
СИСТЕМНАЯ ОРГАНИЗАЦИЯ РАБОТЫ МОЗГА ПРИ ОБЕСПЕЧЕНИИ ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ
Системная нейрофизиология (nan)
Показано, что деятельность, определяющую роль в организации которой играют базовые мозговые механизмы и операции, характеризуется соответствием между увеличением активности и функциональных взаимодействий между структурами их обеспечения. При вовлечени.и операции сравнения с рабочей памятью, когда рассогласование предъявленного стимула с ожидаемым приводит к реактивному торможению уже подготовленного действия.
При усложнении деятельности, обусловленном необходимостью подавления подготовленного действия с использованием удерживаемого в памяти правила сочетания управляющих стимулов, показано, что процессы реактивного торможения обеспечиваются на фоне включения в большой набор взаимодействующих звеньев уже упоминавшегося рострального отдела левой веретенообразной извилины. При этом, уровень BOLD сигнала в этой структуре был снижен относительно сравниваемого простого тестового условия, когда в обеспечение деятельности вовлекалась операция сравнения с рабочей памятью.
Представленные варианты сочетаний локальной активности и дистантных взаимодействий демонстрируют, что сам по себе фактор сложности не оказывает специфического, определенного влияния на системную организацию мозговых систем обеспечения целенаправленной деятельности.
Получено подтверждение существования операции сравнения с рабочей памятью в условиях внешнего управления действиями и свидетельство вовлечения этой операции в процессы реактивного торможения подготовленного моторного действия, характеризующееся повышением локальной активности рострального отдела левой веретенообразной извилины и усилением ее функциональных взаимодействий со структурами обеспечения подавления действий.
Вес уверенности:
КИРЕЕВ Максим Владимирович
ДИССЕРТАЦИЯ на соискание ученой степени доктора биологических наук Санкт-Петербург – 2017
Актуальность темы исследования.
Настоящая работа посвящена изучению организации функциональных взаимодействий структур мозга человека, которые вовлекаются в обеспечение целенаправленной деятельности. В данной работе под функциональным взаимодействием понимается факт влияния активности одной структуры мозга, регистрируемой с помощью функциональной МРТ, на активность в другой. Основной целью исследования является изучение характера реорганизации функциональных отношений между структурами мозга, обеспечивающими текущую деятельность разной степени сложности.
Благодаря широкому распространению технологий томографической нейровизиуализации накоплен большой объем экспериментальных данных о функциональной специализации отдельных структур мозга человека относительно видов высшей нервной деятельности. Сейчас, для любой произвольно выбранной области мозга установлены виды деятельности, а также мозговые механизмы и операции, в обеспечение которых такая область мозга может вовлекаться. Уже давно перестали быть редкостью т.н. мета-аналитические работы, в которых в качестве экспериментального материала используются и обрабатываются большие объемы уже опубликованных результатов (см., например, Eickhoff, et al., 2012). Несмотря на весомый вклад таких работ в изучение физиологии мозга, обратной стороной этого явилась неизбежная множественность экспериментальных данных и, как следствие, их противоречивость. Не в последнюю очередь, из-за неопределенности в том, какой именно нейрофизиологический процесс отражается в регистрируемом с помощью функциональных томографических методов сигнале (см., например, Logothetis, 2008). В том числе и поэтому, на основании анализа множества таких работ можно прийти к выводу, что одни и те же структуры мозга, а также их группы, участвуют в обеспечении множества видов высшей психической деятельности. Сложившаяся ситуация затрудняет дальнейшее развитие представлений о том, как именно работает мозг. Ведь физиологическая сущность наблюдаемых локальных изменений активности мозга остается, в общем, малопонятной. Таким образом, доступные современные методы нейровизуализации позволяют обнаруживать структуры мозга, вовлекаемые в обеспечение исследуемой деятельности, но этого недостаточно для понимания их работы. Во многом это связано с тем, что до сих пор малоизвестно, как именно организованы взаимодействия между структурами мозга и каковы основные характеристики их перестроек при изменении исследуемой деятельности или условий ее реализации. Поэтому решение этой проблемы связано с целенаправленным изучением организации сочетанной работы функционально связанных структур мозга, которые вовлекаются в обеспечение исследуемой деятельности.
Несмотря на то, что методы по изучению функциональных взаимодействий структур мозга с использованием данных, в основном получаемых методом функциональной МРТ, принципиально позволяют исследовать организацию работы мозговых систем в масштабе всего мозга (Friston, et al., 1997, Friston, 2010), такие исследования имеют узконаправленный характер, и их пока что слишком мало для выявления общих закономерностей. Кроме того, руководящей логикой таких исследований является принцип активационных исследований, когда повышение регистрируемой активности рассматривается как отражение относительно большего вовлечения выявленной структуры мозга в обеспечение исследуемого механизма или операции. Аналогичным образом организованы экспериментальные исследования функциональных взаимоотношений, в которых проверка гипотез о вовлечении исследуемых операций и механизмов построена на той же логике. В рамках такого «активационно-ресурсного» подхода подразумевается, что факт относительно большего вовлечения изучаемого механизма доказывается демонстрацией усиления функциональных взаимодействий между вовлекаемыми структурами мозга. На этом основании можно ожидать, что, например, усложнение деятельности на фоне относительного увеличения локальной активности будет сопровождаться усилением взаимодействий со структурами, связанными с обеспечением когнитивного контроля и управлением поведения (как правило, со структурами префронтальной коры). Однако, такой подход не учитывает сложившихся представлений о свойствах организации мозговых систем, которые были выявлены при изучении активности мозга на уровне отдельных нейронных популяций. Так, в ходе целенаправленных исследований системной организации мозга человека Н.П. Бехтеревой и ее сотрудниками были выявлены ключевые принципы работы нейрональных систем. В ходе исследований по изучению активности нейронов с помощью долгосрочных имплантируемых электродов при специализированной методической ориентации именно на вскрытие системной организации мозга было показано, что нейрональные системы состоят из корко-подкорковых звеньев разной степени жесткости (Бехтерева, 1966, Бехтерева, 1974). При этом подчеркивалось, что характеристика того, насколько часто данная структура мозга выступает в качестве участника группы взаимодействующих звеньев, отражает его жесткость или гибкость относительно данной нейрональной системы (т.е. относительно частое или редкое вовлечение соответственно). Показано, что жесткость звена может проявляться и как привязанность к определенной функции, и как участие конкретной области мозга в широком спектре исследуемых видов деятельности. Было установлено, что нейроны ведут себя индивидуально и принципиально полифункциональны (т.е. не закреплены за какой-то одной функцией или видом поведения). Дальнейшие исследования раскрыли важные свойства динамической организации мозговых систем, которые подтвердили концепцию Н.П. Бехтеревой о корково-подкорковых нейрональных системах, состоящих из звеньев различной жесткости (Медведев, 1987, Медведев, Пахомов, 1989). Было показано, что функциональная роль звена системы может меняться не только при переходе от одной деятельности к другой, но и при повторном выполнении одной и той же деятельности (Пахомов, 1983, Гоголицин, и др., 1987). При этом, обеспечивающая решение поведенческой задачи мозговая система в разные моменты времени могла быть разной и по своему звеньевому составу, и по характеру связей между этими звеньями (Медведев, Пахомов, 1989). Данный принцип был справедлив не только для сложноорганизованных, но и для относительно стереотипизированных видов деятельности. Выявленные свойства динамической организации нейрональных систем мозга указывают на принципиальную несводимость внешне одинакового результата деятельности к работе какой-то одной системы. Это соответствует принципу «повторение без повторения», сформулированному в работах Н.А. Бернштейна, в соответствии с которым внешне одинаковые движения в терминах биомеханики их нервно-мышечного обеспечения никогда не повторяются (Бернштейн, 1990). Сформулированные принципы не противоречили и представлениям теории функциональных систем мозга П.К. Анохина (Анохин, 1975).
Работы этого направления доказали перспективность изучения взаимосвязей в активности дистантно расположенных областей мозга (Медведев, Пахомов, 1989). Полученные данные указывали на недостаточность изучения только локальных перестроек активности в звеньях мозговых систем, вовлекаемых в обеспечение исследуемой деятельности, поскольку совпадение формы локальных реакций не всегда означало их функциональную связь. И наоборот, функциональные взаимодействия могли фиксироваться между дистантно расположенными популяциями нейронов без регистрируемых в них локальных реакций. А продемонстрированная, даже в условиях относительно монотонной деятельности, динамичность их образований легла в основу концепции об обеспечении деятельности «распределенной в пространстве и принципиально динамичной во времени системой» (Медведев, 1987), которая отражает фундаментальное свойство работы мозга.
В описанных выше исследованиях, явившихся важными этапами на пути раскрытия принципов функционирования мозга, в основном исследовались популяции нейронов только тех структур мозга, долгосрочная имплантация электродов в которые строго соответствовала клиническим задачам (Бехтерева, 1988, Медведев, Пахомов, 1989, Пахомов, 1983, Гоголицин, и др., 1987). Во-первых, по сравнению с объемом всего мозга их количество было чрезвычайно мало, т.е. изучался лишь фрагмент работающей системы. Во-вторых, до сих пор оставался невыясненным вопрос о том, можно ли указанные закономерности динамической организации системной активности нейронных популяций (микрозвеньев) применять в качестве принципа рассмотрения организации нейрональных систем в масштабе мозга, т.е. когда в качестве звеньев такой системы выступают отдельные структуры мозга. Однако, целенаправленных исследований этого вопроса не проводилось. Хотя это является высокоактуальной проблемой, поскольку функциональная диагностика с применением современных методов медицинской томографической нейровизуализации выявляет именно локальные интегральные энергетические и биохимическое показатели состояния мозга. С учетом вышесказанного очевидно, что ограничиваясь только анализом изменений локальных процессов вовлекаемых структур мозга, сложно полноценно определить влияние отклонений в их функционировании на организацию системной работы мозга при его заболеваниях. В результате, такое положение дел ограничивает информативность и эффективность современных диагностических методов томографической нейровизуализации: активный поиск объективных нейромаркеров психических и неврологических расстройств ведется до сих пор (см. например, Kropotov, et al., 2016). Это означает, что решение высокоактуального вопроса оптимизации существующих и разработки новых методов лечения и диагностики заболеваний мозга тесно связано с развитием представлений об основных характеристиках взаимодействий мозговых структур.
Именно поэтому настоящая работа была направлена на сочетанное рассмотрение показателей изменений локальной активности и дистантных взаимодействий структур мозга, вовлекаемых в обеспечение целенаправленного поведения.
Цели и задачи исследования. Основной целью настоящей работы является выявление характера организации функциональных взаимоотношений между структурами мозга человека, вовлекаемыми в обеспечение целенаправленной деятельности. Для этой цели исследовались виды деятельности разной степени сложности в терминах цели и способов ее достижения. В соответствии с целью исследования были поставлены следующие основные задачи:
1. Выбрать алгоритм обработки данных функциональной томографии, который позволил бы выявлять состав, структуру и характер функциональных взаимодействий между звеньями нейрональных мозговых систем обеспечения целенаправленной деятельности.
2. Определить структуры мозга, являющиеся звеньями нейрональных систем обеспечения целенаправленной деятельности, на примере набора тестовых заданий моделирующих ее усложнение: 1) управление действиями в условиях соответствия управляющих зрительных стимулов (двухстимульный Go/NoGo тест) и вовлечения базового мозгового механизма рабочей памяти, 2) управление действиями в условиях произвольного принятия решения о действии при сознательной лжи, 3) вербальная деятельность по генерации форм глаголов разных классов, соответствующих ментальным грамматическим правилам порождения разной степени стереотипизацииавтоматизации.
3. Определить структуру пространственной организации и характер функциональных отношений обнаруженных звеньев нейрональных систем обеспечения целенаправленной деятельности, а также их реорганизацию при ее усложнении.
4. Оценить соотношение между интегральными показателями локальной активности звеньев, их «энергетического» состояния и характером их вовлечения в систему взаимодействующих элементов нейрональных систем, в зависимости от сложности целенаправленной деятельности. Выяснить вопрос о существовании структур мозга, не видимых в активационных исследованиях, которые проявляют свое участие в обеспечении текущей деятельности только путем изменения функциональных отношений с другими звеньями. Экспериментально проверить характер соответствия между изменениями показателей активности структур мозга и их дистантных взаимодействий, предсказываемых «активационно-ресурсным» подходом рассмотрения организации мозгового обеспечения исследуемой деятельности.
