КОГНИТОМ: В ПОИСКАХ ФУНДАМЕНТАЛЬНОЙ НЕЙРОНАУЧНОЙ ТЕОРИИ СОЗНАНИЯ, К. В. Анохин
Институт перспективных исследований мозга, Московский
государственный университет имени М.В. Ломоносова
Москва, Россия
2 Научно-исследовательский институт
нормальной физиологии им. П.К. Анохина
Москва, Россия
* E-mail: kanokhin@gmail.com
Поступила
в редакцию 27.08.2020
После доработки 05.10.2020
Принята к публикации 05.10.2020
· DOI:
ВВЕДЕНИЕ
Для науки о мозге нет более важной задачи, чем объяснить природу субъективного мира – решить многовековую проблему разум-мозг (mind-brain problem). Мы можем обозначить ее как “главную проблему” (MAIN problem – Mind-brAIN problem), стоящую перед современной нейронаукой. Принимая во внимание всю историю устремлений человека понять сущность собственного “Я”, можно ожидать, что решение “главной проблемы” явится крупнейшим научным достижением, сопоставимым по значению с эволюционной теорией происхождения видов.
Как и в случае дарвиновской революции, это решение вряд ли придет в результате какого-то отдельного наблюдения или эксперимента. Для него потребуется усилие особого рода – разработка фундаментальной теории когнитивных систем, обладающих свойствами разума и сознания. Такая теория должна проникнуть в сущность “главной проблемы”, предложить ее принципиальное решение и выступить в роли концептуального каркаса для исследования универсальных принципов происхождения, организации и функционирования когнитивных систем.
Поиски такой теории активно ведутся в последние годы. Их отличительной чертой стала концентрация усилий на проблеме сознания и его соотношения с мозгом. Стремительный рост числа работ на эту тему отмечается в последние 30 лет (Baars, 1988, 1997; Edelmаn, 1989, 2004; Edelman, Tononi, 2000; Massimini, Tononi, 2018; Dennet, 1991; Crick, 1994; Penrose, 1994; Chalmers, 1996; Searle, 1997; Damasio, 1999; Koch, 2003, 2012, 2019; Gray, 2004; Dehaene, 2015; Аллахвердов, 2000; Соколов, 2008; Васильев, 2009; Дубровский, 2015). Эта проблема сегодня нередко обозначается как “трудная проблема” сознания. Ее суть составляет давно известный вопрос: “каким образом материя мозга производит субъективное явление” (Павлов 1951, с. 247) или, в физикалистской формулировке Д. Чалмерса, предложившего выражение “the hard problem of consciousness”, “вопрос о том, как и почему физические процессы порождают субъективное сознание” (Chalmers, 2010, с. 105). Еще одна формулировка “трудной проблемы” – почему обработка мозгом информации не протекает “в темноте” свободно от внутренних ощущений, почему выполнение мозгом когнитивных и поведенческих функций сопровождается субъективным опытом? (Chalmers, 1996).
Сосредоточение современных исследований именно вокруг проблемы сознания обосновывается двумя причинами. Во-первых, сознание наиболее существенно для всей нашей психической жизни (Chalmers, 1996), включая даже и неосознаваемые ментальные процессы (Searle, 2000). Во-вторых, “сознание – это то, что делает проблему разума и тела действительно трудноразрешимой” (Nagel, 1974, p. 435). Современные исследования проблемы сознание-мозг движимы надеждой, что, решив эту “трудную проблему”, мы автоматически получим решение и “главной проблемы” – разум-мозг. Это было бы элегантным решением. Однако правомерность такого переноса внимания с “главной проблемы” на “трудную проблему” далеко не очевидна и требует специального рассмотрения.
Настоящая работа состоит из четырех частей. В первой выделены основные характеристики предмета современных исследований – феномена сознания. Во второй разбран список требований, которым должно отвечать научное объяснение сознания. В третьей части эти критерии использованы, чтобы оценить наиболее влиятельные современные нейронаучные теории сознания. Критический анализ показывает, что ни одна из них не отвечает требованиям к действенной теории сознания. Их наиболее заметным общим недостатком является то, что они не имеют ясной нейробиологической концепции носителя субъективного опыта – той системы, в которой протекают процессы сознания.
В четвертой части работы предлагается другой подход к решению “трудной проблемы” сознания и “главной проблемы” нейронауки. Я буду утверждать, что успех понимания природы сознания критически зависит от создания развернутой нейронаучной теории носителя сознательного опыта – того, что веками называлось “разум” (mind). Мы не можем расcчитывать полноценно понять процессы в системе, не имея понимания ее устройства и структуры. Разум, в конструктивном истолковании этого термина, как раз и является той особой структурой, в которой протекают специфические лишь для нее процессы – сознание.
Чтобы отделить это значение разума как когнитивной структуры от других смыслов данного термина, я использую понятие “когнитом” (Анохин, 2012). В отличие от нематериального res cogitans, когнитом будет рассматриваться как высокопорядковая структура головного мозга – нейронная гиперсеть (Анохин, 2015). Сознание согласно этому подходу является особой формой динамики в этой гиперсети – широкомасштабной интеграцией ее когнитивных элементов. Объяснить сознание можно лишь имея теорию когнитома – мозга как нейронной гиперсети.
1. СОЗНАНИЕ: ОПРЕДЕЛЕНИЕ ПРЕДМЕТА ИССЛЕДОВАНИЯ
Вначале определим сам предмет настоящего исследования – сознание. Подчеркнем, что нам пока не требуется развернутое научное определение сознания. На данном этапе нас интересует определение не объясняющее, а объясняемое, т.е. не explanans, а explanandum. Другими словами, нам необходимо выделить в понятии “сознание” те его ключевые характеристики, которые были накоплены многовековым опытом употребления этого термина и которым должно быть дано объяснение в научной теории.
В целом этот подход может выразиться в простом изначальном определении сознания, встречающемся в последнее время в философских и нейронаучных работах:
“Часто можно услышать, что “сознание” пугающе трудно определить. Но если мы говорим об определении в терминах здравого смысла, достаточном для определения цели исследования, в противоположность точному научному определению того рода, которое обычно приходит в конце научного исследования, то это слово не кажется мне трудным для определения. Вот определение: Сознание состоит из внутренних, качественных, субъективных состояний и процессов восприятия или осознания. Сознание, определяемое таким образом, начинается, когда мы просыпаемся утром от сна без сновидений, и продолжается до тех пор, пока мы снова не засыпаем, не умираем, не впадаем в кому или иным образом не становимся “бессознательными”. Оно включает в себя огромное разнообразие осознаний, которое мы считаем характерным для нашей бодрствующей жизни. Оно включает в себя все: от ощущения боли до зрительного восприятия объектов, до состояния тревоги и депрессии, до разгадывания кроссвордов, игры в шахматы, попыток вспомнить номер телефона своей тети, споров о политике или просто желания оказаться где-нибудь в другом месте. Сновидения по этому определению являются формой сознания, хотя, конечно, они во многих отношениях весьма отличаются от сознания при бодрствовании” (Searle, 2000, с. 559).
Это определение вполне соответствует нашему интуитивному пониманию сознания. Однако в качестве рабочего определения цели исследования оно все же недостаточно. Чтобы оценить, достигнута ли цель, она должна быть адекватно параметризована. Продвижение в этой проблеме уже само является началом теоретической работы, поскольку для сознания не существует систематического и общепризнанного каталога признаков, которые служили бы предметом объяснения в научной теории (Metzinger, 2003; 2009). Настоящий раздел будет посвящен решению этой задачи.
Отметим сразу, что на данном этапе параметризации мы не должны стремиться к исчерпывающему перечислению всех возможных свойств сознания. В ходе работы над фундаментальной теорией сознания предстоит еще многое узнать о нем, и некоторые выделяемые сегодня детали могут оказаться незначимыми, уйти на второй план или быть поглощенными более общими категориями. Тем не менее для начала этой работы нам нужно иметь достаточно дифференцированное определение сущностных свойств сознания, которое могло бы далее направлять разработку теории.
Прежде чем мы приступим к этому анализу, я хотел бы указать на сопровождающую его проблему. С одной стороны, наше собственное сознание неразрывно сплетено с языком и отражает общественное сознание, наши соотношения с культурой. Но, с другой стороны, свойства субъективного опыта вызревали на протяжении геологических эпох и его базовые формы присутствуют у многих видов животных. Фундаментальная теория сознания должна начинаться с объяснения именно этих основополагающих свойств сознания. Насколько оно может быть успешным на языке, категории которого определялись и формировались в соответствии с запросами специфической для человека социальной практики – отдельный и серьезный вопрос. И все же в данной работе мы будем ставить своей целью понимание именно базовых характеристик сознания, относящихся к универсальным свойствам любого феномена субъективного опыта.
1.1. Начальная параметризация сознания
Многомерность понятия сознания всегда составляла одну из главных сложностей для его объяснения. С этим был также связан долгий негласный запрет на использование термина “сознание” в нейронаучных исследованиях. Один из основателей когнитивной нейронауки Дж. Миллер писал: «Сознание – это слово, затертое миллионами языков… Может быть, нам следует запретить это понятие на десятилетие или два, пока мы не сможем разработать более точные термины для различных употреблений, которые сейчас затушевывает “сознание”» (Miller, 1962). Однако и четверть века спустя в Международном словаре психологии С. Сазерленд был вынужден дать следующее определение: “Сознание: обладание восприятиями, мыслями и чувствами; осведомленность. Этот термин невозможно определить иначе, как в терминах, которые непонятны без предварительного понимания того, что означает сознание… Об этом не было написано ничего стоящего прочтения” (Sutherland, 1996).
Отложим оценочную сторону этого высказывания до результатов анализа конкретных теорий сознания и сосредоточимся на описательной части определения. Отметим вначале, что оно дает характеристику сознания через перечисление трех других ментальных процессов – восприятия, мышления и ощущения. Это, безусловно, придает определению черты циркулярности. Однако обратим внимание, что все три понятия соответствуют одной и той же общей характеристике сознания, данной более века назад У. Джеймсом, использовавшим для нее выражение “поток сознания”: сознание – это не вещь, а процесс, возникающий вследствие взаимодействия мозга, тела и среды (James, 1978).
Заметим также, что это описание сознания устойчиво в веках – оно касается все тех же процессов, которые почти 400 лет назад Р. Декарт понимал под термином cogitatio: “все то, что происходит в нас таким образом, что мы воспринимаем его непосредственно сами собою; и поэтому не только понимать, желать, воображать, но также чувствовать означает здесь то же самое, что мыслить (сogitare)”. (Декарт, 1950, с. 429). Можно видеть, что знаменитое декартовское “je pense…”, cogito, означало нечто большее, чем мышление, и идея существования включала для него более широкий спектр ментальных процессов, ассоциируемых нами сегодня с общим понятием “сознание” (Searle, 2000; Koch, 2019).
В совокупности, мы должны сделать из этого важный вывод, что в состав предмета фундаментальной теории сознания должны входить не только феномены мышления и восприятия, но и все остальные виды субъективных процессов и состояний: чувства, ощущения, эмоции, мотивации.
Этот набор явлений часто объединяют под терминами “опыт”, “субъективный опыт” или “субъективная реальность” – “сознание – это субъективный опыт” (Tononi et al., 2016), “сознание обладает специфическим и неотъемлемым качеством субъективной реальности” (Дубровский, 2015). Интегральное понятие субъективной реальности как раз охватывает большое разнообразие видов явлений: ощущения, восприятия, чувства, мысли, намерения, желания, волевые усилия и т.д. (Дубровский, 2015). Все они попадают в категорию качественных состояний сознания. Другим термином, обозначающим сознание с этой качественной стороны, является понятие “квалиа” (qualia), часто используемое в англоязычной аналитической философии для обозначения субъективного аспекта состояния сознания, его внутренне узнаваемых качественных признаков, особенности “каково это”; а также понятие “феноменальное”, более используемое в европейской философии.
Возвращаясь к определению сознания, данному Сазерлендом, обратим внимание на четвертый термин, отделенный от первых трех точкой с запятой, подчеркивающей его не рядоположенность предыдущим, – осведомленность (awareness). Осведомленность или осознание также характеризует свойство сознания как активного состояния. Однако оно описывает его уже не со стороны его качественного содержания, а как процесс доступа субъекта к этому качественному содержанию. “Ментальное состояние является сознательно доступным, если субъект имеет определенный вид доступа к содержанию этого состояния. Точнее, состояние является осознанным, если в силу наличия этого состояния его содержание доступно для вербального сообщения, для рационального вывода и для сознательного контроля поведения.” (Chalmers, 2010, с. 503). Некоторые исследователи полагают, что именно особенность сознательного доступа является главной определяющей чертой сознания: “Подлинным сознанием следует считать сознательный доступ – тот простой факт, что обычно, когда мы бодрствуем, все, на чем мы решаем сосредоточиться, может стать сознательным.” (Dehaene, 2015, с. 9).
Американский философ Н. Блок зафиксировал эти разные стороны, выделенные Сазерлендом в определении сознания, как две его разных формы: феноменальное сознание (phenomenal consciousness, P-consciousness), обладающее свойствами квалиа, и сознание доступа (access consciousness, A-consciousness) – функциональную доступность той или иной информации всем частям когнитивной системы (Block, 1995, 2004). Его аргумент в пользу такой демаркации заключается в наличии фактов двойной диссоциации этих форм сознания, когда каждая из них может наблюдаться без второй. Сторонники этой точки зрения отождествляют феноменальное сознание с квалиативными свойствами опыта и считают, что проблема его объяснения и есть “трудная проблема” сознания, определяющая “разрыв в объяснении” (Levine, 1983) соотношения сознания и мозга (Young, 2003; Bayne, 2009; Block, 2012). А проблема осознанного доступа, согласно этой точке зрения, относится к числу “легких проблем” сознания и она решаема средствами когнитивной науки и нейронауки. Однако другие исследователи не принимают это разделение сознания на две отдельных формы (Chalmers, 1997; Cohen, Dennett, 2011, Schlicht, 2012; Dehaene, 2015), и по этому вопросу ведется активная дискуссия (Kouider et al., 2012; Block, 2012, Mashour et al., 2020).
1.2. “Что”: качественные свойства сознания
Сознание отличается от всех других известных феноменов своими уникальными качественными свойствами. Обозначим их здесь термином “что”. Современные исследователи приводят перекрывающиеся наборы таких отличительных свойств. Рассмотрим некоторые из них.
I. Сёрл. Важнейшие свойства сознания (Searl, 2000):
A. Важнейшее свойство сознания – сочетание качества, субъективности и целостности.
1. Качественность: каждое сознательное состояние имеет определенное качественное ощущение.
2. Субъективность: сознательные состояния существуют только тогда, когда их переживает какой-то субъект – человек или животное.
3. Целостность: все сознательные переживания в любой момент жизни агента являются частью единого сознательного поля.
B. Некоторые другие свойства:
4. Сознательные состояния обычно обладают “интенциональностью”, тем свойством ментальных состояний, посредством которого они направляются на объекты и ситуации в мире.
5. Сознательные состояния характеризуются различием между центром и периферией внимания.
6. Все сознательные переживания существуют в том или ином настроении.
7. Все сознательные состояния имеют измерение удовольствия / неудовольствия.
8. Сознательные состояния отличаются ситуативностью, они связаны с положением дел в мире.
9. Сознательные состояния имеют гештальтную структуру – организацию вырожденных перцептивных стимулов в когерентные перцептивные формы.
10. Сознательные состояния несут чувство знакомости.
II. Дж. Эдельман. Отличительные свойства сознательных состояний (Edelman, 2003):
1. Сознательные состояния целостны, интегрированы и сконструированы мозгом.
2. Они могут быть чрезвычайно разнообразными и дифференцированными.
3. Они упорядочены во времени, последовательны и изменчивы.
4. Они отражают связывание различных модальностей.
