Что такое Я - схемотехнический подход
Содержание книги основывается на постулате, что природная нейросеть мозга является схемотехнической структурой, т.е. ее работа и функциональность полностью соответствует причинно-следственным взаимодействиям элементов - в точности, как это можно сказать про схемотехнику электронного прибора - при всей огромной разнице в способах реализации.
Обзорная статья: «Что такое Я»
Что такое “Я” - схемотехнический подход
Основы схемотехники природных нейросетей
Петрийчук Николай Дмитриевич
Это – вторая редакция, дополненная данными схемотехнической реализации системы индивидуальной адаптивности на основе жизненных параметров (Beast): fornit.ru/65133.
Описание основных принципов эволюционного совершенствования индивидуальных систем адаптивности живых существ, природных или искусственных - в трилогии "Основы фундаментальной теории сознания": Рефлексы, Субъективное, Бессознательное
Аннотация
Содержание книги основывается на постулате, что природная нейросеть мозга является схемотехнической структурой, т.е. ее работа и функциональность полностью соответствует причинно-следственным взаимодействиям элементов - в точности, как это можно сказать про схемотехнику электронного прибора - при всей огромной разнице в способах реализации. Соответственно, для понимания механизмов нейросетевой схемы необходимо понимать схемотехнические принципы, а для этого нужно быть специалистом–схемотехником так же, как для понимания работы электрической схемы.
На основе схемотехнической концепции и совокупности фактических данных исследований мозга и действующего прототипа индивидуальной системы адаптивности на основе гомеостаза, представлена модель организации взаимодействий элементов мозга, описывающая иерархию эволюционно обусловленных механизмов, задачей которых является все более гибкая индивидуальная адаптивность к новому.
Сам факт создания целостной модели, обобщающей массив данных исследований во взаимосвязь адаптивных механизмов, показывает, что схемотехнический подход оказывается необходимым и достаточным для понимания и формализации всех известных проявлений работы мозга вплоть до психических.
Ключевые слова: психофизиология, нейросети, схемотехника
УДК 159.923
ББК 88.3
П30
Подготовка публикации:
Научный рецензент: Черноризов Александр Михайлович, доктор психологических наук, заслуженный профессор МГУ им. М.В. Ломоносова, заведующий кафедрой психофизиологии факультета психологии МГУ.
Разработчик демонстрационных моделей, участник обсуждений, автор схем и текстов в разделе “Первые управляющие структуры организмов на основе распознавателя”: Парусников Алексей Владимирович.
Художник: Петрийчук Михаил Евгеньевич.

Оглавление
Системы, системный подход, синергетика
О схемотехнике адаптивных нейросетей
Принципиальные элементы эволюции нервной системы
Эволюционное формирование нейросетевых элементов
Принцип выделения составляющих элементов воздействующих факторов.
Еще про распознаватели, кое-что по-настоящему важное
Резюмируем основные схемотехнические принципы
Первые управляющие структуры организмов на основе распознавателя
Химия электротоков и ее воплощение в нейросети
Схемотехнические принципы в модели нейрона
Использование схемотехнических механизмов
Дополнительное пояснение работы схемы компаратора на транзисторе в модели нейрона
Основной принцип организации природных нейросетей
Различие искусственных и природных нейросетей
Условия образования связей между нейронами
Конкретизируем условия образования связей
Причинно-следственная последовательность образования связей
Базовый механизм индивидуального поддержания жизнеспособности
Формируем обобщающую модель адаптивной нейросети
Принципы поддержания оптимального уровня параметра
Условия образования условного рефлекса
Общий механизм возникновения условного рефлекса
Новые реакции на основе старых
Необходимость повторения сочетаний условного и безусловного компонента реакции
Период доверчивого обучения, включающий период инициативы
Новое всегда противопоставляется привычному
О том, что такое цель, подробнее
Смысл, интерпретация, передача смысла
Явление субъективизации образов восприятия-значимости-действия
Автоматизмы, моторные и мыслительные
Свойства и особенности автоматизмов
Особенности мыслительных автоматизмов
Суть субъективного переживания
Контекст понимания смысла. Личность.
Как определить наличие сознания
Функционирование моделей понимания
Сопоставление отдельных смыслов в общий смысл
Произвольность порождается ориентировочным рефлексом
Информационная функция сновидений
Резюмируем уровни модели волевой адаптивности
Схема процесса осознания Beast
Роль слова среди других символов взаимопонимания
Практические следствия модели МВАП
Формирование осознанных навыков
Произвольность оценки происходящего
Современное состояние академической науки о психике
Список использованных источников:
Это – вторая редакция, дополненная данными схемотехнической реализации системы индивидуальной адаптивности на основе жизненных параметров (Beast): fornit.ru/65133.
Аннотация
Содержание книги основывается на аксиоме, что природная нейросеть мозга является схемотехнической структурой, т.е. ее работа и функциональность полностью соответствует причинно-следственным взаимодействиям элементов - в точности, как это можно сказать про схемотехнику электронного прибора - при всей огромной разнице в способах реализации. Соответственно, для понимания механизмов нейросетевой схемы необходимо понимать схемотехнические принципы, а для этого нужно быть специалистом-схемотехником так же, как для понимания работы электрической схемы.
Была осуществлена схемотехническая реализация (программным методом) действующего прототипа природной организации эволюционных механизмов индивидуальной адаптивности на основе жизненных параметров : fornit.ru/65133.
На основе схемотехнической концепции, созданного прототипа и совокупности фактических данных исследований мозга представлена уточненная модель организации взаимодействий элементов мозга, описывающая иерархию эволюционно обусловленных механизмов, задачей которых является все более гибкая индивидуальная адаптивность к новому.
Сам факт создания целостной модели, обобщающей массив данных исследований во взаимосвязь адаптивных механизмов, показывает, что схемотехнический подход оказывается необходимым и достаточным для понимания и формализации всех известных проявлений работы мозга вплоть до психических.
Предисловие
Особенности ссылок на исходные материалы в этой книге – использование коротких интернет-адресов, легко набираемых от руки, например: fornit.ru/13268. Это позволяет тут же посмотреть источник, а не испытывать трудности с классическим списком литературы. В книге много таких ссылок на ресурс fornit.ru, где собрано большое количество вспомогательных материалов и поясняющих текстов, которые никак не могут быть размещены в ограниченном объеме книги. Это не значит, что нужно обязательно переходить по таким ссылкам (их очень много), а смотреть только те, что заинтересуют (в точности как в любой научной статье ищут текст источника из списка литературы только при достаточной важности, но здесь это несравнимо удобнее). Много источников взяты из сторонних публикаций, а для удобства и надежности сохранены с короткими адресами. Вот несколько систематизированных источников фактических данных исследования и их обобщений: fornit.ru/ax1, fornit.ru/psy_arts, fornit.ru/journal, fornit.ru/R1, fornit.ru/1282.
Книга написана не в “академическом стиле” текстов по специальности и вот почему:
1. Это – междисциплинарная популяризация, т.е. минимизированы специальные термины и смысловые особенности, характерные для узкой предметной области.
2. Свободный текст легче воспринимается, он ассоциативен и более очевиден (о доказательных свойствах очевидности см. fornit.ru/7117).
3. Специализированный текст скрывает многие погрешности и создает иллюзию доказательности и обоснованности из-за эффекта распознания “научного текста”: срабатывает стереотип отношения к привычному, надежному и понятному. Академический стиль написания текстов позволяет легко протащить антинаучные (попирающие научную методологию) домыслы, вот как это проделал мистик П.Гаряев (fornit.ru/260), протащив статью в журнал ВАК, за что рвали волосы в президиуме Академии: fornit.ru/245. “Академический стиль” написания текстов легко имитируется (вот простая методика: fornit.ru/2750) даже компьютером, и в солидных журналах редакторы приняли такие сгенерированные, совершенно бессмысленные статьи потому, что они сразу распознавались как написанные “научно”, например: fornit.ru/6384.
4. Просто легче и приятнее читать изложенное литературным языком.
Достаточно очевидно, что вовсе не стиль определяет доказательность, а те доводы, которые представляются очевидно верными, что в случае заинтересованности позволяет самому проверить такие доводы, тем более что уже создан безальтернативный инструмент для моделирования системы адаптивных механизмов любой сложности, с помощью которого и разработана вторая версия прототипа (см. раздел “Новый инструмент”: fornit.ru/64975).
Этот инструмент позволяет строить ЛЮБЫЕ АДАПТИВНЫЕ СИСТЕМЫ на основе заданных жизненных параметров и верифицировать результат по соответствую ожидаемой функции так, что каждый может проверять свои представления об этом.
О чем эта книга
Эта книга представляет несколько разных аспектов понимания сути психики с ее феноменами так, что выходит за рамки традиционных предметных областей исследований, прежде всего потому, что в основе концепции взята аксиома о том, что природная нейросеть мозга является схемотехнической структурой (fornit.ru/41930), т.е. ее работа и функциональность полностью соответствует причинно-следственным взаимодействиям элементов (fornit.ru/24649) - в точности, как это можно сказать про схемотехнику электронного прибора - при всей огромной разнице в способах реализации.
Это в достаточной степени аксиоматически самоочевидно потому, что ничто, кроме причин и следствий взаимодействий в динамике времени не оказывает влияние на работу мозга. Мы сразу отметаем все, что не описывается физико-химической термодинамикой в рамках масштаба строения вещества, где все происходит в метрике пространства и времени (лапласовский детерминизм, но с учетом квантовомеханических явлений: fornit.ru/882). Всякие другие “квантово-голографические”, “трансперсональные”, “аксионные” и т.п. таинственные и туманные отсылки на возможную природу работы мозга отвергаются не только потому, что они неопределенны, не корректны и, по сути, не говорят ни о чем, а потому, что схемотехнический подход полностью исчерпывает необходимое описание и больше никаких лишних сущностей не требуется. Во всяком случае, в схемотехнической реализации действующего прототипа ничего такого не потребовалось.
В книге тесно взаимосвязывается суть абстракций (fornit.ru/103) и эволюция механизмов адаптивности (fornit.ru/1644), в которых на всех уровнях используются отвлечения от реальной действительности для оперирования ее коррелятами. Именно так устроена схема действующего прототипа.
Книга начинается с ознакомления со схемотехническими принципами, которые присутствуют в любых схемах причинно-следственных цепочек независимо от способов реализации. Система таких принципов составляет основу представлений специалиста-схемотехника. При этом для того чтобы этот базис воплотился в эффективный профессиональный опыт, необходимо, обычно, около 3-5 лет практики его применения. За этот срок из студентов получаются дееспособные специалисты и, к сожалению, нет способа уменьшить этот интервал времени потому, что знания из получаемых сведений формируются в процессе собственного применения (невозможно стать хирургом только по книжкам). В книге будет обосновано, почему именно это происходит при становлении индивидуальных способностей.
Это не должно отпугивать тех, кто не обладает навыками схемотехника. Как минимум можно получить понимание, т.е. “книжные знания” и составить целостную модель представлений. Одно дело – понимание принципов, на что требует гораздо меньше времени, а другое – практическое воплощение такого понимания в различных условиях в виде личных знаний. Понимание в основах, позволяющих охватить предлагаемую модель, возникает у заинтересованного читателя, даже если он не является схемотехником и специалистом в нейрофизиологии. Скорее, наоборот, неискушенный читатель легче воспримет материал, без проблем с сопоставлением с уже имеющимися у него знаниями и неизбежными моментами неприятия, когда это сопоставление покажется не в пользу предлагаемых утверждений.
Вот почему после первой главы, посвященной общим схемотехническим принципам, все будет даваться с корректным обоснованием источниками фактических исследований. Хотя в первой главе упоминаются те или иные свойства элементов нейросетей без корректного обоснования, воспринимайте это как бы авансом, в расчете на последующее закрытие всех спорных моментов. Поэтому просьба к нейрофизиологам: побудьте первое время достаточно снисходительными, принимайте спорные утверждения в качестве постулатов, и это потом окупится.
Чтобы представить, какие именно феномены психики станут доступными на уровне механизмов, вот – мотивирующая зарисовка, несколько снимающая напряжение первых настораживающих предупреждений.
Давайте зададимся вопросом: почему вот уже миллионы лет существует человек на Земле и, несомненно, где-то еще во вселенной и, наверняка, будет существовать еще долгое время, но только сейчас каждый из нас осознает себя? При том, что с яйцеклеткой матери слился один из миллионов сперматозоидов. А если бы слился не он и в другое время, а если бы отцом был совсем другой человек, мы бы осознали себя или оставались в небытии как много лет до этого и после этого?
Ни один самый крутой ученый сегодня не может внятно и вообще хоть как-то определенно ответить на этот вопрос, который уже содержит вполне очевидный ответ: да, мы бы существовали при любом раскладе родителей, да, мы существовали всегда и всегда будем существовать, мы существуем прямо сейчас во всех особях животных, обладающих определенным механизмом психики. Все дело в том, что такое – мы. И тут нет никакой мистики, в этой книге мы разберемся с этим с позиции методологии современной науки.
Все люди ощущают свою непостижимую исключительность бытия, и это – реальный феномен самосознания (fornit.ru/861). Такая исключительность - нечто общее, присущее всем независимо от реальных отличий, от любых отличий наследственности или условий развития. Такой вывод ясен формально, при сопоставлении вероятностей и здравого смысла, но пока непонятно, почему именно возникает этот феномен, так же как неясно, что же такое по сути самоощущение вообще.
В книге этот вопрос будет доведен до предельной определенности, но без обещания легкости понимания: слишком сложна и непривычна тема.
Попробуем несколько пояснить утверждения о специфике схемотехнического подхода, с которой, как правило, не сталкиваются нейрофизиологи.
Предположим, что человеку, не склонному к схемотехническому мышлению, поручено изучить работу марсианского компьютера, чтобы суметь создать такой же. Это – зримая аналогия с задачей изучить мозг. Исследователь становится очень хорошим специалистом по тому, что находится в компьютере, какие проводки откуда и куда идут. Он снимает ЭЭГ работы системного блока и вводит щуп для получения наводок в непосредственной близости к отдельным элементам, документируя явные отличия режимов работы. Он делает предположения об устройстве элементов, сошлифовывая их под микроскопом (именно так передирались зарубежные полупроводниковые приборы советскими плагиаторами). Он пишет научные отчеты и строит теории, но общее понимание никак не дается. Все его предположения основаны на обособленных фактах, не являющихся целостной системой, и разные исследователи предполагают очень различные гипотезы, в силу своей специфики и личных особенностей.
Теперь самое интересное. Этим исследователям попался настоящий марсианин – схемотехник компьютерных систем. И тут начинается чудесное: все попытки схемотехника что-то объяснить исследователям компьютера упираются в стену непонимания, ведь те не потратили годы на освоение принципов конструирования компьютеров и не понимают даже самые простые вещи в этой области.
В самом деле, разве есть шанс разобраться в работе сложнейших схем мозга, если возникают непреодолимые трудности даже в том, чтобы разобраться в простейшей схеме музыкального усилителя?
Для кого эта книга
Сказанное выше уже определяет, кому может быть интересна книга: всем, кто всерьез увлечен функциональностью мозга и тем, как организованы механизмы этой функциональности.
Эта книга может быть полезна:
· тем, кто интересуется механизмами адаптивной организации психики (а не просто попытками ее имитации с помощью искусственных нейросетей и семантических моделей)
· нейрофизиологам, которые хотят с большим качеством исследовать природные реализации мозга на разном уровне эволюции,
· и тем, кто просто хочет понять как устроена его субъективность.
При этом те, кто имеет хороший уровень подготовки средней школы, уже во многом смогут разобраться. Причем в некоторых случаях даже более быстро и эффективно, чем имеющие уже предвзятые мнения по описываемым вопросам. Так что неискушенность получает преимущества, если только человек имеет достаточный навык самостоятельного освоения нового и для него это чем-то похоже на игровые квесты прохождения по уровням достигнутого.
Так и стоит воспринимать текст книги: игровое прохождение квестов понимания до полной победы. Это – наиболее эффективная стратегия.
Для поддержания игрового контекста (механизмы которого, кстати, будут рассматриваться) нужно не напрягаясь, легко и со специфическим азартом-интересом вникать в сказанное, не опасаясь никаких негативных последствий.
Методология
Очень полезным было бы верифицировать свои наиболее общие представления с помощью книги “Мировоззрение”: fornit.ru/13268. На это потребуется примерно 10 дней, после чего можно читать эту книгу дальше.
Любой продукт осмысления содержит разного рода ошибки (fornit.ru/1012). Поэтому очень важны методы, критерии и принципы, минимизирующие ошибки.
Прежде всего, в основе рассуждений (сопоставления фактов и их обобщении в контексте нерешенной проблемы) должно лежать уже упомянутое добротное мировоззрение, в своих принципах адекватное объективной реальности.
Затем – научная методология, основывающаяся на мировоззренческом понимании взаимодействий причин и следствий объективной реальности и предоставляющая методы и принципы использования аксиоматической базы фактических данных для сопоставлений и обобщений.
Именно накопленная исследователями аксиоматика подхода к изучению нового станет опорой для формирования модели системы организации механизмов психики.
Решение проблемы понимания сути явления имеет несколько чередующихся стадий:
1) философское осмысление и выбор метрологического подхода (технологии измерений) для целенаправленного исследования.
2) исследование свойств и взаимодействий явления, исследование условий, в которых явление всегда воспроизводится.
3) сопоставление результатов исследования (чужих и/или своих) с их обобщением в виде субъективной модели взаимодействий.
4) экспериментальная проверка предположений и корректировка модели.
5) формализация модели в виде доступных для понимания другими специалистами символов.
Кстати, последний пункт прямо противоречит распространенному заблуждению о том, что математику следует использовать как инструмент познания, а не формализации уже имеющегося понимания. Но сначала понимание, а только потом адекватная ему формализация. Хотя это могут быть промежуточные понимания и их формализации.
Все эти пункты могут реализовываться в произвольной последовательности, т.е. по воле исследователя, в зависимости от представляющейся ему целесообразности. Так, если исследователь в большей степени экспериментатор, то он создает средства и методы для проверки чужих предположений. Если он - теоретик, то сосредотачивается на пунктах 2 и 5, а тот, кто начинает чуть ли не с нуля исследовать совершенно новое явление, проходит все пункты, не раз возвращаясь к предыдущим.