Методология и методы исследования. Для решения задач настоящего исследования были подобраны три варианта экспериментальных тестовых заданий, которые моделировали условия усложнения деятельности в терминах цели деятельности и способов ее достижения. Первым вариантом тестового задания было выбрано двухстимульное Go/NоGo задание, при котором на основе соответствия последовательно предъявляемых зрительных стимулов планируемое действие либо реализуется, либо подавляется. Второй вариант тестового задания, который моделировал ситуацию самостоятельного принятия решения о типе действия, подразумевал чередование испытуемым сознательных ложных и правдивых действий в условиях интерактивной игры с компьютером. В качестве третьего, наиболее сложного, варианта тестового задания была выбрана исследовательская парадигма по порождению глаголов разных классов, которые отличаются в терминах морфологических свойств порождения и частотности глагольных классов. В работе был использован исследовательский подход, при котором результаты анализа локальной активности структур мозга, которые вовлекаются в обеспечение исследуемой деятельности, сопоставлялись с данным о характере их дистантных взаимодействий. Стандартный для активационных исследований подход на основе методов множественной регрессии выявлял структуры мозга уровень нейрональной активности которой отражает ее вовлечение в исследуемой деятельности. Для обнаруженных таким образом структур мозга с помощью метода психофизиологических взаимодействий (Friston, et al., 1997) проводился анализ по выявлению их дистантных взаимодействий.
Научная новизна исследования. В работе, впервые на материале серии исследований, моделирующих усложнение реализуемой деятельности, изучен вопрос о характере изменений функциональных отношений между вовлекаемыми структурами мозга человека. Установлено, что широко распространённый в функциональной томографической нейровизуализации подход, в соответствии с которым при увеличении нейрональной активности структуры мозга судят о более активном ее вовлечении в обеспечение текущей деятельности, что, как следствие, предполагает усиление ее взаимодействий с другими звеньями вовлекаемой системы, не является единственным возможным: продемонстрированы и другие варианты соотношений показателей локальной активности и дистантных взаимодействий мозговых зон. Полученные новые данные свидетельствуют об универсальности сложившихся представлений о динамической организации мозговых систем, которые были сформулированы при изучении нейронных популяций, когда динамические перестройки дистантных взаимодействий наблюдались без значимых изменений их импульсной активности. Выявленные сценарии соотношений локальной активности и дистантных взаимодействий вовлекаемых мозговых структур расширяют представления о системной организации мозговых систем обеспечения целенаправленной деятельности. В результате предложенного исследовательского подхода по сочетанному изучению локальной активности и дистантных взаимодействий впервые удалось продемонстрировать организацию взаимовлияний между вовлекаемыми структурами мозга при обеспечении деятельности, ключевую роль в организации которой играет один из базовых механизмов - механизм детекции ошибок. Получены новые данные, демонстрирующие разную функциональную специализацию звеньев лобно-височной системы обеспечения речевой деятельности в зависимости от особенностей процессов порождения элементов речи, раскрывающие представления о мозговой организации ментального лексикона.
Впервые показано, что снижение BOLD-сигнала, отражающего уровень нейрональной активности, не обязательно отражает выключение данной структуры из системы взаимодействующих звеньев вовлекаемых нейрональных систем. В определенных условиях, на фоне снижения показателей нейрональной активности может наблюдаться увеличение функциональной роли дистантных взаимодействий между работающими структурами мозга. Равно, как и увеличение локальной активности вовлекаемой структуры мозга может сопровождаться ослаблением ее дистантных взаимодействий.
Полученные результаты демонстрируют одну из причин отмечаемых противоречий в накопленных данных функциональной специализации отдельных структур мозга. Использование отработанного в ходе настоящей работы методического подхода к изучению механизмов работы мозга с помощью фМРТ позволит преодолеть выявленные ограничения стандартных подходов, что имеет важное значение для клинической диагностики.
Теоретическая и практическая значимость работы. Полученные в настоящей работе результаты развивают современные представления об организации мозговых систем обеспечения поведения. Представленные в работе новые экспериментальные данные и установленные закономерности могут быть использованы для дальнейшего развития исследований функциональной организации мозга. Предложенный и апробированный исследовательский подход позволяет расширить возможности методов функциональной томографической нейровизуализации.
Продемонстрированная эффективность комплекса использованных методов по изучению функциональных взаимодействий между структурами мозга, вовлекаемыми в обеспечение исследуемой деятельности, обуславливает его востребованность для клинической функциональной диагностики. Некоторые из методов, которые применялись в рамках решения задач данной работы, уже в настоящий момент рутинно применяются в предхирургической диагностике пациентов, целью которой является визуализация функционально-значимых областей мозга. Полученные экспериментальные данные, раскрывающие физиологическую сущность изменений локальной активности, наблюдаемых в рамках рутинно используемых методов обработки фМРТ данных, могут быть полезны для оптимизации существующих методов функциональной диагностики, разработки диагностических критериев сохранности функций или нейромаркеров психических расстройств.
Основные положения, выносимые на защиту.
1. Предложенный исследовательский подход позволил установить относительную независимость организации функциональных отношений между вовлекаемыми структурами мозга от уровня их функциональной активности. Таким образом, на общемозговом уровне рассмотрения организации нейрональных систем продемонстрирована универсальность принципа динамической организации мозговых систем, выявленного ранее при анализе импульсной активности популяций нейронов.
2. Участие базовых мозговых механизмов в обеспечении поведения в качестве определяющего элемента характеризуется сочетанными изменениями уровня нейрональной активности и показателей функциональных взаимодействий структур мозга.
3. Обнаруженная вариативность соотношений уровней нейрональной активности структур мозга и их дистантных взаимодействий отражает изменчивость функциональной специализации отдельных звеньев нейрональных систем.
4. Характер реорганизации функциональных взаимодействий между вовлекаемыми в обеспечение целенаправленной деятельности структурами мозга определяется не сложностью реализации деятельности как таковой, а способом достижения ее цели.
Степень достоверности и апробация результатов. Все результаты, полученные в ходе реализации проекта, соответствуют требованиям, предъявляемым к статистической обработке данных функциональной томографии, и были достигнуты с использованием специализированных программных средств. Достоверность выявляемых изменений функциональной активности мозга оценивалась с помощью дисперсионного анализа, с обязательной коррекцией на множественность сравнений, и методами Баейсовской статистики. Материалы диссертационной работы докладывались в виде устных и стендовых докладов на международных и отечественных конференциях: Organization of Human Brain Mapping Meeting «OHBM 2009» (Сан-Франциско, США, 2009 г.), «OHBM 2010» (Барселона Испания, 2010), Society for Psychophysiological Research 49th Annual Meeting (Берлин, Германия, 2009), World Congress of Psychophysiology (Будапешт, Венгрия, 2010; Пиза, Италия, 2012, Хиросима, Япония, 2014, Гавана, Куба, 2016), 7th FENS Forum (Амстердам, Нидерланды, 2010), Четвёртая международная конференция по когнитивной науке (Томск, Россия, 2010), 11th International Conference on Cognitive Neuroscience (Майорка, Испания, 2011), XVIII Всероссийская конференция «Нейроиммунология. Рассеянный склероз» (Санкт-Петербург, 2011), Всероссийская юбилейная научно-практическая конференция «Актуальные проблемы военной психиатрии» (Санкт-Петербург, 2011), Всероссийская молодежная конференция-школа «Нейробиология интегративных функций мозга» (Санкт-Петербург, 2011; 2013), Всероссийский национальный конгресс по лучевой диагностике и терапии «Радиология 2012» (Москва, 2012), «Поленовские чтения-2013» (Санкт-Петербург, 2013), 11th Symposium of Psycholinguistics (Тенерифе, Испания, 2013), 8th International Morphological Processing Conference (Лондон, Великобритания, 2013), Международный симпозиум по нейролингвистике «Night Whites 2014» (Санкт-Петербург, 2014), BNA2015: Festival of Neuroscience (Эдинбург, Великобритания, 2015), Съезд физиологов СНГ (Ялта, Украина, 2011, Сочи, 2016). Исследования, проводившиеся в рамках данной работы, осуществлялись при поддержке отечественных научных фондов (РГНФ № 10-06- 00726, № 12-06-00706, № 14-06-00915, РФФИ № 12-04-31586, РНФ № 16-18-00040, № 16-18-00041).
Публикации. По материалам диссертации опубликовано 33 печатных работы из них 12 статей в научных изданиях, рекомендованных ВАК Министерства образования и науки РФ.
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, описания методов исследования, трех глав с изложением результатов и их обсуждением, заключения, выводов и списка литературы, включающего 35 отечественных и 327 иностранных источников. Работа изложена на 304 страницах и содержит 19 таблиц и 53 рисунка.
Личный вклад автора в проведенное исследование. Автор предложил основную идею и цель исследования, разработал дизайны исследований, лично провел исследования и осуществил подбор методов по анализу полученных данных. Автором был предложен исследовательский подход, сочетающий современные методы обработки нейровизуализационных данных, выполнен их статистический анализ и оформление полученных результатов.
СОДЕРЖАНИЕ РАБОТЫ Объект и методы исследования. Все исследования было одобрены Комитетом по Этике Федерального государственного бюджетного учреждения науки Института мозга человека им. Н.П. Бехтеревой Российской академии наук. Все участники исследования подписывали информированное согласие на участие в исследовании. В серии состоящей из трех исследований, проводимых в условиях двухстимульного тестового задания Go/NoGo, сознательной лжи и порождения элементов речи, приняло участие 19 (15 женщин и 4 мужчин от 20-32 лет), 24 (14 женщин и 10 мужчин, от 19-44 лет) и 21 (13 женщин и 8 мужчин, от 19-32 лет) здоровых праворуких испытуемых соответственно. Праворукость оценивалась по вопроснику Олдфилда (Oldfield, 1971).
Исследование проводилось на магнитном томографе Philips Achieva, с напряженностью поля 3 Тесла. Структурные T1-взвешенные изображения регистрировались до проведения функционального исследования (T1W3DTFE), со следующими параметрами: поле обзора – 240×240 (FOV); TR – 25 мс.; срезы – 130 аксиальных срезов толщиной 1 мм и размером пикселя размером 1×1 мм.; угол отклонения вектора намагниченности – 30°.
Для оценки уровня нейрональной активности мозга осуществлялась регистрация BOLD-сигнала (blood oxygen level dependent, т.е. сигнала зависимого от уровня насыщения крови кислородом). При функциональном МРТ-сканировании (фМРТ) использовалась эхопланарная одноимпульсная последовательность. Время, за которое происходила регистрация данных со всех 32-х аксиальных срезов (TR), составляло 2 секунды (TE=35 мс.). Поле обзора составляло 208×208, а угол отклонения вектора намагниченности (flip angle) – 90°. Размер пикселя составлял 3×3 мм. Толщина срезов равнялась 3 мм., с промежутком между ними в 0.3 мм. Таким образом, после трехмерной реконструкции изображения, размер единицы объема (воксел, от volume cell) составлял 3×3×3 мм. Каждое фМРТ исследование состояло из нескольких исследовательских сессий, в которых перед запуском регистрации BOLD- сигнала и началом предъявления тестового задания, которые были синхронизированы, выполнялись две так называемые холостые «динамические сканы». Под динамическим сканом понимается BOLD-сигнал, зарегистрированный в 32 срезах за 2 сек (TR). Дополнительно 2 первых динамических скана удалялись из последующего анализа. Данная процедура является стандартным способом избегания, так называемого T1- эффекта, который привносит артефакты в фМРТ данные.
Во всех исследованиях испытуемые находились в лежачем положении и для предъявления стимулов, использовался специальный монитор в составе комплекса для проведения фМРТ исследований (Invivo Eloquence fMRI System), располагавшийся за головой испытуемого. Изображение с монитора проецировалось испытуемому с помощью системы встроенных зеркал. Программирование последовательности предъявления проб, а также всех временных параметров презентации стимулов, а также запись поведенческих параметров действий испытуемых (время реакции и типы действий), осуществлялось на базе программного пакета E-prime 1.1 (Psychology Software Tools Inc., Pittsburgh, PA, USA), совмещенного с исследовательским комплексом Invivo. В фМРТ-исследовании мозгового обеспечения порождения элементов речи дополнительно использовался специальный МРТ-совместимый микрофон (Persaio MRI Noise Cancellation System (Psychology Software Tools, Inc.)), который позволял запись и последующую фильтрацию речи испытуемых на фоне шума от сканирования.
Обработка фМРТ данных проводилась по единой схеме проводилась в два этапа в ходе которых осуществлялась предварительная подготовка данных и их статистический анализ по выявлению локальных изменений BOLD-сигнала (отражающего уровень нейрональной активности) и психофизиологических взаимодействий выявленных структур мозга. При предварительной обработке получаемые индивидуальные данные подвергались следующим преобразованиям: пространственное выравнивание (realignment) для коррекции смещения положения головы относительно первоначального положения, пространственное совмещение функциональных данных (BOLD-изображений) со структурными изображениями (coregistration), сегментация индивидуальных анатомических изображений на паттерны серого и белого вещества головного мозга, расчет параметров пространственной нормализации в стандартное анатомическое пространство и применение этих параметров ко всем функциональным изображениями и пространственное сглаживание (параметры сглаживания – 8 мм. (FWHM)) с применением функции Гаусса (Friston et al., 2007). Предварительная обработка данных и построение статистических параметрических карт BOLD-сигнала проводились в программном пакете SPM8 (http://www.fil.ion.ucl.ac.uk/spm/software/spm8/). При процедуре выравнивания рассчитывалось 6 параметров, соответствующих смещению и вращению относительно начального положения головы по трем осям (x,y,z). Для обеспечения учета изменений сигнала коррелированных с движениями головы, эти параметры учитывались при статистическом анализе (Johnostone et al., 2006).
