Автор: Александр Марков
Сложные генно-инженерные эксперименты, связанные с воссозданием давно исчезнувших «предковых» белков, позволили американским биологам расшифровать последовательность событий, в результате которых у предков пекарских дрожжей усложнилась система регуляции работы генов. Удвоение гена позволило двум копиям многофункционального регуляторного белка Mcm1 накопить мутации, ведущие к частичной потере функциональности. Функции, утраченные каждой из копий, были разными, благодаря чему копии перестали быть избыточными: теперь оба белка, по-разному подпорченные мутациями, стали жизненно необходимы организму. Дальнейшая специализация двух белков подстегивалась тем, что поначалу они конкурировали, мешая друг другу работать. Минимизация конкуренции потребовала закрепления дополнительных мутаций. В итоге система генной регуляции усложнилась (появился новый белок-регулятор), хотя изначальной потребности в этом, по-видимому, не было. Усложнение стало побочным эффектом цепочки взаимообусловленных событий, начало которым положило случайное удвоение гена. Возможно, конкуренция копий удвоившихся генов является важным механизмом, ограничивающим и направляющим процесс формирования эволюционных новшеств.
Дупликация (удвоение) генов с последующим разделением функций между копиями — один из магистральных путей появления эволюционных новшеств (см. ссылки внизу). Новое исследование, выполненное биологами из Калифорнийского университета в Сан-Франциско, проливает свет на два важных аспекта этого процесса.
Во-первых, оно показало, что генные дупликации могут чисто автоматически, через ряд взаимообусловленных промежуточных шагов, приводить к усложнению генно-регуляторных сетей. Это происходит даже в том случае, если в таком усложнении нет очевидной необходимости и организм мог бы продолжать с тем же успехом обходиться предковой, более простой системой генной регуляции.
Во-вторых, работа продемонстрировала, что копиям удвоившегося гена (паралогам) не всегда удается легко и быстро разделить функции, особенно если ген включен в сложную сеть межмолекулярных взаимодействий. В этом случае копии, едва начав делить функции, начинают конкурировать за субстраты и мешать друг другу работать. Для устранения этой конкуренции требуется закрепление дополнительных мутаций, что способствует окончательному превращению паралогов в два специализированных белка со строгим разделением обязанностей.
Авторы изучили последствия дупликации гена Mcm1 у дрожжей. Этот ген есть у всех грибов. Он кодирует важный регуляторный белок (транскрипционный фактор), который, объединяясь в комплексы с другими регуляторными белками (кофакторами), присоединяется к ДНК, чтобы активировать близлежащие гены (рис. 1).
У некоторых видов дрожжей (например, у Kluyveromyces lactis) гены метаболизма аргинина (ARG) регулируются комплексом из двух молекул белка Mcm1 (такие комплексы называют гомодимерами), который, в свою очередь, соединяется с кофактором Arg81 (рис. 1, A). У пекарских дрожжей Saccharomyces cerevisiae и их ближайшей родни белковый комплекс, регулирующий работу ARG-генов, устроен сложнее. Он включает не два, а три разных белка. Вместо гомодимера из двух молекул Mcm1 пекарские дрожжи используют гетеродимер — соединенные молекулы белков Mcm1 и Arg80 (рис. 1, B).
Другая группа генов (α-гены, отвечающие за «половые признаки» дрожжей, относящихся к типу спаривания α, см. Детерминация типов спаривания у дрожжей) у всех дрожжей регулируется одинаково: при помощи гомодимера Mcm1 в комплексе с кофактором Matα1 (рис. 1, C, D).
Таким образом, у S. cerevisae α-гены (и многие другие гены, подконтрольные Mcm1) регулируются так же, как у других дрожжей, в то время как ARG-гены регулируются более сложным способом, с участием дополнительного белка Arg80, которого нет у других дрожжей. Авторы задались целью разобраться в причинах этого различия.
Анализ геномов показал, что Arg80 произошел от Mcm1 в результате генной дупликации. У предков S. cerevisae исходный ген, кодировавший белок Mcm1, подвергся удвоению. После этого в каждой из двух копий закрепились различные аминокислотные замены. Одна из копий (Mcm1 пекарских дрожжей) сохранила большую часть исходных регуляторных функций, а другая (Arg80) изменилась сильнее и специализировалась на регуляции ARG-генов.
Авторы построили эволюционное дерево белков Mcm1/Arg80 и реконструировали аминокислотные последовательности трех предковых белков, занимающих на дереве ключевые позиции (рис. 2).
Первый из «воскрешенных» белков получил название AncMADS. Здесь «Anc» происходит от слова «ancestor» (предок), MADS (см. MADS-box) — название обширной группы регуляторных белков, к которой относятся рассматриваемые белки. AncMADS — это тот белок, ген которого подвергся дупликации у предка пекарских дрожжей (и их ближайшей родни). Второй реконструированный белок, AncMcm1, — это исходный вариант белка Mcm1 пекарских дрожжей и их родни, то есть одного из двух паралогов, сформировавшихся после дупликации и разделения функций. Наконец, AncArg80 — это исходный вариант второго паралога, белка Arg80.