5. Они обладают конструктивными свойствами, включая гештальт, замыкание и феномены заполнения.
6. Они демонстрируют интенциональность с широким содержанием.
7. Они имеют обширный доступ и обладают ассоциативностью.
8. У них есть центр, периферия, окружение и пограничные аспекты.
9. Они подвержены модуляции вниманием, от фокальной до диффузной.
10. Они отражают субъективные чувства, квалиа, феноменальность, настроение, удовольствие и неудовольствие.
11. Они связаны с ситуативностью и положением в мире.
12. Они порождают чувство знакомости или ее отсутствия.
III. А. Дамасио: Определяющие свойства сознательных состояний (Damasio, 2010):
1. Они всегда имеют содержание (они всегда о чем-то).
2. Они всегда воспринимаются как интегрированная совокупность частей.
3. Они всегда выявляют различимые качественные свойства по отношению к различным содержаниям.
4. Они всегда содержат обязательный аспект чувствования – они ощущаются как нечто для нас.
IV. Дж. Тонони. Важнейшие свойства сознания (Tononi, 2017):
1. Сознание существует внутренне: мое переживание (experience) реально.
2. Сознание структурировано: каждое переживание имеет внутреннюю структуру, состоящую из феноменальных различий, связанных вместе различными способами, которые существуют внутри него.
3. Сознание специфично: каждое переживание имеет специфическую форму – определенную композицию специфических феноменальных различий, связанных вместе различными способами и тем самым отличающихся особым образом от других переживаний.
4. Сознание целостно: каждое переживание не сводимо к не зависящим друг от друга компонентам.
5. Сознание определенно: каждое переживание очерчено по своему содержанию и пространственно-временной зернистости.
Объединим эти списки в таблицу из десяти наиболее часто упоминаемых отличительных свойств сознания (). Анализируя ее, можно отметить, что почти все перечисленные в ней свойства могут быть представлены как специфические варианты более базовых биологических категорий. Дифференцированность и интегрированность, специфичность и индивидуальность, определенность и разнообразность, временная структура и вписанность в среду характеризуют многие физиологические процессы и состояния у организмов, не обладающих сознанием или не пользующихся им в этих процессах. Будучи помноженными на одно единственное свойство, выделенное в пятой строке – особое качество субъективных явлений, их квалитативность, квалиа, они приводят ко всем остальным отличительным характеристикам сознания.
Таблица 1.
Отличительные свойства сознания . Characteristic features of consciousness
|
№ |
Свойство сознания |
I |
II |
III |
IV |
Прототипическое свойство |
Специфика у сознания |
Разновидность свойства |
|
1 |
реальность |
ѵ |
|
|
ѵ |
каузальность (К) |
квалитативность |
особый вид К |
|
2 |
субъективность |
ѵ |
ѵ |
|
|
индивидуальность (И) |
квалитативность |
особый вид И |
|
3 |
дифференцированность |
|
ѵ |
|
ѵ |
дифференцированность (Д) |
квалитативность |
особый вид Д |
|
4 |
целостность |
ѵ |
ѵ |
ѵ |
ѵ |
целостность (Ц) |
квалитативность |
особый вид Ц |
|
5 |
качественность |
ѵ |
ѵ |
ѵ |
ѵ |
специфичность (С) |
квалитативность |
особый вид С |
|
6 |
интенциональность |
ѵ |
ѵ |
ѵ |
|
специфичность (С) |
квалитативность |
особый вид С |
|
7 |
разнообразность |
|
ѵ |
|
ѵ |
разнообразность (Р) |
квалитативность |
особый вид Р |
|
8 |
определенность |
|
ѵ |
|
ѵ |
определенность (О) |
квалитативность |
особый вид О |
|
9 |
ситуативность |
ѵ |
ѵ |
|
|
вписанность (В) |
квалитативность |
особый вид В |
|
10 |
временная структура |
|
ѵ |
|
ѵ |
временная структура (ВС) |
квалитативность |
особый вид ВС |
Так, субъективность может быть рассмотрена как квалитативный вариант свойства индивидуальности сложной системы. Дифференцированность сознания может являться частным проявлением общего принципа дифференцированности процессов и состояний организма, который, будучи умноженный на квалитативность составляющих сознание элементов, дает особый вид дифференцированности. Целостность сознания может быть представлена как частный случай интегрированности, объединяющей в данном случае специфические элементы субъективного опыта и т.д.
Это, безусловно, не снимает вопроса о специфике конкретных механизмов интеграции, дифференциации и других свойств в сознательной системе. Это также не означает, что все разнообразие субъективных феноменов, входящих в сложную структуру сознания, должно сводиться исключительно к квалиа. Однако отмеченное обстоятельство позволяет сфокусировать теорию на задаче объяснения, в первую очередь, именно основ квалитативности субъективного опыта. Если теория сможет сделать это в естественно-научных понятиях, то есть надежда, что и другие специфические характеристики сознания получат свое закономерное объяснение.
1.3. “Где” и “когда”: динамические аспекты сознания
В ходе начальной параметризации сознания мы выделили два его базовых свойства. Во-первых, это качественные характеристики феноменальных состояний – “что”. Во-вторых, это специфические операции над этими состояниями, обеспечивающие доступ субъекта к их содержанию. Сами эти процессы недоступны осознанию, но составляют непременные компоненты любого явления сознания, которые обязана объяснить фундаментальная теория. Их можно было бы обозначить общим термином “как”. Более детальное их рассмотрение показывает, что это “как”, по-видимому, следует разделить на “как” в смысле “где” и “как” в смысле “когда”.
I. Где. Аспект “как” в смысле “где” относится к тому факту, что и само субъективное состояние и доступ к нему разворачиваются в некоем пространстве. Что это за пространство, и что в нем в этот момент происходит? Это, возможно, один из самых сложных вопросов в проблеме “сознание и мозг”. Если оставить гипотезы прошлых веков о желудочках мозга и эпифизе, то за последние десятилетия в качестве критических областей, отвечающих за сознание, выдвигался целый реестр мозговых структур, начиная от ретикулярных ядер ствола мозга, ядер таламуса, структур дорзального и вентрального зрительного пути, лимбических структур, гиппокампа, клауструма, до разных регионов неокортекса (Laureys et al., 2015). В отношении локализации сознания в коре в последние годы ведется оживленная дискуссии между сторонниками взглядов, что его можно более прицельно сосредоточить в задних сенсорных или же передних исполнительных областях неокортекса. Вместе с тем против каждой из этих теорий могут быть выдвинуты серьезные контраргументы (Laureys et al., 2015). Одним из способов примирить эти противоречия являются попытки интегрировать перечисленные регионы мозга в единые нейросетевые архитектуры (Hunt, Schooler, 2019).
Однако, возможно, вопрос еще более сложен. Во-первых, обратим внимание, что ответ на вопрос “где” в терминах топографии мозга млекопитающих мало подходит для фундаментальной теории сознания. Он не обладает тем масштабом универсальности, который соответствовал бы распространенности феноменов субъективного опыта в мире животных с разной организацией нервной системы (Edelman et al., 2005, Edelman, Seth, 2009, Burghardt, Bekoff, 2009; Ginzburg, Jablonka, 2019). Если, как считается, субъективные ощущения способны испытывать, например, птицы (Birch et al., 2020) и некоторые виды головоногих моллюсков (Godfrey-Smith, 2016; Mather, 2019; Birch et al., 2020), то ответ на вопрос “где”, выраженный в терминах структур головного мозга млекопитающих, мало что даст для понимания субъективного опыта в нервной системе вороны или осьминога.
Но наибольший недостаток существующих подходов к решению вопроса “где”, часто обозначаемого как поиск нервных коррелятов сознания (Crick, Koch, 1990), состоит в том, что любое перечисление только нейроанатомических структур и протекающих в них нейрофизиологических процессов оставляет нерешенной задачу сцепления “где” с “что” – с особым субъективным содержанием состояния сознания. Это как раз тот самый, отмеченный философами, “разрыв в объяснении” (Levine, 1983). В разработке фундаментальной теории сознания мы будем исходить из того, что она будет успешной, только если укажет на способ этого сцепления. Решение данной задачи может потребовать перехода теории от топографического пространства нервной системы того или иного вида организмов в высокопорядковое топологическое пространство когнитивной системы. В этом пространстве качественные свойства субъективного опыта, рассмотренные ранее под термином “что”, должны сохраняться независимо от изменчивых деталей строения конкретного типа нервной системы.
II. Когда. Аспект “как” в смысле “когда” относится к пороговым условиям сознательного доступа, процессу “глобального воспламенения” (global ignition) (Dehaene, Changeux, 2005; 2011), обеспечивающему появление специфического осознанного состояния. Далеко не все ментальные состояния становятся осознанными – случаи подпорогового восприятия, ложной слепоты (blindsight), бессознательной веры, влияющей на поведение субъекта, иллюстрируют это. “Когда достаточное количество областей мозга соглашается в оценке важности поступающей сенсорной информации, они синхронизируются в крупномасштабное состояние глобальной коммуникации. Обширная сеть воспламеняется вспышкой активации высокого уровня – и природа этого воспламенения объясняет наши эмпирические опознавательные признаки сознания” (Dehaene, 2015). Значительная доля современных нейронаучных исследований сознания посвящена выяснению нейробиологических механизмов этого “когда” и “как” – процессов в нейронной сети мозга, обеспечивающих ее “глобальное воспламенение” (Noy et al., 2015; Levy et al., 2016; Noel et al., 2019; Mashour et al., 2020).
Еще одно рассмотрение аспекта “когда” выдвигается в высокопорядковых (higher-order) теориях сознания, утверждающих, что граница между осознаваемыми и неосознаваемыми состояниями определяется моментом осознания собственных когнитивных процессов. “Именно благодаря восприятию наших собственных восприятий последние становятся феноменально сознательными. … воспринимая свое восприятие красноты, я осознаю ее; и именно последнее осознание делает первое феноменально сознательным.” (Carruthers, 2017). Высокопорядковые теории сознания также ставят перед нейронаукой вопрос о том, когда и при каких условиях низкопорядковые когнитивные процессы преодолевают порог осознания и становятся феноменальными (Carruthers, 2017).
В какой мере “где” и “когда” являются самостоятельными механизмами сознания или же разными аспектами рассмотрения одного и того же процесса сознательного доступа, предстоит понять в ходе исследований. Пока у нас нет достаточно ясного понимания глубинной сущности процессов, стоящих за этими категориями, мы будем разделять их.
Таким образом, мы выделили три базисные стороны любого субъективного опыта, условно обозначенные нами как “что”, “где” и “когда”. Существуют ли другие фундаментальные компоненты, которых недостает в этом описании сознания?
1.4. “Кто”: скрытый ингредиент в определениях сознания
Основной тезис настоящего раздела состоит в том, что все определения сознания опираются на еще одну, как правило, скрытую, но непременную составляющую часть – “кто”. “Кто” служит носителем “что” – источником всех квалиа; определяет “когда” – выступает генератором их надпороговой активации; и является системой, “где” происходит их осознание – служит реципиентом этих квалитативных состояний. Посмотрим с этой точки зрения на некоторые из описаний сознательного опыта.
“Ибо если я скажу “я вижу” и “я иду” и сделаю отсюда вывод, что “я существую”, и буду разуметь действия, совершаемые моими глазами или ногами, то заключение не будет настолько непогрешимым, чтобы я не имел основания в нем сомневаться, так как я могу думать, что вижу или хожу, хотя бы я не открывал глаз и не трогался с места, как бывает подчас во сне и как могло бы быть даже, если бы я вовсе не имел тела. Если же я подразумеваю только действия моей мысли или моего чувства, иначе говоря мое внутреннее сознание, в силу которого мне кажется, будто я вижу или хожу, то заключение настолько правильно, что я в нем не могу сомневаться, ибо оно относится к душе, которая одна лишь способна чувствовать и мыслить каким бы то ни было образом.” (Декарт, 1950, с. 428–429).
“Сознание – это не просто бодрствование. Проснувшись, я не стал рассеянно оглядываться по сторонам, впитывая в себя все виды и звуки, как будто мое бодрствующее сознание никому не принадлежало. Напротив, я почти мгновенно, без малейших колебаний, без малейших усилий, понял, что это я сижу в самолете, возвращаясь домой в Лос-Анджелес с длинным списком дел до конца дня, ощущая странное сочетание усталости от путешествия и энтузиазма по поводу того, что ждет впереди, интересуясь взлетно-посадочной полосой, на которую мы приземлимся, и внимательно следя за регулировкой мощности двигателя, которая доставит нас на землю. Несомненно, бодрствование было обязательным для этого состояния, но бодрствование вряд ли было его главной особенностью. Что это была за главная особенность? Это факт, что бесчисленное множество содержимого, отображаемого в моем сознании, независимо от того, насколько ярко или хорошо оно упорядочено, через невидимые нити, которые свели эти содержания вместе, связано со мной, владельцем моего разума, в движущемся вперед пиршестве, которое мы называем самостью. И не менее важен факт, что связь эта была мною ощутима.” (Damasio, 2010, c. 3).
“Когда я смотрю на красную книгу, я могу сообщить о наличии книги (“есть красная книга”), я могу рассуждать по этому поводу (например, сделать вывод, что я, должно быть, положил ее туда, когда читал вчера), и я могу использовать ее присутствие в сознательном управлении моим поведением (например, когда я беру книгу и кладу ее обратно на полку). Таким образом, мое восприятие красной книги дает мне соответствующий доступ к информации о красной книге. Мое состояние восприятия здесь – сознательный доступ. Можно также сказать, что в таком случае субъект осознает доступ к соответствующему объекту. Итак, здесь я осознаю красную книгу. Точно так же многие из моих перцептивных состояний являются сознательными, как и многие из моих эмоциональных и когнитивных состояний.” (Chalmers, 2010, c. 503).
“Сознание существует внутренне. Мой опыт реален … – например, я вижу свою спальню – это единственный факт, в котором я могу быть немедленно и абсолютно уверен; более того, он существует для меня с моей внутренней субъективной точки зрения. Сознание структурировано. … В рамках одного и того же опыта я могу видеть разные места в визуальном пространстве, разные цвета, разные объекты, объекты определенных цветов в определенных местах и так далее. Сознание специфично. … Мое восприятие спальни здесь и сейчас – это то, что она есть, кровать, тело на ней, книжный шкаф, синяя книга на левой полке книжного шкафа и так далее. Более того, мой текущий опыт неизбежно отличается от других переживаний, содержащих другие объекты и цвета, звуки и запахи, или опыт чистой тьмы и тишины и так далее. Сознание едино. … Я переживаю всю визуальную сцену, а не левую сторону зрительного поля зрения, независимую от правой стороны, и наоборот. Более того, я вижу предмет, лежащий на столе, как книгу, а не как набор признаков.” (Tononi, 2017, c. 243–244).
Эти и многие другие характеристики сознания, данные разными авторами в разные времена и с разными намерениями, содержат один общий компонент: местоимения “я” и “мой”. Мы так часто используем их, что обычно не даем себе отчета в их опорной роли в собственных словах и мыслях. Однако стоит убрать их из приведенных выше описаний сознания, и весь их смысл теряется. Само понятие субъективного опыта уже предполагает существование субъекта, которому принадлежит этот опыт. Как отмечал Г. Фреге, “чувственное впечатление зеленого, которым я обладаю, возникает только благодаря мне: я являюсь его носителем. … Ощущение невозможно без ощущающего. Внутренний мир предполагает того, внутри кого он существует.” (Фреге, 1918/2000).
Это означает, что категория “кто” также обязана входить в число неотъемлемых ингредиентов полноценного описания сознания.