Статистический анализ проводился с использованием основной линейной модели (ОЛМ (Friston, et al., 2007)), в которой в качестве регрессоров (компонентов) использовались временные параметры предъявления стимулов или осуществляемых действий испытуемых. Дополнительно, для учета влияния артефактов движения, в ОЛМ вводились параметры смещений положения головы, рассчитанные на этапе выравнивания функциональных данных. Задача вычисления ОЛМ сводится к тому, чтобы так подобрать статистические параметры ее компонентов, чтобы ОЛМ наилучшим образом соответствовала зарегистрированному в действительности BOLD- сигналу. В данном исследовании такой анализ проводился для каждого элемента фМРТ-изображения (вокселя). При этом одним компонентом ОЛМ моделируется не одиночное событие в заданном интервале времени (временном окне), а сразу вся совокупность исследовательских проб конкретного типа. При вычислении т.н.
статистических контрастов, т.е. относительной разницы в BOLD-сигнале между двумя пробами разного типа, вычисляется линейная комбинация (разница) между рассчитанными параметрами регрессии соответствующих компонентов ОЛМ. На первом этапе (first level analysis) на основе вычисленных параметров регрессии линейной модели рассчитывались t-контрасты между пробами (компонентами ОЛМ), для каждого испытуемого в отдельности. За исключением специально оговариваемых случаев, рассчитывались t-контрасты типа «Экспериментальная проба» > состояние покоя). Полученные контрасты, представляющие из себя линейную комбинацию параметров ОЛМ, подвергались дальнейшему статистическому анализу на втором этапе (second level analysis), который осуществлялся с учетом межсубъектной вариабельности (т.н. random effect analysis) и выполнялся с помощью дисперсионного анализа (Friston, et al., 2007). Иными словами, на групповом этапе анализа данных в качестве переменных использовались вычисляемые на первом этапе статистические параметр (именно поэтому анализ томографических данных называют статистическим параметрическим картированием). Структуры мозга, выявляемые на этой стадии статистического анализа использовались в качестве областей интереса (ОИ, с радиусом 4 мм.) для анализа психофизиологических взаимодействий (psychophysiological interaction, PPI-анализ (Friston, et al., 1997). Предлагаемый метод позволяет получать информацию об изменении функциональной связанности между областями мозга, вовлекаемыми в обеспечение исследуемой деятельности (Gitelman, et al., 2003; Friston, et al., 2011; Ciscler, et al., 2013), в зависимости от типа реализуемой деятельности. Применение этого метода направлено на выявление тех областей мозга, взаимодействие которых с заранее выбранной областью интереса модулируется текущей деятельностью. В основе метода лежит предположение, что психофизиологические взаимодействия происходят на нейрональном уровне (Gitelman et al., 2003). Поэтому, сначала значение BOLD сигнала в ОИ подвергается обратной свертке, процедуре обратной той, которая осуществляется при создании ОЛМ, т.е. свертка функции гемодинамического ответа и вектора, описывающего наступление значимых событий. В результате создается модель нейрональной активности в выбранной ОИ (см. рисунок 1). При этом, каждому типу экспериментальных проб соответствует свой вектор. Далее, полученные данные используются для создания ОЛM, состоящие из PPI-регрессоров, каждый из которых создается путем конволюции смоделированной таким образом нейрональной активности и стандартной формы гемодинамического ответа. В качестве игнорируемых переменных дополнитeльно в модель вводятся регрессоры, моделирующие изменения BOLD сигнала, используемые при стандартном анализе. В результате, вычисление параметров регрессии в такой модели позволяет выявить области мозга, в которых проявляется статистическая зависимость от изменений нейрональной активности в ОИ, связанных с текущей деятельностью. Развернутое математическое описание метода представлено в нескольких относительно недавних публикациях (Cisler, et al., 2014, Friston, et al., 1997, Gitelman, 2003, McLaren, 2012).

Рисунок 1. Схема PPI-анализа (адаптировано из (Cisler, et al., 2014)).
Обозначения: разным цветом обозначены времена предъявления стимулов или анализируемые события двух вариантов тестового задания, Y обозначается реально зарегистрированный BOLD-сигнал в области Б, XA обозначает реально зарегистрированный сигнал в области A, gPPI обозначение использованного в настоящей работе программного пакета (generalized psychophysiological interaction (McLaren, et al., 2012)).
Для того чтобы избежать ложно-положительных результатов, повоксельное построение статистических карт, отражающих как изменения BOLD-сигнала, так и параметров психофизиологических взаимодействий осуществлялось с порогом p<0.05, корректированным на множественность сравнений по методу FWE (Friston, et al., 2007, Nichols, Hayasaka, 2003). При этом учитывались только кластеры размером больше 20 вокселей. Для определения локализации выявленных кластеров в терминах полей Бродманна, использовался программный пакет Anatomy (Eickhoff et al., 2005)).
Тестовые задания.
Исследование организации нейрональных систем обеспечения целенаправленной деятельности в условиях вовлечения механизмов проактивного и реактивного когнитивного контроля на примере двухстимульного тестового задания Go/NoGo. Первый вариант тестового задания использовался для исследования организации нейрональных систем обеспечения целенаправленной деятельности в условиях вовлечения механизмов проактивного и реактивного когнитивного контроля, которое проводилось с использованием двухстимульного тестового задания Go/NoGo. Испытуемым последовательно предъявлялась пара стимулов (С1-С2, т.е. Стимул1-Стимул2), а предварительная
инструкция устанавливала при каком сочетании испытуемый должен действовать или, наоборот, не нажимать кнопку. Схематическое изображение тестового задания представлено на рисунке 2. В качестве стимулов использовались цветные изображения животных (A) или растений (P), которые предъявлялись на экране монитора в течение 100 мс. Межстимульный интервал между инструктирующим сигнальными стимулом и целевым, т.е. между C1 и C2, всегда составлял 1000 мс. Во всех вариантах исследования, если в качестве С1 предъявлялось изображение животного, то испытуемый готовился к появлению С2 и действовал согласно инструкции (80 проб). Если в качестве С1 предъявлялось изображения растения, то испытуемый ожидал появления С2, но заведомо знал, что вне зависимости от содержания С2 нажимать в этой пробе не требуется (80 проб). В зависимости от варианта инструкций различались следующие типы проб, которые затем учитывались при последующем статистическом анализе: 1) нажатие кнопки на второй стимул пары С1-С2 «животное-животное» - AcueAGо (40 проб); 2) подавление нажатия кнопки на второй стимул пары С1-С2
«животное-растение» - AcuePNоGо (40 проб); 3) нажатие кнопки на второй стимул пары С1-С2 «животное-растение» - AcuePGо (40 проб); 4) подавление нажатия кнопки на второй стимул пары С1-С2 «животное-животное» - AcueANоGо (40 проб); 5) отсутствие подготавливаемого действия PcueA (80 проб) и PcueP (80 проб).
Таким образом, моделировалась ситуация, которая подразумевала: 1) в
«Исследовании1» формирование модели сенсорного стимула и модели моторного ответа (sensory/motor preparatory set), при рассогласовании с которой запускается процесс подавления действий; 2) в «Исследовании2» такой модели не формировалось, поскольку ожидался не конкретное изображение, а какой-либо стимул из категории
«растение», а выбор типа действия осуществлялся с учетом правила сочетания стимулов («животное-растение»). В исследовании2 моделировалось относительно большее вовлечение процессов когнитивного контроля. Помимо этого, в каждом исследовании предъявлялись пробы с чередованием стимулов типа PA, PP, в которых предъявление в качестве C1 изображения растения инструктировало испытуемого дожидаться C2 без каких-либо действий после его предъявления.
Статистическая модель для анализа фМРТ-данных каждого испытуемого включала регрессоры соответствующие стимулам (С1-С2) экспериментальных проб и двух экспериментальным сессиям: 1) AcueAGo, AcuePNogo, PcueA, PcueP, AcueAtarget (в качестве C1-стимула предъявляется животное и формировалось ожидание этого же изображения животного), PcueExp1 (в качества С1-стимула предъявляется изображение растения), пробы с ошибками; 2) AcuePGo, AcueANoGo, PcueA, PcueP, PcueExp2 (C1-стимул), AcuePtarget (в качестве C1-стимула предъявляется животное, и формировалось ожидание изображения принадлежащего к категории растение), пробы с ошибками. При групповом анализе в качестве переменных использовались индивидуально рассчитываемые t-контрасты между всеми типами проб и состоянием покоя (П). В «Исследовании1»: AcueAGo>П, AcuePNogo>П, PcueA>П, PcueP>П,
AcueAtarget>П, PcueExp1>П, Ошибка>П. В «Исследовании 2»: AcueANoGo>П, AcuePGo>П, PcueA>П, PcueP>П, AcuePtarget>П, PcueExp2>П, Ошибка> П.
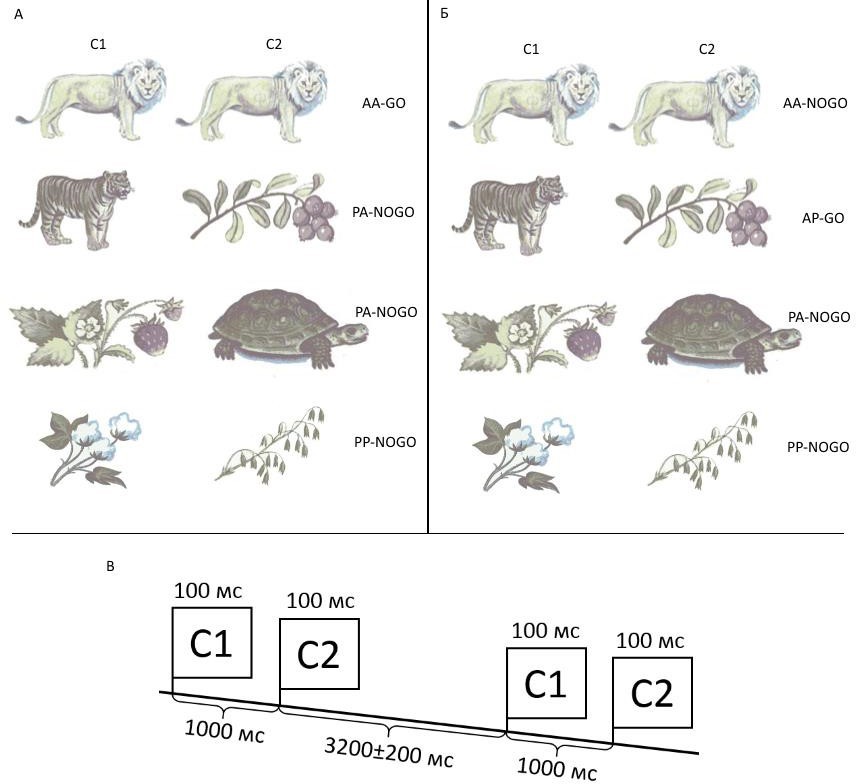
Рисунок 2. Дизайн двухстимульного Go/NoGo тестового задания.
Обозначения: А - варианты последовательностей предъявляемых стимулов и инструкции к ним в «исследовании 1», Б - варианты последовательностей стимулов и инструкции к ним в «исследовании 2», B - схема предъявления стимулов. После появления стимула С2 испытуемый должен был либо действовать (пробы в названии которых слово «Gо»), либо не нажимать конку (пробы в названии которых слово
«NоGо»). Обозначения: мс – миллисекунды, А – изображение животного, Р – изображение растения, С1 – первый стимул пробы (cue), С2 – второй стимул пробы (target), Gо – пробы с инструкцией нажимать кнопку, NоGо – пробы с инструкцией не нажимать кнопку.
Исследование роли базового механизма детекции ошибок в организации нейрональных систем обеспечения сознательных ложных и правдивых действий. Второй вариант тестового задания используемого в настоящей работе, в отличие от Go/NoGo теста, подразумевал самостоятельное принятие решение о том, как именно действовать. Исследования мозгового обеспечения произвольной целенаправленной деятельности в условиях реализации сознательных ложных и правдивых действий проводилось в условиях, при которых испытуемый манипулировал действиями оппонента (компьютера), самостоятельно и сознательно принимая решение лгать или нет в условиях интерактивной игры с компьютером, которой был реализован принцип карточной игры «Верю – не верю» (см. Рисунок 3). В каждой из них предъявлялось по 90 проб. При этом 30 из них были контрольными (К), а остальные 60 – игровыми (т.е.