Понятно, что критически решающим оказывается пункт 3, эффективность реализации которого зависит от уровня подготовленности субъекта, его навыка творческих сопоставлений и обобщений. Это - уровень первичного понимания сути явления, и он не может быть реализован нигде, кроме как в чьей-то отдельной голове. Причем, как правило, в чьей-то одной голове из группы исследователей складывается наиболее полная и адекватная картина. Лишь потом следует межличностное изложение формулы понимания, что и лимитирует одновременность стадий понимания в нескольких головах даже самой сплоченной группы исследователей.
Для каждого из этих пяти пунктов характерны специфические принципы методологии и есть общие для всех.
Общими оказываются все принципы естественнонаучного мировоззрения, о которых уже шла речь и которые еще будут сформулированы.
Одним из важнейших общих принципов является подход выбора очередности рассмотрения проблемы или вариантов ее решения: из всех представимых вариантов объяснения сути явления для исследования в первую очередь нужно выбирать наиболее правдоподобное. Правдоподобность оценивается статистически (fornit.ru/7498) и эвристически (fornit.ru/7498). Этот принцип в частном виде сформулирован как "бритва Оккама" («Не следует привлекать новые сущности без крайней на то необходимости»), но не столь кардинально: не стоит что-то совсем "обрезать", а просто не нужно ставить в первую очередь. И если более вероятное окажется верным, тем более - без обнаружения чего-то более сложного, а при верификации и надежно подтвержденным, то остальное уже можно не рассматривать, это уже точно оказывается "лишней" сущностью.
В простых случаях выбор кажется очевидным, и мы скорее поверим, что наш знакомый не отдает долг из-за денежных затруднений, чем потому, что его захватили инопланетяне. Но есть ситуации, когда такой очевидности нет или человек слишком неопытен, чтобы ее увидеть, и тогда в голову приходят фантастические объяснения. Следует всегда иметь ввиду, что даже если не видно простого и рационального объяснения, оно может быть, но по каким-то причинам не приходит в голову, как механизмы поражающих воображение фокусов профессиональных престидижитаторов. Вот в таких случаях выручает творческая фантазия в попытках измыслить хоть какое-то правдоподобное предположение (в статусе неуверенного объяснения). И эта фантазия должна быть разумно ограничена и направлена (в чем и заключается сила творчества), иначе возникнет бесконечное множество самых чудесных предположений.
Стоит отметить важнейший принцип системности, позволяющий выявить необходимую и достаточную часть системы взаимодействующих факторов в искусственно выделенном из окружающего явлении: функциональность системы не зависит от второстепенного, не относящегося к ее причинно-следственным механизмам.
Этот подход реализуется в организации формирования субъективных моделей понимания, в которых с каждым новым фактом исследования все более уточняются существенные взаимосвязи и отсеивается все второстепенное и случайное.
Этим принципом многие пренебрегают, начиная исследование с дотошного и кропотливого выявления особенностей материалов реализации объекта внимания вместо того, чтобы понять, как именно это работает вне зависимости от того, из какого именно материала это сделано. Можно погрузиться в глубокое изучение метаболизма и особенностей реализации нервных и окружающих клеток, потому как другого фактического материала просто нет, но это имеет побочный эффект: особенности реализации заслоняют возможность увидеть принципиальную сторону функциональности.
Типичное заблуждение, проистекающее из игнорирования принципа системности – убеждение, что данные статистики являются доказательством наличия взаимосвязи. На самом деле ни единичное, ни многократные повторения каких-то результатов исследования не является доказательством наличия связи, т.е. причинно-следственной корреляции между какими-то параметрами, поэтому введен критерий шести сигм (fornit.ru/7506). В статье (fornit.ru/6612) показаны очень впечатляющие корреляции при явно абсурдных сопоставлениях, к примеру: “Влияние количества пиратов на глобальное потепление”. Это озадачивает многих, в том числе ученых, не задумывающихся о данном аспекте научной методологии: какие могут быть сомнения, когда в большом числе опытов с очевидностью показано совпадение. И разве такие данные не дают возможности использовать это в практических целях?
Но нет, статистика, какой бы она ни была сама по себе, не доказывает связь чего-то в явлении. Такую связь может доказать только понимание механизма взаимодействия – конкретно выявленных причин и следствий. И если не выявлен такой механизм, о связи говорить нельзя даже при надежности данный в 6 сигм, да сколько угодно сигм! Вот возьмем неоспоримо очевидное утверждение: “все, кто ест огурцы, умирают”. Практический вывод – огурцы смертельно опасны, их нельзя есть. Безупречная статистика есть, а механизма нет. Именно принцип причинно-следственного воздействия дает реально адекватное представление, которое можно включать в систему понимания происходящего в явлении, т.е. при формализации модели явления. Вот в чем заключается суть принципа системности.
Специально научной методологии, как и мировоззрению, мы не будем дополнительно уделять внимание, это – вынесенные за рамки книги, но совершенно необходимые представления. О научной методологии: fornit.ru/956.
И, наконец, для верификации модели необходимы критерии полноты и верности теории: fornit.ru/7649.
Обоснования утверждений
Книга основывается на обширной аксиоматике фактических данных исследований. Их взаимосвязанная система собрана в fornit.ru/ax1 и использована для обоснований отдельных обобщений при формировании модели организации механизмов психики.
Методом формирования модели представлений является отслеживание строгой последовательности эволюционно усложняющихся механизмов индивидуальной адаптивности, реализуемой структурами мозга.
Сегодня каждый год в мире выпускается несколько сотен тысяч статей по теме, и есть те, кто не упускает ничего из виду, пытаясь выбрать из этих тонн руды хоть что-то полезное. В частности, выдающийся популяризатор и инициатор консолидации в исследовании мозга К. Анохин (fornit.ru/1714, fornit.ru/8104) в докладе о положении науки о мозге за 2019 год очень пессимистически подвел итоги для возможного создания общей теории на этой основе.
И в самом деле, в этой предметной области возникает ощущение тупика, не только в результатах и направлении исследований, в стратегии исследований, но даже в терминологии. Вот слово “когнитом”, используемое К.Анохиным в качестве наиболее характеризующего мозг свойства: когнитивность (познание, сознание, разум). Но мозг простейших насекомых не обладает этим качеством (что станет предельно ясно в этой книге при рассмотрении того, какими именно механизмами обеспечивается это качество и вообще, что это такое), в то же время нейросеть любого мозга строится на совершенно идентичных базовых принципах и целевой функцией мозга является не разумность, а адаптивность, а для термина, обозначающего такое качество мозга, более подходит “адаптом”.
Что касается стратегии исследований, то принцип: “сначала личное понимание (это станет предельно очевидно при рассмотрении механизмов сопоставления и обобщения: fornit.ru/693), а потом формализация” не учитывается в попытках привлечь математику, в надежде обрести понимание по ее моделям, а математика – это всего лишь способ формализации уже понятого.
Если пренебрегать такой стратегией, то будут возникать казусы. Пример – публикации с удивительными утверждениями (неважно, где и кто):
“В среднем мозге и базальных ганглиях позвоночных существует несколько интегративных локусов, которые поддерживают обучение и память, так что организм может обновлять свои реакции на стимулы в соответствии со своим опытом. У насекомых эта способность поддерживается грибовидными телами (Bazhenov, Huerta, & Smith, 2013; Fahrbach, 2006; Galizia, 2014).
...Таким образом, существуют значительные параллели между функциональной организацией мозга насекомого и ядром контрольной поведенческой системы среднего мозга позвоночных.
...Как мы уже говорили, обработка (информации) такого рода поддерживает способность к субъективному восприятию окружающей среды. Обработка в головном мозге насекомых в одинаковой степени унифицирована по аналогичным причинам. По этой причине, мы предполагаем, что мозг насекомого также может поддерживать способность к субъективному опыту.”.
Авторы подобных работ (а их очень много в авторитетных источниках) не определяют, что такое “субъективное”, какими механизмами обеспечивается, какую функциональность выполняет, но применяют слова “субъективность,”, “сознание”, “разум” в качестве определенных понятий.
Можно выделить три методологических принципа, которые при этом нарушаются:
1. по сходству наблюдаемых качеств делается вывод о сходстве явления. Пример: если мяч синий, то другой синий предмет вовсе не обязательно – тоже мяч. Да и синий цвет вовсе не обязательно присущ мячу. Если насекомое корежит от воздействия огня, то это никак не значит, что оно ощущает боль.
2. по наличию первопричин двух явлений делается вывод о сходстве этих явлений. Пример: и дворец и канализация могут быть сделаны из кирпича, но это – совершенно разные вещи. А все вообще сделано из атомов. У высших животных есть уровень древности механизмов, в точности как у насекомых. Это – кирпичи, на основе которых строится более поздние механизмы, без которых субъективное не может существовать, но делать вывод, что раз у насекомых тоже есть древние элементы регуляция гомеостаза, то оно разумное – так же нелепо как говорить, что все из кирпичей – дворцы.
3. Никак не определенное явление (т.е. без понятия, что это такое) применяется в выводах о существовании. Аналогичное утверждение: “Обладают ли насекомые сепуляцией”? Какова должна быть сложность организма, чтобы утверждать такое? Классик сепуления С.Лем давно сформулировал трудную проблему сепуления в повести о звездных приключениях Иона Тихого: «... сепульки очень похожи на муркви, а своей цветовой гаммой напоминают мягкие пчмы. Разумеется, их практическая функция другая, но думаю, Вам, как человеку взрослому, мне не нужно этого объяснять». Опираясь на его теории, можно ли найти признаки сепуления у бабочки? У пчелы? (сепупярность насекомых == разумность насекомых).
Выявление любого из таких погрешностей выводов делает все рассуждение ложным, возможно намеренно ложным. Вот такого качества “обоснований” не будет в этой книге.
С валом работ по исследованиям мозга сегодня вполне возможно разобраться потому, что очень немалое число научных журналистов не пропускают хоть сколько-то многообещающую статью, публикуя свою интерпретацию вместе со ссылками на первоисточник. Вовсе не обязательно самому пытаться проделывать эту огромную коллективную работу, положив все свое время. Сегодня невозможно упустить никакой сколько-то важной новости, она обязательно появится, выделенная фокусом внимания среди остальной публикуемой руды. Но надеяться, что вдруг какая-то из новостей разом все прояснит, и телега легко покатится – нет ни малейшего основания. Все дело не в отдельных фактах, а в эффективном обобщении всего накопленного, и успех такого обобщения зависит от выбора верной точки отсчета.
За много лет из доступных материалов были сформированы большие тематические подборки (fornit.ru/ax1, fornit.ru/1282, fornit.ru/pm, fornit.ru/pp и др.). Это – такое огромное количество взаимодополняющих и ключевых фактов исследований, которые представляют богатый пазл, позволяющий с применением схемотехнического подхода собрать все в убедительную картину без дырок пазла в основе ветвей иерархии общей модели.
В сделанной попытке целостного обобщения в качестве обоснований для уверенности в каждом акте верификации утверждений выступает весь массив фактических данных исследований, обеспечивающих уверенность в достоверности и надежности каждого факта на уровне аксиоматики. Из элементов такого пазла и состоит предлагаемая модель.
Наиболее общий контекст книги
Определенный смысл тексту книги придадут основные аспекты рассмотрения проблематики, которыми можно уже сейчас определить понятие абстракции, понятие, понятие схемотехнического подхода. Все это самым тесным образом будет связано между собой в механизмах системы адаптивности, реализуемой мозгом.
Поэтому в первую очередь определимся с этими понятиями - как принимаемыми постулатами, и затем станет ясно, насколько оправдано и эффективно такое допущение. Сами эти постулаты возникли в результате немалого числа итераций процесса формирования модели.
Психика не бывает без мозга. Почему – вопрос мировоззренческий и здесь не будет рассматриваться, но это проясняется бескомпромиссно адекватно реальности в отдельной книге Мировоззрение, доступной по адресу fornit.ru/13268.
Итак, подходить к проблематике нужно с целевого назначения мозга. А мозг, прежде всего, выполняет управляющие функции, чтобы, используя сенсоры состояния внешней и внутренней среды, в зависимости от текущих условий организовывать адаптирующее поведение особи.
Так, глаз является по своей адаптивной функции оптическим устройством, позволяющим использовать оптические признаки изменений в окружающей среде, но большую часть этой функциональности возлагается на мозг, корректирующий зрительные признаки восприятия. Мозг сформирован для обеспечения общей адаптирующей функциональности, после глаза и других рецепторных органов и до эффекторных органов, обеспечивающих конечные фазы регуляции.
Вся эта система оптимизируется эволюционным отбором в направлении стабильности существования живых особей (fornit.ru/64924) и отдельных фрагментов этих особей (в принципе нет никакой разницы), так же тупо как вода течет туда, где пониже.
Сам по себе этот принцип развития любых систем (живых или неживых) дает очень мощное направление для обобщений и заслуживает отдельного рассмотрения, что и сделано в виде материалов:
· Эволюция живых существ на Земле - fornit.ru/1640
· Наследование признаков - fornit.ru/806
Другими аспектами, определяющими целостность и верность развития модели психики, будут понятия абстракции и контекстов. И все это – в рамках схемотехнического мышления (системы схемотехнических принципов и навыков их применения).
· Абстракции - fornit.ru/103
· Контексты - fornit.ru/610
· Особенности понимания схемотехнических систем - fornit.ru/24649
Понятие адаптивности нивелируется с таким подходом, лишаясь условности выделения именно живого и именно особей среди всего, претерпевающего взаимодействия причин и следствий. Но мы не можем рассматривать процессы на таком базовом уровне как взаимодействия и будем применять понятия вроде адаптивности, при этом не станем забывать про условность этих понятий. И эти условности придется умолчательно или явно иметь ввиду в контексте граничных условий применения терминов, чтобы смысл оставался достаточно определенным.
Поэтому чисто методологически всегда будем иметь ввиду условия корректности применяемых терминов: fornit.ru/1315
Почему именно схемотехнической подход?
Вопрос в том, какая из предметных областей может претендовать на модельное описание системы механизмов организации психики. Вот те, что считаются наиболее подходящими.
Философия давно и традиционно строит предположения в области пока еще недостаточных данных исследований и поэтому является донаучным инструментом первичного понимания (fornit.ru/920).
Математика – “царица всех наук”, на которую многие уповают как на средство, способное сгенерировать верную теорию системы взаимодействий механизмов мозга, на самом деле в принципе не продуцирует системные обобщения, а является лишь способом формализации и верификации уже имеющихся представлений. Невозможно обойти последовательность: сначала понимание, потом – формализация. Хотя математическое моделирование и способно создавать новые формы, в точности как формы дюн в пустыне от ветра, но их смысл (осознаваемая значимость) оказывается понимаем только после того, как такое понимание уже подготовлено субъективно (fornit.ru/693).
Психология изучает эмпирически найденные проявления психики, но их механизмами не занимается. Она собрала огромное количество фактических данных исследований проявлений психики, но ее теории не способны дать системную модель для описания механизмов и при всех упорных попытках не приближается к этому (fornit.ru/7258).
Нейрофизиология изучает все то, что относится к анатомии и функциональности нервных клеток и их связанных состояний. Она предоставила огромное количество фактических данных исследований всего, что касается нервных клеток и их роли в физиологических процессах организма, она выявила многие механизмы функций нервных клеток и их взаимосвязей, проследила связи между отдельными зонами мозга и то, на что такие связи влияют, но ее теории не способны дать системную модель для описания механизмов организации психики и при всех упорных попытках не приближается к этому.
Психофизиология призвана изучать механизмы психики на физиологическим уровне, т.е. используя данные нейрофизиологии, но оказалась неспособной к самостоятельности в принципиальном подходе и оказалась синонимом нейрофизиологии, даже ее частным случаем, ограниченном областью психики.
Все (ученые, следующие научной методологии) понимают общую функцию мозга как систему управления эффекторами в зависимости от состояния рецепторов. Ученые понимают, что любая управляющая система является взаимодействием механизмов причин и следствий, что относит ее к области схемотехнических систем управления. Но множество сомнений появляется на уровне того, что в мозге отнести к элементам принципиальных механизмов управляющих функций: то ли учитывать ДНК ядер клеток, то ли учитывать влияние других клеток, кроме нейронов, то ли считать принципиальным организацию взаимодействий между нейронами.
Но самое главное, что упускается, это то, что понять схему системы взаимодействий механизмов способен только профессионал схемотехник, а если они и есть среди ученых нейрофизиологов, то такие оставляют свой опыт схемотехники за рамками исследований нейрофизиологии потому, что они затрудняются выделить принципиальные элементы механизмов среди того, что обеспечивает их реализацию.
В самом деле, если попытаться объяснить нейрофизиологу, как устроен телевизор по его принципиальной схеме, то он не сможет ничего понять, сначала ему придется пройти путь развития схемотехнических представлений в течение многих лет наработки опыта понимания и личного использования. Но схемотехника мозга – еще сложнее схемотехники телевизора...
Нейрофизиолог видит множество видов нейронов, имеющих разный размер, разную форму, разные виды отростков и разные виды межнейронных соединений. Ему трудно отбросить предположение о разной функции столь различающихся элементов.
Если посмотреть на схемотехнику электроцепей, то там проводники, транзисторы, резисторы, конденсаторы и катушки могут иметь самую разную форму и размеры, различающиеся в сотни раз, но у них одна главная функциональность: проводники проводят электрический ток, транзисторы изменяют ток управляющим потенциалом, резисторы огранивают прохождение тока своим сопротивлением, конденсаторы – это емкости для зарядов, катушки – это индуктивности или накопители энергии полей электрического тока. То, в каком конструктиве все это крепится, так же оказывает влияние на работу схемы (как и влияние астроцитов и других конструктивных элементов мозга).
Схемотехник электроцепей знает, как меняются свойства элементов в зависимости от того, как они сделаны потому, что это определяет характеристики этих элементов в работающей схеме.
То же самое можно сказать и про схемы мозга.
