Физиология центральной нервной системы
Центральную нервную систему составляют спинной и головной мозг. Основными функциями центральной нервной системы являются: 1) регуляция деятельности всех тканей и органов и объединение их в единое целое; 2) обеспечение приспособления организма к условиям внешней среды (организация адекватного поведения соответственно потребностям организма).
Управление различными функциями осуществляется и гуморальным путем (через кровь, лимфу, тканевую жидкость), однако нервная система играет главенствующую роль. У высших животных и человека ведущим отделом центральной нервной системы является кора больших полушарий, которая управляет также наиболее сложными функциями в жизнедеятельности человека — психическими процессами (сознание, мышление, память и др.).
§ 1. Физиология нервной клетки
 Основным структурным элементом нервной системы является нервная
клетка, или нейрон. Через нейроны осуществляется передача информации от одного
участка нервной системы к другому, обмен информацией между нервной системой и
различными участками тела. В нейронах происходят сложнейшие процессы обработки
информации. С их помощью формируются ответные реакции организма (рефлексы) на
внешние и внутренние раздражения.
Основным структурным элементом нервной системы является нервная
клетка, или нейрон. Через нейроны осуществляется передача информации от одного
участка нервной системы к другому, обмен информацией между нервной системой и
различными участками тела. В нейронах происходят сложнейшие процессы обработки
информации. С их помощью формируются ответные реакции организма (рефлексы) на
внешние и внутренние раздражения.
Нейроны разделяются на три основных типа: афферентные, эфферентные и промежуточные нейроны. Афферентные нейроны (чувствительные, или центростремительные) передают информацию от рецепторов в центральную нервную систему. Тела этих нейронов расположены вне центральной нервной системы — в спинномозговых ганглиях и в ганглиях черепно-мозговых нервов.
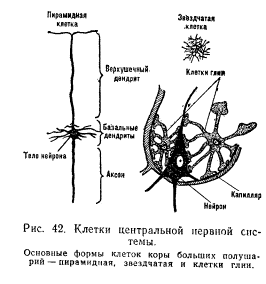
Афферентный нейрон имеет ложноуниполярную форму, т. е. оба его отростка выходят из одного полюса клетки. Далее нейрон разделяется на длинный дендрит, образующий на периферии воспринимающее образование — рецептор, и аксон, входящий через задние рога в спинной мозг. К афферентным нейронам относят также нервные клетки, аксоны которых составляют восходящие пути спинного и головного мозга. Эфферентные нейроны (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим (например, пирамидные нейроны коры больших полушарий — рис. 42) или из центральной нервной системы к рабочим органам (например, в передних рогах спинного мозга расположены тела двигательных нейронов, или мотонейронов, от которых идут волокна к скелетным мышцам; в боковых рогах спинного мозга находятся клетки вегетативной нервной системы, от которых идут пути к внутренним органам). Для эфферентных нейронов характерны разветвленная сеть дендритов и один длинный отросток — аксон. Промежуточные нейроны (интернейроны, или вставочные) — это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного мозга в другие — выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов (например, звездчатые клетки коры — см. рис. 42).
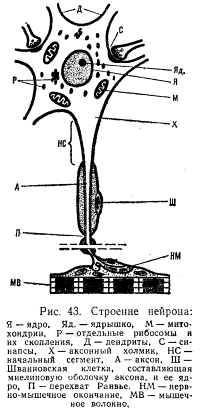
Функциональное значение различных структурных элементов нервной клетки. Различные структурные элементы нейрона имеют свои функциональные особенности и разное физиологическое значение. Нервная клетка состоит из тела, или сомы (рис. 43), и различных отростков. Многочисленные древовидно разветвленные отростки дендриты (от греч. dendron — дерево) служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток аксон (от греч. axis — ось), который передает нервные импульсы дальше — другой нервной клетке или рабочему органу (мышце, железе). Форма нервной клетки, длина и расположение отростков чрезвычайно разнообразны и зависят от функционального назначения нейрона.
 Среди нейронов встречаются самые крупные клеточные элементы
организма. Размеры их поперечника колеблются от 6—7 мк (мелкие зернистые клетки
мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их
расположения в некоторых отделах центральной нервной системы очень велика.
Например, в коре больших полушарий человека на 1 мм3 приходится
почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно
половину объема коры.
Среди нейронов встречаются самые крупные клеточные элементы
организма. Размеры их поперечника колеблются от 6—7 мк (мелкие зернистые клетки
мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их
расположения в некоторых отделах центральной нервной системы очень велика.
Например, в коре больших полушарий человека на 1 мм3 приходится
почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно
половину объема коры.
В крупных нейронах почти 1/3 — 1/4 величины их тела составляет ядро. Оно содержит довольно постоянное количество дезоксирибонуклеиновой кислоты (ДНК). Входящие в его состав ядрышки участвуют в снабжении клетки рибонуклеиновыми кислотами (РНК) и протеинами. В моторных клетках при двигательной деятельности ядрышки заметно увеличиваются в размерах. Нервная клетка покрыта плазматической мембраной—полупроницаемой клеточной оболочкой, которая обеспечивает регуляцию концентрации ионов внутри клетки и ее обмен с окружающей средой. При возбуждении проницаемость клеточной мембраны изменяется, что играет важнейшую роль в возникновении потенциала действия и передаче нервных импульсов. Аксоны многих нейронов покрыты миелиновой оболочкой, образованной Шванновскими клетками, многократно «обернутыми» вокруг ствола аксона. Однако начальная часть аксона и расширение в месте его выхода из тела клетки — аксонный холмик лишены такой оболочки. Мембрана этой немиелинизированной части нейрона — так называемого начального сегмента — обладает высокой возбудимостью.
Внутренняя часть клетки заполнена цитоплазмой, в которой расположены ядро и различные органоиды. Цитоплазма очень богата ферментными системами (в частности, обеспечивающими гликолиз) и белком. Ее пронизывает сеть трубочек и пузырьков — эндоплазматический ретикулюм. В цитоплазме имеются также отдельные зернышки — рибосомы и скопления этих зернышек — тельца Ниссля, представляющие собой белковые образования, содержащие до 50% РНК. Это белковые депо нейронов, где также происходит синтез белков и РНК. При чрезмерно длительном возбуждении нервной клетки, вирусных поражениях центральной нервной системы и других неблагоприятных воздействиях величина этих рибосомных зернышек резко уменьшается.
В специальных аппаратах нервных клеток — митохондриях совершаются окислительные процессы с образованием богатых энергией соединений (макроэргических связей АТФ). Это энергетические станции нейрона. В них происходит трансформация энергии химических связей в такую форму, которая может быть использована нервной клеткой. Митохондрии концентрируются в наиболее активных частях клетки. Их дыхательная функция усиливается при мышечной тренировке. Интенсивность окислительных процессов нарастает в нейронах более высоких отделов центральной нервной системы, особенно в коре больших полушарий. Резкие изменения митохондрий вплоть до разрушения, а следовательно, и угнетение деятельности нейронов отмечаются при различных неблагоприятных воздействиях (длительном торможении в центральной нервной системе, при интенсивном рентгеновском облучении, кислородном голодании и гипотермии).
Обмен веществ в нейроне. Основной особенностью обмена веществ в нейроне является высокая скорость обмена и преобладание аэробных процессов. Потребность мозга в кислороде очень велика (в состоянии покоя поглощается около 46 мл/мин кислорода). Хотя вес мозга по отношению к весу тела составляет всего 2%, потребление кислорода мозгом достигает в состоянии покоя у взрослых людей 25% от общего его потребления организмом, а у маленьких детей — 50%. Даже кратковременное нарушение доставки кислорода кровью может вызвать необратимые изменения в деятельности нервных клеток: в спинном мозгу — через 20 — 30 мин., в стволе головного мозга — через 15 — 20 мин., а в коре больших полушарий — уже через 5 — 6 мин.
Энерготраты мозга составляют 1/6 — 1/8 суточных затрат организма человека. Основным источником энергии для мозговой ткани является глюкоза. Мозг человека требует для обмена около 115 г глюкозы в сутки. Содержание ее в клетках мозга очень мало, и она постоянно черпается из крови.
Деятельное состояние нейронов сопровождается трофическими процессами—усилением в них синтеза белков. При различных воздействиях, вызывающих возбуждение нервных клеток, в том числе при мышечной тренировке, в их ткани значительно возрастает количество белка и РНК, при тормозных же состояниях и утомлении нейронов содержание этих веществ уменьшается. В процессе восстановления оно возвращается к исходному уровню или превышает его. Часть синтезированного в нейроне белка компенсирует его расходы в теле клетки во время деятельности, а другая часть перемещается вдоль по аксону (со скоростью около 1—3 мм в сутки) и, вероятно, участвует в биохимических процессах в синапсах.
Кровоснабжение нервных клеток. Высокая потребность нейронов в кислороде и глюкозе обеспечивается интенсивным кровотоком.
Кровь протекает через мозг в 5—7 раз скорее, чем через покоящиеся мышцы. Мозговая ткань обильно снабжена кровеносными сосудами. Наиболее густая сеть их находится в коре больших полушарий (занимает около 10% объема коры). В отдельных слоях ее средняя длина капиллярной сети достигает у человека 1 м на 1 мм3 ткани. Каждый крупный нейрон имеет несколько собственных капилляров у основания тела клетки, а группы мелких клеток окутаны общей капиллярной сетью. При активном состоянии нервной клетки она нуждается в усиленном поступлении через кровь кислорода и питательных веществ. Вместе с тем жесткий каркас черепа и малая сжимаемость нервной ткани препятствуют резкому увеличению кровоснабжения мозга при работе. Однако это компенсируется выраженными в мозгу процессами перераспределения крови, в результате которых активный участок нервной ткани получает значительно больше крови, чем находящийся в покое. Возможность перераспределения крови в мозгу обеспечена наличием в основаниях артериальных ветвей крупных пучков гладких мышечных волокон — сфинктерных валиков. Эти валики могут уменьшать или увеличивать диаметр сосудов и тем самым производить раздельную регуляцию кровоснабжения разных участков мозга.
Мышечная работа вызывает снижение тонуса стенок мозговых артерий. При развитии физического или умственного утомления тонус артериальных сосудов повышается, что ведет к уменьшению кровотока через нервную ткань.
В головном мозгу имеется богато развитая система анастомозов между различными артериями, между венозными сосудами и между артериями и венами. Эта система уменьшает пульсацию внутричерепного кровотока, обусловленную ритмическими сокращениями сердца и дыхательными движениями грудной клетки. Уменьшение пульсовых колебаний способствует улучшению тканевого кровотока. Благодаря наличию артериовенозных анастомозов пульсовые колебания кровотока передаются с артерий мозга на вены, минуя капилляры. Анастомоз между системами сонных и позвоночных артерии (Виллизиев круг) гарантирует постоянство кровотока в различных отделах головного мозга при любом положении головы по отношению к туловищу и направлению силы тяжести, связанном с изменением положения тела в пространстве.
Клетки глии. В процессах питания нервных клеток и их обмене веществ участвуют также окружающие нейрон клетки глии (глиальные клетки, или нейроглия). Эти клетки заполняют в мозгу все пространство между нейронами. В коре больших полушарий их примерно в 5 раз больше, чем нервных клеток. Капилляры в центральной нервной системе плотно окружены клетками глии, которые покрывают сосуд полностью или оставляют небольшую часть (15%) свободной. Выросты некоторых глиальных клеток расположены частично на кровеносных сосудах и частично на нейроне (см. рис. 42). Полагают, что расположение этих клеток между сосудом и нейроном указывает на их роль в снабжении нервных клеток питательными веществами из крови. Глиальные клетки активно участвуют в функционировании нейрона. Показано, что при длительном возбуждении в нейроне высокое содержание белка и нуклеиновых кислот поддерживается за счет клеток глии, в которых их количество соответственно уменьшается. В процессе восстановления после работы запасы белка и нуклеиновых кислот сначала нарастают в клетках глии, а затем и в цитоплазме нейрона. Глиальные клетки обладают способностью перемещаться в пространстве по направлению к наиболее активным нейронам. Это наблюдается при различных афферентных раздражениях и при мышечной нагрузке. Например, уже через 20 мин. плавания у крыс было обнаружено увеличение числа глиальных клеток вокруг мотонейронов переднего рога спинного мозга.
Возможно, клетки глии участвуют в условно-рефлекторной деятельности мозга и в процессах памяти. Участвуют, но не функционально, а как вспомогательное окружение. Механизмы рефлексов функционально полностью описываются свойствами нейронов и синапсов.
Основные функции нервной клетки. Основными функциями нервной клетки являются восприятие внешних раздражении (рецепторная функция), их переработка (интегративная функция) и передача нервных влияний на другие нейроны или различные рабочие органы (эффекторная функция).
Особенности осуществления этих функций позволяют разделить все нейроны центральной нервной системы на 2 большие группы:
1) клетки, передающие информацию на большие расстояния (из одного отдела центральной нервной системы в другой, от периферии к центру, от центров к исполнительному органу). Это крупные, афферентные и эфферентные нейроны, имеющие на своем теле и отростках большое количество синапсов, как возбуждающих, так и тормозящих, и способные к сложным процессам переработки поступающих через них влиянии;
2) клетки, обеспечивающие межнейроальные связи в пределах ограниченных нервных структур (промежуточные нейроны спинного мозга, коры больших полушарий и др.). Это мелкие клетки, воспринимающие нервные влияния только через возбуждающие синапсы. Эти клетки не способны к сложным процессам интеграции локальных синаптических влияний потенциалов, они служат передатчиками возбуждающих или тормозящих влияний на другие нервные клетки.
Воспринимающая функция нейрона. Все раздражения, поступающие в нервную систему, передаются на нейрон через определенные участки его мембраны, находящиеся в области синаптических контактов. В большинстве нервных клеток эта передача осуществляется химическим путем с помощью медиаторов. Ответом нейронов на внешнее раздражение является изменение величины, мембранного потенциала.
Чем больше синапсов на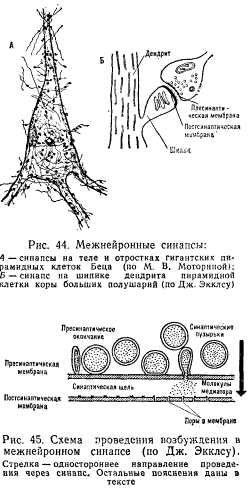 нервной клетке, тем больше
воспринимается различных раздражений и, следовательно, шире сфера влияний на ее
деятельность и возможность участия нервной клетки в разнообразных реакциях
организма. На телах крупных мотонейронов спинного мозга насчитывают до
15000—20000 синапсов. Разветвления аксонов могут образовывать синапсы на
дендритах (аксодендритические синапсы) и на соме (теле) нервных клеток (аксосоматические
синапсы), в ряде случаев—на аксоне (аксоаксональные синапсы). Наибольшее число
(до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние
части и окончания дендритных отростков, причем многие контакты расположены на
специальных шипиковидных выростах, или шипиках (рис. 44), которые еще больше
увеличивают воспринимающую поверхность нейрона. В мотонейронах спинного мозга и
пирамидных клетках коры поверхность дендритов в 10—20 раз больше поверхности
тела клетки.
нервной клетке, тем больше
воспринимается различных раздражений и, следовательно, шире сфера влияний на ее
деятельность и возможность участия нервной клетки в разнообразных реакциях
организма. На телах крупных мотонейронов спинного мозга насчитывают до
15000—20000 синапсов. Разветвления аксонов могут образовывать синапсы на
дендритах (аксодендритические синапсы) и на соме (теле) нервных клеток (аксосоматические
синапсы), в ряде случаев—на аксоне (аксоаксональные синапсы). Наибольшее число
(до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние
части и окончания дендритных отростков, причем многие контакты расположены на
специальных шипиковидных выростах, или шипиках (рис. 44), которые еще больше
увеличивают воспринимающую поверхность нейрона. В мотонейронах спинного мозга и
пирамидных клетках коры поверхность дендритов в 10—20 раз больше поверхности
тела клетки.
Чем сложнее интегративная функция нейрона, тем большее развитие имеют аксодендритические синапсы (в первую очередь те, которые расположены на шипиках). Особенно они характерны для нейрональных связей пирамидных клеток в коре больших полушарий.
Промежуточные нейроны (например, звездчатые клетки коры) таких шипиков лишены.
Приходящие в пресинаптическую часть контакта нервные импульсы вызывают опорожнение синоптических пузырьков с выведением медиатора в синаптическую щель (рис. 45). Веществами, передающими нервные влияния в синапсах нервных клеток, или медиаторами, могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных ганглиях), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Диаметр пузырьков примерно равен ширине синаптической щели. В клетках передней центральной извилины коры больших полушарий у людей 18 — 30 лет синаптические пузырьки имеют диаметр 250 — 300 ангстрем при ширине синаптической щели 200 — 300 ангстрем. Выделение медиатора облегчается тем, что синаптические пузырьки скапливаются вблизи от синаптической щели— в так называемых активных, или оперативных, зонах. Чем больше нервных импульсов проходит через синапс, тем больше пузырьков перемещается в эту зону и прикрепляется к пресинаптической мембране. В результате облегчается выделение медиатора последующими нервными импульсами.
Эффекты, возникающие при активации синапса, могут быть возбуждающими или тормозящими. Это зависит от качества медиатора и свойств постсинаптической мембраны. Возбуждающие нейроны выделяют возбуждающий медиатор, а тормозные — тормозной. Кроме того, один и тот же медиатор может оказывать различное воздействие в разных органах (например ацетилхолин возбуждает скелетные мышечные волокна и тормозит сердечные).
В состоянии покоя мембрана поляризована: снаружи регистрируется положительный заряд, а внутри — отрицательный. В нервной клетке мембранный потенциал покоя равен примерно 70 мв.
При возбуждающих влияниях увеличение проницаемости мембраны обусловливает вхождение положительно заряженных ионов натрия в клетку и, следовательно, уменьшение разности потенциалов по обе стороны мембраны, т. е. ее деполяризацию. В постсинаптической мембране данного участка клетки при этом регистрируется небольшое отрицательное колебание мембранного потенциала с амплитудой около 10 мв, или возбуждающий постсинаптический потенциал (сокращенно ВПСП), нарастающий примерно за 1, 2 мсек. до своего максимума и затем убывающий.
При тормозных воздействиях проницаемость мембраны увеличивается незначительно — главным образом для ионов калия (диаметр гидратированного иона калия меньше, чем диаметр иона натрия). Поскольку ионов калия больше внутри клетки, они выходят из нее наружу, увеличивая поляризацию мембраны, т. е. вызывают ее гиперполяризацию. При этом регистрируется положительное колебание с амплитудой около 5 мв — тормозной постсинаптический потенциал (сокращенно ТПСП). Обычно ВПСП и ТПСП длятся всего несколько миллисекунд. Однако в некоторых нейронах головного мозга постсинаптические потенциалы могут иметь значительно большую длительность: ВПСП — до 80 мсек., ТПСП — более 100 мсек.
Интегративная функция нейрона. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов на теле и дендритах клетки. На мембране нейрона происходит процесс алгебраического суммирования положительных и отрицательных колебаний потенциала. При одновременной активации нескольких возбуждающих синапсов общий ВПСП нейрона представляет собой сумму отдельных местных ВПСП каждого синапса. При одновременном возникновении двух различных синаптических влияний — ВПСП и ТПСП — происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний. Преобладание тормозных синаптических воздействий приводит к гиперполяризации мембраны и торможению деятельности клетки. При сдвиге мембранного потенциала в сторону деполяризации повышается возбудимость клетки. Ответный разряд нейрона возникает лишь тогда, когда изменения мембранного потенциала достигают порогового значения — критического уровня деполяризации. Для этого величина ВПСП клетки должна составлять примерно 10 мв.
В крупных (афферентных и эфферентных) нейронах возбудимость различных участков мембраны неодинакова. В области начального сегмента нервной клетки (аксонного холмика и начальной немиелинизированной части аксона) имеется низкопороговая зона, мембрана которой обладает в несколько раз более высокой возбудимостью, чем на других участках клетки (порог возбудимости мембраны начального сегмента равен 10 мв, а порог возбудимости соматодендритической мембраны—20—30 мв). В этой зоне с момента достижения критического уровня деполяризации начинается лавинообразное вхождение натрия в клетку и регистрируется потенциал действия (ПД). В ПД различают кратковременную высоковольтную часть, или спайк (пик), и длительные низкоамплитудные колебания — следовые потенциалы. ПД мотонейронов имеют амплитуду пика около 80 — 100 мв и длительность его около 1,5 мсек.
Эффекторная функция нейрона. С появлением ПД, который в отличие от местных изменений мембранного потенциала (ВПСП и ТПСП) является распространяющимся процессом, нервный импульс начинает проводиться от тела нервной клетки вдоль по аксону к другой нервной клетке или рабочему органу, т. е. осуществляется эффекторная функция нейрона. Синапсы, расположенные ближе к возбудимой низкопороговой зоне на теле клетки (аксосоматические), оказывают большее влияние на возникновение потенциала действия, чем более удаленные, расположенные на окончаниях дендритов (аксодендритические). Импульсы, приходящие через аксосоматический синапс, как правило, вызывают ответный разряд нейрона, а импульсы, действующие на аксодендритический синапс —лишь подпороговое изменение его возбудимости. Так, разряды мотонейронов спинного мозга и пирамидных нейронов коры, вызывающие двигательные реакции организма, являются ответом на специфические аксосоматические влияния. Но возникнет ли этот ответ или нет, определяется характером воздействий, поступающих через аксодендритические синапсы от других нервных путей. Так складываются адекватные реакции, зависящие от многих раздражении, действующих на организм в данный момент времени, и осуществляется тонкое приспособление поведения к меняющимся условиям внешней среды.
Процессы, происходящие в активном нейроне, можно представить в виде следующей цепи: потенциал действия в пресинаптическом окончании предыдущего нейрона —> выделение медиатора в синаптическую щель —> увеличение проницаемости постсинаптической мембраны —> ее деполяризация (ВПСП) или гиперполяризация (ТПСП) —> взаимодействие ВПСП и ТПСП на мембране сомы и дендритов нейрона —> сдвиг мембранного потенциала в случае преобладания возбуждающих влияний —> достижение критического уровня деполяризации —> возникновение потенциала действия в низкопороговой зоне (мембране начального сегмента) нейрона —> распространение потенциала действия вдоль по аксону (процесс проведения нервного импульса) —> выделение медиатора в окончаниях аксона (передача нервного процесса на следующий нейрон или на рабочий орган).
Таким образом, передача информации в нервной системе происходит с помощью двух механизмов — электрического (ВПСП, ТПСП, потенциалы действия) и химического (медиаторы).
§ 2. Физиологические показатели функционального состояния нейрона
Величина мембранного потенциала является основным параметром, который определяет значения важнейших показателей функционального состояния нейрона — его возбудимость и лабильность.
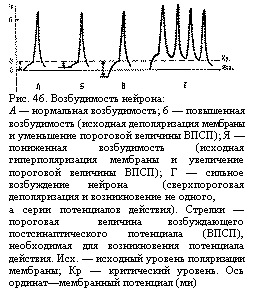
Возбудимость нейрона. Возбудимость нейрона — это его способность отвечать на синоптическое воздействие потенциалом действия. Она зависит от соотношения двух параметров—мембранного потенциала и критического уровня деполяризации (рис. 46). В нормальных условиях деятельности критический уровень деполяризации нейрона относительно постоянный, поэтому возбудимость нейрона определяется в основном величиной мембранного потенциала.
Подпороговые изменения мембранного потенциала. Мембранный потенциал может иметь нормальную величину (для нервной клетки—около 70 мв), уменьшенную (явление деполяризации) или большую (явление гиперполяризации). Для появления ПД при нормальном уровне мембранного потенциала требуется его сдвиг примерно на 10 мв, т. е. амплитуда порогового ВПСП должна быть около 10 мв для достижения критического уровня деполяризации. Эта величина и называется порогом возбудимости нейрона (см. рис. 46, Л).
При активной деятельности нейрона в нем может некоторое время сохраняться состояние подпороговой деполяризации. Такое же состояние возникает при слабых синаптических воздействиях, когда появляются подподпороговые ВПСП. При этом разница между уровнем мембранного потенциала и критическим уровнем деполяризации уменьшается (см. рис. 46, Б), т. е. требуется уже меньшая сила раздражения (меньшая амплитуда порогового ВПСП) для появления ответного ПД. Возбудимость нейрона в этом случае повышена.
При понижении возбудимости нервной клетки разница между уровнем мембранного потенциала и критическим уровнем деполяризации увеличена и, следовательно, требуется значительно большее, чем в норме, раздражение (большая амплитуда ВПСП) для появления ПД (см. рис. 46, В).
Сверхпороговое изменение мембранного потенциала. При сильном возбуждении в нервной клетке могут возникать ВПСП очень большой амплитуды, которая значительно превосходит критический уровень деполяризации и длительное время сохраняется на этом сверхпороговом уровне. В результате в течение всего периода сверхпороговой деполяризации нейрон генерирует потенциалы действия (см. рис. 46, Г). Такие резкие сдвиги возбудимости нейронов возникают, например, под влиянием вышележащих отделов центральной нервной системы. Импульсы, приходящие к мотонейронам спинного мозга из головного мозга, способны вызывать мощные ВПСП с амплитудой 20—25 мв, которые гораздо выше пороговой величины (10 мв). При этом мотонейрон отвечает на одно раздражение залпом нескольких ПД. Этим достигается значительное усиление двигательного ответа при рефлекторных реакциях.
 Величина
деполяризации нервных клеток находится в линейной зависимости от частоты,
раздражающих импульсов. Высшие отделы мозга, посылая импульсы различной частоты
к нейронам нижележащих отделов, регулируют их возбудимость, осуществляя контроль
за ответными реакциями организма.
Величина
деполяризации нервных клеток находится в линейной зависимости от частоты,
раздражающих импульсов. Высшие отделы мозга, посылая импульсы различной частоты
к нейронам нижележащих отделов, регулируют их возбудимость, осуществляя контроль
за ответными реакциями организма.
Изменения возбудимости нейрона при его деятельности. Протекание каждого ПД сопровождается закономерными изменениями возбудимости нейрона. В момент возникновения высоковольтной части — спайка нервная клетка не может ответить на раздражение новым ПД, т. е. является абсолютно невозбудимой. Эта фаза (абсолютная рефракторная фаза) длится примерно 0,5 мсек. Затем, в период постепенного снижения деполяризации мембраны, возбудимость нейрона восстанавливается до исходного уровня (фаза пониженной возбудимости, или относительная рефракторная фаза) и даже некоторое время может его превышать (супернормальная, или экзальтационная, фаза). После этого может наблюдаться еще некоторое понижение возбудимости во время развития следовой гиперполяризации (субнормальная фаза). Длительность описанных фаз изменения возбудимости определяет протяженность периодов повышенной активности (нормальная и супернормальная возбудимость) и пониженной активности (абсолютная и относительная рефракторные фазы, субнормальная фаза) нейронов. Так, большинство нейронов промежуточного мозга и коры больших полушарий после очередного разряда импульсов во время протекания абсолютной и относительной рефракторной фаз неактивны в течение примерно 100 мсек. Следовательно, наиболее удобным ритмом их спонтанной активности может быть ритм около 10 разрядов в 1 сек., который и является одним из факторов, определяющих частоту электрических колебаний на поверхности мозга в состоянии покоя.
Лабильность нейрона. Лабильность нейрона—это скорость протекания элементарных реакций, лежащих в основе его возбуждения. Наиболее удобная мера лабильности нейрона—максимальная скорость протекания ПД. В качестве меры лабильности используют также максимальную частоту импульсов, воспроизводимых нейроном без трансформации в единицу времени, или оптимальную, наиболее устойчивую, частоту импульсов (текущая лабильность). Разные по функциям и величине нервные клетки обладают различными величинами лабильности. Даже в пределах одного нейрона лабильность разных его структур (дендритов, сомы, начального сегмента и аксона) резко различается.
Величина лабильности нейрона определяется уровнем его мембранного потенциала.
Лишь при определенной величине мембранного потенциала достигается оптимальный уровень возбудимости и лабильности нервной клетки, а также наиболее высокий уровень ее ритмической активности, что является важным условием для передачи информации
нервной системе и осуществления целесообразных реакций. Например, наибольший максимальный ритм мотонейронов спинного мозга (до 100 импульсов в 1 сек.) отмечается при средней, оптимальной, величине мембранного потенциала—55 мв (т. е. при повышенной возбудимости мотонейрона).
В естественных условиях деятельности частота разрядов мотонейрона обычно не превышает 50—60 импульсов в 1 сек. Лишь при мощных влияниях вышележащих отделов нервной системы резко изменяется мембранный потенциал мотонейронов и максимальная частота их разрядов может повыситься до 100 и даже до 300 импульсов в 1 сек. Однако разряды с такой частотой очень кратковременны. Они сменяются более медленным и устойчивым ритмом активности. В мелких промежуточных нейронах спинного мозга частота разрядов в начальные моменты активации достигает 600— 800 импульсов в 1 сек., а иногда превышает 1000 импульсов в 1 сек. Такая частота необходима для резкого и быстрого воздействия этих клеток на мотонейроны; через некоторое время она резко замедляется.
Изменения в нейронах при активном и бездеятельном состояниях. Афферентные раздражения, возникающие при различных влияниях внешней и внутренней среды, в том числе при мышечной работе (с рецепторов мышц и сухожилий), повышают возбудимость и лабильность нейронов, их способность к стабильной ритмической активности. Бездеятельное же состояние снижает эти показатели, ухудшая функциональное состояние нервной клетки.
Активная деятельность клетки вызывает не только функциональные изменения, но и значительные структурные и биохимические перестройки различных ее элементов. Прогрессивные изменения наблюдаются в синаптическом аппарате нейронов. Эксперименты с тренировкой животных (тренировка задних конечностей собаки путем массажа, электрического раздражения и пассивной гимнастики, тренировка крыс с помощью плавания) показали, что средняя величина (диаметр) синапсов на мотонейронах поясничного отдела спинного мозга достоверно возрастает (примерно на 35%). С увеличением размеров синапсов улучшается проведение нервных импульсов по рефлекторным путям.
Активное участие корковых нейронов в условнорефлекторной деятельности вызывает усиленное развитие шипикового аппарата на дендритах. Это способствует улучшению межнейронных взаимосвязей в коре больших полушарий. Подавление условнорефлекторной деятельности различными фармакологическими воздействиями приводит к ухудшению рецепторной функции нейронов — нарушению синаптических контактов (исчезновению шипиков), а затем и к Деформации дендритов.
При усилении афферентной импульсации, поступающей к нейрону, в нейрональных синапсах происходят увеличение числа синаптических пузырьков, их усиленное новообразование и интенсивное перемещение в оперативную зону пресинаптической мембраны. Кроме того, в пресинаптических образованиях отмечается еще большее усиление (в 7 раз по сравнению с покоем) синтеза медиатора (ацетилхолина) и выраженное увеличение его выделения (в 70 раз). В нейронах повышается уровень окислительных процессов, возрастает количество белков и РНК в телах нервных клеток и в клетках глии.
При усиленной двигательной активности увеличиваются размеры эффекторных нейронов—пирамидных клеток коры и мотонейронов спинного мозга.
Все прогрессивные структурные, биохимические и функциональные изменения улучшают осуществление нервными клетками их сложных функций. Это лежит в основе совершенствования различных поведенческих реакций целого организма, в том числе двигательной деятельности и психических процессов у человека.