испытуемый либо лгал (Л), либо был правдив (Пр)). Последовательности контрольных и игровых проб не повторялись и были рандомизированы для каждого испытуемого. Каждая проба состояла из целевого стимула (стрелки), действия испытуемого (манипулятивная правда, сознательная ложь и правдивое нажатие по инструкции в К- пробах) и двух стимулов обратной связи. В самом начале Л и Пр -проб в течение 500 мс. на черном фоне предъявлялась стрелка сплошного белого цвета. Испытуемый должен был принимать решение о том, солгать компьютеру о направлении стрелки или нет, и нажать соответствующую кнопку манипулятора, предназначенного для работы в условиях сильного магнитного поля (Invivo button response unit). На это отводилось
4.5 сек., по истечении этого времени следовала реакция компьютера на предложение испытуемого (правдивое или ложное нажатие) – слова «согласен» или «не согласен». Слово предъявлялось на 500 мс. и сменялось вторым стимулом обратной связи такой же длительности. Если компьютер соглашался с ложным предложением испытуемого, т.е. слово «согласен» появлялось после того, когда испытуемый нажимал кнопку мыши, которая не соответствовала направлению стрелки – это считалось выигрышем и сопровождалось появлением чисел « 2» или « 5», отображающих размер суммы, добавляемой к вознаграждению за участие в исследовании. Выигрышем считалась и ситуация, при которой компьютер не был согласен с правдивым действием испытуемого. Испытуемый проигрывал в тех случаях, когда компьютер не соглашался с ложным ответом и соглашался с правдивыми. При проигрыше испытуемому предъявлялась информация о размере штрафа «-2» и «-5» соответственно. Дифференцированная система штрафования и поощрения использовалась для варьирования уровня проигрыша и выигрыша в Пр и Л – пробах между сессиями. Это было сделано для обеспечения вовлечения испытуемого в деятельность и соблюдения одинаковой ценности ложных и правдивых нажатий.

Рисунок 3. Схема тестового задания фМРТ-исследования.
Таким образом, в задачу испытуемого входило выиграть у компьютера в максимально большем количестве случаев. При этом как ложные, так и правдивые нажатия в игровых пробах осуществлялись для манипуляции мнением компьютера – заставить поверить в ложь и не поверить правде. В действительности же, испытуемые вводились в заблуждение относительно игры, так как согласие или несогласие компьютера с предложением об ориентации стрелки испытуемых было случайным. При этом слово «согласен» предъявлялось в 60% игровых проб, что неявно провоцировало к ложным действиям. Временные параметры контрольных К-проб были аналогичны используемым в Л и Пр- пробах. Отличие от игровых проб заключалось в характере окраса первого стимула проб – использовались черные стрелки с белым контуром. Появление такой стрелки было сигналом для испытуемого о том, что данная проба контрольная, т.е. нужно нажимать только ту кнопку, которая строго соответствует направлению стрелки. В случае корректного ответа сразу после слова «принято», появлялся второй стимул обратно связи «*». Если нажатие было некорректным, то это считалось нарушением инструкции и штрафовалось – предъявлялась информация о размере штрафа «-2». Во всех пробах нажатие одной кнопки указательным пальцем правой руки соответствовало ответу испытуемого –
«стрелка направлена вверх», а нажатие другой кнопки большим пальцем правой руки – «стрелка вниз». Правдивыми пробами считались те реализации, в которых испытуемый нажимал кнопку, соответствующую направлении стрелки. В случае если испытуемые нажимали кнопку, которая не соответствовала направлению стрелки (например, при стрелке «вверх» происходило нажатие большим пальцем), то такие пробы обозначались как ложные (Л-пробы). Для обеспечения эффективности дизайна исследования использовался варьирующий интервал между окончанием предшествующей и началом следующей пробы – от 500 до 2500 мс, с шагом 500 мс. Таким образом, общая длительность пробы, в среднем, составляла 10 сек. Такие параметры предъявления были выбраны с учетом представлений об оптимальных характеристиках фМРТ исследований (Amaro, et al., 2006, Wager, Nichols, 2003, Friston, et al., 1999). Статистический анализ фМРТ данных проводился по результатам индивидуально рассчитываемых t-контрастов Л > К, Пр > К и Л > Пр, результаты которых использовались в качестве переменных при групповом анализе данных.
Изучение системной организации мозговых систем обеспечения речи на примере генерации регулярных и нерегулярных форм глаголов. Третий вариант тестового задания использовался для моделирования экспериментальных условий для изучения системной организации мозговых систем обеспечения речи на примере генерации регулярных и нерегулярных форм глаголов. Каждое исследование состояло из трех сессий в каждой из которых предъявлялось 96 слов и 48 "пустых проб".
Временной интервал между пробами (т.е. от начала одной пробы до начала
следующей) составлял 4 сек (среднее значение). Время экспозиции слов - 700 мс, шрифт белый Times New Roman, 40 размер. Фон экрана черный. В остальное время до появления следующего стимула предъявлялось изображение "xxxxxx"). Для обеспечения эффективности дизайна исследования (Dale, 1999), использовался варьирующий интервал между окончанием предъявления предшествующего и началом предъявления следующего стимула: изображение "xxxxxx" в псевдослучайном порядке предъявлялось на 3.1 сек, 3,2 сек, 3.3 сек, 3.4 или 3.5 сек. Средняя длительность межстимульного интервала составляла 3.3 сек. Реальные и квази-глаголы предъявлялись в форме инфинитива. Существительные и квази-существительные предъявлялись в ед. числе. Испытуемые инструктировались давать свой ответ как можно быстрее и вслух: в ответ на предъявляемые стимулы нужно было сгенерировать вслух, либо глагол в первом лице ед. числа прошедшего времени или существительное во множественном числе). Варианты и примеры тестовых заданий представлены в Таблице 1.
Таблица 1. Экспериментальные условия и примеры
Экспериментальные условия | Предъявляемые слова | Корректные ответы |
Регулярные глаголы | кивать | киваю |
Нерегулярные глаголы | колоть | колю |
Регулярные псевдоглаголы | вупать | вупаю |
Нерегулярные псевдоглаголы | хороть' | хорю |
Регулярные существительные | сокол | соколы |
Нерегулярные существительные | посол | послы |
Регулярные псевдосуществительные | мокол | мокол (и моклю) |
Нерегулярные псевдосуществительные | фопол | фоплы (и фополы) |
Всего в исследовании предъявлялось по 35 слов каждого типа. Все устные ответы записывались одновременно с регистрацией фМРТ данных в условиях шума и для этих целей использовалась система шумоподавления (Persaio ™ Psychology
Software Tools Inc., Pittsburgh, PA, USA). Корректность сгенерированных слов оценивалась после исследования. Пробы, в которых сгенерированные испытуемыми слова не соответствовали ожидаемой модели (например, к глаголам регулярного (AJ- класса (применялась другая модель), учитывались как «ошибка» при последующем статистическом анализе фМРТ данных. Важно также отметить, что в данном исследовании выбор стимульного материала осуществлялся таким образом, чтобы избежать многочисленных методических ограничений свойственных предыдущим исследованиям (использование слов разной частотности, фонологической сложности и т.д.). Поэтому используемые в качестве стимульного материала слова были сбалансирован как по частотности, так и фонологической сложности. Кроме того, одновременное использование заданий по генерации глаголов и существительных (в случайной очередности), что позволяло избегать формирования определенной стратегии в ответах испытуемых и эффекта «прайминга» (влияния контекста тестового задания на скорость обработки и опознания стимулов). В результате из-за такой организации предъявления стимулов испытуемые не догадывались о целях исследования. В качестве переменных при статистическом анализе фМРТ данных использовались параметры регрессии, которые вычислялись отдельно для каждого испытуемого путем рассчета t-контрастов между всеми типами проба и состоянием покоя (П): РегГл>П, НерегГл>П, КвазиРегГл>П, КвазиНерегГл>П, Сущ>П, КвазиСущ>П и Ошибка>П.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Исследование организации нейрональных систем обеспечения целенаправленной деятельности в условиях вовлечения механизмов проактивного и реактивного когнитивного контроля на примере двухстимульного тестового задания Go/NoGo.
При изучении мозгового обеспечения проактивного когнитивного контроля, ожидалось, что на стадии подготовки деятельности, будут наблюдаться: 1) увеличение функциональной активности в областях мозга, связанных с обеспечением подготовки действия и формированием моторной программы (ассоциативные области моторной коры); 2) изменения функциональной активности в разных областях мозга при ожидании конкретного изображения и при удержании в памяти правила соответствия С1-С2 стимулов. Опираясь на ранее полученные данные, полученные при анализе потенциалов связанных с событиями (ПСС, (Kropotov, et al., 2011, 2015, 2016), предполагалось, что формирование репрезентации ожидаемого конкретного изображения будет связана с активностью зрительной ассоциативной коры и областей мозга, связанных с краткосрочной зрительной памятью (удержание изображения животного). Для проверки сформулированных выше гипотезах рассчитывался F- контраст соответствующий двухфакторному дисперсионному анализу:1) фактор
«ожидание» (сравнение всех проб с изображением животного в качестве С1 и проб с изображением растения в качестве С1) и фактор «повторение» (сравнение проб первого и второго исследования (Исследование 1 и 2 соответственно)). В результате были выявлены только структуры мозга, в которых уровень BOLD-сигнала в пробах с изображением «животного» в качестве С1 был значимо выше, по сравнению с пробами PA/PP-NOGO (отсутствие подготовки к действию): таламус, прецентральная и постцентральная извилина, дополнительная моторной кора, хвостатые ядра, инсулярная кора, нижняя и средняя лобная извилина. При этом не удалось установить ожидаемых значимых различий в BOLD-сигнале между процессами ожидания конкретного изображения животного С2 (Atarget) или изображения растения (Ptarget), которое должно было сравниваться с С1 (Acue) в рамках применения правила соответствия управляющих стимулов С1-С2 для запуска моторной команды. Вместе с тем статистический анализ поведенческих данных (F(1, 18)=6.63, p=0.02) указывал на то, что для нажатия кнопки мыши в AcuePGo пробах требовалось значимо больше времени (391 мс.), по сравнению с AcueAGo пробами (368.5 мс.). Это показывает, что необходимость применения правила сочетания между Acue (стимул С1) и Ptarget (стимул С2) является более сложной деятельностью по сравнению с ситуацией, когда ожидается конкретное изображение. Поскольку, поведенческие данные подтвердили разный характер участия рабочей памяти (удержание образа изображения или правила сочетания) в подготовке действий, далее проводился анализ функциональных взаимодействий для выявленных структур мозга. В соответствии с проверяемой
гипотезой, основанной на «активационно-ресурсном» подходе, необходимость удержания в памяти правила действия должна характеризоваться усилением функциональных взаимодействий с соответствующими структурами префронтальной коры. Исходя из этого, предполагалось, что проактивный когнитивный контроль в пробах AcuePtarget, будет характеризоваться большими значениями функциональных взаимодействий между выбранными ОИ и областями мозга, связанными с обеспечением планирования действий и, возможно, структурами префронтальной коры, обеспечивающими удержание в памяти правил действий. Последнее необходимо для усиления функциональной интеграции между ассоциативными областями вентрального пути обработки зрительной информации и областей, связанных с планированием, торможением и реализацией действий. Для проверки этих предположений был использован PPI-анализ для областей интереса (ОИ), располагающиеся в обоих полушариях в следующих анатомических структурах: хвостатое ядро (CaudN – caudate nucleous, координаты в левом и правом полушарии соответственно (х, y, z): -9, 8, -2 и 12, 11, 4), дополнительная моторная область (SMA –
supplementary motor area: -3, -7, 55 и 3, -7, 55), нижняя лобная кора/инсулярная кора
(IFG – inferior frontal gyrus: -30, 23, 1 и 36, 23, -2). В результате было показано, что для проб AcuePtarget (применение правила соответствия) практически во всех ОИ, за исключением левого хвостатого ядра, наблюдается повышение функциональных взаимодействий с ассоциативными областями затылочной (в том числе веретенообразной извилиной), височной коры и префронтальных отделов, связанных с обеспечением когнитивного контроля и планирования действий. Только для ОИ в левом хвостатом ядре, при расчете t-контраста статистических параметров функциональной связности AcueAtarget> AcuePtarget, было выявлено усиление функциональных взаимодействий с передней поясной корой (61 воксел, pFWE=0.024). При этом, обратный контраст не выявил значимых изменений функциональных взаимодействий.