Понимая это, возможно выделить основные функции элементов управляющих схем мозга от того, каким способом они реализованы и составить ту же управляющую функциональность на транзисторах, проводниках, сопротивлениях и т.п. или же реализовать схему управления с помощью программных алгоритмов.
Поэтому предметной областью для понимания функциональности психики является «Схемотехника адаптивных нейросетей», анонсированная в fornit.ru/41930.
Только схемотехнический подход дает возможность понять и формализовать в виде принципиальной схемы систему индивидуальной адаптивности на уровне психики. Он основывается на фактических данных исследований природных версий реализации (множества видов животных, обладающих психикой) и выделяет общие принципы схемотехнической функциональности, что и формализовано в виде модели МВАП .
Для того, чтобы дальнейшее изложение не оказалось поверхностным и расплывчато многозначительным, вызывающим протест непонимания или неприятия, необходимым условием является приведение базовых понятий к принимаемым условностям. Это значит, на этом рекомендуется отложить дальнейшее прочтение и прочесть материалы по приведенным ссылкам или оставить это на второй проход книги. Конечно, из-за сложности вопросов это может быть не вполне достаточно, но даст необходимый контекст понимания остального. В данной книге просто не получилось бы втиснуть тексты по всем этим темам, так что такое отвлечение оказывается вынужденным.
Системы, системный подход, синергетика
Понятие “система”, которое обычно используют в работах по нейробиологии, оказывается достаточно расплывчато, что прямо сказывается и в попытках формализовать системные представления о нейросетях.
Система – условная модель представлений о причинно-следственных взаимосвязях выделенных вниманием элементов, в граничных условиях абстрагирования этих элементов в реальном явлении. А такая модель и есть схемотехническое представление совокупности механизмов взаимодействия – формализованная схема взаимодействий, отражающая механизмы взаимодействия всех элементов.
Граничными условиями выделяется только то существенное, что относится к принципу взаимодействий, а не его конкретной реализации. Поэтому кроме принципиальных схем существуют конструкторские, монтажные и т.п. дополнительные схемы. В случае нейросети за границей рассмотрения принципиальной схемы системы оказываются особенности строения элементов нейросети, метаболизм и другие непринципиальные факторы.
Система оказывается целостной и верной (адекватной реальности) тогда, когда описываемая совокупность элементов и механизмов их взаимодействий позволяет практически воплотить такую систему в реальности, выбрав те или иные материалы и способ реализации механизмов взаимодействия, в заданной степени точности описания проявлений работы этой системы.
К примеру о роли выбора граничных условий, система гравитационного взаимодействия тел в пространстве, ограниченная точностью в несколько знаков, описывается формулами классической механики, где массы взаимодействуют обратно пропорционально квадрату расстояния между ними, что отражает трехмерное пространство, в котором площадь увеличивается по закону квадрата расстояния и, значит, так же ослабляется и гравитационное взаимодействие, определяемое именно метрикой пространства. Сами массы несколько изменяют метрику пространства (как и их относительная скорость), но для выбранной точности значений это несущественно. А если говорить не о гравитационном, а внутриядерном взаимодействии, то там метрика взаимодействий имеет большее число измерений, и взаимодействия ослабляются быстрее, чем от квадрата расстояния.
Выбор граничных условий для описания модели взаимодействий элементов нейросети определяется той принципиальной функцией задач адаптивности, которую мы выделяем как целевую роль системы для обеспечения приспособления к окружающим условиям.
Возникает критерий системности описаний нейросети: такая модель должна являться принципиальной схемой описания определенных адаптивных функций, что позволяет реализовать эту модель в виде электронного устройства, в виде программного продукта иди другими способами реализации.
Попытки системного подхода прослеживаются у многих исследователей мозга. У И.Павлова – сигнальные системы, у П.Анохина – функциональные системы.
Чем «схемотехнические» описания отличаются от просто системно ориентированных описаний? Схемотехнический подход это – и есть системный подход, но с обеспечением уверенности в практической реализации принципиальных схем. Если схема формализует предполагаемый механизм взаимодействий, то такая схема может быть реализована и будет работать в реальности, а не только в воображении.
Как схемотехнический подход выглядит на практике?
Представим картину изучения мозга с помощью МРТ-визуализации. На некоторой выборке живых людей в разных состояниях исследователи замечают закономерности возникновения и взаимного поведения активностей нейросети. Это коррелирует с определенными состояниями внимания и сознания. Вот пока и все полученные данные. Исследователи называют проявления таких активностей по их психо-коррелятам “пассивным режимом работы головного мозга (DMN)” и “задней системой внимания (DAT)”, а научные журналисты публикуют (fornit.ru/ext-2) очередной прорыв в понимании сознания и рассуждают про нейронные сети, ответственные за проявления сознания. Но, как уже замечалось на примере со смертью от огурцов, никакая статистическая корреляция не означает на самом деле существующих взаимодействий и, тем более, понимания механизмов взаимодействий.
На самом деле, никакого исследовательского прорыва от бессистемных наблюдений МРТ данных нет, есть просто некие факты наблюдений, а что это такое - так и неясно, хотя часто напрашиваются правдоподобные предположения. Зато показавшиеся важными факты названы научными терминами и становятся трендами в публикациях. Они ничего вообще не проясняют, кроме того, что дают повод выдвигать кучу новых предположений. Точно так же можно заметить в природе множество других закономерностей, ничего не приоткрыв в понимании их механизмов. Вот примеры того, как опасно вообще полагаться на наблюдаемые корреляции без поддержки найденных механизмов взаимосвязей (кроме смерти от огурцов): fornit.ru/6612.
Мы не будем делать предположения на основе корреляций, а будем прослеживать эволюционное усложнение конкретных механизмов адаптивности на уровне мозга от самых простых и, на их основе – всех последующих так, чтобы формировалась все более общая модель взаимодействующих механизмов, имеющих общую направленность – индивидуальная адаптивность. Это и будет построение схемотехнической модели, даже если при этом мы не будет рисовать схем мозга в виде условных элементов, а как-то по-другому формализуем механизмы. Но поначалу мы будем рисовать схемы, пока это позволяют границы допустимого небольшой книги с лаконичным изложением.
В результате все эти специфические нейросети просто уступят место совершенно конкретным функциональным механизмам, обеспечивающим индивидуальную адаптивность, и не будут привязываться к особенностям природной реализации, т.е. будут формализованы в виде принципиальных моделей механизмов взаимодействий.
Синергетика – еще одно популярное слово, которое представляется обязательным в привязке к теории, претендующей на достоверность. Синергетика — это динамика “самоорганизующихся” систем в терминах междисциплинарного описания. Вот именно это и является основой данной книги: описание в стиле междисциплинарной популяризации эволюции развития систем индивидуальной адаптивности и описание динамики этих систем в зависимости от условий.
Дело не в словах и терминах. И не в том, как разграничиваются предметные области исследований, бывает, до такой степени, что, изучая, по сути, одно и то же, например, живые организмы, каждая из дисциплин оказывается в своем изолированном мире специфических абстракций. Настолько, что требуется именно междисциплинарный подход для того, чтобы адекватно и целостно обобщить представления в верифицируемую реальностью модель.
Никакие отдельные биологические, математические, схемотехнические, нейрофизиологические и т.п. подходы в попытках сопоставить все многообразие природной реализации и вывести нечто принципиально общее – несостоятельны в силу недостаточности частного охвата. Раньше не было отдельной предметной области, которая начинает и выигрывает сама. Но схемотехнический подход, который базируется на данных фактических исследований нейробиологов и психофизиологов, позволяет строить действующие прототипы адаптивных явлений, его обобщения могут верифицировать не только нейробиологи и психофизиологи потому, что уже созданы не просто схемы, а действующая модель системы индивидуальной адаптивности на основе жизненных параметров (fornit.ru/65133), названная Beast.
Следующая глава – наиболее базовые основы схемотехнических принципов, которые необходимы для понимания организации нейросетей. Здесь не даются корректные обоснования утверждений, касающихся элементов нейросетей. Основное внимание предлагается уделить принципам, которые проявляются в любой схеме цепочек взаимодействия независимо от природы реализации причинно-следственных процессов.
Чтобы максимально облегчить понимание и сделать очевидными утверждения, в следующей главе используются ясные, зримые, знакомые всем процессы.
О схемотехнике адаптивных нейросетей
Точнее и правильнее было бы сказать "Схемотехника адаптивных систем", потому что реализация принципов индивидуальной адаптивности необязательно должна использовать нейросети (fornit.ru/67666).
Схемотехника — это то, что можно выделить вниманием в любых реальных конструкциях, в отличие от умозрительных. Это – план (структура, модель) реального воплощения таких конструкций, что возможно условно формализовать в виде схемы взаимодействий.
Самое важное – то, что любая схемотехника описывает конструкции на основе однотипных принципов, которые используются в самых разных их видах. В схемотехнике определяюще важным становится не то, из какого материала и каким способом будет выполнена конструкция, а то, какие принципы были заложены для реализации функции данной конструкции. Это нисколько не принижает значение материалов, способов конструирования и т.п. элементов создания конструкций, но выделяет самое главное в механизмах взаимодействия, что обеспечивает выполнение функции, т.е. обеспечивает причины и следствия взаимодействий элементов конструкции.
В природе схемы управления организмом реализуется из биоматериалов и всего того, что оказывается доступным в природном конструировании (конечно же бесцельном, как в приведенном ранее примере направлении течения потока воды).
Так, материалы оптической системы глаза разительно отличаются от материалов системы фотоаппарата, но при этом есть общие принципы, обеспечивающие функцию фокусировки света в зависимости от его источника и регистрацию картинки.
Итак, при изучении любых систем оказывается важно не то, какой материал, источники питания и т.п. ее обеспечивают, а то, какие механизмы позволяют реализовать данную функцию. И это – очень важный принцип, позволяющий отделять систему взаимодействий от того, что прямо не участвует в этом.
Одну и ту же функцию, приводящую к определенному результату, можно реализовать очень многими (бесконечно многими) способами, но принципиальная схемотехника будет одной и той же.
Это очень эффективно способно отсекать из рассмотрения все то, что не принципиально, обеспечивая понимание того, как именно реализованы цепочки причин и следствий в данном механизме. Вот почему в рассмотрении совокупности механизмов мозга и их конструкции основное внимание будет уделено именно схемотехническим принципам.
Нейросетью будем называть систему взаимодействующих идеализированных нейронов, связанных определенным образом через идеализированные синапсы. От реальных элементов мозга идеализированные отличаются наличием только принципиальных свойств и функций, не зависящих от способа реализации так, что нейросеть можно организовать любым способом: с помощью дискретных электро-элементов, программно, математически и т.п..
Понятно, что такая идеализация – критически ответственный процесс, от адекватности которого природной реальности зависят свойства моделей, построенных на ее основе.
Поэтому с особой тщательностью были проделаны работы по обобщению принципиальной модели нейрона и синапса (fornit.ru/6449), но такое утверждение никого не убедит, пока, в конечном счете, не будет с очевидностью показана достигнутая адекватность реальности. И к этому мы постепенно и уверенно придем, а пока что можно в общем виде ознакомиться с вводной статьей (с приведенным большим списком сопоставляемых фактов исследований в конце), формулирующей базовую функцию нейрона: fornit.ru/6449, но можно на эту статью по ссылке не отвлекаться потому, что далее будет очень последовательно и детализовано показана модель нейрона и синапса.
Первые же попытки программной разработки системы индивидуальной адаптивности показали, что и сама функциональность нейрона является вторичной по отношению к последовательности усложняющихся принципов адаптивности так, что в случае программирования нет смысла эмулировать работу нейронной сети, потому что нужные адаптивные функции гораздо правильнее реализуются спецификой программной схемотехники. В случае, если такая модель будет разрабатываться на основе дискретных электронных компонентов, то функциональные аналоги нейрона (нейристоры) станут наиболее оптимальным решением.
В этой книге будет рассмотрена схемотехника на основе функциональности нейронов и синапсов, что приближает к природной реализации.
Важное предупреждение. В последующем тексте главы будут рассматриваться отдельные схемотехнические принципы, которые не зависят от способов реализации схемы (механизма), но при этом будут кратко описываться проявления этих принципов в природной нейросети. Объем и популярный формат книги не позволяют обеспечивать строгую обоснованность сделанных утверждений и это способно вызывать серьезное неприятие у нейрофизиологов, привыкших к доказательности во всем, что как-то затрагивает их рабочие представления. В то же время в последующих главах будут рассматриваться уже реальные нейрофизиологические феномены и механизмы с использованием полученного контекста схемотехнического подхода. И здесь уже будут привлекаться все необходимые обоснования в виде источников опубликованных материалов по фактическим исследованиям.
Поэтому во избежание активного неприятия и потери интереса, стоит воспринимать утверждения этой главы в качестве постулатов, которые будут оправданы до очевидности в последующей общей модели.
Для нейробиологов, не имеющих схемотехнического опыта, этапы такого описания выглядят недоказательными просто потому, что они не видят системы, определяемой схемотехническими принципами. Но эти принципы достаточно жестко и однозначно укладывают данные исследований по своим местам в общем пазле модели и позволяют верифицировать как адекватность самих данных реальности, так и предполагаемые выводы.
Принципиальные элементы эволюции нервной системы
Эволюция нервной системы и, собственно, нейрона описана в большом количестве работ, но для системного вычленения принципов организации нейросети, оказывается, вовсе не обязательно проводить тщательное расследование эволюционных процессов, а достаточно сопоставить уже имеющиеся развитые природные нейросети. Тем более, что окончательных выводов из эволюционных исследований все еще сделать нельзя и попытки делать конечные утверждения такого плана выглядят спорными. И, все же, будет полезным проследить наиболее значимые моменты природной эволюции, тем более что это позволит выявить важные принципы, которые всегда используются в природной эволюции. Эти принципы вытекают из достаточно хорошо формализуемых процессов, выделенных в статье fornit.ru/806:
“Из всей совокупности сопоставленных данных исследований возникает вполне определенное обобщение: решающую роль в реализации наследственных механизмов играют два принципа. Первый – аксиоматический. Этот принцип касается не генетики и даже не биологии, а эволюции в самом широком смысле, в том числе неживых образований.
Вот пример. Возьмите каплю чернил и начните капать ее с одной и той же высоты в 1 см. на салфетку. Вы получите круги, внутри чернильного цвета, далее по радиусу - водяные (граница хроматографии чернил на салфетке). Они все одинаковые. Нужны ли какие-то поля, чтобы записать форму получающихся капель? Нет. Начнем капать на другой вид бумаги - чернильные радиусы станут другими.
Понятно, к чему все клонится? Результат той или иной формы эволюционно вовсе не был задан строго заранее. Он таков, какой выходит из общих законов термодинамики, химии, физики, - из законов причинно-следственных связей. У человека внешний вид таков, каков он получился при развитии в определенных условиях с учетом того, что оставалось не в противоречии этим условиям, иначе это отсеивалось. Форма тела у нас такая, какая получилась при мутациях и отборе наиболее жизнеспособного. Генетический материал в этом плане эволюционно отбракован среди других молекул как нечто, что способно развиться в определенных условиях опять в организм, способный существовать в данных условиях. Но если условия окажутся другими, то и результат, форма станет другой (если только организм еще окажется способным развиваться в таких условиях).
¶Второй принцип - в том, что при развитии от первой клетки, особенности каждого последующего деления определяются уже достигнутым новым состоянием, которое активизирует ту часть генома, которая эволюционно возникла для создания этого состояния организма. Т.е. переход от одноклеточных к многоклеточным потребовал дополнения генома тем, что заставляет клетку делиться вообще, далее все новшества в дальнейшем развитии организма так же постепенно формировали и дополнительные фрагменты в геноме, которые активизируются в момент соответствующего этапа развития организма – как воздействие новых условий соседства разделившихся клеток.
¶Даже не вдаваясь в подробности механизмов реализации этого принципа, понятно как он связан с эволюционными процессами. Кроме того, ясно, что вовсе не нужно запоминать план всего организма на уровне носителей наследственности, а достаточен лишь только тот пусковой механизм, который в данных условиях развития приведет к определенному результату и будет корректироваться на узловых этапах развития, активизируя соответствующие фрагменты генома, - пусковых для особенностей развития, в каждом из участков развития организма.
¶Эти два принципа аксиоматически очевидны в контексте понимания термодинамики взаимодействий любых тел:
1. Форма развития объекта нашего наблюдения (чего бы то ни было) зависит не только от внутренних свойств объекта (генетической программы развития у живых существ), а от того, какое влияние оказывает на это внешняя среда. Только в среде, на которую рассчитана программа, она дает результаты, оставляющие объект в привычной нам форме.
2. Каждый предыдущий шаг последовательности развития создает новые условия для последующего, изменяя форму объекта.”.
Эволюционное формирование нейросетевых элементов
Эволюция нервной системы – довольно обстоятельно описываемая область. В частности, в ранней работе С.В. Савельева «Происхождение мозга» (fornit.ru/5079):
“…клеточные предшественники нервной системы были однотипны. Они отличались от окружающих клеток только способностью быстрее проводить сигналы, что и выделило их в самостоятельную и быстродействующую нервную систему. Однако реакции на внешние раздражители становятся совершеннее при точном распознавании источника воздействия. Чем лучше идентифицирован раздражитель, тем адекватнее будет ответ организма.
…точка зрения братьев Гертвигов (Hertwig, 1878). Их гипотеза состоит в том, что нервные и мышечные клетки происходят из эктодермального эпителия. Они возникают независимо друг от друга и вступают во взаимодействия уже вторично (рис. II-1, а-в). В соответствии с гипотезой Гертвигов источником всех нервных клеток является первичная чувствительная клетка, возникшая из эктодермы и получившая возможность воспринимать раздражения, генерировать и проводить возбуждение. Специализированные отростки этих клеток вступают в связь с независимо возникшими мышечными клетками.
... По представлениям Клейненберга-Заварзина, из эктодермы возникает клетка одновременно с сенсорными и моторными функциями. Затем она дифференцируется на нервную и мышечную системы.