§ 3. Системы нервных клеток
Диффузная нервная сеть и процесс централизации. У простейших одноклеточных животных одна клетка осуществляет разнообразные функции. Передвижение этих животных обеспечивается либо выпячиванием части клетки (ложноножки) и переливанием в нее содержимого клетки (амебоидное движение, характерное и для белых клеток крови человека), либо с помощью специальных образований — ресничек или жгутиков.
Усложнение деятельности организма привело к разделению функций различных клеток — их специализации. Одни из них приобрели способность к сокращению (мышечные клетки), другие — к восприятию внешних и внутренних раздражении, переработке поступающей информации и передаче управляющих сигналов на органы движения и другие органы тела (нервные клетки). Наиболее простая форма организации нервной системы — диффузная нервная сеть низших беспозвоночных животных (губок, гидр, актиний и медуз). Нейроны такой сети имеют многочисленные взаимные связи, по которым возбуждение распространяется диффузно, по всем направлениям.
Более сложной формой является сегментарная организация нервной системы у высших беспозвоночных животных (червей, насекомых). Тело их состоит из имеющих одинаковое строение участков — сегментов, иннервация которых осуществляется расположенными в этих же сегментах нервными клетками.
С усложнением поведенческих реакций происходит развитие у позвоночных животных головного конца нервной системы—энцефалона. В нем сосредоточиваются группы нервных клеток, управляющих важнейшими функциями, — нервные центры. При этом утрачивается автономность отдельных сегментов и все большая часть функций передается вышележащим отделам нервной системы. Этот процесс получил название энцефализации (централизация) функций.
С формированием коры больших полушарий происходит подчинение всех других отделов нервной системы, т. е. процесс кортикализации.
Строение и функции центральной нервной системы у человека отражают этот
эволюционный процесс. Более древние и менее сложно организованные ее отделы
имеют более выраженную, сегментарную организацию. Это спинной мозг, продолговатый
и средний, многие центры которых иннервируют расположенные на том же уровне
участки тела (рис. 47). Находящиеся над ними более мо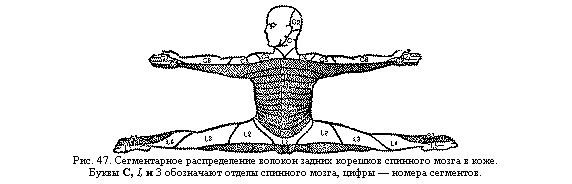 лодые
отделы, так называемые надсегментарные—промежуточный мозг, мозжечок, кора
больших полушарий,—не имеют непосредственной связи с органами тела, а управляют
их деятельностью через сегментарные центры. Надсегментарные отделы приобретают
по мере совершенствования все более важную роль в жизнедеятельности органов.
Они не только подчиняют, но и значительно перестраивают функции нижележащих
структур, оставляя им лишь самые элементарные рефлексы. Так, отделение спинного
мозга от головного вызывает угнетение всех его рефлексов—спинальный шок,
который выражен гораздо слабее и длится меньше (всего несколько минут) У рыб и
земноводных и более глубок и длителен (несколько недель) у млекопитающих. После
исчезновения явлений шока спинальная лягушка (лишенная головного мозга) может
осуществлять довольно сложные координированные движения (например, удаление
пальцами задней лапки кислотного раздражителя с бедра), у спинальных кошек и
собак могут наблюдаться сгибание, разгибание, маятникообразные движения
конечностей, а у человека при перерыве связи головного мозга со спинным —
практически только сгибательные рефлексы (разгибательные движения у него ниже
места перерыва спинного мозга почти не наблюдаются, перекрестный разгибательный
рефлекс выражен намного слабее, чем у спинальной собаки).
лодые
отделы, так называемые надсегментарные—промежуточный мозг, мозжечок, кора
больших полушарий,—не имеют непосредственной связи с органами тела, а управляют
их деятельностью через сегментарные центры. Надсегментарные отделы приобретают
по мере совершенствования все более важную роль в жизнедеятельности органов.
Они не только подчиняют, но и значительно перестраивают функции нижележащих
структур, оставляя им лишь самые элементарные рефлексы. Так, отделение спинного
мозга от головного вызывает угнетение всех его рефлексов—спинальный шок,
который выражен гораздо слабее и длится меньше (всего несколько минут) У рыб и
земноводных и более глубок и длителен (несколько недель) у млекопитающих. После
исчезновения явлений шока спинальная лягушка (лишенная головного мозга) может
осуществлять довольно сложные координированные движения (например, удаление
пальцами задней лапки кислотного раздражителя с бедра), у спинальных кошек и
собак могут наблюдаться сгибание, разгибание, маятникообразные движения
конечностей, а у человека при перерыве связи головного мозга со спинным —
практически только сгибательные рефлексы (разгибательные движения у него ниже
места перерыва спинного мозга почти не наблюдаются, перекрестный разгибательный
рефлекс выражен намного слабее, чем у спинальной собаки).
Соматотопическое представительство функций. Системы нейронов, образующих восходящие пути от рецепторов или нисходящие пути к рабочим органам, расположены в определенном порядке — по типу проекции «точку в точку». Так, каждый участок воспринимающей поверхности глаза (сетчатой оболочки с фоторецепторами) передает свои сигналы определенному участку в зрительной области коры больших полушарий. Эти чувствительные корковые центры расположены таким образом, что они как бы образуют экран, отражающий расположение фоторецепторов на периферии. Следовательно, наш мозг получает объективную информацию о пространственных особенностях внешнего мира. Определенная пространственная организация в нервной системе сигналов от различных органов чувств (от зрительных, слуховых, мышечных, тактильных и других рецепторов) позволяет человеку иметь необходимое представление о схеме пространства.
Таким же образом в мозгу обеспечивается представление о схеме тела. Порядок размещения афферентных волокон в проводящих путях и локализация нервных центров в проекционных областях разных этажей нервной системы соответствуют порядку размещения участков кожи и скелетных мышц в теле. Этому принципу отвечает представительство моторных функций в различных этажах нервной системы. Так, например, в самой верхней части передней центральной извилины коры больших полушарий находятся центры, посылающие импульсы к мышцам нижних конечностей, несколько ниже — моторные центры мышц туловища, еще ниже—моторное представительство мышц верхних конечностей и, наконец, центры мышц шеи и головы. Таков же порядок расположения этих центров в моторных ядрах нижележащих этажей нервной системы (например, в красном ядре среднего мозга).
Такая организация связей в центральной нервной системе, при которой пространственное расположение нейронов, связанных с определенными рецепторами или рабочими органами на периферии, отражает специфику их пространственного расположения в организме, называется соматопическим (от лат. somaticus — телесный и topicus — местный) или экранным представительством функций. В соматопической организации нервной системы важное значение имеют: 1) функциональная значимость отдельного органа тела и 2) роль данного участка мозга в его иннервации. В связи с этим возникает несоответствие размеров органа тела и размеров территории мозга, которая управляет его деятельностью. Например, мышцы пальцев руки выполняют чрезвычайно сложные функции в произвольных движениях человека. В их деятельности основную роль играют импульсы, идущие от коры больших полушарий. Этим объясняется то, что представительство (как сенсорное, так и в особенности моторное) этих мышц в коре занимает значительно большую территорию, чем представительство всех остальных мышц тела.
Механизмы взаимодействия нервных клеток. Нервные клетки функционируют в тесном взаимодействии друг с другом.
Значение нервных импульсов. Все взаимодействия между нервными клетками осуществляются благодаря двум механизмам: 1) влияниям электрических полей нервных клеток (электротоническим влияниям) и 2) влияниям нервных импульсов.
Первые распространяются на очень небольшие территории мозга Электрический заряд нервной клетки создает вокруг нее электрическое поле, колебания которого вызывают изменения электрических полей лежащих рядом нейронов, что приводит к изменениям их возбудимости, лабильности и проводимости. Электрическое поле нейрона имеет сравнительно небольшую протяженность—около 100 мк, оно быстро затухает по мере удаления от клетки и может оказывать воздействие лишь на соседние нейроны.
Второй механизм обеспечивает не только ближайшие взаимодействия, но и передачу нервных влияний на большие расстояния. Именно с помощью нервных импульсов происходит объединение отдаленных и изолированных участков мозга в общую, синхронно работающую систему, что необходимо для протекания сложных форм деятельности организма. Нервный импульс, следовательно, является основным средством связи между нейронами. Высокая скорость распространения импульсов и локальное их воздействие на избранную точку мозга способствуют быстрой и точной передаче информации в нервной системе. В межнейронных взаимодействиях используется частотный код, т. е. изменения функционального состояния и характера ответных реакций одной нервной клетки кодируются изменением частоты импульсов (потенциалов действия), которые она посылает к другой нервной клетке. Общее количество импульсов, отправляемых нервной клеткой в единицу времени, или ее суммарная импульсная активность,—важный физиологический показатель деятельности нейрона.
Значение синапсов. В передаче нервных импульсов от одного нейрона к другому большое значение имеют синапсы. Функционально возбуждение передается только через синапсы. Они выступают как переключатели, которые могут быть еще не включены (до образования долговременной проводимости-памяти) или уже хорошо проводить возбуждение, запуская от пришедшего стимула запомненную связь с ним последующих звеньев, что и обуславливает механизм долговременной памяти. Особенно много их в высших отделах нервной системы и у нейронов с наиболее сложными функциями. С совершенствованием условнорефлекторной деятельности в процессе эволюции животных организмов увеличивается число синаптических контактов на пирамидных нейронах коры больших полушарий, в частности за счет увеличения числа шипиков на их дендритах. Состояние синапсов на этих шипиках коррелирует с состоянием высшей нервной деятельности. При подавлении условных рефлексов некоторыми веществами (например, аминазином) контакты на шипиках разрушаются, исчезают сами шипики, а затем деформируются и дендритные отростки нейрона.
Способность синапсов передавать нервные импульсы непостоянна. Она повышается после активной деятельности синапса и снижается при отсутствии активности. Понижение функциональных возможностей синапсов (гипосинапсия) ведет к ухудшению проведения через них нервных импульсов, а их полное нарушение (асинапсия) вызывает окончательное разобщение нервных клеток.
При отсутствии активации синапса в течение нескольких дней уже наблюдаются уменьшение размера синаптических пузырьков, запасов медиатора, нарастание длительности синаптической задержки и т. п. Бездействие синаптических контактов на мотонейронах спинного мозга, вызванное перерезкой чувствительных путей от соответствующих мышечных волокон, через 2—4 недели уменьшает их возбуждающее влияние на мембрану мотонейрона вдвое по сравнению с влияниями других афферентных путей, оканчивающихся на том же мотонейроне. У кролика через несколько дней содержания в абсолютной темноте, т. е. после полного бездействия рецепторных клеток глаза, уменьшаются синаптические пузырьки в окончаниях их аксонов. Это объясняет постепенное угасание приобретенных навыков (хотя механизм забывания этим не исчерпывается). Длительность этого процессам может быть от нескольких минут до десятков лет, в зависимости от локализации (см ниже).
Активная деятельность нервных клеток способствует улучшению синоптического взаимодействия между ними: росту площади синаптических контактов, ускорению проведения через синапсы и т. д. Это наблюдается, например, при двигательной деятельности. Как показали эксперименты на животных, ежедневная принудительная ходьба их по 1 часу приводит к значительной активации синапсов в соответствующих рефлекторных дугах спинного мозга.
Синапсы, являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения — сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов — их разрушение, в результате чего полностью нарушается нервная деятельность.
§ 4. Свойства нервных центров
Понятие о нервном центре. В сложных многоклеточных организмах животных и человека отдельная нервная клетка не в состоянии регулировать какие-либо функции. Все основные формы деятельности нервной системы связаны с участием в ее функциях определенных групп нервных клеток—нервных центров.
Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния).
Для выявления функций нервного центра используют ряд методов: метод раздражения (усиление его деятельности), метод разрушения или удаления — экстирпации (нарушение регулируемой им функции), электрофизиологический метод — регистрация электрических явлений в нервном центре и др. Так, функциональное значение, а также локализация дыхательного центра в продолговатом мозгу были выявлены путем последовательных перерезок головного мозга. При перерезках выше области его расположения (выше продолговатого мозга) дыхание сохранялось, а при перерезках ниже продолговатого мозга дыхательные движения исчезали.
Сложные реакции в целостном организме обычно связаны с участием многих нервных центров, расположенных в различных этажах центральной нервной системы. Например, произвольное изменение дыхания осуществляется человеком при участии центров коры больших полушарий, особого центра в среднем мозгу, дыхательного центра продолговатого мозга и центров спинного мозга, иннервирующих дыхательные мышцы.
Физиологические свойства нервных центров определяют характер ответных реакций. Эти свойства в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки.
Одностороннее проведение возбуждения. В отличие от нервного волокна, в котором возможно двустороннее проведение волны возбуждения от места нанесения раздражения, в нервных центрах волна возбуждения проводится только в одном направлении: раздражение задних (чувствительных) корешков спинного мозга вызывает возникновение нервного импульса в передних двигательных корешках, однако при раздражении передних корешков волна возбуждения в задних корешках не появляется. Это обусловлено особенностями синаптической передачи возбуждения в отдельных нервных клетках.
Поскольку проведение волны возбуждения от одного нейрона к другому через синапс происходит химическим путем—с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране, — проведение нервных влияний через синапс возможно только от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении (см. рис. 45). В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление: от афферентных нейронов к вставочным и затем к эффекторным — мотонейронам и вегетативным нейронам.
Замедленное проведение возбуждения. В основе проведения нервных
импульсов по цепочке нейронов лежат два различных механизма: электрический
(проведение потенциала действия — ПД — по нервным волокнам) и химический
(передача через синапс с помощью медиатора). Первый осуществляется с большой
скоростью (до 100—140 м/сек), второй—в тысячу раз медленнее. Замедление
проведения связано с затратой времени на процессы, происходящие от момента
прихода пресинаптического импульса в синапс до появления в постсинаптической
мембране возбуждающих или тормозных потенциалов. Этот интервал называется синаптической
задержкой и составляет в мотонейронах и большинстве других клеток Центральной
нервной системы примерно 0,3 мсек. В вегетативной нервной системе длительность синаптической
задержки больше — не менее 10 мсек. За это время пресинаптический импульс
вызывает опорожнение синаптических пузырьков, происходит диффузия медиатора
через синаптическую щель, увеличение под его влиянием ионной проницаемости
постсинаптической мембраны и возникает постсинаптический потенциал. После этого
требуется еще около 1,2 мсек. на развитие возбуждающего постсинаптического
потенциала (ВПСП) до максимума, чтобы возник (в случае достижения критического
уровня деполяризации) ПД. Таким образом, весь процесс передачи нервного
импульса (от ПД одной клетки до ПД следующей клетки) через один синапс занимает
примерно 1,5 мсек. При утомлении, охлаждении и ряде других воздействий
длительность синаптической задержки возрастает. Если же для осуществления
какой-либо реакции требуется участие большого числа нейронов (многих сотен и
даже тысяч), то суммарная величина задержки проведения по нервным центрам — так
называемое центральное время проведения — может составить десятые доли секунды
и даже целые секунды. Поскольку проведение импульсов по нервным 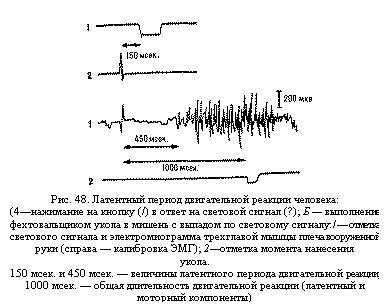 волокнам
от периферических рецепторов в нервные центры и от нервных центров к
исполнительным органам занимает сравнительно небольшое время, общее время от
момента нанесения внешнего раздражения до появления ответной реакции организма
(латентный период рефлекса) определяется центральным временем проведения.
Величина латентного периода рефлекса служит важным показателем функционального
состояния нервных центров и широко используется в практике. Латентный период
двигательной реакции измеряется от момента подачи сигнала до момента начала
движения (или до появления электрической активности соответствующей мышцы). При
осуществлении человеком простых движений в ответ на внешние сигналы, например
нажимание на кнопку при световом сигнале (рис. 48, А), длительность латентного
периода двигательной реакции составляет около 120 – 220 мсек., а при утомлении,
недостатке кислорода и пр. она может увеличиваться до 300 – 500 и более
миллисекунд. При более сложных движениях, например выполнении фехтовальщиком
укола в мишень с выпадом в ответ на световой сигнал, этот период значительно
больше – около 300 – 400 мсек. и более (см. рис. 48, Б).
волокнам
от периферических рецепторов в нервные центры и от нервных центров к
исполнительным органам занимает сравнительно небольшое время, общее время от
момента нанесения внешнего раздражения до появления ответной реакции организма
(латентный период рефлекса) определяется центральным временем проведения.
Величина латентного периода рефлекса служит важным показателем функционального
состояния нервных центров и широко используется в практике. Латентный период
двигательной реакции измеряется от момента подачи сигнала до момента начала
движения (или до появления электрической активности соответствующей мышцы). При
осуществлении человеком простых движений в ответ на внешние сигналы, например
нажимание на кнопку при световом сигнале (рис. 48, А), длительность латентного
периода двигательной реакции составляет около 120 – 220 мсек., а при утомлении,
недостатке кислорода и пр. она может увеличиваться до 300 – 500 и более
миллисекунд. При более сложных движениях, например выполнении фехтовальщиком
укола в мишень с выпадом в ответ на световой сигнал, этот период значительно
больше – около 300 – 400 мсек. и более (см. рис. 48, Б).
 Укорочение
латентного периода одной и той же двигательной реакции человека в процессе
спортивной тренировки может отражать повышение функционального состояния
нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих
в передаче нервных импульсов (укорочение проводящего пути).
Укорочение
латентного периода одной и той же двигательной реакции человека в процессе
спортивной тренировки может отражать повышение функционального состояния
нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих
в передаче нервных импульсов (укорочение проводящего пути).
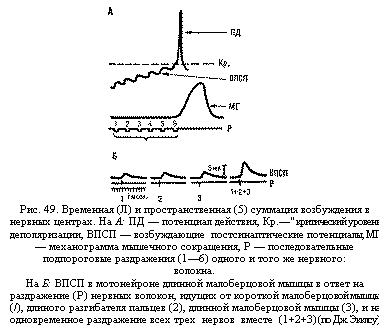
Суммация. В ответ на одиночную афферентную волну, идущую от мышц к мотонейронам, в пресинаптической части синапса освобождается 1 квант нейромедиатора. При этом в пресинаптической мембране мотонейрона обычно возникает ВПСП подпороговой величины – 0,1 – 0,2 мв, недостаточной для вызова ответной реакции клетки. Чтобы величина ВПСП достигла критического уровня (порога возникновения ПД – примерно 10 мв.), требуется суммация на мембране клетки многих подпороговых ВВСП. Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одновременное возбуждение синапсов в различных участках мембраны мотонейрона повышает амплитуду суммарного ВПСП до пороговой величины, возникает ответный импульс мотонейрона и рефлекторно осуществляется двигательная реакция. Так, при синхронном раздражении трех афферентных нервах волокон—от короткой малоберцовой мышцы, длинного разгибателя пальцев и длинной малоберцовой мышцы — в мотонейроне длинной малоберцовой мышцы регистрируется ВПСП, равный сумме отдельных ВПСП, получаемых при изолированном раздражении каждого волокна (рис. 49, Б}. Обычно требуется одновременная активация 50—100 афферентных волокон для получения ответа двигательной клетки спинного мозга. Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражении. Если интервалы между поступающими импульсами достаточно коротки и ВПСП мотонейрона от предыдущего раздражения не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны не Достигнет критического уровня для возникновения ПД (см. рис. 49, Л).
Явления суммации могут наблюдаться при осуществлении отдельных рефлексов (например, рефлексов чихания и кашля в ответ на слабые раздражения слизистой оболочки дыхательных путей). Они лежат в основе формирования нейронных комплексов, определяющих поведение целого организма, в основе развития условных рефлексов.
Фоновая активность. Многие нервные клетки обладают способностью к постоянной импульсной активности. Их мембранный потенциал периодически колеблется, то увеличиваясь, то уменьшаясь. Каждый раз, когда его увеличение превышает критический уровень деполяризации (порог возбудимости клетки), возникает импульсный разряд. Такая автоматическая деятельность называется фоновой активностью нейрона (рис. 50). Ее могут вызывать случайные импульсы, циркулирующие в нервной системе, но она может проявляться и при отсутствии приходящих к нейрону раздражении—в результате изменении ионных и обменных процессов, колебаний температуры и других причин. В последнем случае ее называют спонтанной.
 Фоновая
активность играет важную роль в деятельности целого организма. Во-первых,
импульсы, приходящие по афферентным путям, легче возбуждают клетки со
спонтанной активностью, чем
Фоновая
активность играет важную роль в деятельности целого организма. Во-первых,
импульсы, приходящие по афферентным путям, легче возбуждают клетки со
спонтанной активностью, чем
бездеятельные. Это облегчает протекание многих рефлекторных реакций. Во-вторых, клетки, обладающие постоянным ритмом фоновой активности, могут иметь значение водителя ритма (пейсмекера), т. е. задавать этот ритм другим нервным клеткам. При этом возникают большие комплексы синхронно работающих клеток, в которых благодаря общему ритму особенно хорошо выражено взаимодействие между отдельными нейронами. В-третьих, некоторые такие клетки выполняют роль счетчиков времени в центральной нервной системе. Их деятельность помогает поддерживать определенный ритм движений, субъективно оценивать интервалы времени, через определенные сроки включать необходимые функции и т. п.
Автоматическая деятельность присуща клеткам дыхательного центра. Их спонтанные разряды позволяют поддерживать ритмические дыхательные движения не только при бодрствовании, но и во время сна человека, в условиях наркоза; периодические разряды дыхательного центра регистрируются даже при полной изоляции продолговатого мозга от других отделов центральной нервной системы.
 Вызванная
активность. Импульсные разряды нейрона, возникающие в ответ на внешнее
раздражение, называются вызванной активностью. Ответ нервной клетки может
возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а
также в виде пачек импульсов, появляющихся через определенные интервалы. В
различных нейронах наблюдаются разные типы вызванной активности. Одни клетки
дают разряды только в момент включения раздражителя (эффект включения) —это
наиболее частая реакция клеток, другие — только в момент его выключения (эффект
выключения), третьи—в обоих случаях. В центральной нервной системе имеются
клетки, дающие разряды лишь на новые раздражители, что обеспечивает
ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная
активность при неправильных реакциях организма, т. е. сигнализирующие об
ошибках, что позволяет вносить коррекции в действия человека.
Вызванная
активность. Импульсные разряды нейрона, возникающие в ответ на внешнее
раздражение, называются вызванной активностью. Ответ нервной клетки может
возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а
также в виде пачек импульсов, появляющихся через определенные интервалы. В
различных нейронах наблюдаются разные типы вызванной активности. Одни клетки
дают разряды только в момент включения раздражителя (эффект включения) —это
наиболее частая реакция клеток, другие — только в момент его выключения (эффект
выключения), третьи—в обоих случаях. В центральной нервной системе имеются
клетки, дающие разряды лишь на новые раздражители, что обеспечивает
ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная
активность при неправильных реакциях организма, т. е. сигнализирующие об
ошибках, что позволяет вносить коррекции в действия человека.
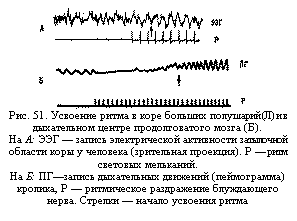
Усвоение и трансформация ритма. При ритмических раздражениях вызванная активность нейрона может настроиться на ритм приходящих импульсов (явление усвоения ритма—рис. 51). Эта настройка деятельности нейрона на заданный извне ритм (реакция усвоения, перестройки или навязывания ритма) имеет большое значение для организации взаимодействия между различными нервными центрами и различными этажами нервной системы, в частности для организации ритмических движений у человека.
Активность нейрона может перестроиться как на более высокий, так и на более низкий ритм. В результате такой способности многие нервные клетки получают возможность сонастраиваться, т. е. работать сообща, в одном комплексе, не мешая друг другу различной по темпу импульсацией.
Впервые вопрос о значении усвоения ритма для протекания реакций организма был поставлен А. А. Ухтомским и его учениками (Н. В. Голиковым, Е. К. Жуковым и др.). Особенно важно, что в процессе ритмического раздражения рефлекторной дуги нервные центры способны усвоить ритм активности, превышающий их первоначальную лабильность. Это явление лежит в основе повышения работоспособности отдельных рефлекторных дуг, а также целостного организма (в частности, это имеет место в спортивной тренировке).
Очень часто вызванная активность нейрона не соответствует ритму приходящей к данной клетке импульсации. Нервные клетки обладают свойством изменять частоту передающихся импульсов, т.е. свойством трансформации ритма. Характер ответного разряда зависит, во-первых, от свойств раздражителя и, во-вторых, от функционального состояния нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях чем сильнее раздражение, тем чаще проявляется ответная импульсация.
При высокой возбудимости нейрона может возникать учащение импульсации (мультипликация ритма). В этом случае афферентная волна вызывает длительные надпороговые ВПСП в клетке, и на .каждое одиночное раздражение клетка отвечает залпом из нескольких ПД (пока ВПСП превышает пороговый уровень). При низком уровне возбудимости и лабильности нейрона чаще происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения ПД. В результате на несколько приходящих импульсов нейрон отвечает лишь одним импульсом.
В некоторых крупных клетках, например мотонейронах спинного мозга, имеются специальные механизмы, регулирующие ритм ответных реакций. После каждого ПД в этих клетках наблюдаются выраженные следовые процессы — следовая гиперполяризация мембраны. В этот момент возбудимость клетки понижена. Следующий разряд возникает в клетке лишь тогда, когда заканчивается этот следовой потенциал. Поэтому ритм разрядов мотонейрона, направляющихся к скелетной мышце, обычно не превышает 60 импульсов в 1 сек. Мелкие клетки, например вставочные нейроны, такого механизма не имеют и могут давать в начальные моменты ответа очень высокую частоту разрядов—до 1600 импульсов в 1 сек., которая, однако, быстро затухает.
Следовые процессы. После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозгу (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы и даже дни) и очень велика в коре больших полушарий (до нескольких десятков лет). Различают явные и скрытые следы прежних раздражении. В явной форме сохраняются лишь кратковременные следовые процессы или последствия. Они могут быть одноименными — продолжение возникшего в клетке состояния (тормозное или экзальтационное последействие) или контрастными (возбуждение вслед за торможением, торможение вслед за возбуждением).
Поддерживать состояние возбуждения или торможения в нервном центре могут импульсы, блуждающие в нервных цепях—задержавшиеся на длинных путях передачи или возвращающиеся повторно к нейрону по замкнутым цепям нейронов.
Значительно сложнее по природе скрытые следы, характеризующиеся отсутствием импульсации в нервных центрах. Предполагают, что длительное сохранение в нервной клетке скрытых следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков (возможно, также и белков глиальных клеток).
Согласно биохимической теории памяти (X. Хиден), в процессе запоминания происходят структурные изменения в молекулах рибонуклеиновой кислоты (РНК), от которой зависит образование белков клетки. В результате этого в нервной клетке начинают синтезироваться «измененные» белки с отпечатками прежних раздражении, составляющие биохимическую основу памяти.
Эксперименты на крысах и других животных показали, что способность их запоминать следы прежних раздражении или способность к обучению коррелирует с изменением содержания рибонуклеиновой кислоты в нервной ткани. Интересные опыты были проведены на червях. У них вырабатывали условные рефлексы — включение света сопровождалось электрическим ударом, вызывающим у червя реакцию сжимания. После ряда сочетаний этих раздражении одно включение света заставляло червей сжиматься. Эту способность реагировать на включение света двигательной реакцией приобретали также и «необученные» черви после того, как их кормили кашицей из растолченных «обученных» червей или вводили им рибонуклеиновую кислоту, полученную из тканей «обученных» червей. Следовательно, приобретенный опыт может передаваться от одного организма к другому с химическими веществами.
При формировании новых двигательных навыков увеличивается содержание измененной рибонуклеиновой кислоты в соответствующих нервных центрах. Такие изменения, сохраняющиеся в двигательных нейронах, могут составлять основу так называемой «мышечной памяти». Известно, что человек, забывший в результате ранения одного из участков мозга начертания букв алфавита, может писать слова автоматически, пользуясь сохранившейся моторной памятью. Мышечная память позволяет человеку вспомнить заученные движения, которые он забыл после длительного перерыва.
Непродолжительные последействия (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные с биохимическими перестройками в клетках,—в основе долгосрочной памяти. Кратковременная память служит человеку недолго (например, спросив по телефону справочной необходимый номер телефона, можно набрать его и сразу забыть). Долгосрочная память обеспечивает длительное сохранение следов. Она имеет важнейшее значение в обучении человека (например, для усвоения определенного объема теоретических знаний, фиксирования в мозгу выработанных двигательных навыков).
§ 5. Координация деятельности центральной нервной системы
Для деятельности центральной нервной системы характерна определенная упорядоченность и согласованность рефлекторных реакций, т. е. их координация. Взаимодействие двух нервных процессов—возбуждения и торможения, лежащих в основе всех сложных регуляторных функций организма, закономерности их одновременного протекания в различных нервных центрах, а также последовательная смена во времени определяют точность и своевременность ответных реакций организма на внешние и внутренние воздействия.
Иррадиация и концентрация нервных процессов. Проведение афферентной волны по рефлекторной дуге вызывает в ее нервных центрах состояние возбуждения или торможения. Эти процессы при определенных условиях могут охватывать и другие рефлекторные центры. Распространение процесса возбуждения на другие нервные центры называют иррадиацией. Она осуществляется благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг, так что при раздражении одного рецептора возбуждение в принципе может распространяться в центральной нервной системе в любом направлении и на любую нервную клетку.
Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Это явление можно наблюдать на спинальной лягушке. Слабое давление на пальцы задней лапки вызывает ответный рефлекс сгибания этой же лапки. Небольшое усиление давления приводит к сгибанию другой задней лапки, хотя рецепторы последней не раздражаются. Этот ответ возникает в результате того, что в сферу возбуждения помимо нервных центров одноименной половины спинного мозга вовлекаются центры другой его половины. При еще более сильном раздражении волна возбуждения охватывает вышележащие и нижележащие нервные центры и вызывает движения верхних конечностей (сначала на стороне тела, подвергшейся раздражению, а затем на противоположной).
Аналогичное явление иррадиации возбуждения можно наблюдать при действии различных раздражении в коре больших полушарий.
Иррадиация через некоторое время сменяется явлением концентрации процессов возбуждения в том же исходном пункте центральной нервной системы. Концентрация происходит в несколько раз медленнее, чем иррадиация нервных процессов.
Процесс иррадиации играет положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Активация большого количества различных нервных центров позволяет отобрать из их числа наиболее нужные для последующей деятельности, т. е. совершенствовать ответные действия организма. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи — условные рефлексы. На этой основе возможно, например, формирование новых двигательных навыков.
Вместе с тем иррадиация возбуждения может оказать и отрицательное воздействие на состояние и поведение организма. Так, иррадиация возбуждения в центральной нервной системе нарушает тонкие взаимоотношения, сложившиеся между процессами возбуждения и торможения в нервных центрах, и приводит к расстройству двигательной деятельности.
Торможение в центральной нервной системе. Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г. Значение этого процесса было рассмотрено им в книге «Рефлексы головного мозга».