Таким образом, при анализе процессов, связанных с проактивным когнитивным контролем, была впервые продемонстрирована зависимость организации взаимодействия дистантно расположенных зон мозга от характера вовлечения процессов рабочей памяти: ожидание конкретного стимула характеризовалось меньшим составом взаимодействующих звеньев. В случае, если реализация планируемого действия зависит от правила сочетания управляющих стимулов, то удержание в памяти правила соответствия между ними проявляется как расширение набора взаимодействующих структур мозга. При этом, функциональная специализация этих структур мозга связана с обеспечением сенсорной (каудальные области веретенообразной извилины и вторичной зрительной коры) и смысловой обработки изображений (передние отделы веретенообразной извилины), планированием действий (дополнительная моторная кора), а также селекции и контроля их реализации (нижняя лобная извилина). Это соответствует данным о специализации обнаруженных структур
префронтальной коры относительно процессов удержания в памяти правил реализации деятельности (Domenech, Koechlin, 2015, Koechlin, et al., 2003). Полученные данные укладываются в исходные предположения о зависимости характера функциональной интеграции от сложности планируемой деятельности и соответствуют логике
«активационно-ресурсного» подхода. При этом, обращает на себя внимание факт, что несмотря на выявленные изменения функциональных взаимодействий, BOLD-сигнал в анализируемых ОИ значимо не отличался между сравниваемыми условиями. Следует отметить, что аналогичные эффекты независимого поведения показателей локальной активности и дистантных взаимодействий наблюдались ранее при изучении импульсной активности нейронный популяций мозга человека (Медведев, Пахомов, 1989). Полученный результат показывает, что анализ психофизиологических взаимодействий не только раскрывает функциональную роль отдельных звеньев мозговых систем в зависимости от условий реализации поведения, но и выявляет участие структур мозга, которые не видны при использовании стандартных методов активационных исследований.
Далее проверялась гипотеза, в соответствии с которой, операция сравнения в рабочей памяти вовлекается в работу механизма реактивного когнитивного контроля, необходимого для обеспечения реактивного торможения. Проводимые ранее ПСС- исследований (Kropotov, et al., 2011, 2015, 2016), указывали на существование гипотетической операции сравнения с рабочей памятью, которая запускается в случае, когда предъявляемый целевой С2 стимул не соответствовал ожидаемому изображению конкретного животного (AcuePNoGo пробы). Для проверки этого отдельно рассматривались пробы с подавлением действий – т.е. AcueANoGo и AcuePNoGo. Напомним, что в пробах типа AcueAtarget изображения животных всегда были одинаковыми. Предполагалось, что в результате такого рассогласования запускаются процессы подавления актуализированной моторной программы (нажимать кнопку). В результате расчета t-контраста AcuePNoGo > AcueANoGo показано (см. Рисунок 5), что операция сравнения с содержимым рабочей памяти ассоциируется с билатеральным повышением BOLD-сигнала в передних отделах левой и правой веретенообразной извилины (pFWE=0.021 и pFWE=0.002 соответственно), а также медиальной области задней поясной коры (pFWE=0.001). При этом, обратный t-контраст AcueANoGo > AcuePNoGo не выявил статистических значимых изменений BOLD-сигнала. С учетом данных о пространственной дифференциации функциональной специализации задних и передних отделов веретенообразной извилины относительно процессов рабочей зрительной памяти (Saygin, et al., 2011, Zang, et al., 2016) полученный результат является независимым экспериментальным доказательством гипотезы о существовании операции сравнения в рабочей памяти (Folstein, Van Petten, 2008, Kropotov, et al, 2011, 2015, 2016). Исходя из этого ожидалось, что вовлечение данной операции для торможения подготовленного действия должно сопровождаться усилениями функциональных взаимодействий ОИ в области веретенообразной извилины и структурами, обеспечивающими подавление действий. Данное предположение опиралось на известные литературных данные о роли стриатума, а именно хвостатых ядер, и нижней лобной извилины правого полушария (Aron, et al., 2014) в обеспечении процессов реактивного тормозного контроля (Li, et al., 2008, Jahanshahi, et al., 2015).
В результате расчета t-контраста AcuePNoGo>AcueANoGo для ОИ в левой веретенообразной извилине был обнаружен кластер увеличения функциональных взаимодействий с правым хвостатым ядром (pFWE=0.001) и талусом (pFWE<0.001), т.е. звенями тормозной стриато-таламо-кортикальной нейрональной системы (Jahanshahi, et al., 2015). Выявленный эффект является прямым свидетельством в пользу взаимодействия между операциями сравнения с рабочей памятью и торможения действий в ситуации, когда предъявленный стимул не соответствует ожидаемому. Как и при изучении проактивного когнитивного контроля, предположения сформулированные на основе «активационно-ресурсного» подхода, получили экспериментальное подтверждение: вовлечение базовых операций характеризуется сочетанным повышением локальной соответствующей структуры мозга и усилением ее функциональных взаимодействий.
Но, вместе с тем, оказалось, что при вычислении обратного контраста AcueANoGo>AcuePNoGo для этой же ОИ в левой веретенообразной извилине (см. Рисунок 6), несмотря на относительное снижение ее локальной активности в условии AcueANoGo (по данным BOLD cигнала), которое моделировало рассогласование с правилом соответствия между управляющими стимулами С1 и С2, было обнаружено усиление функциональных взаимодействий со структурами в ассоциативной зрительной коре (pFWE=0.002), предклинье (pFWE<0.001), задней и средних третях поясной извилины (pFWE=0.004), височной коре и надкраевой извилине (pFWE=0.002), а также в правой префронтальной коре ПБ 10/9 (pFWE=0.25). Аналогичный характер соотношений локальной активности и дистантных взаимодействий был продемонстрирован и для ОИ в задней поясной коре.
Выявленный эффект, усиления функциональных взаимодействий левой веретенообразной извилины и задней поясной коры на фоне снижения BOLD сигнала в них, имеет принципиальное значение. Полученный довольно неожиданный результат является свидетельством того, что изменение функциональной роли отдельного звена нейрональной системы связано не столько с усложнением деятельности как таковой, а сколько с изменениями условий ее реализации в терминах вовлекаемых базовых операций и механизмов. Кроме того, выявленный эффект не укладываются в логику
«активационно-ресурсного подхода» и в большей степени соответствует представлениям о динамической организации мозговых систем, в соответствии с которыми функциональных взаимодействия между дистантно расположенными популяциями нейронов могут меняться независимо от характера их локальной импульсной активности (Медведев, 1987, Медведев, Пахомов, 1989). Таким образом, показано, что в ситуации подавления действия, запускаемого рассогласованием управляющего стимула с правилом действия, передняя область веретенообразной извилины специализирована не относительно операции сравнения с рабочей памятью,
а является одним из звеньев распределенной системы подавления действия с опорой на правило соответствия.
Исходя из этого, можно предполагать, что в случае, если вовлекаемые базовые операции и механизмы играют ведущую роль в организации реализуемой деятельности, как, например, при рассогласовании с содержимым рабочей памяти, которое запускает торможение подготовленной программы действия (AcueNoGo), то повышение локальной активности будет сопровождаться усилением дистантных взаимодействий. И в этой ситуации, «активационно-ресурсный» подход может быть адекватно применен. Однако, в ситуации, когда базовые операции и механизмы перестают быть определяющими в организации поведения, он уже не работает. Таким образом, выявленный феномен имеет важное значение для понимания организации нейрональных систем обеспечения целенаправленного поведения. Наблюдаемый ранее при изучении динамической организации мозговых систем эффект относительной независимости между локальной активностью нейронных популяций и характером их функциональных взаимодействий (Медведев, 1989), проявляется и на макроуровне, т.е. реорганизации функциональных отношений между относительно большими территория/структурами мозга. Также, полученный результат укладывается в представления системной психофизиологии (Александров, 2003), в соответствии с которыми не только ожидаемый и /или планируемый к достижению результат деятельности, но и способы его достижения, отражаются соответствующей организацией активности нейронов (Швырков, 1995).
На следующем этапе исследований, проводившихся в условиях реализации сознательных ложных действий, проверялось предположение о том является ли вовлечение базовых механизмов мозга условием, при котором будет наблюдаться сочетанное изменение локальной активности и дистантных взаимодействий вовлекаемых структур мозга.
Исследование роли базового механизма детекции ошибок в организации нейрональных систем обеспечения сознательных ложных и правдивых действий.
Предполагалось, что реализация сознательных ложных действий, не смотря на их выгодность, будет сопровождаться вовлечением мозгового механизма детекции ошибок, который мониторирует корректность любых действий. Данная гипотеза была сформулирована исходя из известных свойств работы мозговой системы детекции ошибок (Bechtereva, Gretchin, 1968, Бехтерева 1974, Bechtereva et al, 2005), который срабатывает на любое отклонение от стереотипа или принимаемого за норму образа действия. В частности, ожидалось, что ложные действия, по сравнению с правдивыми, будут характеризоваться изменениями локальной активности в структурах мозга, которые связаны с обеспечением работы данного механизма.
Полученные поведенческие данные не выявили предпочтения в пользу ложных или правдивых действий (p<0,93), а среднее групповые значения реализуемых ложных и правдивых действий составило 55.9±12 и 61.04±10.8 соответственно. Среднее время
реакции (ВР) составило 1076 мс. (стандартное отклонение (СО) 297 мс.) при ложных действиях, 1039 мс. (СО – 266 мс.) при правдивых и 878 мс. (СО – 162 мс.) при контрольных действиях. Статистический анализ с использованием параметрической статистики не выявил значимых различий между ВР ложных и правдивых действий (t=2.01, p=0.056), но непараметрический текст Вилкоксона установил, что ВР для ложных и правдивых действий могут считаться достоверно разными (Z=2.51, p=0.01). При этом, попарные сравнения ВР ложных и правдивых действий с ВР для контрольных проб выявили статистически значимые различия: Л>K (t=6.62, p<0.001) и Пр>К (t=4.46, p<0.001).).
В результате анализа BOLD сигнала было установлено два важных экспериментальных факта. Во-первых, реализация и ложных и правдивых действий в условиях интерактивной игры (целью которой является манипуляция мнением оппонента), обеспечивается работой одной и той же лобно-теменной нейрональной системы. Сопряженный анализ t-контрастов Л>К и Пр>К, выявил структуры мозга, которые одинаково вовлекались в обеспечение обоих типов игровых/манипулятивных действий (см. Рисунок 7 A-C). Во вторых, только прямое сравнение проб с ложными и правдивыми действиями (Л > Пр), установило, что ложные действия, по сравнению с правдивыми, характеризуются 6-ю кластерами значимого увеличения BOLD-сигнала, которые локализуются в области хвостатых ядер обоих полушарий, таламусе, прецентральной коре (дополнительная моторная область, ПБ 6) и нижней теменной дольке (ПБ 40). При этом обратное сравнение Пр>Л не выявило значимых изменений BOLD сигнала, связанных с реализацией правдивых действий.
Таким образом, было установлено, что оба типа действия обеспечиваются работой единой лобно-теменной системы, неоднократно наблюдавшейся в исследованиях мозгового обеспечения лжи (Sip, et al., 2010, Greene, Paxton, 2009 см. Spence, et al., 2004, Spence, 2009, Christ, et al., 2009), которая характеризуется широким спектром функциональной специализации: продукция новых (ложных) действий, принятие решений лгать или нет, подавление предпочтительной тенденции отвечать правдиво и также исполнительный контроль. Однако только в данном исследовании была установлена связь между увеличением BOLD-сигнала в хвостатых ядрах и реализацией именно ложных действий (см. Рисунок 7 D).
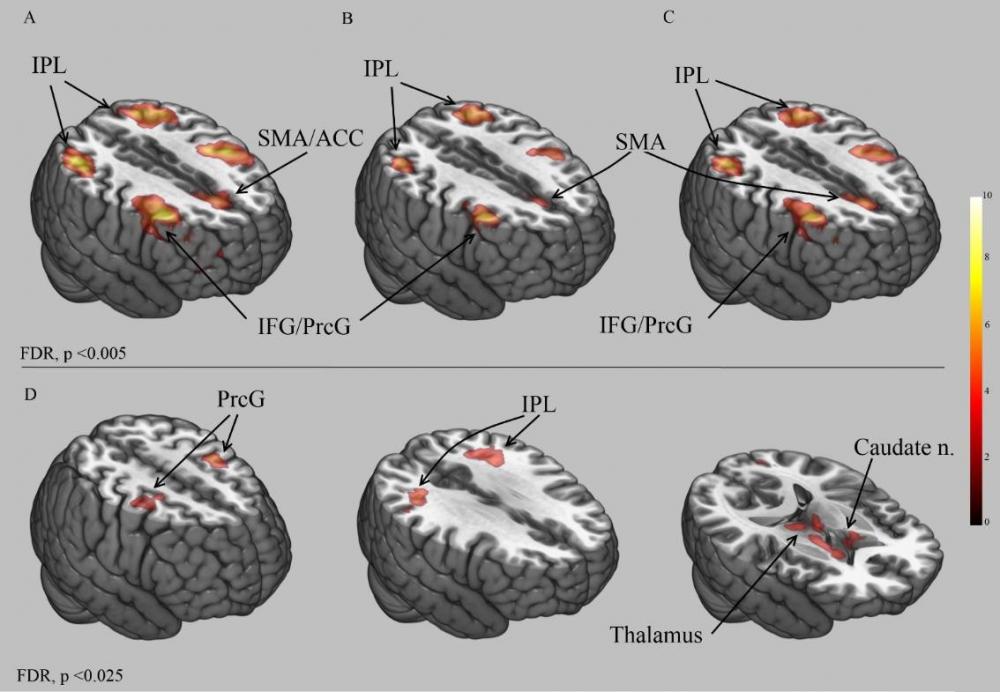
Рисунок 7. Изменения BOLD сигнала, отражающие процессы обеспечения ложных и правдивых действий. Обозначения: IPL – нижняя теменная кора, SMA – дополнительная моторная область, АСС – передняя поясная кора, IFG – нижняя лобная извилина, PreC – прентральная извилина, Caudate n. – хвостатое ядро, Thalamus – таламус, FDR – метод коррекции на множественность сравнения по методу false discovery rate. А) t-контраст Л>К, В) t-контраст Пр>К, С) результат сопряженного анализа по выявлению общих кластеров для Л>К и Пр>К, D) прямое сравнение Л>Пр.