... Самым простым способом интеграции нервных сетей стало окологлоточное нервное кольцо. Его появление вполне оправдано тем, что оно находилось на границе поступления пищи в организм кишечнополостных. ... Самым простым движением, позволяющим проиллюстрировать действие диффузной нервной системы, является реакция на механическое раздражение. Пресноводная гидра (Pelmatohydra oligactis) при малейшем раздражении сжимается в микроскопический комочек. Это происходит за счёт расположенных продольно в эктодерме и поперечно в энтодерме сократимых белков. Кроме Генерализованной реакции, кишечнополостные могут дифференциально пользоваться отдельными щупальцами или их группами. Гидры способны передвигаться, чередуя при переворотах опору на подошву и ротовое отверстие.”
Начнем рассмотрение с ранних этапов эволюции, упрощая несущественное в их реализации.
Вначале были только одноклеточные организмы, возникшие на основе бесклеточных. Каждый из таких организмов пребывал в нише допустимых для его существования условий, приспособившись к этим условиям. За рамками узких условий они не могли существовать. Но разнообразие одноклеточных позволяло существовать одним видам там, где не могли существовать другие. И у всех был уже хорошо развитый механизм мутаций, определяющих пределы вариабельности, что позволяет пусть и очень малому проценту выживать в иных условиях.
Почти сразу начали возникать и способы совместного существования одноклеточных, когда разные адаптивные способности вместе оказывались настолько удачными, что расширяло границы существования по сравнению с существованием порознь. Это давало качественно большее преимущество в конкуренции и поэтому многоклеточные образования, у которых разные клетки брали на себя разные функции, становились все более универсально приспособленными.
Эволюция развития многоклеточных это – то дерево жизни, из которого возникли высшие животные и человек и поэтому нельзя обойти вниманием этот процесс.
В любых сложных организмах универсальной строительной единицей является клетка – при всем разнообразии этих клеток, настолько отличающихся по функциональному назначению, что практически решаются все проблемы адаптивности, необходимые для их существования.
Клетка оказалась настолько универсальным образованием при своей общей принципиальной схеме, что обеспечила все необходимое организму от рецепторов внешнего воздействия, до эффекторов ответного действия при том, что сам организм превратился в надежно отлаженную невероятным количеством попыток реализации систему. И процесс оптимизации все еще идет, кипит огромным разнообразием элементарных составляющих так, что даже у одного вида животных его белковые основы оказываются индивидуально неповторимы и несовместимы с другими индивидами.
И лишь один вид клеток специализировался в распознаватели для управления. Этот вид возник как промежуточный между клетками, способными выдавать электрический сигнал при внешнем воздействии (или выделять вещество, которое в последующих клетках выделяет электрический сигнал) - рецепторы и клетками, способными по электрическому сигналу совершать какие-то действия - эффекторы.
И рецепторов, и эффекторов существует немалое число разных видов (рецепторы цвета, звука, прикосновения, запаха, вкуса, температуры, эффекторы мышечных сокращений и выделения веществ разного вида - железы). Но еще больше рецепторов и эффекторов во всех отделах организма, где используются непосредственные связи рецепторов с эффекторами для различного рода регуляции.
Вначале вообще все рецепторы непосредственно взаимодействовали с эффекторами. Наследственно предопределенные связи отростков рецепторов подходили к приемным органам эффекторов. Но возникали участки, где между рецепторами и эффекторами встраивались промежуточные управляющие клетки. Местоположение таких участков оптимизировалось приносимой пользой выживания особей.
В этом процессе важнейшим эволюционным достижением было образование особенных контактов между отростком рецептора и телом эффектора, который первоначально не обеспечивал воздействие активировавшегося рецептора на эффектор, (т.е. как бы напоминал разрыв в соединении – “синаптическая щель”), но механически закреплял положение отростка на теле эффектора, чтобы он не дрейфовал где-то, а был всегда рядом.
Сейчас мы рассмотрим явление самопроизвольной активации – как схемотехнический принцип.
В механической модели это можно проиллюстрировать легкостью разбалансировки коромысла весов, если они ничем не нагружены и любой ветерок способен заставить их колебаться около равновесия.
В электрических схемах точно так же чувствительный элемент с открытыми входами (“висящими в воздухе”) будет откликаться на малейшие возмущения, продуцируя на выходе колебания потенциала.
В биосистемах описан тот же эффект для эффектора (мышцы, железы или нейрона), который уже созрел до способности проявлять активность, но в отсутствии связей оказывается легко возбуждаем любыми изменениями окружающей среды. И это – проявление все того же, всеобщего принципа легкости активации в отсутствии сдерживающего фактора.
Каждый эффектор без связи похож на коромысло весов, не нагруженное с обеих сторон, которое мотает в обе стороны малейшими воздействиями. Созревшая эффекторная клетка подвергается внешним и внутренним случайным воздействия, которые заставляют ее срабатывать хаотично. Мышца подергивается. Железа выбрасывает продуцируемое вещество в окружающее пространство.
Общее то, что эта активность у любых эффекторов обусловлена клеточным мембранным процессом, отражающимся на состоянии как самой клетки, так и ближайшего окружающего пространства. И если рядом есть отросток рецептора, который в это же время активирован внешним воздействием, то обе эти активности по обе стороны синаптической щели приводят к тому, что она становится все более проводимой так, что эта проводимость не позволяет более клетке эффектора возбуждаться хаотично, как если бы к коромыслу весов прицепили бы грузик.
Теперь только если грузик меняет свой вес, состояние коромысла может также измениться. Вот из-за такой аналогии в персептронах и стали применять понятие весов – как выражение состояния проводимости контакта, пропускающей сигнал: вес 1 будет условно означать, что контакт имеет 100% проводимость, а 0 – нулевую.
Этот эффект самопроизвольного возбуждения – общее правило любых схемотехнических систем, хоть механических, хоть электрических. Он будет в дальнейшем более подробно описан и показано его неизбежное включение в систему нейросети при ее развитии.
Заметим, что любой эффектор фактически выполняет роль распознавателя активности отростков на своем теле и в этом принципе не отличается от нейрона. А рецептор ничем не отличается от нейрона в плане имеющегося отростка для передачи в схемотехнической цепи возбуждения рецептора (если не сама клетка рецептора им обладает, как в случае фоторецептора, то обладает парная с ней клетка, на которую рецептор воздействует химическим путем, но это – не принципиально: просто будем называть такую пару рецептором). Понятно, что возникновение промежуточной клетки, обладающей обеими сущностями, не потребовала от эволюции чего-то качественно нового. Таким образом, нейрон стал одновременно и рецептором - для последующих структур и эффектором - для предыдущих. Условно назовем такую клетку элементарным распознавателем – универсальным элементом для построения любых схем управления.
И мы уже описали способ образования связи между такими клетками в момент созревания эффектора и активности рецепторов. Тут важно одно: эффектор созревает всегда позже рецепторов, которые его иннервируют так, чтобы на время своей специализации в связях уже были активными рецепторы (на самом деле все сложнее, учитывая то, что вначале рецепторы и эффекторы были почти на равных – просто разными типами клеток).
После того как процесс специализации закончится, эта клетка в норме уже оказывается не способной образовывать новые связи (исключение – в третичных зонах мозга, где сохраняется синаптическая пластичность за счет нового эволюционного механизма) и только в случае повреждений запускается механизм регенерации и новой специализации, что является очень важным условием общего последовательного развития цепи связей (другие варианты отсеивались отбором полезного). И первоначально такой период специализации оказывался достаточно коротким (хотя и оптимизируется в зависимости от порядка в последовательной цепочке связей) так, что наследственно предопределенные связи надежно образовывались в нужный момент развития организма, в оптимальной временной последовательности. И после специализации возникал запрет на порчу уже созданной схемы всякими дополнительными связями.
Т.е. весь процесс развития простейшей нейросети определялся последовательностью активизации генетических структур программы развития организма (последовательностью экспрессии генов в зависимости от окружающих условий).
А теперь – выводы из такой организации.
Рецепторы являются клетками с универсальной функцией выработки электрического сигнала и различаются принципиально только тем, как организован запуск этого сигнала.
Эффекторы – клетки со столь же универсальной функцией активации от электрического сигнала и различаются только тем, какое действие совершается при этом.
Третий вид клеток – универсальные рецепторы-эффекторы, у которых вообще нет принципиальной разницы в функциональности потому, как они обладают универсальной функцией рецепторов и универсальной функцией эффекторов – уже без какой-то своей специфики, при любом количестве вариантов реализации такой универсальности.
Этот вид клеток оказывается полезным для организации промежуточных цепей между рецепторами и эффекторами. И в такой сети промежуточных цепей управления особенно эффективно реализуются самые различные схемотехнические принципы, которые можно самим попробовать моделировать в специальном редакторе (fornit.ru/41239).
Важно обратить внимание, что уже на уровне непосредственных связей между рецепторами и эффекторами конкретизируются те критерии их функциональности, которые оказываются необходимыми и достаточными для целей регуляции. Это – уровень и длительность воздействия электрического сигнала рецептора на эффектор для того, чтобы тот выполнил ответное действие, что вполне определяет рассматриваемые границы использования такого промежуточного электрического сигнала: никакой другой “кодировки”, передачи какой-то информации не происходит и все ограничивается просто активацией одного элемента другим. Как говорят электрики – дело в эффективном значении тока воздействия, а не его скважности (длительности и периода повторения).
Этот важнейший вывод полностью относится и к промежуточным клеткам – нейронам. Имеет значение (для результирующего эффекта) только то, откуда пришел сигнал – потому, что это означает определенный признак воздействия параметров среды и то, достаточно ли сильный (продолжительный при данной скважности) это сигнал для активации последующего элемента.
Адекватность воздействию среды реализуется в том, что на эффектор воздействует соответствующий параметрам среды сигнал, что позволяет избирательно реагировать на такое воздействие. В эволюционном развитии структур нейросетей возникли иерархии выделения параметров среды в тех сочетаниях любой сложности, которые оказываются важны для организации ответных действий.
Нейрон от функции распознавания просто входной активности становится распознавателем любых сочетаний воспринимаемых параметров внешней и внутренней среды, при этом ничего не меняя в своей универсальной специализации. Как это происходит – станет ясно чуть позже.
Мы отсекли все несущественное в первичной организации нейросети. Конечно, это не означает, что не могут возникнуть какие-то модуляции сигнала или какая-то еще передача информации, если это окажется полезным. Но 1) для подтверждения необходимо найти конкретные механизмы такой передачи (а такие попытки найти это есть), 2) сам принцип релаксационной генерации электроактивности нейроном не предполагает какую-то возможность модуляции (релаксационный генератор принципиально немодулируем и может только менять частоту импульсации при неизменной длительности импульса) и 3) в ходе дальнейшего рассмотрения развития нейросети будет все более ясно, насколько вообще это может быть в чем-то полезно или оказывается лишней сущностью.
В ходе рассмотрения дальнейшей эволюции нейросети, мы будем продолжать выделять принципы реализации схемного решения механизмов, позволяющие ограничивать условия рассмотрения возможных вариантов описания механизмов (так же как рассмотрение принципа универсальности специализации элементов конструкции), отделяя несистемные составляющие конкретного способа реализации механизма. Это позволит избежать множества тупиковых предположений.
Схемотехнические принципы
Вы запросто можете не читать главы про эти принципы, если нет желания вникать в них, а склонны просто поверить в их определяющую роль в понимании любых систем причин и следствий. В таком случае можно сразу перейти к главе Первые управляющие структуры организмов на основе распознавателя Ошибка! Закладка не определена.. Но при этом потеряете немало интересного и полезного.
Есть умелые люди, которые не затрудняются что-то сделать в доме нестандартное, починить, сотворить новые удобные приспособления. Говорят, что у человека золотые руки. А он в детстве разбирал игрушки на составляющие, понимал, как все там работает и сооружал что-то свое. У него развилось схемотехническое мышление к очень многим практически полезным вещам. Сделав какие-то небольшие устройства, он способен на их основе создавать более сложные. У него в голове сформировалась система понимания взаимодействия отдельных компонентов, а потом – взаимодействия еще более сложных компонентов так, что не затрудняется взять и представить себе мысленно, как можно из имеющихся штуковин соорудить нужную вещь.
Это стало возможным только потому, что у него был достаточный интерес к этому, он этому придавал максимально большое значение, выше еды, когда его мама звала “ну пойди же покушай, наконец!”. Для него это была самая увлекательная игра. И по-другому развить схемотехническое мышление невозможно. Если подумать и вспомнить, как же удалось вам развить до высокой эффективности то, что есть теперь в активе, то окажется, что была большая необходимость и, часто - был игровой, увлеченный подход. То, что вымучивается насильно, без интереса, получается жалким заморышем. Именно поэтому родители гораздо хуже осваивают новые гаджеты, чем их увлеченные дети.
Попробуем создать упрощенный пример для того, чтобы прочувствовать эффективность схемотехнического мышления, то, что буквально все вокруг становится доступным именно при схемотехническом подходе. Этот пример, кстати, – эмулятор релаксационного механизма разряда нейрона в пейсмеккерном режиме.
Все знают, что такое электромагнит. Это катушка провода вокруг ферромагнитного сердечника и, если подать напряжение на концы катушки, по ней потечет ток, возникнет магнитный поток вдоль сердечника и к нему начнет примагничиваться все, что может.
Если приделать железный рычажок, который будет примагничиваться, когда течет ток, а на конце рычажка будет молоточек, бьющий по колокольчику, и одновременно этот рычажок, когда он примагничивается разрывает контакт тока через катушку, то получится звонок, который раньше был на всех квартирных дверях и телефонах. Получается такая причинно-следственная последовательность. Пока мы не включили ток, рычажок подпружинен наверху и его контакты подключают один из проводов катушки к электричеству. Если нажать кнопку звонка, то через контакт рычажка потечет ток в катушку, она резко притянет рычажок и тот ударит в колокольчик. Но при этом его контакт разомкнется, и катушка перестанет магнитить. Рычажок опять распрямится и подаст ток в катушку. Звонок будет звонить, пока на эту автоколебательную систему подается напряжение.
Точно по такому же релаксационному принципу нейроны создают перепад потенциала при возбуждении.
Точно так же работают маятниковые часы, только там подается не ток, а натягивается с постоянным усилием цепочка с гирькой. Вместо гирьки можно приспособить электричество, получатся электро-маятниковые часы, скорость вращения которых зависит от инерционных свойств маятника. Вместо маятника можно поставить другие колебательные элементы, как, например, рычажок звонка или кристалл кварца. Но во всех случаях автоколебаний будет работать один общий схемотехнический принцип положительной обратной связи. Тут очень много завораживающе интересного, что многими воспринимается “да ну нафиг эти сложности!”.
Нового вообще вокруг случается очень много, но для одних это не имеет значения, а для других приоритетно важно. Вот и вся разница между теми, кто станет умельцем и теми, кто просто будет простым пользователем айфона.
Раньше на электрических магнитах, которые управляли контактами (электромагнитные реле), делали вычислительные машины и любые управляющие устройства. Такой огромный агрегат помог определить код немецкой шифровальной машинки Энигмы. Фон Нейман доказал, что на такой механике может быть создано вычислительное или управляющее устройство любой сложности.
На реле можно собрать адаптирующийся мозг. Каждый нейрон будет катушкой, у которой несколько пар проводов. Если подать напряжение только на одну пару, магнитного потока не хватит, чтобы притянуть рычажок, а если запитается достаточное число пар проводов, то общая сила будет достаточной, чтобы притянуть рычажок и нейрон сработает, замкнув контакты пар проводков следующего нейрона. От того, как включены пары, зависит - возбуждающий это вход или тормозной.
Но это будет невероятно громоздкое устройство и лучше его сделать компактнее, на транзисторах и микросхемах. Природа же реализовала это совсем по-другому.
Но неважно, в виде чего реализуется устройство схемы, важен принцип, который при этом используется для взаимосвязи элементов. Он помогает понять устройство.
Итак, для развития схемотехнического мышления, первым делом, нужно придать всему этому высокую значимость в игровом контексте – понять необходимость и создать доминирующую нерешенную проблему, требующую “закрытия гештальта”. Проще говоря, нужно искренне полюбить и сделать привычным этот стиль мышления так же, как половое и пищевое поведение. Тогда откроется возможность понять не только то, как устроены схемы, реализующие психику, а все вокруг приобретет совершенно иной, глубокий смысл.
Далее мы рассмотрим несколько характерных для нейросетей схемотехнических принципов. Это пока еще не подход к серьезному рассмотрению эволюции адаптивных механизмов усложнения нейронной организации мозга, а только – полезное предваряющее отвлечение, которое позволит в новом качестве рассматривать обобщение фактов исследований по развитию мозга.
Обратная связь
Самый вульгарный пример обратной связи это – когда мышь запустили в трубу и конец трубы соединили с началом, как это случалось в детских мультиках. Вот игровая анимация этого процесса: fornit.ru/an-book-18
Связь в самом прямом смысле – обратная: сигнал с выхода подается на вход.
Сигнал – мышь. И связь положительная, т.к. мышь с выхода на вход приходит в неизменной форме.
Предположим, что мы все время запускаем новых мышей по штучке с каждым кругом. Их в трубе становится все больше.
Вот если бы что-то сделало из мыши ее противоположность, то, выскочив из выхода и став негативной, она бы вычиталась из новой запускаемой мыши и мышей бы не было больше одной: если первая аннигилировала, то запущенная вторая мышь пробежала бы круг и тут же аннигилировала с новой мышью.
Почувствуйте разницу: в случае положительной обратной связи сигнал усиливается (не мгновенно, а с некоторой скоростью запуска мышей) пока труба не окажется непроходимой (сопротивление ограничивает ток сигнала).
А при отрицательной обратной связи число мышей стабилизируется.
Вот почему положительная обратная связь вызывает непрерывную циркуляцию процесса. Если где-то что-то вызывает колебания, то можно уверенно говорить: тут есть положительная обратная связь и это стоит запомнить, как принцип. В природной нейросети есть незатухающие колебания, которые регистрируются ЭЭГ.
Люди, мыслящие по схемотехническому принципу обратной связи, придумали беличье колесо, ленту транспортера, беговую дорожку, цепную передачу и многое другое. Т.е. если этот принцип хорошо сидит в автоматизмах, то очень многое можно придумать полезного, до чего не додумается другой.