В философии вопрос что такое “Я” занимает выдающееся место. Как отмечают Гинзбург и Яблонка, точек зрения на природу “Я” существует больше, чем самих философов (Ginsburg, Jablonka, 2019, с. 175). Введение Декартом понятия “вещь мыслящая” (res cogitans), по сути, было попыткой решения именно этого вопроса в его соотношении с субъективным опытом: “А что это такое – вещь мыслящая? Это нечто сомневающееся, понимающее, утверждающее, отрицающее, желающее и не желающее, а также – обладающее воображением и чувствами” (Декарт, 1994, с. 24). Велико также число теорий личности в психологии (Hjelle, Zielgler, 1992). Однако в нейронаучных теориях концепция “кто” практически не разработана. На мой взгляд, это является основной причиной затруднений в современных попытках научного понимания сознания. В заключение статьи я предложу план действий по преодолению этого недостатка.
1.5. “Кто”, “что”, “где” и “когда” эпизода сознания
Итак, сознание имеет особые качественные свойства и специфические операциональные особенности. И те и другие – атрибуты когнитивного агента и не могут быть поняты без понимания устройства последнего. Поэтому описание любого эпизода сознания требует категорий “кто”, “что”, “где” и “когда”. Мы можем изобразить разворачивающийся при этом процесс в виде схемы ().
Рис. 1.
Основные ингредиенты универсального эпизода сознания.
. Major constituents of universal episode of consciousness.
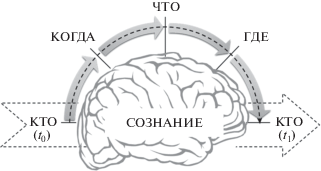
• Любой эпизод сознания развивается в структуре когнитивного агента (КТО).
• В какой-то момент времени часть когнитивных элементов этой системы активируется до надпорогового уровня (КОГДА).
• Их совместная активность создает уникальную по содержанию интеграцию – состояние сознания (ЧТО).
• Такая группа активных элементов обретает каузальный доступ в глобальное пространство других когнитивных элементов системы (ГДЕ).
• Результатом является изменение целостного состояния когнитивного агента (КТО).
Таким образом, каждый элементарный эпизод субъективного опыта – это всплеск специфической по своему субъективному качеству глобальной активности в когнитивном агенте, неважно, порожден ли он внешними или внутренними условиями. Искомая фундаментальная теория обязана объяснить нейронаучную конкретику “кто-что-где-когда”, составляющих этот универсальный сознательный эпизод.
2. ТРЕБОВАНИЯ К НАУЧНОЙ ТЕОРИИ СОЗНАНИЯ
Прежде чем начинать работу над теорией сознания, уже имея очерченный предмет ее объяснения, необходимо определиться с тем, каким требованиям должно соответствовать это объяснение. Что значит объяснить сознание? Как мы узнаем, что уже получили объяснение базовых “кто-что-где-когда” сознания и можем перейти к пониманию его более высокопорядковых свойств? Об этих условиях оценки необходимо договориться еще до начала разработки теории, чтобы иметь возможность объективно определить достижение ею результата в конце пути.
Ниже будет разобран ряд критериев, которым должно соответствовать полноценное научное объяснение сознания. Однако вначале необходимо устранить одно важное заблуждение, препятствующее прогрессу в решении этой задачи.
2.1. Что значит понять сознание?
Как следует из предыдущего анализа, современные представления о сознании сосредоточились на специфических качествах субъективного опыта (Chalmers, 1996, 2010; Block, 2004, Edelman, 2004; Tononi, 2012; Tononi, Koch, 2015; Koch, 2019). Часто это выражается в определении сознания через ощущение “каково-это-быть” (Chalmers, 1996, 2010; Koch, 2019), сформулированном в известной статье Т. Нагеля “Каково быть летучей мышью?”:
“Тот факт, что организм имеет сознательный опыт вообще, в фундаментальном смысле означает, что существует нечто, что значит быть этим организмом … Организм имеет сознательные ментальные состояния, если и только если существует нечто, каково быть этим организмом для самого этого организма. Мы можем назвать это субъективным характером опыта.” (Nagel, 1974).
Нередко вслед за этим делается вывод, перекрывающий любую возможность последующего научного решения проблемы. Его логика примерно такова: даже если наука преуспеет в раскрытии всех эволюционных корней и нервных основ сознания, это ничего не даст для понимания главного свойства сознания – “каково-это-быть”. Другими словами, мы вынуждены признать, что проблема соотношения разума и тела так же когнитивно закрыта для людей, как квантовая механика для зебр (McGinn, 2000).
Эта аргументация, конечно, не нова. Знаменитое “не знаем и никогда не узнаем” Дюбуа-Реймона основывалось на этой же логике. Полтора века назад Ф. Энгельс, разбирая ее, использовал вместо примера летучей мыши с ее своеобразными органами эхолокации, муравья с его специфическими органами химической рецепции. Рассматривая возможность объективного научного познания восприятия муравьем этих химических сигналов, чувственно не доступных нам самим, он отмечает: “Разумеется, мы никогда не узнаем, в каком виде воспринимаются муравьями химические лучи (сигналы). Кого это огорчает, тому уж ничем нельзя помочь” (Энгельс, 1961, с. 554). Л.С. Выготский, приводящий эту цитату полувеком позже, продолжает: “Заметим кстати, что на этом психологическом примере можно видеть, как не совпадают в психологии факт научный и факт непосредственного опыта. Оказывается, можно изучать, как видят муравьи, и даже как они видят невидимые для нас вещи, и не знать, какими эти вещи являются муравьям, т.е. возможно устанавливать психологические факты, отнюдь не исходя из внутреннего опыта, иначе говоря не субъективно. Энгельс даже, видимо, считает это последнее для научного факта неважным: кто этим огорчается, говорит он, тому ничем нельзя помочь” (Выготский, 2000, с. 30).
Сегодня мы знаем о нервных механизмах такого вида чувственного познания и биологических причинах его ограниченности гораздо больше, чем во времена Энгельса и Выготского. Исходя из этого, я разделю принципиальные способы понимания субъективных явлений на два: зеркальный и научный.
Способ I – зеркальное познание. Этот биологически важный способ познания сформировался в эволюции нервной системы у социальных организмов (Panksepp, Panksepp, 2013; Chen et al., 2018). В его основе лежит способность одной когнитивной системы сопереживать психологическое состояние другой родственной ей системы за счет резонансной активации в своем когнитивном аппарате элементов, зеркальных по отношению к активированным когнитивным элементам второй системы (Keysers, Gazzola, 2017; Spunt, Adolphs, 2019; Kingsbury et al., 2019). Такая резонансная активация элементов когнитивного аппарата дает познающему субъекту способность частичного воспроизведения и, таким образом, эмпатического понимания ментальных ощущений, которые испытывает познаваемый субъект. Не только люди, но и другие виды социальных животных используют этот важный инструмент познания (Panksepp, Panksepp, 2013; de Waal, Preston, 2017). В рамках определенной эволюционной гомологии ментальных процессов он способствует даже межвидовому эмпатическому взаимодействию, например, между собакой и человеком (Harris, Prouvost, 2014; Van Bourg et al., 2020).
Однако существенным ограничением зеркального способа познания является необходимость совпадения когнитивных структур и элементов двух субъектов по познаваемому параметру. Это полностью невозможно даже у представителей одного вида. Как заметил один из участников семинаров по сознанию у Ф. Крика: “Я не только не знаю, каково это быть летучей мышью, я даже не знаю, каково это быть моей женой”.
Рассматривая логический перенос от вывода об ограниченности чувственного способа познания к заключению о принципиальной невозможности понять сознание, А. Тьюринг писал: “если следовать этому взгляду, то окажется, что единственный способ убедиться, что данный человек действительно мыслит, состоит в том, чтобы стать именно этим человеком. Фактически эта точка зрения является солипсистской.” (Turing, 1950, с. 446).
Способ II – научное познание. Научное познание существенно расширило горизонты человеческого понимания. В отличие от зеркального, оно охватывает объекты и события, к которым у него нет непосредственного чувственного или, тем более, сочувственного доступа. В этом модусе понять субъективное состояние означает не ощутить “каково это” в виде когнитивного резонанса собственных ментальных состояний, а объяснить данное явление с помощью научной теории. “Когда мы говорим, что понимаем группу природных явлений, мы имеем в виду, что нашли конструктивную теорию, охватывающую эту группу явлений” (Эйнштейн, 1965, с. 247). Понять в этом случае означает “понять эмпирическую закономерность как логическую необходимость” (Эйнштейн, 1965, с. 265), при которой “мы хотим, чтобы наблюденные факты логически следовали из нашей концепции реальности” (Einstein, Infeld, 1938). Применительно к естественно-научному пониманию сознания это означает нахождение теорией таких основополагающих принципов, из которых с неизбежностью следуют факты субъективности, квалиа, ощущений “каково-это-быть”. С помощью подобной теории мы хотим узнать, почему и как природа привела к возникновению таких когнитивных систем, и как она неизменно достигает этого результата в каждом из рождающихся на свет сознательных организмов.
Безусловно, оба способа познания комплементарны и каждый дает то, на что не способен другой. Однако в контексте целей настоящей работы отметим четыре важных обстоятельства.
1) Зеркальное познание, даже в случае его успешности, ограничено. “Кто-то другой может испытывать сострадание ко мне; но при этом моя боль всегда будет принадлежать мне, а его сострадание – ему. Он не испытывает моей боли, а я не испытываю его сострадания.” (Фреге, 2000).
2) Зеркальное познание того или иного конкретного психологического состояния, даже в случае его успешности, не обладает универсальностью. Оно не позволяет перенести достигнутое резонансное понимание на другие случаи субъективного опыта. Для этого необходима единая теория.
3) Зеркальное познание не обладает никакой объяснительной силой при решении “трудной проблемы” сознания. Оно не дает ответа на вопрос – почему выполнение мозгом определенных когнитивных и поведенческих функций сопровождается субъективным опытом, а не протекает “в темноте” (Chalmers, 2010). В лучшем случае, оно позволяет ощутить, каково то или иное субъективное состояние, но не предоставляет каких-либо способов узнать, “почему природа является именно такой, а не другой”. Этот вопрос может быть разрешен только в рамках научного модуса познания.
4) Зеркальное познание бесполезно для понимания субъективного опыта у систем, устройство и эволюция которых сильно отличаются от нашей. Одной из актуальных проблем такого рода является вопрос о существовании сознания у осьминогов, возможность которого активно обсуждается в исследованиях последнего времени (Godfrey-Smith, 2016; Ginsburg, Jablonka, 2019; Mather, 2007, 2019; Birch et al., 2020). Осьминоги имеют ноцицептивные нейронные системы, воспринимающие повреждающие стимулы (Crook et al., 2013, Andrews et al., 2013), – испытывают ли они боль? Они демонстрируют игровое поведение (Kuba et al., 2006; Mather, Anderson, 2013) – испытывают ли они при этом положительные эмоции? Они способны обучаться, наблюдая за поведением других осьминогов (Fiorito, Scotto, 1992), – протекают ли у них при этом резонансные эмпатические процессы? Нервная система осьминога устроена так, что церебральные ганглии и брахиальное сплетение – нервное кольцо, охватывающее основание щупалец, обладают достаточной функциональной независимостью (Godfrey-Smith, 2016; Carls-Diamante, 2017) – может ли быть так, что они обладают и двумя относительно независимыми восприятиями мира? Нейронные сети каждого из щупалец осьминога также относительно автономны, осьминоги предпочитают разные конечности для разных видов своей активности, при этом конечности функционируют отчасти независимо от мозга (Godfrey-Smith, 2016; Schnell et al., 2020) и обладают механизмами самоузнавания, предотвращающими их интерференцию друг с другом (Nesher et al., 2014), – не может ли быть так, что они имеют даже не две, а девять систем опыта и сознания (Birch et al., 2020)?
Зеркальный способ познания не может помочь в ответе на эти вопросы. Научная теория сознания, напротив, призвана решить:
1. Что такое любое субъективное состояние, в чем его сущность?
2. Обладает ли осьминог субъективными состояниями?
3. В каких когнитивных модальностях, регистрах опыта они лежат?
4. Почему у осьминога возникают субъективные состояния?
5. Как нервная система осьминога генерирует эти состояния?
6. При каких условиях и в какие именно моменты они возникают?
7. Где в пространстве нервной системы осьминога они возникают?
8. Насколько эти состояния целостны и исключительны?
9. Варьируют ли они по выраженности и как ее измерить?
Если мы будем иметь фундаментальную научную теорию сознания, то на некоторые из этих вопросов мы получим ответы не только для осьминога, но и для любого сознательного существа, например, человека. Для решения других из них потребуется знание специфики устройства когнитивной системы осьминога, организации его нервной сети, ее архитектуры и связей, особых процессов когнитивной дифференциации и интеграции в ней. Но и здесь только научная теория способна направить исследование, определить необходимые приборы и методы и позволить понять смысл того, что будет наблюдаться в экспериментах. Рассмотрим ключевые требования к такой теории.
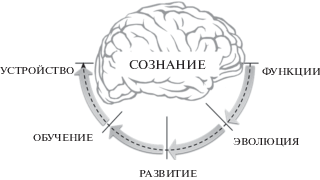
2.2. Пять вопросов для теории сознания
Записи Чарльза Дарвина в период открытия принципов естественного отбора показывают, насколько ясно он представлял, что разум и сознание являются продуктами биологической эволюции (Darwin, 1988, p. 539). Для настоящего анализа это означает, что объяснение сознания должно удовлетворять тем же условиям, что и объяснение других биологических явлений. В данной работе я выделю пять таких требований.
Разбирая, какие главные вопросы должна решать теория тех или иных биологических явлений, С. Бреннер отмечал: “Биологи задают о живом организме только три вопроса: как он работает? Как он был построен? И как он получился таким? Это проблемы, воплощенные в классических областях физиологии, эмбриологии и эволюции.” (Brenner, 2012). Схожий набор ключевых вопросов выдвигали и другие выдающиеся биологи-эволюционисты (Huxley, 1942; Mayr, 1961). Однако наибольшее распространение получила формулировка Н. Тинбергена (Tinbergen, 1963). В знаменитой статье, посвященной 60-летию Конрада Лоренца, он отметил, что биологи, работающие над поведением, поднимают четыре принципиально разных вида вопросов, которые он определил как “ценность для выживания”, “эволюция”, “онтогенез” и “причинность” (Tinbergen, 1963). Хотя Тинбергена интересовало поведение, эти проблемы можно выразить в виде четырех вопросов к любому свойству организма (Bateson, Laland, 2013):
− для чего оно нужно?
− как оно развивалось на протяжении истории вида?
− как оно развивалось в течение жизни индивида?
− как оно устроено и работает?
Вопросы о ценности и эволюции часто обозначаются как вопросы “почему”, а развитийные и механистические – как вопросы “как” (Bateson, Laland, 2013; Nesse, 2018), хотя нередко эти четыре проблемы называют также “четырьмя почему” Тинбергена, потому что они представляют собой четыре способа спросить: “почему это животное ведет себя таким образом?” (Bolhuis, Giraldeau, 2005).
Поскольку мы принадлежим к виду биологических организмов, для которого нет ничего более реального и причинного, чем наше собственное сознание, четыре вопроса Тинбергена распространяемы и на него (Gutfreund, 2018):
1. Каковы функции сознания?
2. Как сознание формируется в эволюции?
3. Как сознание развивается у индивида?
4. Каково устройство сознания?
Однако в отношении сознания мы должны разделить вопрос об онтогенезе на две части. Первая – о видоспецифических закономерностях созревания нервных субстратов сознания в ходе пренатального развития. Это тот этап развития, который был отнесен Бреннером к компетенции эмбриологии. В нем закладываются нервные механизмы и свойства сознания, общие для всех организмов с данным филогенезом.