Опуская лапку лягушки в кислоту и одновременно раздражая некоторые участки головного мозга (например, накладывая кристаллик поваренной соли на область промежуточного мозга), И. М. Сеченов наблюдал резкую задержку и даже полное отсутствие «кислотного» рефлекса спинного мозга (отдергивания лапки). Отсюда он сделал заключение, что одни нервные центры могут существенно изменять рефлекторную деятельность в других центрах, в частности вышележащие нервные центры могут тормозить деятельность нижележащих. Описанный опыт вошел в историю физиологии под названием «Сеченовского торможения».
Тормозные процессы — необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает иррадиацию возбуждения, чем способствует его концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым выключает деятельность ненужных в данный момент органов, осуществляя координационную функцию. В-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
По месту возникновения различают пресинаптическое торможение и постсинаптическое.
Постсинаптическое торможение. Оно возникает в постсинаптической мембране нейрона в результате действия тормозного медиатора и связано с наличием в центральной нервной системе специальных тормозных нейронов. Это особый тип вставочных нейронов, у которых окончания аксонов выделяют тормозной медиатор. Природа тормозного медиатора в настоящее время точно не установлена. Нервные импульсы, подходя к тормозным нейронам, вызывают в них такой же процесс возбуждения, как и в Других нервных клетках. В ответ по аксону тормозной клетки распространяется обычный ПД, но в отличие от других нейронов окончания аксона при этом выделяют не возбуждающий, а тормозной медиатор. Под влиянием этого медиатора возникает кратковременная гиперполяризация постсинаптической мембраны следующего нейрона и регистрируется тормозной постсинаптический потенциал (ТПСП). В результате тормозные клетки не возбуждают, а тормозят те нейроны, на которых оканчиваются их аксоны. Такой вид торможения называют прямым, так как оно возникает сразу, без предварительного возбуждения.
 Специальные
тормозные нейроны. — это клетки Рэншоу в спинном мозгу и корзинчатые клетки в
промежуточном мозгу. Клеткам Рэншоу принадлежит важная роль в координации
деятельности спинного мозга. Большое значение, например, эти клетки имеют при
регуляции деятельности мышц-антагонистов. Они обеспечивают развитие торможения
в мотонейронах мышц-антагонистов (рис. 52, В), что облегчает осуществление
сокращения этих мышц. Клетки Рэншоу участвуют в регуляции уровня активности
отдельных мотонейронов, ограничивая (тормозя) чрезмерное их возбуждение.
Специальные
тормозные нейроны. — это клетки Рэншоу в спинном мозгу и корзинчатые клетки в
промежуточном мозгу. Клеткам Рэншоу принадлежит важная роль в координации
деятельности спинного мозга. Большое значение, например, эти клетки имеют при
регуляции деятельности мышц-антагонистов. Они обеспечивают развитие торможения
в мотонейронах мышц-антагонистов (рис. 52, В), что облегчает осуществление
сокращения этих мышц. Клетки Рэншоу участвуют в регуляции уровня активности
отдельных мотонейронов, ограничивая (тормозя) чрезмерное их возбуждение.
Корзинчатые клетки играют важную роль в регуляции деятельности высших отделов мозга — промежуточного мозга и коры больших полушарий. Они являются как бы воротами, которые пропускают или не пропускают импульсы, идущие в кору больших полушарий.
Это обусловлено тем, что эти клетки вызывают синхронное торможение большого числа связанных с ними нейронов промежуточного мозга, регулируя тем самым прохождение восходящих импульсов через эти нейроны, время поступления их в кору больших полушарий и ритм корковой активности (см. рис. 52, Г).
При возбуждении мотонейрона спинного мозга импульсы поступают по его аксону к мышечным волокнам и одновременно по коллатералям аксона — к тормозной клетке Рэншоу. Аксоны последней «возвращаются» к этому же мотонейрону, вызывая его торможение (см. рис. 52, Б). Следовательно, через клетку Рэншоу мотонейрон может сам себя затормаживать. Чем больше возбуждающих импульсов посылает мотонейрон на периферию (а значит, и к тормозной клетке), тем сильнее это возвратное торможение (разновидность постсинаптического торможения). Такая замкнутая система действует как механизм саморегуляции нейрона, ограничивая степень его возбуждения и предохраняя от чрезмерной активности.
Пресинаптическое торможение. Оно возникает перед синоптическим контактом — в пресинаптической области. Окончания аксонов одной нервной клетки образуют аксоаксональный синапс на окончании аксона другой нервной клетки и блокируют передачу возбуждения в последнем (см. рис. 52, Л). В области такого пресинаптического контакта развивается чрезмерно сильная деполяризация мембраны аксона, которая приводит к состоянию парабиоза (пессимального торможения, по Н. Е. Введенскому). Заторможенный концевой участок аксона прекращает проведение нервных импульсов к синапсу и, значит, к следующей нервной клетке. Этот вид торможения обусловливает ограничение притока афферентных импульсов к нервным центрам.
Доминанта. Активность нервных центров непостоянна, и преобладание активности одних из них над активностью других вызывает заметные перестройки в процессах координации рефлекторных реакций.
Исследуя особенности межцентральных отношении, А. А. Ухтомский обнаружил, что если в организме животного осуществляется сложная рефлекторная реакция, например повторяющиеся акты глотания, то электрические раздражения моторных центров коры не только перестают вызывать в этот момент движения конечностей, но и усиливают и ускоряют протекание начавшейся цепной реакции глотания, оказавшейся главенствующей. Аналогичное явление наблюдалось при отравлении фенолом передних отделов спинного мозга лягушки. Повышение возбудимости мотонейронов приводило к тому, что отравленная лапка отвечала рефлексом потирания (отряхивания) не только на непосредственное раздражение ее кожи кислотой, но и на самые разнообразные посторонние раздражители:
поднимание животного со стола в воздух, удар по столу, где оно сидит, прикосновение к передней лапке животного и др.
Подобные эффекты, когда разнообразные поводы вызывают не адекватный им ответ, а реакцию, уже подготовленную в организме, постоянно встречаются и в поведении человека (смысл этого точно передают, например, такие пословицы, как «у кого что болит, тот о том и говорит», «голодной куме все пирог на уме»).
В 1923 г. А. А. Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров.
Термином доминанта был обозначен господствующий очаг возбуждения в центральной нервной системе, определяющий текущую деятельность организма.
Основные черты доминанты следующие: 1) повышенная возбудимость нервных центров, 2) стойкость возбуждения во времени, 3) способность к суммации посторонних раздражении и 4) инерция доминанты. Доминирующий (господствующий) очаг может возникнуть лишь при определенном функциональном состоянии нервных центров. Одним из условий его образования является повышенный уровень возбудимости нервных клеток, который обусловливается различными гуморальными и нервными влияниями (длительными афферентными импульсациями, гормональными перестройками в организме, воздействиями фармакологических веществ, сознательным управлением нервной деятельностью у человека и пр.).
Установившаяся доминанта может быть длительным состоянием, которое определяет поведение организма на тот или иной срок. Способность стойко поддерживать возбуждение во времени — характерная черта доминанты. Однако далеко не всякий очаг возбуждения становится доминантным. Повышение возбудимости нервных клеток и их функционального значения определяется способностью суммировать возбуждение при поступлении любого случайного импульса.
Восходящие нервные импульсы могут направляться не только по прямому специфическому пути — в соответствующие проекционные зоны мозга, но и через боковые ответвления — в любые зоны центральной нервной системы (см. § 6 этой главы). В связи с этим при наличии в каком-либо участке нервной системы очага с оптимальным уровнем возбудимости этот очаг приобретает возможность повышать свою возбудимость за счет суммирования не только собственных афферентных раздражении, но и посторонних, адресованных другим центрам. Не сила возбуждения, а способность накапливать и суммировать его превращает нервный центр в доминирующий. Явления суммации лучше всего выражены лишь при умеренном, оптимальном, повышении возбудимости нейронов. Это выражается в том, что доминанта легче всего подкрепляется слабыми раздражениями и гасится сильными.
Чем больше нейронов вовлечено в данный очаг возбуждения, тем прочнее доминанта и тем больше она подавляет деятельность других отделов мозга, вызывая в них так называемое сопряженное торможение. Нервные клетки, входящие в доминирующий очаг, расположены не обязательно в одном участке нервной системы. Чаще всего они составляют определенную систему клеток (по А. А. Ухтомскому, «созвездие», или констелляцию, нейронов), расположенных в разных этажах головного и спинного мозга. Такими сложными являются, например, доминанты, обеспечивающие выполнение мышечной работы. Внешним их выражением могут быть стационарно поддерживаемые движение и рабочая поза, а также исключение в этот момент других движений и поз. В эти доминанты включаются клетки разнообразных областей коры больших полушарий и подкорковых отделов, связанных с организацией двигательной деятельности, а также клетки различных эмоциональных и вегетативных центров (дыхательного, сердечно-сосудистого, терморегуляционного и др.).
Объединение большого числа нейронов в одну рабочую систему происходит путем взаимного сонастраивания на общий темп активности, т. е. путем усвоения ритма. Одни нервные клетки снижают свой более высокий темп деятельности, другие повышают низкий темп до какого-то среднего, оптимального, ритма. Доминирующая группа нервных центров, работающих в общем ритме, тормозит центры с иными ритмами активности. Значение явления усвоения ритма как механизма формирования доминирующего очага и механизма его функционального выделения из общей массы нервных клеток подтверждено в последнее время электрофизиологическими исследованиями на животных и человеке.
Важным свойством доминанты является инерция. Однажды возникшая доминанта может длительное время поддерживаться и после удаления первоначального стимула, например при осуществлении цепных двигательных рефлексов. Инерция выражается также в том, что доминанта может надолго сохраняться как следовое состояние (потенциальная доминанта). При возобновлении прежнего состояния или прежней внешней ситуации доминанта может возникнуть снова. Такое воспроизведение доминанты происходит в организме спортсмена условнорефлекторно в предстартовом состоянии когда в известной мере активизируются все те нервные центры, которые входили в рабочую систему во время предыдущих тренировок. Это проявляется в усилении всего комплекса функций, связанных с мышечной работой: центральных, мышечных, выделительных, сосудистых и др. Мысленное выполнение физических упражнений также воспроизводит (актуализирует) доминирующую систему центров, что обеспечивает тренирующий эффект представления движений и является основой так называемой идеомоторной тренировки.
В норме в нервной системе редко отсутствуют какие-либо доминанты. Бездоминантное состояние—это очень слабое возбуждение, разлитое более или менее равномерно по различным нервным центрам. Сходное состояние возникает у спортсменов в процессе полного расслабления, при аутогенной тренировке. Путем такого расслабления добиваются устранения мощных рабочих доминант и восстановления работоспособности нервных центров.
Как фактор поведения доминанта связана с высшей нервной деятельностью, с психологией человека. Доминанта является физиологической основой акта внимания. Она определяет характер восприятия раздражения из внешней среды, делая его односторонним, но зато более целеустремленным. При наличии доминанты многие влияния внешней среды остаются вне внимания, но зато более интенсивно улавливаются и анализируются те, которые особенно интересуют человека. Доминанта — мощный фактор отбора биологически и социально наиболее значимых раздражении.
Возникновение в коре больших полушарий доминантных состояний наблюдается в начале образования временных связей. Условный рефлекс образуется, когда доминантный очаг возбуждения начинает отвечать не на любое афферентное раздражение, а лишь на специфическое раздражение, ставшее сигнальным.
Поскольку доминанта связана с определенной реакцией, она определяет одностороннее выражение поведения. Чем выраженное доминанта, тем больше она тормозит другие текущие рефлексы. Таким образом, из многих степеней свободы выбирается одна — при наличии в определенных двигательных центрах доминанты работает интенсивно лишь та часть мускулатуры, которая управляется этими центрами, а остальная выключена из сферы деятельности в результате сопряженного торможения. При этом заторможены также многие вегетативные центры. В начальный момент интенсивной мышечной работы почти полностью могут исчезать условные рефлексы: слюноотделительные, мигательные и др. Это обеспечивает Целесообразность движений и экономичность энерготрат. Мощная Двигательная доминанта при статических усилиях за счет сопряженного торможения приводит к задержке дыхания и угнетению деятельности сердечно-сосудистой системы.
По мере формирования двигательного навыка система доминирующих нервных центров совершенствуется. Из нее исключаются все лишние нервные центры, остаются лишь те, которые необходимы и достаточны для осуществления двигательной задачи.
§ 6. Взаимодействие процессов возбуждения и торможения в центральной нервной системе.
Важнейшей характеристикой деятельности нервных центров является постоянное взаимодействие процессов возбуждения и торможения как между разными центрами, так и в пределах каждого из них.
Взаимосочетанная (реципрокная) иннервация мышц-антагонистов. Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц- разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — процесс торможения. При возбуждении же центров разгибателей, наоборот, тормозятся центры сгибателей. Такие координационные взаимоотношения между моторными центрами спинного мозга были названы взаимосочетанной или реципрокной иннервацией мышц-антагонистов.
Появление и усиление в нервных центрах процесса торможения при одновременном возбуждении других центров получило по аналогии с физическими процессами название индукции (в данном случае это одновременная индукция). В настоящее время выяснены механизмы проявления реципрокной иннервации. Афферентное раздражение (например, болевое раздражение рецепторов кожи) направляется не только по собственному рефлекторному пути к мотонейронам мышц-сгибателей, но и одновременно через коллатерали активирует тормозные клетки Рэншоу. Окончания этих клеток образуют тормозные синапсы на мотонейронах мышц-разгибателей, вызывая в них торможение.
Реципрокные отношения между центрами мышц-антагонистов не являются постоянными и единственно возможными. В необходимых ситуациях (например, при фиксации суставов, при точностных движениях) они сменяются одновременным их возбуждением. В этом проявляется большая гибкость, целесообразность сложившихся в организме координации.
Показано, что у человека во время ходьбы и бега основной формой координации являются реципрокные отношения, но помимо них имеются фазы одновременной активности мышц-антагонистов голеностопного и, особенно коленного и тазобедренного суставов. Длительность фаз одновременной активности увеличивается с повышением скорости перемещения.
Взаимосочетанные (реципрокные) отношения характерны не только для моторных центров спинного мозга, но и для других центров. Еще в 1896 г. Н. Е. Введенский в опытах на животных наблюдал при раздражении двигательной зоны одного полушария коры головного мозга реципрокное торможение моторных центров другого полушария. При этом сокращение мышц одной половины тела сопровождалось расслаблением одноименных мышц другой.
Реципрокные отношения формируются также при возникновении доминанты, когда при возбуждении одних центров с помощью сопряженного торможения выключается деятельность других, посторонних, нервных центров.
Последовательная смена процессов возбуждения и торможения. Взаимоотношения процессов возбуждения и торможения в центральной нервной системе могут проявляться во времени в виде последовательной смены возбуждения и торможения в одних и тех же нервных центрах.
«Возбуждение вслед за торможением» впервые наблюдал И. М. Сеченов. Он описал резкое усиление у лягушки рефлекторной деятельности после ее торможения сильным раздражителем: резкий прыжок с голосовой реакцией и восстановлением кожной чувствительности («рефлекс Сеченова»). Позднее было обнаружено «торможение вслед за возбуждением» (А. А. Ухтомский). После сильных ритмических раздражении лапки лягушки выключение раздражителя приводит к мгновенному расслаблению лапки—поднятая конечность падает, как плеть. Описанные взаимоотношения процессов возбуждения и торможения часто встречаются в коре больших полушарий при условнорефлекторной деятельности.
Контрастное усиление одного процесса после другого в одном и том же нервном центре получило название последовательной индукции. Оно имеет большое значение при организации ритмической двигательной деятельности, обеспечивая попеременное сокращение и расслабление мышц.
Принцип конвергенции. К одной и той же нервной клетке благодаря многочисленным побочным взаимосвязям рефлекторных дуг могут поступать импульсы от различных рецепторов тела, т. е. сигналы о самых разнообразных раздражениях. Схождение импульсов, поступивших по различным афферентным путям, в каком-либо одном центральном нейроне или нервном центре называется конвергенцией.
В низших отделах нервной системы—спинном и продолговатом мозгу—конвергенция выражена гораздо меньше. Нейроны этих отделов получают информацию от рецепторов сравнительно небольших участков тела — рецептивных полей одного и того же рефлекса. В надсегментарных отделах, особенно в коре больших полушарий, происходит конвергенция импульсов различного происхождения от разных рефлекторных путей. Нейроны надсегментарных отделов могут получать сигналы о световых, звуковых, проприоцептивных и прочих раздражениях, т. е. сигналы разной модальности. На теле нейронов постоянно изменяются «конвергентные узоры» — возбужденные и заторможенные участки. Подсчитано, что размеры рецептивных полей корковых нейронов, т. е. участков тела, от которых к ним могут поступать афферентные раздражения, в 16— 100 раз больше, чем размеры тех же полей для афферентных клеток спинальных рефлекторных дуг. Благодаря такому разнообразию поступающей информации в нейронах вышележащих отделов головного мозга может происходить ее широкое взаимодействие, сопоставление, отбор, выработка адекватных реакций и установление новых связей между рефлексами.
Принцип общего конечного пути. Афферентных нейронов в центральной нервной системе в несколько раз больше, чем эфферентных. В связи с этим многие афферентные влияния поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для них общими конечными путями к рабочим органам. Система реагирующих нейронов образует таким образом как бы воронку («воронка Шеррингтона»). Множество разнообразных раздражении может возбудить одни и те же мотонейроны спинного мозга и вызвать одну и ту же двигательную реакцию (например, сокращение мышц-сгибателей верхней конечности). Английский физиолог Ч. Шеррингтон, установивший принцип общего конечного пути, предложил различать союзные (аллиированные) и антагонистические рефлексы. Встречаясь на общих конечных путях, союзные рефлексы взаимно усиливают друг друга, а антагонистические—тормозят. В первом случае в нейронах общего конечного пути имеет место пространственная суммация (например, сгибательный рефлекс усиливается при одновременном раздражении нескольких участков кожи). Во втором случае происходит борьба конкурирующих влияний за обладание общим конечным путем, в результате чего один рефлекс осуществляется, а другие затормаживаются. При этом освоенные движения выполняются с меньшим трудом, так как в их основе лежат упорядоченные во времени синхронизированные потоки импульсов, которые проходят через конечные пути легче, чем импульсы, поступающие в случайном порядке.
Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.
В таком отборе важную роль играет наличие в центральной нервной системе доминанты. Она обеспечивает протекание главной реакции. Например, ритмический шагательный рефлекс и одиночный, непрерывный рефлекс сгибания при болевом раздражении являются антагонистическими. Однако спортсмен, внезапно получивший травму, может продолжать бег к финишу, т. е. осуществлять ритмический рефлекс и подавлять болевые раздражения, которые, поступая к мотонейронам сгибательных мышц, препятствуют попеременному сгибанию и разгибанию ноги.
§ 7. Специфические и неспецифические системы
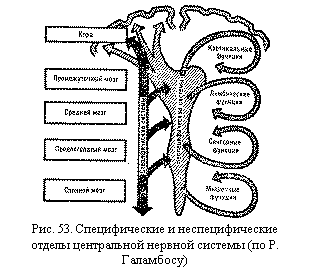 В
нервной системе в настоящее время различают два отдела — специфическую и
неспецифическую системы. Специфическая система расположена в наружных и боковых
частях центральной нервной системы, а неспецифическая занимает срединную ее
часть (рис. 53). Эти системы различаются по строению и функциональным
особенностям.
В
нервной системе в настоящее время различают два отдела — специфическую и
неспецифическую системы. Специфическая система расположена в наружных и боковых
частях центральной нервной системы, а неспецифическая занимает срединную ее
часть (рис. 53). Эти системы различаются по строению и функциональным
особенностям.
К специфической системе относятся все пути и нервные центры. проводящие афферентную импульсацию от различных рецепторов тела и эфферентную импульсацию к рабочим органам. Это пути сенсорных систем и нисходящие пути.
Афферентные пути этой системы проводят сигналы какой-либо специфической чувствительности (например, мышечно-суставной, тактильной, слуховой, зрительной и других) от рецепторов, воспринимающих раздражения определенного качества, к соответствующим нервным центрам, где происходит анализ этих сигналов и возникают ощущения и восприятия.
Эфферентные импульсы от нервных центров к исполнительным органам тела — мышцам и железам—направляются по специфическим нисходящим путям, образующим проводящие пути, или тракты (например, пирамидный тракт), и служат для управления определенными функциями на периферии (например, движениями скелетных мышц, изменением просвета сосудов и т. д.). Это эфферентные отделы, обеспечивающие различные рефлекторные реакции, описание которых дано в соответствующих разделах учебника. Следовательно, деятельность специфической системы связана с анализом различных раздражителей и определенным характером ответных реакций организма. В этой деятельности активное участие принимает также неспецифическая система организма, регулирующая и изменяющая как восприятие организмом специфических раздражении, так и его эфферентную деятельность.
Неспецифическая система не связана с анализом какой-либо специфической чувствительности или с выполнением определенных рефлекторных реакций. Импульсация в эту систему поступает через боковые ответвления от специфических путей, передающих проприоцептивные, слуховые и другие специфические сигналы. К одному и тому же нейрону могут приходить импульсы различного происхождения и от разных рецепторов тела. В результате такого схождения (конвергенции) импульсов создаются широкие возможности для их взаимодействия. Вследствие этого неспецифическая система играет большую роль а процессах интеграции функций в организме.
Характерной особенностью нейронов неспецифической системы являются также обилие и разнообразие их отростков. Они обеспечивают широкую циркуляцию импульсов в неспецифической системе. Благодаря этому здесь возможно взаимодействие одной клетки почти с 30000 других нейронов. По характеру расположения нейронов и обилию их связей неспецифические отделы ствола головного мозга получили название сетевидного образования (или ретикулярной формации).
Функции ретикулярной формации. Различают два основных типа влияния неспецифической системы на работу других нервных центров — активирующее и тормозящее влияния. Оба они могут адресоваться как вышележащим центрам (восходящие влияния), так и нижележащим (нисходящие влияния).
Восходящие влияния. В опытах на животных было показано, что из сетевидного образования среднего мозга исходит мощное активирующее влияние на кору больших полушарий. Электрические раздражения этих отделов неспецифической системы через вживленные электроды вызывали пробуждение спящего животного. У бодрствующего животного подобное раздражение повышало уровень корковой активности, усиливало внимание к внешним сигналам и улучшало их восприятие. Эти опыты свидетельствуют о большом значении импульсов среднемозговых отделов неспецифической системы в повышении функционального состояния нейронов коры больших полушарий. Восходящие влияния из этих областей охватывают диффузно всю кору, вызывая общее изменение ее функционального состояния. Они особенно усиливаются при действии новых раздражений, заранее подготавливая организм к реакции на неожиданную ситуацию. Это всегда имеет место при ориентировочных реакциях и в начальных стадиях выработки условного рефлекса. В результате активирующих воздействий во всех областях коры больших полушарий повышается возбудимость и лабильность корковых нейронов, быстрее возникают ответы на внешние раздражения и легче происходит вовлечение новых нервных клеток в протекающую реакцию. Увеличение числа активированных нейронов позволяет в процессе обучения, тренировки «выбрать» из них клетки, наиболее важные для осуществления ответных действий организма.
Активирующие влияния, исходящие из промежуточного мозга (от неспецифических ядер таламуса), в отличие от среднемозговых, воздействуют лишь на ограниченные области коры. Считают, что такое избирательное повышение активности небольшой территории коры имеет значение в организации внимания, выделяя на общем фоне работу небольшого числа корковых клеток. Обычно активируются нейроны именно тех отделов коры, которые заняты в данный момент наиболее важной для организма деятельностью. Это в значительной мере способствует протеканию специфических функций мозга — восприятию и переработке наиболее нужных сигналов, осуществлению первоочередных двигательных актов.
Неспецифическая система способна оказывать на кору помимо активирующих и тормозящие влияния. Эти влияния различны по характеру: локальные—из области промежуточного мозга и общие (диффузные) — из задних отделов мозга. Первые участвуют в создании «рабочей мозаики» в коре, разделяя ее на более активные ч менее активные участки. Вторые обусловливают диффузное угнетение корковых функций. Широко распространенное по всей коре торможение деятельности корковых нейронов возникает с участием заднестволовых отделов ретикулярной формации — неспецифических нейронов задних частей среднего и продолговатого мозга. Оно имеет место в случае длительной и монотонной работы (например, производственных условиях при работе на конвейере или в спорте при прохождении длинных и сверхдлинных дистанций).
В электрических проявлениях деятельности мозга активирующие влияния проявляются в виде возникновения частой асинхронной активности (десинхронизация), а тормозящие влияния — в виде медленных ритмичных колебаний (синхронизация).
Нисходящие влияния. Все отделы неспецифической системы оказывают помимо восходящих значительные нисходящие влияния. Отделы ствола мозга регулируют (активируют или угнетают) активность нейронов спинного мозга и проприорецепторов мышц (мышечных веретен). Эти влияния совместно с воздействиями из экстрапирамидной системы и мозжечка играют большую роль в регуляции тонуса мышц и обеспечении позы человека. Непосредственные команды к осуществлению движений и влияния, формирующие перестройки тонуса мышц, передаются по специфическим путям. Однако неспецифические влияния позволяют существенно изменить протекание этих реакций. При усилении активирующих воздействий из ретикулярной формации среднего мозга на нейроны спинного мозга увеличивается амплитуда производимых движений и повышается тонус скелетных мышц. Включение этих влияний при некоторых эмоциональных состояниях помогает повысить эффективность двигательной деятельности человека и выполнить значительно большую работу, чем в обычных условиях.
Таким образом, в формировании целостных актов организма участвуют обе системы головного мозга—специфическая и неспецифическая. Проведение и обработку специфической информации, а также управление ответными реакциями осуществляет специфическая система. На эти процессы значительно влияет неспецифическая система. В свою очередь, деятельность этого неспецифического отдела контролируется специфическими системами мозга. В объединении функций обеих систем и общем контроле за их активностью ведущая роль принадлежит коре больших полушарий.
§ 8. Основные рефлекторные центры нервной системы
Спинной мозг. Спинной мозг является низшим и наиболее древним отделом центральной нервной системы. Он имеет значительно меньшую самостоятельность у человека по сравнению с животными. У человека его вес по отношению к головному мозгу составляет всего 2% (у кошек—25%, у кролика—45%, у черепах— 120%).
Надежность сегментарных функций спинного мозга обеспечена множественностью его связей с периферией: каждый сегмент спинного мозга иннервирует 3 метамера (участка) тела — собственный, половину вышележащего и половину нижележащего, а каждый метамер тела получает иннервацию от 3 сегментов спинного мозга. Такое устройство гарантирует осуществление функций спинного мозга при возможных его перерывах и других поражениях.
Распределение функций входящих и выходящих волокон спинного мозга подчиняется определенному закону: все чувствительные (афферентные) волокна входят в спинной мозг через его задние корешки, а двигательные и вегетативные (эфферентные) выходят через передние корешки. В задних корешках волокон гораздо больше, чем в передних (их соотношение у человека примерно 5:1) т. е. при большом разнообразии поступающей информации организм использует незначительное количество исполнительных приборов Основную часть волокон в спинномозговых корешках составляют мякотные волокна. По задним корешкам в спинной мозг поступают импульсы от рецепторов скелетных мышц, сухожилий, кожи, сосудов, внутренних органов. Передние корешки содержат волокна к скелетным мышцам и вегетативным ганглиям.
Задние корешки образованы волокнами одного из отростков афферентных нейронов, тела которых расположены вне центральной нервной системы — в межпозвоночных ганглиях, а волокна другого отростка связаны с рецептором. Общее число афферентных волокон у человека достигает примерно 1 млн. Они различаются по диаметру. Наиболее толстые идут от рецепторов мышц и сухожилий, средние по толщине—от тактильных рецепторов кожи, от части мышечных рецепторов и от рецепторов внутренних органов (мочевого пузыря, желудка, кишечника и др.), наиболее тонкие миелинизированные и немиелинизированные волокна—от болевых рецепторов и терморецепторов. Одна часть афферентных волокон заканчивается на нейронах спинного мозга, другая часть направляется к нейронам продолговатого мозга, образуя спинно-бульбарный путь.
Передние корешки состоят из отростков мотонейронов передних рогов спинного мозга и нейронов боковых рогов. Волокна первых направляются к скелетной мускулатуре, а волокна вторых переключаются в вегетативных ганглиях на другие нейроны и иннервируют внутренние органы.
В составе серого вещества спинного мозга человека насчитывают около 13,5 млн. нервных клеток. Из них двигательные клетки — мотонейроны — составляют всего 3%, а 97% представляют промежуточные клетки (вставочные, или интернейроны). Следует иметь в виду, что по функциональному механизму эти виды нейронов не различаются. Среди мотонейронов спинного мозга различают крупные клетки — альфа-мотонейроны и мелкие клетки — гамма-мотонейроны. От альфа-мотонейронов отходят наиболее толстые и быстропроводящие волокна двигательных нервов, вызывающие сокращение скелетных мышечных волокон. Тонкие волокна гамма-мотонейронов не вызывают сокращения мышц. Они подходят к проприорецепторам — мышечным веретенам и вызывают сокращение их внутренних (интрафузальных) мышечных волокон. При этом сокращении растягиваются рецепторы веретен, повышается их чувствительность, усиливается поток афферентных импульсов от скелетных мышц к нервным центрам. Таким образом, альфа-мотонейроны вызывают двигательные акты, а гамма-мотонейроны регулируют чувствительность мышечных рецепторов, информирующих мозг о выполнении этих движений.
Группу альфа-мотонейронов, иннервирующих отдельную скелетную мышцу, называют ее моторным ядром. Ядра крупных скелетных мышц состоят из мотонейронов, расположенных в 2—3 сегментах спинного мозга. Отростки этих клеток выходят из спинного мозга в составе 2—3 передних корешков. Мелкие же мышцы иннервируются мотонейронами одного сегмента, волокна которого идут в составе одного переднего корешка.
Особое место в деятельности спинного мозга занимают его промежуточные нейроны, или интернейроны. Это в основном мелкие клетки, через которые осуществляются межнейронные взаимодействия в спинном мозгу и координация деятельности мотонейронов. К промежуточным нейронам относятся и тормозные клетки Рэншоу, с помощью которых осуществляются возвратное торможение альфа-мотонейронов и реципрокное торможение центров мышц-антагонистов.
Огромное значение в сложных процессах координации имеют межнейронные взаимодействия на уровне спинного мозга. Это может быть продемонстрировано следующими данными: из огромного количества межнейронных синапсов лишь 10% образовано волокнами, приходящими из головного мозга, и всего около 1 % — афферентными волокнами, т. е. почти 90% остальных синаптических контактов на спинальных клетках образовано волокнами, которые начинаются и кончаются в самом спинном мозгу. Это указывает на существенную роль собственной интегративной деятельности спинного мозга. Благодаря такому множеству существующих связей имеются широкие возможности комбинаций различных нервных клеток для организации любой целесообразной ответной реакции организма.
У человека процессы координации на уровне спинного мозга в значительно большей мере подчинены регулирующим влияниям головного мозга, чем у животных. Нарушение связей спинного мозга с головным приводит к выраженному расстройству протекания спинно-мозговых рефлексов (спинальный шок). На вставочных и моторных нейронах импульсы, приходящие в спинной мозг из головного, взаимодействуют с сегментарными афферентными влияниями. Приказы вышележащих этажей нервной системы увязываются таким образом с текущим состоянием двигательного аппарата.
