередко оказываются неподходящими, и среди них встречаются бессмысленные слоги и сочетания слогов. Примечательно, что даже в тех случаях, когда отдельные слова правильны, высказывание в целом выражает смысл каким-то обходным путем. Так, больной, которого попросили описать картину, изображающую двух мальчиков, ворующих печенье за спиной у женщины, сообщил: «Матери нет здесь, она делает свою работу, чтобы получить ее лучше, но когда она смотрит, два мальчика смотрят в другое место. Она работает в другой раз».
На основе анализа этих дефектов Вернике сформулировал представление о программировании речи в мозгу. За прошедшие 100 лет были получены многочисленные новые данные, но основные принципы, установленные Вернике, все еще сохраняют свое значение. Согласно его модели, основная структура высказывания возникает в зоне Вернике. Затем она по дугообразному пучку передается в зону Брока, где включает детальную и координированную программу вокализации. Эта программа приходит в смежные лицевые области моторной коры, которая активирует соответствующие мышцы рта, губ, языка, гортани и т.д.
Зона Вернике не только принимает участие в речеобразовании, но и играет большую роль в понимании произносимых слов, чтении и письме. Когда слово прослушивается, звук вначале воспринимается первичной слуховой корой, но если его нужно воспринять как словесное сообщение, сигнал должен пройти через смежную зону Вернике. Когда слово читается, зрительный образ (из первичной зрительной коры) передается в угловую извилину, которая производит определенное преобразование, ведущее к появлению звуковой формы слова в зоне Вернике. Написание определенного слова в ответ на некоторую устную инструкцию требует того, чтобы информация прошла по тому же самому пути в обратном направлении: из слуховой коры в зоне Вернике и к угловой извилине.
Эта модель объясняет многие симптомы, характерные для афазий. Повреждение зоны Брока нарушает саму речь, но почти не сказывается на ее понимании. Повреждение же зоны Вернике сказывается на всех аспектах речи. Последствия определенных более редких поражений также согласуются с этой моделью. Так, например, разрушение дугообразного пучка, приводящее к разобщению зон Вернике и Брока, оставляет речь беглой и хорошо артикулированной, но делает ее семантически дефектной. Зона Брока работает, но не получает информации из зоны Вернике. Однако, поскольку последняя также функционирует, понимание произносимой речи и ее написание остаются почти нормальными. Письмо нарушается при всех афазиях, характеризующихся отклонением речи от нормы, однако пути, принимающие участие в письме, в деталях неизвестны.
Повреждение угловой извилины ведет к разъединению систем, участвующих в устной и письменной речи. При повреждении определенных участков этой извилины больные могут нормально говорить и понимать устную речь, но теряют способность писать. Создается впечатление, что понимание написанных слов требует того, чтобы в зоне Вернике возник их слуховой вариант. Повреждение угловой извилины, по-видимому, нарушает сообщение между зрительной корой и зоной Вернике, в связи с чем понимание письменной речи нарушается.
Хотя распределение лингвистических функций между несколькими участками коры сейчас подтверждено многочисленными данными, строгость этих распределений не следует преувеличивать. Пессимистическая точка зрения, согласно которой повреждение ткани в этих зонах неизбежно ведет к необратимому нарушению лингвистических способностей, необоснованна. На самом деле, часто наблюдается значительная степень восстановления. Нервная ткань, разрушенная в результате артериального тромбоза, не способна регенерировать, однако, по всей вероятности, функцию разрушенной области принимают на себя, хотя бы частично, другие отделы. В некоторых случаях восстановление, возможно, отражает существование другой обучающейся системы на противоположной стороне мозга, которая оставалась бездействующей до тех пор, пока доминирующая сторона не была повреждена. В ряде случаев эти функции берут на себя нейроны областей, смежных с поврежденной или окружающих ее. П. Уолл (P. Wall) из Лондонского университетского колледжа показал, что существует кайма из таких бездействующих, но потенциально активных клеток по краю соматосенсорной коры, и кажется весьма вероятным, что подобного рода кайма имеется в мозгу повсеместно. Дж. Mop (J. Mohr), который сейчас работает в Университете Южной Алабамы, вместе со своими сотрудниками показал, что при афазии Брока прогноз в смысле восстановления весьма благоприятен, если разрушенная область не слишком велика. Одна из интерпретаций этого открытия предполагает, что области по границе зоны Брока разделяют ее специализацию в латентной форме.
Хотя детали механизма восстановления неизвестны, было установлено, что у некоторых групп больных восстановление лингвистических способностей происходит быстрее, чем у других. У детей, особенно моложе восьми лет, часто наблюдается прекрасное восстановление, причем у левшей оно бывает чаще, чем у правшей. Даже те правши, у которых среди родственников выявляются левши (родители, братья и сестры или дети), более склонны к восстановлению, чем те, у которых таких родственников нет. Взаимосвязь между доминирующей рукой и восстановлением при повреждениях речевых зон указывает на то, что структуры, определяющие доминирование руки и речевые способности, в какой-то мере связаны.
Сосудистая система мозга играет важную роль в картировании функциональных зон коры. О функциях, которыми данная область коры обладает в норме, часто можно судить по расстройствам или нарушениям поведения, наступающим при повреждении области. Самая распространенная причина таких повреждений — это закупорка артерии, обслуживающей кору, что ведет к гибели тканей, снабжаемых этой артерией. Зоны Брока и Вернике были идентифицированы таким способом около 100 лет назад: в результате вскрытия было обнаружено, что у больных с характерными афазиями, или дефектами речи, имелись повреждения именно в этих областях левого полушария.
Существует мозговое нарушение, поражающее тем, что его проявление очень узко ограничено: это прозопагнозия, или неспособность узнавать лица. Способность узнавать людей по лицу сама по себе замечательна. С одного взгляда можно узнать человека только по чертам лица, даже если они изменились с годами или представлены в сильно искаженном виде, как, например, на карикатуре. У больных с прозопагнозией способность к подобной ассоциации утрачивается.
Что особенно поразительно в этом нарушении, так это его специфичность. Как правило, оно сопровождается малым числом других неврологических симптомов, исключая разве выпадение части поля зрения, иногда на обеих сторонах, а иногда только левой половины. Большинство мыслительных задач, включая те, которые требуют переработки зрительной информации, выполняется без особых трудностей; например, такой больной обычно может читать и правильно называть предметы. Чего он не может делать, так это назвать человека, посмотрев на него или на его фотографию. Иногда он даже неспособен узнать свою жену и детей. Когда знакомый человек заговорит, больной узнает голос и может немедленно назвать имя. Восприятие черт лица также не нарушается, поскольку часто больной может описать лицо в деталях и обычно способен соотнести фотографию, сделанную в фас, с профилем того же человека. Нарушение кажется приуроченным к формированию ассоциации между лицами и личностями.
Поражения, вызывающие прозопагнозию, столь же стереотипны, как и само расстройство. Повреждения обнаруживаются на нижней стороне обеих затылочных долей и простираются вперед к внутренней поверхности височных долей. Можно думать, что некоторая нейронная сеть внутри этой области специализирована для быстрого и надежного узнавания человеческих лиц. Может показаться, что непропорционально большая часть мозговых ресурсов растрачивается на весьма ограниченные задачи. Однако следует иметь в виду, что узнавание людей как индивидуумов - весьма ценная способность для столь высокосоциального биологического вида, как человек, и, возможно, в ее формировании сыграло роль сильное давление отбора.
Сходные способности, возможно, имеются и у других общественных видов. Г. Ван Хесен (G. Van Hoesen) — мой бывший сотрудник по Гарвардской медицинской школе, а теперь работающий в Университетском медицинском колледже в Айове, начал исследовать неврологическую основу узнавания лиц у макаков-резусов. Ему удалось доказать, что обезьяны способны с легкостью различать других обезьян по фотографиям. Однако нейронные структуры, участвующие в решении этой задачи, пока не идентифицированы.
До недавнего времени мало что было известно о физиологической основе памяти — одной из самых важных функций человеческого мозга. Однако посредством изучения некоторых узкоспецифических нарушений удалось выявить области или структуры мозга, участвующие в определенных процессах памяти. Так, например, исследование различных форм антероградной амнезии - неспособности усваивать новую информацию - выявило роль височных долей в процессах памяти. В частности, удивительные расстройства у одного больного, которого Милнер наблюдает более 25 лет, свидетельствуют о важном значении для памяти таких структур, как гиппокамп, находящийся на внутренней поверхности височной доли.
В 1953 г. этот больной перенес радикальную хирургическую операцию, при которой были разрушены значительная часть гиппокампа и ряд связанных с ним структур в обеих височных долях. После операции приобретенные ранее навыки и знания в значительной степени сохранились, и больной в состоянии нормально следить за текущими событиями. Создается даже впечатление, что он способен обычным образом воспринимать ограниченное количество новой информации.
Однако через короткое время большая часть этой информации становится для него недоступной.
Милнер опрашивала и обследовала этого больного через определенные промежутки времени после операции и обнаружила, что отмеченная у него антероградная амнезия мало изменилась за прошедшее время. У больного была выявлена, кроме того, обширная, хотя и выборочная, ретроградная амнезия (касающаяся времени, предшествовавшего операции), но она заметно уменьшилась. В отсутствие отвлекающих моментов больной может, скажем, удерживать в памяти трехзначное число в течение многих минут путем многократного его повторения или с помощью сложной мнемонической схемы. Если же хоть на мгновение отвлечь его внимание, он не может вспомнить ни числа, ни мнемонической схемы, на которую потратил столько усилий; он не может вспомнить даже саму задачу. Живя текущим моментом, больной неспособен заучить свой адрес или запомнить, где в доме хранятся предметы, которыми он ежедневно пользуется. Он не узнаёт людей, которые уже много лет регулярно его посещают. Подобные двусторонние операции, приводящие к таким нарушениям памяти, по очевидным причинам больше не производятся, но сходные поражения внутренней поверхности височных долей время от времени случаются при односторонних операциях на мозге у больных с необнаруженным повреждением противоположной доли. Результатом являются аналогичные нарушения, и, таким образом, роль внутренней поверхности височных долей в функционировании памяти в настоящее время общепризнана. Кроме того, тот факт, что у таких больных обычно сохраняются перцептивные способности, подтверждает правильность выдвигаемой многими исследователями идеи о существовании двух различных видов памяти-кратковременной и долговременной; последняя обеспечивает длительное хранение информации. У описанных выше больных, очевидно, нарушены процессы второго типа; мнения о природе этих нарушений пока противоречивы. Некоторые полагают, что нарушается способность консолидации, закрепления, т. е. передача информации из кратковременной памяти в долговременную; другие считают, что информация передается и сохраняется, но не может быть извлечена. Чтобы решить, кто из них прав, необходимо получить более точное представление о нейронных схемах памяти.
На первый взгляд кажется, что, подобно большинству других органов тела, мозг имеет совершенную билатеральную симметрию. Можно было бы ожидать, что две половины мозга и функционально так же эквивалентны, как две почки или два легких. В действительности многие из более специализированных функций присущи только либо одному полушарию, либо другому. Даже видимая анатомическая симметрия оказывается иллюзорной.
В первичных моторных и сенсорных областях коры распределение обязанностей между двумя полушариями подчиняется следующему простому правилу: каждая сторона мозга связана преимущественно с противоположной стороной тела. Большая часть нервных волокон на некотором участке своего пути переходит на противоположную сторону. Вследствие этого мышцами правой руки или ноги управляет преимущественно левая моторная кора, а сенсорные импульсы с правой половины тела идут преимущественно в левую соматосенсорную кору. Каждое ухо имеет связи со слуховой корой обеих половин мозга, но связи с контралатеральной стороной сильнее. Распределение сигналов от глаз несколько сложнее. Зрительные волокна распределены так. что образы, относящиеся к правой половине пространства, из обоих глаз проецируются на левую зрительную кору, а левая половина поля зрения обоих глаз проецируется в правое полушарие. В результате такой организации контралатеральных связей сенсорные и моторные функции двух полушарий разделены, но они обладают значительной степенью симметрии. Каждая половина мозга обслуживает половину тела и половину поля зрения.
Распределение более специализированных функций совершенно иное — оно глубоко асимметрично. Я указывал выше, что лингвистические способности определяются преимущественно левым полушарием. Есть основания полагать, что правая половина мозга более важна для восприятия мелодий; одним из свидетельств этого служит способность петь, отмеченная у больных с афазией, у которых повреждение локализуется в левом полушарии. Восприятие и анализ невербальных зрительных образов, таких, как перспективные рисунки, осуществляются преимущественно с помощью правого полушария, хотя и левое полушарие вносит свой вполне определенный вклад в эти функции. Асимметрия проявляется также в частичных выпадениях памяти, которые встречаются при одностороннем поражении височной доли. Удаление левой височной доли, нарушая способность запоминать словесный материал, не сказывается на способности запоминать пространственные взаимоотношения, лица, мелодии и абстрактные зрительные образы.
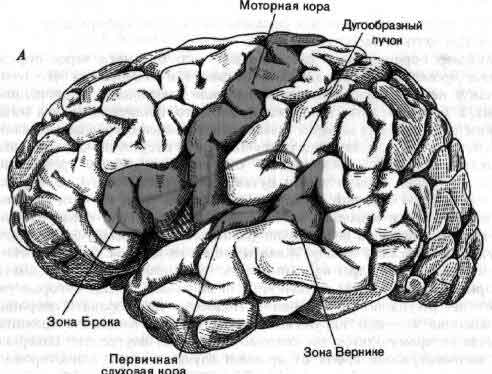
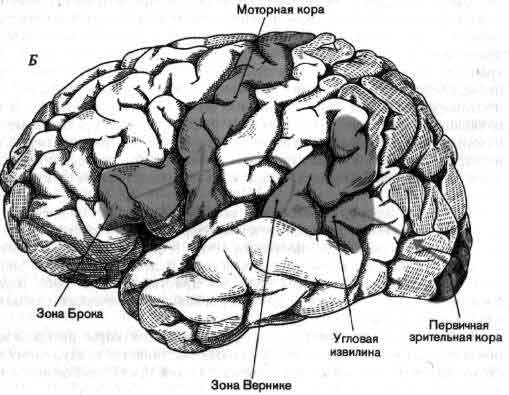
Пользование речью требует кооперации нескольких областей коры. Когда воспринимается слово (А), сигналы идут от уха в первичную слуховую кору, но данное слово не будет понято, если сигнал не подвергнется обработке в расположенной поблизости зоне Вернике. Если слово нужно произнести, некоторое его представление, по-видимому, передается из зоны Вернике в зону Брока посредством группы нервных волокон, называемой дугообразным пучком. В зоне Брока это слово порождает детальную программу артикуляции, поступающую затем в лицевую область моторной коры. Когда написанное слово читается (Б сигналы сначала регистрируются первичной зрительной корой. Затем они, по-видимому, направляются в угловую извилину, которая связывает зрительную форму данного слова с соответствующим слуховым аналогом в зоне Вернике. Произнесение слова затем осуществляется с помощью той же системы нейронов, о которой говорилось выше.
Узнавание лиц — это способность, за которую, по-видимому, ответственны области коры на внутренней стороне височных и затылочных долей обоих полушарий; здесь представлен их вид снизу. Поражение этих областей нарушает способность узнавать человека по лицу, но почти не вызывает других расстройств. Часто наблюдается некоторое ослабление зрения, но больной может читать, называть видимые предметы и даже соотносить портрет, сделанный в фас, с профилем того же человека. Больной может также узнавать людей по голосам. Единственная способность, которая утрачивается — это способность узнавать человека по лицу; расстройство бывает столь резко выраженным, что больной не узнает даже близких родственников.
