Системная нейрофизиология (nan)
Карта местности в мозге
Редактировать |
Электроэнцефалография (ЭЭГ) — широко распространённый и достаточно мощный инструмент изучения активности головного мозга, связанной с различными состояниями организма. Различия между сном и бодрствованием, направленным вниманием и состоянием «рассеянности» были выявлены именно с использованием этого метода. ЭЭГ используется также для изучения пространственного ориентирования и запоминания во время передвижения, однако к человеку этот метод до недавнего времени применять было невозможно из-за множества помех, возникающих при записи ЭЭГ с кожи головы свободно перемещающегося испытуемого. Если у животных допускается снимать энцефалограмму непосредственно с поверхности черепа или даже с коры мозга, то аналогичные исследования на людях не допустимы с точки зрения этики. Исследователи из Калифорнийского университета (США) справились с этой проблемой и выяснили, что параметры электрической активности мозга движущихся людей и грызунов, перед которыми стоит задача исследовать незнакомую местность, весьма схожи.
То, как мозг «обходится» с пространством, по каким принципам строит карту местности, задает скорость перемещения животного, в основном изучалось и изучается на излюбленных модельных объектах нейробиологов — грызунах, чаще всего мышах и крысах. Разнообразие методик велико, благо этика исследований на животных хотя и становится более жёсткой в последние годы, тем не менее допускает проведение операций по вживлению электродов в мозг абсолютно здоровой мыши или крысы или постановку этих электродов непосредственно на поверхность мозговой коры. Электрод, задача которого — регистрация разности потенциалов между определённым участком мозга и «нулевой» (референтной) точкой, должен находиться всё время в одном и том же положении относительно последней, чтобы его показания были верными. Если регистрирующий электрод погружен в ткань мозга, проблемы его перемещения относительно нулевой точки не возникает: плотность расположения нейронов и нейроглии не дает ему сдвинуться с места. К тому же сигнал, воспринимаемый погруженным электродом, достаточно сильный, ведь он фактически возникает там же, где регистрируется.
Проблема закрепления электрода возникает, если он расположен, скажем, на поверхности головы. Скальп, даже человеческий, с его атрофированными подкожными мышцами, подвижен, и даже небольшие изменения выражения лица (не говоря уже о ходьбе и тем более беге, которые у животных в подобных опытах вполне допустимы) вызывают нежелательные смещения электродов на коже головы и как следствие — искажение регистрируемых ими сигналов (так называемые артефакты). К тому же сигнал от нервных клеток, снимаемый со скальпа, ослабляется, ведь, прежде чем попасть на электрод, ему приходится преодолевать толщу мозговой ткани, тонких и грубых оболочек мозга, черепа и, наконец, самой кожи головы. Таким образом, снимать показания мозговой активности у свободно передвигающихся людей фактически не представляется возможным: вся запись из-за обилия движений испытуемого будет состоять только из артефактов.
Впрочем, технологии не стоят на месте, и команда калифорнийских нейрофизиологов смогла найти решение проблемы артефактов записи энцефалограммы при движении. Результаты этой работы были опубликованы в одном из недавних номеров Journal of Neuroscience.
Зачем вообще нужно проводить столь методологически трудоёмкие исследования на человеке? Наверное, наиболее общий ответ — чтобы подтвердить, что закономерности работы мозга, открытые на модельных животных, применимы и к нам самим. Известно, что у грызунов представление о пространстве и скорость передвижения связаны с амплитудой и вариациями частоты тета-ритма — изменения суммарной разности потенциалов клеток гиппокампа. Различные группы клеток гиппокампа, поочерёдно генерируя нервные импульсы, кодируют так называемую «карту пространства» — spatial map в англоязычной литературе. Электрод, расположенный в толще гиппокампа или на поверхности коры или скальпа, регистрирует суммарное изменение потенциала, вызываемое деятельностью не одного нейрона, а их популяции из нескольких тысяч клеток, соседствующих между собой. Сигнал, улавливаемый таким образом, называется потенциалом ближнего поля (local field potential). Параметры таких потенциалов отражают свойства формируемых карт пространства.
Приматы, к которым относимся и мы с вами, отличаются по части «навигации» от грызунов как минимум по двум параметрам. Во-первых, у обезьян значительно большую долю ощущений составляет информация от органов зрения, на которую они полагаются сильнее, чем грызуны («конёк» последних — обоняние). Во-вторых, неокортекс у приматов развит значительно лучше, чем у грызунов.
Эти две особенности строения ЦНС обезьян приводят к тому, что в формировании пространственных карт у них задействован не только гиппокамп, но и кора больших полушарий, конкретнее — теменная кора, поскольку она получает информацию от первичных зрительных областей неокортекса (см.: Мозг сортирует зрительные образы не так, как считалось ранее, «Элементы», 23.08.2013). С точки зрения удобства регистрации мозговой активности это хорошо, поскольку теменная кора находится ближе к поверхности мозга и головы, чем гиппокамп, а значит, сигналы от нее будут более сильными. При построении карт пространства приматы руководствуются внешним видом окружающей обстановки, своим расположением в видимом участке пространства, а также расстоянием между объектами по траектории передвижения.