Рефлексы спинного мозга можно подразделить на двигательные, осуществляемые альфа-мотонейронами передних рогов, и вегетативные, осуществляемые эфферентными клетками боковых рогов. Мотонейроны спинного мозга иннервируют все скелетные мышцы (за исключением мышц лица). Спинной мозг осуществляет элементарные двигательные рефлексы — сгибательные и разгибательные, возникающие при раздражении рецепторов кожи или проприорецепторов мышц и сухожилий, а также посылает постоянную импульсацию к мышцам, поддерживая их напряжение — мышечный тонус.
Мышечный тонус возникает в результате раздражения проприорецепторов мышц и сухожилий при их растяжении во время движения человека или при воздействии силы тяжести. Импульсы от проприорецепторов поступают к мотонейронам спинного мозга, а импульсы от мотонейронов направляются к мышцам, обеспечивая поддержание их тонуса. При разрушении нервных центров спинного мозга или при перерезке нервных волокон, идущих от мотонейронов к мышцам, исчезает тонус скелетных мышц. Участие спинного мозга в двигательной деятельности проявляется не только в поддержании тонуса, но и в организации элементарных двигательных актов и сложной координации деятельности различных мышц (например, согласованной деятельности мышц-антагонистов). Это возможно благодаря мощному развитию системы вставочных нейронов и их богатым взаимосвязям внутри спинного мозга.
Специальные мотонейроны иннервируют дыхательную мускулатуру — межреберные мышцы и диафрагму и обеспечивают дыхательные движения. Вегетативные нейроны иннервируют все внутренние органы (сердце, сосуды, железы внутренней секреции, пищеварительный тракт и др.) и осуществляют рефлексы, регулирующие их деятельность.
Проводниковая функция спинного мозга связана с передачей в вышележащие отделы нервной системы получаемого с периферии потока информации и с проведением импульсов, идущих из головного мозга в спинной. Наиболее важными восходящими путями спинного мозга являются: 1) путь в продолговатый мозг—спинно-бульбарный; 2) в мозжечок—спинно-мозжечковый, несущие импульсы ог проприорецепторов мышц, суставов и сухожилий, частично от рецепторов кожи; 3) в промежуточный мозг—спинно-таламический путь (от тактильных, болевых и терморецепторов). По различным восходящим путям передаются в головной мозг сигналы от интерорецепторов внутренних органов
Продолговатый мозг и варолиев мост. Продолговатый мозг и варолиев мост относят к заднему мозгу. Он является частью ствола мозга. Задний мозг осуществляет сложную рефлекторную деятельность и служит для соединения спинного мозга с вышележащими отделами головного мозга. В срединной его области расположены задние отделы ретикулярной формации, оказывающие неспецифические тормозные влияния на спинной и головной мозг.
Через продолговатый мозг проходят восходящие пути от рецепторов слуховой и вестибулярной чувствительности. Функции нейронов вестибулярных ядер продолговатого мозга разнообразны. Одна часть их реагирует на перемещение тела (например, при горизонтальных ускорениях в одну сторону они увеличивают частоту разрядов, а при ускорениях в другую сторону уменьшают их). Другая часть предназначена для связи с моторными системами. Эти вестибулярные нейроны, повышая возбудимость мотонейронов спинного мозга и нейронов двигательной зоны коры больших полушарий, позволяют регулировать двигательные акты в соответствии с вестибулярными влияниями.
В продолговатом мозгу оканчиваются афферентные нервы, несущие информацию от рецепторов кожи и мышечных рецепторов. Здесь они переключаются на другие нейроны, образуя путь в таламус и далее в кору больших полушарий. Восходящие пути кожно-мышечной чувствительности (как и большая часть нисходящих кортико-спинальных волокон) перекрещиваются на уровне продолговатого мозга.
В продолговатом мозгу и варолиевом мосту находится большая группа черепно-мозговых ядер (от V до XII пары), иннервирующих кожу, слизистые оболочки, мускулатуру головы и ряд внутренних органов (сердце, легкие, печень). Совершенство этих рефлексов обусловлено наличием большого количества нейронов, образующих ядра и соответственно большого числа нервных волокон. Так, только в одном нисходящем корешке тройничного нерва, проводящем болевую, температурную и тактильную чувствительность от головы, содержится во много раз больше волокон, чем в спинно-таламическом пути, содержащем волокна, идущие от болевых и температурных рецепторов остальной части тела.
На дне IV желудочка в продолговатом мозгу находится жизненно важный дыхательный центр, состоящий из центров вдоха и выдоха. Его составляют мелкие клетки, посылающие импульсы к дыхательным мышцам через мотонейроны спинного мозга. В непосредственной близости расположен сердечно-сосудистый центр. Его крупные клетки регулируют деятельность сердца и состояние сосудов. Функции этих центров взаимосвязаны. Ритмические разряды дыхательного центра изменяют частоту сердечных сокращений, вызывая дыхательную аритмию — учащение сердцебиений на вдохе и замедление их на выдохе.
В продолговатом мозгу находится ряд рефлекторных центров, связанных с процессами пищеварения. Это группа центров моторных рефлексов (жевания, глотания, движений желудка и части кишечника), а также секреторных (слюноотделение, выделение пищеварительных соков желудка, поджелудочной железы и др.). Кроме того, здесь находятся центры некоторых защитных рефлексов: чихания, кашля, мигания, слезоотделения, рвоты.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц (см. ниже). Влияния, исходящие из вестибулярных ядер продолговатого мозга, усиливают тонус мышц-разгибателей, что важно для организации позы.
Неспецифические отделы продолговатого мозга, наоборот, оказывают угнетающее влияние на тонус скелетных мышц, снижая его и в мышцах-разгибателях. Продолговатый мозг участвует в осуществлении рефлексов поддержания и восстановления позы тела, так называемых установочных рефлексов (см. ниже).
Средний мозг. Через средний мозг, являющийся продолжением ствола мозга, проходят восходящие пути от спинного и продолговатого мозга к таламусу, коре больших полушарий и мозжечку.
В состав среднего мозга входят четверохолмия, черная субстанция и красные ядра. Срединную его часть занимает ретикулярная формация (см. § 6 этой главы), нейроны которой оказывают мощное активирующее влияние на всю кору больших полушарий, а также на спинной мозг.
Передние бугры четверохолмия представляют собой первичные зрительные центры, а задние бугры—первичные слуховые центры. Ими осуществляют также ряд реакций, являющихся компонентами ориентировочного рефлекса при появлении неожиданных раздражителей. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных—настораживания ушей. Этот рефлекс (по И. П. Павлову, рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции на любое новое воздействие. Он сопровождается усилением тонуса мыщц-сгибателей (подготовка к двигательной реакции) и изменениями вегетативных функций (дыхание, сердцебиения).
Средний мозг играет важную роль в регуляции движений глаз. Управление глазодвигательным аппаратом осуществляют расположенные в среднем мозгу ядра блокового (IV) нерва, иннервирующего верхнюю косую мышцу глаза, и глазодвигательного (III) нерва иннервирующего верхнюю, нижнюю и внутреннюю прямые мышцы нижнюю косую мышцу и мышцу, поднимающую веко, а также расположенное в заднем мозгу ядро отводящего (VI) нерва, иннервирующего наружную прямую мышцу глаза. С участием этих ядер осуществляются поворот глаза в любом направлении, аккомодация глаза, фиксация взгляда на близких предметах путем сведения зрительных осей, зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету).
У человека при ориентации во внешней среде ведущим является зрительный анализатор, поэтому особое развитие получили передние бугры четверохолмия (зрительные подкорковые центры). У животных с преобладанием слуховой ориентации (собака, летучая мышь), наоборот, в большей степени развиты задние бугры (слуховые подкорковые центры).
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук).
В среднем мозгу важные функции осуществляет красное ядро. О возрастании роли этого ядра в процессе эволюции свидетельствует резкое увеличение его размеров по отношению к остальному объему среднего мозга. Красное ядро тесно связано с корой больших полушарий, ретикулярной формацией ствола, мозжечком и спинным мозгом.
От красного ядра начинается руброспинальный путь к мотонейронам спинного мозга. С его помощью осуществляется регуляция тонуса скелетных мышц, происходит усиление тонуса мышц-сгибателей. Это имеет большое значение как при поддержании позы в состоянии покоя, так и при осуществлении движений. Импульсы, приходящие в средний мозг от рецепторов сетчатки глаза и от проприорецепторов глазодвигательного аппарата, участвуют в осуществлении глазодвигательных реакций, необходимых для ориентации в пространстве, выполнении точностных движений.
Промежуточный мозг. В состав промежуточного мозга, который является передним концом ствола мозга, входят зрительные бугры — таламус и подбугровая область — гипоталамус.
Таламус представляет собой важнейшую «станцию» на пути афферентных импульсов в кору больших полушарий.
Ядра таламуса подразделяют на специфические и неспецифические.
К специфическим относят переключательные (релейные) ядра и ассоциативные. Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Это так называемые специфические восходящие пути. Они характеризуются соматотопической организацией. Особенно большое представительство таламусе имеют эфферентные влияния, поступающие от рецепторов лица и пальцев рук. От таламических нейронов начинается путь к соответствующим воспринимающим областям коры — слуховым, зрительных и др. Ассоциативные ядра непосредственно не связаны с периферией. Они получают импульсы от переключающих ядер и обеспечивают их взаимодействие на уровне таламуса, т. е. осуществляют подкорковую интеграцию специфических влияний. Импульсы от ассоциативных ядер таламуса поступают в ассоциативные области коры больших полушарий, где участвуют в процессах высшего афферентного синтеза.
Помимо этих ядер, в таламусе имеются неспецифические ядра, которые могут оказывать как активирующее, так и тормозящее влияние на кору (см. § 6 этой главы).
Благодаря обширным связям таламус играет важнейшую роль в жизнедеятельности организма. Импульсы, идущие от таламуса в кору, изменяют состояние корковых нейронов и регулируют ритм корковой активности. Между корой и таламусом существуют кольцевые кортико-таламические взаимосвязи, лежащие в основе образования условных рефлексов. С непосредственным участием таламуса происходит формирование эмоций человека. Таламусу принадлежит большая роль в возникновении ощущений, в частности ощущения боли.
Подбугровая область расположена под зрительными буграми и имеет тесные нервные и сосудистые связи с прилежащей железой внутренней секреции—гипофизом. Здесь расположены важные вегетативные нервные центры, регулирующие обмен веществ в организме, обеспечивающие поддержание постоянства температуры тела (у теплокровных) и другие вегетативные функции.
Участвуя в выработке условных рефлексов и регулируя вегетативные реакции организма, промежуточный мозг играет очень важную роль в двигательной деятельности, особенно при формировании новых двигательных актов и выработке двигательных навыков.
Подкорковые узлы. Подкорковыми узлами называют группу ядер серого вещества, расположенных непосредственно под полушариями большого мозга. К ним относятся парные образования: хвостатое тело и скорлупа, составляющие вместе полосатое тело (стриатум), и бледное ядро (паллидум). Подкорковые ядра получают сигналы от рецепторов тела через зрительные бугры. Эфферентные импульсы подкорковых ядер направляются к нижележащим центрам экстрапирамидной системы. Подкорковые узлы функционируют в единстве с корой больших полушарий, промежуточным мозгом и другими отделами мозга. Это обусловлено наличием кольцевых связей между ними. Через подкорковые ядра могут соединяться между собою разные отделы коры больших полушарий, что имеет большое значение при образовании условных рефлексов. Совместно с промежуточным мозгом подкорковые ядра участвуют в осуществлении сложных безусловных рефлексов: оборонительных, пищевых и др.
Представляя собой высший отдел мозгового ствола, подкорковые узлы объединяют деятельность нижележащих образований, регулируя мышечный тонус и обеспечивая необходимое положение тела во время физической работы. Бледное ядро выполняет моторную функцию. Оно обеспечивает проявление древних автоматизмов — ритмических рефлексов. С его деятельностью связано также выполнение содружественных (например, движения туловища и рук при ходьбе), мимических и других движений.
Полосатое тело оказывает на двигательную деятельность тормозящее, регулирующее влияние, угнетая функции бледного ядра, а также моторкой области коры больших полушарий. При заболевании полосатого тела возникают непроизвольные беспорядочные сокращения мышц (гиперкинезы). Они обусловливают некоординированные толчкообразные движения головы, рук и ног. Нарушения возникают также в чувствительной сфере — понижается болевая чувствительность, расстраиваются внимание и восприятие.
В настоящее время выявлено значение хвостатого тела в самооценке поведения человека. При неправильных движениях или умственных операциях из хвостатого ядра в кору больших полушарий поступают импульсы, сигнализирующие об ошибке.
Мозжечок. Это — надсегментарное образование, не имеющее непосредственной связи с исполнительными аппаратами. Мозжечок входит в состав экстрапирамидной системы. Он состоит из двух полушарий и червя, находящегося между ними. Наружные поверхности полушарий покрыты серым веществом — корой мозжечка, а скопления серого вещества в белом веществе образуют ядра мозжечка.
Мозжечок получает импульсы от рецепторов кожи, мышц и сухожилий через спинно-мозжечковые пути и через ядра продолговатого мозга (от спинно-бульбарного пути). Из продолговатого мозга в мозжечок поступают также вестибулярные влияния, а из среднего мозга—зрительные и слуховые. Корково-мосто-мозжечковый путь связывает мозжечок с корой больших полушарий. В коре мозжечка представительство различных периферических рецепторов имеет соматотопическую организацию. Кроме того, наблюдается упорядоченность связей этих зон с соответствующими воспринимающими областями коры. Так, зрительная зона мозжечка связана со зрительной зоной коры, представительство каждой группы мышц в мозжечке — с представительством одноименных мышц в коре и т. д. Такое соответствие облегчает совместную деятельность мозжечка и коры в управлении различными функциями организма.
Эфферентные импульсы от мозжечка поступают к красным ядрам ретикулярной формации, продолговатому мозгу, таламусу, коре и подкорковым ядрам.
Мозжечок участвует в регуляции двигательной деятельности. Электрические раздражения поверхности мозжечка вызывают движения глаз, головы и конечностей, которые отличаются от корковых моторных эффектов тоническим характером и большой длительностью. Мозжечок регулирует изменение и перераспределение тонуса скелетных мышц, что необходимо для организации нормальной позы и двигательных актов.
Функции мозжечка изучались в клинике при его поражениях у человека, а также у животных путем удаления (экстирпации мозжечка) (Л. Лючиани, Л. А. Орбели). В результате выпадения функций мозжечка возникают двигательные расстройства: атония— резкое падение и неправильное распределение тонуса мышц, астазия — невозможность сохранения неподвижного положения, непрерывные качательные движения, дрожание головы, туловища и конечностей, астения — повышенная утомляемость мышц, атаксия — нарушение координированных движений, походки и др.
Мозжечок оказывает влияние также на ряд вегетативных функций, например желудочно-кишечного тракта, на уровень кровяного давления, на состав крови.
Таким образом, в мозжечке происходит интеграция самых различных сенсорных влияний, в первую очередь проприоцептивных и вестибулярных. Мозжечок даже ранее считали центром равновесия и регуляции мышечного тонуса. Однако его функции, как оказалось, гораздо обширнее—охватывают также регуляцию деятельности вегетативных органов. Деятельность мозжечка протекает в непосредственной связи с корой больших полушарий, под ее контролем.
§ 8. Кора больших полушарий головного мозга
Общий план организации коры. Кора больших полушарий является высшим отделом центральной нервной системы, который в процессе филогенетического развития появляется позже всего и формируется в ходе индивидуального (онтогенетического) развития позже других отделов мозга. Кора представляет собой слой серого вещества толщиной 2—3 мм, содержащий в среднем около 14 млрд. (от 10 до 18 млрд.) нервных клеток, нервные волокна и межуточную ткань (нейроглию). На поперечном ее срезе по расположению нейронов и их связей различают 6 горизонтальных слоев. Благодаря многочисленным извилинам и бороздам площадь поверхности коры достигает 0,2 м2. Непосредственно под корой находится белое вещество, состоящее из нервных волокон, которые передают возбуждение в кору и из нее, а также от одних участков коры другим.
Корковые нейроны и их связи. Несмотря на огромное число нейронов в коре, известно очень немного их разновидностей. Основными типами их являются пирамидные и звездчатые нейроны. Которые не различаются по функциональному механизму.
В афферентной функции коры и в процессах переключения возбуждения на соседние нейроны основная роль принадлежит звездчатым нейронам. Они составляют у человека более половины всех клеток коры. Эти клетки имеют короткие ветвящиеся аксоны, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендриты. Звездчатые нейроны участвуют в процессах восприятия раздражении и объединении деятельности различных пирамидных нейронов.
Пирамидные нейроны осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца — находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид — их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит, через который в клетку поступают различные афферентные влияния от других нейронов, а вертикально вниз отходит эфферентный отросток — аксон.
Многочисленность контактов (например, только на дендритах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейронов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма.
Для коры больших полушарий характерно обилие межнейронных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними нейронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферентное раздражение (от одного и того же рецептора) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов.
Распространение возбуждения в поперечном направлении—от одной вертикальной колонки к другой — ограничено процессами торможения. Возникновение активности в вертикальной колонке приводит к возбуждению спинальных мотонейронов и сокращению связанных с ними мышц. Этот путь используется, в частности, при произвольном управлении движениями конечностей.
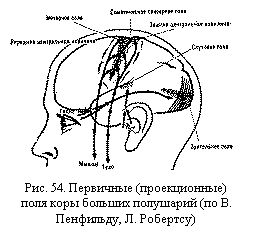 Первичные,
вторичные и третичные поля коры. Особенности строения и функционального
значения отдельных участков коры позволяют выделить отдельные корковые поля.
Первичные,
вторичные и третичные поля коры. Особенности строения и функционального
значения отдельных участков коры позволяют выделить отдельные корковые поля.
Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенезе, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову (например, поле болевой, температурной, тактильной и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и двигательное поле в передней центральной извилине коры) (рис. 54). Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов. При разрушении первичных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Первичные и вторичные поля имеются и у человека, и у животных.
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейроны. Третичные поля находятся в задней половине коры — на границах теменных, височных и затылочных ее областей и в передней половине — в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражений и с учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысленные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
Воспринимая и оценивая все сигналы из внутренней и внешней среды, кора больших полушарий осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций.
Функции коры больших полушарии. Кора больших полушарий выполняет наиболее сложные функции организации приспособительного поведения организма во внешней среде. Это прежде всего функция высшего анализа и синтеза всех афферентных раздражении.
 Афферентные
сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов
(первичные поля), а затем синтезируются во вторичных и третичных полях,
благодаря деятельности которых создается целостное восприятие внешнего мира.
Этот синтез лежит в основе сложных психических процессов восприятия,
представления, мышления. Кора больших полушарий представляет собою орган, тесно
связанный с возникновением у человека сознания и регуляцией его общественного
поведения. Важной стороной деятельности коры больших полушарий является замыкательная
функция — образование новых рефлексов и их систем (условные рефлексы,
динамические стереотипы—см. главу XV).
Афферентные
сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов
(первичные поля), а затем синтезируются во вторичных и третичных полях,
благодаря деятельности которых создается целостное восприятие внешнего мира.
Этот синтез лежит в основе сложных психических процессов восприятия,
представления, мышления. Кора больших полушарий представляет собою орган, тесно
связанный с возникновением у человека сознания и регуляцией его общественного
поведения. Важной стороной деятельности коры больших полушарий является замыкательная
функция — образование новых рефлексов и их систем (условные рефлексы,
динамические стереотипы—см. главу XV).
Благодаря необычайно большой продолжительности сохранения в коре следов прежних раздражении (памяти) в ней накапливается огромный объем информации. Это имеет большое значение для сохранения индивидуального опыта, который используется по мере необходимости.
Электрическая активность коры больших полушарии. Изменения функционального состояния коры отражаются на характере ее биопотенциалов. Регистрация электроэнцефалограммы (ЭЭГ), т. е. электрической активности коры, производится непосредственно с ее обнаженной поверхности (в опытах на животных и при операциях на человеке) или через неповрежденные покровы головы (в естественных условиях на животных и человеке). И, т.о. регистрируют суммарную активность всех ближайших нейронов. Современные электро-энцефалографы усиливают эти потенциалы в 2—3 млн. раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно.
В ЭЭГ различают определенные диапазоны частот, называемые ритмами ЭЭГ (рис. 55). В состоянии относительного покоя чаще всего регистрируется альфа-ритм (8—12 колебаний в 1 сек.), в состоянии активного внимания — бета-ритм (выше 13 колебаний в 1 сек.), при засыпании, некоторых эмоциональных состояниях — тэта-ритм (4—7 колебаний в 1 сек.), при глубоком сне, потере сознания, наркозе — дельта-ритм (1—3 колебания в 1 сек.).
В ЭЭГ отражаются особенности взаимодействия корковых нейронов при умственной и физической работе. Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит к так называемой десинхронизации ЭЭГ — быстрой асинхронной активности (см. рис. 55). По мере формирования двигательного навыка происходит сонастраивание активности отдельных связанных с данным движением нейронов и отключение посторонних.
В ЭЭГ при этом возникают различные формы синхронизации (см. рис. 55, ж, з). Выполнение освоенного и автоматизированного движения может протекать при незначительной активности очень небольшого числа корковых нейронов, находящихся в ограниченных областях коры. При этом почти на всей остальной поверхности коры восстанавливается исходный ритм колебаний — альфа-ритм (см. рис. 55, з).
В процессе спортивной тренировки происходит перестройка и совершенствование функций коры больших полушарий. С ростом спортивного мастерства увеличиваются амплитуда и регулярность проявления фоновой активности — альфа-ритма в состоянии покоя. При развитии качества быстроты (например, у баскетболистов) повышается частота волн альфа-ритма, что способствует ускорению произвольных движений.
В процессе мышечной работы значительно усиливается по сравнению с состоянием относительного покоя взаимосвязанность (синхронность и синфазность) электрической активности различных областей коры. Это облегчает функциональные взаимодействия между различными корковыми центрами. Процесс формирования двигательного навыка сопровождается концентрацией взаимосвязанной активности в ограниченных зонах коры, наиболее важных для текущей деятельности. Между этими зонами устанавливается общий ритм активности. В такие характерные системы взаимодействующих корковых зон включаются не только первичные поля (моторные, зрительные и др.), но и вторичные (например, премоторные и др.) и особенно третичные поля: передние — программирующие лобные области и задние — зоны афферентного синтеза (нижнетеменные и др.).
§ 9. Нисходящие влияния головного мозга на двигательную деятельность
Основные нисходящие системы головного мозга. Высшие отделы головного мозга осуществляют свои влияния на деятельность нижележащих отделов, в том числе спинного мозга, через нисходящие пути.
У млекопитающих животных и человека контроль за рефлекторной деятельностью спинного мозга осуществляется главным образом тремя нисходящими системами: 1) кортико-спинальной, 2) кортико-рубро-спинальной и 3) кортико-ретикуло-спинальной. Пути этих систем идут в спинной мозг непосредственно от нейронов коры больших полушарий, а также через нейроны красного ядра среднего мозга — рубро-спинальный путь, ядра ретикулярной формации продолговатого и среднего мозга — ретикуло-спинальный путь и вестибулярные ядра продолговатого мозга — вестибуло-спинальный путь. Все эти системы функционируют в тесном взаимодействии и под контролем коры больших полушарий. Окончания волокон образуют, как правило, контакты с дендритами спинальных клеток (аксодендритные синапсы).
Нисходящие пути головного мозга группируют обычно в 2 основные нисходящие системы—пирамидную и экстрапирамидную. Под пирамидной системой, или пирамидным трактом, понимают прямые пути от корковых пирамидных нейронов к нейронам спинного мозга (кортико-спинальный путь) и ядрам черепно-мозговых нервов ствола мозга (кортико-бульбарный путь). Экстрапирамидную систему составляют многоступенчатые связи коры больших полушарий со спинным мозгом. К ней относят подкорковые узлы, люисово тело промежуточного мозга, красные ядра и черную субстанцию среднего мозга, мозжечок, ретикулярную формацию ствола мозга и вестибулярные ядра продолговатого мозга. Важнейшими путями этой системы являются кортико-рубро-спинальная и кортико-ретикуло-спинальная системы. Группировка путей на указанные 2 системы в значительной мере условна, так как не соответствует особенностям их морфологии и функционирования.
В настоящее время предлагают подразделять основные нисходящие пути, исходя из расположения проводящих волокон в белом веществе спинного мозга, на следующие 2 системы: 1) латеральную, волокна которой расположены в боковых (латеральных) частях спинного мозга (сюда относят кортико-спинальную и рубро-спинальную системы), и 2) медиальную, волокна которой идут во внутренней (медиальной) части белого вещества (состоящую из вестибуло-спинальной и ретикуло-спинальной систем).
Волокна латеральной системы оканчиваются преимущественно в задних рогах спинного мозга. Они связаны со специальной группой вставочных нейронов, передающих их влияния на мотонейроны в передние рога спинного мозга. Только у обезьян и человека имеются волокна, непосредственно оканчивающиеся на мотонейронах. Латеральные системы — это самые молодые в эволюционном отношении системы, характерные лишь для млекопитающих. Больше всего они развиты у наиболее высокоорганизованных животных и особенно у человека.
Волокна медиальной системы, направляются к сегментарным вставочным нейронам и отчасти мотонейронам медиальной (внутренней) части передних рогов спинного мозга. Это древние системы, которые характерны для всех классов позвоночных животных.
Кортико-спинальная система. Ей принадлежит важнейшая роль о осуществлении произвольных движений у человека. Начало этой системе дают крупные и мелкие пирамидные клетки коры больших полушарий. У человека около 30% волокон пирамидного пути начинается в передней центральной извилине, примерно 20% — в задней центральной извилине, остальные волокна—в других отделах коры и в подкорковых образованиях головного мозга. Большая часть (80%) волокон пирамидного пути перекрещивается на уровне продолговатого мозга, меньшая часть — на соответствующем сегменте спинного мозга. В результате этого корковые клетки левого полушария управляют преимущественно деятельностью мышц правой половины тела, а корковые клетки правого полушария— левой половины тела. У человека количество перекрещивающихся волокон может варьировать, по современным данным, от 0 до 100%, и многие волокна могут идти от корковых клеток к спинному мозгу без перекреста. Кроме того, в составе пирамидного пути имеются волокна с двойным перекрестом — на уровне межполушарных волокон (мозолистого тела) и в продолговатом мозгу. Эти морфологические особенности создают возможность корковой иннервации двигательного аппарата своей стороны тела. Такое дублирование управления мышцами со стороны левого и правого полушарий позволяет компенсировать двигательные функции при односторонних поражениях мозга, а также обеспечивает взаимозамещение деятельности полушарий в процессе длительной мышечной работы.
Волокна пирамидного тракта различаются по диаметру и скорости проведения. У человека в каждом пирамидном пучке на уровне продолговатого мозга содержится около 1 млн. нервных волокон— около 1/3 мякотных и 2/3 безмякотных. В составе пирамидного тракта находятся неоднородные волокна, диаметр которых колеблется от 1 до 25 мк. Большей частью это тонкие волокна. Лишь около 2— 4% волокон имеют диаметр более 10 мк. Это волокна, характеризующиеся наибольшей скоростью проведения—до 120—140 м/сек. Они начинаются от гигантских пирамидных клеток Беца в передней Центральной извилине коры больших полушарий. Примерно такие скорости были обнаружены у человека во время нейрохирургических операций.
Пирамидная система выполняет 3 основные функции: 1) посылает мотонейронам спинного мозга импульсы-команды к движениям (пусковые влияния); 2) изменяет проведение нервных импульсов во вставочных нейронах (влияния, регулирующие протекание спинномозговых рефлексов) и 3) осуществляет контроль за потоком афферентных сигналов в нервные центры (обеспечение обратных связей от работающих органов). Первая функция—двигательная функция кортико-спинальной системы — заключается в преимущественном возбуждении мотонейронов мышц-сгибателей. При этом импульсы, поступающие в спинной мозг, вызывают двоякий эффект — не только возбуждают мотонейроны мышц-сгибателей, но и одновременно затормаживают мотонейроны мышц-разгибателей что обеспечивает беспрепятственное сгибание в соответствующем суставе.
Влияния пирамидной системы больше выражены по отношению к мышцам верхней части туловища и верхних конечностей, чем к мышцам нижних конечностей. Эта неравномерность воздействий отражена в самом устройстве пирамидного пути: значительно большее количество кортико-спинальных волокон проходит в шейной и грудной областях спинного мозга по сравнению с пояснично-крестцовой областью (табл. 3). Двигательные сигналы адресуются преимущественно мотонейронам дистальных мышц конечностей. Этим обеспечивается более тонкое влияние на отдельные мышцы и даже части мышц верхней конечности, особенно на мышцы фаланг пальцев руки.
Развитие специализированного управления дистальными мышцами верхней конечности со стороны коры больших полушарий связано не только с общим совершенствованием движений человека, но главным образом с особой ролью руки — участием ее в сложнейших двигательных операциях, требующих большой точности и быстроты.
Таблица 3
Распределение волокон пирамидного тракта в различных сегментах спинного мозга у человека (по А.Веиль и А.Лассек, 1929; А.Лассек, Л.Дауд и А.Веиль, 1930)
|
Сегменты спинного мозга |
Среднее (для 10 взрослых людей) количество (%) волокон |
Колич. волокон на 1 г мышцы |
|
3-й шейный |
100 |
95 |
|
4—7-й грудной |
81 |
24 |
|
3-й грудной |
55 |
62 |
|
6-й грудной |
46 |
62 |
|
12-грудной |
35 |
62 |
|
4-й поясничный |
20 |
13 |
В процессе эволюционного развития у обезьян и человека появляются прямые пути (без вставочных нейронов) от пирамидных клеток коры к мотонейронам спинного мозга. Чем больше развиты у них дифференцированные движения пальцев верхней конечности, особенно хватательные, тем большее количество корковых волокон оканчивается непосредственно на спинальных мотонейронах. Так, у макаки их число составляет около 2% всех волокон пирамидного пути, у шимпанзе—5%, а у человека—8%. Наличие в этих путях всего одного синапса (моносинаптические пути) ускоряет проведение корковых импульсов к двигательному аппарату.
Считают, что эти пути передают влияния от мозговых механизмов обеспечивающих принятие быстрого решения при произвольных движениях человека.
В процессе филогенетического развития произошло разделение пирамидной системы на 2 подсистемы с различным функциональным значением — «быструю» и «медленную». К «быстрой» подсистеме относятся крупные клетки моторной области коры, аксоны которых обладают высокой скоростью проведения — от 80 до 25 м/сек (средняя скорость—50 м/сек). К «медленной» подсистеме—средние и мелкие клетки моторной и других областей коры, обладающие меньшей скоростью проведения в аксонах—от 25 до 7 м/сек (средняя скорость 14 м/сек). В функциях данных нейронов имеются существенные различия. «Быстрые» пирамидные нейроны значительно быстрее отвечают на афферентное раздражение (латентный период их активации меньше), процесс возбуждения в них короче (меньше длительность пиков потенциала действия и следовых потенциалов). Импульсы, идущие по быстрым волокнам, подходят к клеткам спинного мозга значительно раньше, чем импульсы, передающиеся по медленным волокнам. В состоянии мышечного покоя крупные пирамидные нейроны с быстропроводящими аксонами имеют слабую импульсную активность, а при возникновении движения частота их разрядов резко повышается. Полагают, что «быстрые» нейроны обеспечивают срочную передачу информации от коры к спинному мозгу и контролируют фазные рефлекторные движения. Их называют нейронами движения.