В примере с электрозвонком контакт рычажка размыкается, когда его притянет якорь электромагнита, прерывая цепь тока и рычажок отлипает, снова замыкая контакт. Рычажок – активный элемент, у которого выход – контакт, а вход – магнитный поток. Замыкающийся контакт прямо вызывает магнитный поток, т.е. связь положительная.
Обычно в схемотехнике тип связи достаточно просто идентифицируется. А вот когда мы выходим за границы обыденного, там, где нет метрики пространства и времени, становится сложно: вот что заставляет поддерживать колебания квантов (суперструн)? Это еще не выяснили. В обычной механической струне, по которой идет смычок, есть положительная обратная связь, как и в маятнике. Мало того, она есть и в струне, которую только раз щипнули, и она зазвучала, пока не затихла. Если есть колебания – значит, есть положительная обратная связь. Это – фундаментальный принцип, который может вызывать сомнения в конкретных явлениях.
К примеру, физиологи выяснили, что в сетчатке глаза между ФР и горизонтальными клетками (ГоК) имеется отрицательная обратная связь: в темноте ФР активирует с помощью глутамата ГоК, а ГоК в ответ тормозит ФР с помощью ГАМК. При включении света этот механизм на какое-то время вызывает колебания потенциала ФР. Где же наличие положительной обратной связи? Как раз про затухающие колебания говорилось ранее, в теме про обратную связь: “Если где-то что-то вызывает колебания, то можно уверенно говорить: тут есть положительная обратная связь и это стоит запомнить как принцип”. Разберем этот эпизод подробнее в контексте схемотехнических принципов.
В данном случае включение света порождает крутой фронт активации. Любая динамика процесса раскладывается на составляющие гармоники, суммы которых и формируют фронт. Резкий перепад света формирует самую высокую гармонику, отражающую скорость нарастания света, к ней добавляются гармоники, формирующие все большую продолжительность периода светового воздействия.
При наличии обратной связи резкий фронт вызывает появление затухающих колебаний потому, что обратная связь срабатывает с задержкой и не успевает скомпенсировать резкие изменения. Если бы задержка была такая, что действие обратной связи оказывалось бы синхронным с изменением сигнала на входе, то возникла бы постоянная генерация. Но реакция всего лишь немного запаздывает, оказываясь положительной только для высоких гармоник процесса. Это значит, что всегда можно найти такую плавность нарастания освещения, при которой не будут возникать затухающие колебания. По таким колебаниям можно вычислить время запаздывания обратной связи.
В точности по такому же принципу возникает и “микрофонный эффект”, когда вдруг динамики начинают свистеть, когда обратная связь от их излучения оказывается положительной на какой-то частоте случайного сигнала, спровоцировавшего свист.
В каждом конкретном случае генерации можно вот так найти механизмы причин и следствий, которые приводят к явлению генерации или ее затуханию, или сразу вызывают появление стабильного, сбалансированного обратной связью сигнала.
Рассмотренный эпизод является примером того, как схемотехнический подход укладывает экспериментальные данные в систему взаимосвязанных принципов, позволяя понять суть происходящего явления.
А теперь - многозначительный эксперимент, который можно проделать мысленно потому, как для такого уровня представлений причин и следствий наверняка уже достаточно опыта.
Возьмите обычную шумовку (длинную ложку с широкой пластиной, в которой наделано много дырок). Проденьте в ручку карандаш (можно просто привязать веревку, но карандаш ограничивает возможность качания в разных плоскостях).
Пустите из крана довольно сильную струю воды и приблизьте пластину к струе. Ясно, что ее оттолкнет, шумовка провернется маятником на ручке и снова под своей тяжестью попадет под струю. Начались автоколебания.
Но можно подобрать такую тонкую струйку, которая хоть и слегка отклонит пластинку, но не вызовет автоколебания: положительная обратная связь оказалась недостаточной для накопления нужного для автоколебания количества воды. Ток воды, входящей на пластину, не позволяет накопиться там достаточному количеству кинетической энергии чтобы, отклонив пластину, прервать ток воды на нее. Получается условие генерации, связанное со скоростью подачи тока, так, чтобы обратная связь стала накапливать этот ток.
Возьмем коромысло от весов, на одном конце которого будет гирька (200 грамм для наглядности), а к другому концу прицепим 5 легких стаканчиков на одной оси: ряд стаканчиков – поперек коромысла (под прямым углом). Начнем наполнять стаканчики сразу пятью струйками воды. В какой-то момент равновесие изменится, и стаканчики пойдут вниз. Эти стаканчики были продернуты чуть выше середины так, что легко могут вращаться, если их наклонит и у них есть наклоняющие выступы. Опустившись до края ограничительной пластины, выступы стаканчиков заставят их перевернуться и вылить воду. Если бы стаканчики опускались медленно, то они бы просто затормозились за выступ, но до этого места они уже успели набрать много воды и их скорость (инерция) придала им достаточную кинетическую энергию, чтобы опрокинуть и вылить воду. После чего пустые стаканчики взлетят вверх. Начнется автоколебательный процесс.
Зачем нужны были несколько стаканчиков, а не один? Это был водяной аналог срабатывания нейрона по пяти входам. Пока равновесия не было нарушено достаточным весом воды, нейрон перевешивался гирькой, которая определяла его порог срабатывания.
Следует осмыслить этот принцип порога срабатывания (компарирования), само устройство – компаратор (сравнивающий величину сигнала с порогом). Можно довольно легко найти компараторы в окружающем.
Принцип выделения составляющих элементов воздействующих факторов.
Для адаптивности, адекватной реальности, самое главное – суметь воспринять важные признаки в окружающем, чтобы не махать тупо кулаками вслепую.
Например, выделить положение уязвимого места врага.
Можно это сделать просто по сигналу датчика освещенности, который реагирует на отраженный от врага свет: когда враг близко, станет чуть темнее. Но это потребует вращения головы во все стороны, чтобы определить в каком месте стало темнее. Некоторые виды животных довольствуются такой простотой.
Можно вокруг головы иметь много отдельных датчиков освещенности, чтобы не крутить головой, и они сразу покажут, где динамично меняется освещенность. Но точно прикинуть расстояние или вообще отличить врага от друга так не получится. У насекомых датчики освещенности собраны в большие фасеты с трубками, направленными в разные стороны. Кроме этого, есть и отдельные датчики освещенности. При большом числе фасеточных датчиков получается лучшее ориентирование.
А вот если до сетчатки светодатчиков поставить общую линзу, которая сделает изображение четким, а не размытым, то эффективность возможности распознавания качественно увеличится.
Со зрением такое возможно потому, что длина волны видимого света очень маленькая и позволяет выделить мелкие детали изображения. А вот длина волны звука – большая и невозможно иметь звуковую линзу малого размера, да и детали будут распознаваться только очень крупные (или нужно перейти в ультразвуковой диапазон). Чтобы улучшить ситуацию, звук делится на множество длин волн с помощью резонансных волосков улитки уха, это дает детализацию по высоте звука, что тоже очень важно. А вот детализация по длине волны у глаза человека имеет всего 3 значения.
Вот главная зависимость: чем на большее число составляющих разбить параметр восприятия, тем точнее восприятие и больше его возможности выделить значимые параметры среды. И тогда можно распознавать множество разных сочетаний этих параметров, просто реагируя на каждое данное сочетание.
Каждый такой распознаватель имеет входы от отдельных уже разбитых на детали элементов параметра внешнего воздействия и эти входы так подобраны по реагированию на детали, что общее срабатывание распознавателя будет только когда возникнет определенная картинка в сочетании входных сигналов.
Такой распознающий элемент назвали персептроном (fornit.ru/38065), и нет никакого другого принципиального способа организовать распознавание. Т.е. распознаватель всегда должен работать с уже разбитыми на детали элементами воздействующего параметра внешней среды, от одного до множества. Персептрон со входами от разбитых на детали параметров – важнейший схемотехнический принцип организации распознавания.
Какой бы ни была реализация адаптивной системы, она должна обладать возможностью выделять детали входных параметров и распознавать их сочетание. Исключений не может быть никаких ни в чем и это – важнейшая аксиома: каким бы ни было схемное решение для распознавания чего бы то ни было, это устройство будет иметь компаратор, срабатывающий при превышении порога веса схожести с эталоном. И этот принцип позволяет понять, как организуется природная нейросеть.
Забегая вперед, заметим решающие отличия однослойного, предельно упрощенного персептрона (со связями ограниченного количества рецепторов с распознавателем) в реализации природной нейросети и сложного многослойного персептрона (со связями все со всеми) в реализации современных искусственных сетей.
Если представить цепочку рабочих, перебрасывающих кирпичи, то первому из них была отведена роль распознавателя правильного кирпича, потому как если попадется не такой, как нужно, кирпич, то будет и неправильное его использование, скажем, вместо кладки огнеупорной печки, получится брак, который развалится очень быстро.
А чтобы правильно распознать кирпич, рабочий должен иметь возможность разбить свет, отраженный от него, на мельчайшие части уже сфокусированной картинки, позволяющей по ее деталям отличить огнеупорный кирпич от пористого по характерным распознаваемым сочетаниям деталей.
Проводимость и сопротивление
Проводимость – ключевое свойство в нейросетях, обеспечивающее распознавание.
Проводимость – способность проводить ток. Ток, как уже говорилось – в самом широком смысле: количество кирпичей по цепочке рабочих, количество воды в водопроводе, количество электронов в электропроводе или ионов в жидкости – перемещающихся в единицу времени.
Проводимость – понятие, обратное сопротивлению, т.е. сопротивление – способность проводника ограничивать ток неким количеством носителей в единицу времени.
Тут главное – именно то, в каком отношении находятся составляющие процесса:
ток (будем обозначать i), сопротивление (r) и то усилие, которое заставляет течь ток (напряжение u), проводимость (g).
Не будем называть это математикой, но уметь в уме видеть, в каком соотношении находятся параметры взаимодействующих факторов в процессе – просто необходимо, это – важная часть схемотехнического мышления, позволяющая предвидеть результат причинно-следственной связи взаимодействующих компонентов.
Сейчас у нас 3 главных параметра: сопротивление (или обратная ему величина - проводимость), ток и напряжение. Представим их взаимодействие на примере вытекающей из бака воды.
Чем выше бак, тем сильнее из него вниз по трубе течет вода. Высота бака обеспечивает ток воды, и этот параметр назовем напряжением или разностью потенциалов кинетической энергии на высоте и внизу. В любом промежуточном месте трубы потенциал будет меньше, чем на самом верху, и разность потенциалов – тоже меньше, но так как выше есть давление воды из более высокого потенциала, то она потечет быстрее, чем если бы она выливалась из меньшей высоты.
Таким образом, ток воды ограничивается не только просветом трубы, но и тем, что верхней воде мешает нижняя из-за своей вязкости и инертности. В случае электронов этого фактора нет потому, что электроны не обладают вязкостью и инертностью, ограничиваясь только возможностью материала проводника пропускать электроны. В случае тока дробинок и, еще больше, песка, взаимное трение мешают сильнее, и даже крупные песчинки могут вообще закупорить отверстие. Но в песочных часах отверстие подбирается заведомо крупнее песчинок и сами песчинки обкатаны до шарообразности.
Как бы высоко ни был бак, из-за ограниченной проводимости сливной трубы, вода вытечет не сразу. И наоборот, как бы низок ни был бак, ток воды все равно будет, если только высота не станет равна нулевой и исчезнет разность потенциалов или напряжения.
Из отношения зависимостей можно сказать: ток будет прямо пропорционален напряжению и обратно пропорционален сопротивлению проводника. Это и есть закон Ома. Он справедлив для любого вида тока и любого вида напряжений и сопротивлений.
Он должен стать уже очевидным в своей верности и остается только применить его к самым разным бытовым и не очень явлениям, чтобы это стало автоматизмом понимания, т.е. вошло в систему схемотехнического мышления.
Для нейросетей более важно понятие проводимости потому, что именно оно позволяет с очевидностью представлять принцип действия персептрона, о котором уже говорилось и который, как утверждалось, - единственно возможный способ организовывать распознавание признаков восприятия. Если все это не было воспринято как определяющее важное, то стоит вернуться и убедиться в справедливости утверждения, а также попробовать во многих разных случаях, пусть в мысленных экспериментах, найти его реализации.
Возвращаясь к закону Ома, можно представить его в виде формулы для расчетов:
i=u/r или в случае проводимости i=g*u; Пожалуйста, сделайте несложные математические преобразования, чтобы получить эти зависимости от эвристического представления о пропорциональности параметров r, g, u, i.
Еще про распознаватели, кое-что по-настоящему важное
Представим в качестве распознавателя замок, распознающий подходящий для него ключ.
Сможет ли он провернуться через прорезь, сделанную для его язычка или нет.
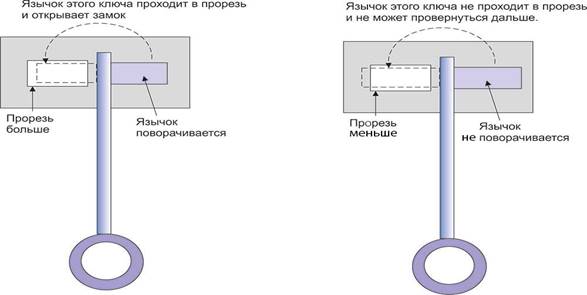
Сейчас важно определить, сколько же признаков необходимо для распознавания и какими должны быть эти признаки.
У классического ключа есть язычок: если его длина и расположение при воткнутом ключе оказывается точно напротив прорези, то вращение ключа становится возможным и язычок, пройдя прорезь, отключит замок, т.е. произойдет распознавание с компарированием схожести язычка и прорези и выдачей сигнала в виде свободы вращения ключа.
Для любого причинно-следственного процесса нужна энергия, позволяющая осуществлять движение. Такую энергию дает усилие вращения от руки. Любые другие схемотехнические системы не имеют в этом исключений.
Итак, язычок может быть один или их может быть несколько, длины язычка тоже могут быть разными. Компарирование будет успешным, когда вся совокупность этих параметров окажется в соответствии с профилем выреза замка.
И здесь есть один важный момент. Если язычок будет длиннее прорези для нее, то ключ не провернется. А вот если меньше - то легко провернется, но хватит ли его для того, чтобы зацепить открывающий механизм? Обратите внимание, что величина в сторону увеличения оказывается критичной, а длина в сторону уменьшения от длины прорези – нет. Другими словами, позитивная величина параметра распознавания оказывается существенной, а негативная – влияет уже после валидации длин язычков прорезями – на втором этапе компарирования.
Этот принцип оказывается определяюще важным в нейронных природных распознавателях потому, как позволит сузить границы предположений для уточнения модели распознавателей в природной нейросети.
Получается, что замок можно обмануть, укорачивая язычки, но не до степени, когда они перестанут открывать механизм. И на первом этапе работают только положительные параметры (в терминах нейрона назовем их возбуждающими) и только потом срабатывает сравнение длины язычка с длиной рычажка открытия замка.
Попробуйте придумать, как организовать сразу распознавание с равноценным использованием как возбуждающего, так и тормозного параметра в случае классического замка (т.е. чтобы укорачивание язычка так же надежно не давало провернуть ключ, как лишняя длина). Попробуйте настойчиво до успешного решения, чтобы полностью прочувствовать проблему. Вы убедитесь, что это – очень непросто. А вот в два этапа распознать подходящий ключ несравненно проще в любом плане, как в плане сложности конструкции, так и в плане расходуемых материалов и в плане надежности распознавания. Это – принцип, определяющий направление совершенствования нейросети.
Может ли распознаватель оперировать только с одним параметром? Казалось бы, это – абсурд, раз параметр один, то чего его распознавать? Но распознаватель – не только валидатор соответствия параметра профилю распознавания, но его функцией, без которой он теряет смысл, является активация – сигнал распознавания, который может как-то использоваться.
Возьмем сито с круглыми дырками определенного диаметра. Параметры среды – песчинки разного размера, цель сита – отсеять все, что мельче диаметров его дырок. Параметр распознавания один, а польза очень существенна. И, заметим опять, что сито работает только с положительными параметрами и не может отсеять более мелкие, чем дырки, песчинки. Для этого понадобится вторичные процессы с более мелкими дырками. Или какой-то другой распознаватель, но работающий только с теми песчинками, которые меньше диаметра его профиля распознавания.
Делаем вывод: распознаватель имеет смысл, начиная уже с одного единственного параметра распознавания. И даже в случае, если у него вообще нет ограничений на функцию распознавания, и он срабатывает от всего, что может заставить сработать его компаратор, превысив какой-то порог, в чем-то можно извлечь пользу.
Это означает, что распознаватель является наиболее универсальным устройством для создания любых схемотехнических последовательностей. Достаточно будет модифицировать только его профиль распознавания.
Идеальный математический распознаватель – персептрон. Он настолько идеален, что обладает возможностью распознавать с использованием как положительных (возбуждающих его), так и отрицательных параметров.
Системы на основе идеального персептрона реализуют так называемые искусственные нейросети, которые, по признанию самих разработчиков, не имеют ничего общего с природными нейросетями.
Причина этого не только в том, что для реализации учета отрицательных параметров нужны сложные и затратные ухищрения, а процесс эволюции бездумен и, как вода в арыке, идет по наиболее покатому руслу. Сейчас мы рассмотрим одну из этих причин.
Вопрос: возможно ли одним ситом отделить конкретную фракцию, только разделив кучу на две части: с большими и меньшими относительно ячейки сита гранулами.
Нет, конечно. Одно сито имеет граничные условия распознавания от диаметра своей дырки до нуля. Обратите внимание: каждое сито – это распознаватель такого широкого диапазона.
Говоря же о колонке простейших распознавателей-сит с дырками 1,2,3,4,5,6,7,8,9 см, вроде бы понятно, что кроме собственно распознавания каждым ситом от диаметра его дырки до нуля, стало возможным формировать последующие распознаватели любых сочетаний, скажем только от 2 до 5.
Альтернатива: создание сложных (непонятно как устроенных) распознавателей для каждого такого сочетания, т.е. вместо девяти - девять в квадрате.
Это как пытаться делать сразу распознаватели лица каждого конкретного человека вместо того, чтобы воспользоваться более простыми графическими примитивами для их сочетаний только в случае нужного распознавания (запоминания данного человека).