Вместе с тем у когнитивных организмов, обладающих долговременной пластичностью, морфогенез мозга продолжается и после рождения. В этот период их фенотипическая пластичность выражается в особом виде процессов – обучении и формировании долговременной памяти, накоплении индивидуального опыта. Эта фаза развития и ее специфические механизмы, протекающие под когнитивным контролем, требуют от теории отдельного объяснения. Поэтому в итоге для организмов, обладающих сознанием, мы можем выделить пять принципиальных вопросов, на которые должна ответить объясняющая его научная теория:
1. Каковы функции сознания?
2. Как сознание формируется в эволюции?
3. Как сознание созревает в ходе эмбриогенеза?
4. Как сознание развивается в процессах обучения?
5. Каково устройство сознания?
Мы можем представить эти пять вопросов как опорные для всех явлений сознания (). Их значение, действительно, очень велико. Из ответов на вопросы об устройстве и функциях сознания должна стать понятна его качественная уникальность – его специфические механизмы и его особая причинная сила. Из ответов на вопросы о происхождении должно вытекать понимание, почему сознание обладает такими уникальными свойствами. Как существование биологических видов невозможно понять без теории их происхождения, точно так же существование сознания невозможно по-настоящему понять без теории его происхождения. Если научная теория даст убедительные ответы на все эти пять вопросов, она решит “трудную проблему” – объяснит сознание.
Рис. 2.
Ключевые вопросы, стоящие перед теорией сознания.
. Key questions for theory of consciousness.
2.3. Циркулярная ловушка вопросов о сознании
Итак, мы разобрали ряд базовых свойств сознания и сжали их до четырех ингредиентов: “кто”, “что”, “где”, “когда”. Рассматривая требования к самой теории, мы определили пять ключевых вопросов “как” и “почему”, на которые она обязана ответить. В итоге этого анализа мы можем определить круг вопросов, с которыми должна иметь дело любая успешная теория сознания:
1. Почему и как сознание возникало в эволюции?
2. Как сознание созревает в развитии?
3. Как сознание развивается при обучении?
4. Кто является источником состояний сознания?
5. Когда и как они возникают?
6. Что и как определяет их уникальные свойства?
7. Где и как они реализуются?
8. Кто и почему является их бенефициаром?
Представим их в виде круговой диаграммы, каждая из осей которой соответствует одному из ключевых вопросов ().
Рис. 3.
“Циркулярная ловушка” сознания.
. A “circular trap” of consciousness.
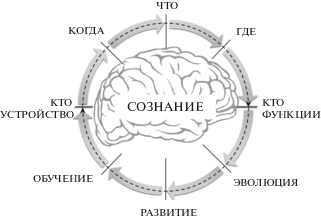
В совокупности они образуют то, что можно было бы назвать “циркулярной ловушкой” феномена сознания. Ее суть состоит в следующем:
− невозможно по-настоящему понять сущность субъективного опыта (“что”), не понимая устройство его носителя – разума (“кто”);
− невозможно по-настоящему понять устройство разума, не понимая законы его формирования в процессах обучения;
− невозможно по-настоящему понять процессы обучения, не понимая принципов развития нервной системы в онтогенезе;
− невозможно по-настоящему понять принципы развития нервной системы в онтогенезе, не понимая закономерности ее эволюции в филогенезе;
− невозможно по-настоящему понять закономерности эволюции нервной системы в филогенезе, не понимая роль в них нервных механизмов поведения и субъективного опыта (“что”).
В этом замкнутом кольце решение каждого из вопросов зависит от ответа на предыдущий, а последний всегда замыкается на первый. Действенная теория сознания обязана охватить весь этот цикл в целом – другого способа выхода из циркулярной ловушки просто не существует. Это одно из самых сложных требований к теории сознания. Если теория терпит неудачу на каком-то одном из этапов этого цикла, она не может полноценно объяснить и последующий, а это значит, что она не может по существу объяснить ничего.
2.4. Первые принципы теории сознания
Список разбираемых критериев будет недостаточен, если он не будет включать в себя и общие требования к фундаментальной теории, преломленные в отношении специфики теории сознания:
1) Фундаментальная теория должна обладать широким кругозором. Искомая теория сознания должна объяснять и предсказывать как можно большее количество явлений в трех областях ее компетенции, которые можно условно обозначить как М1, М3 и N3:
М1 – ментальные явления, наблюдаемые от первого лица, т.е. субъективно;
М3 – ментальные явления, наблюдаемые от третьего лица, т.е. объективно;
N3 – нервные явления, наблюдаемые от третьего лица, т.е. объективно.
То, что теория сознания должна одновременно распространяться на все эти три вида феноменов, нетривиально – большинство теорий не выходят за пределы каких-то двух из них. Учитывая обсуждение предыдущих разделов, подобные решения не могут быть признаны удовлетворительными.
2) Фундаментальная теория должна быть компактной. Если четыре базисных ингредиента субъективного опыта “кто-что-где-когда” умножить на объяснение их в терминах пяти “как” и “почему”, мы получим двадцать необходимых ответов. Если прибавить к этому требование объяснить и десять выделенных качественных свойств сознания, число требующихся ответов превысит шестьдесят. Если же учесть, что в объяснение сознания должны войти не только онтологический, но и гносеологический (знание), аксиологический (ценность), праксеологический (активность) ракурсы его рассмотрения (Дубровский, 2009), плюс должны быть решены многочисленные вопросы соотношения сознания с бессознательными процессами, восприятием, вниманием, памятью и другими когнитивными категориями, и все эти аспекты необходимо описать для нервных систем столь различных организмов, как, например, головоногих моллюсков, птиц и млекопитающих, то число требуемых объяснений начнет исчисляться сотнями.
Очевидно, что этот объем не может быть обеспечен за счет формулировки самостоятельного ответа на каждый из вопросов. Специальные ответы, вводимые только для решения отдельного вопроса, противоречат идеалу научного объяснения. Его общим принципом является простота фундаментальной теории. Разбирая это требование в отношении физической теории, Эйнштейн отмечал: “Теория преследует две цели: 1. Охватить по возможности все явления в их взаимосвязи (полнота); 2. Добиваться этого, взяв за основу как можно меньше логически независимых понятий и произвольно установленных соотношений между ними (основных законов или аксиом)” (Эйнштейн, 1965, с. 264).
3) В описании Эйнштейна первая цель соответствует нашему понятию кругозора теории, а вторая – ее компактности. В максимально общем виде мы можем свести эти условия к единственному – компрессии. Успешная компрессия в теории проявляется в том, что “теория производит тем большее впечатление, чем проще ее предпосылки, чем разнообразнее предметы, которые она связывает, и чем шире область ее применения” (Эйнштейн, 1965, с. 143). Это требование можно конкретизировать в отношении теории сознания следующим образом:
фундаментальная теория сознания должна с необходимостью выводить максимально большее число феноменов М1, М3 и N3 из максимально меньшего числа основополагающих принципов.
Если принять во внимание справедливую критику, что сознание сейчас можно определить лишь в терминах, которые непонятны без предварительного понимания самого смысла сознания (Sutherland, 1996), мы можем сформулировать это требование еще более жестко:
фундаментальная теория сознания должна выводить максимально большее число явлений М1, М3 и N3 из наименьшего числа основополагающих принципов, которые сами не выражены в терминах субъективного опыта.
Только такие принципы можно назвать первыми принципами фундаментальной теории сознания.
2.5.Что означает фундаментальность теории сознания?
Сформулированное выше требование имеет определяющее значение для решения принципиального вопроса: что такое фундаментальная теория сознания? К каким объяснениям она должна стремиться в отличие от не фундаментальных теорий?
На этот счет возможны две точки зрения. Обе они исходят из того, что такая теория объясняет фундаментальные явления – те, которые не сводимы ни к каким другим более простым. Однако затем они диаметрально расходятся.
Первая из них утверждает, что в фундаментальной теории сознания субъективный опыт должен быть понят как несводимая сущность, заложенная на самом глубоком физическом уровне, подобно таким свойствам, как время, масса и пространство. Согласно одному из сторонников такой точки зрения Д. Чалмерсу, такая “теория сознания будет иметь больше общего с теорией физики, чем с теорией биологии. Биологические теории не включают в себя никаких фундаментальных принципов, поэтому биологическая теория имеет определенную сложность и беспорядок; но теории в физике, поскольку они имеют дело с фундаментальными принципами, стремятся к простоте и элегантности” (Chalmers, 1995, с. 210). Примером одной из научных теорий, основанной на таком понимании сознания, является разбираемая ниже теория интегрированной информации (Tononi, 2017). Авторы, придерживающиеся такой позиции, как правило, разделяют ту или иную из версий панпсихизма (Chalmers, 2010; Tononi, Koch, 2015; Tononi, 2017; Koch, 2019). Элементарными формами субъективного опыта и определенным уровнем сознания должны, согласно этим теориям, обладать протоны и нейтроны (Koch, 2012, c. 132), фотодиоды (Tononi, 2008, c. 236), термостаты (Chalmers, 1996, c. 294-295), а простые двумерные решетки из большого числа физических элементов могут вообще обладать уровнем сознания, превосходящим человеческий (Tononi, 2014, 2017a, c. 254; 2017b, c. 623–624).
Я буду исходить из принципиально иной позиции. Как и первая, она исходит из нередуцируемости фундаментальных явлений и принципов. Как и первая, она придерживается идеалов простоты и элегантности таких принципов. Но далее второй взгляд радикально отличается от первого. Его можно сформулировать в виде следующих двух положений:
1. В ходе эволюции возникают новые уровни структурной организации материи, имеющие свойства и качества, не объяснимые в терминах чего-либо более простого. В этом смысле эти сущности являются фундаментальными для данного уровня.
2. В составе эволюционирующей организации данного уровня имеются принципы, которые при достаточном времени их действия способны привести к возникновению систем более высокого уровня с их новыми фундаментальными свойствами и качествами.
Таким образом, в рамках данной парадигмы свойства более высоких уровней не сводимы к более низким, но исторически выводимы из них.
Конечно, в русскоязычной литературе эта система взглядов узнаваема как преломление исторического и диалектического материализма (см., напр., Пономарев, 1983; Швырков, 2006; Дубровский, 2009). Конкретизируем ее применительно к рассматриваемой в данной работе проблеме.
Мы будем считать фундаментальной только такую теорию сознания, первые принципы которой принадлежат сугубо биологическому уровню, но при достаточном времени их действия неизбежно приводят к возникновению нередуцируемой специфики когнитивных систем и субъективных явлений. Именно такая фундаментальная теория будет интересовать нас в данном поиске.
Подобный ориентир исключает из нашего дальнейшего рассмотрения все теории сознания, апеллирующие за объяснением к физике (Beshkar, 2020), необычной физике (Penrose, 1994) и тем более выходящим за пределы физики (Eccles, 1990). Отдельно отметим, что вопрос о построении систем с искусственным сознанием в рамках этой методологии обсуждаем, однако принципиально иначе, чем при первой точке зрения.
2.6. Схема оценки теорий сознания
Суммируем все содержание настоящего раздела в виде простой последовательности утверждений:
1. Объяснить сознание означает создать научную теорию сознания.
2. Эта теория должна отвечать на пять вопросов “как” и “почему” в отношении отличительных свойств и ключевых ингредиентов субъективного опыта.
3. Ответы на эти вопросы должны быть совместимы с требованием выхода из “циркулярной ловушки” сознания.
4. Фундаментальная теория сознания должна охватывать своим объяснением максимальное количество феноменов М1, М3 и N3, исходя при этом из минимального числа первых принципов.
5. Первые принципы фундаментальной теории сознания должны основываться на понятиях биологического уровня, из которых должно закономерно выводиться возникновение свойств когнитивного уровня.
Составленную нами диаграмму вопросов к научной теории сознания можно использовать для сравнения различных теорий сознания. С этой целью отложим на каждой из ее осей пять потенциальных уровней обращения теории к вопросу ():
Рис. 4.
Диаграмма для оценки объяснительной силы теории сознания.
. Diagram for evaluation of explanatory power of theory of consciousness.
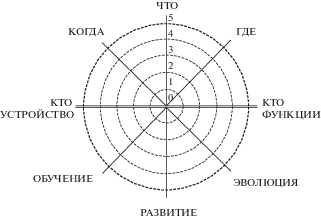
0 – теория не упоминает данный вопрос;
1 – теория ставит данный вопрос, но не предлагает решения;
2 – теория ставит данный вопрос и предлагает его решение;
3 – теория предлагает решение, привлекая к этому известные факты;
4 – теория предлагает решение и объясняет им известные факты;
5 – теория предлагает решение и делает с его помощью предсказания.
В итоге мы обладаем схемой, с помощью которой можно провести сравнительный анализ полноты и объяснительной силы различных теорий сознания. При необходимости объяснительный потенциал теории можно подвергнуть измерению и в отношении выделенных в десяти отличительных свойств сознания, оценив их по такой же пятибалльной шкале.
3. НАУЧНЫЕ ТЕОРИИ СОЗНАНИЯ
В современной литературе существует большое число научных теорий сознания (см., напр., Дубровский 2009; Bayne et al., 2009; Schneider, Velmans 2017). В то же время имеется мнение, что “об этом не было написано ничего стоящего прочтения” (Sutherland, 1996). Это едкое высказывание Сазерленда далеко не исключение, и другие авторы тоже считают, что “существует огромное количество теорий о сознании, но очень немногие из них являются обоснованными или даже полезными” (Dehaene, 2011, с. 218).
Настоящий раздел не ставит своей целью систематический обзор и классификацию всех имеющихся научных теорий сознания. Мы применим разработанный набор критериев для оценки лишь четырех наиболее влиятельных нейробиологических теорий. Задачей этого анализа будет выявить те типологические пробелы существующих решений, которые необходимо преодолеть при разработке новой теории.
3.1. Теория селекции нейронных групп
Теория селекции нейронных групп (ТСНГ) – наиболее разносторонняя и детальная из разбираемых здесь теорий. Первоначально она была сформулирована Дж. Эдельманом в виде гипотезы механизма сознания на основе селекции нейронных групп и фазной повторной сигнализации (Edelman, 1978) и затем развита в трилогии монографий (Edelman, 1987, 1988, 1989) и ряде последующих публикаций (Edelman, 1993, 2003, 2004; Tononi, Edelman, 1998; Edelman et al., 2011; Edelman, Gally, 2013).
В исходной работе Дж. Эдельман предложил принцип соматического отбора нейронных групп в головном мозге, обеспечивающий пластичность мозга при взаимоотношениях организма с окружающей средой (Edelman, 1978). При этом он отметил сходство этой модели с принципами селекции в иммунной системе. В монографии “Neural Darwinism” эта идея была существенно развита и представлена в качестве общей селекционной теории мозга (Edelman, 1987). В основу теории были положены три принципа: (1) отбор в развитии, (2) отбор на основе опыта и (3) сигнализация путем повторного входа. Единицами отбора в мозге, согласно теории, являются нейронные группы – совокупности от сотен до тысяч нейронов, действующих в нервной системе как функциональные единицы (Edelman, 1987). Отбор в развитии приводит к формированию первичного ассортимента таких нейронных групп в головном мозге, отбор на основе опыта приводит к формированию из них вторичного ассортимента, а механизм повторного входа обеспечивает пространственно-временное увязывание активности этих сформировавшихся нейронных групп при взаимодействии организма с окружающим миром и является нейробиологической основой для возникновения состояний или “сцен” сознания (Edelman, 1987).
Впоследствии были опубликованы две другие части данной трилогии (Edelman, 1988, 1989). В “Topobiology” (Edelman, 1988) излагалась теория эмбриологических механизмов образования первичного репертуара и их отбора при развитии, в отсутствие когнитивных взаимодействий организма и среды. В монографии “Remembered Present. A Biological Theory of Consciousness” (Edelman, 1989) была представлена развернутая нейронаучная теория сознания – расширенная ТСНГ. Теория утверждает, что основой состояний сознания является особый нервный процесс повторного входа. На критическую роль данного нервного механизма обращают внимание и другие теории (Иваницкий, 1996; Сергин, 1998, 2016; Damasio, 1999; Lamme, 2006; Grossberg, 2013), которые по этому признаку нередко обознаются как “рекуррентные” теории сознания. Согласно ТСНГ повторный вход характеризуется постоянным двунаправленным обменом сигналами вдоль реципрокных аксональных волокон, связывающих две или более области мозга. Это позволяет широко распределенным в мозге группам нейронов достигать интегрированного и синхронизированного возбуждения, которое и является основой сознания (Edelman, 1989, 1993; Edelman, Gally, 2013). Предполагается, что эти процессы фазной повторной сигнализации связывают нейронные сети сенсорных задних областей коры, осуществляющих перцептуальную категоризацию, с передними областями и лимбическими структурами, поддерживающими ценностные категории организма.
Первоначальная версия ТСНГ подверглась резкой критике со стороны Ф. Крика, не нашедшего в ней, с одной стороны, достаточной оригинальности, с другой – достаточной убедительности (Crick, 1989). Позже теория была расширена и включила гипотезу “динамического ядра” (dynamic core), которая описывает таламокортикальную область мозга как функциональный кластер – интеграционный центр сознания (Tononi, Edelman, 1998; Edelman, 2003, 2004). Взаимодействие между различными нейронными группами в динамическом ядре определяет осуществляемые им высокопорядковые дискриминации – квалиа, иными словами, качественную природу сознательного опыта.
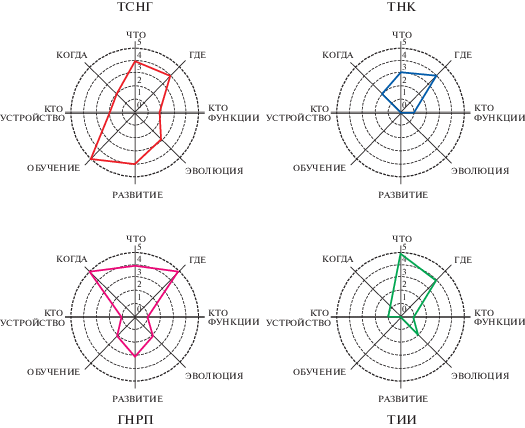
Система представлений Эдельмана затрагивает почти весь набор сформулированных нами требований к полноценной теории сознания ( (а)). Она обращается к механизмам морфологической эволюции нервной системы, молекулярным и клеточным механизмам развития, формированию репертуара функциональных нейронных групп в эмбриональном периоде, нервным механизмам раннего обучения и обучения во взрослом возрасте, в том числе, ведущего к формированию новых категорий и концепций, роли языка в формировании высокопорядкового сознания у человека, нейрональным основам квалиа и механизмам их связывания в целостные сцены субъективного опыта, функциональной роли этих субъективных состояний в естественном отборе. Наиболее заметным слабым звеном этой сложной теории является недостаточная проработка ею модели “кто” – самого субъекта, являющегося источником и носителем субъективного опыта. В отличие от других рассматриваемых ниже теорий, ТСНГ обращается к этому вопросу, говоря, что система “кто” представлена регуляторными системами внутреннего гомеостаза тела, преломляющегося через подкорковые гипоталамические центры и лимбические структуры в системы ценностной памяти организма, которые вступают через передние, височные и теменные области коры во взаимодействия с первичными и вторичными сенсорными регионами коры. Согласно теории, именно эти рекуррентные взаимодействия через механизмы повторного входа и процессы перцептуальной категоризации и ведут к явлениям первичного сознания (Edelman, 1989, 2003, 2004). Однако теория не предлагает разработанной модели того, как в “ценностных системах” нейронов голубого пятна, ядер шва и вентральной покрышки среднего мозга, секретирующих норадреналин, серотонин и дофамин, воплощаются качественные категории “ценностей” (Edelman, 1989), в пептидергических нейронах, миндалине и других подкорковых структурах возникают качественные категории “эмоций” (Edelman, 2003, 2004), и как их циклические взаимодействия с сенсорными регионами неокортекса производят в конечном счете качественную категорию “квалиа”.
Рис. 5.
Объяснительная сила ведущих нейронаучных теорий сознания. ТСНГ – теория селекции нейронных групп; ТНК – теория нейронных коалиций; ГНРП – теория глобального рабочего пространства; ТИИ – теория интегрированной информации.
. Explanatory power of main neurobiological theories of consciousness. ТСНГ – The Theory of Neuronal Group Selection; ТНК – The Neuronal Coalition Theory; ГНРП – The Global Neuronal Workspace Theory; ТИИ – The Integrated Information Theory
3.2. Теория нейронных коалиций
Теория нейронных коалиций была предложена Ф. Криком и К. Кохом первоначально как теория нервных коррелятов сознания, имеющих в своей основе механизмы синхронизации на частоте 40 Гц у обширных популяций нейронов коры головного мозга во время состояний зрительного осознания (Crick, Koch, 1990). Однако позже авторы отошли от утверждения, что гамма-синхронизация является достаточным условием возникновения сознания, и развили более общие взгляды о нейронной коалиции как нейронном корреляте сознания (Crick, Koch, 1998, 2003, 2007; Koch, 2004; Koch et al., 2016). Согласно этой концепции, многие типы нейронов коры головного мозга, как возбуждающие, так и тормозные, образуют кратковременные объединения, члены которых поддерживают друг друга тем или иным образом, увеличивая активность своих нейронов-собратьев. Победившая коалиция приобретает некоторую устойчивость и воплощает в себе содержание сознания.
Согласно теории, такие нейронные коалиции могут различаться по размеру и характеру (Crick, Koch, 1998, 2007). Так, нейронная коалиция, генерируемая зрительным воображением с закрытыми глазами, может быть менее обширной, чем коалиция, создаваемая ярким и устойчивым зрительным входом. Коалиции во сне могут несколько отличаться от коалиций, формирующихся во время бодрствования. Нейронные коалиции в передней части коры могут иметь несколько иной характер, чем те, которые формируются в задней части коры. В частности, фронтальные коалиции могут отражать такие чувства, как настроение и, возможно, ощущение “авторства”, связанное с чувством воли. Эти ощущения могут быть более диффузными по своей локализации и продолжаться более длительное время, чем коалиции в задних отделах коры (Crick, Koch, 2007).
Крик и Кох не разбирают детального устройства клеточных коалиций, отмечая только, что сознательный образ конкретного аспекта зрительной сцены предполагает существование небольшой группы нейронов, реагирующих на этот признак как детектор (Crick, Koch, 2007). Корковая сеть при этом может быть представлена как состоящая из узлов. Каждый ее узел необходим для появления того или иного аспекта восприятия, который не может стать осознанным, если для него не существует данного критического узла. Чтобы конкретный признак мог достичь осознания, нейронная активность составляющего его узла должна достичь надпорогового уровня. Это маловероятно, если она не становится членом общей успешной коалиции (Crick, Koch, 2007).
Авторы теории пишут, что идея нейронных коалиций опирается на представление о нейронных ансамблях, высказанное Д. Хеббом (Hebb, 1949). В этом смысле теорию нейронных коалиций можно рассматривать как представителя целого класса “ансамблевых” теорий сознания. Крик и Кох также подчеркивают, что ряд других теорий, среди которых они называют теорию селекции нейронных групп и гипотезу “динамического ядра” Дж. Эдельмана, а также теорию нейронального глобального рабочего пространства С. Дехана и Ж-П. Шанже, в значительной степени перекрываются с их теорий (Crick, Koch, 2007).
Оценивая данную теорию в рамках сформулированного нами списка требований ( (б)), можно видеть, что она адресуется лишь к узкому кругу вопросов, связанных, во-первых, с условиями возникновения состояний сознания, во-вторых, с условиями осознания этих состояний, в-третьих, с нервными коррелятами сознательных состояний и, лишь косвенно, с вопросом о качественном содержании субъективного опыта или с проблемой “смысла в широком плане”, по определению авторов. Таким образом, специализация данной теории лежит в области вопросов “где”, “когда” и, отчасти, “что”. Теория не решает проблему “кто” и не упоминает всего блока вопросов, связанных с историческими факторами – формированием сознания в эволюции, развитии и вследствие обучения.
3.3. Теория глобального нейронного рабочего пространства
Теория глобального нейронного рабочего пространства была выдвинута С. Деханом и соавт. (Dehaene, Nacchache, 2001; Sergent, Dehaene, 2004; Mashour et al., 2020), одним из которых является выдающийся французский нейробиолог Ж.-П. Шанже (Dehaene, Changeux, 2005, 2011). Эта теория представляет собой нейробиологическое расширение когнитивной теории глобального рабочего пространства, ранее сформулированной Б. Баарсом (Baars, 1988).
Центральный тезис исходной теории глобального рабочего пространства состоял в том, что элементы перцептивной информации становятся сознательными только тогда, когда они широко транслируются через мозг к другим процессорам информации. Такое глобальное вещание подразумевает, что информация в рабочем пространстве становится доступной для многих локальных процессоров, и именно широкая доступность этой информации, как предполагается, составляет основу для сознательного опыта (Baars, 1988). В качестве структуры мозга, определяющей глобальное рабочее пространство, Баарс предложил расширенную ретикулярно-таламическую активирующую систему.
Гипотеза глобального нейронного рабочего пространства (ГНРП) расширила и детализировала нейробиологическую сторону этих представлений (Sergent, Dehaene, 2004; Dehaene, Changeux, 2005, 2011; Mashour et al., 2020). В дополнение к локализованным, специализированным модульным кортикальным областям, обрабатывающим специфическую информацию, теория вводит второе нейронное рабочее пространство. Оно состоит из широко распределенных по коре длинноаксонных возбуждающих нейронов, образующих взаимосвязанные тракты. Эти нейроны способны избирательно мобилизовать или подавлять вклад специфических процессорных нейронов.
Согласно теории ГНРП, сознание – это общемозговой процесс распределения и совместного использования информации (Dehaene, 2015). При этом субъективное состояние определяется устойчивым возбуждением на несколько сот миллисекунд коалиции из подмножества нейронов рабочего пространства. Данные нейроны кодируют разные аспекты субъективного образа, которые составляют единое интегрированное целое за счет синхронизующего воздействия нейронов префронтальной коры, имеющих длинноаксонные нисходящие проекции в сенсорные области в задней части коры (Dehaene, Changeux, 2011, Dehaene, 2015, Mashour et al., 2020). Состояние сознания возникает нелинейным образом, называемым в теории “воспламенением” (ignition) (Dehaene, Changeux, 2005). “Воспламенение” характеризуется внезапной когерентной активацией подмножества длинноаксонных нейронов рабочего пространства, кодирующих текущее содержание сознания. Этому процессу сопутствуют волна P3b и вспышка высокочастотных осцилляций, при этом остальная часть нейронов рабочего пространства ингибируется. “Воспламенение” может быть вызвано внешним стимулом, например, при когнитивной задаче, или может произойти спонтанно в состоянии покоя (Dehaene, 2015, Mashour et al., 2020).
Таким образом, главным предметом теории ГНРП являются вопросы “когда” – при каких обстоятельствах и посредством каких механизмов происходит возникновение эпизодов сознания, “где” – посредством каких нервных процессов и где в мозге происходит осознание перцептуальной информации и, отчасти, “что” – в аспекте нервных носителей информации, составляющей содержание сознания ( (в)). Теория обращается к вопросам развития сознания в постнатальном онтогенезе человека и других млекопитающих (Dehaene, 2015, Mashour et al., 2020), но не разбирает эволюционные механизмы, развитие нервных субстратов сознания в эмбриогенезе, а также специфические механизмы обучения и памяти, ответственные за формирование индивидуального содержания сознания. Также как другие рассмотренные ранее теории, она не делает попытку ответить на вопрос: “кто” является тем “я”, которое генерирует сознательные состояния и воспринимает возникающую в них когнитивную информацию. Присущая ей и другим рассмотренным здесь теориям проблема магического участия “я” в потоке физиологической активности структур мозга хорошо видна из следующего фрагмента изложения теории:
“Под осознанием определенной информации мы подразумеваем то, что она достигла уровня обработки в мозгу, на котором ею можно поделиться. Поскольку она доступна для совместного использования, ваша область Брока (или ее часть, участвующая в выборе слов, которые вы собираетесь произнести) информируется о содержании того, что вы видите, и вы получаете возможность назвать то, что вы видите. В то же время ваш гиппокамп, возможно, получает информацию о том, что вы только что видели, поэтому вы можете сохранить это представление в памяти. Ваши теменные области также получают информацию о том, что вы видели, чтобы они могли направить внимание или решить, что это не то, чем вы хотите заниматься… и так далее и тому подобное. Критерий совместного использования информации отсылает к нашему ощущению, что всякий раз, когда часть информации осознается, мы можем делать с ней очень широкий спектр вещей. (Dehaene 2011, p. 226–227).
3.4. Теория интегрированной информации
Теория интегрированной информации (ТИИ) была предложена Дж. Тонони, который на протяжении ряда лет являлся сотрудником Дж. Эдельмана и разработал с ним концепцию “динамического ядра” сознания (Tononi, Edelman, 1998). В отличие от других перечисленных здесь теорий, берущих начало либо от нейронауки, либо от психологии, ТИИ исходит из фактов чистой феноменологии, обозначаемых в теории как аксиомы сознания. Теория выделяет пять таких аксиом, исчерпывающе описывающих, по утверждению Дж. Тонони, любое явление субъективного опыта. Для этих аксиом в теории формулируются соответствующие им постулаты – утверждения об устройстве физических механизмов, определяющих указанное феноменологическое свойство и в совокупности необходимых и достаточных для реализации любого из субъективных состояний (Tononi, 2004, 2008, 2012; 2017, Oizumi et al., 2014; Tononi, Koch, 2015; Tononi et al., 2016).
Аксиомы ТИИ гласят, что сущностные свойства сознания таковы. Каждый субъективный опыт: (1) существует внутренне; (2) он структурирован; (3) специфичен; (4) един; (5) определен. Далее ТИИ утверждает, что для каждого из этих специфических свойств феноменального существования должно иметься соответствующее свойство физического существования. Такой физический субстрат сознания (ФСС): (1) должен обладать причинно-следственной силой, обращенной на самого себя; (2) составляющие его части также должны обладать причинно-следственной силой внутри ФСС; (3) ФСС должен предопределять причинно-следственную структуру, которая является специфичной; (4) единой; (5) определенной.
Исходя из этих постулатов, ТИИ описывает ФСС как совокупность элементов, называемых “комплексом”, задающих внутреннюю причинно-следственную структуру определенной и максимально несводимой формы. Эта форма называется в ТИИ “концептуальной структурой”. Концептуальная структура – это совокупность “концептов”, которые сами являются максимально несводимыми причинно-следственными репертуарами, связанными каузальными отношениями. Причинно-следственный репертуар определяет то, как физические единицы, реализующие концепцию, ограничивают вероятность входных и выходных состояний комплекса в причинно-следственном пространстве. Общая несводимость концептуальной структуры измеряется интегральной информацией “большой фи” (Φ max), а концептов – “малой фи” (φ max). Связь концептов между собой осуществляется через такие же максимально нередуцируемые отношения.
Основываясь на этих аксиомах и постулатах, ТИИ выдвигает центральное тождество теории – утверждение, что субъективный опыт идентичен концептуальной структуре. Согласно теории это означает, что каждое свойство существующего опыта должно соответствовать какому-то свойству существующей концептуальной структуры, и наоборот.