В повседневной жизни эта латерализация функций редко обнаруживается, поскольку информация легко переходит из полушария в полушарие через несколько комиссур, включая мозолистое тело. Даже когда эти связи перерезаны, проявления доминирования полушарий в полной мере можно наблюдать только в лабораторных условиях, где можно точно гарантировать, что сенсорная информация поступает в данное время только в одно полушарие и что моторная реакция идет только от одного полушария. В таких условиях наблюдаются удивительные отклонения в поведении. Если какой-нибудь предмет помещают в левую руку больного или предъявляют только левой половине поля зрения, больной не может назвать предмет. Дело не в том, что он не узнает предмета — он безошибочно указывает на родственные объекты; причина кроется в том, что восприятие, осуществляемое правым полушарием, не ассоциируется с названием, которое «известно» только левому.
Определенные процессы памяти представляются связанными со структурами на внутренней поверхности височных долей, такими, как гиппокамп (окрашено). Установлено, что двухстороннее повреждение этих областей вызывает сильное и длительное расстройство памяти, характеризующееся неспособностью усваивать новую информацию. В случае поражений такого типа способность воспринимать, по-видимому, не снижается, но в значительной степени нарушается способность переводить новую информацию на хранение в долговременную память. Острые поражения этой области на одной височной доле иногда вызывают сходные, но менее стойкие расстройства памяти, которые отражают различную специализацию полушарий: характер информации, запоминание которой нарушается, зависит от того, какая сторона повреждена.
Однако специализацию отдельных полушарий не следует переоценивать. Правое полушарие все же имеет некоторые зачаточные лингвистические способности. Кроме того, без сомнения, имеется много задач, в которых оба полушария обычно действуют сообща. В одном тесте, проводимом после хирургического разобщения полушарий, предлагается воспроизвести простой узор из цветных блоков. В ряде случаев ошибки совершаются часто независимо от того, какой рукой выполняется задача-левой или правой, но ошибки эти имеют характерные отличия. По-видимому, ни одно из полушарий не в состоянии справиться с этой задачей самостоятельно и им приходится кооперироваться.
В мозгу человека обнаружена анатомическая асимметрия коры, которая может быть соотнесена с различной функциональной специализацией полушарий. Одно из проявлений асимметрии легко наблюдать на интактном мозге: сильвиева борозда, которая определяет верхнюю границу височной доли, на правой стороне мозга поднимается более круто. Еще более впечатляющая асимметрия обнаружена на planum temporale, которая формирует верхнюю поверхность височной доли и которую можно увидеть только тогда, когда сильвиева борозда расширена. Задняя часть planum temporale обычно намного больше на левой стороне. Увеличенная область является частью зоны Вернике, что позволяет предполагать связь этой асимметрии с доминированием левого полушария в отношении речи. Распределение указанных асимметрий зависит от доминирующей руки.
Одно из самых удивительных последних открытий — это установление того факта, что повреждения правого и левого полушарий мозга вызывают различные эмоциональные реакции. Поражения большинства областей левого полушария сопровождаются ощущением потери, которое можно ожидать при любой тяжелой травме. Больного угнетает его беспомощность и он часто бывает в подавленном настроении. При повреждении в большей части правого полушария больной иногда остается в неведении относительно своего состояния. Г. Гайнотти (G. Gainotti) из Католического университета в Риме тщательно собирал факты, касающиеся этих различий в эмоциональных реакциях.
Эмоции и «расположение духа» часто связывают со структурами лимбической системы, но в последние годы было обнаружено, что кора, и особенно кора правого полушария, также вносит сюда свой значительный вклад. Поражения правого полушария не только вызывают неадекватные эмоциональные реакции больного на его собственное состояние, но и нарушают способность узнавать эмоции собеседника. При левостороннем поражении больной иногда неспособен понять утверждение, но во многих случаях может все-таки оценить эмоциональную окраску, с которой оно высказано. Больной с повреждениями правого полушария обычно понимает смысл того, что сказано, но часто неспособен определить, как это сказано — с раздражением или с юмором. Хотя о явлениях доминирования полушарий большого мозга у человека известно более ста лет, сопоставимые асимметрии у других видов были обнаружены всего несколько лет назад. Пионером в этой области явился Ф. Ноттебом (F. Nottebohm) из Рокфеллеровского университета, который исследует нервные механизмы пения у певчих птиц. У большинства видов, изученных им к настоящему времени (но не у всех), левая половина мозга более важна для пения. Примеры доминантности у млекопитающих, помимо человека, также были описаны, хотя и намного менее детально. Как показал В. Дененберг (V. Denenberg) из Коннектикутского университета, при определенных условиях повреждения правой половины мозга у крыс меняют эмоциональное поведение животных. Доминирование коры левого полушария для некоторых слуховых задач открыл у одного вида обезьян Дж. Дьюсон III (J. Dewson III), который сейчас работает в Стэнфордском университете. М. Петер-сен (М. Petersen) и другие исследователи из Мичиганского и Рокфеллеровского университетов показали, что у японских макаков, которые используют необычайное разнообразие звуковых сигналов, при узнавании видоспепифических криков доминирует левое полушарие. Однако у ближайших родственников человека — человекообразных обезьян - до настоящего времени никаких определенных примеров функциональной асимметрии описано не было.
В течение многих лет среди неврологов господствовало мнение о том, что функциональная асимметрия мозга не коррелирует с анатомической асимметрией. Считалось, что если бы существовали сколько-нибудь существенные различия между полушариями, они давно были бы замечены хирургами или патологоанатомами. Около 10 лет назад мой коллега В. Левицкий (W. Levitsky) и я решили пересмотреть этот вопрос заново, исходя из более ранних наблюдений немецкого невропатолога Р. Пфайфера (R. Pfeifer). Мы исследовали planum temporale человеческого мозга в 100 случаях; эта область расположена на верхней поверхности височной доли и уходит внутрь сильвиевой борозды. Наше исследование касалось только макроанатомии, и мы не пользовались никакими сложными инструментами, ограничившись фотоаппаратом и линейкой; тем не менее было обнаружено недвусмысленное доказательство асимметрии. Как правило, длина и ориентация сильвиевоей борозды на левом и правом полушариях неодинаковы. И что более существенно - задняя часть planum temporale, которая входит в состав зоны Вернике, обычно больше на левой стороне. Эти различия не слишком малы, их легко заметить невооруженным глазом.
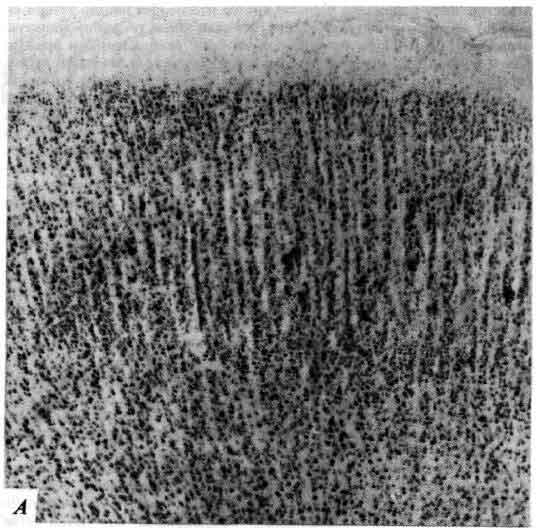
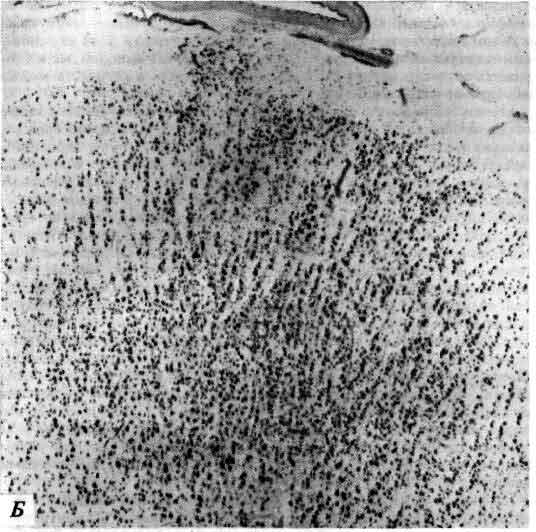
В речевой зоне у больного с врожденным дефектом способности к чтению была обнаружена аномальная клеточная организация. А. Срез нормальной коры на задней части planum temporale - области, которая составляет часть зоны Вернике. Можно выделить несколько слоев и отметить наличие характерных колонок. Б. Срез из той же области мозга больного с дизлексией. Одной из особенностей является присутствие тел нейронов в самом поверхностном слое (у верхнего края фотографии), где они в норме не встречаются. Кроме того, по всей ткани расположение клеток нарушено. Эта аномалия была обнаружена А. Галабурдой из Гарвардской медицинской школы и Т. Кемпером из Медицинской школы Бостонского университета.
Вслед за нами Дж. Вада (J. Wada) из Университета Британской Колумбии показала, что асимметрию planum temporale можно обнаружить и у человеческого плода. Таким образом, оказалось, что увеличение левой planum temporale не может быть реакцией на развитие лингвистических способностей в детстве. Наоборот, «лингвистическое превосходство» левого полушария, по-видимому, имеет анатомическую основу. Не так давно мой коллега А. Галабурда (A. Galaburda) установил, что левая planum temporale не только больше по размеру, но отличается и по клеточной организации. На planum temporale есть область с характерным расположением клеток, обозначаемая Tpt. Галабурда обнаружил, что протяженность этой области значительно больше в левом полушарии; у первого же мозга, который был исследован, эта область на левой стороне более чем в семь раз превосходила такую же область справа.
А. Галабурда и Т. Кемпер (Th. Kemper) из Медицинской школы Бостонского университета исследовали мозг пострадавшего от несчастного случая, у которого была стойкая дизлексия. Они обнаружили, что области Tpt в двух полушариях были примерно одинаковой величины. Кроме того, клеточная структура области Tpt слева была аномальна. В нормальной коре нейроны располагаются последовательными слоями, для каждого из которых характерна своя популяция клеток. У больного с дизлексией эти слои были нарушены, причем бросалось в глаза присутствие тел нейронов в самом поверхностном слое коры, где их обычно не бывает. Островки корковой ткани были также обнаружены в белом веществе мозга, которому они не принадлежат. Хотя на основании единичного случая нельзя делать определенных выводов, все же создается впечатление, что в случае лингвистических расстройств можно обнаружить структурные нарушения в речевых зонах.
Новое направление исследований асимметрии мозга было позднее открыто моей сотрудницей М. ЛеМэй (М. LeMay). Она разработала несколько методов обнаружения анатомической асимметрии у живых людей. Один из этих методов — церебральная артериография: в кровяное русло вводят рентгеноконтрастное вещество и исследуют его распределение в сосудах мозга. Артериографию часто используют для диагностики опухолей и других заболеваний мозга, и артериограммы, которые исследовала ЛеМэй, были получены с диагностической целью. Одна из внутричерепных артерий (средняя мозговая) идет вдоль сильвиевой борозды; ЛеМэй показала, что вид этой артерии на артериограмме отражает длину и ориентацию борозды. Оказалось, что у большинства людей правая средняя мозговая артерия идет круче и в конце концов поднимается выше, чем соответствующая артерия на левой стороне.
ЛеМэй обнаруживала асимметрию мозга также с помощью компьютерной аксиальной томографии (КАТ)-метода, при котором из набора рентгеновских проекций реконструируется картина мозга в поперечном сечении. На полученных изображениях видны характерные отклонения от билатеральной симметрии. У правшей правая лобная доля обычно шире, чем левая, но зато левая теменная и затылочная доли шире, чем правые. Внутренняя поверхность самого черепа сильнее вдавлена в правой лобной и левой затылочной областях в соответствии с этими выпуклостями.
ЛеМэй даже сообщила об обнаружении асимметрии на слепках ископаемых черепов неандертальцев и других гоминидов. На внутренней поверхности черепа есть гребень, соответствующий сильвиевой борозде; в тех случаях, когда этот гребень сохранился достаточно хорошо для того, чтобы отпечататься на слепках, ЛеМэй нашла те же проявления асимметрии, которые наблюдаются у современного человека; это заставляет предполагать, что доминирование полушарий возникло по крайней мере 30000 лет назад. ЛеМэй и я показали, что аналогичная асимметрия сильвиевых борозд существует и у крупных приматов, но не у низших обезьян. Об аналогичных открытиях сообщили Г. Йени-Комшиан и Д. Бенсон (G. Yeni-Komshian, D. Benson) из Медицинской школы Университета Джонса Гопкинса. Если удастся обнаружить функциональные корреляты этих анатомических отклонений, можно будет составить представление о доминировании полушарий у человекообразных обезьян.
Одно из самых банальных проявлений доминирования полушарий является в то же время и одним из самых поразительных: это феномен доминирования руки. У многих животных наблюдаются какие-то проявления этого свойства; например, если заставить обезьяну выполнять некоторую задачу только одной рукой, она всегда будет использовать одну и ту же руку. Однако в любой большой популяции обезьян левши и правши встречаются одинаково часто. В человеческой популяции левши составляют не более 9 процентов. Возможно, что такой значительный сдвиг в сторону доминирования правой руки служит выражением уникальной специализации человеческого мозга.
Генетика и наследование доминирования руки являются предметом живой полемики. Р. Коллинз (R. Kollins) из Джексоновской лаборатории в Бар-Харборе показал, что продолжительный инбридинг мышей с доминирующей правой лапой не приводит к увеличению числа таких особей в потомстве. У человека положение совершенно иное. М. Аннетт (М. Annett) из Ланчестерского политехнического института в Англии выдвинула теорию, согласно которой один аллель из генной пары благоприятствует развитию праворукости, однако комплементарного ему аллеля леворукости не существует. В отсутствие аллеля, благоприятствующего развитию праворукости, доминирующая рука определяется случайным образом.
Исследования, предпринятые ЛеМэй и ее сотрудниками, показали, что распределение асимметрий у левшей иное, чем у правшей. У правшей и, значит, у большинства людей правая сильвиева борозда расположена выше левой в 67% случаев, левая борозда выше в 8%, и обе борозды находятся на одинаковой высоте в 25% случаев. У 71% левшей сильвиевы борозды примерно симметричные; у остальных правая борозда чаще расположена выше (21% против 7). Асимметрии, наблюдаемые с помощью томографии, также по-разному распределяются у правшей и левшей. У части населения с доминирующей левой рукой асимметрия менее выражена. Эти данные находятся в качественном согласии с теорией Аннетт.
Если такие узко ограниченные функции, как узнавание лиц, обеспечиваются специфическими нейронными сетями мозга, то кажется вероятным, что многие другие функции представлены аналогичным образом. Так, например, одна из главных целей воспитания детей — выучить их набору высоко дифференцированных реакций на эмоциональные стимулы, такие, как гнев и страх. Ребенок должен также научиться подходящим реакциям на стимулы из своей внутренней среды, такие, как голод или ощущение наполнения мочевого пузыря и кишечника. Большинство детей научается этим типам поведения точно так же, как они научаются языку, на основании чего можно предположить, что и тут существуют процессоры специального назначения. К настоящему времени о таких нейронных системах мало что известно. На самом деле, даже если картирование специализированных зон будет продолжаться, на повестку дня должна быть поставлена следующая главная задача-задача описания их внутренних операций.
С. КИТИ
Заболевания человеческого мозга
Они могут быть следствием наследственного нарушения обмена, сосудистого заболевания, инфекции, опухоли, травмы. При исследовании психических заболеваний важны отношения между генетическими факторами и факторами внешней среды
В такой сложной структуре, как человеческий мозг, может возникнуть множество нарушений. Удивительно то, что у большинства людей мозг работает эффективно и непрерывно дольше шестидесяти лет. Это говорит о пластичности, избыточности и самовосстанавливающейся природе его механизмов. Но дело в том, что в мозгу иногда нарушается его структурная архитектоника или электрические и химические процессы. Более ста лет назад патологи уже умели обнаруживать заболевания, связанные с повреждением крупных анатомических структур мозга и возникающие в результате кровоизлияния, компрессии, смещения, воспаления, дегенерации и атрофии. Микроскоп и избирательное окрашивание дали возможность увидеть, как морфологическое повреждение вызывает голодание, дегенерацию и гибель нейронов.
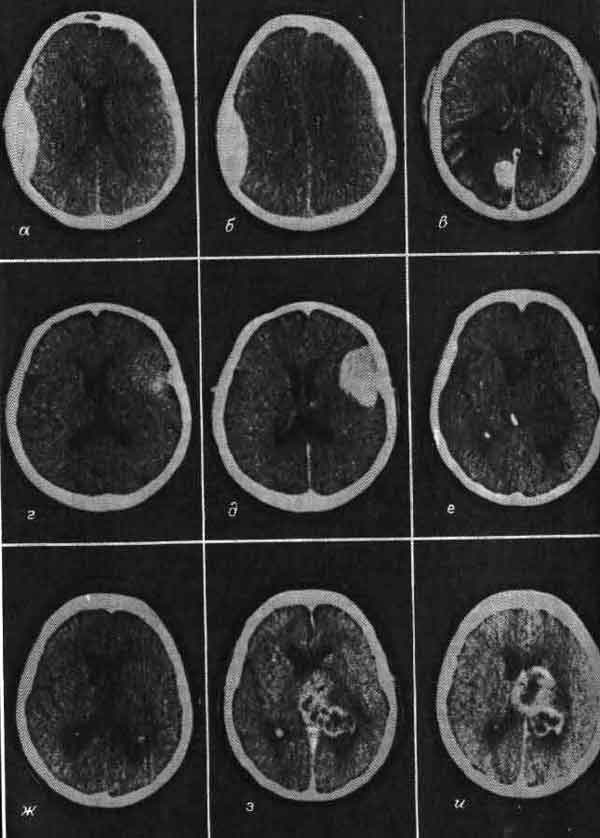
Компьютерная аксиальная томография (КАТ) использует рентгенограммы, сделанные под разными углами, с целью получения картины головного мозга в поперечном разрезе. Инъекция йодистого препарата в венозную систему усиливает контраст. На кадрах а и б для выявления плотного сгустка крови в пространстве между мозгом и черепом иод был не нужен. Кровоизлияние произошло от удара тупым орудием по этой части черепа. По диагонали от сгустка видно кровоизлияние на поверхности мозга или непосредственно под поверхностью; это кровоизлияние вызвано повреждением мозга по механизму «противоудара» (contre-coup). Желудочки (в центре) сужены из-за набухания ткани мозга. На кадре в препарат иода выявил опухоль (в центре внизу) у больного с метастазами рака. Справа от узла опухоли — нормальная вена, которая отчетливо видна благодаря контрастному веществу — введенному в кровь иоду. Желудочки смещены набуханием тканей вокруг опухоли. На кадре г менингиома (доброкачественная опухоль) без иода едва видна. Внутри опухоли лежит небольшой островок кальция. Гиперостоз, разрастание кости вблизи опухоли, характерно для менингиомы. На кадре д под действием иода та же самая опухоль стала видна гораздо лучше. Тонкая белая линия, пересекающая опухоль, проведена сканирующим аппаратом с целью измерения. На кадрах е и ж злокачественная опухоль (в середине) без иода едва видна, но на кадрах з и и, полученных с применением иода, она ясно выделяется в виде пятнистого участка. Желудочки смещены, и границы их зазубрены. На кадре е кальцинированный эпифиз (в середине) тоже слегка смещен. Усиленные иодом белые кольцевые зоны характерны для злокачественной опухоли. Девять кадров КАТ любезно предоставлены Ф. Ходжесом III (F. Hodges III) из Медицинской школы Университета Джонса Гопкинса.
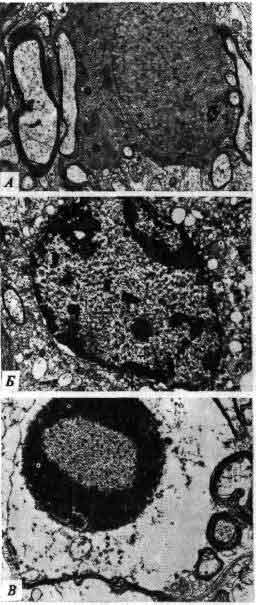
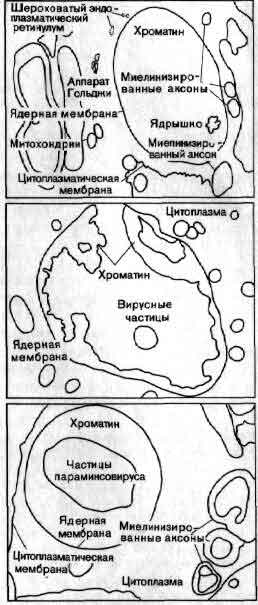
Глиальная клетка, которая вырабатывает и обновляет жироподобный слой миелина, образующий оболочку аксонов в центральной нервной системе, может играть роль в расстройствах функции головного мозга. А. Электронная микрофотография нормальной глиальной клетки; это сравнительно темная клетка с плотно упакованными внутриклеточными органеллами, в том числе митохондриями, шероховатым эндоплазматическим ретикулумом и четко очерченным аппаратом Гольджи (рядом дана карта органелл). В этой нормальной клетке хроматин - носитель генетического материала - равномерно распределен по всему ядру. На периферии клетки лежат части нескольких миелинизированных аксонов; миелин является прямым продолжением специализированной цитоплазматической мембраны глиальной клетки. Б. Глиальная клетка больного хроническим лимфатическим лейкозом. Органеллы клетки сильно разрушены. Клетка, находящаяся в ткани, взятой при аутопсии, отличается увеличенным и смещенным ядром, конденсированным хроматином и наличием многочисленных вирусных частиц, уничтожавших клетку. В результате миелин аксонов не обновлялся. Постепенная демиелинизация основных нервных путей привела к развитию симптомов болезни. Приблизительно за четыре месяца до смерти больной жаловался на ухудшение зрения (что закончилось слепотой в левом поле зрения), на то, что он не различает лиц и не может читать. В конце наступила полная слепота, некоторая спутанность сознания с двусторонним парезом. В. Глиальная клетка в ткани, взятой при аутопсии у больного подострым склерозирующим панэнцефалитом. Отдельные органеллы в клетке неразличимы, хотя близлежащие миелинизированные аксоны еще видны. Хроматин образовал комки и сдвинут частицами, характерными для парамиксовируса. Электронные микрофотографии получены Дж. Волынским (J. Wolinsky) из Медицинской школы Университета Джонса Гопкинса.
В течение многих лет препятствием для изучения заболеваний мозга служило отсутствие способов исследования живого мозга. То немногое, что было известно о таких болезнях, явилось результатом посмертных исследований. Открытие рентгеновских лучей в конце XIX в. позволило ученым заглянуть в живой мозг. Теперь есть возможность обнаруживать грубые структурные разрушения, касающиеся желудочков головного мозга, при помощи пневмоэнцефалографии, т.е. рентгенологической методики, при которой жидкость, окружающая мозг и наполняющая его желудочки, замещается воздухом, что позволяет получить картину их формы. При другом методе — церебральной ангиографии - в кровь вводят рентгеноконтрастное вещество и это дает возможность видеть на рентгенограмме патологическое смещение кровеносных сосудов мозга. Обычное рентгенологическое исследование при всей своей ценности страдает одним крупным недостатком: на проявленной пленке рентгенографические проекции патологически измененных областей могут наложиться на проекции нормальных структур, из-за чего трудно или даже невозможно отличить их друг от друга. Это особенно дает себя знать при одинаковой проницаемости соседних структур для рентгеновских лучей, как, например, в случае опухоли, окруженной нормальной тканью такой же плотности.
Создание метода компьютерной аксиальной томографии (КATI позволило преодолеть этот недостаток. КАТ-сканирование - это особая методика, при которой сопоставляются многочисленные рентгеновские снимки, сделанные под разными углами, с целью получения изображения внутренней структуры головного мозга в поперечном сечении. Такое сканирование выявляет увеличение или атрофию нормальных структур и любые патологические образования.
В середине нашего столетия возникли электрические методики, ставшие важными инструментами исследования мозга. Сообщения, получаемые мозгом от органов чувств, директивы, которые он им посылает, и сообщения между миллиардами нейронов в самом мозгу — все они передаются с помощью электрических сигналов. Электрические поля у поверхности мозга улавливаются и усиливаются электроэнцефалографом. Это позволяет определить специфическую локализацию источников нарушения электрической активности.
В течение последних двух десятилетий изучение функции мозга распространилось на химические процессы. Как мозг использует энергию, можно исследовать, измеряя кровоток и использование кислорода из глюкозы. Благодаря недавним работам Н. Лассена (N. Lassen) из Клиники Биспебьерга в Копенгагене и Д. Ингвара (D. Ingvar) из Копенгагенского университета появилась возможность наблюдать на рентгеновском экране, как при таких специфических видах умственной деятельности, как чтение вслух или чтение про себя, быстро меняется кровообращение в разных участках мозга. Л. Соколов (L. Sokoloff) с сотрудниками из Национального института охраны психического здоровья разработали приемы для количественной оценки метаболизма глюкозы в любой точке мозга. Поскольку функциональная активность тесно связана с интенсивностью кровотока и использованием глюкозы, такие методики служат средством для картирования живого мозга в отношении функционирования его компонентов.
На уровне нейрона мозговые расстройства могут возникнуть из-за аномальных химических процессов, действующих в синапсах между нейронами. Нарушения синтеза, высвобождения или инактивации определенного химического медиатора или нарушения чувствительности пост-синаптических рецепторов медиатора могут привести к дисфункции синапса. Такая дисфункция необязательно сопровождается морфологическими изменениями на макро- или микроуровне. Разработанные недавно гистофлуоресцентные и иммунофлуоресцентные методики, т. е. по существу методы окрашивания на специфические медиаторы или ферменты, позволяют обнаруживать и измерять действие медиатора на отдельные нейроны. Новые химические методики с применением радиоактивных изотопов дают возможность определять число и чувствительность постсинаптических рецепторов, а мощными аналитическими приемами можно исследовать жидкость в головном мозгу, спинномозговую жидкость, кровь и мочу на присутствие ничтожных следов медиаторов и их метаболитов.
Для успешного протекания таких сложных психических процессов, как восприятие, мышление, суждение и эмоции, необходимы идеально настроенные и бесперебойно функционирующие синапсы. Поскольку при психических заболеваниях такие процессы часто нарушаются, расширение представлений об этих процессах должно помочь раскрыть тайны нарушений психики. Лишь недавно эти новые способы изучения химизма синапса были применены в случаях психических заболеваний — шизофрении и маниакально-депрессивного психоза. Мне представляется вполне возможным, что эти орудия исследования будут для психиатрии тем, чем старые методики были для невропатологии.
Патологические процессы вызываются в мозгу большим разнообразием близких и отдаленных причин, которые часто делят на генетические факторы и факторы внешней среды. Поскольку всякое свойство живого организма в конечном счете определяется сложным взаимодействием генетических влияний и влияний внешней среды, попытка расчленить их может показаться бесплодной. Но все же есть возможность их дифференцировать, если проследить, каков вклад каждого из этих влияний в изменчивость данного конкретного признака. Например, способность к речи требует высокоразвитого механизма в головном мозгу, механизма, который совершенно очевидно зависит от генетических процессов. Но тот конкретный язык, на котором говорит человек, не обусловлен генетически, а почти полностью определяется фактором среды — принадлежностью к той или иной культуре и т.п. С другой стороны, существуют генетические дефекты головного мозга, которые выявляются только в условиях специфического питания. Если эти условия повсеместны, то отличие аномального индивидуума от нормального будет полностью обусловлено генетическим фактором; поэтому такое нарушение относят к генетическим болезням. Разумеется, большинство человеческих свойств не поддается классификации с такой легкостью. По-видимому, они оказываются где-то между двумя крайними точками.
Гены определяют последовательности аминокислот, образующих белки. Именно эти белковые молекулы, синтезируемые в особых местах и в особое время, служат структурными компонентами и ферментативными катализаторами, которые обусловливают развитие и работу мозга. Известно, что многие нарушения в центральной нервной системе, в особенности те, которые проявляются умственной отсталостью, обусловлены генетически. Например, фенилкетонурия и галактоземия вызываются генетически обусловленной недостаточностью определенных ферментов. При той и другой болезни из-за дефицита определенного фермента становятся токсичными некоторые компоненты обычной пищи. У ребенка с фенилкетонурией отсутствует фермент - фенилаланингидроксилаза, - который обеспечивает дальнейший метаболизм фенилаланина. В результате эта незаменимая аминокислота в избытке накапливается в крови и тканях, что нарушает развитие и работу мозга.
При галактоземии новорожденный ребенок кажется нормальным, но через несколько дней или недель кормления молоком у него появляются анорексия и рвота, он начинает отставать в весе и погибает от истощения. Дети, выжившие без лечения, остаются карликами и умственно отсталыми. Галактоземия вызывается отсутствием фермента, необходимого для дальнейшего метаболизма сахара галактозы, который поэтому в избытке накапливается в организме. Вредные последствия этих болезней можно устранить внешним воздействием, а именно исключением из рациона ребенка (по крайней мере в раннем детстве, в период интенсивного роста и развития головного мозга) того вещества, метаболизм которого в организме нарушен (фенилаланина или галактозы).
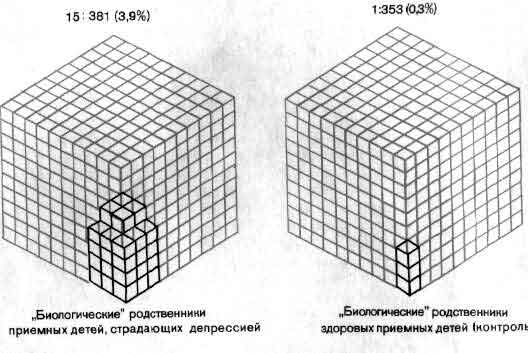
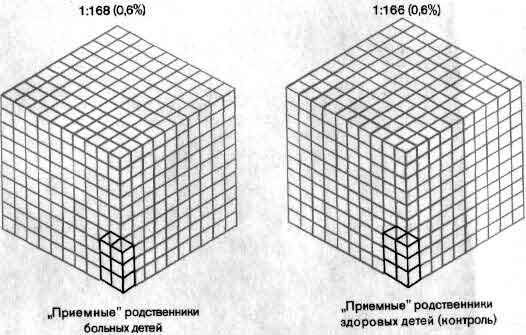
Больший процент самоубийств установлен среди биологических родственников тех приемных детей, которые страдали депрессией, по сравнению с числом самоубийств среди их приемных родственников и родственников приемных детей контрольной группы, не страдавших психическим заболеванием, что заставляет предполагать в основе этих самоубийств генетический фактор. На каждом графике показано число родственников-самоубийц по отношению к общему числу родственников. Данные взяты из работы, проведенной автором и Д. Розенталем (D. Rosenthal) из Национального института охраны психического здоровья. Ф. Шульзингером (F. Schulsinger) из Копенгагенского университета и П. Уэндером (P. Wender) из Университета Юты.
Некоторые генетически обусловленные болезни сцеплены с полом. Синдром Леша-Нихана, характеризующийся бесцельными и неуправляемыми движениями, умственной отсталостью и психотическим поведением («самоповреждение»), возникает из-за отсутствия определенного фермента, что в свою очередь обусловлено отсутствием или дефектом гена в Х-хромосоме. Этот синдром встречается только у мужчин, у которых всего одна Х-хромосома. У женщин, у которых имеется две Х-хромосомы, отсутствие или дефект гена в одной из них компенсируется нормальным геном другой.
Мозговые расстройства возникают не только от недостатка генетического материала, но и от его избытка. Наличие добавочной Х- или Y-хромосомы сопровождается синдромами незначительных нарушение интеллекта и личности. При синдроме Дауна, который наблюдается у одного из каждых 700 новорожденных, имеется лишняя хромосома 21. Такие дети отстают в развитии — как физическом, так и умственном.
Генетические болезни не всегда обнаруживаются при рождении. Например, симптомы хореи Гентингтона, а именно неконтролируемые движения и снижение интеллекта, появляются впервые в возрасте 30—50 лет. Эта болезнь, которая наследуется как доминантный генетический признак, связана с резко выраженной атрофией мозолистого тела и дегенерацией нейронов в хвостатом ядре и других глубоких ядрах, а также в лобной коре больших полушарий.
В 1968 г. Л. Полинг (L. Pauling) предположил, что существуют генетически обусловленные индивидуальные различия в потребности в витаминах. Он полагал, что эти различия связаны с заболеваниями центральной нервной системы, включая головной мозг. Примерами, свидетельствующими в пользу этой гипотезы, служат некоторые редкие неврологические синдромы у детей, при которых генетически обусловленная неспособность усваивать или перерабатывать определенный витамин создает тяжелую витаминную недостаточность, которая неблагоприятно сказывается на функции центральной нервной системы. Возникающие патологические синдромы успешно излечиваются большими дозами соответствующего витамина. Полинг предположил подобный механизм в основе шизофрении, однако повышенная потребность в том или ином витамине при шизофрении не доказана.
Психозы, в том числе шизофрения и аффективные расстройства, встречаются значительно чаще упомянутых выше заболеваний. В этих психозах главную роль, по-видимому, играют генетические факторы, хотя их биологические последствия еще не определены, в отличие от того, как это сделано для многих других заболеваний мозга. С тех самых пор, как около 100 лет назад были впервые описаны шизофрения и маниакально-депрессивный психоз, известно, что они носят характер семейных болезней. Около 10% родственников больного тоже страдают этим заболеванием. Поэтому такие болезни часто считали наследственными.
Однако членов одной семьи объединяют не только генетические влияния, но и общие влияния среды, и поэтому семейный характер заболевания еще мало говорит о его этиологии. Пеллагра - болезнь, обусловленная витаминной недостаточностью, которая в первые десятилетия нашего века была причиной 10% психических заболеваний в США, тоже поражает целые семьи, и поэтому одно время считалась наследственной. В 1915 г. Дж. Голдбергер (J. Goldberger) из Службы здравоохранения США показал, что главной причиной пеллагры является острый недостаток в пище ниацина - витамина группы В. Поскольку члены одной семьи обычно едят одинаковую пищу, болезнь носит семейный характер. При помощи соответствующего питания пеллагру удалось почти полностью искоренить.
Доказательство генетической этиологии шизофрении и маниакально-депрессивного психоза получено при исследовании частоты данных заболеваний у однояйцовых и разнояйцевых близнецов (у первых все гены одинаковые, а у вторых одинаковы около половины генов). У однояйцовых обе болезни дают большой процент совпадения (конкордантность около 50%), тогда как у разнояйцевых он такой же, как вообще у потомков (около 10%). Работы последних лет сосредоточены на изучении приемных детей, генетическую конструкцию которых можно выяснить обследованием кровных («биологических») родственников, а влияние среды — обследованием их «приемных» родственников.
Такие исследования показали, что и шизофрения и маниакально-депрессивный психоз чаще встречались в биологических, а не в приемных семьях. Частота психических нарушений среди приемных, родственников больного ребенка оказалась такой же, как в общей популяции. Этот факт наряду с более высокой конкордантностью по этим заболеваниям у однояйцовых близнецов свидетельствует о роли генетических факторов в этиологии шизофрении и маниакально-депрессивного психоза, хотя и не исключает возможной роли факторов среды. Поскольку выражение гена осуществляется только через биохимические механизмы, значение генетических факторов в этиологии психических болезней является неопровержимым доказательством роли биохимических субстратов, хотя эти субстраты точно не идентифицированы.
Изучение биологических и приемных семей выявило важную роль генетического фактора при хроническом алкоголизме, при котором фактор среды, а именно алкоголь, играет очевидную роль, а также при самоубийствах, при которых несомненно играют роль разнообразные внешние влияния. Генетическими факторами таких заболеваний может объясняться то обстоятельство, что не каждый, кому случается пить спиртные напитки, становится алкоголиком, и что только немногие из людей, попавших во внешне безвыходные положения, кончают с собой.
Поскольку все негенетические этиологические факторы должны представлять собой факторы внешней среды, эта последняя категория охватывает широкий диапазон влияний, различных по качеству, силе и тем периодам жизни индивидуума, когда они проявляются. Такие факторы впервые начинают действовать в период внутриутробного развития. В сложных превращениях оплодотворенного яйца в новорожденного ребенка действуют химические и физические процессы, благодаря которым реализуется генетическая программа. Все виды недостаточности и нарушений внешней среды могут помешать этой реализации и нарушить нормальное развитие центральной нервной системы.
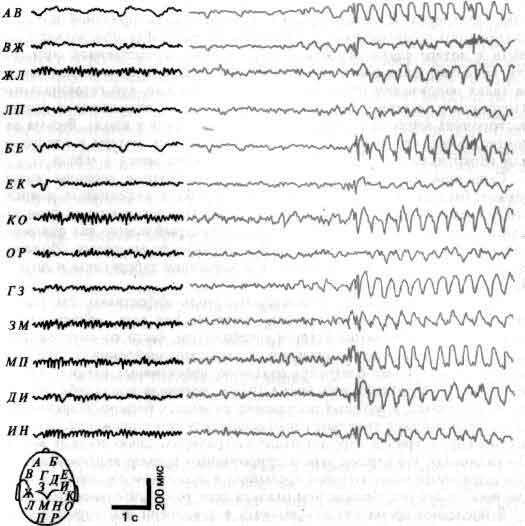
Электроэнцефалограмма больного эпилепсией (цветная): видна нормальная электрическая активность головного мозга до припадка, начало которого приблизительно соответствует средней точке цветных кривых. Их первая половина схожа с электроэнцефалограммой человека, не страдающего эпилепсией (черная). Каждая кривая является выражением того, как меняется во времени потенциал между двумя участками головы (обозначенными двумя буквами, которые соответствуют участкам на схеме). Под кривыми дана калибровка, на которой горизонтальная линия соответствует 1 секунде, а вертикальная - 200 микровольтам. Кривые зарегистрированы Р. Янгом (R. Young) из Гарвардской медицинской школы.
Многие формы церебрального паралича и умственной отсталости возникают как результат нарушений факторов среды, окружающей плод, или процесса родов. Если в первые три месяца беременности плоду передается от матери краснуха, это может привести к умственной отсталости и, возможно, к детскому аутизму. Если в мозг плода проникает цитомегаловирус, который широко распространен у взрослых и обычно безвреден, то у плода это может привести к глухоте и снижению интеллекта. Ядерная желтуха - желтуха плода, причиной которой является его несовместимость с матерью по резус-фактору, может привести к потере слуха, церебральному параличу и умственной отсталости, хотя благодаря диагностическим и профилактическим мерам частота таких осложнений резко снизилась. Установлено, что гормональные нарушения у матери и воздействие некоторых медикаментов и других посторонних веществ вызывают аномалии развития у плода. Весьма вероятно также, что недоедание или приверженность матери к алкоголю или наркотикам нарушает нормальное развитие мозга у плода.
Серьезной причиной заболеваний мозга в разные периоды жизни служат бактериальные инфекции, хотя борьба с инфекциями посредством антибиотиков позволила почти полностью искоренить многие тяжелые болезни, например общий паралич и острый психоз как осложнения сифилиса, а также менингит, или воспаление мозговых оболочек (часто со смертельным исходом) как последствие туберкулеза и других бактериальных инфекций.
Вирусные инфекции не излечиваются столь эффективно, как бактериальные, и иногда такие вирусные болезни, как корь, свинка, грипп и герпес, сопровождаются острым энцефалитом, часто со смертельным исходом. Полиомиелит - вирусное поражение мотонейронов - почти полностью ликвидирован благодаря созданию эффективных вакцин. Эпидемия гриппа, охвативная весь мир в 1918 г., оставила после себя несметное число людей, у которых постепенно развилась болезнь Паркинсона, характеризующаяся тяжелым нарушением регуляции движений. Данный штамм вируса гриппа «предпочитал» экстрапирамидную систему мозга. По-видимому, это первый зарегистрированный пример вирусной инфекции нервной системы, которая пребывала в латентном состоянии в течение нескольких лет, прежде чем оказала свое разрушительное действие.
В последнее время удалось выявить в этиологии некоторых других болезней латентные или медленно развивающиеся вирусные инфекции нервной системы. Примером служит куру - медленно развивающееся неврологическое заболевание, распространенное только среди небольшого племени в Новой Гвинее. Эта болезнь носит семейный характер, и потому ей приписывали генетическую природу, пока Д. Гайдушек (D. Gajdusek) не установил его вирусную этиологию.
Болезнь Крейцфельдта-Якоба - редко встречающаяся деменция (слабоумие) в среднем возрасте - еще одно медленно прогрессирующее вирусное заболевание. Болезнь Альцгеймера - более распространенный тип старческого слабоумия - сходна с болезнью Крейцфельдта-Якоба по многим клиническим проявлениям. Несколько лабораторий занимаются проверкой предположения о вирусной этиологии болезни Альцгеймера. Вирус герпеса и цитомегаловирус служат примерами еще двух вирусов, которые могут годами находиться в организме в латентном состоянии, а затем проявиться неврологическими или психическими симптомами. Есть данные о вирусной природе некоторых форм шизофрении.
История психиатрии и невропатологии знает много случаев заболеваний мозга, вызванных разного рода токсинами. Сумасшедший Шляпник был не просто плодом воображения Льюиса Кэрролла, а введенной в повествование жертвой профессионального заболевания, распространенного в XVIII и XIX веках. Шляпники, подвергаясь ежедневно действию ртути, применяемой при изготовлении войлока, нередко страдали токсическим психозом. Известно, что и другие тяжелые металлы разрушают нервную систему. Марганец вызывает одну из форм паркинсонизма, а свинец разрушает периферические нервы и головной мозг. Свинцовое отравление у детей (главным образом, от заглатывания изготовленной на свинце краски) может привести к нарушению поведения и утрате способности к обучению.
В средние века хлеб из ржаной муки, зараженной спорыньей, вызывал «эпидемии сумасшествия». Как теперь известно, это паразитическое растение содержит несколько алкалоидов, которые оказывают огромное влияние на нервную систему. Синтетическим производным одного из них является сильно действующий галлюциноген LSD.
Кроме экзогенных ядов и лекарственных средств токсичными для мозга могут быть эндогенные вещества, которые вырабатываются в самом организме при тех или иных заболеваниях. Нелеченный диабет и тяжелая или смертельная почечная или печеночная недостаточность нередко сопровождаются появлением в организме токсичных веществ, которые вызывают спутанность сознания, бред и, в конце концов, кому. Порфирия представляет собой редкое расстройство обмена генетического происхождения, при котором образование порфирина нарушает умственную деятельность. Сумасшествие короля Георга III объясняют порфирией.
Хотя процессы иммунитета служат для защиты организма от инфекции, иммунная реакция иногда искажается, что приводит к различным расстройствам, в которые иногда вовлекается мозг. Самыми распространенными заболеваниями такого рода являются астма и сенная лихорадка. Такие болезни, как ревматический артрит, красная волчанка и другие коллагенозы также связаны с патологической иммунной реакцией, направленной против собственных нормальных тканей. Окончательно установлено, что такой аутоиммунный процесс лежит в основе myasthenia gravis - тяжелого нервно-мышечного расстройства, которое характеризуется приступами утомления и слабости мышц. Болезнь поражает нервно-мышечные соединения, где выделяется медиатор ацетилхолин, который действует на рецепторы мышечных клеток, запуская сокращение. Последние данные показывают, что при этой болезни аутоиммунный процесс нарушает функцию рецепторов и снижает эффективность нервно-мышечного соединения. Возможно, что причиной рассеянного склероза тоже является аутоиммунный процесс. Данные в пользу этого получены в лабораторном эксперименте на модели аутоиммунного заболевания, названного экспериментальным аллергическим энцефаломиелитом, который напоминает рассеянный склероз.
Наиболее распространенные болезни мозга связаны с недостаточным кровоснабжением. Потребность головного мозга в энергии — одна из самых высоких в организме. Мозгу требуется одна пятая всего объема крови, выбрасываемой сердцем, и одна пятая кислорода, потребляемого организмом в покое. Хотя известны и врожденные аномалии сосудов головного мозга, чаще всего встречается их атеросклеротическое поражение. Атеросклероз поражает не только сосуды мозга, но и сердца, почек и конечностей. Атеросклероз приводит к сужению просвета, а иногда к тромбозу сосуда, что создает постепенное или внезапное сокращение кровоснабжения какой-либо области мозга, нарушая ее функцию. Возможен также разрыв пораженного сосуда и кровоизлияние в ткань мозга. Симптомы тромбоза или кровоизлияния бывают весьма разнообразными в зависимости от того, какие области мозга поражены и какие процессы затронуты. Некоторые симптомы кровоизлияния являются результатом повышения внутричерепного давления, которое нарушает кровоток, смещает отделы мозга и искажает его архитектонику. Такое же действие могут оказывать опухоли и травмы головы.
Эпилепсия, характеризующаяся одновременной ритмической импульсацией большого числа нейронов, представляет собой нервно-психическое заболевание, этиология которого остается пока неясной. Один из видов эпилепсии возникает в результате формирования на поверхности мозга рубца, как следствия травмы или инфекции. Рубцовая ткань служит очагом патологической электрической активности, которая распространяется по обширной области соседних нормальных нейронов. Однако в большинстве случаев эпилепсии никаких рубцов или других повреждений не обнаруживается. Для этих случаев накапливается все больше данных о химической природе дефекта, который касается одного или нескольких медиаторов, в частности ГАМК, - главного тормозного медиатора в центральной нервной системе.
Что касается наиболее часто встречающихся психозов — шизофрении и маниакально-депрессивного психоза, - то их этиология еще не выяснена. В тяжелой классической форме они могут быть сходны с органическими поражениями мозга. Отдельные симптомы или весь синдром психического заболевания иногда обнаруживается на ранних стадиях хорошо известной нервной болезни - хореи Гентингтона.
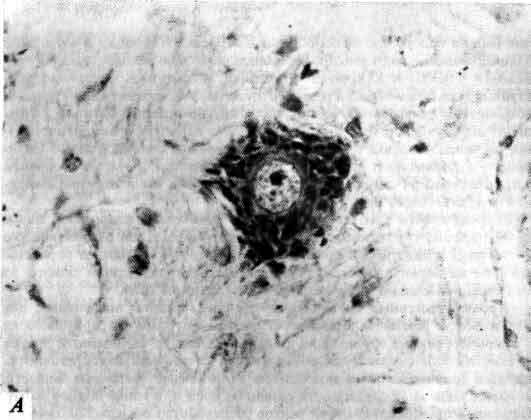
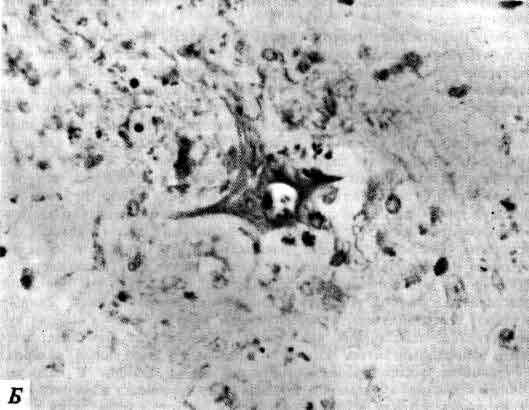
Полиомиелит - вирусная инфекция мотонейронов - почти ликвидирован благодаря созданию специфических вакцин. А. На электронной микрофотографии показана нормальная клетка из переднего рога спинного мозга макака-резуса; для нее характерны крупные тельца Ниссля в цитоплазме, центральное положение ядра и диффузно рассеянный в нем хроматин. Б. Необратимо поврежденная клетка из спинного мозга обезьяны на поздней стадии полиомиелита. Микрофотографии получены Д. Бодиеном (D. Bodian) из Медицинской школы Джонса Гопкинса.
О химической природе психических болезней яснее всего говорят упомянутые мною исследования, которые выявили значение генетических факторов в этиологии психозов. Биохимические процессы, лежащие в основе выражения соответствующих генов, точно не установлены, хотя имеются определенные сведения о том, что они протекают на уровне некоторых синапсов головного мозга. Фармакологические препараты,действующие на мозг, способны смягчить симптомы депрессии и шизофрении, но могут также и вызвать их. Такие препараты оказывают специфическое действие на синаптические процессы. Препараты, которые удаляют из синапсов определенные медиаторы (дофамин, норадреналин и серотонин), вызывают депрессию, тогда как все препараты, применяемые для лечения клинической депрессии, обычно повышают содержание этих медиаторов или усиливают их действие. Гипотеза, согласно которой некоторые формы депрессии возникают вследствие недостатка этих медиаторов или других, взаимодействующих с ними, представляется вероятной, и в ее пользу говорят исследования метаболитов в моче и спинномозговой жидкости.
Кроме того, известно, что все медикаменты, снимающие симптомы шизофрении, ослабляют активность дофаминовых синапсов; тогда как фенамин, который вызывает токсический психоз, очень близкий к шизофрении, повышает уровень дофамина в этих синапсах. Прямых доказательств избыточного содержания дофамина в мозгу больных шизофренией у нас нет; вполне возможно, что существуют другие факторы, дающие такой же эффект, как избыток дофамина; таким фактором могла бы быть чрезмерная чувствительность дофаминовых рецепторов или недостаточная активность другого медиатора, который в норме противодействует дофамину. Все это необходимо еще исследовать.
В этиологии психических заболеваний важную роль безусловно играют психологические и социальные факторы. Нет сомнения в том, что такие влияния изменяют проявления, тяжесть, длительность и течение этих болезней и во многих случаях вызывают их. Выдвинуто несколько вероятных гипотез относительно возможного взаимодействия между факторами среды и биологическим предрасположением к психической болезни. К внешним влияниям, которые играют некоторую этиологическую роль в психических заболеваниях, относятся, помимо психологических факторов, физиологические сдвиги в предродовом периоде и при родах, инфекции и некоторые токсины. Такие влияния имеют разное социальное распределение. Они чаще наблюдаются у представителей бедных слоев общества в связи со скученностью, плохими гигиеническими условиями и недостаточной медицинской помощью. Известно, что среди бедняков, живущих в больших городах, шизофрения встречается вдвое чаще, чем в общей популяции.
Успешное лечение или предупреждение того или иного заболевания мозга во многом определяется тем, что о нем известно. Симптомы опухоли, или атеросклероза кровеносных сосудов, можно иногда смягчить паллиативными воздействиями, но более специфическое лечение или профилактика болезни требует понимания механизмов, лежащих в ее основе. Болезнь Паркинсона, проявления которой делают инвалидами около 50000 американцев в год, может служить хорошим примером того, как знание нейробиологических процессов, вызывающих определенную болезнь, дает возможность создать успешный метод ее лечения. В прежние времена это прогрессирующее хроническое заболевание означало для его жертв смерть или тяжелую инвалидность. Болезнь начинается с непроизвольной дрожи в руках. Атаксичесхий тремор распространяется на все конечности, шею и челюсть. Спина деревенеет, мышцы становятся ригидными, походка странной, шаркающей. На терминальных стадиях больной прикован к постели.
В работе А. Карлсона (A. Carlson) из Гетеборгского университета и О. Хорникевича (О. Hornykiewicz) из Венского университета обнаружено, что причина этой болезни лежит в прогрессирующем разрушении нейронных связей, для которых характерен медиатор дофамин. Следствием этого, как было установлено, является нехватка дофамина в синапсах проводящих путей полосатого тела, - структуры, которая лежит близ середины мозга и модулирует движение.
Эти сведения позволили исследователям разработать эффективное лечение. Было известно, что, хотя сам дофамин не проникает через гематоэнцефалический барьер, для его химического предшественника L-ДОФА этот барьер проходим. Препарат L-ДОФА после приема внутрь (через рот) попадает из кишечника в кровоток и достигает головного мозга, где захватывается и преобразуется в дофамин. При клинических испытаниях больших доз L-ДОФА Дж. Котциас (G. Cotzias) из Национальной лаборатории в Брукхевене установил выраженное смягчение симптомов болезни. Терапевтические дозы L-ДОФА довольно высоки (до 8000 мг в день), что вызывает возникновение многочисленных побочных эффектов. Недавно разработаны модифицированные формы лечения, которые не дают серьезных осложнений.
Эффективная терапия болезни Паркинсона - одна из самых крупных побед медицины, основанных на фундаментальных исследованиях в области биохимии и нейробиологии. Но даже до того, как становится понятным механизм заболевания мозга, иногда случайно удается открыть способ его эффективного лечения. Так было с антидепрессантами и антипсихотическими лекарственными препаратами, которыми ознаменовалось развитие психофармакологии и которые произвели революцию в психиатрии. Понимание того, как эти препараты действуют на химические синапсы, облегчает создание более эффективных лекарственных средств с меньшим побочным действием. Эпилепсия, причины и механизмы которой еще неизвестны, успешно лечится дилантином — препаратом, который был открыт в результате систематического поиска с применением электрофизиологических методик.
По мере того как в процессе тщательных исследований и накопления фундаментальных знаний растет понимание конкретных болезней, создаются новые способы их лечения и в конце концов методы их предупреждения. Так произошло с такими инфекционными заболеваниями мозга, как полиомиелит, менингит и некоторые виды энцефалита, когда установление этиологического агента сделало возможными профилактическую вакцинацию и терапию, антибиотиками. Еще раньше так было с пеллагрой. Есть основания надеяться, что то же самое произойдет с теми болезнями мозга, которые пока еще окутаны тайной.
Ф. КРИК
Мысли о мозге
Раздумывая о себе самом, человеческий мозг открыл некоторые поразительные факты. Чтобы понять, как он работает, очевидно, нужны новые методики его исследования и новая система понятий
В предыдущих статьях этого издания читатель, вероятно, видел, как мозг изучается на многих уровнях — от молекул в его синапсах до сложных форм поведения — и путем различных подходов — химического, анатомического, физиологического, эмбриологического и психологического - к нервной системе разнообразных животных, от примитивных беспозвоночных до самого человека. И все же читатель, вероятно, заметил также, что, несмотря на непрерывное накопление детальных сведений, то, как работает человеческий мозг, по-прежнему окутано глубокой тайной. Издатели "Scientific American" попросили меня как новичка в нейрофизиологии сделать некоторые общие замечания о том, как воспринимает эту проблему сравнительно посторонний наблюдатель. Я интересуюсь нейробиологией более 30 лет, но только последние года два пытаюсь заняться ею серьезно.
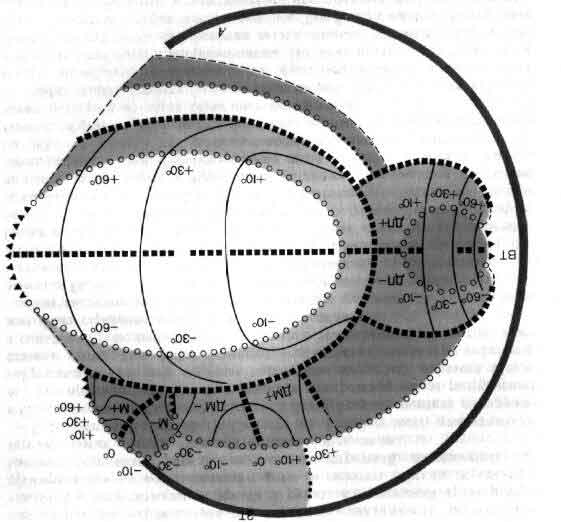
Зрительная кора ночной обезьяны служит примером того, как кора больших полушарий образует «карту» из областей, геометрически связанных с их функциями. В коре этого животного левое полушарие состоит из девяти областей, которые представляют собой организованные в соответствующем порядке карты поля зрения обезьяны (и трех областей, которые отвечают на стимулы, лежащие в поле зрения, но, по-видимому, не воспроизводят его в определенном порядке). А. На весьма схематичном изображении зрительная кора, занимающая заднюю треть коры больших полушарий, развернута таким образом, что ее можно видеть сверху. Геометрические отношения между полем зрения и разными участками зрительной коры были установлены в электрофизиологических опытах, где посредством микроэлектродов, введенных в зрительную кору, регистрировали ответы небольших групп нейронов на стимулы, предъявляемые в определенных участках поля зрения. Б. Здесь показана правая половина поля зрения. Квадратами обозначен ее горизонтальный меридиан, кружками — вертикальный, а треугольниками - крайняя периферия поля. Эти символы наложены на те участки мозга, которые реагируют на части поля, представленные символами. Девять организованных зрительных областей следующие: первая зрительная (Зр 1), вторая зрительная (Зр 2), дорсолатеральный серп (ДЛ), медиальная височная (MB), дорсальная промежуточная (ДП), дорсомедиальная (ДМ), медиальная (М), задняя вентральная (3В), передняя вентральная (ПВ). Три, по-видимому, неорганизованные зрительные области следующие: задняя теменная (ЗТ), височно-теменная (ВТ) и нижнетеменная (НТ). Плюсы обозначают верхнюю часть поля зрения, минусы - нижнюю часть. В. На дорсолатеральной поверхности мозга показано положение левого полушария и зрительных областей в нем. Картирование коры ночной обезьяны произвел Дж. Олмен (J. Allman) из Калифорнийского технологического института и И. Каас (J. Kaas) из Университета Вандербильта.
Приступая к новой дисциплине, полезно попытаться отделить те вопросы, которые хотя еще далеко не разгаданы, но по крайней мере могут, по-видимому, быть изучены тем или иным обычным способом, - от таких, для которых в настоящее время даже в общих чертах не предвидится готового объяснения. (Именно такой анализ привел нас с Дж. Уотсоном к поискам структуры ДНК.) К первой категории я бы отнес такие вопросы, как химическая и электрическая природа нейронов и синапсов, привыкание и сенситизация одиночных нейронов, действие медикаментов на нервную систему и т. д. Собственно говоря, я включил бы сюда почти всю нейроанатомию, нейрофармакологию и значительную часть нейрофизиологии. Даже развитие мозга не представляется мне по существу таинственным, несмотря на то что мы ничего не знаем о тех конкретных процессах, которые протекают в развивающемся эмбрионе.
В то же время некоторые функции человека, как мне кажется, недоступны пониманию на современном уровне наших знаний. Мы чувствуем, что есть нечто трудно объяснимое, но мы почти не в состоянии ясно и четко выразить, в чем состоит трудность. Это наводит на мысль, что весь наш способ мышления о таких проблемах, возможно, ошибочен. Из таких проблем я бы выдвинул на передний план восприятие, хотя другие, может быть, заменили бы его пониманием, воображением, волей или эмоциями. Все они имеют то общее, что составляет часть нашего субъективного опыта и что в них, вероятно, принимает участие множество сложно взаимодействующих нейронов.
Для того чтобы постичь эти высшие уровни нервной деятельности, очевидно хорошо было бы узнать как можно больше о более низких уровнях, особенно тех, которые доступны прямому эксперименту. Но само по себе такое знание еще недостаточно. Представляется несомненным, что мы должны рассмотреть теории, которые касаются непосредственно переработки информации в больших и сложных системах, будь то информация, поступающая от органов чувств, или инструкции, посылаемые мышцам и железам, или же поток информации, содержащийся в обширной нервной активности между этими двумя крайними членами.
Причина, по которой я выдвигаю на первый план восприятие и, в частности, зрительное восприятие, состоит в том, что, как ясно показывают Д. Хьюбел и Т. Визель (см. ), оно более доступно прямому эксперименту. Кроме того, наш внутренний образ внешнего мира и точен и ярок, что неудивительно, поскольку человек относится к животным, у которых зрение особенно хорошо развито. Чувство обоняния у человека, напротив, гораздо более смутное. Любопытно, что во многом зрительный образ строится способами, требующими от нас лишь незначительных усилий. Когда мы пытаемся думать об истинно трудных задачах, мы обычно выбираем что-нибудь вроде шахмат или математики, или изучения иностранного языка. Мало кто осознает, каким поразительным достижением является сама по себе способность видеть. Главный вклад в сравнительно новую область искусственного интеллекта состоял не столько в решении проблем обработки информации, сколько в том, чтобы показать, как невероятно трудны эти проблемы. Когда задумаешься над тем, какие расчеты должны быть произведены для опознания даже такой обычной картины, как человек, переходящий улицу, то поражаешься тому, что такое необыкновенное число последовательных детальных операций может быть осуществлено без всякого усилия за такое короткое время.
Создание более крупных, более быстрых и более дешевых компьютеров - достижение, еще далекое от конечной цели, - дало нам некоторое представление о том, чего можно достичь быстрым вычислением. К сожалению, аналогия между компьютером и мозгом хотя и полезна в некоторых отношениях, но может и ввести в заблуждение. В компьютере информация обрабатывается при быстром темпе импульсов и последовательно. В мозгу темп гораздо медленнее, но информация может быть обработана параллельно в миллионах каналов. Детали современной вычислительной машины очень надежны, но исключение одной или двух из них может нарушить все вычисление. По сравнению с ними нейроны мозга несколько ненадежны, но порча даже немалого их числа вряд ли приведет к сколько-нибудь заметному изменению поведения. Компьютер работает по строго двоичному коду, мозг же пользуется менее точными способами сигнализации. Зато он, по-видимому, приспосабливает сложными и тонкими приемами число и эффективность своих синапсов к тому, чтобы его операции соответствовали прошлому опыту. Поэтому не удивительно, что, хотя компьютер точно и быстро производит длинные и запутанные арифметические вычисления — в чем человек довольно слаб, - человек способен опознавать изображения такими приемами, к которым ни один из современных компьютеров еще и не начал приближаться.
Было бы не слишком удивительно, если бы орудием теоретического подхода к таким проблемам оказалась теория информации. Пока что применение ее к зрительному восприятию ограничивалось главным образом ранними этапами зрительного пути. Так, например, для объяснения того, как информация, посылаемая по ограниченному каналу (такому, как зрительный нерв), может в принципе быть представлена более подробно в зрительной коре головного мозга, из области обработки информации были взяты теорема отсчетов и метод пересечения нуля (по Логану). Кроме того, Д. Марр (D. Магг) из Массачусетского технологического института схематически представил ход вычислительных операций, какие должен выполнить мозг, чтобы мы видели предметы так, как мы это делаем. Это дало нам представление о сложности проблемы, но пока что не привело к сколько-нибудь значительному выяснению действующих механизмов. В частности, до сих пор мы не располагаем каким-либо описанием сознательного восприятия, которое осветило бы наш непосредственный опыт такого восприятия. Как можно полагать, такие феномены основаны на том, что пути вычислений действуют каким-то образом сами на себя, но как именно это происходит, неизвестно. Поскольку эта центральная проблема ускользает от решения, нам остается только обратиться к более локальным и частным вопросам в надежде, что, трудясь над их решением, мы натолкнемся на правильный подход к более сложным глобальным проблемам.
Какой, в общем смысле, аппарат дает мозгу возможность выполнять свою замечательную деятельность? Число компонентов (нейронов) в мозгу составляет, вероятно, около 1011. Число синапсов, или контактов между нейронами, равно, возможно, 1015. В среднем на каждом нейроне имеется несколько тысяч отдельных входов, и он сам посылает связи ко многим другим нейронам. Физическое распределение большинства компонентов не особенно отчетливое. Дендриты, или короткие волокна, соседних нейронов сложно переплетены, хотя обычно они не соприкасаются друг с другом. Между дендритами ветвятся аксоны, или длинные волокна нейронов; многие из них часто обладают тысячами контактов. Монтажная схема таких волокон, если бы возможно было ее создать, разумеется, была бы весьма запутанной.
Как пробираться через эти немыслимые джунгли? Традиционный способ состоит в том, что удаляют части мозга и смотрят, как меняется поведение. Это можно проделать на экспериментальных животных, хотя лишь немногие операции достигают желаемой точности. Над людьми такие «опыты» вопреки нашему желанию совершают инсульты, опухоли и травмы, но в этих случаях трудно точно определить характер и степень повреждения. Тем не менее такие «опыты» принесли много ценных сведений. Возникают два важных вывода. Первый состоит в том, что разные области мозга выполняют разную работу. Нарушения деятельности мозга вследствие повреждения одной области часто совсем иные, чем нарушения, возникающие при повреждении другой. Второе заключение несколько неожиданно. Некоторые опыты показывают, что мозг перерабатывает информацию способами, совершенно отличными от тех, какие можно было бы предполагать. Такие процессы, как узнавание букв и узнавание цифр, которые, казалось бы, должны протекать в одном и том же месте, по-видимому, осуществляются в разных местах. Справедливо и обратное: некоторые процессы, представляющиеся раздельными, нарушаются при повреждении одной и той же области.
Это приводит нас к одному из самых серьезных предостережений, какое должен принять исследователь мозга. На всех уровнях нас вводит в заблуждение наша интроспекция. Это относится не только к сложным процессам, но и к процессам, которые представляются простыми. Человек, не размышлявший о таких вещах, возможно, полагает, что видит в деталях одновременно все части зримого мира. Хотя он и знает, что те предметы, на которые он смотрит «краем глаза», видны не очень четко, но не отдает себе отчета в том, насколько в действительности узко то окно ясного видения, через которое он создает себе большую часть картины внешнего мира. Поскольку движением глаз человек по желанию выделяет детали в любой части видимого мира, у него создается впечатление, что она «все время перед ним».
Быть может, наиболее яркой иллюстрацией сказанного служит наличие слепого пятна в каждом глазу. Далеко не все осознают его присутствие, хотя это легко доказать. Примечательно, что в нашем поле зрения мы не видим дыры. Происходит это отчасти потому, что у нас нет способов обнаружить ее края, а отчасти потому, что мозг заполняет дыру зрительной информацией, получаемой из ее ближайшего окружения. Наша способность к самообману по поводу работы собственного мозга почти безгранична, главным образом потому, что часть, о которой мы можем сообщить, составляет лишь ничтожную долю того, что происходит у нас в голове. Вот почему философия в значительной части бесплодна уже более 2000 лет и, вероятно, останется такой же, пока философы не научатся понимать язык переработки информации.
Это не значит, однако, что следует вовсе отказаться от изучения нашей умственной деятельности путем интроспекции, как это попытались сделать бихевиористы. Поступить так, значило бы устранить один из наиболее значительных атрибутов того, что мы стараемся изучить. Но факт тот, что никогда нельзя принимать на веру свидетельства интроспекции. Им следует давать объяснения иные, чем только те, которые дает она сама.
Основная проблема состоит в том, что почти всякий процесс, который можно изучать, наблюдая общее поведение (например, чтение), включает сложное взаимодействие многих областей мозга, каждая из которых перерабатывает информацию по-своему. Мы знаем только в самых общих чертах, как распознать и классифицировать эти разные области. Хотя наши сведения о том, как они взаимосвязаны, быстро расширяются, но ни качество, ни количество этих сведений еще далеко не достаточны. Кроме того, мы редко знаем, какую операцию производит каждая область, т.е. как соотносятся выходы с входами, а в некоторых случаях мы даже не имеем ни малейшего представления о том, что происходит.
В этом лежит главная причина того, почему по мерке строгой науки чистая психология довольно бесплодна. Это не значит, что она не может быть количественной. Одна ее отрасль, довольно забавно именуемая психофизикой, безусловно, использует количественный подход и притом часто разумно и с воображением. Основная трудность состоит в том, что психология пытается обращаться с мозгом, как с черным ящиком. Экспериментатор изучает входы и выходы и из полученных результатов старается вывести структуру и операции внутри ящика. Такой подход необязательно плох. Многие годы предмет генетики был черным ящиком. Она старалась, не без известного успеха, вывести структуру и функцию генетического материала из изучения результатов выведения животных и растений. И, действительно, немало хорошего в биологии создано методом черного ящика. Это возможно на любом уровне. Для предыдущего поколения биохимиков фермент был черным ящиком. В наше время многие энзимологи изучают структуру фермента с целью связать ее с его поведением. То, что для одного является черным ящиком, для другого составляет задачу исследования.
Трудность подхода, исходящего из принципа черного ящика, состоит в том, что если только ящик по сути своей не совсем прост, то скоро наступает стадия, когда наблюдаемые результаты одинаково хорошо объясняются несколькими соперничающими теориями. Попытки выбрать одну из них часто оказываются безуспешными, поскольку по мере проведения новых опытов выявляются все большие сложности. И если вопрос тем или иным способом должен быть решен, то здесь нет иного выбора, как постучаться внутрь ящика.
То, что мы знаем о мозге — как это здесь показано, - говорит нам о двух вещах. Мозг явно настолько сложен, что возможности предсказывать его поведение только на основании изучения его частей еще слишком далеки, чтобы стоило принимать их во внимание. Эта же сложность служит нам предостережением, что если применяемый чистой психологией подход по принципу черного ящика не потерпит поражения, то это будет для него большой удачей. Психология необходима. Что делает организм в действительности, мы можем узнать, только наблюдая его. Однако одна только психология скорее всего окажется бесплодной. Она должна сочетать изучение поведения с параллельным исследованием того, что происходит внутри мозга. Хорошим примером служит исследование Р. Сперри и его сотрудников из Калифорнийского технологического института, проведенное у больных с «расщепленным мозгом», у которых нарушены связи между двумя полушариями большого мозга. Другим примером является применение дезоксиглюкозы для введения метки в те участки мозга, в которых уровень активности выше среднего, когда подопытное животное выполняет какую-нибудь задачу. Таким образом, изучение нейроанатомии и нейрофизиологии может сочетаться с исследованиями поведения. Мы должны изучать и структуру, и функцию, но изучать их внутри черного ящика, а не только снаружи.
Новичок в таких вопросах бывает рад, когда, отпрянув в ужасе от сложностей нейроанатомии, он потом обнаруживает, что этот предмет в последнее время стал одним из самых захватывающих в нейробиологии. Это объясняется тем, что хотя, вообще говоря, схема соединений мозга очень запутанна, но работы последнего времени показали, что эти соединения гораздо более упорядоченны, чем можно было думать
всего несколько лет назад. Такой переворот произошел в значительной мере благодаря новым экспериментальным методикам, пришедшим из биохимии, в частности благодаря использованию аминокислот, меченных радиоактивными изотопами, и фермента пероксидазы хрена для прослеживания связей, а также благодаря использованию меченой дезоксиглюкозы для тех участков, где нейроны особенно активны. Кроме того, плодотворным оказалось применение специфических антител для «окраски» определенного класса нейронов. Есть надежда, что эффективная новая методика моноспецифических (моноклональных) антител, которую только начали применять, даст еще больше для классификации нейронов по типам, имеющим определенное значение, и для обнаружения их трехмерного распределения. Однако задачи эти столь грандиозны, что, как можно сказать с большой долей уверенности, без разработки еще и других новых методов движение вперед будет медленным. Резкий подъем молекулярной биологии оказался возможным благодаря продуманным поискам новых методов (например, методов быстрого определения последовательности оснований в ДНК).
Подобно нейроанатомии нейрофизиология неуклонно развивается, особенно с тех пор, как появилась возможность регистрировать импульсацию отдельных нейронов, а не групп клеток. Хорошим примером этого служит классическая работа Хьюбела и Визеля на зрительной коре. Однако можно предполагать, что и здесь нужно будет изменить темпы. В частности, возможно, понадобится отводить активность многих нейронов одновременно и независимо друг от друга, используя для этого новые возможности, какие дает нам микроэлектроника. Это позволило бы также более эффективно изучать ответы на более сложные комбинации стимулов, например ответ на две или три параллельные линии в поле зрения, а не только на одну.
Как в нейроанатомии, так и в нейрофизиологии накопление новой информации идет медленно по сравнению с общим ее количеством, содержащимся в системе. Поэтому важная роль теоретической нейробиологии состоит не просто в попытках создать правильные и детальные теории нервных процессов (что может оказаться очень трудной задачей), но и в том, чтобы указать, какие свойства полезнее всего изучать и в особенности измерять, чтобы понять, какого рода теория требуется. Бесполезно требовать невозможного, например точной схемы соединений в одном кубическом миллиметре ткани мозга или определения характера импульсации всех нейронов в нем. Задача в том, чтобы установить, какая легко доступная информация была бы самой полезной, а также (что труднее), какие данные можно получить в разумные сроки, если ввести осуществимые новые методики. Иногда ясная формулировка требования - это уже полдела для понимания того, как его выполнить. Например, цены бы не было методике, которая позволила бы вводить в нейрон вещество, способное четко окрасить все связанные с ним нейроны и только их. То же самое относится к методу, при помощи которого можно было бы инактивировать все нейроны строго одного типа, сохраняя все остальные более или менее неизмененными.
Как далеко все это привело нас в настоящее время? Помещенные в данном выпуске статьи дают хорошее общее представление о достигнутых успехах. Чего заметно не хватает, так это широких идейных рамок, в пределах которых можно интерпретировать все эти различные подходы. В таком состоянии находились биохимия и генетика до того, как произошла революция в молекулярной биологии. Дело не в том, что большинство нейробиологов не имеют какого-либо общего представления о том, что происходит. Беда в том, что представление это не сформулировано точно. Стоит притронуться к нему, как оно разваливается. Если привести лишь несколько примеров, то природа восприятия, нейронные корреляты долговременной памяти, функция сна — все они носят такой характер.
Как же в таком случае следует строить общую теорию мозга? На ее пути стоят, по-видимому, три важных ограничительных условия. Первое - это природа физического мира. Повседневный мир, в котором мы живем, не является чем-то аморфным и беспорядочным. Он состоит из предметов, которые обычно занимают определенное пространство и которые, хотя и перемещаются относительно других предметов, сохраняют свои размеры и форму. Зрительно предмет обладает поверхностями, очертаниями, цветом и т. п. Он может испускать звук или запах. Не вникая в детали, безусловно можно ожидать, что какова бы ни была переработка информации, поступающей в мозг, она будет связана с инвариантами и полуинвариантами во внешнем мире, доступными органам чувств.
Эксперимент показывает, что так это и происходит, притом часто неожиданным образом. Хорошим примером является такого рода восприятие цвета, какое показано в опытах Э. Ленда (Е. Land). Можно было бы думать, что когда дело идет о большом цветном пятне, то воспринимается окраска света, который действительно исходит от пятна и падает на сетчатку. На самом же деле цвет в большинстве случаев создается сочетанием окраски света, падающего на предмет, и света, отражаемого поверхностью предмета. Как это ни удивительно, мозг способен извлекать эту последнюю информацию из поступающих зрительных сигналов. Она может в значительной степени компенсировать характер падающего света. То, что мы воспринимаем, больше соответствует отражению от поверхности, которое является свойством самого предмета.
Поразительная демонстрация такого рода феномена проводится в Эксплораториуме в Сан-Франциско, хотя в этом случае поверхности имеют разные оттенки серого — от почти белого до совершенно черного. Освещение, источник которого скрыт, здесь неровное — оно гораздо сильнее в нижней части экспозиции, чем в верхней. Один участок внизу выглядит очень черным, другой участок наверху кажется почти белым. С помощью узких трубок эти участки можно рассматривать каждый по отдельности. И если смотреть в них таким образом, то видно, что оба участка имеют в точности один и тот же оттенок серого. Когда моя жена - художница - у видела этот экспонат, она была поражена и заявила, что это фокус. Она не отдает себе отчета в том, что в известном смысле все, что она видит, это фокус, который показывает ей ее мозг.
Второе ограничительное условие налагается биохимией, генетикой и эмбриологией. Нервная система сделана не из металла или неорганических полупроводников, а из специализированных клеток. Импульс, бегущий по аксону, движется с умеренной скоростью по сравнению со скоростью света (даже несмотря на то, что нейрон может при помощи разных фокусов ускорять его движение); это ограничение налагается на него биохимией. Хотя важную роль в нейронной активности играют такие неорганические ионы, как натрий и калий, не вызывает удивления тот факт, что передача нервного импульса от одного нейрона к другому производится мелкими органическими молекулами, потому что многие такие молекулы синтезируются с легкостью. Удивительно то, что один и тот же медиатор действует в столь многих различных местах - ограничительное условие, вероятно, налагаемое эволюцией.
Дальний конец аксона находится далеко от ближайшего пункта синтеза белка (не считая митохондрий внутри аксона), и это обстоятельство может налагать ограничения на скорость, с которой в нем происходят некоторые биохимические изменения. Вероятно, существуют одни типы нейронных цепей, с которыми организм справляется сравнительно легко, и другие, которые для него непосильны. Насколько мы знаем, генам высшего животного может быть трудно придавать схеме нейронных связей большую точность, особенно если в схеме участвует очень много клеток. Например, точного распределения связей, необходимого для правильного стереоскопического зрения, трудно достичь без того благотворного влияния, какое оказывает некоторый контакт с реальным внешним миром, может быть, потому, что системы, связанные с двумя глазами, вероятно, не могут быть построены с требуемой точностью.
Возможны и другие ограничения. Как предположил много лет назад Г. Дэйл, нейрону, по-видимому, трудно так устроить свои дела, чтобы одна веточка его аксонного дерева выделяла один медиатор, а другая - другой. Этот принцип, возможно, лежит в основе случая, описанного Э. Кэнделом в настоящем выпуске (см. статью «Малые системы нейронов»), когда медиатор, продуцируемый одной и той же клеткой, возбуждал одни клетки, тормозил другие и оказывал смешанное влияние на клетки третьего типа.
Третье ограничительное условие налагается математикой, в частности теорией связи. На первый взгляд может показаться, что некоторые такие результаты противоречат ожиданиям. При известных обстоятельствах распределение, или общая структура (pattern), прекрасно воссоздается на основе небольшой выборки, если производить ее через правильные промежутки. Информация может быть заложена в память в распределенной форме подобно голограмме таким образом, что извлечение части хранимой информации не устраняет какой-либо части картины, хотя снижает до некоторой степени качество всей картины.
Возникает искушение ввести четвертое ограничительное условие, но опыт показал, что оно ненадежно. Это условие налагается эволюцией. Несомненно, все организмы и их компоненты образовались в результате долгого эволюционного процесса, и это обстоятельство никогда нельзя забывать. Однако неразумно было бы угверждать (разве только в самых общих выражениях), что эволюция не могла сделать того-то или должна была сделать то-то. Биолог должен руководствоваться хорошим рабочим правилом, что эволюция гораздо умнее его. Это не значит, что сравнительные биологические исследования не могут показать, что определенная структура часто связана с определенной функцией. Таксономическое сопоставление результатов эволюции может привести к полезным предположениям, но такие предположения всегда должны быть подтверждены прямым экспериментом.
Как бы то ни было, но первые три ограничительных условия мы понимаем лишь частично. Процесс разложения окружающего нас мира на его существенные признаки не всегда протекает прямолинейно. Многие важные вопросы эмбриологии еще не получили ответа. Теория информации - сравнительно новая отрасль знания. Поэтому, хотя и есть возможность получать какие-то наводящие указания, рассматривая все три условия, исследователь редко встречается с таким обилием ограничений, что это практически позволило бы сделать выбор между теориями. Существует так много способов, какими наш мозг мог бы перерабатывать информацию, что без существенной помощи прямых экспериментальных фактов (а они обычно малочисленны) мы вряд ли сделаем правильный выбор.
Существуют ли какие-то идеи, которых следует избегать? Я думаю, что одна, по крайней мере, есть - это идея гомункулуса. Недавно я пытался разъяснить одной умной женщине проблему, которая состоит в том, как понять, что мы вообще воспринимаем что бы то ни было, но мне это никак не удавалось. Она не могла понять, в чем тут проблема. Наконец, в отчаянии я спросил ее, как она сама считает, каким образом она видит мир. Женщина ответила, что, вероятно, где-то в голове у нее есть что-то вроде маленького телевизора. «А кто же в таком случае, - спросил я, - смотрит на экран?» Тут она сразу же поняла, в чем проблема.
Большинство нейробиологов считает, что в мозгу нет гомункулуса. К несчастью, легче констатировать ошибку, чем не впасть в нее. Это происходит потому, что мы несомненно питаем иллюзию существования гомункулуса — нашей личности. Вероятно, сила и прочность этой иллюзии имеют свои основания. Возможно, она отражает некоторые аспекты общего управления мозгом, но какова природа этого управления, мы еще не узнали.
Следовало бы избегать еще одной общей ошибки. Ее можно было бы назвать «ошибкой премудрого нейрона». Представим себе нейрон, который посылает сигнал на некоторое расстояние по своему аксону.
Что этот сигнал сообщает воспринимающему синапсу? Сигнал, разумеется, закодирован частотой нервных импульсов, но что означает его сообщение? Легко усвоить привычку считать, что оно содержит в себе больше, чем это есть в действительности.
Возьмем, например, нейрон в зрительной системе, который считается цветочувствительным. Предположим, что лучше всего он разряжается при стимуляции пятнышком желтого света. Мы склонны думать, будто он сообщает нам, что свет в этой точке желтый. Однако на самом деле это не так, потому что большинство рецепторов цвета имеют широкую кривую ответов и генерируют импульсы — во всяком случае, в известной степени — в довольно широком диапазоне длин волн. Поэтому данная частота импульсации может быть вызвана и слабым желтым и сильным красным светом. Кроме того, на импульсацию данного нейрона могли повлиять объем движения светового пятна и его точные форма и размеры. Короче говоря, множество разных, хотя и связанных между собой входов вызовут импульсацию одной и той же частоты.
Поскольку относящийся к данному стимулу сенсорный вход в нейрон обладает многими признаками, а выход (грубо говоря) только один, то информация, передаваемая одним нейроном, обязательно неоднозначна. Однако следует иметь в виду, что мы можем, кроме того, извлечь информацию, сравнивая импульсацию одного нейрона с импульсацией другого или нескольких других нейронов. Посредством одного только типа рецепторов (палочек) мы вовсе не можем воспринять цвет, а видим только оттенки серого. Чтобы свет в нашем восприятии был окрашен, нужно не меньше двух типов рецепторов, причем у каждого из них кривая ответов на разные длины волн отличается от кривой для другого нейрона. Опытами показано, что это именно так: мы можем воспринимать цвет палочками при участии хотя бы одного типа колбочек.
Следует иметь в виду, что это соображение относится не только к цвету. Одиночный «детектор границы» в действительности не сообщает нам о наличии границы. Он обнаруживает, говоря приблизительно, «свойство границы» во входных сигналах, т. е. особого типа неоднородность изображения на сетчатке, которая может быть создана многими разными предметами. Одна из задач теоретической нейробиологии состоит в том, чтобы постараться превратить такие неопределенные понятия, как «свойство границы», в математически точные описания.
Это общее соображение относится ко всем уровням нервной системы. Оно ясно показывает, почему для извлечения полезной информации из входной нервной активности нам нужно перерабатывать ее столь многими различными способами.
Но все же, нельзя ли нарисовать некую общую картину переработки информации? Пожалуй, легче всего это сделать, рассматривая зрительную систему и, в частности, зрительную кору большого мозга. На первый взгляд, карта зрительных ответов в стриарной коре (поле 17) представляет собой топологически соответствующую картину половины поля зрения. Но надо сказать, что картина эта определенным образом искажена: большая ее часть соответствует центральной области сетчатки с ее высокой остротой зрения (центральной ямке) и меньшая часть — периферии. Тем не менее эта карта достаточно упорядочена и довольно точна.
Более внимательное рассмотрение показывает, что дело обстоит не так просто. Входы от левого глаза переплетаются в форме полос со входами от правого глаза. Кроме того, каждый глаз обладает входами трех разных типов (от латерального коленчатого тела): один — для Y-клеток сетчатки (которые дают довольно короткие ответы) и остальные два входа — для Х-клеток. Кроме того, .имеются мощные входы от таких образований, как подушка, а также от других областей зрительной коры. Выходов тоже много. От этого поля идет не один выход, а множество, отчасти к нейронам, образующим карты в других зрительных областях, и отчасти к подкорковым областям, в том числе сравнительно крупный выход обратно к латеральному коленчатому телу, откуда первоначально пришли многие входы. Таким образом, этот участок коры представляет собой область множественных входов и множественных выходов. Каждый вход или выход включает множество отдельных аксонов (миллионы), распространяющихся по всей области.
Какую переработку информации выполняет стриарная кора (первичная зрительная область)? Как показано в статье Хьюбела и Визеля, главная функция стриарной коры состоит в ответах на разную ориентацию форм в поле зрения. В каждой «колонке» стриарной коры нейроны отвечают только на одну ориентацию, хотя их ответ может относиться к любому из множества типов. Преобразованная таким образом входная активность затем направляется в другие пункты.
Как широко процесс обработки информации распространяется по коре в разные стороны? Поразительно, что он весьма локален. За пределами нескольких миллиметров уже сравнительно мало взаимосвязей, если не считать аксонных связей диффузного характера, идущих от ствола мозга. Каждый маленький участок содержит множество взаимосвязанных нейронов (каждый квадратный миллиметр поверхности коры включает около 100000 нейронов), но они почти не имеют каких-либо прямых связей с теми нейронами, которые реагируют на более далекие части поля зрения. (У макака общая поверхность стриарной коры в одном полушарии составляет приблизительно 1400 квадратных миллиметров.)
Следует отметить, что пока еще нет убедительных свидетельств о наличии внутри области дискретных модулей. Отношения здесь скорее такие, как между жителями воображаемого города, которым запрещено удаляться от дома более чем на полмили. Они могут общаться со своими соседями на расстоянии одной мили, особенно со своими ближайшими соседями, но не имеют никаких прямых контактов с теми, кто живет подальше.
Примечательно, что это описание, в его самом общем виде, применимо к большей части коры, если не ко всей. Известно множество отдельных зрительных полей, каждое с какой-либо «картой» поля зрения. В коре у ночной обезьяны по меньшей мере восемь, а может быть, и больше, преимущественно зрительных полей. Если мы взглянем на слуховые или на соматосенсорные области, то увидим то же самое. Слуховая кора этой обезьяны состоит из четырех отдельных полей, «картированных» по частоте и, вероятно, по амплитуде. Поверхность тела обезьяны картирована несколько раз в соматосенсорных областях. Во всех случаях входы, большая часть которых идет к средним слоям коры, состоят из чередующихся полос того или иного типа. Эти входы смешиваются при переработке информации, которая происходит в верхних и нижних слоях. После переработки выходная активность направляется упорядоченным образом к нескольким другим пунктам как в коре, так и в подкорковых областях. Вполне оправдана гипотеза, согласно которой в случае такой связи между двумя полями выход одного из них картируется упорядоченно, но необязательно однородно на поверхности второго поля. Кроме того, часто наблюдаются обратные отношения. У обезьяны поле 17 посылает картированную проекцию к полю 18, а это последнее тоже посылает часть своей выходной активности обратно к полю 17. Эти обратные связи, вероятно, не диффузны, а образуют обратную карту. Насколько точно прямая и обратная карты соответствуют друг другу, еще неизвестно. Можно думать, что соответствие окажется довольно точным.
Из того правила, что в корковых слоях не обнаружено отчетливых клубочков нейронов, имеется одно интересное исключение. Это бочонки, исследованные Т. Вулси (Т. Woolsey) с сотрудниками в Медицинской школе Вашингтонского университета и упомянутые в настоящем выпуске в статье У. Коуэна (см. «Развитие мозга»}. Каждый бочонок имеет в поперечнике приблизительно 200 ммк, и, хотя соседние бочонки соединены между собой, связи внутри бочонка гораздо богаче. Это служит иллюстрацией того, как кора справляется с мелкими, пространственно дискретными, а не непрерывными входами.
Возможно ли, что кора содержит фиксированное, определимое число отдельных полей? Это, очевидно, верно для сенсорных и, вероятно, для моторных полей. На приведенном мною рисунке изображена карта зрительной коры ночной обезьяны, составленная недавно Дж. Олменом (J. Allman) с сотрудниками из Калифорнийского технологического института. На ней показаны восемь отдельных полей, картированных в этой области. Как можно видеть, хотя каждое поле и имеет на карте достаточно очерченные границы и определяется в этих случаях вполне однозначно, в коре в этих точках отсутствуют отчетливые изолирующие границы. В обоих полушариях кора представляет собой непрерывный слой с одним только краем. В нем нет «щелей». Неудивительно поэтому, что на границах в картировании имеется, говоря приближенно, локальная плоскость зеркальной симметрии. Иными словами, две карты, по одной с каждой стороны границы, взаимосвязаны. При продвижении вдоль границы эти карты по обе стороны от нее одинаковы. При удалении от нее под прямым углом направление движения в поле зрения на одной карте такое же, как при удалении от границы на другой карте. Если внимательно рассматривать рисунок, то видно, что это правило нарушается только в одном месте. Правило локальной плоскости зеркальной симметрии сохраняет свою силу на картах как слуховой, так и соматосенсорной системы, опять-таки с несколькими исключениями. Этого как раз и следовало ожидать при наличии ряда отдельных карт, которые в то же время до известной степени взаимодействуют по краям. Это позволяет думать, что в процессе эволюции новые функциональные поля головного мозга возникают попарно.
Распространяется ли такое картирование в виде обособленных полей также на остальные части коры, в частности на лобные области, и на то, что мы называем ассоциативными зонами? В настоящее время никто этого не знает даже про обезьян, не говоря уже о человеке. Но уже теперь ясно, что у обезьян большую часть коры можно картировать таким образом. Хотя легко представить себе, как можно было бы разрушить представление о полях (понятие поля полезно только при условии, если несколько разных его определений однозначно приводят к одному и тому же делению на поля), я склоняюсь к тому, что это представление правильно почти для всей коры. А если оно правильно, то сколько же полей в коре человека? Больше 50? Может быть, меньше 100? Если бы каждое поле можно было посмертно четко окрасить так, чтобы точно увидеть, сколько всего полей, каких размеров каждое из них и как именно оно соединено с остальными полями, это явилось бы большим шагом вперед.
До сих пор я говорил только о коре большого мозга, но кора мозжечка очевидно с ней сходна. Здесь также входы упорядочены и образуют более одной карты. Оба главных входа, по-видимому, расположены в виде полос. Есть что-то в эмбриологии, чему «нравятся» полосы. Это очень ярко показала М. Константин-Патон (М. Constantine-Paton) из Принстонского университета, которая путем эмбриологических манипуляций создала лягушек с третьим глазом. В норме на каждый tectum opticum проецируется только один глаз, но в этом случае к нему могут идти проекции от обоих глаз. Когда это происходит, входы располагаются полосами, чего не бывает у нормальных лягушек.
Далее, если взять такие подкорковые области, как таламус, то здесь мы снова найдем некоторые свидетельства упорядоченного картирования. Каждое кортикальное поле обычно содержит карту своего собственного участка таламуса, часто в виде пятен. Упорядоченные карты, очевидно, имеются и в других частях нервной системы - базальных ганглиях, стволе мозга, спинном мозгу и т. д. В каждом таком случае нам нужно точно знать, как раздробить обширную совокупность нейронов на мельчайшие, имеющие определенное значение единицы, даже если эти единицы взаимодействуют до известной степени со своими соседями. Во многих случаях они представляют собой слои или части слоев, в других случаях имеют более компактную форму. Их входы и выходы не всегда расположены так аккуратно, как в коре, и поэтому задача не всегда будет легкой.
Снабженные весьма приблизительной картиной высших отделов головного мозга, мы теперь можем обратиться к общим вопросам о природе связей. Для этого нам нужны два довольно простых понятия: прецизионная схема связей и ассоциативные сети.
В то время как иногда кажется, что в головном мозгу все соединено со всем, в прецизионной схеме связи расположены определенным, упорядоченным образом. Лишь определенные клетки связаны с другими определенными клетками, а общая структура связей часто одинакова у отдельных животных. Прецизионное распределение связей обычно обнаруживается там, где в нем участвует небольшое число клеток, как, например, у сравнительно примитивного беспозвоночного аплизии, описанного в статье Кэндела (см. стр. 59). Хорошим примером служит также небольшой круглый червь Caenorhabditis elegans, которого изучал С. Бреннер (S. Brenner) с сотрудниками в Лаборатории молекулярной биологии Медицинского исследовательского совета в Кембридже в Англии. У этого вида сеть состоит точно из 279 нейронов, которые соединены между собой в точности одинаково у всех особей. Более многочисленные нейроны тоже могут быть связаны прецизионно, в особенности там, где структура из клеток повторяется многократно, что характерно, например, для глаза мухи. Прецизионное распределение связей не исключает возможности обучения, что ясно показано в статье Кэндела, потому что сила связей может быть изменена опытом.
В то же время, если рассматривать схему распределения связей (в той мере, в какой она нам известна) в области головного мозга у более сложного животного, скажем поля в зрительной коре обезьяны, то обнаружатся два обстоятельства. Клеток здесь гораздо больше, а распределение связей между ними, очевидно, гораздо менее прецизионное. В одном полушарии головного мозга у обезьяны оно, безусловно, не точно такое же, как в другом. Тем не менее связи, идущие от глаза к зрительной коре, распределены вовсе не случайно. Как пишут в своей работе Хьюбел и Визель, эти связи образуют, хотя и не точную, топографическую карту. Нейроны относятся здесь ко многим типам, которые связаны между собой не случайно, хотя точную степень упорядоченности этих связей установить трудно. Создается впечатление, что в пределах одного небольшого участка точные связи отчасти являются делом случая. Кроме того, один предъявляемый глазу простой сигнал — скажем, короткая линия в одном пункте поля зрения — возбудит не один-единственный детектор границы, а, возможно, несколько тысяч таких детекторов. Одним словом, структура связей не только рассчитана на извлечение из входных сигналов их признаков, но, по-видимому, обладает также некоторыми свойствами ассоциативной сети.
Ассоциативная сеть — это абстрактная схема соединений, изучаемая теоретиками - Марром (Marr), К. Лонге-Хиггинсом (Ch. Longuet-Higgins), Л. Купером (L. Cooper) и другими. Такая сеть имеет набор входных каналов (иногда несколько наборов) и набор выходных каналов. Каждый входной канал связан со всеми выходными, но сила связей неодинакова. Точное распределение зависит от типа рассматриваемой сети. Сила связи отрегулирована «на основании опыта» по определенным точным правилам, обычно так, что те проводящие пути, которые часто активируются совместно, каким-то образом усиливаются.
Такие сети служат для тонкой настройки системы с частично прецизионным распределением связей или способствуют вызову сложного выхода, когда приходит входная активность (или еще лучше - частичная входная активность) от чего-либо, с ним связанного. Взглянув на лицо человека, вы вспоминаете его имя (хотя, увы! не всегда). Вы в состоянии вспомнить человека, даже если увидели лишь часть его лица.
Высшая нервная система представляет собой чрезвычайно хитроумную комбинацию прецизионного распределения связей с ассоциативными сетями. Она построена не так, чтобы каждый входной канал был прямо связан со всеми остальными входными каналами. Кроме того, у высших животных она, по-видимому, не построена прецизионно. Для достижения своих целей система использует две стратегии. Одна из них - стратегия множественного и последовательного картирования (включая реципрокное картирование); она является компонентом, аппроксимирующим прецизионное распределение связей. Но, кроме того, система, по-видимому, организует связи таким образом, что локально - в пределах маленького участка — она, грубо говоря, соединяет все со всем.
Каждый участок содержит целое семейство локальных перекрывающихся ассоциативных сетей. Поэтому на ранней стадии переработки некоторые сигналы (скажем, одна группа от глаза и другая — от уха) не соотносятся друг с другом. Но по мере перехода сигналов от одной карты к другой первоначальное картирование становится и более диффузным, и более отвлеченным (например, ответ на ориентацию лучше, чем на пятно), и, таким образом, сигнал анализируется последовательно все более сложными способами в сочетании с сигналами от других входов.
При таком описании системы мы сразу видим, что это не просто одна огромная ассоциативная сеть. Построить такую сеть, в которой каждый нейрон реагирует непосредственно со всеми другими нейронами, было бы гораздо труднее и она заняла бы гораздо больше места. Поэтому сеть разделена на множество мелких подсетей; одни из них связаны параллельно, другие более последовательно. Кроме того, дробление на подсети отражает внешнюю и внутреннюю структуру окружающего мира, а также наши взаимоотношения с ним. Каждая локальная сеть приспособлена для выполнения тех специальных операций на своем входе, которые нужнее всего в этом пункте для извлечения значимой новой информации. При рассмотрении под таким углом зрения многие общие свойства головного мозга — многочисленные функциональные области, множественные связи с каждым нейроном — начинают приобретать некий смысл.
Разумеется, деятельность головного мозга еще гораздо шире, чем те процессы, которые я обрисовал выше. Должны существовать механизмы внимания, в особенности сосредоточенного внимания, усиливающие активность малых частей мозга. Должна существовать какая-то общая управляющая система. Чтобы мое чрезмерно упрощенное описание не ввело читателя в заблуждение, я советую ему обратиться к статье У. Науты и М. Фейртага (W. Nauta, M. Feirtag), где ясно показано, насколько сложно в действительности все устройство (см. «Организация мозга»). Тем не менее нарисованная мною схема покажет хотя бы кое-что из того, что можно надеяться увидеть при более детальном исследовании нервных процессов.
Какая система или какой уровень вероятнее всего окажется наиболее доступным экспериментальному изучению? Ответить на этот вопрос всегда трудно. Как указывает Кэндел, беспозвоночные животные с их крупными клетками, связанными между собой довольно точно, представляют много преимуществ, и можно не сомневаться, что некоторые созданные в процессе их исследования методики, полученные данные и проникновение в их смысл окажутся полезными также для понимания нервной системы высших и более сложных животных. Сомнительно, однако, чтобы на низших животных можно было получить ответы на все вопросы. Трудно также сказать, какое животное послужит наилучшей моделью человека и какой отдел мозга легче всего изучать. Зрительная система макака, очевидно, очень сходна с нашей. Зрительная система кошки меньше похожа на нее, но кошки обладают другими достоинствами для эксперимента. Много лет не утихает спор о том, какая система более пригодна для изучения — мозжечок или зрительная кора. (Поступающие в зрительную кору сигналы легче контролировать, но упорядоченность ее нейронного строения — ничто по сравнению с упорядоченностью структуры мозжечка.)
Как решить, например, что лучше - детально изучить одну или несколько областей коры, с тем чтобы точно объяснить наблюдаемые в них процессы с позиций нейроанатомии и нейрофизиологии, или же считать все кортикальные области маленькими черными ящиками и сосредоточиться на отношениях между ними? Вполне можно считать, что области, контролирующие более диффузные «активационные» системы, например систему проекций от locus coeruleus, более доступны изучению, чем системы, детально перерабатывающие информацию. На чем бы ни остановиться, за что ни взяться в первую очередь, очевидно, предстоит еще долгий путь, пока мы достигнем, хотя бы в общих чертах, такого понимания работы мозга, которое будет прочно опираться на эксперимент и на теорию.
Каковы в таком случае ближайшие перспективы для нашего понимания мозга? По мере накопления сведений можно ли надеяться на своего рода «прорыв»? Это всегда возможно, но перспективы здесь не очень обнадеживающие. Иногда забывают, что нейробиология уже совершила несколько таких прорывов. Одним из них было открытие, что нервный импульс распространяется по аксону в форме «спайков» приблизительно одинаковой амплитуды и с одинаковой скоростью. Другой прорыв произошел, когда стало ясно, что в большинстве синапсов происходит химическая передача и, в особенности, что синапсы бывают как возбудительные, так и тормозные. Оба этих открытия касаются феноменов, возникших на ранней стадии эволюции животных. Несколько отрезвляет то обстоятельство, что в молекулярной биологии основные прорывы тоже были связаны с механизмами, возникшими уже очень давно. Часто сложные природные явления основаны на простых процессах, но эволюция обычно украшала их всякими видоизменениями и добавлениями в стиле барокко. Разглядеть скрытую под ними простоту, которая в большинстве случаев появлялась достаточно рано, часто бывает чрезвычайно трудно.
Существует еще одна проблема. Анализируя, какие разделы молекулярной биологии развивались быстрее всего, мы видим, что это те части, которые относятся к одномерным приспособлениям (например, определение последовательности оснований в нуклеиновой кислоте или аминокислот в белке), или зависят от возможности отделить небольшую часть системы (например, фермент) и изучать ее сравнительно оторванно от всей остальной системы. Те проблемы, которые связаны с множеством одновременных взаимодействий (как, например, предсказание характера укладки полипептидной цепи), мало продвинулись вперед. Такой анализ предвещает мало хорошего проблемам изучения высших отделов нервной системы, относящимся преимущественно к этому последнему типу.
Но один прорыв, совершенный генетикой и начатый Менделем, явился результатом подхода с позиций представления о черном ящике (изучение наследования признаков у растений) и дал сведения о высоком уровне организации (о хромосоме). Поэтому если в исследовании головного мозга действительно произойдет прорыв, то, вероятно, это будет на уровне общего управления системой. Если бы система была такой хаотичной, какой она иногда кажется, мы не могли бы выполнять удовлетворительно даже самые простые задачи. Если взять возможный, хотя и маловероятный, пример, то мощным прорывом явилось бы открытие, что работа мозга производится фазически, каким-то периодическим часовым механизмом, подобно компьютеру.
В настоящем выпуске «Scientific American» показано, что головной мозг успешно изучается под многими углами зрения и что проделана большая увлекательная работа. Только поразмыслив о том, как запутана вся система и как сложны многочисленные разнообразные операции, которые она должна выполнять (в этой статье мы коснулись только некоторых из них), мы понимаем, что перед нами еще долгий путь. Но новые методы дают новые результаты, а новые результаты порождают новые идеи, так что нам не следует падать духом. Нет области науки более жизненно важной для человека, чем исследование его собственного мозга. От нее зависит все наше представление о Вселенной.
Мозг
Ramon у Cajal S. Histologie du systeme nerveux de Phomme et des vertebres, Consejo
Superior de Investigaciones Ceintificas, Institute Ramon у Cajal, 1952. Katz B. Nerve, muscle and synapse, McGraw-Hill Book Company, 1966. [Имеется
перевод: Катц Б. Нерв, мышца и синапс.-М.: Мир, 1968.] Clarke E. S., O'Malley С. В. The human brain and spinal cord, University Of
California Press, 1968.
Нейрон
Huffier S. W., Nicholls J.G. From neuron to brain: A cellular approach to the function of the nervous system, Sinauer Associates, Inc., Publishers, 1976. [Имеется перевод: Куффлер С., Николе Дж. От нейрона к мозгу.-М.: Мир, 1979.]
Steinbach J.H., Stevens С. F. Neuromuscular transmission. In: Frog Neurobiology: A Handbook (R. Llinas and W. Precht, eds.. Springer-Verlag, 1976.
Hille B. Gating in sodium channels of nerve, Annual Review of Physiology, vol. 38, 139-152, 1978.
Stevens Ch.F. Interactions between intrinsic membrane protein and electric field: An approach to studying nerve excitability, Biophysical J., vol. 22, No. 2, 295-306, May, 1978.
Fambrough D.M. Control of acetylcholine receptors in skeletal muscle, Physiological Reviews, vol. 59, No. 1, 165-227, January, 1979.
Малые системы нейронов
Benzer S. Genetic dissection of behavior, Scientific American, vol. 229, No. 6, 24-37,
December, 1973. Nicholls J. G., van Essen D. The nervous system of the leech, Scientific American, vol.
230, No. 1, 38-48, January, 1974. Bentley D., Hoy R.R. The neurobiology of cricket song, Scientific American, vol. 231,
No. 2, 34-44, August, 1974. Kandel E.R. Cellular basis of behavior: An introduction to behavioral neurobiology,
W. H. Freeman and Company, 1976. [Имеется перевод: Кэндел Э. Клеточные
основы поведения.-М.: Мир, 1980.У Kandel E.R. Cellular insights into behavior and learning. In: Harvey Lectures, Series
73, 29-92, 1979.
Организация мозга
Herrick C.J. Neurological foundations of animal behavior, Henry Holt and
Company, 1924. Nauta W. J. H., Karten H. J. A general profile of the vertebrate brain with sidelights
on the ancestry of cerebral cortex. In: The neurosciences: Second study program
(F.O. Schmitt, ed.), Rockefeller University Press, 1970. Neurology. In: Gray's Anatomy. 35th British edition (R. Warwick and P.L. Williams,
eds.), W.B. Saunders Company, 1973.
Развитие мозга
Hunt R.К. Development programming for retinotectal patterns. In: Cell patterning:
Ciba foundation symposium 29, Associated Scientific Publishers, 1975. Rakic P. Cell migration and neuronal ectopias in the brain. In: Birth Defects:
Original articles Series, vol. 11, 95-129, 1975. Hubel D.H., Wiesel T.N., Levay S. Plasticity of ocular dominance columns in monkey
striate cortex. In: Philosophical transactions of the royal society of London, series
B, vol. 278, No. 961, 377^109, April 26, 1977. Cowan W.M. Aspects of neural development. In: International Review of physiology,
vol. 17: Neurophysiology III (R. Porter, ed.), University Park Press, 1978. Jacobson M. Developmental neurobiology, Plenum Press, 1978.
Химия мозга
Hall Z. W., Hildebrand J.G., Kravitz E.A. Chemistry of Synaptic transmission: Essays
and sources, Chiron Press, 1974. Iversen L.L., IversenS.D., Snyder S.H. Handbook of psychopharmacology, Plenym
Press, 1975-1978. Cooper J. R., Bloom F. E., Roth R. H. The biochemical basis of neuropharmacology,
Oxford University Press, 1978.
Hughes J. (ed.). Centrally Acting Peptides, University Park Press, 1978. Iversen S.D., Iversen L.L. Behavioral pharmacology, Oxford University Press, second
edition in press.
Центральные механизмы зрения
Mountcastle V.B. Modality and topographic properties of single neurons of cat's
somatic sensory cortex, J. of Neurophysiology, vol. 20, No. 4, 408-434, July, 1957. Hubel D. H., Wiesel T. N. Receptive fields and functional architecture of monkey
striate cortex, J. of Physiology, vol. 195, No. 2, 215-244, November, 1968. Hubel D.H., Wiesel T.N. Ferrier lecture: Functional architecture of macaque monkey
visual cortex, Proceedings of the Royal Society of London, Series B, vol. 198, 1-59,
1977. Hubel D.H., Wiesel T.N., Stryker M.P. Anatomical demonstration of orientation
columns in macaque monkey, J. of Comparative Neurology, vol. 177, No. 3,
361-379, February 1, 1978.
Механизмы головного мозга, управляющие движением
StelmachG.E. (ed.) Motor control: Issues and Trends, Academic Press, 1976.
Phillips C. G., Porter R. Corticospinal neurones: Their role in movement, Academic Press, 1977.
Kots Ya.M. The organization of voluntary movement: Neurophysiological mechanisms, Plenum Press, 1977.
Granit R. The purposive brain, The MIT Press, 1977.
Miles F. A., Evarts E. V. Concepts of motor organization, Annual Review of Psychology, vol. 30, 327-362, 1979.
Специализация человеческого мозга
Gainotti G. Emotional behavior and hemispheric side of the lesion, Cortex, vol. 8, No.
1, 41-55, March, 1972. Geschwind N. Selected papers on language and the brain, D. Reidel Publishing Co.,
1974.
Gazzaniga M.S., Ledoux J.E. The integrated mind, Plenum Press, 1978. Galaburda A.M. LeMay M., Kemper Th.L., Geschwind N. Right-left asymmetries in
the brain, Science, vol. 199, No. 4311, 852-856, February 24, 1978.
Заболевания человеческого мозга
Penrose L. S. The biology of mental defect, with a preface by J. B. S. Haldane, Grune and Stratton, 1962.
Shyder S. H. Madness and the brain, McGraw-Hill Book Company, 1974.
Ingvar D.H., Lassen N.A. (eds.) Brain work: The coupling of function, metabolism
and blood flow in the brain, Munksgaard, 1975. Kety S.S. The biological roots of mental illness: Their ramifications through cerebral
metabolism, synaptic activity, genetics and the environment, The Harvey Lectures,
Series 71, pp. 1-22, 1978. Kinney O.K., Matthysse S. Genetic transmission of schizophrenia, Annual Review of
Medicine, vol. 29, 459^73, 1978.
Мысли о мозге
Allman J.M., Kaas J.H., Lane R.H., Miezen P.M. A representation of the visual field
in the inferior nucleus of the pulvinar in the owl monkey (Aotus trivirgatus), Brain
Research, vol. 40, No. 2, 291-302, May 26, 1972. Brenner S. The Genetics of caenorhabditis elegans, Genetics, vol. 77, No. 1, 71-94,
January, 1974. Constantine-Paton M., Capranica R. R. Central projection of optic tract from
translocated eyes in the leopard frog (Rana pipiens), Science, vol. 189, No. 4201,
480-482, August 8, 1975.
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении, качестве перевода и другие просим присылать по адресу: 129820, Москва, И-110, ГСП 1-й Рижский пер., д. 2, издательство «Мир».
Мозг
Ст. научи, редактор Е. А. Яновская Мл. научн. редактор О. А. Горгун Художник Е. Ю. Баранская Художественный редактор Л. М. Кузнецова Технический редактор М. А. Страшнова Корректор Т. И. Стифеева
ИБ № 2763
Сдано в набор 15.06.81.
Подписано к печати 24.11.81.
Формат 60 х 90 Vie-
Бумага офсетная № 1.
Гарнитура тайме. Печать офсетная.
Объем 8,75 бум. л.
Усл. печ. л. 17,50. Усл. кр.-отт. 49,24.
УЧ.-ИЗД л. 19,35, Изд. № 4/1394.
Тираж 15000 экз. Зак. 409. Цена 1 р. 70 к.
ИЗДАТЕЛЬСТВО «МИР» Москва, 1-й Рижский Vp.s 2.
Можайский полиграфкомбинат Союзполиграфпрома при Государственном комитете СССР по делам издательств, полиграфии и книжной торговли, г. Можайск, ул. Мира, 93.