И хотя активность хвостатых ядер при лжи интерпретировалась авторами как отражение когнитивных усилий, направленных на применение специальной инструкции по сигналу «фальсифицировать» автобиографическую информацию (Nunez, et al., 2005), подавление, мониторинг и выбор некорректных действий (Lee, et al., 2002, Menon, et al., 2001, Wager, et al., 2003), выявленный эффект рассматривается как свидетельство вовлечения мозговой системы детекции ошибок. Тот факт, что именно в хвостатых ядрах были впервые продемонстрированы изменения напряжения кислорода, отражающие некорректные пробы в тестовом задании на распознавании ранее запоминаемых слов (Bechtereva and Gretchin 1968), подтверждает изначальную гипотезу о срабатывании мозгового механизма детекции ошибок на ложные действия как на некорректные. В пользу именно такой функциональной роли хвостатых ядер соответствуют данные, в соответствии с которыми эта структура мозга функционально интегрирована с префронтальной, передней поясной корой и дополнительной моторной корой в конфликтных условиях, подразумевающих переключение между двумя типами заданий (Fan, et al., 2008).
Далее для решения задач настоящего исследования проверялось предположение, в соответствии с которым, срабатывание механизма детекции ошибок на ложные действия будет сопровождаться усилением взаимодействий хвостатого ядра со структурами префронтальной коры, обеспечивающими процессы выбора и реализаций действий. Проверка данной гипотезы, соответствующей логике «активационно- ресурсного» подхода, должна была продемонстрировать правомочность предположения о сочетанном изменении локальной активности и дистантных взаимодействий вовлекаемых структур мозга в ситуации, когда в реализации деятельность определяющую роль играют базовые мозговые механизмы. В результате PPI-анализ, с ОИ в области хвостатых ядер, выявил кластер со значимыми изменениями параметров PPI-регрессоров (показатели функциональной связности) для Л и Пр проб (t=4.95, FWE на уровне кластера, p<0.016, см. Рисунок 8). Групповой анализ PPI-данных показал, что пробы с ложными действиями характеризуются большими значениями параметров PPI-регрессоров (усредненными по всем вокселям выявленного кластера), по сравнению с правдивыми действиями.
Выявленный эффект указывает на то, что реализация сознательных ложных действий характеризуется относительным усилением функциональной связанности между головкой хвостатого ядра (ОИ) и латеральной частью нижней лобной извилины левого полушария (ПБ 45). Выявленные особенности функциональной интеграции хвостатых ядер и префронтальной коры соответствуют как имеющимся литературным данным об их структурной связанности (Alexander, Crutcher, 1990), так и представлениями о функциональной роли мозгового детектора ошибок (Bechtereva, Gretchin, 1968, Bechtereva, et al., 1991, 2005). Полученный результат подтверждает предложенную нами гипотезу о характере функциональных взаимодействий, лежащих в основе мозгового обеспечения лжи, ключевую роль в реализации которой играет срабатывание мозгового механизма детекции ошибок. Выявленный феномен является первым экспериментальным доказательством изменений функциональной интеграции мозговых структур при лжи и раскрывает их характер: на фоне ложных действий возрастает функциональное взаимодействие между префронтальной корой и базальными ганглиями.
Дополнительно изучался вопрос о характере вовлечения мозговых механизмов управления действиями в обеспечение ложных действий. В соответствии с доминирующими представлениями реализация ложных действий тесно связана с операцией подавления действий, которая необходима для обеспечения воздержания от тенденции отвечать правдиво (Verschuere, et al., 2012, Debey, et al., 2014, 2015). Учитывая экспериментальные данные, свидетельствующие о важной роли правой нижней лобной извилины (rIFG) в процессах обеспечения операции подавления действий (Aron, et al., 2014), можно ожидать что реализация ложных действий будет характеризоваться взаимодействиями со структурами этой области. Вместе с тем, другими исследователями отмечалось, что теоретически довольно сложно отделить процессы подавления действий от процессов их селекции (Mostofsky, Simmonds 2008). Поэтому более правдоподобным представляется, что именно процессы выбора действий в большей степени вовлекаются в мозговое обеспечение реализации ложных действий, которые, как считается, обеспечиваются структурами левой нижней лобной извилины (Moss, et al., 2003, Goghari, MacDonald, 2009). Следовательно, можно ожидать, что ложные действия будут характеризоваться усилением функциональных взаимодействий между передними отделами префронтальной коры и/или левой нижней лобной извилиной. Для проверки этих гипотез была выбрана ОИ в средней лобной извилине левого полушария, функциональная активность которой была одинаковой активной как при ложных, так и правдивых действиях. В результате прямого сравнения проб Л и Пр, был выявлен кластер в области левой нижней лобной извилины (p<0.013, c коррекцией FWE на уровне кластера), демонстрирующий усиление взаимодействий со средней лобной извилиной левого полушария (см. Рисунок 9). В соответствии с имеющимися данными, средняя лобная извилина левого полушария при лжи может вовлекаться в обеспечение обще-когнитивных процессов рабочей памяти, тормозного контроля, переключения между заданиями и генерацию/формирование ложной информации (Abe, et al., 2006, Christ, et al., 2009, Ito, et al., 2011, Sip, et al., 2010; Vartanian, et al., 2013). С другой стороны, функциональная роль нижней лобной извилины левого полушария при лжи, как правило, рассматривается в контексте обеспечения функции исполнительного контроля (т.е. подавления правдивых действий при лжи (Spence, et al., 2008, Lee, et al., 2009)), селекции действий (Langleben, et al., 2005) и переключения между задачами (Christ, et al., 2009, Fullam, et al., 2009). Учитывая вышесказанное, можно прийти к выводу, что полученные данные указывают на то, что ложные действия обеспечиваются при усилении взаимодействия между процессами удержания в памяти цели деятельности и способов ее достижения. При этом, факт большей функциональной связности между ОИ в средней лобной извилине и нижней лобной извилиной для Л-проб, рассматривается как свидетельство в пользу относительно большего вовлечения процессов селекции действий.
Выявленный факт усиления взаимодействий между средней и нижней лобными извилинами соответствует структурно-функциональной модели ростро-каудальной организации префронтальной коры, отражающей процессы контроля над механизмом выбора действий с учетом цели деятельности (Badre, 2008, Blumenfeld, et al., 2013, Domenech, Koechlin, 2015, Koechlin, Summerfield, 2007). Структуры префронтальной коры, расположенные относительно рострально, связаны с обеспечением удержания/актуализации в рабочей памяти текущих целей деятельности и абстрактных правил (способов) их достижения. При этом, области мозга расположенные относительно более каудально связаны с обеспечением удержания подцелей (конкретных моторных актов) и процесса селекции действий.
В соответствии с известными литературными данными и предполагаемым характером вовлечения механизма детекции ошибок (ДО) при реализации ложных действий, можно предполагать, что определяющую роль в их обеспечении роль играет взаимодействие между механизмом ДО и операциями выбора и подавления действий. В результате такого взаимодействия, время от времени, допускается реализация действий, которые некорректны с точки зрения их сенсомоторной репрезентации (т.е. представлений о корректном соответствии между нажатием кнопки и ориентаций стимула). Это означает, что при реализации ложных действий, срабатывание механизма детекции ошибок, сигнализирующего о некорректности, в некотором роде, преодолевается этим селективным механизмом обеспечение которого осуществляется на уровне нижней лобной извилины. С целью экспериментальной проверки данных гипотез, полученные фМРТ- данные анализировались с помощью метода динамического причинно-следственного моделирования (dynamical causal modeling, DCM Friston, et al., 2003, 2011; Stephan, et al., 2004). Для этого по данным активационного исследования и анализа психофизиологических взаимодействий были выбраны три ОИ. Две их них располагались в префронтальной коре левого полушария (IFG (-51, 14, 4), IFGcaud (-39,17, 19)), а третья в области головки хвостатого ядра (Cd
(-12, 11, 13). Было создано 20 моделей возможных взаимодействий между этими ОИ, взаимовлияния между которыми модулировались реализацией ложных и правдивых действий. Далее для всех 24 испытуемых был проведён DCM анализ по поиску
совокупностей моделей, которые наилучшим образом соответствуют зарегистрированным в анализируемых ОИ BOLD-сигналу. В результате было установлено, что наиболее вероятной является совокупность моделей, при которых и ложные и правдивые действия, за счет прямых и обратных взаимовлияний, влияют на взаимодействия между всеми анализируемыми областями интереса. Продолжение DCM-анализа моделей, входящих в выбранную совокупность, выявило «победившую модель» (см. Рисунок 10), в соответствии с которой и ложные и правдивые действия усиливают влияние хвостатого ядра на области интереса в нижней лобной извили (ОИ в IFGcaud и IFG), а также влияние IFGcaud на IFG. Причем отмечается восходящий характер влияний от хвостатого ядра на префронтальную кору, который соответствует предполагаемому характеру вовлечения мозгового механизма. Продемонстрированные взаимовлияния указывает на определяющую роль механизма детекции ошибок в организации поведения – структура мозга, связанная с обеспечением работы ДО модулирует активность областей мозга обеспечивающих выбор и реализацию действий.
Таким образом, на примере изучения вовлечения мозгового механизма детекции ошибок в обеспечение ложных действий показано, что соответствие между характером локальной активности и дистантными взаимодействиями структур мозга обеспечивающих целенаправленную деятельность может наблюдаться в ситуации, когда определяющую роль в ее организации играют базовые механизмы и операции. В этом отношении, характер выявленных взаимовлияний между вовлекаемыми структурами мозга позволяет говорить о том, что физиологический смысл выявленного функционального взаимодействия является активирующим: звено обеспечения
детекции ошибок модулирует активность в префронтальных структурах исполнительного (когнитивного) контроля действий. При этом, однако, остается открытым вопрос о том, сохранится ли характер выявленных на этом этапе закономерностей функциональных отношений между вовлекаемыми структурами мозга в условиях, когда целенаправленная деятельность обеспечивается вовлечением специализированных и развивающихся в ходе жизни индивида нейрональных систем. Проверке этого вопроса был посвящен следующий этап исследований, который проводился в условиях реализации деятельности по порождению элементов речи.
Изучение системной организации мозговых систем обеспечения речи на примере генерации регулярных и нерегулярных форм глаголов.
В соответствии с одной из распространенных теорий (т.н. двусистемных подход (Marslen-Wilson, Tyler, 1997, 1998, Pinker, Prince, 1988)), предполагалось что образование регулярных форм обеспечивается специализированной лобно-височной нейрональной системой, а в обеспечение генерации нерегулярных глаголов вовлекаются структуры височно-теменной коры, также связанные с обеспечением лексической памяти. В качестве регулярных форм использовались глаголы самого частотного продуктивного класса (таких, как читать – читаю, т.е. «АЙ» класса), которые характеризуются свойствами «дефольности», т.е. по модели «АЙ» класса в русском языке используется по умолчанию. Глаголы данного класса осваиваются первыми и более часто используются в детском возрасте (до 5-и лет). В качестве нерегулярных форм использовались глаголы из самых низкочастотных непродуктивных классов (таких, как колоть – колю, мести – мету и пр.). Предполагалось, что порождение регулярных и нерегулярных форм глаголов будет сопровождаться увеличением BOLD-сигнала в соответствующих структурах мозга: например, вовлечение области Брока в обеспечение генерации регулярных форм глаголов.