Авторы синтезировали гены реконструированных предковых белков и внедрили их в клетки пекарских дрожжей, лишенных собственных белков Mcm1 и Arg80. Утрата Mcm1 смертельна для дрожжей при любых обстоятельствах, а без Arg80 у них нарушается метаболизм аргинина, что становится критичным в среде, где аргинин (или его предшественник орнитин) является единственным источником азота.
Оказалось, что до-дупликационная версия предкового белка (AncMADS) прекрасно справляется с работой обоих современных белков (Mcm1 и Arg80). Дрожжи, лишенные одного из этих белков или обоих сразу, выживают и нормально размножаются, если в их геном вставлен ген белка AncMADS. Предковый белок AncMcm1 успешно заменяет современный Mcm1, но не может возместить утрату Arg80. Соответственно, предковый белок AncArg80 компенсирует потерю Arg80, но не спасает дрожжи, лишенные Mcm1.
Эти результаты в совокупности с другими экспериментальными данными показали, что вскоре после дупликации две копии исходного гена AncMADS накопили мутации, которые привели к частичной потере функциональности. Один из паралогов (AncMcm1) утратил способность соединяться с кофактором Arg81, а другой (AncArg80) разучился связываться с кофактором Matα1 (рис. 3).
Авторам удалось найти конкретные аминокислотные замены, изменившие структуру «кофактор-связывающего кармана» белковой молекулы, что и привело к частичной утрате функциональности. AncMcm1 потерял способность взаимодействовать с Arg81 благодаря одной замене, которая была унаследована всеми современными вариантами белка Mcm1 «пост-дупликационных» дрожжей. AncArg80 лишился способности взаимодействовать с Matα1 из-за трех других аминокислотных замен (которые тоже сохранились у современных потомков этого белка).
Полученные результаты показывают, что разделение функций между паралогами было основано на так называемой «субфункционализации», то есть частичной утрате разных аспектов исходной функции двумя копиями удвоившегося гена. Многие теоретики и раньше считали такой сценарий разделения функций наиболее вероятным.
После удвоения гена мутации, нарушающие функциональность одной из копий, перестают быть вредными и не отсеиваются отбором — ведь остается вторая копия, успешно справляющаяся с той работой, которую больше не может выполнять первая копия. После того как одна из копий утратила какую-то функцию, отбор уже не допустит утраты той же самой функции второй копией. Что, впрочем, не мешает второй копии потерять какую-нибудь другую функцию, пока еще сохранившуюся у первой копии. Если это произойдет, копии перестанут быть «избыточными». Теперь ни одна из них не может быть утрачена без резкого снижения приспособленности (а значит, отбор не позволит им потеряться). В итоге вместо одного «неразборчивого», многофункционального белка организм получает два специализированных.
Самое интересное, что это усложнение молекулярной организации вовсе не обязано быть «полезным», то есть повышать приспособленность организма. Вполне возможно, что после дупликации и разделения функций организм будет чувствовать себя ничуть не лучше, чем до этих событий. Например, в описанных экспериментах дрожжи, у которых белки Mcm1 и Arg80 были заменены на предковый AncMADS, росли не хуже обычных. В отдаленной перспективе дупликация и субфункционализация могут открыть новые эволюционные возможности, ведь две копии исходного гена (и их функции) могут теперь эволюционировать более или менее независимо. Но естественный отбор ничего не знает об отдаленных перспективах, он ориентируется только на «здесь и сейчас». Усложнение молекулярной организации в данном сценарии является не более чем побочным (хотя и закономерным) следствием случайного удвоения гена.
Не остановившись на достигнутом, авторы решили проверить гипотезу, согласно которой после дупликации многофункциональных белков, таких как AncMADS, вовлеченных к тому же в сложную сеть взаимодействий с другими белками, возникшие паралоги могут конфликтовать и мешать друг другу работать. Например, новообразовавшийся паралог AncArg80, разучившийся взаимодействовать с кофактором Matα1, мог бы «по ошибке» присоединяться к регуляторной области α-генов, занимая то место на ДНК, куда должен прикрепляться другой паралог, AncMcm1. В результате регуляция α-генов оказалась бы нарушена. Если такой конфликт между паралогами действительно когда-то существовал, то в дальнейшем отбор должен был поддержать и закрепить мутации, снижающие его остроту.
В поисках таких мутаций авторы обратили внимание на ту часть молекулы AncArg80, которая отвечает за прикрепление белка к ДНК. Здесь обнаружилось пять аминокислотных замен, которые делают связь белка с ДНК менее прочной. Может быть, именно благодаря этим заменам белок AncMcm1 и его потомки, сохранившие способность крепко цепляться за ДНК, и одерживают верх над Arg80 в конкуренции за право присоединиться к регуляторной области α-генов? Чтобы проверить эту смелую гипотезу, авторы сконструировали версию AncArg80, лишенную вышеупомянутых пяти замен в ДНК-связывающей области. Ген этого белка внедрили в геном дрожжей вместо изъятого оттуда гена Arg80. Результат подтвердил ожидания исследователей: у модифицированных дрожжей регуляция α-генов оказалась полностью нарушена. Когда пять мутаций вернули на место, α-гены снова стали работать нормально.
