
Нужны ли эмбрионам гены?
|
Вопреки расхожему мнению о том, что индивидуальное развитие есть реализация заложенной в геноме «программы», ранние стадии эмбрионального развития животных идут при отключенном геноме. Вплоть до стадии или даже зародыш синтезирует все необходимые ему белки на основе матричных РНК, полученных от матери. Когда гены зародыша наконец начинают работать, материнские РНК уничтожаются. Механизмы и биологический смысл этих явлений остаются во многом загадочными.
Принято считать, что индивидуальное развитие () — это постепенная «реализация» той генетической информации, которая заключена в геноме оплодотворенной яйцеклетки () и которая в итоге окончательно «воплощается» в строении взрослого организма. Все понимают, что путь от к труден и извилист, но лишь немногие эволюционисты всерьез пытаются объяснить ключевые закономерности эволюции особенностями тех сложнейших процессов самоорганизации, которые составляют суть онтогенеза (уж слишком трудна задача). Поэтому обычно проблему пытаются упростить, сведя всё к вопросу о том, каким образом те или иные изменения генотипа (например, случайные мутации) могут отразиться на процессе развития зародыша.
Согласно традиционным взглядам, геном рассматривается как активное начало (в нём всё изначально «закодировано», он «руководит» развитием). Развивающийся зародыш, напротив, считается чем-то вроде пассивного «результата» деятельности генома. Дело несколько осложняется тем обстоятельством, что сам геном в процессе онтогенеза явно находится под контролем: в разных клетках эмбриона одни гены включаются, другие выключаются в строгой последовательности, определяемой, в частности, химическими сигналами, которыми обмениваются между собой клетки и ткани зародыша. Кто кем управляет, становится не совсем ясно. Некоторые теоретики по этому поводу даже заявляют, что геном — это не «программа развития зародыша», а скорее некий набор инструментов, которыми зародыш пользуется (или не пользуется) по своему усмотрению.
Для управления собственными генами зародыш использует множество разных механизмов: это и регуляция транскрипции (считывания генов) при помощи малых РНК и специальных регуляторных белков — транскрипционных факторов (знаменитые HOX-гены тоже кодируют транскрипционные факторы), и особые «эпигенетические» механизмы (см. С. А. Назаренко. ), в том числе метилирование генов при помощи специальных ферментов ДНК-метилтрансфераз и ацетилирование гистонов — белков, на которые «намотаны» молекулы ДНК. В отличие от обычных регуляторов-переключателей, «эпигенетические» часто передаются по наследству от родительской клетки к дочерним, то есть их положение (вкл./выкл.) может сохраняться неизменным в ряду поколений делящихся клеток.
Еще одно обстоятельство, которое делает сомнительным тезис о полной и однозначной обусловленности онтогенеза геномом зиготы, состоит в том, что у подавляющего большинства многоклеточных животных на ранних стадиях развития геном вообще не функционирует. Он просто-напросто отключен, все гены молчат и матричные РНК («считываемые» с генов матрицы для синтеза белка) не производятся.
|
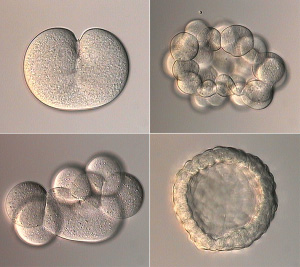
Зародыш между тем претерпевает сложные превращения. Яйцеклетка начинает дробиться, число клеток эмбриона растет в геометрической прогрессии: 2, 4, 8, 16, 32... Наконец формируется однослойный шар из клеток (бластула). Клетки, находящиеся на одном из полюсов бластулы, мигрируют внутрь, давая начало второму зародышевому листку (энтодерме), из которой позже разовьется кишечник. На этой стадии двуслойный зародыш называется гаструлой. Только на этом этапе у многих животных начинают наконец включаться гены, унаследованные от папы с мамой. У других это происходит чуть раньше — на стадии бластулы. И только млекопитающие — группа уникальная во многих отношениях — включают свои гены еще раньше (например, мышь делает это на стадии двух клеток).
Как удается эмбриону развиваться без всякого «генетического контроля» вплоть до стадии гаструлы? Почему гены зародыша так долго остаются выключенными? Какие механизмы обеспечивают отключение генов в зиготе, а затем их своевременное включение? Обзорная статья, опубликованная в журнале Science, рассказывает о последних достижениях ученых, пытающихся разгадать эти загадки.
Ответ на первый вопрос более или менее ясен. Яйцеклетка содержит большое количество матричных РНК, унаследованных от материнского организма. Эти мРНК считываются с материнских генов заблаговременно, в процессе созревания яйцеклетки. Именно они обеспечивают синтез белков, необходимых для ранних стадий онтогенеза. В определенный момент материнские мРНК начинают уничтожаться. Это происходит как раз тогда, когда зародыш начинает сам производить мРНК, то есть включает свои гены. Этот довольно быстрый процесс замены зародышем материнских мРНК на свои собственные называется maternal-zygotic transition (MZT).
