
Параллельная эволюция эусоциальности у пчел: принцип общий, эволюционные маршруты — разные
Автор: Александр Марков
|
Сравнительный анализ геномов 10 видов одиночных и общественных пчел показал, что поэтапное развитие эусоциальности, происходившее параллельно в нескольких эволюционных линиях, было основано на изменении разных наборов генов. Главной общей чертой всех рассмотренных случаев является усложнение регуляторных генных сетей, о чем можно судить по увеличению числа участков ДНК, служащих для прикрепления регуляторных белков (транскрипционных факторов), а также числа генов, подвергающихся метилированию. В отличие от регуляторных участков, белок-кодирующие участки многих ключевых генов, управляющих развитием нервной и эндокринной систем, с переходом к социальности стали эволюционировать медленнее. Скорее всего, это объясняется возросшей многофункциональностью этих генов. Общая направленность генетических изменений при переходе к эусоциальности говорит о расширении нормы реакции, или, иными словами, о росте «интерактивности» генома, то есть его способности генерировать разные фенотипы в зависимости от получаемых сигналов, в том числе социальных.
Эусоциальность — высшая форма общественной организации, при которой часть особей отказывается от собственного размножения, чтобы помогать другим, чаще всего — собственным родителям. Эусоциальность появлялась много раз независимо в разных группах животных. Больше всего таких случаев отмечено у перепончатокрылых насекомых, которые исходно вели одиночный образ жизни, но затем разные их представители (осы, пчелы, шмели, муравьи) независимо друг от друга «изобретали» эусоциальность.
Происхождение эусоциальности у перепончатокрылых — излюбленная тема эволюционных биологов (см. ссылки в конце новости). Интерес к ней подстегивается тем, что переход к эусоциальности — это крупное эволюционное событие, сопряженное с глубокими изменениями поведения, морфологии и физиологии организмов. Кроме того, с разделением на репродуктивную и рабочую касты естественный отбор начинает действовать на уровне групп (семей), а не отдельных особей. Поэтому характер эволюции должен существенно различаться у эусоциальных и одиночных видов. Теоретические модели, описывающие эти предполагаемые различия, разработаны достаточно подробно. Более сложной задачей является эмпирическая проверка модельных предсказаний. Большие надежды в этом плане возлагаются на сравнительную геномику, и надежды эти начинают оправдываться.
Большой международный коллектив, в состав которого вошли ученые из США, Китая, Дании, Испании, Швейцарии, Германии, Великобритании, Бразилии и Канады, опубликовал результаты сравнительного анализа геномов 10 видов одиночных и общественных пчел. Пять геномов были отсеквенированы ранее, пять других прочтены авторами специально для данного исследования. Виды были подобраны таким образом, чтобы эволюционное дерево, отражающее их родственные связи, включало несколько независимых событий приобретения и усовершенствования эусоциальности (рис. 1), а именно два независимых перехода от одиночной жизни к примитивным формам эусоциальности (просто организованные семьи с небольшим числом рабочих) и два перехода от простой эусоциальности к сложной (с огромными семьями и четким разделением труда между рабочими).
Теория предсказывает, что переход к эусоциальности должен сопровождаться усложнением генной регуляции. Дело в том, что у социальных насекомых один и тот же геном должен обеспечивать формирование разных фенотипов (каст) в зависимости от условий (сигналов, получаемых в процессе развития), а также сложные системы социальных взаимодействий, коммуникацию и обучаемость. Чтобы проверить это предсказание, авторы предприняли целенаправленный поиск регуляторных участков генома (сайтов связывания транскрипционных факторов), к которым прикрепляются белки, регулирующие экспрессию генов (транскрипционные факторы, ТФ). Для этого в промоторных участках генов искали участки ДНК с характерными последовательностями нуклеотидов, к которым прикрепляется тот или иной ТФ. Учитывали 188 ТФ, которые имеются у всех рассматриваемых видов пчел и для которых точно известны нуклеотидные последовательности сайтов связывания. Выяснилось, что по мере развития социальности число таких регуляторных участков действительно росло. Иными словами, у социальных видов на каждый регуляторный белок приходится в среднем больше регулируемых генов, чем у одиночных, а у видов со сложно организованными семьями — больше, чем у видов с простыми формами социальности.