Пока еще мы никак не затрагивали схемотехнику в программировании. А базовые представления о программировании потребуются для понимания многих моментов в книге. Те, у кого с этим плохо, предлагаю не пожалеть пару дней на блиц-курс по основам: fornit.ru/1156
Программные продукты не реализуют природные причины и следствия, а лишь моделируют, эмулируют их, основываясь на логике алгоритмов.
Так, оператор под названием IF является программным распознавателем соответствия условия (параметра) и действующего фактора. Если компоненты совпадают, то происходит действие (срабатывание распознавателя), если нет, то - игнорирование. В этом операторе можно сравнивать как отдельные параметры, так и любые их совокупности.
Так строился ИИ старых (и многих новых) компьютерных игр. Например, если персонаж игрушки говорил игроку: “Ну, привет, бро!”, то выдавались ограниченное число возможных вариантов ответа на выбор, например, 1) О, привет, дружище! 2) Да пошел ты… В случае совпадения выбранного варианта 1 в операторе IF (IF answer == 1) возникало продолжение дружеской беседы, а при IFanswer == 2 начиналась мочиловка.
Создатели игрушек усложняли условия, пока у них хватало мозгов все отследить (а сложность кода нарастала лавинообразно), например, добавлялось время действия и время суток. Если все происходило в уютной комнате вечером с героиней игрушки, то ясен пень какое было продолжение, а если встреча была на пустоши среди коварных врагов, то мочиловка могла начаться по любому поводу.
Понятно, что все ограничивалось только уже предусмотренными вариантами и не более той сложности, после которой игрушка больше глючит, чем работает (контролировать все переходы по оператору IF становится лавинообразно сложно по мере их используемого количества в совместном акте распознавания).
Но главное вот что.
Оператор IF сравнивает объекты, а не их составные части. Так, даже если задано условие IF parameter == 3.14, то число 3.14 – это не какой-то дробный огрызок, а объект с таким вот свойством. Никакие другие числа не пройдут это условие. Т.е. входным параметром реально действующего распознавателя должен быть и реальный в своих свойствах признак, обладающий (или не обладающий) свойством, на которое настроен распознаватель. Именно реальные признаки окружающей действительности важны для реагирования.
Пока не будем обращать внимание на то, что тем самым признак выделяется из реальности в том виде, в каком он в реальности не существует, образуя некий аналог абстракции, но заметим, как рано начинают формироваться отвлечения от реальности при адаптивном распознавании.
Если уже достаточно ясен принцип идеального персептрона, то очевидно, что на входы его суммирующего компаратора подаются строго дозированные части входных параметров за счет ограничения их проводимости, что в персептронах называется весами (На коромысло весов со стаканчиками наливается вода и вес каждого стаканчика влияет на компарирование). Получается, что идеальный персептрон работает не с реальными входными параметрами, а с заданными программистом какими-то их частями, которые подбираются программистом так, чтобы персептрон срабатывал только на данный набор входных параметров. Это та самая сложная заморочка, которая была при попытке заставить замок реагировать на уменьшение длины язычка так же, как на его увеличение.
Вот почему искусственные нейросети такие непонятно сложные даже для самих их создателей: нет возможности осмысленно отследить, что же происходит там внутри. И нет возможности найти в самом деле независимый от программиста способ обучения. А искусственное возбуждение нейрона в реальной нейросети примитивов восприятия даст появление определенного образа.
Вывод: реальный универсальный распознаватель должен работать с реальными же входными параметрами, а не с их весовыми частями. И не пытаться сразу в одном флаконе распознать как позитивную, так и негативную составляющую входных параметров, а проще и надежнее это делать поочередно.
Но, понятно, что организовывать профиль распознавания, настроенный на реальные параметры (на свет, на звук и т.п.) – невозможно. Этим занимаются только самые первичные распознавали физических воздействий в сетчатке глаза или в улитке уха. А оттуда идут уже цельные (а не взвешенные) сигналы, но вторичные по очереди распознавания, что проявляется в примерно одинаковой амплитуде выходных импульсов.
Это - еще довод против того, что сигналы нейроном как-то модулируются для полезного использования.
Универсальный распознаватель реального действия похож на ту водяную модель с коромыслом весов, на одной стороне висит грузик, определяющий порог срабатывания, а на другой до 10000 нановедерок для заливки в них воды по трубочкам примерно одной пропускной способности (конкретная проводимость уже не важна, важно только идет по ним вода или нет), идущих от первичных распознавателей физического воздействия.
Так, если появился звук определенной частоты, то откроется клапан соответствующей трубочки и в ней вода начинает литься в ведерки тех распознавателей, которые должны определять, например, что вместе с этим звуком еще есть и красный свет в какой-то фасетке датчика света. И распознаватель такого сочетания реальных признаков быстро наполнит свои ведерки, они перевесят, и весы откроют клапан чтобы вода по трубочке от этого распознавателя полилась дальше в очередь последующих распознавателей. Будет воспринят образ: красный свет в сопровождении определенного звука, как это бывает при взрыве, а синий цвет с таким же звуком сопровождает молнию.
Уже, наверное, заметно, как постепенно образуется целостная система важнейших схемотехнических принципов, без которой невозможно разумно пытаться отгадать механизмы природной нейросети.
Вы, должно быть, уже прочувствовали, что каждый из этих принципов важен и универсален вне зависимости от способа реализации последовательностей процессов. И, возможно, многие из рассмотренных принципов предстали в довольно неожиданном ракурсе и свойствах, но потом все стало взаимосвязано и понятно.
Но эти все важные принципы (и то многое, о чем пока еще не говорили) до сих пор практически никак не учитываются физиологами, пытающими понять работу нейросети, и в этом причина всех их неудач даже в определении функции нейрона.
К примеру, в (довольно старой уже) работе fornit.ru/816 (с не менее старыми комментариями в тексте) Ю.И. Александрова “Научение и память: системная перспектива” делается попытка формализовать функциональность нейрона с такими утверждениями:
· нейрон, как и организм, реагирует на стимулы
· Последовательность событий в деятельности нейрона становится аналогичной той, которая характеризует активный целенаправленный организм, а его импульсация - аналогичной действию индивида.
· активность нейрона, как и поведение организма, рассматривается не как реакция, а как средство изменения соотношения со средой,
· на уровне отдельных нейронов, достижение результата выступает как удовлетворение метаболических «потребностей» нейронов и прекращает их импульсную активность.
· Заметим, что в рамках представлений, отрицающих подход к нейрону как к проводнику возбуждения, сам термин «медиатор» или «нейротрансмиттер» (используемый в значении «биологически активное вещество, являющееся посредником в процессе передачи возбуждения через осуществление синаптического влияния») представляется неадекватным.
· В настоящее время становится общепризнанным, что многие закономерности модификации функциональных и морфологических свойств нейронов, а также регуляции экспрессии генов, лежащие в основе научения у взрослых, сходны с теми, которые определяют процессы созревания, характеризующие ранние этапы онтогенеза]. Это дает авторам основание рассматривать научение как реактивацию процессов созревания, имеющих место в раннем онтогенезе. Формирование нового поведенческого акта в любом возрасте есть формирование новой системы - системогенез.
· Селекция нейронов зависит от их индивидуальных свойств
· Из сказанного следует, что описать механизмы поведения с системных позиций – значит выяснить, каковы «паттерны нейронных специализаций» разных структур мозга, т.е. относительно каких конкретно систем специализированы нейроны этих структуры, как формируются системные специализации.
и т.п.
Попробуем дать чуть более образное пояснение принципа целостного параметра воспроизведения, а не использование дробных и отрицательных (тормозных) сигналов (как используются тормозные воздействия будет показано позже и эти воздействия чрезвычайно важны).
Какой бы ни была чувствительность сенсорной клетки, по ее сигналу становится возможно ориентироваться об окружающем, породившем этот сигнал.
Например, представим, что слепой видит только одной точкой сетчатки, переводя ее с одного на другое. Он переходит дорогу, наводит глаз на светофор, точка показывает красный цвет. Неважно, какая чувствительность точки, главное, что она достаточная, чтобы передать информацию: на светофоре красное. Для этого не нужны никакие веса, нужен просто распознаватель цвета этой точки. От сенсора идет три проводка: красный, зеленый и синий. Все: воспринимаемый параметр – цельное свойство, выделенное из реальности, а не что-то дробное или отрицательное.
Но интенсивность – тоже важная характеристика, точнее не абсолютная яркость, а относительная, и эта относительность так же распознается.
На этом нужно будет остановиться до полной ясности, а будет еще много слов… Хотя принцип ненужности точного учета веса отдельных параметров на входе персептрона был уже как-то обоснован, но, наверняка, недостаточно убедительно, так что буду пытаться находить очевидные доводы.
В многослойном персептроне нет выделенных примитивов первого и последующих уровней. Там работает вся сеть так, что погрешности первого уровня компенсируются последующими в общую картину на выходе. Многослойный персептрон и несколько параллельных последовательных однослойных с несколькими последовательными такими слоями, где каждый нейрон работает самостоятельно (если не считать тормозного взаимовлияния в каждом слое) – совершенно разные схемы, с разными свойствами.
Для целей нахождения модели универсального распознавателя мы рассмотрим только однослойный персептрон (fornit.ru/945) - аналог одного нейрона.
Этот универсальный распознаватель имеет много особенностей в развитии в зависимости от того, в какой области мозга (по эволюционной последовательности) он находится. Наиболее разительная разница – развитие распознавателя в древних структурах, определяемых практически полностью наследственными особенностями и развитие в составе новой коры, где специализация зависит от сенсорной активности внешней среды. В первом случае возможны самые прихотливые сочетания, во втором развитие происходит строго в составе текущего созревающего слоя нейронов и там очень важно их взаимное тормозное влияние.
В любом случае нейроны-персептроны характеризуются суммацией всех входных сигналов и компарированием превышения этой суммы определенного порога (в виде релаксационного процесса, порождающего скачок потенциала).
В случае новой коры каждый нейрон первичного слоя не связан сразу со всеми рецепторами данной модальности (как это всегда есть в искусственных нейросетях, где используются многослойные системы типа все связи со всеми), а связан лишь с некоторым их количеством прямо перед собой (в зоне неширокого ветвления дендритов) и соседний нейрон во многом перекрывает это поле, заходя чуть в сторону. Тормозное влияние соседей приводит к тому, что в процессе специализации возникает эффект контрастирования – четкого выделения активных нейронов под воздействием более размытого характера стимула. Это – основное условие формирования примитивов восприятия, без какого-либо корректирующего ошибки процесса. Контрастирующий эффект можно посмотреть с помощью эмулятора: fornit.ru/38103.
На этом этапе нет никакой значимости распознаваемых образов, они просто создаются, отражая характерные условия окружения, что и нужно для того, чтобы можно было использовать эти признаки восприятия для адаптивности.
Теперь наша задача – определить принципиальную модель наиболее универсальной единицы схемотехники нейросетей, необходимую и достаточную, чтобы с ее помощью возможно было создавать любые схемы управления чем-то в зависимости от воспринятого. Это – универсальный распознаватель.
Распознаватели располагаются между сенсорными датчиками, преобразующими самые разные виды воздействия в совершенно безликие сигналы да-нет или 1-0 (в природной нейросети – есть активность и нет активности).
1 – данный вид воздействия (в механической модели - течет вода по трубке), 0 – нет такого воздействия (не течет вода по трубке). Т.е. какими бы ни были виды воздействия: звук, свет, прикосновение, запах и т.п., а выходной сигнал для всех одного вида: есть или нет такое воздействие. И этот сигнал не дробный (не весовой). В условной трубке проводника сигнала вода может течь или капать, но ее наличие говорит о том, что есть такое вот определенное воздействие.
А как же градуальные сигналы, например, ФР и биполяров сетчатки? Градуальные изменения ВПСП, ТПСП и частоты ПД? Чем выше частота испульсации, тем быстрее активируется последующий элемент. Может ли он таким образом адекватно определить силу воздействия на рецептор?
Реально нет возможности оценить абсолютную интенсивность сигнала, она регистрируется относительно соседних, которые оказывают тормозное влияние на все окружающие сигналы. Поэтому частота импульсации не отражает абсолютную интенсивность, а оказывается в составе общего образа, состоящего из всех поступающих сигналов. Этот момент – достаточно сложен в понимании картины принципа распознавания профиля активности, и мы его будет уяснять постепенно. Пока что стоит просто принять выделенное утверждение к сведению.
Этот принцип обеспечивает передачу всего диапазона градаций силы воздействия, от порога чувствительности до максимально возможного. В любом случае каждый сенсор выдает только да или нет. Градация же обеспечивается тормозным воздействием соседних элементов так, что воспринимается только самый сильный сигнал, а другие подавляются. И только когда нет сигналов, то сенсор приобретает свою пороговую чувствительность.
В этом принципе – качественная разница между распознавателями нейросети и искусственной нейросетью: в первой есть только два уровня (да или нет), во второй используется плавное изменение градаций силы сигналов (весовые коэффициенты по каждому входу) и во второй нет взаимного торможения соседними элементами (к тому же там используются связи “все со всеми”).
Искусственные нейросети реализуют идеальный распознаватель, которому требуется обучение, а универсальный распознаватель природной нейросети – очень прост, без обучения на ошибках, но при этом способен организовать сколь угодно гибкую и эффективную систему адаптивного управления.
Понятно, что чем грубее датчик, тем он менее информативен. Мы уже говорили о том, почему светочувствительность развивалась через множество отдельных светочувствительных датчиков на голове, потом через фасеточные трубки насекомых, торчащие во все стороны, позволяющие точнее определять направление и вид светового воздействия, а потом – зрительную матрицу-сетчатку глаза. Во всех случаях каждый сенсор (часто через систему промежуточной обработки) выдает на выходе только 1 или 0. У нас была механическая модель: когда есть воздействие, оно открывает краник и по трубочке начинает течь вода, наполняя ведерки коромысла-распознавателя.
Еще мы говорили, что лучше делать очень простые распознаватели, чем сразу сложные потому, что из набора простых можно следующим шагом получить любые сложные сочетания. А вот сразу получить распознаватель сложного сочетания очень трудно, да и неневозможно заготовить все возможные сочетания так же, как заготовить все фразы языка вместо того, чтобы пользоваться буквами, и на их основе - словами и только потом – фразами.
Вырисовывается картина, что универсальный распознаватель, во-первых, использует и универсальные сигналы на входе 1 или 0, в зависимости от того, что означает вода в трубочке данного входа (от какого сенсора идет трубочка) и, во-вторых, элементарные распознаватели, в случае новой коры, образуют колонки плавно меняющихся примитивов своего распознавания всегда очень простых сочетаний прямых воздействий (без дробных частей от каждой трубочки и без обратного значения параметра по каждой трубочке – аналогия с ситами).
Сам по себе любой распознаватель ничего не значит, значит только то, на какое внешнее воздействие откликается он, как он распознает соответствие с внешним параметром воспринимаемого (абстрагируясь от реального воздействия в виде внутренних коррелятов). Тварь выжила потому, что трубочки данного воздействия привели к срабатыванию данного нановедерочного распознавателя и удачно включила подходящее действие, спасшее тварь, и теперь все ее потомки передали такой механизм по наследству. Это – самое примитивное управление, предопределенное в генах.
Понятно, что большинство не настолько удачливых тварей подохло в данной ситуации, а только несколько со случайно подходящими процессами управления выжили, но теперь возникла популяция, способная справляться с напастью, убившей всех остальных.
Как видите, никакой проблемы обучения управляющих распознавателей здесь не требуется. Но, создав элементарный распознаватель, природа стала применять его во все более изощренных схемах, так же как она поступила, создав клетку – универсальный строительный материал тела.
После освоения самого принципа организации элементарного распознавателя, мы будем рассматривать способы его специализации уже не наследственным отбором, а в период индивидуального развития особи – с помощью новой коры мозга. Это – уже очень продвинутый этап эволюции и очень не у многих животных есть колончатая кора примитивов распознавания, которая формируется у животных по сложности начиная с птиц и рыб (fornit.ru/40141). Системное обобщение данных по колончатой организации собрано в статье fornit.ru/43642.
Насекомые от простейших как бабочка со всего лишь считанным числом цепочек реагирования в зависимости от обстоятельств, выбивались в более продвинутые с каждым новым совершенно случайным мутантным приобретением еще какой-то цепочки. И у пчел-муравьев этих цепочек накручивалось очень, очень много, для многих случаев. Автомат любой сложности можно собрать из таких цепочек, проверяя жизнями огромного числа особей за огромное количество времени. Поэтому пчелы и муравьи демонстрируют очень сложное поведение.
Так происходило пока не возникла еще одна мутация, породившая зачатки “новой коры” с возможностью формировать дополнительные “внутренние” псевдо-органы чувств, т.е. кроме глаз, ушей и т.п еще и примитивы сочетаний разных образов восприятия, которые оказываются сразу не связанными с цепочками реакций, так же как сразу не связаны с ними рецепторы глаз или ушей. Они составляли наборы признаков, отражающих внешнее, на которые модно было полагаться уже для реагирования с зависимости от их сочетаний.
Вот это и был качественный скачек от насекомых к более сложным тварям. У насекомых нет развитых коллекций примитивов восприятия, на основе которых можно развивать условные рефлексы, или есть в самом зачаточном состоянии. Конечно, четкую грань между принципами нейрорегуляции сложными насекомыми и более сложными животными провести невозможно, так что эта классификация очень условна.
Рассмотренные принципы организации универсального распознавателя и сети управления на его основе настолько важны, что стоит остановиться на отдельных моментах.
Про принцип простоты (сита, ключа и т.п.).
Мы не знаем заранее, какие диапазоны камней нам потребуется отсеивать и поэтому в хозтоварах покупаем 9 сит с дырками 1,2, ... 9 см. Теперь мы можем выделить любую фракцию, например, если нам нужны камни от 2 до 5 см. или только мельче 2 см. или только крупняки больше 7 см. Никто не будет спрашивать продавца, а есть ли у вас сито, отсеивающее строго от 5 до 6 см. А если спросят, продавец покрутит пальцем у виска (мысленно) и предложит взять два сита 4 и 7 см и сначала просеять вторым, потом первым. А наивному ребенку объяснит, что придумывать одно сито на 5 и 6 никто не станет потому, что 1) это непростая и дорогая конструкция, 2) гораздо проще и удобнее сделать два простых сита, чем одно такое вот навороченное.