Название теории происходит от используемого в ней специфического понятия информации, смысл которого принципиально расходится с классическим понятием Шенноновской информации. В ТИИ вводится представление о внутренней информации. Она отождествляется с описанным выше понятием концептуальной структуры – формы в причинно-следственном пространстве. Эта форма, заданная элементами ФСС, и есть интегрированная информация – содержание сознания (Tononi, 2004, 2008). Такое понятие информации в корне отличается от традиционных представлений о “кодировании”, “обработке” и “передаче” информации в нейронауках и психологии (Oizumi et al., 2014; Tononi, Koch, 2015; Tononi et al., 2016). ТИИ утверждает, что концептуальную структуру (внутренняя информация) невозможно передать через канал связи (внешняя или Шенноновская информация), поскольку ее информационная емкость превышает пропускную способность любого канала. Поэтому сознательная информация согласно ТИИ – это не то, что может передаваться из одной части мозга в другую, как это постулируется, например, в теории ГНРП (Tononi et al., 2016).
В принципе, постулаты ТИИ могут быть использованы для определения того, является ли система элементов сознательной, насколько и каким образом. Однако в силу высокой вычислительной сложности интегрированной информации это невыполнимо даже для простейших из биологических нервных систем. В качестве приближения к решению этой задачи Тонони и соавт. был предложен индекс сложности возмущения (perturbation complexity index – PCI) (Casali et al., 2013), вычисляемый как нормализованная сложность пространственно-временного паттерна активации коры головного мозга, вызванного воздействием транскраниальной магнитной стимуляции. PCI вычисляет алгоритмическую сложность реакции мозга на возмущение и определяет через это два важных компонента сложности текущего состояния таламокортикальной системы: дифференцированность и интегрированность. Исследования с использованием PCI в качестве опосредованной меры интегрированной информации установили, что потеря и восстановление сознания связаны с распадом и восстановлением способности к интеграции информации по показателю PCI. Это было показано для различных состояний сна (Massimini et al., 2010), наркоза с использованием анестетиков с различными механизмами действия (Sarasso et al., 2015), а также у пациентов с травмами головного мозга (Casali et al., 2013). Однако были высказаны и критические замечания, что PCI не имеет прямого отношения к интегрированной информации и не может служить ее достоверным индикатором (Sitt et al., 2013).
Из данного описания видно, что ТИИ значительно отличается от других теорий своей математической ориентированностью и высокой степенью абстракции. Она проявляется, в частности, в приверженности теории аксиоматическому методу построения, а также в использовании при расчетах интегрированной информации гипотетического субстрата сознания в виде небольших систем из 3–4 логических элементов (Oizumi et al., 2014). Теория делает при этом глобальные выводы о сознании в природе (Tononi, Koch, 2015), в частности, заключение о возможности существования сознания у неорганических систем, таких как фотодиод (Oizumi et al., 2014), или у простых двумерных решеток из большого числа физических элементов (Tononi, 2017).
ТИИ предполагает, что сознание как интегрированная информация является внутренней сущностью системы и, в этом смысле, солипсично (Tononi, 2008). В принципе, оно может существовать само по себе, без всякой функции и цели, даже у полностью пассивной системы, не имеющей никаких контактов с внешним миром (Tononi, 2008). Однако в ходе эволюции, эпигенеза и отбора информационные отношения во внешнем мире могут канализировать эволюцию соответствующих им информационных отношений в системе, способствуя возникновению систем с более высокой интегрированной информацией (Tononi, 2012). Это утверждение теории проверялось в исследованиях на компьютерных моделях искусственных существ – аниматов, эволюционировавших на протяжении десятков тысяч поколений в виртуальной проблемной среде. Результаты симуляции подтвердили предсказание ТИИ: более адаптированные аниматы имели более высокий уровень интегрированной информации в их модельном мозге, и уровень интегрированной информации у аниматов был тем выше, чем выше была сложность среды (Albantakis et al., 2014; Albantakis, Tononi, 2015).
Однако при этом ТИИ не содержит в числе своих постулатов таких основополагающих функций когнитивных систем, как обучение и память (Tononi et al., 2016). Она также не предлагает решения вопроса: “кто” является субъектом в системах, обладающих сознанием, и как именно этот субъект возникает в индивидуальном развитии ( (г)).
3.5. Итоги анализа
Задачей настоящего раздела было оценить, в какой степени основные нейронаучные теории сознания предлагают решение для сформулированных нами вопросов. Рассмотрение с этой точки зрения четырех наиболее влиятельных теорий показывает, что:
1. Среди них есть те, которые удовлетворяют некоторым из ключевых требований, но нет ни одной, которая отвечала бы им всем.
2. Ни одна из рассмотренных теорий, за исключением теории селекции нейронных групп, не пытается сколько-либо последовательно связать эволюционные механизмы в филогенезе, развитие нервной системы в эмбриогенезе, преобразование нейронных сетей при обучении и процессы генерации субъективного опыта у взрослого организма, чтобы объяснить, почему сознание закономерно формируется в результате эволюции, развития и обучения.
3. Особенно заметен дефицит всех теорий в области решения проблемы “кто”: научного объяснения устройства и возникновения носителя субъективного опыта – генератора и реципиента состояний сознания.
4. СОЗНАНИЕ И КОГНИТОМ
Располагая всем полученным материалом, вернемся к задачам настоящей работы. Ее целью являлось нахождение путей принципиального решения “главной проблемы” нейронауки – проблемы разум-мозг (Mind-brAIN problem – MAIN problem). Как мы видели, современные искания ответов на этот вопрос сосредоточились в области “трудной проблемы” сознания. Оценка наиболее часто обсуждаемых нейронаучных теорий сознания показала при этом, что ни одна из них не отвечает всему кругу сформулированных нами требований. Исходя из жесткой формулировки феномена “циркулярной ловушки”, никакая из этих теорий не дает удовлетворительного объяснения сознания. Проанализируем, чего именно не хватает в них для решения проблемы?
4.1. Фундаментальная сущность когнитивных систем
Отметим прежде всего, что рассмотренные теории не обладают фундаментальностью. Правда, теория интегрированной информации претендует на нее, но ценой столь существенной трансформации самого понятия сознания, что оно оказывается распределенным и по неорганическим системам, существовавшим еще до возникновения жизни (Tononi, 2008, 2017b; Tononi, Koch, 2015; Koch, 2012, 2019). В итоге теория объясняет нечто, но это вовсе не то, что подразумевается под сознанием другими исследователями или соответствует общечеловеческому пониманию этого феномена (Searle, 2013; Cerullo, 2015).
Наиболее приближается к критерию фундаментальности и решению проблемы “циркулярной ловушки” теория селекции нейронных групп. Она делает попытку вывести возникновение сознания из биологических закономерностей эволюции, развития и обучения организмов, обладающих нервной системой, гомеостазом и физиологическими потребностями. Эти системы взаимодействуют с окружающей средой в процессах соматического и естественного отбора, меняющих их морфологию и структуру их нервной системы. Таким образом, эта теория выводит сознание из биологии и не прибегает к физическому фундаментализму (Edelman, 2003). Поэтому мы можем выбрать ее в качестве прототипа фундаментальной теории в обозначенном нами ранее смысле и проанализировать, каких именно объяснений в ней недостает для полного соответствия нашим требованиям.
Коротким заключением будет, что эта теория, как и все остальные, обладает “слепым пятном” в области проблемы “кто”. Как только речь заходит о том, где разворачиваются когнитивные, ментальные, сознательные процессы, теория обращается к языку анатомических областей и физиологических процессов нервной системы, причем лишь нервной системы млекопитающих. Она оперирует понятиями возбуждения нейронных групп, корреляции клеточных сигналов в слоях, полотнах и нейронных цепях коры, таламических ядер, стриатума, гиппокампа, других подкорковых структур, активности в связывающих их нейронных проекциях. Этот вполне осознанный шаг теория предпринимает, чтобы изгнать из мозга гомункулуса – мистического “кто”, обитающего в пространстве мозга. Теория строится на убеждении, что при наличии процессов отбора в рекуррентных нейронных цепях головного мозга “не нужно вызывать гомункулуса, маленького человечка, живущего в мозгу, чтобы интерпретировать значение восприятия” (Edelman, 2004, c. 46–47).
Аналогичное отношение к проблеме “кто” свойственно и другим теориям, за исключением некоторых взглядов Ф. Крика и К. Коха (Crick, Koch, 2000, 2003, Koch, 2004). “Ужас гомункулуса” (Edelman, 1992, p. 79) выступает устрашением для подавляющего большинства нейронаучных исследователей сознания. Основания для этого очевидны: проблема quis custodiet ipsos custodes – кто наблюдает за наблюдающим, проблема регресса в бесконечность. Выход из этого видится единственным – убить гомункулуса или изгнать его из нервной системы. Но необходимо проследить за тем, что далее логически следует из этого решения.
Разберем это на примере теории селекции нейронных групп. Сознание после изгнания гомункулуса представляется в ней как процессы нейрофизиологического уровня, протекающие в “динамической сердцевине” таламокортикального комплекса. Только они, обозначаемые как С'-процессы, могут обладать, согласно теории, причинной силой. А процессы сознания – C-процессы, которые сопровождают С’-процессы, “сами не могут быть причинными” (Edelman, 1992, c. 78). “Таким образом, мы должны заключить, что наша вера в то, что сознание способно выступать причиной событий, представляет собой одно из полезных заблуждений. На самом деле тем, что движет индивидом и ментальными реакциями, является подлежащая нервная активность. … Философы находят этот набор заключений проявлением эпифеноменализма – что сознание ничего не делает. В действительности же оно служит для информирования нас о наших состояниях мозга и поэтому центрально для нашего понимания.” (Edelman, 2006, c. 92).
Если после знакомства с этими итогами изгнания гомункулуса складывается впечатление сгущающегося тумана, то, на мой взгляд, это ровно то, что на самом деле и происходит. Обратим внимание лишь на наиболее заметные признаки возникающих здесь противоречий: (1) причинную роль играют только нейрофизиологические С'-процессы, (2) а сознательные С-процессы сами не могут быть причинными, (3) но они при этом выполняют функцию информирования “нас” – т.е. того “кто”, который был изгнан как гомункулус из мозга, (4) о “наших”, т.е. “его” состояниях мозга, (5) и поэтому центральны для “нашего”, т.е. “его” понимания.
Другие иллюстрации неизбежности такого присутствия “кто” в определениях сознания были приведены в разделе 1.4.
В настоящей работе мы примем другой подход. Согласно нему, проблема “кто” не решается ни умерщвлением гомункулуса, ни изгнанием его из нервной системы, ни запретом на использование психологических понятий при описании операций мозга, ни сведением психологических процессов к нейрофизиологическим. “Кто” – это не просто совокупность анатомических структур и физиологических процессов центральной нервной системы. “Кто” – это новый фундаментальный уровень существования биологических систем, поднимающий их до категории когнитивных систем. Проблема “кто” может быть действенно решена только реконструкцией этого пути, проделанного природой – логическим выведением специфических структур и процессов субъективного уровня из закономерностей эволюции, индивидуального развития и поведенческих операций нервной системы у адаптирующихся биологических организмов.
Иными словами, “кто” должен быть не изъят из мозга, а объяснен специальной теорией, адекватно охватывающей его сущность во всей ее специфичности и причинно-следственной силе. Понятия этой теории, сохраняя погруженность в структуры и процессы биологического уровня, должны отражать качественную специфику структур и процессов когнитивного уровня, точно так же, как понятия молекулярной биологии, сохраняя погруженность в структуры и процессы химического уровня, отражают суть структур и процессов биологического уровня. И эти новые понятия теории должны находить эквиваленты среди сложившихся на основе обыденного опыта терминов, обозначающих субъективные явления и структуры.
Итак, мы имеем две разные парадигмы. Одна – распространенное сегодня объяснение сознания и субъективных процессов на языке анатомии и физиологии мозга, а также методологически связанный с этим поиск нервных коррелятов сознания. Вторая – поиск теории особой высокоуровневой организации мозга и протекающих в этой структуре специфических процессов, которые составляют фундаментальную нередуцируемую сущность когнитивных систем.
Принципиальная разница между двумя этими системами взглядов нелегка для понимания и ускользает от восприятия порой даже выдающимися исследователями. Особенно пагубной для продвижения в решении проблемы сознания оказывается следующая ошибка категоризации: а) имеется понимание бесперспективности первого подхода; б) однако отсутствует понимание сути второго подхода и его кардинального отличия от первого; в) из-за этого осуществляется перенос отрицания первого подхода на второй, а вернее, в силу непонимания качественного отличия второго, на нейронаучный подход в целом.
Чтобы яснее обозначить критическую разницу между выделенными подходами, я рассмотрю ее на конкретном примере – принципиальном различии парадигм физиологии высшей нервной деятельности и теории функциональных систем, составлявшем предмет напряженных, а нередко и ожесточенных дискуссий на протяжении значительной части прошлого века.
4.2. Физиология высшей нервной деятельности и теория функциональных систем
Целью заложенной И.П. Павловым новой дисциплины – физиологии высшей нервной деятельности – являлось строгое научное исследование того, “каким образом материя мозга производит субъективное явление” (Павлов 1951, с. 247). Методологической основой физиологии высшей нервной деятельности выступало прямое соотнесение психологических понятий с классическими рефлекторными физиологическими процессами, а ее концептуальной основой – их отождествление. И.П. Павлов считал, что “здесь имеется полное слитие, полное поглощение одного другим, отождествление” (Павлов, 1949, с. 521). Поведение организмов при этой логике определялось не будущими событиями психологического уровня – целями, а прошлыми событиями физиологического уровня – стимулами из внутренней и внешней среды. Согласно известному утверждению И.П. Павлова, “надо показывать пальцем: где было раздражение, куда оно перешло? Если вы живо себе это представите, тогда вы поймете всю силу и правду учения об условных рефлексах, которое совершенно исключило из своего круга психологические понятия, а все время имеет дело только с объективными фактами, т.е. с фактами, существующими во времени и в пространстве.” (Павлов, 1949, c. 385).
Таким образом, физиология высшей нервной деятельности представляла собой первую из разобранных в предыдущем разделе стратегий. Соответственно, она обладала и всеми свойственными ей недостатками, характерными также и для современных нейронаучных теорий сознания. В рамках этой парадигмы механизмы детерминации поведения оставались чисто физиологическими, а психологическая сфера в канве этих физиологических реакций оказывалась параллельным и поэтому вынужденно недейственным бытием. Решение “главной проблемы” в этих условиях сводилось либо к редукции психологии к физиологии, либо к признанию причинной незначимости, эпифеноменальности психики, либо к дуализму, той или иной версии психофизического взаимодействия.
Теория функциональных систем создавалась в качестве альтернативы этому подходу (Анохин, 1926, 1949, 1973, 1974). Она явилась развитием павловской теории условных рефлексов, но одновременно и ее отрицанием, что обусловило многие исторические особенности ее становления.
Разработка теории функциональных систем исходила из того, что “между классической физиологией и психологией … всегда существовала пропасть, к заполнению которой почти не делалось никаких попыток. … Как бы ни были тонки и искусны все наши экспериментальные достижения по линии аналитической физиологии, как бы мы точно ни изучили отдельные фазы возбуждения и его последовательных состояний, мы никогда не сможем только с помощью этого вооружения перебросить мост между физиологией и психологией, … если мы не произведем некой промежуточной операции, заключающейся в таком синтезе всего физиологического материала, который помог бы увидеть принципы, свойственные только целостной организации. Попытка создания такого промежуточного понятия была сделана в нашей лаборатории в виде понятия функциональной системы. … Функциональной системой мы называем всякую организацию нервных процессов, в которой отдаленные и разнообразные импульсы нервной системы объединяются на основе одновременного и соподчиненного функционирования, заканчивающегося полезным приспособительным эффектом для организма. … По своему масштабу функциональные системы организма могут быть весьма различны. Одни из них охватывают огромные комплексы процессов нервного и гуморального характера, как например, дыхательная система, другие сведены до незначительного движения одним-двумя пальцами по направлению к какому-либо предмету.” (П.К. Анохин, 1947, с. 32).