Анализ поведенческих данных показал, что испытуемые совершали меньше ошибок при порождении регулярных глаголов по сравнению с нерегулярными глаголами (в 22 из 735 и в 71 из 735, что соответствует 3.0% и 9.7% соответственно). Еще больше эта разница проявилась для квазиглаголов. При порождении квазирегулярных глаголов ошибки были совершены в 13.1% от всего набора слов (96 из 735). Больше всего ошибок совершалось при порождении нерегулярных квазиглаголов – в 320 из 735 (45.7%). Двухфакторный дисперсионный статистический анализ для повторных измерений выявил значимое влияние фактора «Лексичность» (т.е. сравнение количества ошибок при генерации реальных и квазиглаголов, F(1, 20) =
83.23; p < 0.001; g2 = 0.81) и фактора «Регулярность» (F(1, 20) = 32.29; p < 0.001; g2 = 0.62, т.е. сравнение регулярных и нерегулярных форм). Значимого взаимодействия этих факторов обнаружено не было. Факт большего количества ошибок при порождении нерегулярных квазиглаголов отражает объективную сложность использования непродуктивного и нерегулярного паттерна словоизменения.
Анализ фМРТ данных показал, что порождение нерегулярных глаголов характеризуется повышением BOLD-сигнала в области Брока, передней поясной коре, дополнительной моторной коре и теменной коре (см. Рисунок 11). Полученный результат первоначально был интерпретирован как свидетельство в пользу альтернативного, так называемого односистемного подхода, в рамках которого порождение всех форм обеспечивается единой системой (Hare, Elman, 1995, MacWhinney, Leinbach, 1991, Marchman, 1993, Marchman, et al., 1997, McClelland, Patterson, 2002, Plunkett, Marchman, 1993). Кроме того, принимая во внимание тот факт, что порождение нерегулярных глаголов характеризовалось относительно большим количеством ошибок, выявленное увеличение BOLD-сигнала в области Брока было интерпретировано как отражение усиление контроля за реализацией речевой деятельности. Это соответствовало демонстрируемой в литературе зависимости между уровнем активности области Брока и возрастанию сложности воспринимаемой информации до уровня синтаксиса (Blank, et al., 2016) и ряду теоретических моделей мозговой организации речи, в которых области префронтальной коры (в том числе и область Брока) не связаны с обеспечением реализации лингвистических операций как таковых, а ответственны за обеспечение операции выбора действий и удержания целей деятельности (De Baene, et al., 2015, Bornkessel-Schlesewsky, Schlesewsky, 2013; Koechlin, Summerfield, 2007, Novick, et al., 2010).

Рисунок 11. Изменения BOLD-сигнала, отражающие мозговое обеспечение порождения нерегулярных глаголов/квазиглаголов, по сравнению с регулярными глаголами/квазиглаголами. Обозначения: SMA – дополнительная моторная кора; IPL/SPL – нижняя/верхняя теменная долька; Precentral g. – прецентральная извилина; IFG – нижняя лобная извилина; MFG – средняя лобная извилина, красный цветом обозначены кластеры; выявленные в результате анализа t-контраста НерегГл>РегГл; зеленым цветом обозначены кластеры выявленные в результате анализа t-контраста КвазиНерегГл> КвазиРегГл; синим цветом обозначены области перекрытия между выявленными кластерами значимых изменений BOLD –сигнала.
Исходя из вышесказанного, предполагалось, что в соответствии с
«активационно-ресурсным» подходом, будет наблюдается усиление взаимодействий области Брока при порождении нерегулярных глаголов. Для проверки этого проводился анализ психофизиологических взаимодействий для трех областей интереса соответствующих локальным максимумам кластера, отражающего относительное увеличение BOLD-сигнала при порождении нерегулярных глаголов и распологавшегося в области левой нижней лобной извилины (см. Рисунок 12, А). В результате сравнения статистических параметров функциональной связности, рассчитываемых в t-контрасте НерегГлаг>РегГлаг с целью проверки сформулированных гипотез, не было выявлено значимых различий. Однако, обратный контраст РегГлаг>НерегГлаг выявил три кластера, расположенных в передней трети верхней височной извилины левого (p=0.007, FWE на уровне кластера) и правого полушария (p<0.001, FWE на уровне кластера). Указанные кластеры были выявлены только для одной из трех ОИ (для двух остальных значимых изменений функциональной связности обнаружено не было). Дополнительный расчет средних по группе параметров статистической связности по данным средних значений всех вокселах выявленных кластеров показал усиление функциональных взаимодействий с левой нижней лобной извилиной для условий с порождением регулярных глаголов, по сравнению с нерегулярными. Таким образом оказалось, что усиление функциональных взаимодействий височных структур с областью Брока, уровень активности которой относительно снижен, наблюдается при порождении регулярных глаголов. В результате не получило подтверждение предположение, сформулированное исходя из
«активационно-ресурсного» подхода.
Строго говоря, полученные нами данные полностью не укладываются в теоретические представления ни «двойной системы» (DS-гипотеза), ни «единой системы» (SS-гипотеза). В соответствии с гипотезой о существовании только единой системы порождения, основную роль играет частота морфологического паттерна. Соответственно, хотя в данной работе и в исследовании (Stamatakis, et al., 2005) использованные слова являлись регулярными, данные о функциональных взаимодействиях расходятся. Аналогично, DS-гипотеза предполагает наличие
«дефолтного» правила порождения для регулярных форм (автоматизированного до стереотипа) и активное вовлечение памяти для всех остальных форм. Следовательно, и с этих позиций не должно было быть наблюдающихся отличий в мозговом обеспечении порождения регулярных форм. Одно из наиболее вероятных объяснений может быть найдено в гибридной модели Янга (Yang, 2002), подразумевающей существование нескольких правил порождения (для регулярных и нерегулярных форм), что подтверждается данными поведенческих исследований, проводившихся на материале русского языка (Gor, Chernigovskaya, 2001, 2003, 2005, Gor, et al., 2008, 2013). Можно предположить, что в русском языке процессе порождения нерегулярных форм все же связан с морфологической декомпозицией (отбрасыванием суффикса
инфинитива). А некоторая комбинация процессов извлечения из памяти и морфологической декомпозиции, в определенном смысле, делает порождение нерегулярных форм более сложным по сравнению с регулярными. Порождение нерегулярных глаголов английского языка с этих позиций будет осуществляться сравнительно легче, поскольку оно не предполагает вовлечения именно морфологической обработки слов. Далее, с целью выяснения возможных сценариев организации функциональных отношений между выявленными структурами мозга, которые продемонстрировали разный характер взаимодействий между структурами мозга вовлекаемыми в обеспечение порождения регулярных и нерегулярных форм, был применен метод по динамическому-причинно-следственному моделированию (DCM-анализ). Конкретно, проверялся вопрос о том, вовлекается ли левосторонняя речевая лобно-височная система в порождение именно регулярных форм (по символическому правилу, согласно т.н. двусистемному подходу), или, в действительности, все глаголы порождаются единой билатеральной височно-теменной системой (т.н. односистемный подход).
В соответствии с этим, были созданы четыре DCM-модели описывающие возможные варианты организации прямых и обратных взаимовлияний между ОИ в области Брока и двух ОИ в верхних височных извилинах, которые были выявлены при PPI-анализе. В результате было установлено, что в соответствии с наиболее вероятной моделью порождение регулярных глаголов осуществляется на фоне влияния структуры в левой верхней височной извилине на область Брока. В то же время, порождение нерегулярных глаголов характеризуется влиянием структур височной коры обоих полушарий на область Брока (см. Рисунок 13).

Рисунок 13. Наиболее вероятная DCM-модель, описывающая направления и характер взаимовлияний между взаимодействующими структурами мозга, вовлекаемыми в порождение слов. Обозначения: LSTG – область интереса в верхней височной извилине левого полушария, RSTG – область интереса в верхней височной извилине правого полушария, LIFG – область интереса в области Брока, РегГлаг – порожение регулярных глаголов, НерегГлаг – порождение нерегулярных глаголов.
Таким образом, получены новые данные, раскрывающие работу мозговых структур звеньев нейрональных систем обеспечения порождения элементов речи и организацию ментального лексикона. Установлено, что в зависимости от типа морфологического процесса меняется режим функционирования и конфигурация такой системы. Получено подтверждение вовлечения левосторонней лобно-височной системы в процессы, связанные с порождением регулярных форм, которые подразумевают составление словоформы из морфем. На основании полученных данных, можно утверждать, что наблюдаемый нами ранее эффект регулярности при изучении функциональной связности в действительности отражает процесс порождения по правилу и обеспечивается взаимодействиями с областью Брока. Тогда как билатеральная система вовлекается в обеспечение процессов, подразумевающих работу лексической памяти.
Групповой анализ параметров, отражающих характер взаимовлияний, установил, что порождение регулярных глаголов осуществляется при негативном
влиянии левой височной извилины на активность в области Брока. Порождение же нерегулярных глаголов, напротив, характеризуется позитивной модуляцией активности области Брока со стороны верхних височных извилин обоих полушарий и соответствует как бы большему вовлечению этой области. И, хотя выявленный характер взаимовлияний соответствует тому, что наблюдалось при анализе изменений уровня активности вовлекаемых структур, в целом наблюдается расхождение между показателями их локальной активности и дистантных взаимодействий.
По показателю количества ошибок порождения, генерация нерегулярных глаголов является более сложной деятельностью по сравнению с порождением регулярных, что соответствующим образом отражается фактом относительного повышения активности в области Брока. Однако, взаимодействие этой области со структурами височной коры относительно слабое (по сравнению с регулярными глаголами). Данный факт не укладывается в логику «активационно-ресурсного» подхода, в соответствии с которым стоило ожидать усиление функциональных взаимодействий при повышении локальной активности вовлекаемой структуры мозга. Аналогично, порождение регулярных глаголов обеспечивалось на фоне усиленных взаимодействий между этими же структурами (областью Брока и структурами височной коры) при относительно сниженных показателях их активности (по данным BOLD-сигнала). Таким образом, полученные данные в большей степени соотносятся с выявляемыми ранее принципами организации динамической активности нейронных популяций (Кроль, Медведев 1982, Медведев, 1987, Медведев, Пахомов, 1989) и усилением локальной синхронии при ослаблении дистантной (Свидерская, 1987). Кроме того, выявленный эффект воспроизводит наблюдавшееся в условиях теста Go/NoGo несоответствие между уровнем активности вовлекаемой структуры мозга и характером ее функциональных взаимодействий. Напомним, что данный эффект наблюдался для области интереса в правой передней поясной коре при сравнении относительно сложных и простых проб с реализацией действий, в соответствии с которым были обнаружены следующие соотношения: AcuePGo>AcueAGO для BOLD- сигнала и AcuePGo
«активационно-ресурсного» подхода не всегда правомочна и не может объяснить такие эффекты.
Вместе с тем, основываясь на известных представлениях о свойствах динамической организации систем мозга (Бехтерева, 1974, Медведев, 1987, Медведев, Пахомов, 1989) и принципах организации функциональных систем (Анохин, 1975, Александров, 1989) можно высказать ряд предположений о явлениях, лежащих в основе наблюдаемых расхождений между характером взаимодействий и уровнем функциональной активности вовлекаемых структур мозга. Так, например, при порождении элементов речи усиление взаимодействий на фоне пониженной активности может рассматриваться как эффективная интеграция области Брока и верхней височной извилины, которая достигается за счет минимизации территории (Бехтерева, 1974) и, одновременно, относительного увеличения количества вовлекаемых жестких звеньев. В силу того, что класс регулярных глаголов осваивается самым первым, навык по их генерации является относительно более автоматизированным, и нейрональная система их обеспечения является более дифференцированной (развитой) по сравнению с генерацией нерегулярных глаголов. Иными словами, можно говорить о том, что в этом проявляется принцип историзма в формировании функциональных систем (см. Александров, 1989), в соответствии с которым относительно рано, в онтогенезе, формирующиеся функциональные системы являются наиболее дифференцированными. Отличительной их особенностью также может быть преобладание жестких звеньев. И в терминах системной психофизиологии, полученные данные могут отражать функциональную специализацию области Брока не относительно речевой функции как таковой, а относительно более или менее дифференцированных систем обеспечения речевой деятельности. Исходя из этого, можно ожидать, что звенья системы обеспечения порождения нерегулярных глаголов (элементы менее частотного и непродуктивного глагольного класса, порождаемого не по правилу и осваиваемому гораздо позже регулярного класса) характеризуются относительно большим содержание гибких звеньев. Вероятно, именно за счет большого количества вовлекаемых гибких звеньев и динамизма, присущего работе мозговых систем, (Медведев, Пахомов, 1989) наблюдается повышение локальной активности, но вместе с этим и снижение дистантных взаимодействий. Эффект, подобный тому, наблюдался ранее при усложнении деятельности (Свидерская, 1987) и заключался в преобладании локальной синхронии над дистантной. Таким образом, на примере результатов данного исследования видно, как усложнение деятельности приводит не к изменениям количественных характеристик (например, состава и степени вовлечения структур мозга), а качественной перестройке функциональной организации вовлекаемых нейрональных систем.