Именно сопоставление возможности использовать набор сит с разными дырками или альтернативу сложной конструкции позволяет увидеть, что в первом случае мы получаем неограниченную гибкость любых сочетаний разных сит, а во втором решение не только требует повышенной технологии и затрат, но и узко ограничено данным диапазоном отсеивания, так что требуется много таких специализированных устройств там, где все решалось легко и просто.
Про принцип простого и универсального распознавателя.
Конструкторы искусственных нейросетей понадеялись на максимально математичную и мощную модель идеального многослойного персептрона, т.е. элемента, который максимально широко способен выполнять свою функцию, но при условии многих привнесенных факторов: способности регулировать проводимость входных сигналов в зависимости от ошибки, а, значит, способности определять ошибку. Его можно настроить на распознавание лиц, но такая сеть будет бессильна при распознавании кошек. Можно научить распознавать и лица, и кошек, но, чтобы она была сильна и в случае петухов, сеть нужно опять кропотливо обучать, при этом корректируя для всех других объектов распознавания. Выучить такую сеть распознавать сразу много разных объектов принципиально неосуществимо.
В природе все происходит совершенно по-другому. Когда созрел слой нейронов, то еще нет последующих распознавателей, нет ничего далее, что способно выявлять ошибочность распознавания, и нет того, что способно регулировать проводимость входов в зависимости от ошибок (какого-то вычислителя “обратного распространения ошибки” и регулятора).
Первоначально у созревшего нейрона на теле есть только не проводящие промежутки (щели синапсов), прерывающие контакт от предшествующих рецепторов. А когда эти синапсы становятся проводимыми, то 1) эта проводимость довольно грубая и постоянно варьируется от внешних условий (т.е. природная проводимость синапса рассчитана не на точную дозировку, а на “да” или “нет”), 2) эта проводимость в норме уже не меняется в предшествующих структурах.
Вот чем природный распознаватель отличается от реального персептрона, и поэтому мы не будет называть нейроны персептронами.
Тут стоит уточнить, что в зависимости от того, в какой зоне формируется нейрон и его синапсы, их организация проводимости может существенно различаться. В более древних зонах это и более древний механизм, обслуживающий наследственно предопределенные связи, а в более молодых структурах появляются все более новые и гибкие варианты, наработанные эволюцией для этой специфики. Но суть нейрона и синапсов остается той, какой она была в принципе определена в самых первых клетках, проявлявших базовую функциональность нейрона и синапса. Это стоит иметь ввиду и держаться за этот принцип вне зависимости от огромного множества вариантов реализации. И тогда этот принцип проведет через дебри множества несистемного в попытках построить модель понимания.
Вспомним, что между сенсорами и распознавателями воспринимаемых признаков нет тормозных связей, тогда как в персептроне могут быть как положительные, так и отрицательные веса. Зато есть то, чего нет в персептронах: боковое взаимное торможение, что решает проблему интенсивности стимула при максимальной чувствительности в тишине и контрастирует стимул, выделяя его среди всего мешающего.
Чуть отвлекаясь, заметим, что для природных нейросетей на всех уровнях характерны обратные связи (fornit.ru/5213) в силу такой находки эволюции, как принцип двусторонних связей, что уже на ранних стадиях развития мозга обеспечивало практически любые сочетания связей с богатыми поведенческими эффектами. Ветвление аксонов на фронте эволюционной изменчивости ничем не ограничивается и лишь выбраковывается в случае неудач.
Универсальный распознаватель по своей функциональности нужно представлять именно как сито, а не как персептрон потому, как в реальной нейросети нет персептронов, точнее, они реализованы в предельно простой версии.
Но, конечно, стоит разобраться в принципе работы однослойного, одноэлементного персептрона, что сегодня не представляет сложности из-за огромного числа поясняющих материалов:
· Однослойный персептрон - fornit.ru/945
· Персептрон Розенблатта - статья fornit.ru/1038
· Что такое нейронная сеть и многослойный персептрон - fornit.ru/1038
и еще вот демо-анимации:
· Работа распознавателей простых примитивов - fornit.ru/an-book-2
· Работа более сложных распознавателей - fornit.ru/an-book-3
Резюмируем основные схемотехнические принципы
· Принцип превышения порога – встречается повсеместно.
· Компарирование или принцип использования результата сравнения – важнейший для организации управляющих последовательностей, связанный с принципом превышения порога.
· Обратная связь, положительная, отрицательная и промежуточная (сдвиг фазы).
· Отсюда напрямую следуют принцип условия автогенерации и затухания. И прямое использование: если вы видите автоколебания любого вида и любой реализации, то там есть положительная обратная связь.
· Закон Ома, как это ни покажется странным, применим для любых систем, где есть движущий потенциал, есть то, что им движется и есть сопротивление (или обратное явление проводимости) этому движению.
· Принцип выделения составляющих элементов воздействующих факторов (чем на большее число составляющих разбить параметр восприятия, тем точнее и правильнее восприятие) так же дает практическое правило: самое главное – выделить важные признаки в окружающем, чтобы не махать кулаками вслепую.
· Принцип наиболее универсального распознавания – незаменим для эффективных систем управления и показывает, как следует обучаться, чтобы не пропустить важные звенья понимания, без чего остается только верить уже готовым рецептам или пробовать методом тыка.
Для нейросети получается следующая картина.
1. Нейрон как специализированная клетка сформировался на основе имевшихся ранее рецепторных клеток, которые способны выдавать электрический сигнал на раздражение и на основе эффекторных клеток, которые в ответ на электрический сигнал совершают некое действие (мышечное сокращение или продуцирование гормонов). Нейрон стал промежуточным элементом управления на основе схем непосредственного воздействия рецепторов на эффекторы.
2. Как и любая клетка, нейрон проходит стадию развития до созревания, когда он становится способен проявлять нужную функциональность, а именно: под воздействием электрического сигнала (потенциала на его мембране, запускающего регенеративную трансформацию) он выдает так же электрический сигнал и в этом похож на функции электротехнических устройств управления. И, так же как и в электротехнике в случае с чувствительными элементами, он оказывается в нестабильном состоянии, если его входы ни с чем не соединены. Это похоже на коромысло чувствительных весов, на концах которых нет никаких грузов. Такой нейрон проявляет спонтанную электрическую активность, что подтверждается данными исследований.
3. Когда нейрон формируется, он выпускает отростки в сторону рецепторов, проявляющих электрическую активность с характерно меняющимся химическим составом вокруг. Эти отростки или дендриты непосредственно касаются аксонов рецепторов или непосредственно тела предшествующего нейрона и на месте контакта возникают синапсы – потенциальные электрические соединители. Это – первый этап адаптивности к условиям среды во время раннего развития мозга, которая проявляется в активности рецепторов.
Чуть отвлечемся. Не только в раннем онтогенезе, а и во взрослом мозге описаны факты нейрогенеза (до сих пор оспариваемые, но опять обнаруживаемые в последнее время: fornit.ru/5232, fornit.ru/5360, fornit.ru/7222, fornit.ru/5287, fornit.ru/8410, fornit.ru/5429, fornit.ru/5386, fornit.ru/16232) в третичных (теменной и лобной) зонах мозга, там, где необходимо формировать новые структуры для адаптации к новым условиям (fornit.ru/5292). В то же время нейрогенез в принципе не может допускаться в уже сформировавшихся распознавательных структурах потому, как на них основаны все последующие структуры и какие-то изменения в предыдущих будет иметь эффект изменения опоры для всей конструкции.
Образующиеся потенциальные связи от нейрона с предшественниками в случае неокортекса (именно здесь возникает новое качество формирования примитивов распознавания) веерообразно перекрываются, для соседних нейронов слоя. Это – очень важный момент: в результате возникают не единичные нейроны-детекторы, а сразу ряд детекторов, частично или даже полностью дополняющих один другого. В отличие от классических схем управления, где каждое звено оказывается единственным и его повреждение - фатальным, в нейросети возникает множественное дублирование. В представленной демонстрационной модели fornit.ru/34235 это осуществляется заданными профилями вкладки “SET”, где нейроны перекрывают связями соседей.
4. Синапсы постепенно увеличивают электрическую эффективность в случае, если по обе стороны их щели есть электрическая активность. Это – простейшее условие образования связи (будем разбирать подробно). Нейрон на стадии пейсмеккерной активности самопроизвольно генерирует сигналы и если это совпадает с активностью рецепторов, связанных с ними, то нарастает эффективность связи (за счет увеличения количества нейромедиаторных пузырьков (везикул), но природный механизм не принципиален). Важно то, что процесс идет не моментально, а требует времени и за это время связь усиливается.
5. Из-за двойственной роли нейрона, он выполняет роль эффектора для предыдущего слоя рецепторов и, одновременно, роль рецептора – для последующих.
6. Во всех слоях рецепторов и эффекторов оказалось эволюционно эффективно наличие дополнительных нейронов между ними, которые обеспечивают взаимное торможение активных соседних нейронов (боковое или латеральное торможение). Это дает качественные преимущества при специализации.
7. Следующий слой созревает после того, как специализировался предыдущий. Это – принципиально важно. В природной нейросети каждый последующий слой имеет критический период развития, по длительности превышающий предыдущий потому, что частота стимулов на этом уровне оказывается значительно меньше (и еще по некоторым причинам).
Дополнительно важно то, что все выходные сигналы любых рецепторов идентичны (как в природной нейросети, так и в МНС) и сами по себе ничего не кодируют, различаясь лишь продолжительностью активности и своевременностью. Их назначение в том, откуда они вышли и куда пришли, в точности, как сигналы в цифровых устройствах.
Эта - концепция автоформирования специализации распознавателей, что обеспечивает индивидуальную адаптацию к текущим окружающим условиям, формируя характерные для них примитивы восприятия.
Первые управляющие структуры организмов на основе распознавателя
В настоящее время нервные системы простейших организмов (виноградная улитка, бабочки) исследованы очень подробно в плане организации управляющих связей между рецепторами и эффекторами. И такие связи буквально открывают глаза на то, на каких базовых принципах организованы взаимодействия между нейронами в простейших системах: это – самые тривиальные автоматы с некоторой адаптивной пластичностью.
Этот вывод очень важен, настолько же, как и вывод о том, что все организмы строятся на клеточной основе и эта основа сохраняется от самых простых до самых сложных, не меняя своего принципа межклеточных взаимодействий при всем разнообразии их воплощений. Как уже отмечалось, природа всегда использует удачное универсальное решение далее, строя на его основе новые вариации адаптивных структур.
Это очень сильный контекст для того, чтобы не привлекать к пониманию функциональности нейронов нечто, выходящее за рамки принципов организации межнейронных взаимодействий, изученных для простейших организмов. Нет никакого основания предполагать то, без чего можно обойтись в отслеживании эволюционных усложнений межнейронных взаимодействий (лишние сущности). Поэтому сейчас остановимся на древнем эволюционном этапе простых межнейронных связей.
Самое эффективное для адаптивности изобретение природной эволюции это – способность плодить новые варианты (мутации) на основе уже достигнутого. Это организуется на генетическом уровне так, что в каждом организме возникают свои специфичные, неповторимые в некоторой части бурлящих новшеств особенности. Белки одного организма оказываются несовместимыми с белками другого. Одни и те же механизмы пробуются несколько разными способами (из-за разных белков, которые – в основе всего строительного материала). То, что уже оказалось полезным и вошло в генетический код выживших, становится все более общим, универсальным и менее подверженным мутациям (fornit.ru/50319), но области новых приобретений всегда есть то, что постоянно варьируется.
Даже у очень сложного организма многое основывается на нескольких вариантов реализации, так, у людей - 50 видов гемоглобина и все они прошли испытание на жизнь, обладая теми или иными преимуществами и недостатками.
Это обеспечивают возможность выживания в новых условиях – за счет гибели всего, что к этим условиям оказывается не приспособленным.
Еще это обеспечивает постоянное усложнение потому, что ранее отработанные случайными и удачными мутациями решения, дополняются новыми полезными механизмами. Т.е. усложнения возникают не из какой-то целенаправленности развития, а просто потому, что новые качества изредка дают преимущество в выживании.
Ранее мы рассмотрели, как на основе специализированных терминальных сенсоров и исполнителей возник новый вид промежуточных клеток, которые являются и рецепторами, и эффекторами одновременно – по отношению к предыдущим и последующим по взаимосвязи клеткам. Это было случайное новшество, но оно дало преимущество большей гибкости управления поведением и осталось в генах выживших.
Недавно вышла статья (fornit.ru/b11), где показан закономерный статистический результат изменчивости и отбора к новым условиям: чем выше сложность адаптивных механизмов, тем менее вероятна полезность мутаций (в статье это сформулировано не очень корректно):
“>> вновь возникающие полезные мутации приносят в среднем тем меньше пользы, чем выше текущее значение приспособленности. ”.
Тут – формальное противоречие: если последствие мутации уже оценено как полезное для организма, то это уже не зависит от сложности адаптивной системы. Лучше было сказать: со сложностью адаптивных систем вероятность того, что новая мутация окажется полезной становится меньше. И такое утверждение очевидно верное: чем сложнее система, тем меньше вероятности, что случайное изменение в чем-то окажется полезным потому, что для адаптивности такой системы требуются и более сложные, направленные на соответствие конкретного фактора среды механизмы.
В живых остаются те организмы, которые не только оказывались в данных условиях более адекватными реальности своими адаптивными механизмами, но и те, которые способны препятствовать постоянному вреду от своей же изменчивости. И чем сложнее организм, тем более важна эта задача борьбы с вредом своей изменчивости в уже сформированных предшествующих структурах. Причем, если изменчивость затрагивает какие-то основы строения организма, то все последующие развитие идет наперекосяк и, в лучшем случае получается какой-то монстр-уродец, а чаще – просто нежить.
Вот он – еще один важный для обобщений принцип, казалось бы, тривиальный, но часто упускаемый из виду: все, на чем основывается последующее, не должно меняться, иначе это приведет к фатальной невозможности выполнять свою функцию, основанную на данных от предыдущих структур.
Отсюда, в частности, вытекает, что никаких переделок предыдущих слоев нейронов первичных зон, которые ответственны за распознавание определенных примитивов восприятия, не должно происходить (никакой коррекции, никакого нейрогенеза в этих зонах и т.п.), иначе все последующие нейроны, для которых предыдущие составляли профиль возбуждения, оказываются не у дел.
И отсюда прямо вытекает вывод о строгой по очередности развития структур нейросети, что и наблюдается как поэтапное созревание в соответствии с порядком эволюционных приобретений.
Вот почему клеточные механизмы восстановления генетического кода и борьбы с мутациями на ранних стадиях развития организма нарабатывались особенно эффективно. Так что мутация имеет шанс оказаться полезной для организма только в области самых последних эволюционных приобретений.
Такая картина эволюционной стратегии (точнее тенденции выживания наиболее приспособленных) позволяет понимать то, как появляются новые механизмы, как они развиваются, что влияет на появление новых свойств. Это – очень полезный и достаточно строгий контекст для верификации предположительной модели понимания организации управляющей сети мозга, чтобы уберечься от субъективных фантазий в этом направлении (которыми особенно отличаются всякие современные теории организации мозга), так что стоит как можно естественнее следовать тому, через что прошла природа.
В природе все тупо идет по пути соответствия новым условиям методом природного тыка, а не какой-то изобретательской смекалки. И поэтому не оправдано предполагать те механизмы, которые не следуют данному уровню эволюционных заплаток, а каждый механизм достаточно четко относится к определенному уровню последовательных усложнений.
При разработке Beast именно в силу данного принципа стало ясно, что многие свойства, придаваемые условному рефлексу классические описания в академической науке, не могут быть на том уровне, на котором формируются условные рефлексы. Это позволило очень четко определить реально возможные и необходимые свойства таких рефлексов. Стало очевидно, что то, что в академической науке приписывалось условным рефлексам – возможно только на уровне формирования более сложных структур и относится к реакциям другого типа, которые были названы автоматизмами (не путать с психологическим термином). Это изложено в отчете подробно, в разделе Условные рефлексы: fornit.ru/64975.
Так что останемся в полном соответствии с “доводом” религиозных миссионеров: вихрь на свалке случайно не может собрать самолет. Все развивается мелкими шажками, а не единым актом творения, на основе успешных предыдущих, и это является вектором эволюционных свершений.
Никаких персептронов нет в природе (это – математическая абстракция), а есть лишь отшлифованное почти бесконечными пробами и выборкой удачного устройство для взвешивания накапливающегося потенциала на входе клетки распознавателя до перевешивания порога срабатывания.
При этом нет никаких прецизионно выверенных дробных весов от рецепторов потому, что:
1) нет никакой такой возможности организовывать и поддерживать точные значения в условиях переменной внутренней среды мозга (хотя у высших животных делается все, чтобы сделать эту среду как можно более постоянной)
2) как мы уже рассматривали ранее, каждый рецептор (без исключения, начиная с самого первичного сенсора) имеет какой-то абстрагированный коррелят определенных параметров внешней (по отношению к мозгу) среды (т.е. среда организма – тоже внешняя относительно рецепторов). Но некоторая детализация весов синапсов есть, и это оказывается полезным в некоторых случаях.
Поэтому самые первые сенсоры уже могли воздействовать в полезном для выживания направлении на ответные эффекторы. Потемнело – пора скрываться, посветлело – пора выползать охотиться и плодиться, - что-то в таком роде зависимостей.
Такой эффектор мог использовать информацию (воспринимаемые данные, которые имеют для организма определенную значимость) не одного, а нескольких сенсоров, например, чтобы уползать не тогда, когда чуть потемнело на короткое время, а когда достаточно надолго потемнеет.
Когда мутация сделала из конечного эффектора не сократитель мышц или выделитель регулирующего вещества, а клетку, способную как рецептор выдавать в ответ потенциал, то возможность усложнения схем управления получила новый качественный уровень. Теперь случайные комбинации связей образовывали как вредные, так и бесполезные связи с эффекторами через таких посредников, среди которых в новых условиях ответные действия оказывались выигрышно полезными для выживания.