Еще один довод в пользу усложнения генной регуляции у социальных видов авторы получили, оценив характер метилирования ДНК, которое, наряду с ТФ, является важным способом регуляции работы генов. Уровень метилирования генов оценивали по косвенным признакам, связанным с частотой встречаемости нуклеотидов Ц и Г и динуклеотидов ЦГ. Дело в том, что метилированию чаще всего подвергаются цитозины, за которыми в цепочке ДНК следует гуанин (см. ). Метилированные цитозины часто мутируют, спонтанно превращаясь в тимин. В результате гены, подвергающиеся усиленному метилированию, обычно имеют пониженное количество CpG-сайтов по сравнению с ожидаемым на основе количества Ц и Г в данном участке ДНК. Надежность такой оценки уровня метилирования проверялась на медоносной пчеле Apis mellifera, у которой метилирование многих генов было измерено напрямую. В итоге выяснилось, что по мере развития эусоциальности уровень метилирования генома действительно увеличивался.
Проанализировав скорость эволюции белок-кодирующих участков генов, авторы показали, что развитие эусоциальности сопровождалось ускоренной эволюцией многих белков, участвующих в регуляции транскрипции, сплайсинга и трансляции. Это опять-таки свидетельствует об изменениях регуляторных генных сетей. Впрочем, лишь 2/3 таких быстро эволюционирующих белков у социальных видов меняются под действием направленного (движущего) отбора, то есть путем закрепления мутаций, повышающих приспособленность. У оставшейся трети, судя по соотношению синонимичных и значимых замен (см. ), повышенный темп эволюционных изменений объясняется ослаблением очищающего отбора, отбраковывающего вредные мутации. Этот результат тоже является ожидаемым, поскольку переход к эусоциальности автоматически ведет к резкому снижению эффективной численности популяции (), то есть числа особей, участвующих в размножении. Это, в свою очередь, должно снижать эффективность очищающего отбора (Romiguier et al., 2014. ).
В то время как одни белки с переходом к эусоциальности стали меняться быстрее, другие, наоборот, замедлили свою эволюцию. Среди них оказалось много ключевых регуляторов развития нервной и эндокринной систем. Авторы предполагают, что причиной замедления их эволюции является возросшая многофункциональность. Чем больше функций у белка, тем меньше шансов, что случайное изменение аминокислотной последовательности не принесет вреда и не будет отбраковано отбором. На первый взгляд может показаться, что замедление эволюции подобных генов противоречит представлению об усложнении генных сетей. На самом деле замедление эволюции некоторых белок-кодирующих участков — прямое следствие ускоренной эволюции регуляторных участков. Например, белок-кодирующая часть гена, кодирующего транскрипционный фактор, будет меняться всё с большим и большим трудом по мере того, как в геноме будет расти число сайтов связывания данного ТФ. Ведь рост числа сайтов означает, что всё больше генов попадает под его контроль, следовательно растет его многофункциональность. Можно также ожидать, что чем больше в промоторной области гена сайтов связывания различных ТФ, тем больше у гена функций, а значит, эволюция его белок-кодирующей части должна идти медленнее. И действительно, авторы обнаружили, что гены, эволюция которых замедлилась с переходом к эусоциальности, отличаются ускоренным ростом числа сайтов связывания ТФ в своей промоторной области.
До сих пор речь шла об эволюционных тенденциях, общих для обоих рассматриваемых случаев перехода от одиночной жизни к простой эусоциальности и для обоих переходов от простой эусоциальности к сложной. Еще одной их общей чертой, по-видимому, было уменьшение числа и разнообразия всевозможных «эгоистических элементов» — транспозонов и ретротранспозонов (причины этого неясны, ведь снижение эффективной численности популяции должно способствовать, наоборот, накоплению генетического мусора).