В отличие от них «медленные» нейроны характеризуются постоянной ритмической импульсацией в состоянии покоя. При движениях их активность изменяется медленно и незначительно. Считают, что эти нейроны обеспечивают регуляцию позы тела — ее организацию и приспособление к условиям внешней среды. Соответственно их называют нейронами положения.
Функции «медленной» и «быстрой» подсистем кортико-спинального пути взаимосвязаны. Усиление активности «медленных» нейронов облегчает появление разрядов «быстрых» нейронов.
Вторая функция пирамидной системы заключается в активации вставочных нейронов спинного мозга, передающих импульсы от чувствительных нейронов к двигательным. Передача возбуждения в многонейронных (полисинаптических) рефлекторных дугах спинного мозга при этом облегчается. Важно, что повышение возбудимости вставочных нейронов наступает под влиянием коры до начала Движения и сохраняется в течение всего периода работы мышц, облегчая ее протекание. Однако корковые влияния носят избирательный характер—проведение улучшается не во всех рефлекторных дугах, а лишь в наиболее нужных для данного момента.
Таким образом, спинной мозг самостоятельно осуществляет присущую ему рефлекторную деятельность со всеми особенностями процессов координации (сгибательные реакции сопровождаются вневременным торможением центров мышц-антагонистов, возникают перекрестные рефлексы и т. п.). Роль коры в этом случае проявляется в регуляции этой деятельности (усилении или ослаблении отдельных рефлексов, выборе и запуске адекватных двигательных реакций).
Третья функция пирамидного тракта связана с выключением излишних афферентных воздействий на нервные центры. Аксоны кортико-спинальной системы образуют окончания на аксонах чувствительных нервных клеток, передающих импульсы от рецепторов на периферии в спинной мозг (аксоаксональные синапсы). В месте их контакта развивается под влиянием корковых импульсов пресинаптическое торможение (сильная деполяризация чувствительных волокон), которое блокирует проведение афферентных импульсов к нервным центрам спинного и головного мозга. Главной особенностью этого торможения является то, что, подавляя все виды афферентных импульсов, оно не распространяется на импульсы, идущие от проприорецепторов мышц. В результате этого в процессе движения особенно усиливается роль сигналов от работающих мышц, анализ которых необходим для совершенного управления двигательными актами. Одновременно ослабевает импульсация от других рецепторов тела, посторонних для выполняемой работы.
Поражения пирамидного тракта у человека (кровоизлияния, травмы и др.) приводят к невозможности выполнять произвольные движения. Однако через некоторое время эти движения могут восстановиться за счет деятельности других нисходящих систем, связывающих кору головного мозга со спинным мозгом и способных дублировать функции пирамидной системы. Частично утраченными остаются лишь тонкие движения пальцев. При разрушении патологическим очагом пирамидного тракта на одной стороне тела возможна также компенсация двигательных функций за счет гипертрофии (увеличения диаметра) пирамидных волокон на другой стороне тела.
Кортико-рубро-спинальная система. Корковые нейроны образуют прямые связи с клетками красного ядра среднего мозга, от которых начинаются волокна рубро-спинальной системы. Окончания этих волокон находятся в латеральных частях серого вещества спинного мозга — в основании его задних рогов. У человека этот путь развит в меньшей степени, чем у других млекопитающих. Он образован преимущественно тонкими волокнами, идущими от мелких клеток красного ядра, которые связаны с мотонейронами спинного мозга лишь через специальную систему вставочных нейронов. Основная часть волокон рубро-спинальной системы заканчивается в шейных отделах спинного мозга. В состав кортико-рубро-спинального пути входят волокна, скорость проведения которых значительно выше, чем в большинстве волокон пирамидного пути, —до 105—124 м/сек (средняя скорость — около 70 м/сек). Эти волокна могут пропускать импульсы с очень высокой частотой —свыше 500 в 1 сек.
Афферентные импульсы поступают к красным ядрам от рецепторов кожи, суставных сумок и болевых рецепторов. Красные ядра являются важным промежуточным этапом между корой и спинным мозгом, где происходит подключение регулирующих влияний от мозжечка, промежуточного мозга и подкорковых ядер. В красном ядре хорошо выражено соматотопическое представительство функций: эфферентные клетки рубро-спинальных путей активируются лишь раздражениями, поступающими от того участка тела, с двигательным аппаратом которого они связаны.
Функции кортико-рубро-спинальной системы подобны функциям кортико-спинальной (пирамидной) системы. Это—преимущественное возбуждение мотонейронов мышц-сгибателей, повышение активности вставочных нейронов и контроль за афферентной импульсацией, идущей по восходящим путям. Однако, в отличие от пирамидной системы, здесь нет подразделения на функционально различные подсистемы.
Деятельность кортико-рубро-спинальной системы, ограничивается лишь тоническим контролем. По рубро-спинальным волокнам в клетки спинного мозга постоянно поступает тоническая импульсация с частотой около 40—60 имп/сек. Во время движения головы или конечностей частота ее возрастает (до 100—200 имп/сек). Таким образом, функционально кортико-руброспинальный путь представляет собой единое целое с «медленной» подсистемой кортико-спинального пути, регулируя уровень активности спинальных клеток и тонус соответствующих скелетных мышц. Эта система, обобщая влияния от коры, мозжечка, промежуточного мозга и подкорковых ядер, играет большую роль в организации позо-тонических реакций организма. Формирование этих реакций важно не только в состоянии покоя, но и при выполнении произвольных движений. Еще до начала двигательного акта должна начаться подготовка к нему, в частности в виде изменения позы тела. В связи с этим наличие быстропроводящих волокон в кортико-рубро-спинальной системе может обеспечить своевременное перераспределение тонуса соответствующих мышц.
Медиальная система (ретикуло-спинальный и вестибуло-спинальный компоненты). Медиальная, более древняя, система служит главным образом для тонических воздействий на нейроны спинного мозга. Она подразделяется на ретикуло-спинальную и вестибуло-спинальную. Обе системы контролируются корой больших полушарий. Нисходящие влияния, передаваемые ими, отчетливо различаются по своему влиянию на мотонейроны мышц сгибателей и разгибателей.
Кортико-ретикулярные волокна идут в составе пирамидного тракта от клеток сенсомоторной и других областей коры к ядрам варолиева моста и отчасти продолговатого мозга. От этих ядер начинаются волокна ретикуло-спинального пути к вставочным и двигательным нейронам спинного мозга (в медиальной части переднего рога). Количество волокон в этой системе значительно меньше, чем в пирамидной (около 3000—4000). Они не перекрещиваются и иннервируют нейроны спинного мозга на своей стороне. Связи этих волоков с мотонейронами большей частью полисинаптические — через множество вставочных клеток. Никакой соматотопической организации этого пути не обнаружено. Основная функция кортико-ретикулярного пути — тоническое повышение возбудимости мотонейронов мышц-сгибателей, что приводит к усилению тонуса этих мышц.
Вестибуло-спинальный компонент медиальной системы передает импульсы от вестибулярных ядер продолговатого мозга (например от ядра Дейтерса) к вставочным и двигательным нейронам переднего рога спинного мозга. Основная функция этого пути — тоническое повышение возбудимости мотонейронов мышц-разгибателей, что обеспечивает усиление тонуса этих мышц.
Таким образом, среди нисходящих систем, осуществляющих функцию контроля активности спинного мозга, три системы обеспечивают повышение возбудимости мотонейронов мышц-сгибателей:
кортико-спинальная, кортико-рубро-спинальная и кортико-ретикуло-спинальная, а вестибуло-спинальная система тормозит эти мотонейроны. Лишь одна часть кортико-спинальной системы обусловливает фазную двигательную деятельность. Остальные же нисходящие системы обеспечивают регуляцию тонуса мышц и позных реакций организма.
Роль спинного и головного мозга. Несмотря на совершенство процессов координации в спинном мозгу, они находятся под постоянным контролем головного мозга, в первую очередь коры больших полушарий.
В организме имеются специальные механизмы, обусловливающие преимущественное воздействие коры больших полушарий на общие конечные пути к мышцам—спинальные мотонейроны. Большая эффективность кортико-спинальных влияний по сравнению с сегментарными афферентными влияниями обеспечивается, во-первых, наличием прямых путей из коры к мотонейронам спинного мозга и, во-вторых, возможностью особенно быстрой их активации корковыми импульсами. Электрофизиологическими исследованиями показано, что ритмические воздействия из двигательной области коры вызывают чрезвычайно резкое нарастание суммарной амплитуды возбуждающих постсинаптических потенциалов спинальных мотонейронов. Амплитуда каждого последующего возбуждающего постсинаптического потенциала увеличивается примерно в 6 раз больше, чем при поступлении к тем же мотонейронам импульсов от проприорецепторов по афферентным путям. Таким образом, достаточно уже 2—3 импульсов, идущих от коры, чтобы деполяризация в мотонейроне достигла порогового уровня, необходимого для возникновения ответного разряда в скелетную мышцу. В результате кора больших полушарий может вызывать двигательные действия быстрее, чем периферические раздражения, и часто даже вопреки им.
§ 10. Нервная регуляция эмоций и вегетативных функций
Значение эмоции и их нервная регуляция. Возникновение эмоций связывают с деятельностью лимбической системы, в которую входят некоторые подкорковые образования и участки коры. Корковые отделы лимбической системы, представляющие ее высший отдел находятся на нижних и внутренних поверхностях больших полушарий (поясная извилина, гиппокамп и др.). К подкорковым структурам лимбической системы относят гипоталамус, некоторые ядра таламуса, среднего мозга и ретикулярной формации. Между всеми этими образованиями имеются тесные прямые и обратные связи образующие «лимбическое кольцо».
Лимбическая система участвует в самых разнообразных проявлениях деятельности организма. Она формирует положительные и отрицательные эмоции со всеми двигательными, вегетативными и эндокринными их компонентами (изменением дыхания, сердцебиения кровяного давления, деятельности желез внутренней секреции, скелетных и мимических мышц и др.). От нее зависит эмоциональная окраска психических процессов и изменения двигательной активности. Она создает мотивацию поведения (определенную предрасположенность). Возникновение эмоций имеет «оценочное влияние» на деятельность специфических систем, так как, подкрепляя определенные способы действий, пути решения поставленных задач, они обеспечивают избирательный характер поведения в ситуациях со многими выборами.
Лимбическая система участвует в формировании ориентировочных и условных рефлексов. Благодаря центрам лимбической системы могут вырабатываться даже без участия других отделов коры оборонительные и пищевые условные рефлексы. При поражениях этой системы затрудняется упрочение условных рефлексов, нарушаются процессы памяти, теряется избирательность реакций и отмечается неумеренное их усиление (чрезмерно повышенная двигательная активность и т. д.). Известно, что так называемые психотропные вещества, изменяющие нормальную психическую деятельность человека, действуют именно на структуры лимбической системы. Таким образом, лимбическая система задает общий контекст поведения, в зависимости от условий, переводя в нужное предрасположенное состояние- эмоцию. Направленность эмоции (положительная или отрицательна) и определяет вид формирующегося рефлекса и более сложной реакции.
Электрические раздражения различных участков лимбической системы через вживленные электроды (в эксперименте на животных и в клинике в процессе лечения больных) выявили наличие центров удовольствия, формирующих положительные эмоции, и центров неудовольствия, формирующих отрицательные эмоции. Изолированное раздражение таких точек в глубоких структурах мозга человека вызывало появление чувства «беспричинной радости», «беспредметной тоски», «безотчетного страха».
В специальных опытах с самораздражением на крысах животное приучали нажимом лапы на педаль замыкать цепь и производить электрическое раздражение собственного мозга через вживленные электроды. При локализации электродов в центрах отрицательных эмоций (некоторые области таламуса) животное стараюсь избегать замыкания цепи, а при их расположении в центрах положительных эмоций (гипоталамус, средний мозг) нажимы лапой на педаль следовали почти непрерывно, доходя до 8 тыс. раздражений в 1 час.
Велика роль эмоциональных реакций в спорте (положительные эмоции при выполнении физических упражнений—«мышечная радость», радость победы и отрицательные — неудовлетворенность спортивным результатом и др.). Положительные эмоции могут значительно повышать, а отрицательные — понижать работоспособность человека. Большие напряжения, сопровождающие спортивную деятельность, особенно во время соревнований, создают и эмоциональное напряжение—так называемый эмоциональный стресс. От характера протекания в организме реакций эмоционального стресса зависит успешность двигательной деятельности спортсмена.
Вегетативная нервная система. Регуляция деятельности внутренних органов осуществляется нервной системой через специальный ее отдел — вегетативную нервную систему.
Особенности строения вегетативной нервной системы. Все функции организма можно разделить на соматические, или анимальные (от лат. animal — животное), связанные с деятельностью скелетных мышц, — организация позы и перемещение в пространстве, и вегетативные (от лат. vegetativus — растительный), связанные с деятельностью внутренних органов,—процессы дыхания, кровообращения, пищеварения, выделения, обмена веществ, роста и размножения. Деление это условно, так как вегетативные процессы присущи также и двигательному аппарату (например, обмен веществ и др.); двигательная деятельность неразрывно связана с изменением дыхания, кровообращения и пр.
Раздражения различных рецепторов тела и рефлекторные ответы нервных центров могут вызывать изменения как соматических, так и вегетативных функций, т. е. афферентные и центральные отделы этих рефлекторных дуг общие. Различны лишь их эфферентные отделы.
Совокупность эфферентных нервных клеток спинного и головного мозга, а также клеток особых узлов (ганглиев), иннервирующих внутренние органы, называют вегетативной нервной системой. Следовательно, эта система представляет собой эфферентный отдел нервной системы, через который центральная нервная система управляет деятельностью внутренних органов.
Характерной особенностью эфферентных путей, входящих в рефлекторные дуги вегетативных рефлексов, является их двухнейронное строение. От тела первого эфферентного нейрона, который находится в центральной нервной системе (в спинном, продолговатом или среднем мозгу), отходит длинный аксон, образующий предузловое (или преганглионарное) волокно. В вегетативных ганглиях — скоплениях клеточных тел вне центральной нервной системы—возбуждение переключается на второй эфферентный нейрон, от которого отходит послеузловое (или постганглионарное) волокно к иннервируемому органу (рис. 56).
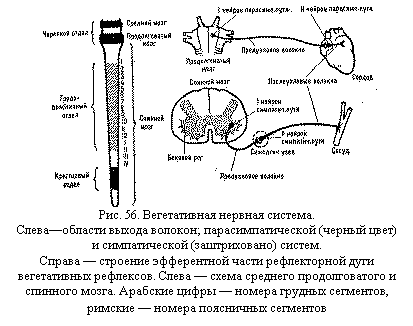 Вегетативная
нервная система подразделяется на 2 отдела — симпатический и парасимпатический.
Эфферентные пути симпатической нервной системы начинаются в грудном и
поясничном отделах спинного мозга от нейронов его боковых рогов. Передача
возбуждения с предузловых симпатических волокон на послеузловые происходит в
ганглиях пограничных симпатических стволов с участием медиатора ацетилхолина, а
передача возбуждения с послеузловых волокон на иннервируемые органы — с
участием медиатора адреналина, или симпатина. Эфферентные пути парасимпатической
нервной системы начинаются в головном мозгу от некоторых ядер среднего и
продолговатого мозга и от нейронов крестцового отдела спинного мозга.
Парасимпатические ганглии расположены непосредственной близости от
иннервируемых органов или внутри их. Проведение возбуждения в синапсах
парасимпатического пути происходит с участием медиатора ацетилхолина.
Вегетативная
нервная система подразделяется на 2 отдела — симпатический и парасимпатический.
Эфферентные пути симпатической нервной системы начинаются в грудном и
поясничном отделах спинного мозга от нейронов его боковых рогов. Передача
возбуждения с предузловых симпатических волокон на послеузловые происходит в
ганглиях пограничных симпатических стволов с участием медиатора ацетилхолина, а
передача возбуждения с послеузловых волокон на иннервируемые органы — с
участием медиатора адреналина, или симпатина. Эфферентные пути парасимпатической
нервной системы начинаются в головном мозгу от некоторых ядер среднего и
продолговатого мозга и от нейронов крестцового отдела спинного мозга.
Парасимпатические ганглии расположены непосредственной близости от
иннервируемых органов или внутри их. Проведение возбуждения в синапсах
парасимпатического пути происходит с участием медиатора ацетилхолина.
Роль вегетативной нервной системы в организме. Вегетативная нервная система, регулируя деятельность внутренних органов, повышая обмен веществ скелетных мышц, улучшая их кровоснабжение, повышая функциональное состояние нервных Центров и т. д., способствует осуществлению функций соматической и нервной системы, которая обеспечивает активную приспособительную деятельность организма во внешней среде (прием внешних сигналов, их обработку, двигательную деятельность, направленную на защиту организма, на поиски пищи, у человека—двигательные акты, связанные с бытовой, трудовой, спортивной деятельностью и пр.). Передача нервных влияний в соматической нервной системе осуществляется с большой скоростью (толстые соматические волокла имеют высокую возбудимость и скорость проведения 50— 140 м/сек). Соматические воздействия на отдельные части двигательного аппарата характеризуются высокой избирательностью. вегетативная нервная система участвует в этих приспособительных реакциях организма, особенно при чрезвычайных напряжениях (стресс).
Другой существенной стороной деятельности вегетативной нервной системы является ее огромная роль в поддержании постоянства внутренней среды организма.
Постоянство физиологических показателей может обеспечиваться различными путями. Например, постоянство уровня кровяного давления поддерживается изменениями деятельности сердца, про. света сосудов, количества циркулирующей крови, ее перераспределением в организме и т. п. В гомеостатических реакциях наряду с нервными влияниями, передающимися по вегетативным волокнам имеют значение гуморальные влияния. Все эти влияния в отличие от соматических передаются в организме значительно медленнее и более диффузно. Тонкие вегетативные нервные волокна отличаются низкой возбудимостью и малой скоростью проведения возбуждения (в предузловых волокнах скорость проведения составляет 3— 20 м/сек, а в послеузловых—0,5—3 м/сек).
Согласно представлениям И. П. Павлова и Л. А. Орбели, все нервные влияния делятся на пусковые, включающие деятельность органа, и трофические, изменяющие его обмен веществ и функциональное состояние. Многие влияния вегетативной нервной системы можно рассматривать как трофические.
Функции симпатического отдела вегетативной нервной системы. С участием этого отдела протекают многие важные рефлексы в организме, направленные на обеспечение его деятельного состояния, в том числе двигательной деятельности. К ним относятся рефлексы расширения бронхов, учащения и усиления сердечных сокращений, расширения сосудов сердца и легких при одновременном сужении сосудов кожи и органов брюшной полости (обеспечение перераспределения крови), выброс депонированной крови из печени и селезенки, расщепление гликогена до глюкозы в печени (мобилизация углеводных источников энергии), усиление деятельности желез внутренней секреции потовых желез. Симпатический отдел нервной системы снижает деятельность ряда внутренних органов: в результате сужения сосудов в почках уменьшаются процессы мочеобразования, угнетается секреторная и моторная деятельность органов желудочно-кишечного тракта, предотвращается акт мочеиспускания (расслабляется мышца стенки мочевого пузыря и сокращается его сфинктер). Повышенная активность организма сопровождается симпатическим рефлексом расширения зрачка.
Огромное значение для двигательной деятельности организма имеет трофическое влияние симпатических нервов на скелетные мышцы. Раздражение этих нервов не вызывает сокращения мышц. Однако сниженная амплитуда сокращений утомленной мышцы может снова увеличиться при возбуждении симпатической нервной системы—эффект Орбели—Гинецинского. Усиление сокращений можно наблюдать и на неутомленной мышце, присоединяя к раздражениям двигательных нервов раздражения симпатических волокон (Я. Б. Лехтман). Более того, симпатические влияния на скелетные мышцы в целостном организме возникают раньше, чем пусковые влияния двигательных нервов, заранее подготавливая мышцы к работе. Л. А. Орбели подчеркивал важнейшее значение симпатических воздействий для приспособления (адаптации) организма к работе, к различным условиям внешней среды, что отражено в его учении об адаптационно-трофической роли симпатической нервной системы.
Функции парасимпатического отдела вегетативной нервной системы. Этот отдел нервной системы принимает активное участие в регуляции деятельности внутренних органов, в процессах восстановления организма после деятельного состояния.
Парасимпатическая нервная система осуществляет сужение бронхов, замедление и ослабление сердечных сокращений; сужение сосудов сердца; пополнение энергоресурсов (синтез гликогена в печени и усиление процессов пищеварения); усиление процессов мочеобразования в почках и обеспечение акта мочеиспускания (сокращение мышц мочевого пузыря и расслабление его сфинктера) и др.
Парасимпатическая нервная система в противоположность симпатической преимущественно оказывает пусковые влияния: сужение зрачка, включение деятельности пищеварительных желез и т. д.
§ 11. Общие принципы регуляции двигательной деятельности
Теория функциональной системы. Современные представления об организации целостных актов поведения живых организмов отражены в теории функциональной системы (П. К. Анохин). Для удовлетворения своих потребностей в условиях постоянного изменения внешней среды организму необходимо ставить перед собой определенные задачи и в своей поведенческой деятельности добиваться достижения намеченного результата.
Согласно теории П. К. Анохина, именно полезный результат является решающим фактором поведения и для его достижения в нервной системе формируется группа взаимосвязанных нейронов — так называемая функциональная система. Сколько нервных клеток из огромного их числа будет включено в эту систему, какой уровень активности этих клеток необходим в данный момент, какие взаимосвязи должны возникнуть между ними и какие возможные взаимосвязи должны быть исключены — все это определяется намечаемым результатом. Следовательно, важная задача деятельности системы — выявление и оценка результата действия.
Организм в своем поведении использует далеко не все структурные и функциональные свои возможности (степени свободы), а оставляет лишь необходимые. Без этого деятельность его была бы хаотичной, нецелесообразной. Создание функциональной системы необходимо для того, чтобы отдельные ее элементы действовали не самостоятельно и независимо друг от друга, а подчинялись общей задаче — получить искомый результат.
Деятельность функциональной системы можно разбить на отдельные последовательные этапы: 1) обработка всех сигналов поступающих из внешней и внутренней среды организма — так называемый афферентный синтез; 2) принятие решения; 3) создание (на основе принятого решения) представления об ожидаемом результате действия и формирование конкретной программы действий для достижения этого результата; 4) анализ полученного результата и уточнение программы действий.
На первом этапе (афферентный синтез) в нервных центрах одновременно взаимодействуют 4 типа поступающих к ним раздражении:
1) пусковая афферентация—сигналы, вызывающие действие; 2) обстановочная афферентация — все остальные внешние раздражения, создающие афферентный фон; 3) мотивация — собственные потребности организма, доминирующие в данный момент: биологические мотивации — жажда, голод, страх и другие, а также высшие мотивации, формирующиеся у человека под влиянием социальных факторов (желание добиться какой-либо цели в жизни, достичь определенного результата в спорте и др.); 4) память—имеющиеся в нервной системе следы прежних раздражении, накопленный опыт.
Все эти виды сигналов должны одновременно обрабатываться в нервных клетках и сопоставляться друг с другом. При оценке текущих афферентных раздражении производится их сопоставление со следами прежних раздражении, хранящихся в памяти. Большую роль в отборе сигналов играют доминирующие мотивации, так как они повышают восприимчивость организма к наиболее важным в данный момент сигналам.
На основании афферентного синтеза принимается решение («что надо делать») и вырабатывается программа действий («как надо делать»). При этом производится оценка всех возможных путей решения возникших перед организмом задач и ответных реакций организма и выбирается лишь одно необходимое решение. Одновременно в мозгу создается представление о том, какой результат должен быть получен.
Динамическая функциональная система — это организованная система, результат действия которой сообщается в центральную нервную систему.
Без постоянной оценки результата действия невозможно выполнить задуманную программу, вносить в нее поправки для улучшения результата. Например, совершенствование техники выполнения упражнения невозможно без постоянной информации о его результатах.
Представление о функциональной системе П. К. Анохина имеет универсальное значение, так как помогает понять различные стороны деятельности организма (механизмы его поведения, закономерности формирования различных функций в процессе индивидуального развития, особенности их нарушения при патологии и возможные пути восстановления).
Циклическая многоуровневая система регуляции движений. Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах центральной нервной системы. Каждый из нервных центров, входящих в данный комплекс, может воздействовать как на нижележащие, так и на вышележащие центры. При этом между ними могут возникать циклы взаимных влияний от высших центров к низшим и обратно. Например, очень часто наблюдается циркуляция импульсов между корой и промежуточным мозгом: импульсы из коры больших полушарий могут влиять на активность нейронов промежуточного мозга, а те, в свою очередь, могут воздействовать на корковые нейроны. В каждом подобном цикле роль задающих звеньев принадлежит преимущественно вышележащим центрам, а активность нижележащих центров является регулируемым звеном. Эта активность поддерживается на заданном уровне в зависимости от потребностей организма.
Наличие циклических взаимосвязей между различными отделами нервной системы позволяет говорить о замкнутой системе регулирования, или замкнутом цикле регулирования.
В процессе двигательной деятельности одновременно возникает множество замкнутых циклов регулирования (например, между корой и ретикулярной формацией, между корой и мозжечком, между мотонейронами спинного мозга и мышцей и др.).
Таким образом, система регуляции движения является многоэтажной (многоуровневой) системой. Важную роль в этой сложной функциональной системе играет кора больших полушарий, которая управляет процессами, протекающими в этих циклах регулирования, и направляет их на решение общей задачи—достижение рабочего эффекта, или цели движения.
Программирование. Для решения поставленных задач в центральной нервной системе вырабатываются программы двигательных действий. Они определяют набор и последовательность включения двигательных актов. В соответствии с этой программой включаются команды к эфферентным нейронам и далее—команды к мышцам.
Программы двигательных действий у человека разнообразны. Они обеспечивают бытовую, производственную и спортивную его деятельность. При этом команды к действию (например, выстрел стартового пистолета, команда «марш!») являются лишь сигналами о начале работы, сама же мышечная работа развертывается по имеющейся программе. В ряде случаев программирование протекает в условиях крайнего дефицита времени, (например при спортивных играх, фехтовании, борьбе и др.). В этих случаях требуется высокая скорость переработки поступающей информации и выполнения ответных действий.
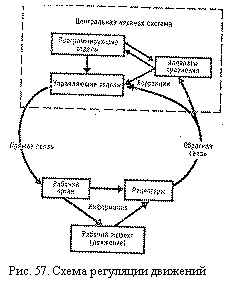 Программы
двигательных действий могут быть очень сложными, особенно при нестандартной
работе, так как для достижения одного и того же результата в изменяющихся
условиях (например, для успешного выполнения броска в баскетбольную корзину при
разных игровых ситуациях) могут потребоваться включение различных мышечных
групп, различная тактика поведения.
Программы
двигательных действий могут быть очень сложными, особенно при нестандартной
работе, так как для достижения одного и того же результата в изменяющихся
условиях (например, для успешного выполнения броска в баскетбольную корзину при
разных игровых ситуациях) могут потребоваться включение различных мышечных
групп, различная тактика поведения.
Обратные связи. Для функциональной системы регуляции движений недостаточно лишь наличия программирующих и управляющих нервных центров. Характерной особенностью подобных систем регуляции является наличие обратной афферентации, или так называемой обратной связи, от рабочего органа к регулирующим нервным центрам (рис. 57). Осведомление центральной нервной системы о деятельности, вызванной на периферии ее управляющими импульсами, происходит при помощи афферентных сигналов, отправляемых органами зрения проприорецепторами мышечной суставного аппарата и другими рецепторами.
Таким образом, с помощью обратных связей производится оценка выполненных движений, определяется их эффективность.
Аппараты сравнения. Эти нервные аппараты, получая обратную сигнализацию, сличают заданную программу с реальными результатами ее выполнения (П. К. Анохин, Н. А. Бернштейн и др.). Имеются данные, что такие аппараты находятся в лобных долях, подкорковых ядрах (хвостатые тела) и других образованиях.
Если есть расхождения фактического и требуемого действия, то в аппаратах сравнения возникают сигналы об ошибках—так называемые импульсы рассогласования. Соответственно этим импульсам в систему, управляющую движением, вносятся необходимые поправки — коррекции. В результате этого в процессе выполнения движения можно сохранить его основные черты, целесообразность, несмотря на изменение условий его осуществления. Так, сохраняется, например, индивидуальный характер походки человека при различиях в обуви, грунте, скорости движения, положении центра тяжести и т. д.
§ 12. Позно-тонические реакции
Общее представление о двигательной деятельности человека.
Двигательная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы.
В основе двигательной деятельности человека лежат 2 формы механической реакции мышечных волокон: 1) их фазное (тетаническое) сокращение и 2) длительное тоническое напряжение. Фазная деятельность обеспечивает быстрые и сильные сокращения мышц при выполнении движений, а также коррекции позы. Эти сокращения осуществляются быстрыми двигательными единицами. Тоническое напряжение отличается от фазного медленным развитием, большей экономичностью, преобладанием изометрического режима и преимущественным участием медленных двигательных единиц. Оно лежит в основе поддержания определенной позы тела. Если при поддержании позы требуется значительное усилие мышц (например для выполнении угла в упоре), то включаются фазные сокращения мышц.
Процессы поддержания позы и двигательные акты скелетных мышц тесно связаны между собой. С одной стороны, движения всегда происходят на фоне определенной позы тела; с другой стороны, процессы поддержания позы обеспечиваются не только тоническим напряжением мышц, но и их фазной деятельностью.
В двигательной деятельности человека различают произвольные и непроизвольные движения и позы. Произвольные движения вырабатываются с участием сознания, а по мере их освоения могут происходить и бессознательно. Ведущую роль в осуществлении произвольных движений играют высшие интегративные области коры больших полушарий (лобные доли и др.). Еще до их выполнения в мозгу создается модель предстоящего движения, его программа. Человек сознает цель этого движения или необходимость принятия определенной позы. В процессе выполнения двигательного акта он, внося коррекции в его выполнение, добивается запрограммированного результата действия.
Непроизвольные движения и позы, выполняемые бессознательно, являются преимущественно безусловными рефлексами (ориентировочными, защитными и др.). Непроизвольными также становятся те произвольные движения и позы, которые автоматизируются при многократном их повторении. Непроизвольная двигательная деятельность может осуществляться различными отделами нервной системы.
Рефлекторная природа мышечного тонуса. В поддержании тонического напряжения мышц участвует сложная многоуровневая система нервных центров, находящаяся под контролем коры. больших полушарий. Многочисленные опыты на животных, а также наблюдения за нарушениями мышечной деятельности у человека в клинике позволили выявить значение отдельных участков мозга в осуществлении и регуляции мышечного тонуса.
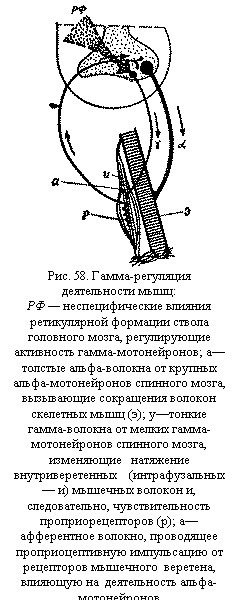 По
своей природе мышечный тонус является рефлекторным актом.
По
своей природе мышечный тонус является рефлекторным актом.