Менее ясен вопрос о том, что движет процессом MZT. Предполагается три возможных механизма:
- По мере роста числа клеток в зародыше начинает не хватать тех веществ (что бы они из себя ни представляли), которые не позволяют генам зародыша включиться. Ранние стадии эмбриогенеза животных не случайно называют «дроблением»: зигота именно дробится, клетки эмбриона после каждого деления становятся всё мельче, поскольку между клеточными делениями отсутствует стадия роста клеток. Общее количество цитоплазмы не растет, тогда как количество клеточных ядер, а следовательно и ДНК, увеличивается в геометрической прогрессии. Если предположить, что яйцеклетка заранее запаслась какими-то ингибиторами транскрипции, то количество этих гипотетических ингибиторов, приходящихся на каждую клетку, должно быстро убывать, и в конце концов их остается так мало, что они уже не могут сдерживать транскрипцию.
- Не исключено, что в зиготе изначально имеет место целенаправленное блокирование некоторых ключевых генов, работа которых инициирует транскрипцию. Так, показано, что искусственное введение в эмбрион мощных активаторов транскрипции (например, так называемого ТАТА-связывающего белка, TBP) может вызвать преждевременное частичное включение эмбрионального генома.
- Наконец, сама по себе быстрая череда клеточных делений может мешать транскрипции. Ведь каждому делению должно предшествовать удвоение ДНК (репликация). В ходе дробления репликация должна происходить, по сути дела, непрерывно. Между тем известно, что репликация может мешать транскрипции, а во время клеточного деления (митоза) может происходить обрыв и уничтожение тех мРНК, синтез которых еще не закончился. Может быть, клетки эмбриона просто физически не успевают транскрибировать свои гены? Искусственное замедление процесса дробления действительно может вызвать преждевременное включение эмбрионального генома. Кстати, у млекопитающих дробление протекает сравнительно медленно, не этим ли объясняется раннее включение генов зародыша?
Впрочем, ни одна из этих теорий не объясняет всей совокупности имеющихся фактов. Например, они не в состоянии объяснить, почему эмбриональные гены включаются постепенно, в строго определенном порядке, а единичные гены могут быть включены уже на самых ранних стадиях дробления.
Вторым существенным аспектом MZT является уничтожение материнских мРНК. Тут, как выяснилось, всё предусмотрено заранее: материнские мРНК помечены особой последовательностью нуклеотидов, расположенной на нетранслируемом (то есть не кодирующем белок) «хвостике» этих молекул. Среди первых генов, которые эмбрион включает в процессе MZT, находятся гены особых белков и маленьких РНК, которые распознают эту последовательность, прикрепляются к ней и тем самым инициируют уничтожение материнских мРНК.
До полного понимания всех этих процессов науке еще очень далеко, но дело движется. Когда основные механизмы генной регуляции в ходе раннего онтогенеза будут расшифрованы, биологи смогут вплотную заняться следующим принципиальным вопросом, а именно: зачем всё это надо? почему многоклеточные животные не доверяют своему геному контроль над ранними стадиями развития, а потом в какой-то момент вдруг «переключаются» с материнских транскриптов на свои собственные?
Между прочим, данная проблема имеет не только теоретическое, но и практическое значение. Трудности, с которыми сталкиваются работы по клонированию животных, во многом определяются тем, что мы еще слишком мало знаем о работе генов в ходе раннего развития. Клонирование животных осуществляется путем пересадки ядра из соматической (неполовой) клетки одного животного в яйцеклетку другого. Однако для нормального развития необходимо, чтобы гены зародыша поначалу помалкивали, а они в донорском ядре вполне активны. Цитоплазма яйцеклетки должна каким-то образом «перепрограммировать» ядро, выключить геном, а потом в нужный момент снова включить его. Пока мы не знаем, как ей в этом помочь, больших успехов в клонировании ожидать трудно.
Источник: Alexander F. Schier. // Science. 2007. V. 316. P. 406–407.
Комментарии:
Ment
А гены эмбрионам все-таки нужны. Ведь материнские мРНК, а точнее продукты трансляции оных, определяют лишь самое начало развития зародыша. Согласно концепции, существующей на сегодняшний день (насколько мне она известна), они являются своеобразными "сигналами"
для некоторых генов зародыша, которые обуславливают определенную степень экспрессии, либо репрессии зародышевых генов. Например, у дрозофилы основное "стартовое" решение в становлении передне-задней оси (грубо: где голова будет) играет градиент концентрации материнской мРНК гена bicoid (bcd), продуктом которого является регулятор транскрипции. Данная мРНК в зиготу попадает лишь с одного конца зародыша от окружающих материнских фоликулярных клеток, что и создает градиент. Этот регулятор активирует, либо реприссирует экспрессию других регуляторов, распределение которых играет важную роль в сегментации тела. Благодаря градиенту bcd-мРНК, экспрессия этих регуляторов неодинакова в разных частях эмбриона и соотношение их продуктов определяет степень экспрессии других генов, многие из которых тоже являются регуляторами и т.д.
Основной проблемой эмбрионального развития является выявление всех участвующих в развитии генов и установление их взаимодействия. Причем настоящие проблемы появляются как раз-таки на более поздних этапах, т.к. идет постоянное увеличение числа вовлеченных регуляторов.
Я к сожалению не имею доступа к Science, но судя по "абстракту" авторы разбирали, каков механизм MZD, а не его биологический смысл, который в общем-то понятен: чем сложнее организм, тем больше усилий необходимо преложить, чтобы все было на своем месте.
 http://www.scorcher.ru/xml/news.rss
http://www.scorcher.ru/xml/news.rss
|