Иллюстрация организации памяти мозга
Иллюстрация организации памяти мозга
Со времени публикации была проделана работа по развитию прототипа системы индивидуальной адаптивности Beast (fornit.ru/beast) и появилось много новых данных и материалов с которыми можно ознакомисться в трилогии «Основы фундаментальной теории сознания» и точки входа теории МВАП «Организации механизмов мозга человека».
Цель данной иллюстрации - наглядно показать основные принципы организации памяти мозга с образованием слоев рецепторных детекторов:
1. - совершенно одинаковой био-схемотехники, но обладающей собственной функцией распознавания,
2. - использование одних и тех же признаков для организации на их основе колонок более сложных детекторов, что обеспечивает возможность распознавания любых сочетаний признаков и независимость от повреждений отдельных зон детекторов,
3 - практически мгновенное распознавание даже сложных образов, вплоть до выбора наиболее предпочтительного варианта поведения в данных условиях, определяемого личным жизненным опытом.
Для понимания приведенных схем предварительно необходимо ознакомиться с основными принципами принципами организации мозга (fornit.ru/66411), в частности, иметь представление о реализации адаптивных механизмов распознавания.
Далее предлагается схемотехническая иллюстрация принципов распознавания образов зрительных примитивов.
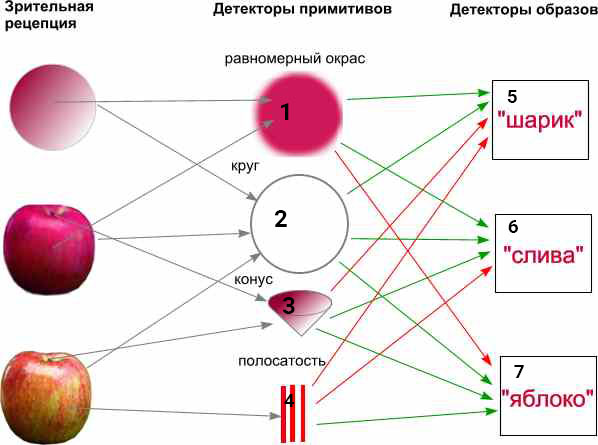
На первой схеме условно (только для понимания принципа за счет упрощения и утрирования) представлен вариант детекции зрительных примитивных фигур с использованием собственной функции распознавания, которые являются рецепторами для более сложных детекторов образов, распознающих объект зрительного восприятия.
Здесь показано то, что каждый предыдущий слой детекторов выполняет рецепторную функцию для последующих слоев и эффекторную функцию - для предыдущих. Т.е. рецепторами для первичного слоя зрительного анализатора являются непосредственно первичные зрительные рецепторы (они претерпевают довольно сложную предварительную обработку уже на уровне сетчатки глаз). Для слоя сложных зрительных образов (шарик, яблоко, слива) рецепторами являются предыдущие детекторы (красные стрелки - тормозные связи, зеленые - возбуждающие).
Если пронумеровать подряд каждый из образов (кроме первой колонки исходных изображений), то видно, что более сложный образ шарика №5 состоит из предшествующих образов 1 и 2. Образ яблока №7 можно раскрыть как совокупность образов 2, 3 и 4.
У каждого образа есть свой уникальный номер в нейросети - который можно присвоить выходу распознавателя данного образа и в дальнейших операциях с образами использовать только эти цифры, не раскрывая (без необходимости уточнения) цифры всех предшествующих образов. Это позволяет оперировать даже с самыми сложными образами (например, детектором бабушки) просто как с уникальными числами - идентификаторами образов (ID). А все иерархия такого принципа распознавания сводится к построению ветвящегося дерева образов, где каждая конечная ветка состоит из ID предыдущих. Такое дерево позволяет очень быстро выбирать нужный образ в определенном контексте. Оно оперирует только целыми положительными цифрами. Так организованы колонки коры мозга (fornit.ru/43642), что объясняет фиксированные количества слоев и эффективность выборки памяти. Так организованы все иерархические структуры контекстов любых условий и признаков (fornit.ru/66797), в том числе так называемая "эпизодическая память" (fornit.ru/67560).
Главным в принципе организации памяти оказываются ID образов, а не то, как именно осуществляется распознавание. В мозге распознающим элементом является однослойный персептрон - именно таким свойством обладает любой (даже вставочный) нейрон (fornit.ru/6449). В искусственной реализации это может быть нейристор (любого удобного вида) или, в программной реализации - структура образа, имеющая свой уникальный ID (fornit.ru/67666), т.е. нейронные сети - не являются принципиально необходимыми для реализации адаптивной системы.