ЗАКЛЮЧЕНИЕ
Предложенный исследовательский подход по сочетанному рассмотрению изменений уровня активности вовлекаемой структуры мозга и ее дистантных взаимодействий позволяет на новом уровне изучать системную организацию работы мозга в условиях реализации целенаправленной деятельности. В результате проведенных в настоящей работе исследований было показано, что концепция об обеспечении деятельности принципиально динамичными во времени и пространстве мозговыми системами (Медведев, 1987), предложенная по результатам анализа активности отдельных нейронных популяций, применима и для описания закономерностей функционирования нейрональных систем в масштабе всего мозга. Установлено, что с одной стороны, реорганизация дистантных функциональных взаимодействий между вовлекаемыми в обеспечение текущей деятельности структурами мозга происходит независимо от уровня их функциональной активности, а с другой, соотношение этих показателей позволяет судить о роли таких структур в работе системы. Поскольку считается, что регистрируемый BOLD-сигнал в основном отражает уровень синаптической активности относительно больших территорий мозга (Logothetis, 2008, Shmuel, Maier, 2015), то выявленное принципиальное непротиворечие полученных в настоящей работе данных с выявленными ранее динамическими характеристиками импульсной активности нейронных популяций больного мозга является свидетельством в пользу фундаментальности обнаруженных закономерностей системной организации мозговых систем.
Обнаруженные варианты соотношений между локальными характеристиками BOLD-сигнала и свойствами статистической зависимости между его показателями в дистантно расположенных структурах мозга расширяют преставления о пространственном и временном динамизме нейрональных систем (Медведев, Пахомов, 1989) обеспечения текущей деятельности. Было показано, что деятельность, определяющую роль в организации которой играют базовые мозговые механизмы и операции, характеризуется соответствием между увеличением активности и функциональных взаимодействий между структурами их обеспечения. При вовлечении операции сравнения с рабочей памятью, когда рассогласование предъявленного стимула с ожидаемым приводит к реактивному торможению уже подготовленного действия, относительное повышение BOLD-сигнала в ростральном отделе левой веретенообразной извилины сопровождалось усилением взаимодействий с хвостатым ядром правого полушария. Аналогично, срабатывание механизма детекции ошибок (ДО, Bechtereva, Gretchin, 1968) на ложные действия как некорректные проявлялось повышением локальной активности левого хвостатого ядра и усилением взаимодействий этой структуры с отделами левой нижней лобной извилины. Анализ взаимовлияний между выявленными взаимодействующими звеньями продемонстрировал модулирующий характер влияния хвостатого ядра на
активность структур нижней лобной извилины, которые, как считается, связаны с обеспечением реализации, выбора и планирования действий. Это доказывает гипотезу о ведущей роли механизма детекции ошибок в организации работы мозга (Медведев, и др., 2003, Bechtereva, et al., 2005).
Если вовлечение базовых механизмов и операций проявлялось соответствием между анализируемыми в рамках предложенного «активационно-коннекционисткого» исследовательского подхода показателями активности мозга, то усложнение реализуемой деятельности приводило к их рассогласованию. Так, на примере экспериментальной ситуации, когда при усложнении деятельности, обусловленном необходимостью подавления подготовленного действия с использованием удерживаемого в памяти правила сочетания управляющих стимулов, показано, что процессы реактивного торможения обеспечиваются на фоне включения в большой набор взаимодействующих звеньев уже упоминавшегося рострального отдела левой веретенообразной извилины. При этом, уровень BOLD сигнала в этой структуре был снижен относительно сравниваемого простого тестового условия, когда в обеспечение деятельности вовлекалась операция сравнения с рабочей памятью. Аналогичный по своей сути эффект был продемонстрирован в условиях порождения регулярных глаголов, когда при относительном снижении уровня активности области Брока отмечалось усиление ее взаимодействий со структурами височной коры. А в относительно сложном тестовом задании генерации глаголов относительно менее частотного глагольного класса, наоборот, отмечалось ослабление этих взаимодействий при повышении активности в области Брока. Представленные варианты сочетаний локальной активности и дистантных взаимодействий демонстрируют, что сам по себе фактор сложности не оказывает специфического, определенного влияния на системную организацию мозговых систем обеспечения целенаправленной деятельности. Пользуясь терминологией системной психофизиологии (см. работы Александрова Ю.И. (Александров, 2003)), наблюдаемые отличия в функциональных взаимовлияниях между областью Брока и височной корой могут отражать ее функциональную специализацию относительно функциональных систем разной степени дифференцированности. Так, генерация регулярных глаголов, осуществляемая с применением правила порождения регулярных форм, обеспечивается относительно более дифференцированной и развившейся ранее в онтогенезе левосторонней лобно- височной системой. Тогда как генерация нерегулярных глаголов в большей степени осуществляется вовлечением лобно-височной билатеральной системы, связанной с обеспечением лексической памяти. Полученные новые данные о разном направлении взаимовлияний между структурами лобно-височной системы при внешне схожих, но морфологически различающихся речевых процессах, укладываются в представления, в соответствии с которыми не только сам ожидаемый результат, но и способы его достижения отражаются соответствующей организацией активности нейронов (Швырков, 1995). А факт выявленных перестроек функциональных взаимодействий в
процессе реализации одинаковой деятельности по порождению элементов речи позволяет говорить о динамичности функциональной специализации вовлекаемых структур мозга.
В результате проведенных исследований убедительно показано, что логика
«активационно-ресурсного» подхода оправдана далеко не во всех случаях, а отдельный анализ показателей локальной активности не учитывает системные характеристики работы мозга. Предложенный исследовательский подход позволяет выявлять не только новые звенья мозговых систем, не видимые в активационных исследованиях, но и специализацию структуры мозга как одного из взаимодействующих звеньев вовлекаемых мозговых систем.
Таким образом, полученные данные демонстрируют важность развития исследований системной организации мозгового обеспечения поведения в рамках подхода по сочетанному анализу локальной активности и дистантных взаимодействий, также и при заболеваниях мозга. Высокая актуальность развития этого направления исследований обусловлена рутинным использованием в медицинской нейровизуализации в основном только показателей локального уровня функциональной активности (Медведев, 2008). Что, как показывают результаты настоящей работы, является препятствием на пути исследований по оптимизации существующих и развитию новых методов диагностики и терапии.
ВЫВОДЫ
- Предложенный исследовательский подход по сочетанному анализу изменений локальной активности и дистантных взаимодействий вовлекаемых мозговых структур выявил относительную независимость перестроек их функциональных взаимодействий от показателей изменения уровня нейрональной активности.
- Получено подтверждение существования операции сравнения с рабочей памятью в условиях внешнего управления действиями и свидетельство вовлечения этой операции в процессы реактивного торможения подготовленного моторного действия, характеризующееся повышением локальной активности рострального отдела левой веретенообразной извилины и усилением ее функциональных взаимодействий со структурами обеспечения подавления действий.
- Выявлена пространственная дифференциация функциональной специализации веретенообразной извилины, в соответствии с которой ее каудальный и ростральный отделы вовлекаются в обеспечение неспецифической по отношению к реализуемой деятельности операции сенсорной дискриминации и базовой операции сравнения с рабочей памятью соответственно.
- Выявлено, что функциональная специализация рострального отдела веретенообразной извилины левого полушария относительно процессов реактивного когнитивного контроля характеризуется изменчивостью, которая определяется характером взаимодействий процессов рабочей памяти и подавления подготовленного
действия. Показано, что при изменениях условий реализации деятельности, т.е. при переходе от торможения действий, вызванного рассогласованием между предъявленным и ожидаемым целевым стимулом, к их подавлению, с опорой на правило соответствия между сигнальным и целевым стимулами, наблюдается реорганизация и расширение набора звеньев, с которыми усиливается взаимодействие этой структуры на фоне понижения уровня BOLD сигнала в ней.
- Эффект зависимости реорганизации сети взаимодействующих звеньев от характера вовлечения рабочей памяти, при отсутствии значимых изменений BOLD сигнала в вовлекаемых структурах, продемонстрирован и для мозгового обеспечения механизма проактивного когнитивного контроля, необходимого для подготовки действия. Таким образом показано, что изменение конфигурации взаимодействующих элементов нейрональной сети зависит как от цели деятельности, так и от способов ее достижения, определяемых характером использования рабочей памяти.
- Подтверждена гипотеза о вовлечении мозговой системы детекции ошибок в обеспечение сознательных ложных и правдивых действий в условиях взаимодействия с оппонентом, которое свидетельствует об участии данного механизма в мониторинге корректности любых действий. Выявленный больший уровень функциональной активности левого хвостатого ядра именно при ложных действиях (по сравнению с правдивыми) является доказательством срабатывания механизма детекции ошибок на ложные действия как на некорректные.
- На примере вовлечения мозгового механизма детекции ошибок в обеспечение сознательных ложных действий продемонстрирована ведущая роль базовых механизмов в организации функциональной активности мозговых систем. Выявлена восходящая структура модулирующих влияний от хвостатых ядер до префронтальной коры, которая наблюдается на фоне совпадения показателей локальной активности и усиления функциональных взаимодействий.
- Получено доказательство существования мозговых речевых систем, специализированных относительно морфологически разных процессов порождения элементов речи, которые реализуются с опорой на относительно автоматизированное правило порождения (регулярные глаголы) или на процессы выбора подходящего варианта из лексической памяти (нерегулярные глаголы). Выявлена изменчивость функциональной роли области Брока, негативный или позитивный характер модуляции активности которой со стороны височных областей коры определяется типом нейрональной системы, в состав которой она вовлечена.
- Совокупность полученных данных показывает, что характер реорганизации функциональных взаимодействий между вовлекаемыми в обеспечение текущей деятельности структурами мозга определяется не столько сложностью реализации деятельности как таковой, но также и характером вовлечения базовых механизмов и специализированных систем.
- Полученные новые экспериментальные данные демонстрируют, что применяемый анализ психофизиологических взаимодействий позволяет обнаруживать новые не видимые стандартным активационным анализом структуры мозга, которые вовлекаются в качестве одного из взаимодействующих звеньев нейрональной системы обеспечения деятельности.
- Киреев, М.В. Особенности функционального взаимодействия хвостатого ядра и нижней лобной извилины в процессе обеспечения сознательных ложных действий / М.В. Киреев [и др.] // Физиология человека. – 2015. – Т. 41, № 1. – С. 29.
- Kireev, M.. Changes in functional connectivity within the fronto-temporal brain network induced by regular and irregular Russian verb production / M. Kireev [et al]// Front. Hum. Neurosci. – 2015. – Vol. 9. – 36.
- Slioussar, N. An ER-fMRI study of Russian inflectional morphology / N. Slioussar,
4. Список опубликованных работ по теме диссертации
M.V. Kireev, T.V. Chernigovskaya, G.V. Kataeva, A.D. Korotkov, S.V. Medvedev // Brain and Language. – 2014. – V. 130. – P. 33-41.
- Kireev, M. Possible role of an error detection mechanism in brain processing of deception: PET-fMRI study / M. Kireev [et al.] // International Journal of Psychophysiology. – 2013. – V.90, № 3. – P. 291-299.
- Катаева, Г.В. Факторная структура значений регионарного мозгового кровотока и скорости метаболизма глюкозы как инструмент исследования режима спонтанной активности мозга в состоянии оперативного покоя / Г.В. Катаева, А.Д. Коротков, М.В. Киреев, С.В. Медведев // Физиология человека. – 2013. – Т. 39, № 1. – С. 60.
- Киреев, М.В. Патология передней поясной извилины при обсессивно- компульсивных расстройствах / М.В. Киреев [и др.] // Физиология человека. – 2013. – Т. 39, № 1. – С. 67.
- Киреев, М.В. Современные методы функциональной томографической нейровизуализации в исследовании функций больного и здорового мозга / М.В. Киреев [и др.] // Российский физиологический журнал им. И.М. Сеченова. – 2013. – Т.99. – С. 53-63.
- Киреев, М.В. Исследование методом функциональной магнитно-резонансной томографии мозгового обеспечения сознательной лжи / М.В. Киреев [и др.] // Физиология человека. –2012. – Т. 38. – С. 41-50.
- Киреев, М.В. Мозговой механизм детекции ошибок – ПЭТ исследование / М.В. Киреев [и др.] // Российский физиологический журнал им. И.М. Сеченова. – 2011. – Т.97. – С. 1060-1065.
- Киреев, М.В. ПЭТ исследование функционального состояния мозга при навязчивых состояниях / М.В. Киреев [и др.] // Профилактическая и клиническая медицина. –2011. – Т.40. – С. 245-248
- Киреев М.В. Этапы мозгового обеспечения заведомо ложных ответов / М.В. Киреев [и др.] // Физиология человека. – 2007. – Т. 33,№ 6. – С. 5-13
- Киреев М.В. Исследование механизмов мозгового “детектора ошибок” при ложных ответах в норме и под воздействием алкоголя / М.В. Киреев [и др.] // Физиология человека. – 2008. – Т. 34, №2. – С. 13-22.
Источник:
Дата создания: 12.03.2024
Последнее редактирование: 12.03.2024
Относится к аксиоматике: Системная нейрофизиология.
Другие страницы раздела "Целевая мотивация":
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя