Мозговая основа субъективных переживаний
Мозговая основа субъективных переживаний
Выделения в тексте - мои.<br>Мои коммнтарии включены синим цветом.
http://www.aha.ru/~geivanit/SUBJ.html
Иваницкий А.М.
Мозговая основа
субъективных переживаний:
гипотеза информационного синтеза
Содержание:
Информационный синтез как
основа ощущений
Идея повторного входа
возбуждения как мозговой основе психики
Исследование механизмов
мышления. Фокусы взаимодействия
Проблема "я"
Сознание и речь
Функциональное значение
психических переживаний
Статья посвящена изложению принципов организации мозговых процессов, лежащих в основе субъективных переживаний. Согласно развиваемой концепции, они возникают как результат синтеза различных видов информации в ключевых для данной психической функции зонах коры. В информационном синтезе участвует информация, исходящая из внешней среды, извлекаемая из памяти и поступающая от центров мотиваций. Концепция построена на основе исследований мозговых механизмов восприятия и мышления. Показано, что ощущение возникает в результате синтеза на нейронах проекционной коры сведений о физических и сигнальных свойствах стимула, который обеспечивается кольцевым движением возбуждения из проекционной в ассоциативную кору, гиппокамп и мотивационные центры с возвратом в проекционную кору. Установлено также, что при мышлении происходит конвергенция корковых связей к определенным центрам, названным фокусами взаимодействия. Их топография специфична для различных мыслительных операций: при образном мышлении фокусы располагаются в теменно-височной, а при абстрактном мышлении - в лобной коре. Предполагается, что в фокусах взаимодействия осуществляется сопоставление и синтез приходящей по нервным связям информации, что и лежит в основе нахождения решения. Последний раздел статьи посвящен вопросу о функциональном значении психических феноменов и их возможном влиянии на процессы мозга.
Происхождение и функциональный смысл субъективных переживаний - одна из загадок нашего мозга. Нужны ли они только для того, чтобы жизнь имела свою ценность (вспомним слова А.С.Пушкина "я жить хочу, чтоб мыслить и страдать"), или они составляют необходимый компонент работы мозга, обладая свойством изменять или, как предполагают многие исследователи, даже направлять течение физиологических процессов.
Опыт веков показывает, что эти вопросы не могут быть решены путем чистой дедукции и философских построений. Путь к разгадке изучение работы мозга объективными методами и, прежде всего, анализ того, как возникают субъективные переживания, каков их мозговой механизм. История науки учит, что в основе возникновения нового качества лежит, как правило, достаточно простой принцип организации. Примером может служить двойная спираль и кодирование наследственной информации триплетами пуриновых и пиримидиновых оснований. Поскольку мозг - это орган обработки информации, логично предположить, что возникновение психики связано с некоторым принципом организации информационных процессов мозга.
В последние годы все большее признание получает идея о том, что субъективный опыт возникает за счет сопоставления имевшийся ранее информации с новой, отражающей изменения во внешней или внутренней среде. Сама по себе эта идея не нова. Еще Д.Юм /33/ писал о том, что чувство "я", важнейший элемент субъективного опыта, возникает в результате движения восприятий вдоль событий прошлого. Исторический анализ проблемы однако выходит за рамки настоящей статьи. Обращаясь к новейшим исследования по физиологическим механизмам психического, следует сказать об информационной теории эмоций П.В.Симонова /27/, в соответствии с которой эмоции возникают как результат рассогласования между имеющейся и необходимой для удовлетворения потребности информации. Позднее этим автором были описаны и корковые структуры, ответственные за интеграцию каждой из информационной посылок (соответственно передние отделы левого и правого полушарий /29/).
Информационный синтез как основа ощущений
Идея об информационном синтезе как мозговой основе возникновения субъективных переживаний была выдвинута нами в 1976 году /7, 14/ в результате исследований физиологических механизмов ощущений - одного из наиболее элементарных психических проявлений. Психологам с 20-х годов известно, что оно возникает довольно поздно, после 100 мс от момента стимула, то есть значительно позднее прихода сенсорных импульсов в кору. В работе была поставлена задача понять, что происходит за это время и какой стадии мозговых процессов соответствует возникновение субъективного образа. В проведенных экспериментах на одни и те же стимулы записывались объективные показатели мозговой деятельности в виде вызванных потенциалов и определялись количественные показатели восприятия. Для получения последних использовались методы теории обнаружения сигнала /61/, которая описывает процесс восприятия с помощью двух независимых переменных: показателя сенсорной чувствительности d' и критерия решения, определяемого мотивационными факторами. Субъект должен был различать стимулы по их силе и нажимать кнопку правой или левой рукой, в зависимости от интенсивности возникающих ощущений. Затем вычислялись корреляции между показателя физиологии и психологии, а именно амплитудой волн вызванного потенциала и психофизическими индексами. Исследования проведены на соматосенсорном /12/ и на зрительном /11/ анализаторах, причем полученные на обеих модальностях результаты были принципиально сходными.
Было установлено, что амплитуда ранних волн вызванного потенциала (ВП) обнаруживала статистически достоверную корреляцию с показателем d', а поздних волн - с критерием решения. Промежуточные волны с латентностью 140 мс для соматосенсорного и 180 мс для зрительного анализатора коррелировали с обоими перцептивными индексами, причем такая двойная корреляция обнаруживалась только для волн проекционной коры. Амплитуда этих волн определялась, таким образом, как сенсорными свойствами стимула, так и его значимостью. На основании имеющихся данных о генезе отдельных волн вызванного потенциала был описан механизм, обеспечивающий указанную двойную корреляцию.
В основе этого механизма лежало кольцевое движение нервных импульсов с "центральной станцией" в проекционной коре. Сначала возбуждение поступало от проекционной к ассоциативной коре. Для зрительных стимулов это были отделы височной коры, играющей важную роль в опознании стимулов, а для кожных раздражений вторичные и третичные зоны соматосенсорной коры. Затем нервные импульсы поступали к структурам лимбико-гиппокампального комплекса и подкорковым центрам эмоций и мотиваций. До этой стадии поступательное движение возбуждения от рецептивных зон к исполнительным центрам полностью укладывалось в схему рефлекса. Однако процесс на этом не заканчивался: он включал еще одно важное звено, превращающее дугу в кольцо, а именно возврат возбуждения в кору, включая и ее проекционные отделы, по системе диффузных проекций. Это звено представляло собой, таким образом, обратную связь от исполнительных к афферентным центрам. Благодаря возврату возбуждения нервные импульсы, приходящие из мотивационных центров накладывались на нейронах проекционной коры на следы сенсорного возбуждения. На это этапе или несколько раньше в процесс включалась и лобная кора, что проявлялось в синхронизации рисунка ВП в интервале от 100 до 200 мс после стимула.
Было высказано предположение, что указанные промежуточные компоненты ВП отражали синтез на корковых нейронах двух видов информации: наличной информации о физических характеристиках стимула и извлекаемых из памяти сведений о его значимости.
До сих пор исследование развивалось по законам рационального, дедуктивного мышления. Но в этом месте нас ожидал сюрприз, как бы заготовленный природой для тех, кто пойдет выбранным нами путем. Оказалось, что латентность волн ВП, обнаруживших двойную корреляцию с психофизическими показателями, точно совпала со временем ощущения, измеренном ранее в психологических экспериментах /4, 45, 56/. Было сделано предположение, что синтез двух видов информации: наличной и извлекаемой из памяти - и составляет тот ключевой механизм, который лежит в основе ощущения как феномена уже не физиологического, а психического уровня (рис.1). Тем самым был сделан шаг к преодолению барьера между этими двумя уровнями мозговой интеграции. Согласно предложенной концепции, последовательное поступление информации от рецепторов приводит к повторному движению возбуждения по указанному кольцу, обеспечивая постоянное сопоставление сигналов, приходящих из внешней и внутренней среды, что и составляет психический мониторинг происходящих изменений. Этот процесс осуществляется с периодом квантования приблизительно в 100-150 мс, составляющих минимальную длительность ощущений /36/.
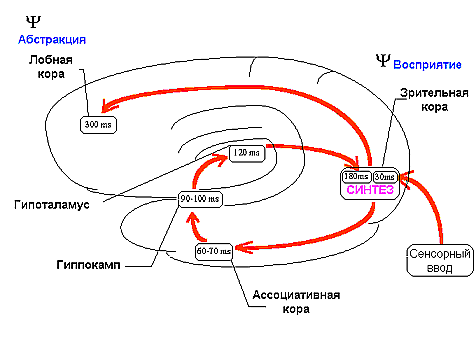 Рис.1. Схема кольцевого движения
возбуждения при возникновении ощущений. Ведущее звено в этом процессе - синтез
информации о физических и сигнальных свойствах стимула на нейронах проекционной
коры.
Рис.1. Схема кольцевого движения
возбуждения при возникновении ощущений. Ведущее звено в этом процессе - синтез
информации о физических и сигнальных свойствах стимула на нейронах проекционной
коры.
В то время мы не имели возможности досконально проверить данную гипотезу, включая верифицированное описание движения возбуждения по структурам мозга человека, в связи с понятными ограничениями этического характера. Однако в настоящее время есть смысл вновь возвратиться к этим представлениям, так как за истекшие годы в литературе появились данные, прямо или косвенно подтверждающие наши представления . Эти подтверждения касаются как описанного нами кольца возбуждения, так и значения этого механизма для возникновения субъективных феноменов.
Для ответа на первый из поставленных вопросов важны исследования М.Мишкина /53/, исследовавшего процесс образования памятного следа действующего раздражителя у обезьян. По данным этого автора, в ответ на стимул возбуждение из проекционной коры распространяется на ростральную темпоро-инсулярную область, откуда проекции подходят к медиальной темпоральной зоне, представленной ринальной корой (а не гиппокампом, как предполагалось ранее). Затем возбуждение переходит на медиальные диэнцефальные структуры и возвращается в кору к медиально-префронтальным ее отделам. Возбуждение последних активирует базальную холинэргическую систему, иннервирующую весь мозговой плащ, что составляет последний, завершающий этап обработки стимульной информации. При повторном действии стимулов синаптическая проводимость задействованных хеббовских нейронов повышается за счет процессов фосфорилизации и синтеза синаптических протеинов, что и лежит в основе памятного следа. Схема, предложенная М.Мишкиным, значительно более детализирована, по сравнению с нашей, что объясняется возможностью непосредственного отведения активности от подкорковых отделов мозга у обезьян. Но нетрудно видеть, что в своих принципиальных положениях она достаточно близка к предложенному нами кольцу возбуждения. В своей работе М.Мишкин не касается психических аспектов описываемого процесса, что и понятно, учитывая использованный им объект исследования.
Идея повторного входа возбуждения как мозговой основе психики
Идея о повторном входе возбуждения в нервные структуры как базисном механизме возникновения субъективного опыта была высказана в последние годы, независимо от нас, рядом авторов. В большинстве случаев авторы основывали при этом свои взгляды на общетеоретических соображениях, а не на результатах прямого сопоставления физиологических и психологических данных в одном эксперименте, как это было сделано в проведенном нами исследовании. Наиболее полно идея повторного входа была развита в трудах Дж.Эдельмена /32, 43/, а созданная на ее основе теория сознания получила достаточно широкую известность. По мнению этого автора, в основе возникновения субъективных феноменов лежит один и тот же механизм повторного входа возбуждения в те же нейронные группы после дополнительной обработки информации в других группах или поступления сигналов из внешней среды. Этот повторный вход (английский эквивалент - reentering) дает возможность сравнивать имевшиеся ранее сведения с изменениями, происшедшими в течение одно цикла. Постоянный мониторинг этих изменений в результате повторных входов, и лежит, по мнению автора, в основе континуума психических переживаний. Подкорковые структуры, которые участвуют в циклическом движении возбуждения различны для разных функций. В качестве ведущих называются гиппокамп (явления памяти), мозжечок (двигательная сфера) и таламус (восприятие).
Сходная концепция сознания разрабатывается в нашей стране В.Сергиным /24/, который также придает решающее значение в возникновении субъективных переживаний циклическому движению процесса возбуждения, что создает феномен внутреннего видения, составляющего суть сознания. Гипотеза о возврате возбуждения в первичную коры как механизме возникновения зрительных ощущений была высказана также П.Стериг и С.Брандтом /60/. Авторы считают, что эти обратные проекции менее дифференцированы и носят диффузный характер, снабжая информацией различные звенья зрительной системы, способствуя их интеграции. В исследованиях на обезьянах /38/ было показано, что компонент соматосенсорного ВП N1 с латентностью 50 мс отражает возврат в первичную проекционную кору возбуждения из вторичных полей, что, по мнению авторов, лежит в основе "осознаваемых" тактильного ощущения. Последнее доказывается тем, что эта волна исчезала при сне и анестезии и соответствовала последующему поведенческому ответу после обучения обезьян различать предъявляемые стимулы.
Сходная схема мозговой организации сознания была описана Дж.Десмедтом и К.Томбергом /42/, которые считают, что в основе осознаваемых феноменов лежит повторный вход возбуждения из дорсолатеральных отделов префронтальной коры к местам проекции сенсорных сигналов. При этом происходит синхронизация потенциалов на частоте в 40 гц.
Достаточно хорошо проработанная с концептуальной и нейрофизиологической точки зрения концепция сознания была выдвинута в последнее время Дж.Греем /47/. В соответствии с этой концепцией, содержание сознания определяется активностью субикулярного (отдел гиппокампа) компаратора вместе с обратной связью от данного компаратора к тем наборам нейронов в перцептивной системе, которые снабжают вход данного компаратора с учетом результатов текущего процесса сравнения. Идея о важной роли лимбических структур в генезе психики согласуется и с данными /54/, что эти структуры тесно связаны с эксплицитной памятью, узнаванием и вспоминанием. В то же вряд ли гиппокампу может принадлежать главная роль в высших функциях мозга. Поэтому представления о синтезе информации в коре как ведущем звене в механизмах психики, с нашей точки зрения, заслуживают предпочтения, тем более, что они хорошо согласуются и с данными других авторов, приведенными выше.
Для оценки предложенных концепций важное значение может иметь соотнесение временной шкалы описываемых ими мозговых процессов со временем субъективно переживаемых событий. По Дж.Эдельмену, время одного цикла прохождения возбуждения составляет 100 - 150 мс. Если прибавить к этому время, необходимое для прихода сенсорных импульсов в кору, то в сумме получается время, что достаточно близкое к тому, которые было найдено в наших экспериментах. Дж.Грей считает, что в квантовании процессов сознания ведущую роль играют процессы, связанные с частым тета-ритмом, что дает время 1000:6 = 166мс. О значении частот тета-ритма в данном аспекте в свое время писал и П.В.Симонов /26/. В цитированной выше работе Дж.Десмедта и К.Томберга процесс синхронизации на частоте 40 гц развивался в течение 100 мс после появления потенциала в первичной коре и до начала волны Р300, то есть также в интервале 100-200 мс.
Представляют также интерес следующие экспериментальные факты. В исследованиях Б.Либета и соавт. /51/ с регистрацией вызванных потенциалов на электрокожные стимулы с поверхности коры во время нейрохирургических операций было показано, что на слабые, подпороговые стимулы в коре регистрируются только ранние волны ответа с латентностью до 100 мс. При усилении раздражений в ВП появляются и более поздние колебания с латентностью 150 мс, что сопровождается появлением субъективных ощущений и практически совпадает с латентностью волн ВП, обнаруживших в наших исследованиях двойную корреляцию с перцептивными индексами. Правда, основываясь на результатах прямой электростимуляции соматосенсорной коры, Б.Либет делает вывод, что субъективные события запаздывают по отношению к мозговым событиям на 500 мс. Г.Хесслоу /48/ однако считает, что это время слишком велико и что при непосредственном раздражении коры деятельность тонких нейронных схем, лежацих в основе ощущений, может быть нарушена.
В исследованиях Б.Базияна /2/, изучавшего феномен подавления зрительных образов при движениях глаз, было установлено, что ранние компоненты зрительного ВП при этом не изменяются, зато более поздние волны с латентностью более 100 мс достоверно уменьшаются по амплитуде. Наконец, в исследованиях И.Циглера и Г.Чибра /40/ было показано, что при предъявлении серии зрительных стимулов на раздражители, отличающиеся по параметрам от большинства сигналов, в задних отделах полушарий возникает отрицательное колебание в интервале 140-180 мс после раздражения. Данное колебание авторы связывают со зрительным вариантом известного феномена "негативности процессинга" /55/, отражающего, в частности, процессы селективного внимания и сравнения наличных свойств стимула с памятью. Все приведенные данные, как нам представляется, хорошо согласуются с результами наших исследований.
У человека возникшее ощущение затем опознается, категоризируется, что происходит при ведущем участии лобных отделов коры. В проведенных нами исследованиях это составляло третий этап восприятия, который характеризовался корреляцией показателя критерия решения в поздними волнами ВП, включая волну Р300. В этих процессах значительную роль играют и вербальные отделы коры. Последнее, в частности, было показано в работе Р.Салмелина и соавт. /58/. Авторы регистрировали магнитные поля мозга при рассматривании субъектом картинок различного содержания. При этом, даже если называть изображенный объект не следовало, реакция переходила на вербальные зоны левого полушария. Это происходило на 400 мс после стимула, то есть на 200 мс позднее возникновения ощущений, которому в описываемых экспериментах соответствовала реакция зрительных и теменно-височных отделов коры с латентностью около 200 мс.
В работе по исследованию ВП при опознании зашумленных стимулов, содержащих или не содержащих элементы изображения животного /63/ было показано, что различия в рисунке ВП начинаются с 150 мс, что можно трактовать как начало процесса опознания возникшего ощущения. Б.Баарс /34/ на основе анализа психологической литературы приходит к выводу, что образы возникают в течение первых 200 мс после действия стимула, а в последующем (200-500 мс) происходит их категоризация. Можно поэтому предполагать, что на протяжении "одного кванта" субъективных переживаний человек находится на довербальной стадии психического. Временная развертка, следовательно, показывает, что более сложные психические процессы не отменяют более простых форм, а как бы надстраиваются над ними, о чем подробнее будет сказано ниже.
Наконец, еще о двух работах, которые могут рассматриваться как косвенно подтверждение нашей гипотезы. Речь идет об исследованиях, в которых показано, что возбуждение нейронов первичной коры, вопреки распространенному убеждению, не достаточно для возникновения ощущения, хотя и является его необходимым условием. Согласно гипотезе Ф.Крика и К.Коха /39/, для возникновения зрительного ощущения необходима согласованная работа зрительной коры и гиппокампа, а также прямые связи с лобной корой. Наличие таких прямых связей показано для полей V4 и MT (и, возможно, для более ранних полей), но не поля V1. Все указанные отделы зрительной системы, которые захватываются обратными проекциями, согласно данным П.Стериг и С.Брандта.
В доказательство своих взглядов Ф.Крик и К.Кох приводят данные ряда психофизических экспериментов. Так установлено, что при предъявлении зрительных стимулов в виде решеток разной частоты имеет место маскировочный эффект более частой решетки по отношению к более редкой, хотя первая и воспринимается в виде серого фона. Авторы объясняют это тем, что маскировка происходит на уровне высокоразрешающих нейронов первичной коры, которые сами по себе не продуцируют ощущений. Известно также, что изображения на сетчатке и соответственно на ее проекциях в первичной коре не идентичны для обоих глаз, что и лежит в основе бинокулярного зрения. Тем не менее мы видим одно изображение, которое возникает позднее поля V1.
В работе Е.В.Дамянович /6/, проведенной на больных с потерей кожной чувствительности после мозговых инсультов во внутреннюю капсулу, было показано, что у части этих больных в проекционной коре может быть зарегистрирован соматосенсорный ВП, начиная с ранних его компонентов (правда, без ранней волна N19, которая в исследованиях В.Б.Стрелец обнаружила высокую корреляцию с d'). Это может служить прямым доказательством того, что приход импульсов в первичную кору еще не достаточен для появления ощущения. Важность этих работ в том, что они показывают, что психические процессы на отслеживают соответствующие физиологические, а требуют особой организации и взаимодействия мозговых структур. Об этом же говорят и приведенные в работе П.Стериг и С.Брандта /60/ данные о том, что при некоторых поражениях первичной зрительной коры имеет место характерное нарушение зрительных функций, названное слеповидением, в отличие от слепоты (blindsight vs. blindness). При этом больной говорит, что он ничего не видит, в то время как у него могут быть выработаны реакции на определенное положение и цвет стимула. Эти работы свидетельствуют, что вопрос о генезе психики не только теоретический, его решение необходимо практике для понимания происхождения неврологических синдромов, не говоря уже о психических заболеваниях.
Таким образом, можно констатировать, что экспериментальный материал, хотя он и был получен разными методами, достаточно однозначен, а созданные на его базе концепции мозговых основ психики весьма близки друг к другу, хотя и были созданы учеными, принадлежавшими к разным школам. Все это внушает оптимизм и может рассматриваться как признак приближения к истинному пониманию того ключевого механизма, который лежит в основе появления субъективного опыта: психическое возникает на базе сопоставления, синтеза наличной информации и сведений, извлекаемых из памяти, что обеспечивается кольцевым движением нервных импульсов с повторным входом возбуждения в места первоначальной проекции. В генезе ощущений ключевым звеном является синтез информации в проекционной коре, благодаря чему ощущение соединяет в себе физические характеристики стимула и его чувственную окраску. Существенно при этом, что значимость стимула, хотя она и принимает участие в формировании ощущений, представлена в нем на данном этапе восприятия в неявной, имплицитной форме. Отметим также, что представление о генезе ощущений как результате синтеза различных свойств стимула близко и к идеям П.К.Анохина /1/ о природе психического как обобщении имеющейся информации, благодаря чему оно и выступает как важная детерминанта поведения.
Исследование механизмов мышления. Фокусы взаимодействия корковые динамические образования, осуществляющие синтез информации
Гипотеза информационного синтеза позднее была развита нами применительно к изучению мозговых механизмов мыслительных операций. Эти исследования были проведены, главным образом, с помощью картирования потенциалов мозга созданным в лаборатории методом, получившим название картирования внутрикоркового взаимодействия /9/. Метод развивает известные представления о том, что синхронизация потенциалов способствует установлению связи между структурами мозга /19, 23/. Метод основан на следующих теоретических предпосылках. (1) Группы нейронов в коре имеют функциональную специализацию, и каждая из них вносит свой вклад в обработку информации. Эта специализация определяется как корковой топографией группы, что особенно выражено для проекционных областей, так и приобретенными в онтогенезе в процессе обучения свойствами. Эти идеи близки к теории селекции нейронных групп Д.Эдельмена. Теоретические и экспериментальные данные указывают, что психические функции возникают, в результате объединения этих нейронных групп в единую систему. (2) Клетки внутри группы соединены системой прямых и обратных связей таким образом, что группа приобретает свойство нейронного осциллятора /15, 16, 35, 44, 52/, который характеризуется присущей ему частотой разрядов. Как правило, эта частота значительно ниже частоты разрядов отдельных нейронов и приближается к частотам ЭЭГ. Вот и серьезное обоснование мозговых регистрируемых ритмов.
Часть корковых нейронов обладает также свойством пейсмекеров. Методы частотного анализа ЭЭГ и ВП, такие, как быстрое преобразование Фурье, выявляют работу основных корковых осцилляторов, что проявляется в виде компонентов спектра. (4) Точное совпадение частотных характеристик частотных осцилляторов способствует установлению между ними функциональной связи. Это объясняется тем, что при этом сигналы от одной нейронной группы достигают другой в одной и той же фазе ее возбудимости. В случае фазы экзальтации порог возбудимости нейронов второй группы оказывается минимальным, что способствует ее вовлечения в совместную с первой группой деятельность. В случае фазы рефрактерности связь приобретает латентный, тормозный характер. Предполагается, что переход от одного к другому виду связи может быть достаточно быстрым и включать несколько циклов за достаточно короткий интервал времени, сопоставимый со скоростью протекания психических функций.
Основываясь на этих положениях, можно таким образом придти к выводу, что точное совпадение частотных пиков спектра разных областей коры свидетельствует о том, что там находятся группы нейронов, функционально связанные друг с другом. Делая акцент на на совпадении частотных характеристик спектра, метод таким образом как бы инвариантен к изменению фаз, выявляя достаточно устойчивые как возбудительные, так и тормозные взаимодействия между корковыми полями. В этом заключается принципиальное отличие данного метода от достаточно широко используемого метода когерентности, который выявляет связанные на базе синхронизации активности корковые пункты лишь в том случае, если разность фаз за это время оставалась неизменной. В противном случае связь будет утеряна, что, по мнению Буллока /37, может привести к ошибкам.
Идеи метода картирования внутрикоркового взаимодействия были реализованы в соответствующих компьютерных программах. Метод был использован для изучения корковых связей при различном типе мыслительных операций. Испытуемым предъявлялись на экране монитора задачи на образное, пространственное и абстрактно-вербальное мышление. В первом случае субъект должен был опознать эмоцию на фотографии лица (использовались четыре базовые эмоции: радости, страха, гнева и горя и смешанные состояния),во втором - сравнение двух геометрических фигуры целью определения их идентичности или зеркальной симметрии. В качестве вербальных задач использовано решение анаграмм или выбор из четырех слов одного, относящегося к другой смысловой группе.
Было установлено, что простой и достаточно симметричный рисунок связей, характерный для состояния покоя, при умственной работе изменялся. Связи начинали сходиться к определенным областям коры, образуя как бы узлы или центры связей, названные фокусами взаимодействия. При этом топография фокусов взаимодействия оказалась специфичной для мыслительных операций разного знака. Так при образном мышлении фокусы локализовались преимущественно в теменно-височной области, а при абстрактно-вербальном мышлении - в лобных отделах коры /9, 20, 21, 25/ (рис.2).
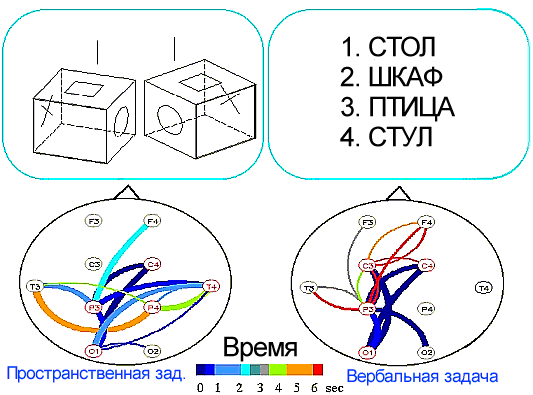
Рис.2. Корковые связи в диапазоне частот бета-ритма при двух мыслительных операциях, образцы которых представлены вверху рисунка: связи при задаче на пространственное мышление, которая заключалась в сравнении двух геометрических фигур, предъявляемых на экране монитора, с целью определения их идентичности или зеркальной симметрии, что решалось путем мысленной ротации. Справа связи при задаче на вербальное мышление, которая состояла в том, что из четырех слов на экране монитора субъект должен был выбрать одно, относящееся к другой смысловой группе. На рисунке даны статистически значимые связи при сравнении со зрительно-моторным контролем, включавшим предъявление изображений и двигательный ответ, но не требовавшим мыслительных операций. Средние данные по группе из 43 человек.
На основе обобщения полученных данных было сформулировано представление о двух когнитивных системах мозга. Первая из них, связанная с височно-теменными областями коры, ответственна за процессы образного мышления. Вторая система, система абстрактно-вербального мышления, локализуется в лобных отделах коры. Системы расположены, таким образом, вдоль передне-задней оси мозга, и такое деление носит эволюционно более древний характер, чем специализация полушарий. Различие между системами, помимо их функции и топографии, заключается еще и в том, что образное мышление преимущественно интуитивно, имплицитно, происходит без ясного осознания последовательности мыслительных операций. Абстрактно-вербальное мышление, напротив, рационально, эксплицитно, так как мыслительные операции как бы контролируются субъектом.
Существенно, что включение "не той" когнитивной системы, что проявляется в локализации фокусов в лобной коре при задачах на образное мышление и в височно-теменной коре при вербальных задачах приводило к тому, что решение не могло быть найдено.
Специализация полушарных функций, связанная с возникновением речи, как бы накладывается на особенности функционирования когнитивных систем. В исследованиях, где испытуемому давалось задание мысленно построить зрительный образ из ограниченного набора простых элементов, было показано, что у лиц с преобладанием первой сигнальной системы по И.П.Павлову фокусы взаимодействия локализовались преимущественно в правом, а у лиц с преобладанием второй, речевой системы - в левом полушарии.При этом на этапе нахождения образа фокусы были расположены в затылочных и височных (зона опознания) отделах полушарий, а на этапе конструирования образа - в лобной коре /13/. Нахождение решение при всех типах задач, даже если речевой ответ не требовался, сопровождалось (или определялось?) включением в функция вербальной левой височной зоны.