Исследователи сделали синхронные записи электроэнцефалограммы, движений глаз, головы и тела. Для регистрации движений глаз (саккад) воспользовались электроокулографическим методом, для регистрации движений тела применялась трёхмерная видеозапись с помощью оптоэлектронных сенсоров и датчиков инерции. Испытания проходили в два этапа.
В первый день испытуемые должны были за 10 минут изучить комнату размером 4 на 5 метров, построенную в системе виртуальной реальности и смоделированную как помещение авианосца. Вдоль стен и на полу комнаты были расположены 39 различных предметов. Затем испытуемые в течение пяти сессий по 10 минут выполняли задания. Предметы, находившиеся в комнате, от них скрыли непрозрачными серыми шарами, и они должны были по очереди подходить к тем из них, которые подсвечивались (в случайном порядке) зеленым цветом. От «прикосновения» шарик пропадал, и участник опыта видел, что находится перед ним. В качестве дополнительного отвлекающего задания нужно было еще оценивать интересность увиденных предметов. Сессия заканчивалась, когда испытуемый обследовал все 39 предметов. Для каждой сессии порядок обхода предметов выбирался случайно, но сами предметы оставались на своих местах. Это обеспечивало разнообразие маршрутов испытуемых внутри комнаты и лучшее запоминание расположения предметов.
На второй день участникам снова предстояло пройти пять сессий. Каждый раз все предметы были закрыты шариками, а часть из них перемещали на новые места (в каждой сессии перемещались разные наборы предметов). Испытуемые снова должны были по очереди подходить к подсвеченным зеленым шарикам и смотреть, какой предмет скрывался за шариком. После этого им нужно было ответить, находится ли этот предмет на том же месте, что и в первый день, или он перемещен. Также нужно было оценивать степень уверенности в ответе.
Таким образом, изначально перед испытуемыми не стояло задачи запоминать расположение объектов. Такое обучение называется unsupervised learning, в русском варианте — обучение без учителя. Однако для данного эксперимента скорее было важным то, что человек обучался выполнять задачу (вспомнить расположение предмета) спонтанно, без предварительной установки на запоминание информации специально для этого задания. Такой тип обучения предъявляет особенно большие требования к пространственной памяти и невозможен без использования карты пространства, которая, как было сказано выше, формируется в том числе и в теменной коре. Поскольку объём полученных данных, которые еще нужно было очищать от многочисленных артефактов, был велик, ученые анализировали только те записи электроэнцефалограммы, которые были получены с электрода, расположенного на центральной линии черепа над теменной корой (обозначается Pz помеждународной системе 10-20). К тому же, точка наложения этого электрода находится дальше остальных от источников энцефалографических помех (глаз, мимических мышц и пр.).
Исследователи выбирали те отрезки энцефалограммы, которые были записаны во время движения испытуемых, разбивали их на участки одинаковой фиксированной длины и «вычитали» из усреднённой записи с точки Pz то, что было зарегистрировано окулографическими электродами. Моменты, в которые испытуемый двигался, определялись сравнением синхронизированных записи ЭЭГ и информации от датчиков движения (рис. 2).
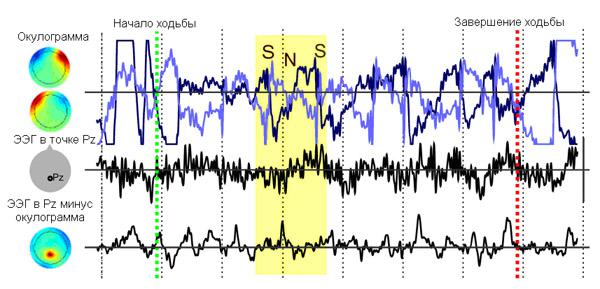 Рис. 2. Пример записи ЭЭГ испытуемого во время ходьбы (длительность 3 секунды) и ее очистка от артефактов. Слева — гистограммы распределения выраженности исследуемого ритма ЭЭГ, в данном случае тета-ритма, по поверхности скальпа. Точки, в которых данный ритм выражен наиболее сильно, имеют красный цвет, наименее сильно — синий. S — саккада (резкое и быстрое небольшое движение глаз), N — поворот головы вслед за направлением саккады (взгляд при этом остается на месте). Момент саккады и поворота головы выделен желтым. Изображение из обсуждаемой статьи вJournal of Neuroscience
|
Далее требовалось соотнести «карту» комнаты с освобожденной от артефактов записью тета-ритма. Для этого электроэнцефалограмму подвергали вейвлет-анализу на частоте тета-ритма — 6 Гц. С помощью этого анализа выявлялся так называемый знак ритма ЭЭГ (в нашем случае — тета-ритма). Далее этот полученный знак на отрывке энцефалограммы накладывали на траекторию движения испытуемого по комнате, которая, как было сказано ранее, регистрировалась с помощью видеокамер (рис. 3).