Для возникновения тонуса в мышцах достаточна уже рефлекторная деятельность спинного мозга. Однако тонкое перераспределение тонуса между различными мышцами, необходимое для протекания реакций целостного организма, осуществляется более высокими Этажами центральной нервной системы, а его произвольная регуляция — корой больших полушарий.
В наиболее простом случае появление тонуса обусловлено сокращением мышцы в ответ на раздражение проприорецепторов при ее растяжении. Такой рефлекс, осуществляемый спинным мозгом называется миотатическим рефлексом на растяжение. Если это растяжение длительно, то рефлекторное сокращение носит тоже длительный тонический характер. При растяжении мышцы возбуждаются чувствительные нервные окончания в мышечных веретенах. Импульсы от них направляются по толстым афферентным волокнам в передние рога спинного мозга, где непосредственно (без участия вставочных нейронов) передаются на альфа-мотонейроны и вызывают сокращение мышц. Такая же двухнейронная рефлекторная дуга лежит в основе аналогичного сухожильного рефлекса — сокращения мышцы при ударе по ее сухожилию. К сухожильным рефлексам относят рефлекс ахиллова сухожилия, коленный рефлекс и др.
В естественных условиях основным раздражителем рецепторов мышечных и сухожильных веретен является сила тяжести, растягивающая скелетные мышцы, особенно мышцы-разгибатели. Поражение спинного мозга приводит к исчезновению тонуса мышц.
Гамма-регуляция мышечного тонуса. Степень тонического напряжения мышцы зависит от частоты импульсов, посылаемых к ней тоническими альфа-мотонейронами. Частота разрядов альфа-мотонейронов, в свою очередь, регулируется импульсами от проприорецепторов той же самой мышцы. Получается замкнутое кольцо между мышцей и иннервирующими ее мотонейронами. Однако потоки импульсов в этом кольце могут регулироваться вышележащими этажами нервной системы с помощью гамма-мотонейронов спинного мозга (рис. 58). Разряд гамма-мотонейронов повышает чувствительность мышечных веретен. В результате увеличивается поток импульсов от рецепторов к альфа-мотонейронам и от альфа-мотонейронов к мышце. Тем самым повышается мышечный тонус.
Изменение напряжения мышц под влиянием деятельности гамма-мотонейронов называется гамма-регуляцией. Активность гамма-мотонейронов находится под контролем ретикулярной формации ствола мозга. В регуляции их деятельности, а следовательно, и в регуляции тонуса скелетных мышц участвуют мозжечок, подкорковые и другие структуры экстрапирамидной системы. Высший контроль осуществляется корой больших полушарий.
Значение различных отделов головного мозга в регуляции тонуса скелетных мышц. Различные отделы головного мозга, посылая импульсы к вставочным нейронам и мотонейронам спинного мозга, могут регулировать тонус скелетных мышц, изменяя тем самым позные и двигательные реакции организма. Влияния на мышечный тонус оказывают обе системы головного мозга — как специфическая, так и неспецифическая.
Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел среднего мозга, а угнетение — тормозящий отдел продолговатого мозга.
Специфические системы действуют избирательно — на отдельные группы мышц. Усиление тонуса мышц-сгибателей вызывают кортико-спинальная, кортико-рубро-спинальная и частично кортико-ретикуло-спинальная системы (последняя оказывает также неспецифическое диффузное влияние). Одновременно эти системы снижают тонус мышц-разгибателей. В противоположность им вестибуло-спинальная система повышает тонус мышц-разгибателей и тормозит тоническое напряжение мышц-сгибателей. Изменения тонуса могут происходить очень быстро, нередко опережая двигательные акты. Это позволяет организму подготовиться к необходимому движению, заранее изменив позу тела.
В опытах на животных было показано, что при перерезке головного мозга между продолговатым и средним мозгом резко повышается тонус мышц-разгибателей. Это явление было названо децеребрационной ригидностью (децеребрация — лишение животного всех отделов головного мозга выше продолговатого, а ригидность — это твердость). После децеребрации у кошки напряжены мышцы спины, голова запрокинута назад, а хвост и конечности вытянуты. Этого не происходит в целостном организме, так как влияния из вестибулярных ядер продолговатого мозга, усиливающие тонус мышц-разгибателей, уравновешиваются в необходимой степени воздействиями других нисходящих систем, усиливающих тонус мышц-сгибателей. Правильное распределение тонуса различных мышц, которое лежит в основе организации определенной позы, обеспечивается деятельностью мозжечка, подкорковых ядер (полосатого тела и бледного ядра) и коры больших полушарий.
Мозжечок не имеет прямых связей со спинным мозгом, но через красные ядра среднего мозга повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга усиливает тонус Мышц-разгибателей. Поражения мозжечка нарушают правильное перераспределение тонуса скелетных мышц, что изменяет не только позные, но и двигательные реакции организма. При мозжечковых расстройствах вследствие снижения и ненормального распределения тонуса мышц конечностей нарушается походка (атаксия). Бледное ядро угнетает тонус мышц, а при его поражениях тонус усиливается. Полосатое тело, регулируя деятельность бледного ядра, снижает его угнетающее действие. Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы организма, обеспечивается ее соответствие двигательным задачам.
Непосредственное отношение к регуляции тонуса имеют медленные пирамидные нейроны коры — так называемые нейроны положения, образующие медленную часть кортико-спинального пути (пирамидного тракта). По этому пути передаются прямые корковые воздействия на спинной мозг. Кроме того, изменяя активность ретикулярной формации ствола мозга, подкорковых ядер, мозжечка, ядер среднего и продолговатого мозга, кора больших полушарий оказывает через них параллельное влияние на тонус мышц.
Позы. Позой называется закрепление частей скелета в определенном положении.
Значение поз в двигательной деятельности человека. Организация позы необходима для преодоления силы земного притяжения. Поза человека служит для сохранения равновесия тела в состоянии двигательного покоя и при выполнении статической или динамической работы, а также для фиксации ряда суставов, необходимой для осуществления движения в других суставах.
При многих видах трудовой и спортивной деятельности человеку приходится принимать новые необычные позы, без которых невозможно движение. Поза, принимаемая при работе, называется рабочей позой. Правильная ее организация способствует повышению работоспособности человека. Наконец, большое значение имеют предрабочие изменения позы, которые заранее компенсируют возможные изменения центра тяжести тела и предотвращают его падение, а также препятствуют действию реактивных сил. Реакции поддержания позы обеспечиваются чаще всего тоническим напряжением мышц, а при больших нагрузках и при коррекциях, исправляющих позу,— фазными сокращениями мышц.
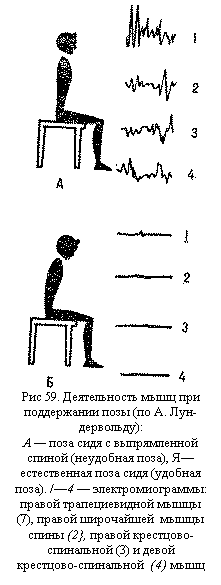 Во
время отдыха или выполнения работы человек выбирает наиболее удобную позу, при
которой возможно минимальное напряжение скелетных мышц (рис. 59). Организация
такой оптимальной позы часто требует длительной тренировки. Например, при
стрельбе из положения стоя только у высококвалифицированных стрелков тоническая
активность мышц, обеспечивающих стояние, минимальна (на электромиограмме
регистрируется наименьшая электрическая активность мышц). У спортсменов низкой
квалификации и новичков наблюдается значительное напряжение этих мышц.
Во
время отдыха или выполнения работы человек выбирает наиболее удобную позу, при
которой возможно минимальное напряжение скелетных мышц (рис. 59). Организация
такой оптимальной позы часто требует длительной тренировки. Например, при
стрельбе из положения стоя только у высококвалифицированных стрелков тоническая
активность мышц, обеспечивающих стояние, минимальна (на электромиограмме
регистрируется наименьшая электрическая активность мышц). У спортсменов низкой
квалификации и новичков наблюдается значительное напряжение этих мышц.
Частным случаем позных реакций является поддержание заданного усилия мышцы и заданного суставного угла. Усилия мышц не сохраняются постоянными. Каждый раз, когда мышца расслабляется и возникают отклонения от заданного усилия или от определенного положения костей, появляются небольшие фазные сокращения работающих мышц, исправляющие эти отклонения. Нарастание усилия мышц достигается благодаря увеличению числа активных альфа-мотонейронов спинного мозга. Таким образом, сохранение достоянного усилия и суставного угла поддерживается как тоническими, так и фазными сокращениями мышц.
Известно, что человек может (особенно в результате мышечной тренировки) очень точно дозировать усилия своих мышц и определять положение суставов. Информацию о положении конечностей сообщают в кору больших полушарий многие рецепторы — мышечные и сухожильные, зрительные, кожные, но особенно большое значение имеют рецепторы суставных, сумок.
Позно-тонические реакции мышц часто бывают кратковременными, так как возникают только в определенные фазы движения. Они способствуют, в частности, фиксации суставов при отталкивании от опоры в процессе ходьбы. Электромиографические исследования ходьбы показали, что наряду с работой мышц-антагонистов, вызывающих сгибания и разгибания нижних конечностей (например, передней большеберцовой мышцы и икроножной мышцы ноги), в фазу отталкивания носком от опоры происходит тоническое напряжение мышц стопы, фиксирующих ее свод.
Установочные рефлексы. Эти рефлексы способствуют сохранению позы. К ним относятся статические и статокинетические рефлексы, в осуществлении которых большое значение имеет продолговатый и средний мозг.
Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменении положения головы в пространстве — это так называемые лабиринтные рефлексы. возникающие в результате раздражения рецепторов вестибулярного аппарата; 2) при изменении положения головы по отношению к туловищу — шейные рефлексы, с проприорецепторов мышц шеи и 3) при нарушении нормальной позы тела — выпрямительные рефлексы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус сгибателей (лабиринтный рефлекс). Подобные реакции имеют место при выполнении многих физических упражнений. Так, выполнение гимнастом стойки на кистях облегчается при отклонении головы назад выполнение группировки во время прыжков в воду или акробатических прыжков — при наклоне головы вперед. Выпрямительные рефлексы — это последовательные сокращения мышц шеи и туловища которые обеспечивают возвращение тела в вертикальное положение теменем кверху. У человека они проявляются, например, во время ныряния.
Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. Например, при быстром подъеме усиливается тонус сгибателей, и человек приседает, а при быстром спуске усиливается тонус разгибателей, и человек выпрямляется — это так называемый лифтный рефлекс. При вращении тела реакции противовращения проявляются в отклонении головы, тела и глаз в сторону, противоположную движению. Движение глаз со скоростью вращения тела, но в противоположную сторону и быстрое возвращение в исходное положение — нистагм глаз — обеспечивают сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
В двигательной деятельности человека часто возникают ситуации, когда требуется подавить эти установочные рефлексы. Произвольное подавление врожденных установочных рефлексов продолговатого и среднего мозга обеспечивается тормозящими влияниями из коры полушарий. Например, для спринтера невыгодно раннее выпрямление туловища при стартовом разбеге, поэтому корой больших полушарий тормозится выпрямительный рефлекс.
§ 13. Двигательная деятельность
Движения человека являются комплексом различных двигательных актов, которые обеспечиваются как сравнительно простыми двигательными рефлексами, так и сложнейшими их комбинациями.
Двигательные акты различной сложности. Рассматривая различные двигательные акты человека, можно выделить элементарные двигательные рефлексы, более сложные—ритмические рефлексы и, наконец, особенно сложные формы двигательной деятельности, обеспечивающие поведение человека.
Элементарные двигательные рефлексы. При сложных движениях человека используются элементарные двигательные рефлексы, осуществляемые спинным мозгом. К простым безусловным двигательным рефлексам спинного мозга относятся: 1) рефлексы, на растяжение (миотатические и сухожильные рефлексы—рис. 60, А), 2) сгибательные рефлексы на раздражение кожных рецепторов (рефлексы удаления от раздражителя — см. рис. 60, Б) и 3) рефлексы отталкивания (рефлексы сближения с опорой—см. рис. 60, В).
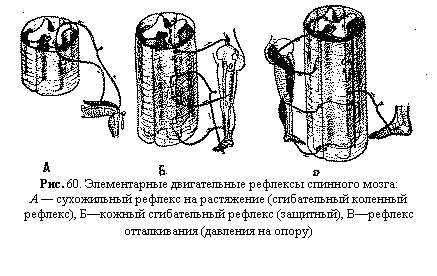 Миотатические
и сухожильные рефлексы возникают при растяжении рецепторов мышц и сухожилий.
Быстрое растягивание этих рецепторов рефлекторно вызывает фазное сокращение
скелетных мышц. Это рефлекс активного противодействия мышцы ее растяжению.
Рефлекторная дуга рефлекса на растяжение — моносинаптическая Она состоит всего
из двух нейронов — афферентного и эфферентного, между которыми имеется лишь
один синапс. Простота рефлекторной дуги обеспечивает быструю и точную реакцию
мышцы, меньшую утомляемость рефлекторных ответов. Рефлексы на растяжение имеют
большое значение при статической деятельности мышц для поддержания определенной
позы, а также при осуществлении локомоторных актов (ходьбы, бега и др.). В
произвольной двигательной деятельности человека иногда необходимо подавлять эти
рефлексы (например, выполнение гимнастического шпагата требует подавления
рефлекса на растяжение).
Миотатические
и сухожильные рефлексы возникают при растяжении рецепторов мышц и сухожилий.
Быстрое растягивание этих рецепторов рефлекторно вызывает фазное сокращение
скелетных мышц. Это рефлекс активного противодействия мышцы ее растяжению.
Рефлекторная дуга рефлекса на растяжение — моносинаптическая Она состоит всего
из двух нейронов — афферентного и эфферентного, между которыми имеется лишь
один синапс. Простота рефлекторной дуги обеспечивает быструю и точную реакцию
мышцы, меньшую утомляемость рефлекторных ответов. Рефлексы на растяжение имеют
большое значение при статической деятельности мышц для поддержания определенной
позы, а также при осуществлении локомоторных актов (ходьбы, бега и др.). В
произвольной двигательной деятельности человека иногда необходимо подавлять эти
рефлексы (например, выполнение гимнастического шпагата требует подавления
рефлекса на растяжение).
При раздражении рецепторов кожи (болевых, температурных и др.) возникает сокращение мышц-сгибателей, которое позволяет отдернуть конечность от повреждающего раздражителя. Этот сгибательный кожный рефлекс носит защитный характер. В его основе лежит полисинаптическая рефлекторная дуга. В отличие от моносинаптического рефлекса на растяжение при сгибательном рефлексе быстрее наступает утомление; время этого рефлекса в связи с наличием вставочных нейронов значительно больше.
Одна из разновидностей элементарных рефлексов спинного мозга—рефлекс отталкивания, описанный Ч. Шеррингтоном. Этот Рефлекс возникает при раздражении кожи стопы опорой. Однако, в Отличие от сгибательного рефлекса, он приводит не к отдергиванию конечности от раздражителя, а к сближению с раздражителем. В первом случае мы имеем защитный рефлекс, предохраняющий организм от разрушения. Во втором случае—биологически целесообразный акт, обеспечивающий контакт с опорой при стоянии и отталкивание от нее при передвижениях. Этот рефлекс лежит в основе сложных локомоций: ходьбы, бега, прыжков в длину ил' высоту и др.
Ритмические рефлексы. Составной частью различны» сложных двигательных действий, как произвольных, так и непроизвольных, часто являются ритмические рефлексы. Они особенно выражены при выполнении циклических движений. Их возникновение и протекание связаны с проявлением механизмов взаимосочетанной (реципрокной) иннервации мышц-антагонистов.
Эта форма рефлексов — одна из древних и относительно простых форм — имеет большое значение в организации многих сложных движений. Например, включение ритмических циклоидных движений в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи; то же самое происходит при освоении акта ходьбы — переход от отдельных шагов к ритмической походке (Н. А. Бернштейн).
Наиболее простая форма ритмического рефлекса — чесательный рефлекс животных, который обеспечивается попеременным сокращением и расслаблением одних и тех же мышц одной конечности. Возбуждение центров-сгибателей сопровождается одновременным торможением центров мышц-разгибателей, в чем проявляется реципрокная их иннервация. В последующей фазе движения в этих центрах возникают противоположные процессы—возбуждение вслед за торможением и торможение вслед за возбуждением. При этом проявляется так называемый рефлекс отдачи: после акта сокращения наступает расслабление, а затем снова акт сокращения. Рефлекс отдачи — это рефлекс на растяжение с проприорецепторов мышц и сухожилий, в результате которого возникает сокращение растянутого разгибателя. Такая простая форма ритмического рефлекса часто встречается в двигательной деятельности человека (например, при забивании молотком гвоздей).
Более сложная форма ритмического рефлекса — шагательный рефлекс, лежащий в основе ходьбы, бега и других локомоций. Для осуществления шагательных движений требуется участие уже не одной, а двух симметричных конечностей. Подключение второй конечности к координированному двигательному акту происходит в результате осуществления перекрестного рефлекса. Он хорошо виден на спинальном животном. Раздражение рецепторов кожи вызывает сгибание раздражаемой конечности и одновременное разгибание одноименной конечности на противоположной стороне тела. Этот рефлекс осуществляется с участием вставочных тормозных нейронов, вызывающих торможение мотонейронов на противоположной стороне спинного мозга. В целостном организме человека сгибание одной ноги вызывает перекрестный разгибательный рефлекс другой ноги, принимающей на себя тяжесть тела при стоянии, ходьбе и пр.
Следовательно, механизмы шагательных движений заложены уже на уровне спинного мозга. У спиналыюго животного в первой фазе движения болевое раздражение рецепторов кожи вызывает сгибательный рефлекс раздражаемой конечности (табл. 4). Одновременно при участии тормозных клеток Рэншоу происходит торможение центров мышц-разгибателей этой же конечности и торможение центров мышц-сгибателей симметричной конечности. Во вторую фазу движения возникает рефлекс отдачи с проприорецепторов растянутых мышц-разгибателей раздражаемой конечности и мышц-разгибателей другой стороны тела. Импульсы от этих рецепторов вызывают рефлекторное сокращение мышц-разгибателей и одновременное торможение центров мышц-сгибателей той же конечности и мышц разгибателей симметричной конечности. Последовательная смена этих двух фаз движения и составляет ритмический акт шагания.
Включение этого спинального механизма производится с более высоких этажей нервной системы. В так называемой «локомоторной области» среднего мозга имеются центры, электрическое раздражение которых вызывает шагательные движения конечностей.
В осуществлении шагательного рефлекса принимает участие мозжечок. Удаление одного его полушария у животных приводит к искажению движений (особенно передней конечности). Движения сильно варьируют по амплитуде, заметно искажаются при малейших посторонних воздействиях, происходит чрезмерное сгибание локтевого сустава в фазе переноса конечности.
Таблица 4
Процессы возбуждения (+) и торможения (—) в нервных центрах спинного мозга при осуществлении шагательного рефлекса
|
Рецепторы, вызывающие рефлекс |
Левая лапа |
Левая лапа |
||
|
сгиба- |
разгиба- |
сгиба- |
разгиба- |
|
|
Болевые рецепторы кожи правой лапы |
+ |
— |
— |
+ |
|
Проприорецепторы разгибателей правой лапы и сгибателей левой |
— |
+ |
+ |
— |
|
Проприорецепторы сгибателей правой лапы и разгибателей левой |
+ |
— |
— |
+ |
Протекание ритмических рефлексов, в том числе шагательного, связано с деятельностью различных отделов неспецифической системы. При активирующих влияниях происходит нарастание амплитуды движений, а при угнетающих влияниях — ее снижение.
Важное значение в регуляции ритмических рефлексов имеют подкорковые ядра — бледное ядро и полосатое тело, обеспечивающие их автоматическое протекание, содружественные движения конечностей, вспомогательные реакции (фиксацию суставов и др.).
Высшим регулятором рефлексов является кора больших полушарий, особенно ее премоторная область. Благодаря коре ритмические движения (например, простой акт ходьбы) приобретают определенное смысловое значение, включаются как составной элемент сложные акты поведения.
Об участии различных корковых областей в регуляции ритмических рефлексов свидетельствует появление на электроэнцефалограмме медленных потенциалов, синхронных по частоте с темпом движений. Протекание ритмических рефлексов облегчается тем, что между нервными центрами и работающими мышцами возникают согласованные во времени потоки нервных импульсов. Прохождение импульсов от нервных центров на периферию к рабочим органам и от мышц обратно к нервным центрам осуществляется в определенном темпе — в темпе рабочих движений. Налаживание этого темпа и стабильное его поддержание, включение древних ритмических автоматизмов в систему регуляции движений значительно упрощают управление ритмическими рефлексами, делают эту двигательную деятельность более экономичной.
Сложные формы двигательной деятельности. В целостном поведении простые рефлексы, сочетаясь, обусловливают сложные двигательные действия. Социальные условия жизни человека намного усложняют его двигательную деятельность, приводя к появлению специально человеческих форм движений: бытовых, производственных, спортивных. Простые и сложные ритмические рефлексы лежат в основе циклической двигательной деятельности человека: ходьбы, бега, плавания, гребли, ходьбы на лыжах, езды на велосипеде и пр. Большое значение в их осуществлении имеют механизмы реципрокной иннервации мышц-антагонистов (рис. 61).
Однако в целом ряде движений требуется одноименная работа мышц-антагонистов — в одной конечности или даже в обеих. В этих случаях реципрокные отношения, характерные для спинного мозга, подавляются центрами головного мозга. Особенно часто встречаются симметричные (а не реципрокно-перекрестные) движения в деятельности верхних конечностей человека, т. е. происходит одновременное сокращение одноименных мышц левой и правой рук. Наиболее сложные формы движений представляют собой многофазную цепь рефлексов, которая основана уже не на элементарных реципрокных отношениях, а является целостным двигательным навыком, образующимся в процессе обучения по механизму условных рефлексов.
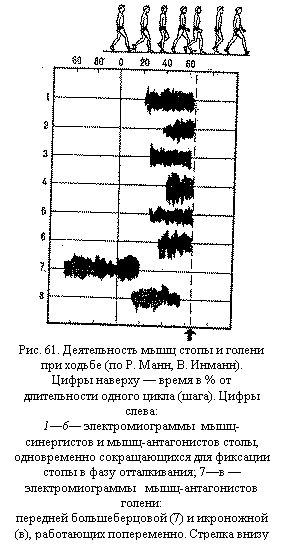 Значение
коры больших полушарии в регуляции произвольных движений. Ведущую роль в
регуляции произвольных движений играет кора больших полушарий, особенно ее
высшие отделы—зоны перекрытия анализаторов, или третичные поля. В коре больших
полушарий происходит выработка цели и задачи движений, соответственно этому
строится и программа конкретных действий, которые нужны человеку для
осуществления цели. В сложные поведенческие акты включаются не только
моторные компоненты, но и необходимые вегетативные компоненты. Еще до начала
движении кора больших полушарий повышает активность тех вставочных и моторных
нейронов спинного мозга, которым предстоит участвовать в движении. В
предстартовый период перед началом циклических движений в электрической
активности коры происходит настройка на темп предстоящих движений. В тот
момент, когда производится движение, кора тормозит деятельность всех
посторонних афферентных путей и оказывается особенно восприимчивой к сигналам
от рецепторов мышц, сухожилий и суставных сумок.
Значение
коры больших полушарии в регуляции произвольных движений. Ведущую роль в
регуляции произвольных движений играет кора больших полушарий, особенно ее
высшие отделы—зоны перекрытия анализаторов, или третичные поля. В коре больших
полушарий происходит выработка цели и задачи движений, соответственно этому
строится и программа конкретных действий, которые нужны человеку для
осуществления цели. В сложные поведенческие акты включаются не только
моторные компоненты, но и необходимые вегетативные компоненты. Еще до начала
движении кора больших полушарий повышает активность тех вставочных и моторных
нейронов спинного мозга, которым предстоит участвовать в движении. В
предстартовый период перед началом циклических движений в электрической
активности коры происходит настройка на темп предстоящих движений. В тот
момент, когда производится движение, кора тормозит деятельность всех
посторонних афферентных путей и оказывается особенно восприимчивой к сигналам
от рецепторов мышц, сухожилий и суставных сумок.
В организации двигательного акта участвуют самые различные отделы коры больших полушарий. Моторная зона коры (поле 4) посылает импульсы к отдельным мышцам, преимущественно к дистальным мышцам конечностей. Объединение отдельных элементов движения в целостный акт осуществляют вторичные поля (6-е и 8-е) премоторной области. Они определяют последовательность двигательных актов, формируют ритмические серии движении, регулируют тонус мышц. Задняя центральная извилина коры — общечувствительная область—обеспечивает субъективное ощущение Движения. Здесь имеются нейроны, сигнализирующие только о возникновении движений в суставе, и нейроны, постоянно информирующие мозг о положении конечности (нейроны движения и нейроны положения).
К пространственной организации движений прямое отношение имеют задние третичные поля — нижнетеменные и теменно-затылочно-височные области коры. С их участием производится оценка удаленности и расположения предметов, оценка расположения отдельных частей собственного тела в пространстве и др. При поражении этих областей у человека теряется представление о «схеме тела» (о том, где находится нос, глаз, ухо, предплечье, спина, как опустить, например, «руки по швам»). Нарушается также представление о «схеме пространства», пространственная ориентация движении у А. Р. Лурия). Трудности возникают при выполнении самых простых актов: человек видит стул и узнает его, но садится мимо чего; он не понимает, откуда идет звук, что означает «влево», «вправо» «вперед», «назад», не может правильно есть (например, ложка с супом попадает мимо рта) и т. д. Становится невозможным использование каких-либо орудий для трудовой или спортивной деятельности.
Области коры, относящиеся к лимбической системе (нижние и внутренние части коры), обеспечивают эмоциональную окраску движений и управляют вегетативными реакциями организма при работе.
В высшей регуляции произвольных движений важнейшая роль принадлежит лобным долям. В третичных полях лобной коры про. исходит сознательное программирование произвольных движений определение цели поведения, двигательных задач и необходимых для их выполнения двигательных актов, а также сопоставление намеченной программы с результатами ее реализации. При регуляции лобными долями движений используется вторая сигнальная система. Движения программируются в ответ на поступающие извне словесные сигналы (словесные указания тренера, спортивные команды я пр.), а также благодаря участию внешней и внутренней речи (мышления) самого человека.
При поражениях лобных долей человек утрачивает способность ставить перед собой задачи, находить пути их решения, сличать выполнение действия с замыслом. При этом сохраняется способность осуществлять любой моторный акт, непосредственно повторяя или копируя его при движениях других людей. Однако сознательно использовать эти движения для достижения какой-либо определенной цели уже невозможно. Появляется стереотипность движений: происходит автоматическое и монотонное повторение одних и тех же однородных двигательных актов, не осознается ошибочность действий, сильно затрудняются реакции выбора.
Таким образом, произвольные движения человека — результат объединенной деятельности самых различных отделов центральной нервной системы. В регуляции таких действий участвует многоэтажная и многозвенная функциональная система, состоящая из многих сотен, тысяч и миллионов нейронов. Работа этой системы сводится к определению оптимальных способов решения двигательных задач: удачного момента для начала движения, наиболее подходящей его структуры (сочетание мышц, степень и скорость их напряжения, порядок включения и т. п.), оптимального уровня функционирования вегетативных систем, постоянной и возможно более эффективной коррекции движений по ходу его выполнения и др.
Речевая регуляция движений. Спецификой регуляции движений у человека является то, что они подчинены речевым воздействиям (как внешним — звуковая и письменная речь, так и внутренним — собственная внутренняя речь). Речевая регуляция движений связана в первую очередь с функцией лобных долей. Исследования покали, что на первых этапах развития — у ребенка 2—3 лет — регуляция произвольных двигательных реакций соответственно речевым инструкциям оказывается недостаточной. Он не может управлять движением в полном соответствии с указаниями другого лица и еще не в состоянии полностью подчинить движения собственным речевым командам. К 3—4 годам ребенок осваивает речевые навыки и сам произносит «надо» или «не надо» делать движения. Эти речевые отчеты помогают ему в регуляции действий. Афферентная импульсация, идущая от речевой мускулатуры, служит важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц. Речевые самоотчеты помогают осмысливать и направлять движения. Постепенно простые речевые реакции усложняются и образуют сложную систему саморегуляции. Внешняя речь ребенка (произносимые им слова) сначала заменяется шепотом, а затем переходит во внутреннюю речь.
Внутренняя речь (процессы мышления) становится важным регулятором действий взрослого человека. При поражении лобной доли у него может сохраняться речь, но нарушается ее регулирующая роль в двигательных действиях. Больной не может выполнять словесные инструкции, исходящие от другого лица, или производить движения согласно собственной речи (например, под собственную команду «раз», «два»). Нарушается сличение результата движения и задуманной программы его выполнения. Движения становятся стереотипными, инертными, теряют свой смысл.
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ (УСЛОВНЫЕ РЕФЛЕКСЫ)
§ 1. Роль высшей нервной деятельности в адаптации организма к изменяющимся условиям среды
Высшая нервная деятельность обеспечивает индивидуальное приспособление организма к изменяющимся условиям внешней и внутренней среды. Она детерминирована (от лат. determinare — определять) совокупным действием многих факторов. К ним относятся, с одной стороны, афферентная импульсация, которая поступает в центральную нервную систему от рецепторов, воспринимающих эндогенные (от греч. endon — внутри, genos — род, происхождение) и экзогенные (от греч. ехо — снаружи) раздражения, т. е. раздражения из внутренней и внешней среды, с другой стороны — следовые явления от прежней деятельности нервной системы, т. е. память. Важная роль в афферентной импульсации принадлежит сенсорным коррекциям (обратным связям), передающим через внутренние и внешние анализаторы сигналы о характере и эффективности реакций организма (в спорте, например, о перемещениях звеньев тела, траектории перемещающихся снарядов при метаниях).
На основе анализа и синтеза афферентной импульсации (в том числе сенсорных коррекций) и следовых процессов формируются новые рефлекторные акты и целостное поведение организма.
Высшая нервная деятельность имеет важнейшее значение в процессе приобретения новых двигательных навыков и адаптации к различным физическим упражнениям.
Механизмы высшей нервной деятельности у высших животных и человека связаны с деятельностью ряда отделов головного мозга, Основная роль в этих механизмах принадлежит коре больших полушарий (И. П. Павлов). Экспериментально показано, что у высших представителей животного мира после полного оперативного удаления коры высшая нервная деятельность резко ухудшается. Они теряют способность тонко приспосабливаться к внешней среде и самостоятельно существовать в ней.
Отсутствие активных пищедобывательных рефлексов и защитных реакций на дистантные (от лат. distantia — расстояние) раздражители может привести к гибели от голода или биологических врагов.
У человека кора больших полушарий головного мозга выполняет роль «распорядителя и распределителя» всех жизненных функции (И. П. Павлов). Это обусловлено тем, что в ходе филогенетического развития происходит процесс кортикализации функций. Он выражается во все большем подчинении соматических и вегетативных отправлений организма регуляторным влияниям коры мозга. В случае гибели нервных клеток в значительной части коры головного мозга человек оказывается нежизнеспособным и быстро погибает при заметном нарушении гомеостаза важнейших вегетативных функций.
§ 2. Безусловные и условные рефлексы
И. П. Павлов разделил все рефлексы на 2 группы — безусловные и условные рефлексы. Они представляют собой сложные многокомпонентные реакции, включающие деятельность ряда как соматических, так и вегетативных органов. Среди компонентов рефлекса можно выделить главные и дополнительные. Например, в оборонительном рефлексе главным является двигательный компонент, в пищевом — двигательный и секреторный.