На этом сходство генетических изменений, сопровождавших четыре эволюционных преобразования, заканчивается. Конкретные наборы генов, замедливших или ускоривших свою эволюцию, транскрипционных факторов, вовлеченных в процесс, и сайтов их связывания во всех случаях были разными, почти не пересекающимися, хотя и связанными с одним и тем же кругом регуляторных функций (рис. 2).
|
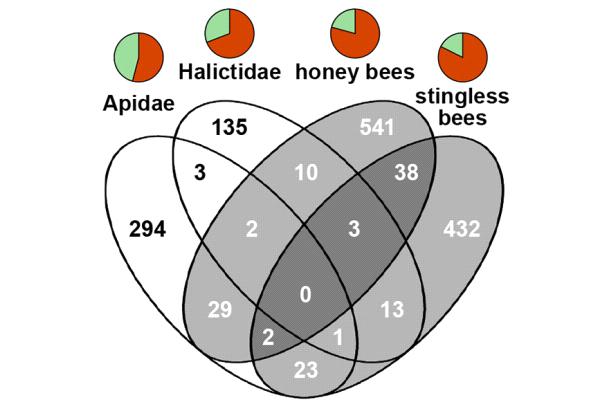
Это значит, что к одному и тому же результату — эусоциальности — эволюция может прийти множеством разных генетических «маршрутов». Ярко выраженный эволюционный параллелизм на уровне фенотипа в данном случае был основан на разных изменениях на уровне ДНК. Этим эволюция эусоциальности разительно отличается от некоторых других «расшифрованных» на генетическом уровне случаев конвергенции (см.: , «Элементы», 15.10.2013). Впрочем, есть и похожие примеры (см.: , «Элементы», 01.02.2012).
Подытоживая полученные результаты, авторы отмечают, что если бы эволюцию перепончатокрылых можно было «проиграть заново», эусоциальность опять каждый раз возникала бы на основе разных генетических механизмов. Но во всех случаях, по-видимому, наблюдалось бы усложнение систем генной регуляции.
Источник: Karen M. Kapheim, Hailin Pan, Cai Li, Steven L. Salzberg, Daniela Puiu, Tanja Magoc, Hugh M. Robertson, Matthew E. Hudson, Aarti Venkat, Brielle J. Fischman, Alvaro Hernandez, Mark Yandell, Daniel Ence, Carson Holt, George D. Yocum, William P. Kemp, Jordi Bosch, Robert M. Waterhouse, Evgeny M. Zdobnov, Eckart Stolle, F. Bernhard Kraus, Sophie Helbing, Robin F. A. Moritz, Karl M. Glastad, Brendan G. Hunt, Michael A. D. Goodisman, Frank Hauser, Cornelis J. P. Grimmelikhuijzen, Daniel Guariz Pinheiro, Francis Morais Franco Nunes, Michelle Prioli Miranda Soares, Érica Donato Tanaka, Zilá Luz Paulino Simões, Klaus Hartfelder, Jay D. Evans, Seth M. Barribeau, Reed M. Johnson, Jonathan H. Massey, Bruce R. Southey, Martin Hasselmann, Daniel Hamacher, Matthias Biewer, Clement F. Kent, Amro Zayed, Charles Blatti III, Saurabh Sinha, J. Spencer Johnston, Shawn J. Hanrahan, Sarah D. Kocher, Jun Wang, Gene E. Robinson, and Guojie Zhang. Genomic signatures of evolutionary transitions from solitary to group living // Science. Published online 14 May 2015.
Об эволюции социальности у перепончатокрылых см. также:
1) , «Элементы», 05.02.2014.
2) , «Элементы», 21.01.2014.
3) , «Элементы», 21.09.2010.
4) , «Элементы», 08.11.2006.
5) , «Элементы», 16.05.2006.
 http://www.scorcher.ru/xml/news.rss
http://www.scorcher.ru/xml/news.rss
|
Рис. 1. Эволюционное дерево, показывающее родственные связи 10 видов пчел, рассматриваемых в обсуждаемой статье. По горизонтальной оси — время в млн лет назад. Справа, рядом с изображениями насекомых, указано примерное число особей в колонии, характерное для данного вида. Белые кружки — два случая перехода от исходного одиночного образа жизни к простым формам эусоциальности, серые кружки — два независимых перехода к высокоразвитой эусоциальности. Названия видов написаны разными цветами в соответствии с уровнем социальности: синие — одиночный образ жизни, зеленые — факультативная простая эусоциальность, желтые — облигатная простая эусоциальность, красные — сложная эусоциальность. Рисунок из обсуждаемой статьи в Science