Деятельность целого организма в соответствии с этими взглядами представлялась производной от многообразных функциональных систем. Репертуар из таких сформированных в фило- и онтогенезе специализированных систем определяет поддержание баланса внутренней среды организма и его взаимодействие с окружающим миром. В качестве единицы такого взаимодействия в теории выделялся элементарный поведенческий акт – базовый цикл соотношения целостного организма со средой, в котором рассматриваются не афферентные и эфферентные, связанные с локальными анатомическими субстратами процессы, а системные процессы организации активности элементов различной анатомической принадлежности в единое целое, поведенческую функциональную систему (П.К. Анохин, 1973; Швырков, 2006). Формирование таких систем в эволюции, эмбриональном развитии и при обучении подчиняется особым закономерностям не органо- или морфогенеза, а системогенеза, теория которого является специальным разделом общей теории функциональных систем (П.К. Анохин, 1948; Швырков, 2006).
С точки зрения задачи решения “главной проблемы”, в теории функциональных систем следует выделить три принципиальных момента:
1) Согласно теории, достижение организмом приспособительных поведенческих результатов обеспечивается специфическим комплексом общемозговых механизмов. Эти системные процессы, включающие афферентный синтез, принятие решения, формирование акцептора результата действия и программы действия, само действие и получение его результата, обратную афферентацию от него, ее сличение с акцептором результата действия и последующую модификацию исходной системы были объединены в теории понятием операциональной архитектоники поведенческого акта. Важным тезисом теории является принципиальный изоморфизм операциональной архитектоники любых поведенческих актов. Это означает, что те или иные психические и сознательные процессы могли получать свое эволюционное развитие, только вписываясь в то или иное место системной архитектоники адаптивного поведенческого акта (П.К. Анохин, 1949, 1973, 1974).
2) Ключевым понятием в теории является приспособительный результат функциональной системы. Эта извлекаемая из индивидуальной памяти модель знакомого, но еще не наступившего в среде события, выступает системообразующим фактором для организации всех исполнительных механизмов функциональной системы. За счет этого именно накопленное в памяти “опережающее отражение” (П.К. Анохин, 1962) будущих соотношений со средой, то есть субъективный опыт, определяет поведение когнитивных организмов и влияет таким образом на их адаптивную эволюцию. Это делает детерминацию их поведения психической, определяемой их опытом, а не физиологической, определяемой рефлекторными процессами в нервной системе, как постулировалось в парадигме физиологии высшей нервной деятельности.
3) С позиции теории функциональных систем подлежащими нейрофизиологическому исследованию становятся не пути проведения возбуждения в нервной системе, не афферентные и эфферентные, сенсорные и моторные структуры, не процессы в тех или иных “центрах”, а закономерности организации клеточных физиологических процессов в качественно специфические системные процессы, отражение в этой организации структуры среды, детерминация этой организацией двигательной активности организма и т.п. Таким образом, кардинально меняется сама проблематика изучения нервных основ субъективного опыта и сознания (Швырков, 2006). Разработанная далее В.Б. Швырковым системно-эволюционная теория применила эти принципы к экспериментальному изучению клеточных основ субъективного опыта (Швырков, 2006). Основным итогом этих исследований стали данные, что отдельные нейроны различных областей мозга животных специализированы относительно не каких-либо отдельных физиологических или психологических функций, а целостных элементов субъективного опыта. Это в частности означало, что изучение подобных нейронных специализаций может служить методом объективного изучения структуры субъективного опыта, а анализ активности нейронов с известной специализацией – объективным методом изучения его динамики (Швырков, 2006). Предлагаемый ниже подход к решению “главной” и “трудной” проблем является прямым продолжением и развитием этой линии.
4.3. Когнитом: сознание в нейронной гиперсети мозга
Выше был подвергнут критике подход, удаляющий гомункулуса из объяснения сознания, за возникающий при этом “разрыв в объяснении” (Levine, 1983) – туман в зоне контакта нейрофизиологических концепций с представлениями о “кто” носителе, генераторе и реципиенте субъективных процессов. Будем справедливыми и отметим и противоположный полюс проблемы: для конкретно мыслящего нейроморфолога, нейробиолога или нейрофизиолога разговоры о нередуцируемой сущности ментальных явлений, качественной специфике системных процессов не находят достаточно конструктивной и надежной опоры в его знаниях о нервной системе.
Главный тезис настоящей работы состоит в том, что в рамках существующих сегодня представлений об устройстве и работе мозга это и невозможно. Чтобы это смогло произойти, вначале требуется радикальная перестройка наших представлений о самом мозге. Иными словами, нужна новая фундаментальная теория мозга – теория, позволяющая увидеть строение и работу мозга на максимуме его причинно-действенного потенциала, не раскрытого пока в современных нейробиологических представлениях.
Таким образом, основное заключение настоящей работы по текущей проблемной ситуации состоит в том, что устранить существующий “разрыв в объяснении” мозга и разума можно только подняв представления о мозге на этот нередуцируемый до традиционной анатомии и физиологии качественно новый уровень, где он становится тождественным представлениям о разуме.
Подробное изложение такого рода теории будет дано в специальной публикации. Здесь же перечислена лишь тезисная последовательность шагов, необходимых для решения как “главной проблемы” разум-мозг, так и “трудной проблемы” сознание-мозг. Для этого потребуется:
− во-первых, провести отчетливую демаркацию между понятиями разума (mind) как специфической структуры и сознания (consciousness) как специфического процесса, протекающего внутри этой структуры;
− во-вторых, научиться ясно различать проблемы “разум и мозг” и “сознание и мозг” как две принципиально разные проблемы, одна из которых о соотношениях понятий о двух структурах, а вторая о процессах, протекающих в определенной структуре;
− в-третьих, принять установку, что проблема “разум и мозг” должна быть решена прежде, чем проблема “сознание и мозг”;
− в-четвертых, увидеть, что основным сегодняшним препятствием на пути решения проблемы “разум и мозг” является кажущееся самоочевидным, но, тем не менее, ошибочное представление о мозге как о типовом биологическом органе, сложной нейронной сети, сущность которой может быть исчерпывающе описана в понятиях нейроанатомии и нейрофизиологии;
− в-пятых, осознать, что истинная природа мозга может быть понята только исходя из принципа, согласно которому истинная сущность сложных многоуровневых систем всегда определяется лишь верхним из их уровней – тем, который осуществляет их максимальные по мощности причинно-следственные взаимодействия;
− в-шестых, понять, что исходя из этого принципа, мозг в своей максимально нередуцируемой сущности является когнитивным органом, состоящим из специфических когнитивных элементов, объединенных специфическими когнитивными связями и реализующим специфические когнитивные процессы и взаимодействия обладающего им когнитивного агента – то есть разумом (mind);
− в-седьмых, сформулировать на основе данного понимания новую теорию мозга, описывающую этот качественно специфический уровень его организации и когнитивного функционирования, не сводимый к традиционным понятиям нейроанатомии и нейрофизиологии;
− в-восьмых, опираясь на эту новую теорию, заново переформулировать традиционные вопросы проблемы “сознание и мозг”, соотнося теперь все субъективные феномены не с физиологическими процессами в нейронной сети – коннектоме, а с когнитивными процессами в составляющей максимальную сущность мозга нейронной гиперсети – когнитоме.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 19-115-50540.
Список литературы
1. Аллахвердов В.М. Сознание как парадокс. СПб: ДНК, 2000. 528 с.
2. Анохин К.В. Когнитом: теория реализованных степеней свободы мозга. Пятая международная конференция по когнитивной науке: Калининград, 2012. Т. 2. с. 429–430.
3. Анохин К.В. Когнитом: гиперсетевая модель мозга. XVII Всероссийская научно-техническая конференция с международным участием “Нейроинформатика-2015”, 2015. Сб. научн. тр. в 3 частях. Под общ. ред. Трофимова А.Г. с. 14–15.
4. Анохин П.К. Диалектический материализм и вопросы “психического”. Человек и природа. 1926. 1: 81–90.
5. Анохин П.К. Теория функциональной системы как основа для понимания компенсаторных процессов организма. Ученые зап. МГУ, Психология. 1947. т. 2. 32–41.
6. Анохин П.К. Системогенез как общая закономерность эволюционного процесса. Бюлл. эксп. биол. мед. 1948. 8 (2): 81–99.
7. Анохин П.К. Узловые вопросы в изучении высшей нервной деятельности. Проблемы высшей нервной деятельности. М., 1949. с. 9–128.
8. Анохин П.К. Опережающее отражение действительности. Вопр. филос. 1962. 7: 91–111.
9. Анохин П.К. Принципиальные вопросы общей теории функциональных систем. Принципы системной организации функций. М.: Наука, 1973. с. 5–61.
10. Анохин П.К. Роль системного аспекта в разработке пограничных проблем нейрофизиологии и психологии. Нейрофизиологические механизмы психической деятельности человека. Под ред. Бехтеревой Н.П. Ленинград: Наука, 1974. с. 10–16.
11. Васильев В.В. Трудная проблема сознания. М.: Прогресс-Традиция, 2009. 272 с.
12. Выготский Л.С. Исторический смысл психологического кризиса. М. ЭКСМО-Пресс. 1008 с.
13. Декарт Р. Избранные произведения. М.: Государственное издательство политической литературы, 1950. 712 с.
14. Декарт Р. Сочинения в двух томах. Т. 2. М.: Мысль, 1994. 640 с.
15. Дубровский Д.И. Проблема сознания: опыт обзора основных вопросов и теоретических трудностей. Проблема сознания в философии и науке. Под ред. Дубровского Д.И. М.: Канон+, 2009. С. 5–52.
16. Дубровский Д.И. Проблема “сознание и мозг”: теоретическое решение. М.: Канон+, 2015. 208 с.
17. Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза. Журнал высшей нервной деятельности им. И.П. Павлова. 1996. 46 (2): 241–252.
18. Павлов И.П. Избранные произведения М., Л.: Издательство Академии наук СССР, 1949. 650 с.
19. Павлов И.П. Полное собрание сочинений. Т. 2, книга 2. М., Л.: Издательство Академии наук СССР, 1951. 589 с.
20. Пономарев Я.А. Методологическое введение в психологию. М. Наука, 1983. 205 с.
21. Проблема сознания в философии и науке. Под ред. Дубровского Д.И. М.: Канон+, 2009. 472 с.
22. Сергин В.Я. Психофизиологические механизмы осознания: гипотеза самоотождествления. Журнал высшей нервной деятельности им. И.П. Павлова. 1998. 48 (3): 558–571.
23. Сергин В.Я. Автоотождествление паттернов нейронной активности как физиологический механизм осознания. Журнал высшей нервной деятельности им. И.П. Павлова. 2016. 66 (2): 1–20.
24. Соколов Е.Н. Очерки по психофизиологии сознания. М.: МГУ, 2010. 255 с.
25. Фреге Г. Мысль. Логическое исследование. М.: 1997. 128 с.
26. Эйнштейн А. Физика и реальность. М.: Наука, 1965. 359 с.
27. Энгельс Ф. Диалектика природы. К. Маркс и Ф. Энгельс. Полное собрание сочинений. Т. 20. М.: Издательство политической литературы, 1961.
28. Швырков В.Б. Введение в объективную психологию. Нейрональные основы психики. Избранные труды. Под ред. Александрова Ю.И. М: Институт психологии РАН, 2006. 592 с.
29. Albantakis L., Hintze A., Koch C., Adami C., Tononi G. Evolution of integrated causal structures in animats exposed to environments of increasing complexity. PLoS Comp. Biol. 2014. 10 (12): e1003966.
30. Albantakis L., Tononi G. The intrinsic cause-effect power of discrete dynamical systems – from elementary cellular automata to adapting animats. Entropy. 2015. 17 (8): 5472.
31. Andrews P.L.R., Darmaillacq A.-S., Dennison N., Gleadall I.G., Hawkins P., Messenger J.B., Osorio D., Smith V.J., Smith J.A. The identification and management of pain, suffering and distress in cephalopods, including anaesthesia, analgesia and humane killing. J. Exp. Mar. Biol. Ecol. 2013. 447: 46–64.
32. Baars B. A cognitive theory of consciousness. Cambridge University Press. 1988. 424 p.
33. Baars B. In the theater of consciousness: The workspace of the mind. New York: Oxford University Press, 1997. 193 p.
34. Bartolomei F., Naccache L. The global workspace (GW) theory of consciousness and epilepsy. Behav. Neurol. 2011. 24 (1): 67–74.
35. Bateson P., Laland K.N. Tinbergen’s four questions: an appreciation and an update. Trends Ecol. Evol. 2013. 28 (12): 712–718.
36. Bayne T. Access consciousness. The Oxford companion to consciousness. Eds. Bayne T., Cleeremans A., Wilken P. Oxford: Oxford University Press, 2009. 1–3 pp.
37. Beshkar M. The QBIT Theory of Consciousness. Integr Psychol Behav Sci. 2020. 54 (4): 752–770.
38. Birch J., Schnell A.K., Clayton N.S. Dimensions of animal consciousness. Trends Cogn. Sci. 2020. 24 (10): 789–801.
39. Block N. On a confusion about a function of consciousness. Behav. Brain Sci. 1995. 18 (2): 227–247.
40. Block N. Consciousness. In: The Oxford companion to the mind. Ed. Gregory R.L. Oxford: Oxford University Press, 2004. 1024 p.
41. Block N. Response to Kouider et al.: Which view is better supported by the evidence? Trends Cogn. Sci. 2012. 16 (3): 141–142.
42. Bolhuis J.J., Giraldeau L-A. Introduction: Mechanisms of animal behaviour. In: The behavior of animals: Mechanisms, function and evolution, Eds. Bolhuis J.J., Giraldeau L-A. (Blackwell, Oxford, 2005. 1–10.
43. Brenner S. Turing centenary: Life’s code script. Nature. 2012. 482 (7386): 461.
44. Burghardt G.M., Bekoff M. Animal consciousness. The Oxford companion to consciousness. Eds. Bayne T., Cleeremans A., Wilken P. Oxford: Oxford University Press, 2009.
45. Carls-Diamante S. The octopus and the unity of consciousness. Biol. Philos. 2017. 32: 1269–1287.
46. Carruters P. Higher-order theories of consciousness. The Blackwell companion to consciousness. Eds. Schneider S., Velmans M. Chichester: John Wiley & Sons, 2017. 848 p.
47. Casali A.G., Gosseries O., Rosanova M., Boly M., Sarasso S., Casali K.R., Casarotto S., Bruno M.A., Laureys S., Tononi G., Massimini M. A theoretically based index of consciousness independent of sensory processing and behavior. Sci. Transl. Med. 2003. 5 (198): 198ra105.
48. Cerullo M.A. The problem with Phi: A critique of integrated information theory. PLoS Comput Biol. 2015. 11 (9): e1004286.
49. Chalmers D.J. Facing up to the problem of consciousness. J. Conscious. Stud. 1995. 2, 200–219.
50. Chalmers D.J. The conscious mind: in search of a fundamental theory. Oxford: Oxford University Press, 1996. 432 p.
51. Chalmers D.J. The character of consciousness. New York: Oxford University Press, 2010. 596 p.
52. Chen C., Martínez R.M., Cheng Y. The developmental origins of the social brain: Empathy, morality, and justice. Front. Psychol. 2018. 9: 2584.
53. Cohen M.A., Dennett D.C. Consciousness cannot be separated from function. Trends Cogn. Sci. 2011. 15 (8): 358–364.
54. Crick F. Neural Edelmanism. Trends Neurosci. 1989. 12 (7): 240–248.
55. Crick F., Koch C. Towards a neurobiological theory of consciousness // Seminars in Neurosci. 1990. 2: 263–275.
56. Crick F. The astonishing hypothesis. The scientific search for the soul. New York: Scribner, 1994. 336 p.
57. Crick F., Koch C. Consciousness and neuroscience. Cereb. Cortex. 1998. 8: 97–107.
58. Crick F., Koch C. The Unconscious Homunculus. Neuro-Psychoanalysis. 2000. 2: 2–59.
59. Crick F., Koch C. A framework for consciousness. Nat. Neurosci. 2003. 6 (2): 119–126.
60. Crook R.J., Hanlon R.T., Walters E.T. Squid have nociceptors that display widespread long-term sensitization and spontaneous activity after bodily injury. J. Neurosci. 2013. 33 (24): 10021–10026.