Следовательно, адаптивный смысл пяти замен, ослабивших связь Arg80 с ДНК, скорее всего, действительно состоял в том, чтобы сгладить конфликт между паралогами. Благодаря этим заменам Arg80 не лезет не в свое дело и не мешает другому паралогу, Mcm1, регулировать работу α-генов. Можно сказать, что закрепление этих пяти замен окончательно превратило копии удвоившегося AncMADS в два разных белка. Приняв такую формулировку, можно согласиться с авторами, полагающими, что в данном случае конфликт между паралогами способствовал усложнению системы генной регуляции (усложнение, напомним, состояло в том, что ARG-гены, для управления которыми ранее хватало двух белков-регуляторов, теперь стали контролироваться тремя).
Кроме того, ослабление связи Arg80 с ДНК объясняет, почему ARG-гены регулируются гетеродимером Mcm1+Arg80, а не гомодимером Arg80+Arg80. Возможно, белок Mcm1 просто-напросто помогает Arg80 удерживаться на ДНК (на регуляторных участках ARG-генов). Что мешает гомодимерам Mcm1+Mcm1 присоединяться к этим участкам, конкурируя с гетеродимерами и мешая работе ARG-генов, — пока неясно.
По мнению авторов, конфликты между паралогами могут быть важным эволюционным фактором, ограничивающим и направляющим процесс формирования эволюционных новшеств путем дупликации генов.
Источник: Christopher R. Baker, Victor Hanson-Smith, Alexander D. Johnson. Following Gene Duplication, Paralog Interference Constrains Tranional Circuit Evolution // Science. 2013. V. 342. P. 104–108.
См. также об эволюционной роли дупликации генов:
1) Многофункциональные гены — основа для эволюционных новшеств, «Элементы», 30.06.2008.
2) В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшества, «Элементы», 25.09.2012.
3) Процесс появления новых ферментов прослежен в эволюционном эксперименте, «Элементы», 23.10.2012.
4) Удвоение генов может приводить к видообразованию, «Элементы», 04.02.2009.
5) Прослежена эволюционная история одного из человеческих генов, «Элементы», 17.06.2008.
6) Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных, «Элементы», 23.06.2008.
7) Обоняние и цветное зрение в эволюции млекопитающих развивались в противофазе, «Элементы», 18.06.2008.
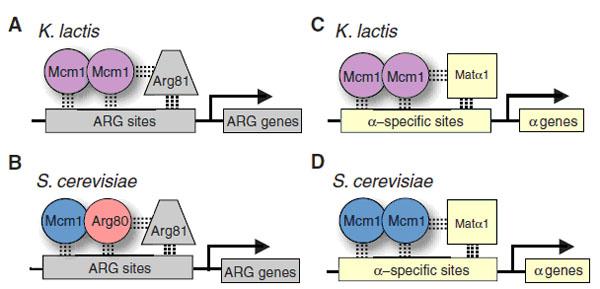
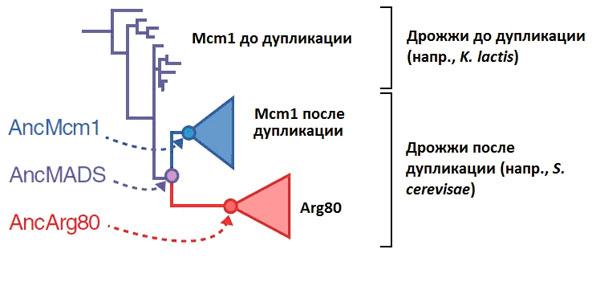
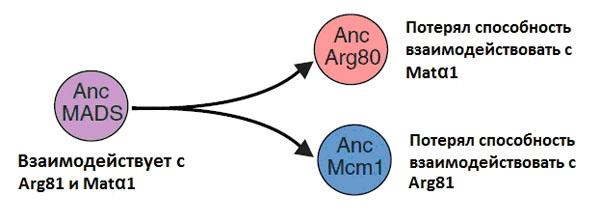
Рис. 1. Схема регуляции работы генов метаболизма аргинина (ARG genes) и генов, связанных с половым размножением (α genes), у дрожжей Kluyveromyces lactis и Saccharomyces cerevisiae. Комплексы белков-регуляторов присоединяются к регуляторным участкам ДНК (ARG sites, α-specific sites) возле контролируемых генов. У K. lactis обе группы генов (ARG и α) регулируются комплексами из двух молекул белка Mcm1 (гомодимерами). У S. cerevisae гены α регулируются такими же гомодимерами, а для регуляции генов ARG используется гетеродимер, состоящий из Mcm1 и дополнительного регуляторного белка Arg80, которого нет у K. lactis. Ген Arg80 появился у предков S. cerevisae в результате дупликации исходного гена Mcm1 и последующего разделения функций. Рисунок из обсуждаемой статьи в Science