Важно то, что одни и те же признаки используются для самых разных детекторов. Отсутствие такого первичного признака (врожденная патология или отсутствие опыта его формирования в определенный критический период развития) приводят к невосполнимому его отсутствию во всех последующих. Но вовсе не означающему невозможность развития адаптивных структур без него. Даже полное отсутствие зрительного восприятия не препятствует этому за счет других типов рецепторов.
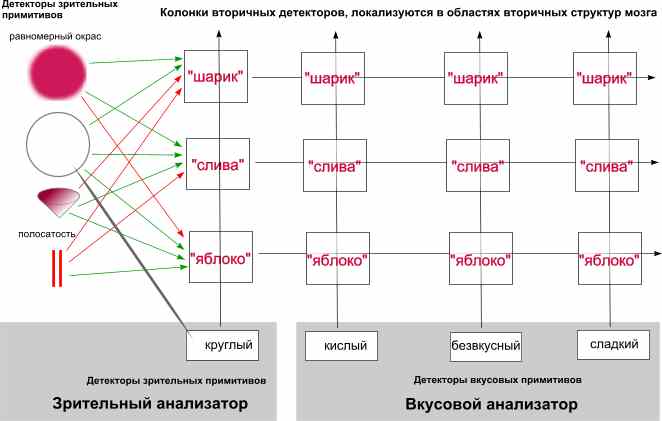
На второй схеме показано как образуются еще более сложные детекторы сочетаний входных признаков, использующие разные типы входных анализаторов (зрительный и вкусовой). При этом, используя одни и те же первичные признаки, образуются колонки детекторов, распознающих более специфичные случаи восприятия, такие как кислое, сладкое или безвкусное яблоко и т.п.
При этом возможно образование локального детектора-символа такого сочетания в виде отдельного элемента нейронной сети (нейрона), настроившего в ходе обучения соответствующую функцию распознавания, или же использование всей совокупности признаков без образования такого детектора.
В первом случае обеспечивается большая степень абстрагируемости от самых первых входных признаков, что обладает всеми преимуществами использования символов вместо всего того, что обозначается такими символами.
Такие детекторы локализуются в определенных (обычно местоположением первичных рецепторных детекторов) зонах мозга. Потеря в результате патологического процесса какого-то из них (например: маленькое, кислое, сморщенное яблочко) или даже целой группы, может практически никак не быть замечены в текущем восприятии вплоть до тех пор, пока условия восприятия не окажутся точно соответствующими потерянному детектору. Но и это не будет критичным просто потому, что вместо символьного детектора будут просто использоваться совокупность обозначаемого им образа, т.е. вместо того, чтобы просто в голове промелькнула мысль о таком-то яблоке, возникнет его полный яркий образ.
Это то, что заставило некоторых исследователей предполагать "голографический" принцип организации памяти мозга. На самом деле ничего общего со свойствами голографии в такой организации памяти нет.
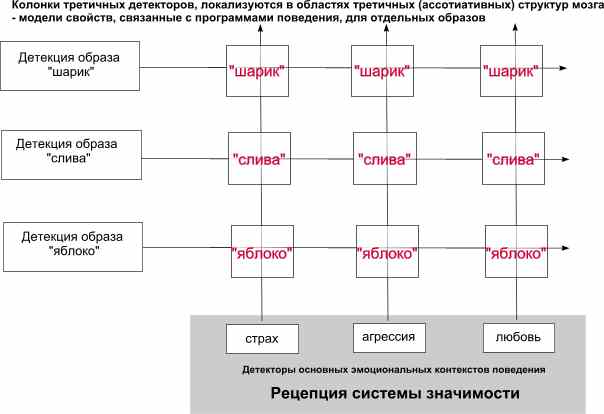
На третьей схеме показаны потенциальные области (страх, агрессия, любовь) для выделения вниманием колонок, содержащих ассоциации со всеми видами рецепторных и эффекторных детекторов, образующие варианты поведенческих программ действий, основанные на моделях свойств детектированных образов (шарик, слива, яблоко). Внимание выделяет (активирует) те из них, которые соответствуют текущему преобладающему эмоциональному контексту. Механизм выбора преобладающего контекста описан в статье Механизм ориентировочного рефлекса.
Каждая из моделей приобретает за счет постоянно обновляемого жизненного опыта, свою специфику реализации поведенческих программ в различных эмоциональных контекстах.
Текущее восприятие активизирует общие для всех моделей признаки, тем самым моментально активизируя и все детекторы, использующие эти признаки. Так же как и признаки наиболее подходящего для текущий условий эмоционального контекста, тем самым очень быстро и эффективно выделяя наиболее подходящую по стилю реагирования группу поведенческих программ.