 Рис. 3. Траектория движения одного из испытуемых по комнате во время одной из экспериментальных проб с наложенными на неё знаками тета-ритма (красный изелёный). На примере отдельно выбранного участка пространства комнаты (врезка справа) демонстрируется, что знаки тета-ритма изменяются не спонтанно, а в зависимости от положения субъекта в пространстве комнаты. Участки траектории с одинаковыми знаками ритма образуют прямые линии, что также показано на врезке слева (красные и зелёные стрелки). Изображение из обсуждаемой статьи в Journal of Neuroscience |
Наложение знаков тета-ритма ЭЭГ на траекторию движения испытуемых выявило, что между упомянутыми параметрами существуют корреляции: знаки тета-ритма изменяются не спонтанно, а в зависимости от положения субъекта в пространстве комнаты. Участки траектории с одинаковыми знаками ритма образуют прямые линии, проходящие через всю комнату. При этом знак тета-ритма в конкретной точке комнаты не зависит от того, в какой момент времени испытуемый был там, но зависит от того, какой знак имел тета-ритм у этого человека близко к этому месту. Нечто похожее наблюдалось и при анализе активности «клеток места» у грызунов (см.: Представление о пространстве является врожденным, «Элементы», 21.06.2010).
Однако сила упомянутых корреляций была невысока, и нейробиологи решили провести дополнительный анализ полученных с теменной части головы записей. Во-первых, поскольку вейвлет-анализ энцефалограммы проводился на частоте 6 Гц, то, возможно, корреляции знаков тета-ритма этой ЭЭГ и положения человека в пространстве — «дело рук» конкретной частоты. В таком случае при наложении на траекторию движения испытуемого синусоидальной волны частотой 6 Гц корреляции с положением человека должны сохраниться. Однако этого не произошло, а значит, зависимость знаков тета-ритма от координат испытуемого была вызвана чем-то другим. Поэтому было проведено еще несколько тестов, из которых самым показательным было наложение ЭЭГ одного испытуемого на траекторию движения другого испытуемого.
Несмотря на то что по условиям задания участники эксперимента должны были проходить весьма близкие по длине участки пути и делать примерно одни и те же действия (рассматривать предмет, отмечать свой интерес к нему, оценивать, не был ли предмет перемещен на следующий день), порядок предметов, с которыми испытуемым нужно было провзаимодействовать, отличался, а значит, не были одинаковыми перемещения относительно объектов в комнате. Наложение ЭЭГ от одного человека на траекторию перемещения другого приводило к исчезновению корреляций между знаком тета-ритма и пространственным положением испытуемого в момент снятия энцефалограммы. Получается, зависимость знака тета-ритма от координат человека индивидуальна и не является артефактом.
Кроме того, по силе корреляций можно было определить, насколько хорошо человек запомнил положение предметов в первый день. У тех, кто лучше отвечал на вопросы типа «Был ли перемещен тот или иной объект во второй день?», зависимость знака тета-ритма ЭЭГ, записанной над теменной корой в первый день, от положения испытуемого относительно обстановки комнаты была более сильной. Однако такой зависимости между числом верных ответов и ЭЭГ, снятой во второй день уже во время воспроизведения запомненного, не было. Это говорит еще и о том, что карты пространства вносят свой вклад в работу памяти. Что, кстати говоря, согласуется с предыдущими данными похожих исследований на людях и на других приматах, а также на грызунах.
Разумеется, методы исследования, которые не требуют проникновения в организм испытуемого, такие как электроэнцефалография, еще во многом уступают по информативности инвазивным методам. Однако представленная работа — несомненный технический прорыв. С ее помощью удалось представить первое «живое» и полученное в максимально естественных условиях доказательство того, что соотношение тета-ритма ЭЭГ и положения организма в пространстве относительно других объектов при движении отражает работу групп клеток коры по построению карты пространства, окружающего субъект.
Источник: Joseph Snider, Markus Plank, Gary Lynch, Eric Halgren, Howard Poizner. Human Cortical Theta during Free Exploration Encodes Space and Predicts Subsequent Memory // The Journal of Neuroscience. September 18, 2013. V. 33(38). P. 15056–15068. Doi:10.1523/jneurosci.0268-13.2013.
Источник:
Чтобы оставить комментарии нужно авторизоваться.
Рис. 1. Экспериментальная обстановка. A1 — виртуальная модель комнаты, разработанная специально для эксперимента, A3 — как ее видит испытуемый. A2 — реальный вид помещения, в котором проводился эксперимент (справа — испытуемая в очках виртуальной реальности, слева — экспериментатор, который держит шнур одного из регистрирующих приборов). Изображение из обсуждаемой статьи в Journal of Neuroscience