Безусловными рефлексами называются врожденные рефлексы. К моменту рождения у животных и человека закладывается основной наследственный фонд таких рефлексов. Но некоторые из них, в частности половые, формируются после рождения, по мере соответствующего морфологического и функционального созревания нервной, эндокринной и других систем.
Безусловные рефлексы обеспечивают первое, грубое приспособление организма к изменениям внешней и внутренней среды. Так, организм новорожденного адаптируется к среде за счет безусловных рефлексов дыхания, сосания, глотания и др.
Безусловные рефлексы отличаются стабильностью, которая обусловливается наличием в центральной нервной системе готовых стойких нервных связей для проведения рефлекторного возбуждения. Эти рефлексы носят видовой характер. Представители одного и того же вида животных имеют примерно одинаковый фонд безусловных рефлексов. Каждый из них проявляется при раздражении определенного рецептивного поля (рефлексогенной зоны). Например, глоточный рефлекс возникает при раздражении задней стенки глотки, рефлекс слюноотделения — при раздражении рецепторов полости рта, коленный, ахиллов, локтевой рефлексы — при раздражении рецепторов сухожилий определенных мышц, зрачковый — при действии на сетчатку резкого изменения освещенности и т. д. При раздражении других рецептивных полей эти реакции не вызываются.
Большинство безусловных рефлексов могут возникать без участия коры больших полушарий и подкорковых узлов. Вместе с тем Центры безусловных рефлексов находятся под контролем коры больших полушарий и подкорковых узлов, которые оказывают субординационное (от лат. sub — подчинение, ordinatio — приведение в порядок) влияние.
К сложным безусловным рефлексам относятся инстинкты. Животным присущи четыре основных вида инстинктов: пищевой, половой, родительский, оборонительный. В отличие от простых безусловных рефлексов, инстинкты обычно регулируются стимулами из внутренней среды организма, в том числе гормональными: голодом,. Жаждой, болью и др. Такого рода эндогенные мотивации (от франц. motif — побудительная причина, повод к действию) стимулируют поисковую и другие виды деятельности животных в среде их обитания и направлены на удовлетворение возникающих побуждений.
Инстинкты весьма прочны. Сложные формы поведения, связанные с их проявлением, нередко свидетельствуют об их высоком приспособительном значении. Например, у щенков из породы охотничьих собак без предварительного обучения в условиях охоты проявляются в поведении многие нюансы, характерные для тренированных животных.
При росте и развитии организма система безусловнорефлекторных связей все же оказывается ограниченной, инертной, неспособной обеспечить достаточно подвижные адаптационные реакции соответствующие колебаниям внешней и внутренней среды. Более совершенная адаптация организма к постоянно изменяющимся условиям существования происходит благодаря условнорефлекторным, т. е. индивидуально приобретенным, реакциям. Условнорефлекторные механизмы головного мозга имеют отношение ко всем видам деятельности организма (к соматическим и вегетативным функциям, к поведению), обеспечивая приспособительные реакции, направленные на сохранение целостности и стабильности системы «организм—среда». И. П. Павлов назвал условный рефлекс временной связью раздражителя с ответной деятельностью, образующейся в организме при определенных условиях. Поэтому в литературе вместо термина «условный рефлекс» часто используется термин «временная связь», который включает и более сложные проявления деятельности животных и человека, представляющие собою целые системы рефлексов и поведенческие акты.
Условные рефлексы не являются врожденными и приобретаются в процессе жизни в результате постоянного общения организма с внешней средой. Они не отличаются столь выраженной стабильностью, как безусловные рефлексы, и исчезают при отсутствии подкрепления. При этих рефлексах ответные реакции могут быть связаны с раздражением самых различных рецептивных полей (рефлексогенных зон). Так, условный пищевой секреторный рефлекс можно выработать и воспроизвести при раздражении разных органов чувств (зрение, слух, обоняние и др.).
§ 3. Механизм и условия образования условных рефлексов
Механизм образования условных рефлексов. Если интенсивность раздражения рецепторов достигает в различных рефлексогенных зонах пороговой или надпороговой силы, в них возникает возбуждение, которое, распространяясь по чувствительным нервам, приходит в центральную нервную систему и вызывает ответную рефлекторную реакцию.
Рефлекторное возбуждение, возникающее в какой-либо рефлексогенной зоне, адресуется из центров чувствительных нервов не всем, а строго определенным эффекторным (двигательным или секреторным) нервным центрам. В безусловных рефлексах эта связь между чувствительными и эффекторными центрами является врожденной.
Условные рефлексы характеризуются тем, что каждый раздражитель (световой, звуковой, проприоцептивный и др.) может при известных условиях приобрести сигнальное значение и стать раздражителем, вызывающим специальную ответную реакцию организма: двигательную, секреторную, пищевую, оборонительную и т. д. Например, если индифферентный раздражитель — звук звонка сочетается с действием пищи на рефлексогенную зону безусловного слюноотделительного рефлекса, то после нескольких сочетаний возбуждение, возникающее в слуховом чувствительном центре, будет распространяться на эфферентные центры слюноотделения (рис. 62). Обстановка спортивных состязаний, многократно сочетаемая с выполнением физических упражнений, также может стать условным раздражителем, т. е. сигналом, вызывающим изменение частоты сердечных сокращений, кровяного давления, интенсивности обмена веществ и т. д. Изменения физиологических функций в организме, наблюдаемые при стартовом состоянии, представляют собой примеры условных рефлексов, образовавшихся при сочетании воздействия обстановки стадиона с выполнением физических упражнений в условиях состязаний.
И. П. Павлов объясняет механизм образования условных рефлексов следующим образом. Если в центральной нервной системе возникают два очага возбуждения, то более сильный из них «притягивает» к себе возбуждение из менее сильного. Если такого рода взаимодействие сильного и слабого очагов возбуждения сочетать повторно несколько раз, может образоваться условный рефлекс. Так, после нескольких сочетаний относительно слабого очага возбуждения в зрительном центре (при действии света) с более сильным очагом возбуждения в пищевом центре (при кормлении) возбуждение из зрительного центра будет распространяться на пищевой центр. В результате действие светового раздражителя будет вызывать пищевую реакцию в виде слюноотделения даже без принятия пищи, т. е. образуется условный рефлекс.
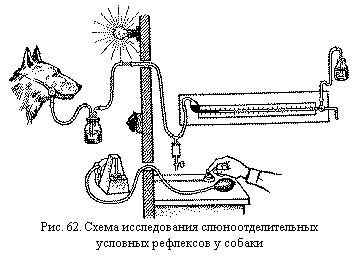 Электрофизиологический
анализ формирования условных рефлексов показал, что для механизмов замыкания
имеет значение пространственная синхронизация. Она заключается в сходстве
протекания (по времени и фазе) биопотенциалов в различных пространственно
удалённых друг от друга констелляциях (от лат constellation
— плеяда, совокупность) нервных клеток (М. Н. Ливанов, Е. Б. Сологуб и др.).
Электрофизиологический
анализ формирования условных рефлексов показал, что для механизмов замыкания
имеет значение пространственная синхронизация. Она заключается в сходстве
протекания (по времени и фазе) биопотенциалов в различных пространственно
удалённых друг от друга констелляциях (от лат constellation
— плеяда, совокупность) нервных клеток (М. Н. Ливанов, Е. Б. Сологуб и др.).
Образование условных рефлексов тесно связано с нервными механизмами памяти.
Как уже отмечалось в главе IV, существует 2 основных вида памяти: кратковременная (краткосрочная) и долговременная (долгосрочная). Информация, поступающая в нервную систему, сначала фиксируется в кратковременной памяти. В дальнейшем она или быстро утрачивается или «передается на хранение» в долговременную память.
Информация, фиксированная в долговременной памяти, может сохраняться дни, недели, месяцы и даже многие годы. Но такая передача информации в долговременную память происходит не всегда. Например, если человеку сообщить незнакомый семизначный номер телефона, то в большинстве случаев он может правильно повторить его. Но через несколько минут (особенно несколько десятков минут) этот номер забывается. Передачу информации из кратковременной памяти в долговременную можно искусственно прекратить путем соответствующих воздействий на центральную нервную систему. Так, если. при начальном формировании условного рефлекса сразу после его выработки дать наркоз, то образовавшийся ранее условный рефлекс исчезает после снятия наркоза. Если же такой наркоз дать через несколько часов или дней после образования условнорефлекторной связи, то рефлекс сохраняется.
Считается, что фиксация информации связана с длительной (на протяжении десятков минут) реверберацией (от лат. reverberare — отражать), т. е. с циркуляцией возбуждения по нейронным цепям. Наркоз прекращает реверберацию. Поэтому если через 5—20 мин. после выработки условного рефлекса дать наркоз, то длительность реверберации оказывается меньше необходимой. Это препятствует фиксации информации в структурах долговременной памяти. Видимо, структуры, связанные с фиксацией кратковременной и долговременной памяти, различны.
В процессах фиксации информации большая роль отводится рибонуклеиновой кислоте (РНК). В опытах на крысах было показано, что применение препаратов, вызывающих снижение содержания РНК в нервной ткани, приводит к значительному снижению эффективности обучения, т. е. выработки условных рефлексов.
У низших животных образование условных рефлексов может осуществляться за счет подкорковых отделов головного мозга -— мозжечка (рыбы), полосатого тел а (птицы) и пр.
У млекопитающих и у человека в формировании условнорефлекторных связей важнейшая роль принадлежит коре больших полушарий. Но при этом связи между условнорефлекторными центрами осуществляются как за счет межкортикальных проводящих путей (т.е. между разными зонами коры больших полушарий головного мозга), так и за счет путей, соединяющих кору с различными под-поковыми образованиями (ретикулярной формацией и др.). Так, после удаления больших полушарий у собак сохраняются и могут образовываться только самые простые условные рефлексы. Они вырабатываются очень медленно, отличаются непрочностью и отсутствием целенаправленности. Последнее проявляется, например, в развитии в ответ на условный сигнал беспорядочной двигательной активности (Э. С. Асратян, М. М. Хананашвили, Н. Ю. Беленков и др).
Основные условия образования условных рефлексов. Условные рефлексы хорошо образуются только при определенных условиях. Главнейшими из них являются: 1) повторное сочетание действия ранее индифферентного условного раздражителя с действием подкрепляющего безусловного или ранее хорошо выработанного условного раздражителя; 2) некоторое предшествование во времени действия индифферентного агента действию подкрепляющего раздражителя; 3) бодрое состояние организма; 4) отсутствие других видов активной деятельности; 5) достаточная степень возбудимости безусловного или хорошо закрепленного условного подкрепляющего раздражителя; 6) надпороговая интенсивность условного раздражителя.
Совпадение действия индифферентного раздражителя с действием подкрепляющего раздражителя (безусловного или хорошо ранее закрепленного условного раздражителя), как правило, должно повторяться несколько раз. При образовании новых условных рефлексов в одной и той же обстановке процесс формирования этих рефлексов ускоряется. Например, у собаки в опытах в камере первый условный рефлекс образуется после 10—20 сочетаний, последующие же — значительно быстрее. У человека многие условные рефлексы, в особенности на словесные раздражители, могут образовываться после одного сочетания.
Длительность времени предшествования действия нового условного раздражителя действию подкрепляющего не должно быть значительным. Так, у собак рефлексы особенно хорошо вырабатываются при длительности этого времени предшествования 5—10 сек. При сочетании в обратном порядке, когда подкрепляющий раздражитель начинает действовать ранее индифферентного, условный рефлекс не вырабатывается.
Образование условнорефлекторных связей, легко протекающее в условиях бодрого состояния организма, затрудняется при его заторможенности. Так, у животных, находящихся в сонливом состоянии, условные рефлексы или не образуются совсем, или же образуются медленно, с трудом. Заторможенное состояние затрудняет формирование условных рефлексов и у человека.
При доминировании в центральной нервной системе центров, не связанных с образованием данных условных рефлексов, формирование этих рефлексов затрудняется. Так, если у собаки возникло резкое возбуждение, например, при виде кошки, то в этих условиях образование пищевого слюноотделительного рефлекса на звук звонка или свет лампочки не происходит. У человека, поглощенного каким-либо делом, образование условных рефлексов на другие виды деятельности в это время также резко затрудняется.
Условные рефлексы образуются только при наличии достаточной возбудимости центров этих подкрепляющих рефлексов. Например, при выработке у собак пищевых условных рефлексов опыты ставятся при условии высокой возбудимости пищевого центра (голодном состоянии животного).
Возникновение и закрепление условнорефлекторной связи происходит при определенном уровне возбуждения нервных центров. В связи с этим сила условного сигнала должна быть достаточной— выше пороговой, но не чрезмерной. На слабые раздражители условные рефлексы не вырабатываются совсем или формируются медленно и отличаются неустойчивостью. Чрезмерно сильные раздражители вызывают развитие в нервных клетках охранительного (запредельного) торможения, что также затрудняет или исключает возможность образования условных рефлексов.
Явления генерализации (обобщения) при образовании условных рефлексов. Характерной особенностью вновь образованных условных рефлексов является их генерализация. Ее сущность заключается в том, что условные раздражители обобщаются и условный рефлекс, образовавшийся на определенный раздражитель, воспроизводится и при действии других, сходных с ним, раздражителей. Процесс возбуждения, возникающий при действии какого-либо одного раздражителя, в результате иррадиации переходит и на корковые центры других, близких по характеру, раздражителей. Например, при образовании условного пищевого слюноотделительного рефлекса на тон 1000 гц условнорефлекторное слюноотделение в первое время после установления временной связи будет вызываться и при действии других тонов: 200 гц, 5000 гц и т. д. В этом случае генерализация выражена при действии близких по частоте тонов больше, чем при действии тонов более отдаленных. Если тоны 700—800 гц дают такой же эффект, как и условный раздражитель, на который вырабатывается рефлекс (1000 гц), то тон 200 гц вызывает значительно меньшее условнорефлекторное слюноотделение.
В двигательных условных рефлексах, например при образовании двигательных навыков, феномен генерализации проявляется в участии в двигательном акте значительного числа мышц, сокращение которых не является необходимым.
Электрофизиологически иррадиация возбуждения выражается в десинхронизации электрокортикограммы (П. К. Анохин) и распространении первичных вызванных ответов по горизонтальным связям коры мозга (А. Б. Коган, Н. Н. Ткаченко).
Явления концентрации при упрочении условных рефлексов. Упрочившиеся условные рефлексы генерализованы в меньшей степени чем только что образованные. Это объясняется тем, что процесс возбуждения при многократном повторении условных раздражении концентрируется и меньше иррадирует на другие центры. Пои этом чем больше упрочивается условный рефлекс, тем меньше становится выраженность явлений и генерализации. Вследствие этого пои упрочившемся условном рефлексе генерализация сказывается лишь на отношении раздражителей, весьма близких друг другу. Например, у собаки, имеющей упроченный условный рефлекс на тон в 1000 гц, генерализация может проявиться при раздражении тоном 900 гц, но ее не будет при раздражении тоном 200—500 гц.
При образовании условных рефлексов у человека явления генерализации выражены несравненно меньше, чем у животных. Процессы концентрации, проявляющиеся у человека в значительно большей степени, чем у животных, дают возможность сразу же устранить условные рефлексы на воздействия, существенно отличающиеся от раздражителя, на который вырабатывается данный условный рефлекс.
§ 4. Сенсорные и оперантные условные рефлексы
Каждый рефлекс содержит афферентные (сенсорные) и эфферентные (исполнительные) компоненты (звенья). В одних случаях формирование новых условных рефлексов относится к образованию только новых сенсорных компонентов, в других же— к образованию и эфферентных, т. е. исполнительных, компонентов рефлекса. Вследствие этого условные рефлексы могут быть двух видов — сенсорные и оперантные (эффекторные).
В сенсорных условных реакциях (названных Ю. Конорским условными рефлексами 1-го рода) ответные акты являются или унаследованными (пищевыми, оборонительными, ориентировочными, половыми и другими безусловными рефлексами), или ранее хорошо закрепленными условными рефлексами (условные рефлексы высших порядков). Следовательно, они характеризуются формированием только афферентной части рефлекса, при которой индифферентный раздражитель превращается в активный. Ответная же реакция на условный раздражитель остается той же, что и при безусловном или ранее хорошо выработанном условном раздражителе. Например, при выработке оборонительного условного рефлекса на запах в центральной нервной системе устанавливаются связи между афферентными клетками, воспринимающими раздражение обонятельного анализатора, и болевым центром. При этом характер ответной условной и безусловной реакций совпадает. В обоих случаях начинается слюноотделение. Аналогично вырабатываются и проявляются другие сенсорные условные реакции (в частности, оборонительные двигательные условные рефлексы в виде отдергивания руки, подкрепляемые болевым раздражением, сухожильные, зрачковые, мигательные рефлексы).
Условные рефлексы этого рода не всегда могут в достаточной мере изменить взаимоотношения организма с окружающей средой и полноценно обеспечить адекватное приспособление, так как при этом не организуются новые формы самих ответных реакций Более адекватное приспособление обеспечивается тем, что животные и человек способны кардинально изменять характер своих эффекторных реакций при взаимоотношениях с окружающей средой
Оперантные условные рефлексы (по классификации Ю. Конорского, рефлексы 2-го рода) характеризуются новой (не унаследованной от предков или не имевшейся в ранее индивидуально при. обретенном фонде) формой ответной реакции. Эти рефлексы называются также «инструментальными», так как при их осуществлении используются разные предметы (инструменты). Например, животное конечностью открывает задвижку на дверце и достает находящуюся за ней пищу. Поскольку при формировании таких рефлексов создается сложный комплекс вновь образованных движений, эти рефлексы называют также «манипуляционными».
В образовании соответствующих условных оперантных рефлексов важнейшая роль принадлежит импульсам, поступающим из двигательного аппарата. Элементарная модель условного оперантного рефлекса наблюдалась при. выработке классического пищевого условного рефлекса на пассивное сгибание лапы у собаки (Ю. Конорский). У животного было выявлено два типа условных реакций — условнорефлекторное слюноотделение в ответ на пассивное сгибание лапы (сенсорный условный рефлекс, или рефлекс 1-го рода) и многократное активное сгибание конечности, являвшееся не только сигналом, но и способом получения пищи (оперантный условный рефлекс, или рефлекс 2-го рода).
Одним из видов оперантных рефлексов являются опыты с самораздражением мозговых структур, при котором получается «чувство удовлетворения потребностей» (см. главу IV, § 10).
При формировании оперантных условных реакций важнейшая роль принадлежит обратной связи между клетками в нервных центрах безусловных или ранее хорошо выработанных условных рефлексов и клетками центров двигательного анализатора. Этому способствует высокий уровень возбудимости двигательных центров, обусловленный потоком афферентных импульсов от проприорецепторов сокращающихся мышц.
Таким образом, одним из условий выработки оперантных моторных условных рефлексов является обязательное включение в систему раздражителей импульсации, возникшей спонтанно или вызванной активным или пассивным движением. Образование этих рефлексов зависит от характера подкрепляющего агента. Оперантные условные рефлексы составляют основу двигательных навыков. Закреплению их способствуют обратные связи, осуществляемые через проприорецепторы мышц, выполняющих движение, и через рецепторы ряда других анализаторов. Благодаря этой афферентации центральная нервная система сигнализирует о результатах выполнения движения.
Формирование новых движений, т. е. не унаследованных от предков, наблюдается не только у человека, но и у животных. Но для человека этот процесс имеет особенно большое значение, так как почти все сложные двигательные акты (в частности, физические упражнения в различных видах спорта) формируются именно в результате обучения.
§ 5. Виды условных рефлексов
В зависимости от особенностей ответных реакций, характера раздражителей, условий их применения и подкрепления и т. д. выделяют различные виды условных рефлексов. Эти виды классифицируются, исходя из различных критериев, в соответствии с поставленными задачами. Некоторые из этих классификаций имеют большое значение как в теоретическом, так и в практическом отношении, в том числе при спортивной деятельности.
Натуральные (естественные) и искусственные условные рефлексы. Условные рефлексы, образующиеся на действие сигналов, характеризующих постоянные свойства безусловных раздражителей (например, запах или вид пищи), называются натуральными условными рефлексами.
Иллюстрацией закономерностей образования натуральных условных рефлексов являются опыты И. С. Цитовича. В этих опытах щенки одного и того же помета содержались на различном пищевом рационе: одних кормили только мясом, других — только молоком. У животных, которых кормили мясом вид и запах его уже на расстоянии вызывали условную пищевую реакцию с ярко выраженными моторным и секреторным компонентами. Щенки, получавшие лишь молоко, первый раз реагировали на мясо только ориентировочной реакцией (т. е., по образному выражению И. П. Павлова, рефлексом «Что такое?») —обнюхивали его и отворачивались. Однако уже однократное сочетание вида и запаха мяса с едой полностью устраняло это «безразличие». У щенков выработался натуральный пищевой условный рефлекс. Образование натуральных (естественных) условных рефлексов на вид, запах пищи и свойства Других безусловных раздражителей характерно и для человека. Натуральные условные рефлексы характеризуются быстрой выработкой и большой стойкостью. Они могут удерживаться всю Жизнь при отсутствии последующих подкреплений. Это объясняется тем, что натуральные условные рефлексы имеют большое биологическое значение, особенно на ранних этапах приспособления организма к окружающей среде. Именно свойства самого безусловного раздражителя (например, вид и запах пищи) являются теми первыми сигналами, которые действуют на организм после рождения.
Но условные рефлексы можно выработать также на различные индифферентные сигналы (свет, звук, запах, изменения температуры и др.), не обладающие в естественных условиях свойствами раздражителя, вызывающего безусловный рефлекс. Такого рода реакции, в отличие от натуральных, называются искусственными условными рефлексами. Например, запах мяты не присущ мясу. Однако если этот запах несколько раз сочетать с подкармливанием мясом то образуется условный рефлекс: запах мяты становится условным сигналом еды и начинает вызывать слюноотделительную реакцию без подкрепления. Искусственные условные рефлексы медленнее вырабатываются и быстрее угасают при неподкреплении.
Примером выработки условных рефлексов на искусственные раздражители может быть образование у человека секреторных и двигательных условных рефлексов на сигналы в виде звучания звонка, ударов метронома, усиления или ослабления освещения прикосновения к коже и т. д.
Условные рефлексы первого и высших порядков. Реакции, образуемые на основе безусловных рефлексов, называются условными рефлексами первого порядка, а реакции, вырабатываемые на основе ранее приобретенных условных рефлексов,—условными рефлексами высших порядков (второго, третьего и т. д.). При выработке условных рефлексов высших порядков индифферентный сигнал подкрепляется хорошо упроченными условными раздражителями. Если, например, раздражение в виде звонка подкреплять едой (безусловная реакция), то вырабатывается условный рефлекс первого порядка. После упрочения условного рефлекса первого порядка можно выработать на его основе условный рефлекс второго порядка, в частности на свет. На базе условного рефлекса второго порядка можно образовать условный рефлекс третьего порядка, на базе рефлекса третьего порядка.—рефлекс четвертого порядка и т. д.
 Образование
условных рефлексов высших порядков зависит от совершенства организации нервной
системы, ее функциональных свойств и биологической значимости безусловного
рефлекса, на базе которого выработан условный рефлекс первого порядка.
Например, у собак в искусственных условиях на фоне повышенной пищевой
возбудимости можно выработать слюноотделительный условный рефлекс третьего
порядка. В случае двигательно-оборонительной реакции у этих же животных
возможно образование условных рефлексов четвертого порядка. У обезьян, стоящих
на более высокой ступени филогенетической лестницы, условные рефлексы высших
порядков образуются легче, чем у собак. Для человека процесс образования
условных рефлексов, высших порядков оказывается наиболее адекватным. При
наличии повышенной возбудимости центральной нервной системы даже у детей в
возрасте до года вырабатываются условные рефлексы пятого и шестого порядка (Н.
И. Красногорский). При развитии функции речи порядковый диапазон этих реакций
значительно расширяется. Так, подавляющее большинство двигательных условных
рефлексов у человека образуется путем подкрепления не каким-либо безусловным
раздражителем, а различными условными сигналами в виде словесных инструкций,
объяснений и др.
Образование
условных рефлексов высших порядков зависит от совершенства организации нервной
системы, ее функциональных свойств и биологической значимости безусловного
рефлекса, на базе которого выработан условный рефлекс первого порядка.
Например, у собак в искусственных условиях на фоне повышенной пищевой
возбудимости можно выработать слюноотделительный условный рефлекс третьего
порядка. В случае двигательно-оборонительной реакции у этих же животных
возможно образование условных рефлексов четвертого порядка. У обезьян, стоящих
на более высокой ступени филогенетической лестницы, условные рефлексы высших
порядков образуются легче, чем у собак. Для человека процесс образования
условных рефлексов, высших порядков оказывается наиболее адекватным. При
наличии повышенной возбудимости центральной нервной системы даже у детей в
возрасте до года вырабатываются условные рефлексы пятого и шестого порядка (Н.
И. Красногорский). При развитии функции речи порядковый диапазон этих реакций
значительно расширяется. Так, подавляющее большинство двигательных условных
рефлексов у человека образуется путем подкрепления не каким-либо безусловным
раздражителем, а различными условными сигналами в виде словесных инструкций,
объяснений и др.
Биологическое значение условных рефлексов высших порядков состоит в том, что они обеспечивают сигнализацию о предстоящей деятельности при подкреплении не только безусловными, но и условными раздражителями. В связи с этим более быстро и полно происходит развертывание адаптационных реакций организма.
Положительные и отрицательные условные рефлексы. Условные рефлексы, в динамике которых проявляется активность организма в виде двигательных или секреторных реакций, называются положительными. Условные реакции, не сопровождающиеся внешним двигательным и секреторным эффектами в связи с их угнетением, относят к отрицательным, или тормозным, рефлексам. В процессе приспособления организма к изменяющимся условиям среды оба вида рефлексов имеют большое значение. Они тесно взаимосвязаны, так как проявление одного вида деятельности сочетается с угнетением других видов. Например, при оборонительных двигательных условных рефлексах тормозятся условные пищевые реакции и наоборот. При условном раздражителе в виде команды «Смирно!» вызывается деятельность мышц, обусловливающих стояние в определенном положении и торможение других условных двигательных реакций, которые осуществлялись до этой команды (например, ходьба, бег).
Такое важное качество, как дисциплинированность, всегда связано с одновременным сочетанием положительных и отрицательных (тормозных) условных рефлексов. Например, при выполнении некоторых физических упражнений (прыжки в воду с вышки, гимнастическое сальто и др.) для подавления реакций самосохранения и чувства страха требуется торможение сильнейших отрицательных оборонительных условных рефлексов.
Наличные и следовые рефлексы. Условные рефлексы, в которых условный сигнал предшествует безусловному раздражителю, действует совместно с ним и заканчивается одновременно или на несколько секунд раньше или позднее прекращения безусловного раздражения, называются наличными (рис. 63). Как уже отмечалось, для образования условного рефлекса необходимо, чтобы условный сигнал начал действовать раньше начала действия подкрепляющего Раздражителя. Интервал между ними, т. е. степень отставления подкрепляющего раздражителя от условного сигнала, может быть различным. В зависимости от длительности отставления безусловно подкрепления от начала действия условного сигнала наличные Условные рефлексы у животных, например пищевые, классифицируются как совпадающие (0,5 – 1 сек.), короткоотставленные (3 – 5 сек.), нормальные (10 – 30 сек.) и запаздывающие (1 – 5 мин. и более).
При следовых условных рефлексах условный раздражитель подкрепляется после прекращения его действия (см. рис. 63) Между затухающим очагом возбуждения в коре от индифферентного агента и очагом возбуждения в корковом представительстве подкрепляющего безусловного или ранее хорошо выработанного рефлекса образуется временная связь. Следовые условные рефлексы формируются при коротком (10—20 сек.) и длинном (позднем) отставлениях (1—2 мин. и более). К группе следовых условных рефлексов относится, в частности, рефлекс на время, выполняющий роль так называемых «биологических часов».
Наличные и следовые условные рефлексы с большим отставлением являются сложными формами проявления высшей нервной деятельности и доступны лишь животным с достаточно развитой корой головного мозга. Выработка подобных рефлексов у собак связана с большими трудностями. У человека следовые условные рефлексы образуются легко.
Следовые условные реакции имеют большое значение при физических упражнениях. Например, в гимнастической комбинации, состоящей из нескольких элементов, следовое возбуждение в коре мозга, вызванное действием первой фазы движения, служит раздражителем для программирования цепи всех последующих. Внутри цепной реакции каждый из элементов является условным сигналом для перехода к следующей фазе движения.
Экстероцептивные, проприоцептивные и интероцептивные рефлексы. В зависимости от анализатора, на основе которого выработаны условные рефлексы, последние делятся на три вида. Реакции, выработанные при раздражении внешних анализаторов (зрительного, слухового и др.), называются экстероцептивными, а выработанные при раздражении рецепторов мышц — проприоцептивными, рецепторов внутренних органов — интероцептивными.
Главным средством связи организма с внешней средой являются экстеро- и проприоцептивные условные рефлексы. Реакции, имеющие большую биологическую значимость, вырабатываются быстрее и лучше дифференцируются. В то же время они достаточно динамичны и могут угасать при изменении сигнального значения раздражителей и их неподкреплении.
Интероцептивные условные рефлексы вырабатываются и дифференцируются значительно медленнее, отличаются большой инертностью, не угасая при неподкреплениях в течение длительного времени. Афферентная импульсация от интерорецепторов может многократно совпадать по времени с осуществлением ответных соматических и вегетативных реакций, возникающих при воздействии на организм определенных сигналов внешней среды. При этом интероцептивные раздражители приобретают для соответствующих реакций сигнальное значение. В целом интероцептивные раздражения стимулируют координирующее влияние нервных центров, в особенности коры больших полушарий, на взаимодействие факторов внутренней и внешней среды, в результате чего развиваются тонкие условнорефлекторные приспособительные реакции. При мышечной деятельности интенсивность проявления вегетативных функций повышается (кровообращение, дыхание и др.). Импульсация со стороны интерорецепторов в центральную нервную систему заметно усиливается, в связи с чем создаются более благоприятные условия для образования интероцептивных условных рефлексов. Определенный характер изменения вегетативных функций в процессе выполнения спортивной работы может сочетаться по механизму условных рефлексов с конкретной двигательной деятельностью и тем самым способствовать более эффективному ее выполнению.
Условные рефлексы на комплексные раздражители. Образование условных рефлексов может происходить при действии не только одиночных, но и комплексных раздражителей, относящихся к одной или разным сенсорным системам. Комплексные раздражители могут действовать одновременно и последовательно. При комплексе одновременно действующих раздражителей поступают сигналы от нескольких раздражителей. Например, условный пищевой рефлёкс может вызываться одновременным воздействием запаха, формы и цвета раздражителя. При комплексе последовательно действующих раздражителей первый из них, например световой, сменяется вторым, например звуковым (в виде высокого тона), затем третьим, например звуком метронома. Подкрепление же следует только после действия всего этого комплекса.
§ 6. Первая и вторая сигнальные системы
Эти представления не выдержали испытания временем.