61. Damasio A. The feeling of what happens: body and emotion in the making of consciousness. New York: Harcourt Brace, 1999. 386 p.
62. Damasio A. Self comes to mind: constructing the conscious brain. New York: Pantheon Books, 2010. 367 p.
63. Darwin C. Charles Darwin’s notebooks, 1836–1844. Cambridge: Cambridge University Press, 1988. 747 p.
64. de Waal F.B.M., Preston S.D. Mammalian empathy: behavioural manifestations and neural basis. Nat. Rev. Neurosci. 2017. 18 (8): 498–509.
65. Dehaene S. Signatures of consciousness. In: The Mind. Ed. Brockman J. New York: Harper Perennial, 2011. 217–238.
66. Dehaene S. Consciousness and the brain. Penguin Books. 2015. 336 p.
67. Dehaene S., Changeux J.P. Ongoing spontaneous activity controls access to consciousness: a neuronal model for inattentional blindness. PLoS Biol. 2005. 3 (5): e141.
68. Dehaene S., Changeux J.P. Experimental and theoretical approaches to conscious processing. Neuron. 2011. 70 (2): 200–227.
69. Dehaene S., Naccache L. Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition. 2001. 79 (1–2): 1–37.
70. Dennett D.C. Consciousness explained. Boston: Little, Brown and Company, 1991. 511 p.
71. Eccles J. A unitary hypothesis of mind-brain interaction in the cerebral cortex. Proc R Soc Lond B Biol Sci. 1990. 240 (1299): 433–451.
72. Edelman D.B., Baars B.J., Seth A.K. Identifying hallmarks of consciousness in non-mammalian species. Conscious. Cogn. 2005. 14 (1): 169–187.
73. Edelman D.B., Seth A.K. Animal consciousness: a synthetic approach. Trends Neurosci. 2009. 32: 476–484.
74. Edelman G.M. Group selection and phasic reentrant signaling: a theory of higher brain function. The Mindful Brain: cortical organization and the group-selective theory of higher brain function. Eds. Edelman G.M., Mountcastle V.B. Boston: MIT Press, 1978. 51–98.
75. Edelman G.M. Neural Darwinism: The theory of neuronal group selection. New York: Basic Books, 1987. 240 p.
76. Edelman G.M. Topobiology: An introduction to molecular embryology. New York: Basic Books, 1988. 371 p.
77. Edelman G.M. The remembered present: A biological theory of consciousness. New York: Basic Books, 1989. 346 p.
78. Edelman G.M. Bright air, brilliant fire. On the matter of the mind. New York: Basic Books, 1992. 280 p.
79. Edelman G.M. Neural Darwinism: Selection and reentrant signaling in higher brain function. Neuron. 1993. 10 (2): 115–125.
80. Edelman G.M. Naturalizing consciousness: a theoretical framework. Proc. Natl. Acad. Sci. U.S.A. 2003. 100 (9): 5520–5524.
81. Edelman G.M. Wider than the sky. The phenomenal gift of consciousness. New Haven: Yale University Press, 2004. 201 p.
82. Edelman G.M. Second nature. Brain science and human knowledge. New Haven: Yale University Press, 2006. 203 p.
83. Edelman G.M., Gally J.A., Baars B.J. Biology of consciousness. Front. Psychol. 2011. 2: 4.
84. Edelman G.M., Gally J.A. Reentry: a key mechanism for integration of brain function. Front. Integr. Neurosci. 2013. 7: 63.
85. Edelman G.M., Tononi G. Consciousness: How matter becomes imagination. London: Allen Lane, 2000. 274 p.
86. Einstein A., Infeld L. The evolution of physics. London: The Scientific Book Club, 1938. 335 p.
87. Fiorito G., Scotto P. Observational learning in Octopus vulgaris. Science. 1992. 256(5056): 545–547.
88. Ginsburg S., Jablonka E. The evolution of the sensitive soul: Learning and the origins of consciousness. Cambridge: MIT Press, 2019. 640 p.
89. Godfrey-Smith P. Other minds: The octopus, the sea and the deep origins of consciousness. New York: Farrar, Strauss and Giroux, 2016. 257 p.
90. Gray J. Consciousness: Creeping up on the Hard Problem. Oxford: Oxford University Press, 2004. 341 p.
91. Griffin D.R. The question of animal awareness: evolutionary continuity of mental experience. New York: Rockefeller University Press, 1981. 209 p.
92. Grossberg S. Adaptive Resonance Theory: how a brain learns to consciously attend, learn, and recognize a changing world. Neural Netw. 2013. 37: 1–47.
93. Güntürkün O., Bugnyar T. Cognition without cortex. Trends Cogn. Sci. 2016. 20 (4): 291–303.
94. Gutfreund Y. The mind-evolution problem: the difficulty of fitting consciousness in an evolutionary framework. Front. Psychol. 2018. 9: 1537.
95. Haig D. Proximate and ultimate causes: How come? and What for? Biol. Philos. 2013. 28: 781–786.
96. Harris C.R., Prouvost C. Jealousy in dogs. PLoS One. 2014. 9 (7): e94597.
97. Hjelle L.A., Ziegler D.J. Personality theories: Basic assumptions, research, and applications. New York: McGraw-Hill Book Company, 1992. 603 p.
98. Hunt T., Schooler J.W. The easy part of the hard problem: A resonance theory of consciousness. Front. Hum. Neurosci. 2019. 13: 378.
99. James W. The writings of William James: a comprehensive edition. Chicago: University of Chicago Press, 1978. 912 p.
100. Kemmerer D. Are we ever aware of concepts? A critical question for the Global Neuronal Workspace, Integrated Information, and Attended Intermediate-Level Representation theories of consciousness. Neurosci. Conscious. 2015. 2015 (1): niv006.
101. Keysers C., Gazzola V. A plea for cross-species social neuroscience. Curr. Top. Behav. Neurosci. 2017. 30: 179–191.
102. Kingsbury L., Huang S., Wang J., Gu K., Golshani P., Wu Y.E., Hong W. Correlated neural activity and encoding of behavior across brains of socially interacting animals. Cell. 2019. 178 (2): 429–446. e16.
103. Koch C. The quest for consciousness: a neurobiological approach. Englewood: Roberts & Company Publishers, 2004. 429 p.
104. Koch C. Consciousness. Confessions of a romantic reductionist. The MIT Press, 2012. 181 p.
105. Koch C. The feeling of life itself: why consciousness is widespread but can’t be computed. Cambridge: MIT Press, 2019. 280 p.
106. Koch C., Massimini M., Boly M., Tononi G. Neural correlates of consciousness: progress and problems. Nat. Rev. Neurosci. 2016. 17 (5): 307–321.
107. Kouider S., Sackur J., de Gardelle V. Do we still need phenomenal consciousness? Comment on Block. Trends Cogn. Sci. 2012. 16 (3): 140–141.
108. Kuba M.J., Byrne R.A., Meisel D.V., Mather J.A. When do octopuses play? Effects of repeated testing, object type, age, and food deprivation on object play in Octopus vulgaris. J. Comp. Psychol. 2006. 120 (3): 184–190.
109. Lamme V.A.F. Towards a true neural stance on consciousness. Trends Cogn. Sci. 2006. 10 (11): 494–501.
110. Lenoble F., Carlier P. A possible contribution of phenomenology to ethology: application to a behaviour pattern in the mouse. Acta Biotheor. 1996. 44 (1): 75–83.
111. Levine J. Materialism and qualia: the explanatory gap. Pacif. Philos. Quart. 1983. 64(4): 354–361.
112. Levy J., Vidal J.R., Fries P., Démonet J.F., Goldstein A. Selective neural synchrony suppression as a forward gatekeeper to piecemeal conscious perception. Cereb. Cortex. 2016. 26 (7): 3010–3022.
113. Mashour G.A., Roelfsema P., Changeux J.P., Dehaene S. Conscious processing and the global neuronal workspace hypothesis. Neuron. 2020. 105 (5): 776–798.
114. Massimini M., Ferrarelli F., Murphy M., Huber R., Riedner B., Casarotto S., Tononi G. Cortical reactivity and effective connectivity during REM sleep in humans. Cogn. Neurosci. 2010. 1 (3): 176–183.
115. Massimini M., Tononi G. Sizing up consciousness: Towards an objective measure of the capacity for experience. Oxford: Oxford University Press, 2018. 224 p.
116. Mather J. What is in an octopus’s mind? Anim. Sentience. 2019. 26: 1.
117. Mather J.A. Cephalopod consciousness: behavioural evidence. Conscious. Cogn. 2008. 17 (1): 37–48.
118. Mather J.A., Anderson R.C. (1999) Exploration, play and habituation in octopuses (Octopus dofleini). J. Comp. Psychol. 113: 333.
119. Mayr E. Cause and effect in biology. Science. 1961. 134: 1501–1506.
120. McClelland T., Bayne T. Concepts, contents, and consciousness. Neurosci. Conscious. 2016. 2016 (1): niv012.
121. McGinn C. The Mysterious flame: Conscious minds in a material world. Basic Books, 2000. 256 p.
122. Metzinger T. Being no one: The self-model theory of subjectivity. Cambridge: MIT Press, 2003. 714 p.
123. Metzinger T. The ego tunnel: The science of the mind and the myth of the self. New York: Basic Books, 2009. 276 p.
124. Miller G.A. Psychology: the science of mental life. New York: Harper & Row, 1962. 388 p.
125. Nagel T. What is it like to be a bat? Philos. Rev. 1974. 83 (4): 435–450.
126. Nesher N., Levy G., Grasso F.W., Hochner B. Self-recognition mechanism between skin and suckers prevents octopus arms from interfering with each other. Curr. Biol. 2014. 24 (11): 1271–1275.
127. Nesse R.M. Tinbergen’s four questions: Two proximate, two evolutionary. Evol Med Public Health. 2018. 2019 (1): 2.
128. Noel J.P., Ishizawa Y., Patel S.R., Eskandar E.N., Wallace M.T. Leveraging nonhuman primate multisensory neurons and circuits in assessing consciousness theory. J. Neurosci. 2019. 39 (38): 7485–7500.
129. Noy N., Bickel S., Zion-Golumbic E., Harel M., Golan T., Davidesco I., Schevon C.A., McKhann G.M., Goodman R.R., Schroeder C.E., Mehta A.D., Malach R. Ignition’s glow: Ultra-fast spread of global cortical activity accompanying local “ignitions” in visual cortex during conscious visual perception. Conscious Cogn. 2015. 35: 206–224.
130. Oizumi M., Albantakis L., Tononi G. From the phenomenology to the mechanisms of consciousness: Integrated information theory 3.0. PLoS Comp. Biol. 2014. 10(5): e1003588.
131. Panksepp J., Panksepp J.B. Toward a cross-species understanding of empathy. Trends Neurosci. 2013. 36 (8): 489–496.
132. Penrose R. Shadows of the mind. A search for the missing science of consciousness. Oxford: Oxford University Press, 1994. 457 p.
133. Sarasso S., Boly M., Napolitani M., Gosseries O., Charland-Verville V., Casarotto S., Rosanova M., Casali A.G., Brichant J.F., Boveroux P., Rex S., Tononi G., Laureys S., Massimini M. Consciousness and complexity during unresponsiveness induced by propofol, xenon, and ketamine. Curr. Biol. 2005. 25 (23): 3099–3105.
134. Schlicht T. Phenomenal consciousness, attention and accessibility. Phenomenol. Cogn. Sci. 2012. 11 (3): 309–334.
135. Schnell A.K., Amodio P., Boeckle M., Clayton N.S. How intelligent is a cephalopod? Lessons from comparative cognition. Biol. Rev. Camb. Philos. Soc. 2020. Epub.
136. Searle J. The mystery of consciousness. New York: The New York Review of Books, 1997. 224 p.
137. Searle J.R. Consciousness. Annu. Rev. Neurosci. 2000. 23: 557–578.
138. Searle J.R. Can information theory explain consciousness? New York Review of Books, 2013. https://www.nybooks.com/articles/2013/01/10/can-information-theory-explain-consciousness/
139. Sergent C., Dehaene S. Neural processes underlying conscious perception: experimental findings and a global neuronal workspace framework. J. Physiol. Paris. 2004. 98 (4–6): 374–384.
140. Seth A.K., Baars B.J., Edelman D.B. Criteria for consciousness in humans and other mammals. Conscious. Cogn. 2005. 14: 119–139.
141. Seth A.K., Izhikevich E., Reeke G.N., Edelman G.M. Theories and measures of consciousness: an extended framework. Proc. Natl. Acad. Sci. U.S.A. 2006. 103 (28): 10799–10804.
142. Sitt J.D., King J.R., Naccache L., Dehaene, S. Ripples of consciousness. Trends Neurosci. 2013. 17: 552–554.
143. Spunt R.P., Adolphs R. The neuroscience of understanding the emotions of others. Neurosci. Lett. 2019. 693: 44–48.
144. Sutherland N.S. The international dictionary of psychology. New York: Crossroad, 1996. 515 p.
145. The Blackwell companion to consciousness. Eds. Schneider S., Velmans M. Chichester: John Wiley & Sons, 2017. 848 p.
146. The neurology of consciousness. Eds. Laureys S., Gosseries O., Tononi G. New York: Elsevier, 2015. 462 p.
147. The Oxford companion to consciousness. Eds. Bayne T., Cleeremans A., Wilken P. Oxford: Oxford University Press, 2009.
148. Tinbergen N. On aims and methods of ethology. Z. Tierpsychol. 1963. 20: 410–433.
149. Tononi G., Edelman G.M. Consciousness and complexity. Science. 1998. 282 (5395): 1846–1851.
150. Tononi G. An information integration theory of consciousness. BMC Neurosci. 2004. 5 (1): 42.
151. Tononi G. Consciousness as integrated information: a provisional manifesto. Biol. Bull. 2008. 215 (3): 216–242.
152. Tononi G. Integrated information theory of consciousness: an updated account. Arch. Ital. Biol. 2012. 150 (2–3): 56–90.
153. Tononi G. Why Scott should stare at a blank wall and reconsider (or the conscious grid). 2014. http://www.scottaaronson.com/blog/?p=1823
154. Tononi G. The integrated information theory of consciousness. An outline. The Blackwell companion to consciousness. Eds. Schneider S., Velmans M. Chichester: John Wiley & Sons, 2017. 243–256.
155. Tononi G. Integrated information theory of consciousness. Some ontological considerations. The Blackwell companion to consciousness. Eds. Schneider S., Velmans M. Chichester: John Wiley & Sons, 2017. 621–633.
156. Tononi G., Boly M., Massimini M., Koch C. Integrated information theory: from consciousness to its physical substrate. Nat. Rev. Neurosci. 2016. 17 (7): 450–461.
157. Tononi G., Koch C. Consciousness: here, there and everywhere? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015. 370(1668): 20140167.
158. Turing A. Computing machinery and intelligence. Mind. 1950. LIX (236): 433–460.
159. Van Bourg J., Patterson J.E., Wynne C.D.L. Pet dogs (Canis lupus familiaris) release their trapped and distressed owners: Individual variation and evidence of emotional contagion. PLoS One. 2020. 15 (4): e0231742.
160. Young A.W. Dissociable aspects of consciousness. The science of consciousness: psychological, neuropsychological and clinical reviews. Ed. Velmans M. London: Routledge, 2003. 118–139.