Изложенные представления о двух когнитивных системах мозга хорошо согласуются и с данными других авторов, в частности М.Познера и М.Ротбарта /57/, использовавших метод позитронно-эмиссионной томографии. Имея высокое пространственное разрешение и возможность получения трехмерных изображений, эти исследования вносят значительный вклад в понимание мозговых основ высших психических функций. Но определенные преимущества имеют и исследования первичных эффектов нервного возбуждения в виде электрических потенциалов или магнитных полей мозга. Помимо высокого разрешения во времени, эти исследования позволяют ответить на вопрос не только "где", но и "как" происходит обработки информации в мозге. При этом через исследование рисунка нервных связей можно выйти и на описание принципов организации нервных процессов, лежащих в основе психики.
Как уже говорилось, одним из принципов такой организации является конвергенция нервных связей к определенным центрам фокусам взаимодействия. При этом связи, подходящие к фокусу, устанавливаются на разных частотах, собственно именно это обстоятельство и лежит в основе образования фокуса, так как связи на одной частоте образовывали бы однородную сеть, не имеющую центров. Можно предполагать, что каждая из связей приносит к центру из определенной области коры или подкорковых образований свою информацию. В фокусе эта информация может быть сопоставлена и перекомбинирована определенным образом. Основную функцию фокуса взаимодействия составляет, таким образом, информационный синтез, то есть процесс, сходный с тем, который мы наблюдали в проекционной коре при возникновении ощущений.
Основная разница заключалась в том, что место сенсорного сигнала здесь могла занять информация, хранящаяся в оперативной памяти (например, об условиях решаемой задачи), а ведущую роль в процессах информационного синтеза принадлежала не проекционной, как при возникновении ощущений, а ассоциативной коре. В фокусе оперативная информация сопоставлялась с информацией, извлекаемой из долговременной памяти и сигналами, приходящими из мотивационных центров. Предполагается, что на основе происходящего в фокусе сопоставления и достигается конечная цель мыслительного процесса в виде нахождения решения. Субъективно все это переживается как процесс думания и нахождения ответа.
С физиологической точки зрения, фокус выполняет, таким образом, функции, аналогичные командному нейрону у низших животных /30, 50/. Однако сложность и многообразие приходящей информации у человека и высших животных требуют и соответствующей более сложной структуры. Гипотетически эта структура состоит из групп нейронов с различной частотной характеристикой, настроенных на изолабильные им группы на периферии. Эти связи по своей природе должны быть двусторонними, то есть как прямыми, так и обратными: если две группы имеют одну частоту, они в равной степени способны и воспринимать, и передавать информацию связанной с ней группе, в зависимости от соотношения фаз колебаний. Одно кольцо при ощущении заменяется здесь, таким образом, как бы системой колец, замыкающихся на один центр.
Внутри фокуса группы нейронов должны быть объединены связями, образованными на ином принципе: так как они работают на разных частотах, принцип изолабильности здесь не применим. Очевидно, это должны быть жесткие связи, основанные на структурных изменениях в синапсах. Данные связи эффективны в любой фазе цикла возбудимости нейрона или нейронного осциллятора, за исключением абсолютной рефрактерной фазы (рис.3). Идея об обеспечении психической функции за счет сочетания жестких и гибких связей было высказано впервые Н.П.Бехтеревой /3/.
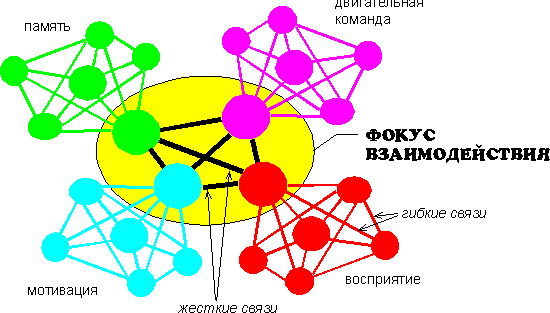
Рис.3. Гипотетическая схема фокуса взаимодействия. Фокус состоит их групп нервных клеток, отличающихся различными частотными параметрами и связанных с группами периферических нейронов гибкими связями на основе идентичности частотных характеристик. Группы внутри фокуса соединены жесткими связями, основанными на структурных изменениях в синапсах. Благодаря такой структуре фокус объединяет и синтезирует информацию, циркулирующую в различных нейронных сетях, на основе чего достигается решение.
Высказанные представления о фокусах взаимодействия и их функциональном значении хорошо согласуются с данными Х. и А. Дамасио /41/, которые, используя метод функционального магнитного резонанса, пришли к выводу, что выявляемые ими при выполнении субъектом различных психологических тестов активные зоны мозга представляют собой, на самом деле, лишь зоны конвергенции различных видов информации. Термин "фокуса" в сходном аспекте используют в своей работе и А.Гевинс и соавт. /46/. Все сказанное, как нам представляется, дает возможность объединить данные, полученные при изучении механизмов восприятия и мышления, единым принципом информационного синтеза как мозговой основы возникновения нового качества в виде субъективных переживаний.
Предложенная нами и использованная в концепции фокусов взаимодействия схема нейронной сети, состоящей из нейронов различной лабильности и построенной по иерархическому принципу, имеет ряд преимуществ, по сравнению с однородной и изолабильной нервной сетью. Главный выигрыш здесь - в высокой информационной емкости, что преодолевает одну из главных трудностей гипотезы о кодировании психических состояний гомогенной нервной сетью. Идея о фокусах взаимодействия хорошо согласуется и с представлениями И.Пригожина и Стенгерс /22 / о так называемых диссипативных структурах, возникающих из хаоса на основе принципа самоорганизации. Соответственно фокусы взаимодействия могут возникать на основе самоорганизации нейронных групп различной лабильности в процессе обучения определенным навыкам, составляя корковое ядро соответствующей психической функции. Вокруг этого жесткого ядра образуется система гибких связей, совокупность которых определяет качественное своеобразие и неповторимость переживаемого психического состояния.
Проблема "я"
По мнению Г.Свенссона /61/, любая теория, которая претендует на объяснение природы психического, должна включать объяснение феномена "видения от первого лица (first-person viewpoint)", что близко посмыслу к феномену "я". Согласно изложенным взглядам (см. также /8, 14/). чувство "я" возникает в мозге как результат сопоставления информационных потоков. При ощущениях это сопоставление информации из внешнего мира и памяти. В самом деле, "я", по существу, ни что иное как воспоминание о своих впечатлениях и поступках в прошлом (по словам А.Теннисона, "я часть всего, с чем встретиться пришлось"). Представления об "я" как хранящейся в памяти и распределенной в мозге информации дает возможность преодолеть одну из главных трудностей проблемы, которая получила название проблемы ренрессии гомункулуса. Так если признать, что "я" имеет определенную мозговую локализацию, необходимо будет объяснить, как это "я" может общаться с внешним миром. Для этого придется поместить внутри гомункулуса соответствующие "сенсорные" системы, также как и структуры, отвечаюшие за мотивации и т.д., то есть практически весь мозг. Все дело в том, что, продолжая ту же логику рассуждений, внутри этого гомункулуса необходимо будет поместить другого, причем этот ряд не имеет конца /18/. В действительности "я" представляет собой актуализированную часть памяти. Но для того, чтобы перевести ее в сферу психических переживаний, необходим триггер в виде внешнего сигнала. При этом гомункулус использует имющиеся сенсорные сисмемы, получая способность "видеть". В равной степени и внешний сигнал, чтобы быть воспринятым, должен найти свой памятный отклик и соединиться с ним.
Данные представления, таким образом, близки к взглядам Д.Юма /33/. Однако Д.Юм видел в своих представлениях одну непреодолимую, как ему казалось, трудность. По Д.Юму, восприятия слишком мимолетны и не связаны друг с другом, и поэтому они не могут обеспечить постоянство чувства "я" (Д.Юм называл это проблемой тождества личности). Данные о том, что в действительности восприятие включает в себя память, как нам представляется, указывают на возможность преодоления этой трудности. Тот же принцип соединения информационных потоков может лежать в основе чувства "я" при мышлении. Однако при этом, как уже говорилось, место внешнего сигнала занимает оперативная память, которая сопоставляется с долговременной памятью.
Необходимым составным элементом "я" является и мотивация третий главный компонент информационного синтеза. Возможно, этим объясняются относительно большие связи "я" с правым полушарием. По П.В.Симонову /27/, правое полушарие ставит задачу, а левое ищет пути ее решения. Входящая в формулу эмоций необходимая ("для себя") информация в большей степени представлена в правом полушарии. Показана также особая роль теменных отделов правого полушария в поддержании схемы тела /5/.
Интересно проследить, как модифицируется в субъективной сфере соотношение между "я" и внешней средой, в зависимости от корковой локализации центра информационного синтеза. В ощущении внешний мир представлен субъекту и не зависит от него. При образном мышлении решение приходит к субъекту как бы само собой, спонтанно в виде опознания, узнавания. Наконец, при абстрактном мышлении субъект выступает в качестве действующего начала, ведущего направленный поиск решения, о чем говорилось уже при описании двух когнитивных систем. Таким образом работа эволюционно более поздних отделов коры (лобных по сравнению с теменно-височными и тем более проекционными) переживается как более активная и контролируемая со стороны "я". Помимо эволюционного фактора, эти различия представляют собой, очевидно, и развитие общего принципа строения центральной нервной системы с ее задними воспринимающими и передними исполнительными функциями.
Сознание и речь
В предыдущем изложении мы почти не касались такого важного элемента психической сферы, какой является речь. Это связано с тем, что главной задачей работы был анализ глубинных, довербальных механизмов психического, того, что по классификации Д.Эдельмена /43/, составляет первичное, "биологическое" сознание. Мы полагали, что поиск мозговых основ психики целесообразно начать с более простых ее форм (ощущения, образное, а также и элементы абстрактного мышления), которые существуют и на довербальном уровне, что показано в исследованиях на высших животных. У человека они также, несомненно, сохраняют свое значение, о чем уже говорилось выше при описании механизмов возникновения ощущения с последующей его идентификацией. Подобная последовательность процессов может сохраняться и при более слодных психических проявлениях. Известны, например, слова А.Эйнштейна, что его теоретические построения сначала возникали у него в виде неясных образов и лишь потом облекались в законченную форму. В наших исследованиях мышления связи с речевыми центрами, как правило, возникали на более поздних этапах, ближе к моменту нахождения решения.
Речь, равно как и другие формы абстракции не могут заменить и непосредственно чувственного восприятия, такого, например, как ощущения голубого цвета. Лежащий в основе ощущений механизм не идентичен механизму сознания. Из уже упомянутой работе Е.Дамянович /6/ вытекает, что этот механизм может быть нарушен при сохранении сознании, как это имело место у исследованных ею больных.
Представляя собой надстройку над этими более простыми проявлениями психики, речь создает феномен человеческого сознания. По определению П.В.Симонова, сознание прпедставляет собой знание, которое в абстрактной форме может быть передано другим людям, а само сознание возникает в процессе общения между людьми, то есть имеет коммуникативную природу /27, 28/. Эти идеи в настоящее время разделяются достаточно широким кругом исследователей /см. 17, 60/.
Вместе с тем, представление о вербализации как базовом механизме осознания само по себе еще не решает вопроса о механизмах субъективного ощущения, переживания речевых функций, то есть о возможности слышать, чувствовать как слова другого человека, так и свою собственную внутреннюю речь. Этот интригующий вопрос был предметом специальных исследований. По одной из гипотез внутренняя речь основана на проприоцептивных ощущениях, возникающих при небольшом непроизвольном сокращении артикуляционных мышц во время вербального мышления. Эта гипотеза однако была отвергнута на основании исследования, в котором испытуемым вводили курареподробные препараты, полностью блокировавшие сокращение мышц, что однако никак не отражалось на их возможности думать и использовать внутреннюю речь /59, 64/. Против данной гипотезы можно привести и следующие сообржания. Передача сигнала на мышцы, их сокращение и обратное поступление сенсорных сигналов в кору потребовало бы не менее 300 мс. Это вызвало бы значительное рассогласование по времени между самой мыслью и ее воспроизведением в субъективной сфере, что сделало бы цепь внутреннюю речь невозможной, а сами эти переживания ненужными, так как они лишь отслеживали бы мысли, а не лежали в их основе. Механизм обеспечения психических переживаний и для речевых функций должен быть поэтому внутримозговым и иметь своей основой единую интегрированную систему связей ассоциативных зон коры с речевыми. При этом базовые принципы организации довербальных форм психики могут быть, при некотором усложнении, использованы и в механизмах сознания. Эта мысль разделяется и Д.Эдельменом /43/, который полагает, что в основе "сознания высшего порядка" лежит тот же принцип повторного входа возбуждения в поля лобной, височной и теменной коры, ответственные за выполнение отдельных функций, с реализацией речевыми центрами полученной информации в соответствующих фонемах.
Функциональное значение психических переживаний
Наконец, еще об одной проблеме, о которой говорилось в начале статьи - о функциональном смысле субъективных переживаний, их роли в поведении. Представляя собой результат синтеза информации, психические феномены содержат интегрированную оценку ситуации, способствуя тем самым выбору поведенческого ответа. Элементы обобщения содержатся в самых простых психических феноменах, таких как ощущение. При мышлениии информационный синтез включает не только соединение, но и перекомбинацию иже известных сведений, что и лежит в основе нахождения решения. Это относится и к так называемому перцептивному решению, то есть опознанию стимула, что составляет последний, третий этап восприятия, и тем более к решению о поведенческом ответе.
Возникновение в процессе эволюции речи и связанного с ней человеческого сознания принципиально изменяет возможности мозга, как органа управлением поведением. Кодирование мира внутренних переживаний в виде абстрактных символов как бы делает доступным этот мир с его мыслями и чувствами для других людей, создавая единое духовное пространство, открытое для общения и накопления знаний. Благодаря этому каждое новое поколение людей живет не так, как предыдущее, что составляет резкий контраст в жизнью животных, образ жизни которых не меняется тысячелетиями. Биологическая эволюция, с ее законами выживания, таким образом, заменяется эволюцией (и революцией), совершаемой в умах людей.
Все эти положения кажутся достаточно очевидными, хотя и несколько общими. Более труден для физиолога вопрос о роли психических феноменов как факторов, влияющих на протекание мозговых процессов или даже управляющих ими. Можно, конечно, предполагать, что устойчивые системы связей, обеспечивающих процессы информационного синтеза и лежащие в основе психики, представляют собой то целое, что определяет движение нервных процессов, канализируя их по наработанным системам связей. Однако такое решение вопроса лишь кажущееся. При этом остается неясным, в чем же роль именно психического начала, так как центры интеграции в виде фокусов взаимодействия или иных структур представляют собой хотя и сложные, но находящиеся на физиологическом уровне образования, и речь идет таким образом не о влиянии психики на мозг, а физиологически более сложного на более простое.
Решение проблемы, очевидно, заключается в том, что возникающие на базе синтеза информации психические явления обладают новым качеством, по сравнению с процессами физиологического уровня. Это новое качество определяет и другую логику развития, последовательности явлений, которые подчиняются иным закономерностям высшего порядка. (Элементы такого нового качества, очевидно, присущи и неосознаваемым психическим явлениям, что, собственно, и может быть критерием для их отнесения к категории психического.) Таковы законы дедукции или, что еще более очевидно, правила грамматики, определяющие построение фразы. Так род существительного управляет формой глагола или прилагательного, определяя отдачу соответствующих команд артикуляционным мышцам. Мнение о том, что психические события привносят в протекание лежащих в их основе физиологических процессов иную логику, разделяется рядом авторов /31, 60/. По Г.Свенссону /61/, мозг без сознания был бы просто рефлекторной машиной, и цепь мыслей была бы невозможной. Говоря с другим, человек научается говорить с собой - так возникает сознание. Эта мысль Г.Свенссона, по существу, повторяет концепцию коммуникативной природы сознания П.В.Симонова, о которой говорилось выше. "Я", по П.В.Симонову, представляет собой интериозированный образ другого (это концептуальное "я", как мы полагаем, надстраивается над первичным чувстве "я", о котором говорилось выше).
Развивая выше изложенные представления об особой логике психического и ее примате над физиологическим, можно сделать и следующий шаг. Не приобретает ли в этом случае понятие "свободы воли" свой реальный, а не метафорический смысл? При такой трактовке принятие решения происходит на основе определенной логики психических событий, субъективной, но реально существующей оценки важности того или иного фактора или мотива поведения при возможности выбора из ненсльких альтернатив.
На первый взгляд, такой подход может показаться противоречащим принципу детерминизма - этого краеугольного камня точной науки. В действительности, это является отступлением только от привычного восходящего детерминизма, в соответствии с которым целое полностью зависит от движения частей. Но если признать такую детерминацию единственно возможной, все явления природы могут рассматриваться как заранее предопределенные, начиная с момента космического взрыва, создавшего вселенную. На самом деле мир устроен более сложно. Возникающие в процессе развития материи новые формы ее организации приобретают новые качества целого, которые влияют на поведение образующих его частей. Принцип детерминизма при таком подходе не отвергается, а лишь заменяется понятием двусторонней восходящей (bottom-up) и нисходящей (top-down) детерминации. (Вся "мистика" нисходящей детерминации сразу исчезает, если вспомнить пример Р.Сперри с колесом, которое катится с горы (вот уж, поистине, "top-down"), увлекая вниз молекулы вещества, из которых оно сделано.)
Эти идеи особенно актуальны при решении проблемы соотношения психики и мозга наиболее сложной в современном естествознании. Предлагаемая гипотеза мозговых основ психики как результата синтеза информации, закодированной в мозговых процессах, благодаря чему эти процессы получают новое содержание и логику развития - один из вариантов реализации этих идей.
Список литературы
1.Анохин П.К. Психическая форма отражения действительности // Избр. труды. (Философские аспекты теории функциональной системы). М.: Наука, 1978. С. 338-360.
2.Базиян Б.Х., Любимов Н.Н. Вызванные потенциалы при фиксации взора и саккадических движениях глаз человека // Физиология человека. 1990. Т. 16. В. 3. С. 137-145.
3. Бехтерева Н.П. Здоровый и больной мозг человека. Л.: Наука, 1988. 262 с.
4. Бойко Е.И. Время реакции человека. М.: Медицина, 1964. 440 с.
5.Гурфинкель В.С., Левик Ю.С. Концепция схемы тела и моторный контроль // Интеллектуальные процессы и их моделирование. Организация движений. Москва. Наука. 1991. С. 59-105.
6. Дамянович Е. Функциональная организация соматосенсорного проведения в условиях нормальной и измененной реактивности мозга у человека //Автореф. дисс. на соискание ученой степени канд. мед. наук. М. 1996.
7.Иваницкий А.М. Мозговые механизмы оценки сигналов. М. Медицина. 1976. 264 с.
8.Иваницкий А.М. "Я" и мозг. Человек // 1990. В.4. С.16.
9.Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза // Журн. высш. нервн.деят. 1996. Т. 46. N 2. C. 241.
10.Иваницкий А.М., Ильюченок И.Р. Картирование биопотенциалов мозга при решении вербальной задачи // Журн. высш.нерв. деят. 1992. Т.42. N 4. С.625-633.
11.Иваницкий А.М., Матвеева Л.В. Взаимоотношения между параметрами вызванного потенциала и структурой сенсорно-перцептивного процесса // Физиология человека. 1976. Т.2. С.386-399.
12.Иваницкий А.М., Стрелец В.Б. Вызванный потенциал и психофизические характеристики восприятия // Журн. высш. нервн. деят. 1976. Т.24. N 4. С. 793-800.
13.Иваницкий А.М., Стрелец В.Б. Функциональные связи различных отделов коры больших полушарий при восприятии внешнего раздражителя// Журн.высш.нерв.деят. 1979. Т.29. N 5. С.1071-1074.
14.Иваницкий А.М., Стрелец В.Б., Корсаков И.А. Информационные процессы мозга и психическая деятельность. М.: Наука, 1984. 200 с.
15.Казанович Я.Б., Борисюк Р.М. Синхронизация в нейронной сети фазовых осцилляторов с центральным элементом // Математическое моделирование. 1994. Т.6. N 8. С.45-60.
16. Киселева И.В., Медведев А.В., Фролов А.А. Анализ статичтичвеских характеристик биопотенциалов головного мозга // Журн. высш. нерн. деят. 1989. Т. 39. No. 4. С. 783.
17.Костандов Э.А. Узловые проблемы психофизиологии сознания. Журн. высш. нерв. деят. 1994. Т.44. N 6. С. 899-908.
18. Крик Ф. Мысли о мозге // Мозг. Под ред. П.В.Симонов. М. Мир. 1982. С. 257.
19.Ливанов М.Н. Пространственная организация процессов головного мозга. М.: Наука, 1972. 181 с.
20.Николаев А.Р. Исследование этапов мысленной ротации сложных фигур методом картирования внутрикоркового взаимодействия // Журн. высш. нерв.деят. 1994. Т. 44,N 3. С. 441-447.
21.Николаев А.Р., Иваницкий Г.А., Кошеварова О.В., Анохин А.П., Иваницкий А.М. Простанственные и временные изменения ритмов ЭЭГ при различных видах мышления. Журн. высш. нерв. деят, 1996, Т,46. N 5. С.831-848.
22.Пригожин И., Стенгерс И. Порядок из хаоса. М.: Прогресс, 1986, 584 с.
23.Русинов В.С. Доминанта. Электрофизиологические исследования. М.: Медицина,1969.231 с.
24.Сергин В.Я. Сознание как система внутреннего видения // Журн. высш. нерв. деят. 1994. Т.44. N 4-5. С. 627-639.
25.Сидорова О.А., Костюнина М.Б. Участие корковых зон мозга в процессах восприятия и воспроизведение эмоциональных состояний человека // Журн.высш.нерв.деят. 1991. Т.41. N 6. С.1094-1101.
26.Симонов П.В. Память, эмоции и доминанта // Гагрские беседы. Т. 7. Нейрофизиологические основы памяти. Под ред. Т.Ониани. Тбилиси: Мецниереба, 1979. С. 358-377.
27.Симонов П.В. Эмоциональный мозг. М.: Наука, 1981. 215 с.
28.Симонов П.В. Созидающий мозг. Нейробиологические основы творчества. М.: Наука, 1993. 111 с.
29.Симонов П.В. Функциональная асимметрия фронтального неокортекса и эмоции. Докл. Академии наук. 1994. Т. 338. N 5. С. 698-699.
30.Соколов Е.Н. Концептуальная рефлекторная дуга // Гагрские беседы. Т.7. Нейрофизиологические основы памяти. Под ред. Т.Ониани. Тбилиси: Мецниереба, 1979. С. 104-115. 31.Сперри Р.У. Перспективы менталистской революции. Возникновение нового научного мировоззрения // Мозг и разум. Москва: Наука, 1994. С.20-44.
32.Эдельмен Дж. Селекция групп и фазная повторная сигнализация: теория высших функций головного мозга // Эдельмен Дж., Маунткасл В. Разумный мозг. Под ред. Е.Н.Соколова. М.: Мир, 1981. С. 68-131.
33. Юм Д. Трактат о человеческой природе // Соч. Т.1. М. Мысль. 1965. С.--- .
34.Baars B. Cognitive theory of consciousness. New York: Cambridge University Press, 1993. 424 pp. 35. Basar E., Schurmann M. Alphs rhythm in thge brain: functional correlates. // News in Physiological Sciences.1996. V.11. P. 90.
36.Blumenthal A.L. The process of cognition. Prentice Hall. N.J: Engelwood Cliffs. 1977.
37. Bullock T.H. et al. Temporal,fluctuations in coherence of brain waves // Proc. Natl. Acad. Sci. 1995. V. 92. P. 11568.
38. Cauller l.j., Kulics A.T. The neural basis of the behaviorally relevant N1 component of the somatosensory-evoked potential in S1 cortex of awake monkeys: evidence that backward cortical projections signal touch sensation // Exp.Brain Res. 1991. V724. P.607.
39. Crick F., Koch C. Are we aware of neuronal activity in primary visual cortex? // Nature. 1995. V. 375. No 625. P. 121.
40.Czigler I. and Csibra G. Event-related potentials and identification of deviant visual stimuli // Psychophysiology. 1992. V.29. No.4. Pp. 471-485.
41.Damasio A. Descartes' error: emotion, reason and the human brain. Grosset/Putham. 1994. Pp.312.
42.Desmedt J., Tomberg C. Neurophysiology of preconscious and conscious mechanisms of the human brain // Abstracts of the Xth International Congress of Electromyography and Clinical Neurophysiology. Kyoto, Japan, October 15-19, 1995. S4.
43.Edelman G.M. The remembered present. A biologocal theory of consciousness. New York: Basics Books, 1989. 346 pp.
44. Franarzczuk P.J., Blinovska K.J. Linear model of brain electrical activity - EEG as a superposition of damped oscillatory modes // Biol.Cybern. 1985. V.53. P.19-25.
45.Froehlich F.W. Die Empfindungszeit: Ein Beitrag zur Lehre von der Zeit-Raum und Bewegungsempfindung. Jena, 1929.
46. Gevins A., Cutillo B., Desmond J., Ward M., Bressler S., Barbero N., Laxer K. Subdural grid recordings of distributed neocortical networks involved with somatosensory discrimination. EEG Clin. Neurophysiol. 1994. V. 92 P. 282-90.
47.Gray J.A. The contents of consciousness: A neuropsychological conjecture // Behav. Brain Sci. 1995, Dec.
48. Hesslow G. Will neuscience explain consciousness? // Journ. of Theoretical Biology. 1994. V.171. No. 3. P.29.
50.Kupferman I., Weiss K.R. The command neuron concept // Behav.Brain Sci. 1978. V.1. N 1. Pp. 3-10. 51.Libet B., Alberts W.W. et al. Responses of human somatosensory cortex to stimuli for conscious sensation // Science. 1967. V.158. Pp.1597-1600.
52. Madler C., Schwender D., Poppel E. Neuronal oscillators in auditory evoked potentials // Intern. Journ. Psychphysiol. 1991. V.11. P. 55.
53.Mishkin M. What is recognition memory and what neural circuits are involved? // Abstracts. XXXII Congress of the International Union of Physiological Sciences. Aug.1st-6th, 1993. Glasgow. Sunday. Pp.42-43.
54.Mishkin M., Horn G., Schacter D. Cerebral memory systems // Third IBRO Congress of Neuroscience. August 4-9, 1991. Montreal, Canada. Abstacts. S19. P.4.
55.Naatanen R. Processing negativity, evoked potential reflection of selective attention // Psychological Bulletin. 1982. V.92. Pp.605-640.
56.Pieron H. La sensation. Paris: Presse univ. France, 1960.
57. Posner M.I., Rothbart M.K. Constructing neuronal theories of mind // High level neuronal theories of the brain. Ed. C.Koch and J.Davis. Cambirdge, Mass. MIT Press. 1994. P.183.
58.Salmelin R., Hari R., Lounasman O.V. and Sams M. Dimanics of brain activation during picture naming // Nature. 1994. V.368. N 6470. Pp. 463-465.
59. Smith S.M., Brown H.O., Toman J.E.P., Goodman I.S. lack of cerebral effects od D-tubocurarine // Anesthesiology. 1947. V.8. P.1.
60. Stoerig P., Brandt S. The visual system and levels of perception: properties of neuromental organization // Theoretical Medicine. 1993. V.14. No. 2. P. 117.
61. Swensson G. Reflections on the problem of identifying mind and brain // Journ. of Theoretical Biology. 1994. V.171. No.1. P.93.
62.Swets Y., Tanner W., Birdsall T. Decision process in perception // Psychol.Rew. 1961. V. 68. P.15. Пер. в кн.: Инженерная психология. М.,: Прогресс, 1964, стр. 269-335.
63. Thorpe S., Fize D., Marlot C. Speed of processing in the human visual system // Nature. 1996. V. 381. P. 520.
64. Weisberg R.W. Memory, thought and behavior.N.Y., Oxford. Oxford Univ. Press.1980. 458 pp.
Исследования, составившие предмет настоящей публикации, были возможны частично благодаря гранту N M9Q000, полученному от Междунарожного научного фонда (International Science Foundation), и гранту N M9Q300 от Международного научного фонда и Российского правительства, гранту N от РФФИ и гранту N от РГНФ.