Взаимоотношения организма со средой осуществляются на основе сигналов, поступающих в нервную систему в результате непосредственного воздействия предметов и явлений внешнего мира на рецепторы. Этот тип сигнализации И. П. Павлов назвал первой сигнальной системой. В животном мире первая сигнальная система является единственным каналом информации организма о состоянии среды. Различные предметы внешнего мира, их физические и химические свойства (звук, цвет, форма, химический состав и др.) приобретают значение условных сигналов, оповещают организм о наступающих вслед за ними явлениях, вызывая тем самым приспособительные реакции. Например, дремлющее травоядное животное при звуке шагов или запахе хищника спасается бегством, так как эти раздражители сигнализируют об опасности.
Первая сигнальная система высших животных обеспечивает достаточно совершенное отражение внешнего мира и в связи с этим гонкое и точное приспособление их к среде. И. П. Павлов рассматривал первую сигнальную систему как систему восприятия, впечатлений от всех воздействий внешнего и внутреннего мира, сигнализирующих о биологически полезных или вредных для организма раздражителях. Он писал: «Для животного действительность сигнализируется почти исключительно только раздражениями и следами их в больших полушариях, непосредственно приходящими специальные клетки зрительных, слуховых и других рецепторов организма. Это то, что мы имеем в себе как впечатления, ощущения и представления от окружающей внешней среды, как общеприродной так и нашей социальной, исключая слово, слышимое и видимое. Это первая сигнальная система действительности, общая у нас с животными».
Сигналы первой сигнальной системы являются конкретными и относятся к определенному предмету.
Образование условных рефлексов через первую сигнальную систему составляет у высших животных физиологическую основу их элементарного конкретного, или предметного, мышления. Первая сигнальная система — одинаковая у человека и животных. В условиях обычной жизни у человека она изолированно функционирует только в первые шесть месяцев жизни.
При воспитании человека развивается вторая сигнальная система, характерная только для человека. Это переводит высшую нервную деятельность человека на более высокую ступень. Она приобретает новые качества, обусловливающие расширение возможностей общения с внешним миром и многогранность его проявлений. И. П. Павлов назвал вторую сигнальную систему «чрезвычайной прибавкой» к механизмам высшей нервной деятельности человека. Вторая сигнальная система — это речь, слово, видимое, слышимое, произносимое мысленно. Это высшая система сигнализации окружающего мира. Она состоит в словесном обозначении всех его сигналов и в речевом общении. Вторая сигнальная система развилась у человека под влиянием социальной среды в процессе труда. Большую роль в этом сыграли кинестезические раздражения мозга, возникающие в результате трудовых процессов. Слово для человека служит таким же физиологическим раздражителем, как предметы и явления окружающего мира.
Словесные сигналы обобщают раздражители первой сигнальной системы. Одно и то же слово «стол» сигнализирует не только об определенном столе, но и о многих других столах, разных по величине, форме, цвету и др. В этом факте выражено не только обобщение, но и отвлечение от конкретных предметов действительности, т. е. переход человека от предметного мышления к абстрактному. Для того чтобы слово «стол» указывало на конкретный стол, необходимо уточнение—«этот стол». В пределах второй сигнальной системы обобщаются раздражители не только первой, но и самой второй сигнальной системы. Например, слово узкого значения «осина» обобщает конкретные раздражители первой сигнальной системы, а слово более широкого значения «дерево» — раздражители второй сигнальной системы.
Таким образом, вторая сигнальная система является всеобъемлющей, способной заменить, абстрагировать и обобщить все раздражители первой сигнальной системы. Благодаря всей предшествующей жизни взрослого человека слово связано со всеми внешними и внутренними раздражениями, приходящими в нервную систему, обо всех них сигнализирует и все их заменяет, вызывая такие же действия, как они.
Другое исключительно важное значение второй сигнальной системы заключается в том, что она резко увеличивает объем информации — за счет использования не только индивидуального, но и коллективного опыта всего человечества. Получаемая человеком от других лиц словесная информация — устная и в особенности письменная — имеет чрезвычайно широкий диапазон (это может быть информация не только от ныне живущих лиц, но и от многих предшествующих поколений). Так, совершенствование спортсмена лишь частично происходит за счет личного его опыта, через словесную информацию он широко использует опыт своего тренера и огромного числа других лиц, изложенный в методических пособиях, учебниках, статьях и т. д.
Первая и вторая сигнальные системы функционально взаимосвязаны. Сигналы первой сигнальной системы, поступающие из различных частей тела и окружающей среды, непрерывно взаимодействуют с сигналами второй сигнальной системы. При этом образуются условные рефлексы второго и высших порядков, функционально связывающие сигнальные системы в единое целое. Кроме того, связь между двумя сигнальными системами, основанная на элективной (избирательной) иррадиации возбуждения, позволяет воспроизводить через вторую сигнальную систему условные рефлексы, выработанные на основе первой сигнальной системы (А. Г. Иванов-Смоленский).
Вторая сигнальная система составляет физиологическую основу абстрактного речевого мышления, присущего только человеку. Афферентные сигналы, поступающие в центральную нервную систему от речевых органов, через слуховой и зрительный анализаторы, формируют у человека сложные рефлексы, обусловливающие звуковую и письменную речь.
Локализация функций второй сигнальной системы в коре головного мозга выяснена еще не полностью. В ее осуществлении участвуют структуры правого и левого полушарий. Доминирующая роль при этом у большинства людей (правшей) принадлежит левому полушарию. Сравнительно обширные его участки осуществляют сложные функции, связанные с пониманием смысла слов, координацией речедвигательного аппарата при их произнесении и другими процессами.
§ 7. Значение торможения в условнорефлекторной деятельности и его виды
Значение торможения в условнорефлекторной деятельности. Наряду с процессами возбуждения существенная роль (как при безусловнорефлекторной, так и при условнорефлекторной деятельности) принадлежит процессам торможения, т. е. угнетения, подавления деятельного состояния. Возбуждение и торможение, непрерывно взаимодействуя, обусловливают возможность осуществления одних реакций и задержки других (нарушающих координацию безусловно- или условнорефлекторной ответной реакции или не являющихся необходимыми). Следовательно, образование любого условного рефлекса в виде координированного ответного акта требует возбуждения одних корковых нервных центров и торможения других.
Виды торможения. Наблюдаемые при условнорефлекторной деятельности виды торможения можно разделить на две основные группы — внешнее торможение и внутреннее.
Внешнее торможение условных рефлексов. Внешнее торможение по своей природе является врожденным видом торможения, присущим всем отделам центральной нервной системы и свойственным как безусловным, так и условным рефлексам. Оно возникает «с места» — без предварительной выработки. К этому виду торможения относятся внешнее торможение и запредельное. Первое из них основано на взаимодействии центров условного рефлекса и постороннего раздражителя. Например, если в момент действия условного сигнала начинает действовать сильный посторонний раздражитель, то условная реакция тормозится. Сильное возбуждение, возникающее в центре постороннего раздражителя, по закону индукции, резко снижает возбудимость корковых центров данного условного рефлекса.
Внешнее торможение постоянно встречается в практике физической культуры и спорта. Например, условнорефлекторные связи, обеспечивающие в обычной обстановке на тренировочном занятии отличное выполнение гимнастических комбинаций, могут нарушаться под влиянием сильных раздражителей (соревнование, присутствие посторонних лиц, резкие замечания тренера и др.), действующих как внешний тормоз. При повторных воздействиях внешнее торможение угасает или проявляется незначительно. Из практики спорта известно, что прочно закрепившиеся условные рефлексы меньше нарушаются под влиянием внешнего торможения. Степень выраженности внешнего торможения определяется функциональным состоянием и типом нервной системы. У лиц с уравновешенным типом нервной системы внешнее торможение при одних и тех же условиях выражено слабее, чем у представителей неуравновешенного типа или у имеющих функциональные расстройства нервной деятельности.
Запредельное (охранительное) торможение, развивающееся при действии сверхсильных или обычных, но длительно продолжающихся раздражении, представляет собой защитный механизм, предотвращающий нервные клетки от истощения и способствующий восстановлению :их химического состава и нормального функционального состояния. По своей физиологической природе охранительное торможение может вызываться факторами, связанными как с безусловно- так и с условнорефлекторной деятельностью. Травмы, редкие и длительные раздражения анализаторов и др. способны вызывать охранительное торможение.
Необходимо указать, что внешнее торможение значительно сильнее сказывается на относительно мало закрепленных условных рефлексах. Из спортивной практики известно, что чем прочнее в провесе тренировки закреплены двигательные условные рефлексы, тем меньше они нарушаются при наличии внешнего торможения.
Внутреннее торможение условных рефлексов. Если торможение условных рефлексов первично возникает в их центрах то такого рода торможение называют внутренним. Существует несколько видов внутреннего торможения, к которым относятся: угасательное, дифференцировочное и торможение при запаздывании подкрепления условного раздражения. Внутреннее торможение, постепенно захватывающее всю кору больших полушарий, приводит к развитию сонного состояния.
Угасательное торможение. Если раздражители, вызывающие условный рефлекс, не подкреплять действием безусловных рефлексов, то они постепенно угасают (табл. 5).
Таблица 5
Угашение у собаки условного рефлекса на вид мясного порошка при отсутствии подкрепления едой (по И. П. Павлову)
|
Время |
Род раздражителя, действующего в течение 1 мин. |
Колич. выделившейся слюны на условный раздражитель (мл) |
|
11 час. 48 мин. 11 час. 51 мин. 11 час. 54 мин. 11 час. 56 мин. 12 час. 00 мин. 12 час. 03 мин. 12 час. 06 мин. 12 час. 09 мин. |
Вид мясного порошка -----"------- -----"------- -----"------- -----"------- -----"------- -----"------- -----"------- |
0,8 0,7 0,5 0,3 0,2 0,1 0.0 0,0 |
Угашение условных рефлексов является не полным разрывом установившихся временных связей, а лишь их торможением. Во-первых, после перерыва (например, на следующий день) угашенный рефлекс самостоятельно постепенно восстанавливается. Лишь при многократном повторном неподкреплении условного раздражителя безусловным рефлексом угашение становится стойким. Во-вторых, угашенный рефлекс сразу же восстанавливается, если во время действия угашенного условного раздражителя внезапно воздействовать каким-либо сильным посторонним раздражителем. Это явление, связанное с устранением торможения условных рефлексов (например, в результате угашения), называется растормаживанием.
Если после угашения восстанавливать временные связи, подкрепляя условные раздражители безусловным рефлексом, то при повторных опытах без подкрепления условные рефлексы будут угашаться все быстрее и быстрее.
У человека при воздействии через вторую сигнальную систем угашение ряда условных рефлексов или их компонентов может происходить очень быстро, даже после однократной инструкции тренера. Однако это не относится к прочно зафиксированным двигательным условным рефлексам. В таких случаях для получения желаемого эффекта при переучивании, связанном с угашением старых и выработкой новых условных рефлексов, требуется длительная тренировка.
Угашение условных рефлексов при неподкреплении соответствующими безусловными или условными (при рефлексах высших порядков) раздражителями имеет большое биологическое значение так как при этом устраняются именно те условные раздражители' которые потеряли свое сигнальное значение для приспособления к окружающей среде.
Дифференцировочное торможение. Приспособление организма к условиям окружающей среды характеризуется тончайшим соответствием ответных реакций особенностям раздражения. Это соответствие приобретается благодаря возможности осуществлять высокую степень дифференцирования (различения) условных раздражителей внешней среды.
Условные рефлексы вначале образуются, как уже указывалось, в генерализованной форме. В дальнейшем происходит процесс дифференцирования, т. е. специализирования, условных раздражителей, который связан с торможением реакции на раздражители, не подкрепляемые безусловными или ранее выработанными условными рефлексами. Например, при образовании условного рефлекса на тон 1000 гц при первой пробе рефлекс будет обнаруживаться и на высокие (2500 гц) и более низкие (500 гц) тоны. Но если тон 2500 и 500 гц не подкреплять, рефлекс на них постепенно угаснет.
Дифференцирование простых раздражителей у животных может быть не только не хуже, но даже лучше, чем у человека. Например, собаки прекрасно различают запахи, тонкие оттенки серого цвета, разницу в высоте различных тонов и пр., которые совершенно недоступны человеку. Однако по способности к образованию дифференцировок при сложных комплексных раздражениях человек превосходит животных. Специальные опыты показали, что если, например, для человека дифференцирование таких двух комплексов, как «шипение — высокий тон — низкий тон — звонок» и «шипение — низкий тон — высокий тон — звонок» не представляет никаких трудностей (различение наблюдается после двух-пяти сочетаний), то для собак такая дифференцировка является уже предельной и для образования ее требуются многие месяцы и даже годы работы.
Сложнейшие дифференцировки образуются у человека при действии сигналов второй сигнальной системы. Он неодинаково реагирует не только на различные слова, но и на интонации, с которыми эти слова сказаны, на место того или иного слова в предложении и др.
Сложные процессы дифференцирования происходят при образовании двигательных условных рефлексов. Например, при обучении различным физическим упражнениям руководитель подкрепляет правильное выполнение двигательного акта и не подкрепляет неправильное Такого рода словесные подкрепления способствуют дифференцированию правильного выполнения упражнения и неправильного.
Скорость образования дифференцировок зависит от степени филогенетического и онтогенетического развития организма, индивидуальных его особенностей, функционального значения раздражений которые необходимо дифференцировать, и от других факторов. У животных, стоящих на низком уровне филогенетического развития сложные дифференцировки образуются, как правило, с большим трудом, чем у животных с более высокой организацией нервной системы. У человека в раннем детском возрасте дифференцировки также вырабатываются труднее, чем в более позднем. Большое значение имеет и функциональное состояние нервной системы. Например, при резком утомлении, при заболевании дифференцировки вырабатываются у человека труднее, ранее же выработанные могут несколько нарушаться.
Раздражители, действующие на различные анализаторы (например, свет и звук, запах и прикосновение), дифференцируются «с места», т. е. сразу же при первом применении. Раздражители того же анализатора, но резко отличающиеся друг от друга по своему характеру (например, тон 3000 гц и шум), также дифференцируются «с места». Раздражители, сходные по характеру (например, тон 1000 и 2000 гц), требуют для дифференцирования нескольких сочетаний, более же близкие по характеру (например, тон 1000 и 1100 гц) будут различаться только после значительного числа сочетаний.
В отдельных случаях дифференцирование сложных и вместе с тем близких по характеру раздражении требует огромного труда, особенно в процессе выработки дифференцировок при образовании двигательных рефлексов. Например, для образования дифференцировок в речедвигательных условных рефлексах, позволяющих говорить без акцента на иностранном языке, нужна большая практика. Она необходима и при обучении многим видам физических упражнений, когда определенный характер движений, необходимый для Достижения максимального результата, нужно дифференцировать с Другими (близкими по форме, но менее эффективными) движениями.
При наличии внешнего торможения дифференцирование условных раздражителей ухудшается. Поэтому, например, нарушение правильности движений при выполнении физических упражнений в момент действия внешнего торможения объясняется расстройством не только положительных условных рефлексов, но и дифференцировок.
Особым видом дифференцировочного торможения является так называемый условный тормоз. Если новый раздражитель, действующий непосредственно перед хорошо выработанным условным раздражителем, не будет подкрепляться безусловным рефлексом, то он превращается в условный тормоз. Например, после выработки слюноотделительного условного рефлекса на звук метронома к нему присоединяют какой-либо новый раздражитель — свет, непосредственно после которого начинает звучать метроном. Если действие метронома всегда подкреплять введением в рот кислоты, а при действии последовательной комбинации «свет — метроном» не давать такого подкрепления, то свет станет условным тормозом, к этом случае действие метронома станет вызывать условный рефлекс. Если же перед, этим будет действовать свет («свет + метроном») то рефлекс окажется заторможенным.
Условное торможение часто проявляется при выполнении физических упражнений, например в спортивных играх (запрещение некоторых действий в определенных зонах игры).
Торможение при запаздывании подкрепления условного раздражения. Торможение может развиваться при выработанных условных рефлексах с большим отставлением от начала действия условного раздражителя до подкрепления безусловным рефлексом. Удлинение скрытого периода условного рефлекса при длительном оставлении подкрепления безусловным или ранее хорошо выработанным условным раздражителем основано на развитии торможения в первый период действия условного раздражителя.
Торможение при запаздывании подкрепления условного рефлекса возникает не сразу, а вырабатывается постепенно. При этом, чем сильнее условный раздражитель, тем с большим трудом вырабатывается этот вид торможения. Существенное значение имеет и сила подкрепляющего раздражителя. Например, при повышении после голодания возбудимости пищевого центра происходит растормаживание и скрытый период условных рефлексов резко уменьшается.
У человека многие условные рефлексы имеют запаздывающий характер. Кратковременные явления запаздывания наблюдаются, например, при строевых командах. Подготовительная часть команд служит условным раздражителем, связанным с развитием запаздывания, а исполнительная часть—условным раздражителем, требующим немедленного исполнения.
§ 8. Динамический стереотип
Различные условные рефлексы постоянно взаимодействуют друг с другом. Если раздражители повторяются в определенном порядке, то между ними формируется взаимосвязь, характеризующаяся стереотипной последовательностью возникновения ответных реакций. При этом рефлексы соответствуют не столько данному раздражителю, сколько месту раздражителя в последовательной цепи их.
Стереотип внешних проявлений реакций в виде секреции или движения был назван И. П. Павловым динамическим стереотипом или функциональной системностью. Термин «динамический» подчеркивает функциональный характер этого стереотипа (формирование и закрепление его только после соответствующих упражнений, возможность его переделки, угасание при длительных перерывах, ухудшение при утомлении, сильных эмоциях, заболеваниях и пр.). Динамический стереотип в отношении сенсорных условных рефлексов ярко проявляется при пищевых рефлексах. Например, если в опытах на собаке применять в течение длительного времени систему положительных и отрицательных условных раздражителей, чередуя их через определенные интервалы времени в строгой последовательности, то это фиксируется нервной системой. Каждому раздражителю, примененному на строго фиксированном для него в стереотипе месте, в соответствии с законом силы раздражителя, соответствует определенная величина ответной реакции (Э. А. Асратян, П С. Купалов и др.). Стереотип может выявляться при замене всех условных раздражителей одним из них, обычно средней силы. Этот раздражитель, примененный в стереотипе вместо других условных раздражителей этого стереотипа, вызывает ответы, величина которых соответствует ответам на замененные раздражители. На месте же отрицательного раздражителя положительный сигнал вызывает очень слабую условную (в частности, слюноотделительную) реакцию.
В двигательной деятельности спортсмена стереотип проявляется, например, в последовательности фаз сложных гимнастических, тяжелоатлетических и других стандартно выполняемых движений.
Переделка стереотипа в некоторых случаях представляет для нервной системы трудный процесс. Чтобы выработать новый стереотип, необходимо сначала угасить старый. Но хорошо закрепленный стереотип трудно подается угашению и может проявляться вновь при возникновении условий, которым он соответствовал.
Динамический стереотип может быть связан не только с отдельными вегетативными или двигательными функциями, но и с целостней деятельностью организма, режимом жизни человека. Формирование таких динамических стереотипов имеет большое значение для человека. Социальная среда, воздействующая на него,— быт, учеба, работа, как правило, в течение более или менее длительного времени остается относительно постоянной (домашний и рабочий режимы, их темп и др.). За счет следового возбуждения клеток в нервных центрах стереотип запечатлевается в них в виде сложной функциональной системы, в которой все воздействующие компоненты среды сливаются в единый синтетический комплекс. Таким образом, стереотип можно характеризовать как систему условных рефлексов на совокупность раздражителей естественной среды. Системность облегчает деятельность. Человек, привыкший изо дня в день делать одну и ту же работу, обычно выполняет ее с большей легкостью.
Однако образование прочного динамического стереотипа может наряду с положительным значением иметь и отрицательное. Привычка действовать по. определенному стандарту затрудняет приспособление к новым условиям выполнения работы, к новому режиму жизни. В некоторых случаях при изменении ситуации, прочный динамический стереотип задерживает приспособление организма к реакциям, более соответствующим новым условиям труда .и быта. Изменение привычных форм работы, режима жизни переживается тяжело и может привести к нарушениям некоторых функций организма, особенно у лиц пожилого возраста. Поэтому, как указывал И. П. Павлов, установление динамического стереотипа является положительным при стандартных условиях деятельности и отрицательным при варьировании этих условий и резком их изменении. Это относится и к физическим упражнениям. Для стандартно выполняемых движений установление стереотипа полезно, для изменяющихся (спортивные игры, единоборство) — нежелательно.
§ 9. Экстраполяция в высшей нервной деятельности
В организме в течение жизни формируется огромное количество различных сенсорных и оперантных условных рефлексов, образующихся в результате действия многочисленных конкретных раздражителей и их комплексов. Вместе с тем число их все же весьма мало по сравнению с огромным разнообразием тех стимулов, которые в процессе жизни воздействуют на организм и требуют экстренных соответствующих обстановке ответных реакций.
Адекватное реагирование организма на возникающие новые раздражения «с места», т. е. без предварительной выработки данного конкретного условного рефлекса, обеспечивается способностью нервной системы к экстраполяции. Она обусловлена способностью нервной системы в новых, в том числе ранее не встречавшихся, условиях правильно реагировать на основании предшествующего наследственного или индивидуально приобретенного опыта и эффективно решать возникающие задачи.
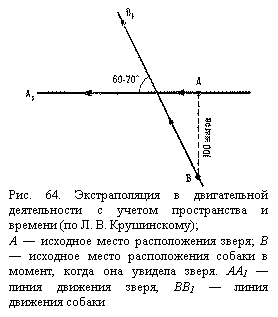 Экстраполяция
наблюдается как при условнорефлекторной, так и при безусловнорефлекторной
деятельности, причем в некоторых случаях даже за счет функций низших отделов
центральной нервной системы.
Экстраполяция
наблюдается как при условнорефлекторной, так и при безусловнорефлекторной
деятельности, причем в некоторых случаях даже за счет функций низших отделов
центральной нервной системы.
В безусловнорефлекторной деятельности экстраполяция четко проявляется в следующем опыте. Обезглавленная (спинальная) лягушка лапкой рефлекторно снимает с кожи спины бумажку, смоченную кислотой. Этот рефлекс всегда осуществляется движением задней лапки той же стороны. Если ее привязать, то для снятия бумажки используется новый рефлекс — в виде движения задней лапки противоположной стороны. При привязывании обеих задних лапок возникают рефлекторные попытки снять раздражающий агент передними лапками.
Экстраполяция достигает особенно высоких форм при наличии значительного фонда условных рефлексов и связана как с сенсорной, так и с оперантной стороной высшей нервной деятельности.
Одним из проявлений экстраполяции в сенсорных компонентах условных рефлексов можно считать обобщение близких по характеру раздражителей. Если, например, на тон в 1000 гц вырабатывается условный рефлекс, то соответствующая реакция, как известно, возникает и при звучании более высоких и низких тонов. Чтобы отдифференцировать от них тон в 1000 гц, требуется специальное угашение действия более высоких или низких тонов.
В результате образования различного рода оперантных условных рефлексов не только человек, но и животные могут правильно реагировать на создавшиеся новые конкретные условия. Например, собака, заметив на охоте бегущего в определенном направлении зверя, экстраполирует скорость и направление передвижения, чтобы бежать к месту встречи по кратчайшему расстоянию. Вследствие этого она направляется не к тому месту, где был замечен зверь, а к месту предстоящего пересечения путей (рис. 64).
Своих наивысших форм экстраполяция достигает у человека. Она постоянно проявляется и при умственной, и при мышечной работе, в частности при спортивной деятельности.
На основании полученного опыта, т. е. приобретения ряда конкретных связей, человек путем экстраполяции правильно решает новые задачи, родственные в определенной мере ранее усвоенным (подробнее об экстраполяции в двигательных навыках см. главу XVI, § 9).
§ 10. Типы высшей нервной деятельности
Эти представление так же устарели.
На основании изучения условных рефлексов и сопоставления их с динамикой внешнего поведения И. П. Павлов выделил четыре основных типа нервной системы, совпадающие с классификацией темпераментов, предложенной более 2 тысяч лет назад Гиппократом. В основу классификации положено три функциональных показателя свойства процессов возбуждения и торможения: сила, уравновешенность и подвижность (табл. 6).
Сильный неуравновешенный (безудержанный) тип характеризуется сильными нервными процессами с резко выраженным преобладанием возбуждения. Представители этого типа очень активны. в них быстро вырабатываются положительные и медленнее тормозные условные рефлексы. Последние являются недостаточно стойкими. У представителей сильного уравновешенного подвижного и сильного уравновешенного инертного типов быстро вырабатываются и стабилизируются положительные и тормозные условные рефлексы.
Различие между этими типами обнаруживается при изменении сигнального значения положительных и тормозных раздражителей, кода положительные перестают подкрепляться, а тормозные подкрепляются безусловными или хорошо выработанными условными рефлексами. Представители сильного уравновешенного подвижного типа быстро приспосабливаются к изменившемуся значению раздражителей и вырабатывают соответственно изменившимся условиям новые условные рефлексы. Для представителей сильного уравновешенного инертного типа перестройка является более сложным процессом и осуществляется медленнее.
Таблица 6 Схема четырех типов нервной системы (по И. П. Павлову)
|
Тип нервной системы |
Характеристика по показателям свойств нервной системы |
Соответствие темпераментам по Гиппократу |
||
|
по силе |
по уравновешенности |
по подвижности |
||
|
Сильный неуравновешенный (безудержный) |
Сильный |
Неуравновешенный. Преобладание возбуждения над торможением |
— |
Холерик |
|
Сильный уравновешенный подвижный |
» |
Уравновешенный |
Подвижный |
Сангвиник |
|
Сильный уравновешенный инертный |
» |
» |
Инертный |
Флегматик |
|
Слабый |
Слабый |
Неуравновешенный. Преобладание торможения над возбуждением |
— |
Меланхолик |
Слабый тип характеризуется легкой тормозимостью даже при самых незначительных внешних влияниях. Представителей этого типа отличает замедленная выработка и неустойчивость положительных условных рефлексов. Тормозные рефлексы вырабатываются у них быстро и характеризуются стабильностью. Животные со слабым типом нервной системы даже при многолетнем пребывании в лаборатории проявляют пассивно-оборонительную реакцию.
Классификация типов высшей нервной деятельности у детей (Н. И. Красногорский, А. Г. Иванов-Смоленский) укладывается в схему четырех типов нервной системы, описанных И. П. Павловым. Критерии оценки типологических особенностей в этих классификациях не выходят за рамки силы, уравновешенности и подвижности нервных процессов. Признаки одного из четырех типов нервной системы наблюдаются у большинства взрослых людей, обследуемых по методике двигательных условных рефлексов на речевое подкрепление.
Однако в резко выраженном виде эти четыре типа встречаются относительно редко. Большинство людей относится к промежуточным формам, число вариантов которых весьма велико. Существенное значение при этом принадлежит воспитанию.
И. П. Павлов рассматривал тип нервной системы как сплав генотипа, т.е. наследственной основы нервной деятельности (обусловливающей конституциональные особенности организма, в том числе и темперамент), с фенотипом, т. е. приобретенными в результате воспитания свойствами.
Таким образом, в процессе воспитания к врожденным свойствам нервной системы добавляются новые, связанные с многообразным действием социальной среды. Свойства нервной системы, приобретенные под влиянием внешней среды, могут доминировать и подавлять генетические задатки. Это проявляется даже в опытах на животных. Так, внешне трусливые, проявляющие пассивно-оборонительную реакцию животные нередко обнаруживают в экспериментальных условиях все черты сильного типа, а внешне активные — черты слабого типа.
У человека, для которого социальная среда служит решающим фактором, определяющим характер высшей нервной деятельности, исключительно важное значение в формировании типа нервной системы имеет воспитание.
§ 11. Фазовые состояния
Здоровая нервная система в бодром состоянии характеризуется нормальным взаимодействием (балансом) между процессами возбуждения и торможения, благодаря чему обеспечивается правильное соотношение величины ответных реакций со значением и физиологической силой раздражителей. Но в некоторых случаях, когда организм оказывается в трудных условиях (например, при переутомлении, заболевании, решении ответственных или особенно трудных задач), баланс между возбуждением и торможением нарушается и в нервной системе возникают фазовые состояния. Признаками развития этих состояний являются неустойчивость структуры и нарушение закона силы в условнорефлекторных реакциях.
Закон силы заключается в том, что реакции организма на раздражители средней величины оказываются больше, чем на слабые раздражители, а реакции на сильные больше, чем на средние. Например, на слабый световой раздражитель реакция является наименьшей, на средний – несколько большей, и на сильный – самой большой. Эта разница реакции при разной физической интенсивности раздражителей, часто наблюдаемая в опытах на жирных, не всегда имеет место в человеческой практике. У людей интенсивность реакции в основном связана со смысловым значением раздражителей, в особенности при словесных сигналах. Тихо сказанное слово, например об опасности, имеет значительно большее значение, чем громко сказанная индифферентная фраза. Поэтому во взаимоотношениях людей физическая сила раздражителя только в некоторых случаях соответствует интенсивности ответной реакции. Значительно чаще эти ответные реакции, т. е. проявление закона силы обусловлены смысловой значимостью раздражителей. Нарушение закона силы в условнорефлекторных реакциях протекает по типу парабиоза Н. Е. Введенского и проявляется в возникновении фазовых состояний.
Первая фаза — уравнительная. Она характеризуется тем, что сильные, средние и слабые раздражители вызывают реакции одинаковой величины.
Вторая фаза — парадоксальная: слабые условные сигналы по величине реакции превосходят сильные.
Третья фаза — ультрапарадоксальная: при отсутствии условных реакций на положительные сигналы развивается положительный эффект на тормозные раздражители.
Четвертая фаза — тормозная. Она характеризуется полным торможением условнорефлекторных реакций.
Сравнительно недавно описана наркотическая фаза, характеризующаяся равномерным уменьшением величины условнорефлекторных ответов на слабые и сильные раздражители до полного их выпадения.
Четкая последовательность в развитии фазовых состояний иногда может нарушаться вследствие выпадения той или другой фазы. При переходе от парабиотического состояния к нормальному фазы следуют в обратном порядке.
Пограничные фазовые состояния особенно хорошо выражены при медленном погружении в гипнотический сон. Последний представляет собой частичное торможение коры, захватывающее относительно большие участки, особенно сенсомоторную область. При этом ряд центров может находиться в состоянии парадоксальной фазы, когда словесный раздражитель действует сильнее, чем сам предмет. В связи с этим предметы воспринимаются в соответствии со словесным раздражением.
Во время гипноза и сна «дежурные» пункты коры, настроенные на восприятие определенных сигналов внешней среды, свободны от фазовых явлениях и обладают высокой степенью возбудимости. При гипнозе эти корковые, центры, связанные с реагированием на голос гипнотизера, обеспечивают речевую связь (так называемый «рапорт»). Во сне эти центры воспринимают раздражители, имеющие для организма специальное значение (например, для матери — плач ребенка).
Нарушение нормального баланса нервных процессов при неврозах, перетренированности и др. может вызвать срыв высшей нервной деятельности и фазовые состояния. При преобладании процессов возбуждения нарушаются все условнорефлекторные реакции, связанные с торможением: дифференцировки, запаздывание и др. При преобладании процессов торможения исчезают многие из ранее выработанных условных рефлексов. Особенно легко срыв высшей нервной деятельности происходит у лиц с сильным неуравновешенным и у лиц со слабым типом нервной системы.
Для восстановления нормального состояния нервных процессов применяется ряд средств, важнейшими из которых являются отдых, облегчение условий работы, перемена обстановки, обеспечение нормального сна и питания, фармакологические средства.