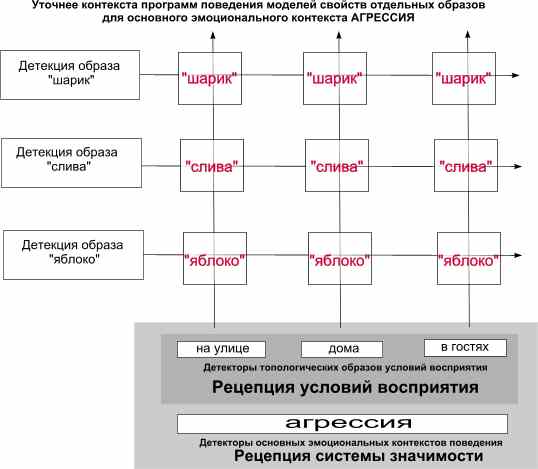
На четвертой схеме показана еще большая конкретизация контекста, в котором выбирается подходящее поведение в определенном эмоциональном контексте (агрессия). Здесь происходят сочетания и с различными вариантами возможных поведенческих реакций (не показаны) с использованием основных детектированных образов - моделей для текущих условий.
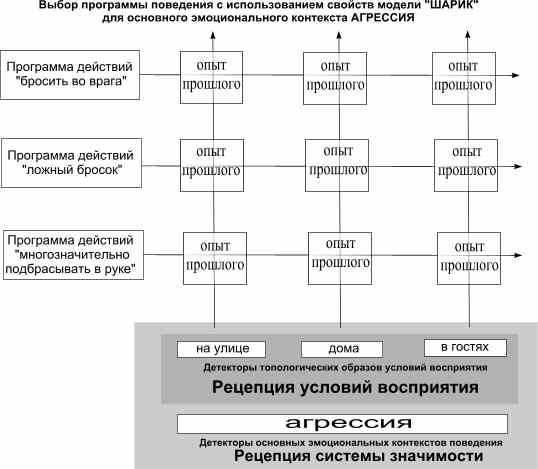
На пятой схеме выбрана только модель "Шарик". Каждый из возможных вариантов ответной реакции, возбужденный признаками ее большей актуальности, вызывает возбуждение и памяти с впечатлением о прошлом опыте такого действия в подобных условиях - эффект прогнозирующего опережающего возбуждения. Положительная оценка такого результата активизирует с вою очередь выбранную поведенческую программу, отрицательная - блокирует ее. Выбранная программа действия, при появлении пускового стимула (после какого-то события возникает пора реагирования), запускает программу действия на исполнение, открывая каналы соответствующих эффекторов (программа разворачивается, получая сигналы о текущей стадии движения и запуская следующую стадию).
Теперь - образное представление того, как, используя одни и те же нейроны в самых разных мыслеобразах, мы оказываемся способны различать их, - на примере зрительных картинок. В течение жизни запоминаются следы неимоверного числа зрительных картинок, в которых принимают участие одни и те же детекторы первичных зрительных примитивов,- картинка не запоминается "в пикселях как на фотографии". Представим, что это - лампочки. Если бы они загорелись все, то изображения перемешались бы неразличимо.
Но есть нечто, что имеет связь к каждым набором таких лампочек в картинке так, что он начинает светиться если пустить ток по этому набору. Если пустить ток по связям другого набора - то засветится другая картинка. Такую схемку очень просто смакетировать (развязав цепи наборов диодами). Роль таких управляющих связей играют признаки остального набора условий, который неизбежно сопутствует любому восприятию: когда, в каком месте и в каком эмоциональном состоянии эта картинка воспринималась и все другие сопуствующих признаки условий. Только при повторении такого сочетания мы сможем ее вспомнить.
Роль места и времени во время вспоминания играет активность детекторов признаков времени и места, которая возникает когда мы мысленно "окажемся там" - т.е. работа активного внимания - воли, которая является следствием преобладания значимости данной активной зоны над значимостями других активных зон в существующих в этот момент условиях (см. Мотивация).
Все рассмотренные примеры очень упрощены и утрированы. Они предполагают, что жизненный опыт уже обладает выбором программ действия, не рассматривается механизм выбора альтернативных программ при произвольном управлении вниманием (осознанный перебор вариантов поведения) и многие другие особенности. Не показывается циклическая активность зон, удерживающих образ восприятия в актуальном состоянии (кратковременная память), не показаны также принципы и условия закрепления следов кратковременной памяти в постоянную систему связей, обеспечивающих все функции распознавания.
Но структурные принципы организации памяти и выделение вниманием актуальных зон, иллюстрируются вполне наглядно.