Вот так тупо формировались наследственно предопределенные реакции: никакого обучения, никаких “глубоких многослойных персептронных сетей” с элементами, связанными все со всеми. Просто – случайные связи с полезным или вредным последствиями, или никаким эффектом.
И этот процесс продолжается, но уже с базы достигнутых изменений – только в области нового. Поэтому те виды животных, что проскочили данную стадию мутагенной вариабельности, больше уже никогда не смогут использовать то, что в данном месте получили другие виды животных. Вот почему из обезьяны и других уже сформировавших внутренние структуры видов никогда уже не возникнет человек.
Исследователи замечали те рефлексы, которые проявляли себя в моторных, наблюдаемых действиях. Но не меньшая работа эволюции шла и для структур, активность которых влияла не на моторные функции. Рефлексы мозжечка были замечены как следствие очень большого по объему скопления нейронов в этом органе и по тому, как, опять же, такие рефлексы проявлялись в моторных действиях. Но механизм образования и функционирования рефлексов мозжечка очень специфичен и не похож на другие наследуемые рефлексы типа стимул-действие. Так же возникает и множество других, пока не обнаруженных исследователями в определенной функциональности наследуемых структур мозга.
У человека и высших животных в области лобной коры наследственно возникали особые структуры, обеспечивающие обработку активностей предшествующих по развитию структур, то, что в отчете о Beast было названо “информационными функциями” (их роль описана в резюме по отчету: fornit.ru/65133). Каждое появление такой структуры давало широкие вариации реализованных надстроек, что усложняли такие структуры, совершенствуя полезность их функционирования. Их совершенствование шло точно так же и по тем же принципам, что совершенствование инстинктов (длинных цепочек безусловных рефлексов), только инстинкты проявляли свою функцию в моторных действиях, а инфо-функции – во “внутренних” регулировках активностей, позволяя работать принципиально не алгоритмизированным (заранее не прошитым). Виды животных, которые нарастили поверх данной функции что-то еще, уже не могли совершенствовать эту функцию. В конечном счете потенциал адаптивных возможностей зависит от числа различных видов инфо-функций и их эффективности. И такое различие есть не только у разных видов животных, но и у разных подвидов, что и определяет наследственные предрасположенности к “уму и сообразительности” у разных людей и других животных разных пород (хорошие-плохие гены).
Вернемся к основам формирования наследственно предопределенных структур.
Описанный принцип образования наследуемых связей базируется на имеющихся свойствах промежуточных клеток (нейронов): спонтанной активности при созревании, но отсутствии эффективных связей, и механизме увеличения проводимости синапсов при одновременной активности с двух сторон. Этот механизм сам в свое время прошел отбор из вариантов, большинство из которых просто ни к чему полезному не приводили, так что остались те варианты, которые реально обеспечивали связи с предыдущим рецептором и возможность образовывать цепи воздействующих групп рецепторов на промежуточные нейроны и конечные эффекторы.
Таких вариантов реализации проводимости синапса очень немало. Кроме того, что варьируются вещества, обеспечивающие проводимость синапса (нейромедиаторы), но и самих механизмов организации передачи возбуждения возникло несколько, вплоть до того, что вместо передачи возбуждения сигнал от рецептора может оказывать не возбуждающее, а тормозное воздействие. И это тормозное действие оказалось очень полезным для схемотехники управления нейронами потому, что:
1) позволяет организовывать намного более гибкую логику
2) позволяет бороться с паразитными возбуждениями, которые могут привести вся сеть нейронов в перевозбужденное состояние, т.е. вместо управления устроить эпилептический припадок.
Особенно полезным тормозное влияние оказалось для соседних нейронов: если возбудился какой-то нейрон, то соседи притормаживаются, уже не срабатывая от случайных шумов чувствительных рецепторов.
Теперь мы подходим к пониманию схемотехники, обеспечивающей автоматическую (без коррекции ошибки и какого-либо задатчика цели) специализацию нейронов.
Когда связь формируется между активным рецептором и созревшим, спонтанно активным эффектором, проводимость нарастает весь период активностей рецептора, пока есть спонтанная активность эффектора. Как только связь окажется достаточной, чтобы прекратить спонтанную активность эффектора (вспомним легкость разбалансировки ненагруженных коромысел или склонность к возбуждению электронных элементов с незадействованным входом), то на этом процесс и останавливается.
Дальнейшая модификация связей на этом уровне становится невозможной, если не соблюдается условие: активность с обеих сторон синапса. Повреждение ткани мозга приводит к разрыву связей и, значит, опять провоцирует спонтанную активность, что может быть сигналом для регенерационных процессов.
Если же активными одновременно бывают несколько рецепторов, связанных пока непроводящими синапсами с созревшим эффектором, то так же проводимости нарастают, пока эффектор не прекратит самопроизвольную активность и тогда получается, что к этому моменту каждая из связей нарабатывает проводимость меньше, чем если бы была только одна связь. Теперь эффектор возбудится только при одновременной активности всех рецепторов, которые участвовали в формировании связей. Возник специализированный образ сочетаний входных активностей. Чуть отвлекаясь, заметим, что если по какой-то причине нейрон оказывается активированным вне своего специализированного профиля рецепторов, то он опять оказывается способным образовывать связи и корректировать специализацию. Но такое не характерно для первичных структур.
Процессу специализации предшествует механизм, когда у созревающего эффектора начинают тянуться выросты (дендриты) в сторону ближайшей активности (туда, где соответственно изменился химический состав среды) и там образуются синапсы.
Формирование новых дендритов так же прекращается после специализации, что еще более стабилизирует получившуюся конфигурацию связей.
Так появлялись все адаптивные механизмы простейших животных, и основы этих механизмов остались преемственны для человека.
Итак, и внешний мир, и внутренняя среда организма – это внешние по отношению к мозгу области получения данных о значимости пользы и вреда, когда польза означает возможность жить в данных условиях, а вред – гибель организма и обрыв передачи генетического кода потомкам. Смерть – главный двигатель эволюции животных.
Внешняя среда отслеживается первичными сенсорами без какой-то оценки того, что это за данные (из внешнего мира или внутренней среды), просто мутации перестраивают его в некоторой наиболее вариабельной (мутагенной) части механизмов, и особи, которым посчастливилось выжить, передают генетический код (fornit.ru/38136).
В анимации “Простейшая нейросеть управления крыльями бабочки” (fornit.ru/an-book-9) показан принцип реализации участка такой сети. И можно себе вообразить, сколько же нужно было времени и погибших бабочек, чтобы методом случайных мутаций организовать такую сеть из уже обкатанных клеток распознавателей и синапсов между ними. Но количество бабочек и количество времени эволюции очень велико, а скорость смены поколений у насекомых – очень высока (у одноклеточных и вирусов – еще больше такое преимущество).
Конечно, все накопления полезного не возникли сразу как самолет из мусора в притчах религиозных миссионеров, а оптимизировались по шагам отдельных попыток в огромном количестве удачных и неудачных последовательных шажков при огромном разнообразии вариантов.
Схемы, наподобие анимации для бабочки, – хороший тренинг для развития схемотехнического мышления в представлениях моделей простейших нейросетей. И это важно для того, чтобы подготовиться к пониманию более сложных механизмов, которые возникли в ходе эволюции и придали новое качество возможности управления. Вот этим мы некоторое время и займемся.
Для развития схемотехнического мышления в игровом стиле создан Конструктор нейросхем с проверкой их работы: fornit.ru/41239. В нем заготовлены демонстрационные модели, иллюстрирующие простейшие реализации схемотехники нейросетей. Этот эмулятор позволяет моделировать и проверять различные фрагменты нейронных соединений.
В схемах нейроны (распознаватели достаточного веса активирующих их сигналов) будем обозначать кружками, синапсы – щелью (похожей на обозначение конденсатора в эл. схемах), а отростки клеток к этим связям – линиями.
Каждый может сам попробовать воплотить разные моменты простейших поведенческих реакций насекомых, в том числе и уползание улитки, когда наступает темнота и, уже сложнее, - распознавание врага по резкому изменению освещенности. Для наглядной эмуляции таких схем создан специальный инструмент: fornit.ru/34235.
Другой нейросимулятор fornit.ru/ns1) позволяет исследовать поведение нейросетей, задавая их основные параметры: веса активирующих и тормозных связей, и, что очень важно – уровень взаимного торможения в слое (латеральное торможение). Это позволяет верифицировать схемотехнические предположения и интерпретировать поведение реальных нейросетей. Описание нейросимулятора: fornit.ru/40662.
Далее последуют некоторые примеры практического воплощения принципиальных схем, интересные тем, что позволяют наглядно увидеть проблемы, решаемые эволюцией и то, какую наиболее правдоподобную модель можно представить для конкретного назначения адаптивного механизма.
В качестве практического примера рассуждений над возможным наследственно предопределенным механизмом, приведем принципиальную схему позиционирования головы насекомого так, чтобы попавший в поле зрения фасеточного глаза объект всегда находился в центре сенсорной матрицы. Для этого нужно дать команду сокращения одной из 4 мышц, управляющих глазом, для его поворота вверх, вниз, вправо, влево соответственно. Поворачиваться голова будет дискретно, то есть один командный импульс – один шаг поворота (неважно, как это устроено на самом деле). Для начала возьмем минимальную матрицу 3х3. Задача – в зависимости от положения в ней активного элемента задать направление перемещения, чтобы активным стал центральный. Для наглядности направление перемещения будем считать, как направление смещения пятна на матрице, хотя на самом деле надо конечно делать наоборот, ведь не пятно движется, а поворачивается голова: чтобы пятно сместилось вниз, голова должна повернуться вверх, для левого смещения – поворот вправо и т. д.
![]()
Линейка с верхними квадратами – номера ячеек сенсорной матрицы 3х3, кружок – нейрон, квадраты внизу – эффекторы, обозначенные как П (мышца для поворота вправо), Л – влево, Н – вниз, В – вверх. По схеме видно, что для угловых ячеек 1, 3, 7, 9 нужна одновременная работа пары мышц, то есть для перемещения из сектора 1 в центральный сектор 5 нужно задействовать эффекторы П и В. Это означает, что к некоторым ячейкам сенсора будет подключено по два входа от соответствующих управляющих нейронов. Такая схема имеет 2 серьезных недостатка:
1.Подключение к сенсору более одного входа от нейронов означает распределение по ним сигнала, потерю мощности – нужно ведь по обоим направлениям подать одинаково мощный сигнал.
2.Схема подразумевает строгое подключение каждого рецептора к своему нейрону. Если представить вероятность попадания в ходе отбора путем случайных мутаций для матрицы 1000х1000 – даже без расчетов очевидно, что она будет очень низкой. Стало быть, нужна схема с менее жесткой привязкой, с большей степенью свободы установления связей.
Для этого сделаем перемещение от угловых ячеек к ортогональным. Тогда каждая угловая ячейка получит две степени свободы. Например, от ячейки 1 можно двинуться вверх или вправо – в любом случае попадаешь на центральную ячейку 4 или 2, а оттуда уже строго к центру. То есть вместо одного строго определенного движения получили два, но зато равновозможных. При этом заодно избавились от двойного подключения к рецептору от разных нейронов.
Теперь к угловым ячейкам рецептора может без потери функциональности устройства равноценно прицепиться один из двух нейронов. Это для матрицы 3х3 создаст 128 возможных схем соединения вместо одной единственной.
Понятно, что чем больше размер сенсорной матрицы, тем больше количество схем подключения. Но можно еще добавить хаоса. Например, в матрице 1000х1000 раскидаем примерно равномерно ячейки, к которым может цепляться вообще любой нейрон. Тогда даже если при реакции сенсора движение будет в противоположную сторону, соседняя ячейка направит ее в нужном направлении. Это, понятно, замедлит общую скорость стабилизации положения, но здесь только нужно найти ту степень хаоса, при которой это затормаживание будет в пределах допустимого.
В итоге получили схему стабилизации головы насекомого, слежение за объектом. Но, по сути, это схема стабилизации любого параметра, например гомеостаза. Если вместо ячеек сенсорной матрицы подразумевать нейроны-рецепторы, а вместо мышц – железы-эффекторы или что-то иное, что требуется совершить для прихода в норму. Главное, что становится очевидно – такая схема вовсе не требует жесткой юстировки (настройки) параметров. Она допускает довольно широкий их разброс, что позволяет теоретически собрать ее эволюционным путем естественного отбора.
При том в голову приходит, что хотя у насекомых практически вся нейросхема предопределяется геномом, но это не жесткий шаблон, а весьма вольная трактовка. Это объясняет, как так удается передавать по наследству довольно замысловатое поведение. А оно и не передается в точном исполнении, но в виде каркаса с допуском вольностей исполнения. В итоге все тараканы при включении света на кухне стремительно сваливают под плинтус – но каждый по своей, замысловатой траектории, но с одинаковым стремлением к ближайшему укрытию.
В подобных принципиальных схемах нейросети возможны две основных логических схемы соединения: логика И и логика ИЛИ. Этим устанавливаются граничные условия возможной реализации тех или иных механизмов.
![]() Логика И
Логика И
Логика И (и тут, и там) - означает, что нейрон сработает только если на всех его входах будет сигнал, в сумме достаточный, чтобы превысить порог. В данном случае такая сумма будет 1,2 — это выше порога 1. Порог это - не 1V - а значение напряжения, при котором срабатывает устройство. На схеме это Uп (напряжение порога).
На схеме слева показаны рецепторы 1, 2, 3. От них к нейрону через синапсы поступает сигнал. И тут особенность схемы "И": раз мы имеем 3 входа, то, чтобы в сумме они набрали выше порога, надо разделить 1 на 3 и чуть добавить (для надежности, но не обязательно). В итоге получили коэффициент 0,4 (0,333... округленное в большую сторону). Было бы 10 входов - на каждом будет по 0,1. А если 1000 - 0,001. Но в этом случае потенциал будет настолько низок, что это будет, по сути, случайные флуктуации - не получится добиться надежного срабатывания устройства. Стало быть, несложно догадаться, что для его успешной работы не стоит использовать слишком много входов.
Но тогда возникает вопрос: известно, что у нейрона их может быть до 10 0000 и даже больше. Но тут уже работает другая логика - ИЛИ (или тут, или там).
![]() Здесь, как видим, синапс не уменьшает сигнала. Стало быть, если хоть один из входов получит сигнал — это уже 1 и нейрон срабатывает. Если больше одного - тем более сработает.
Здесь, как видим, синапс не уменьшает сигнала. Стало быть, если хоть один из входов получит сигнал — это уже 1 и нейрон срабатывает. Если больше одного - тем более сработает.
В случае некоей мухи: если один датчик зафиксировал тень от объекта (предположительно хищник), а другой показывает свет (солнечный) день - тут налицо логика срабатывания "И", реакция бегства при наличии двух обязательных факторов.
Важное следствие из всего сказанного: у нейронов природной сети в отличие от искусственной не нужно устанавливать (подгонять) точные веса синапсов, хотя они и имеют некоторые градации – как следствие того, насколько успела укрепиться данная связь пока нейрон пребывал в режиме формирования усредненного профиля активности. Они все имеют примерно одинаковый вес. Потому, что каждый слой просто определяет наличие или отсутствие "своего" примитива в реакции предыдущего слоя. Для этого достаточно простейшей логики И / ИЛИ.
О роли «подгонки весов» (формировании профиля синаптических связей) в природных сетях - «теория векторного кодирования» (Соколов Е.Н.(2003). «Восприятие и условный рефлекс. Новый взгляд»; Соколов Е.Н. (2010). Очерки по психофизиологии сознания). И вопрос о том, с какой точностью они формируются, остается открыт ровно до тех пор, пока не будет определена верная модель процесса специализации нейрона, а она оказывается одним из базовых принципов в организации нейросети. Но очевидно, что большая точность веса не может обеспечиваться из-за не идеальности природного механизма реализации и определенной нестабильности среды, которая, к тому же, влияет и нас порог срабатывания нейрона.
В итоге постоянного бурления наследственных версий организмов (чем большая численность популяции – тем эффективнее) может возникать и возникает множество разных конфигураций, в том числе абсурдные и сложные, но все это не проходит отбор на эффективность адаптивности. В целом формируется иерархия контекстов представительства тех или иных признаков, а не стоит задача распознавания сразу сложного их сочетания.
Так же и коллатерали аксонов и их ветвление к целям – дело изменчивости и проб, хотя среди таких проб и остается нечто реально полезное.
Такое эволюционное разнообразие проявляется буквально во всем.
У разных животных разное число цветовых сенсоров, у человека их 3. Так что кодировка цвета (оттенки) у человека возникает как сочетание этих трех “RGB”. С яркостью все намного сложнее, потому как она зависит от активности соседних распознавателей: если они молчат (темнота), то возбуждение даже от одного фотона может дать вспышку (на фоне некоторого спонтанного шума молчащих, но то и дело возбуждаемых всякими флуктуациями окружающей среды рецепторами). Из-за взаимного бокового торможения в слое реагирование нейрона на яркость притупляется и поэтому воспринимаемая яркость оказывается относительной величиной.
Даже у одного вида животных несколько различается предварительная обработка (предопределяемая наследственно) от сенсорной сетчатки, а именно здесь упрощается и корректируется поток информации, поступающей в зрительную кору (до которой мы еще не добрались).
У более простых животных, у которых нет “новой” коры мозга, происходит непосредственное использование той упрощающей предобработки, которая оптимизировалась миллионы лет. Как именно у разных животных это организовалось, до сих пор нет определенных сведений, кроме самого факта, что предобработка есть.
Можно нарисовать множество вариантов предварительной обработки, и каждый из них дает какие-то преимущества в определенных условиях.
И только на уровне новой коры мозга возникли более однотипные и универсальные механизмы формирования примитивов восприятия.
В целом вся нейросеть обменивается стандартными по амплитуде сигналами, не имеющими никакой корректировки кроме того, что данный сигнал означает корреляцию с неким набором окружающих факторов, которые можно учитывать в организации ответа – в виде абстрактных (выделенных) признаков восприятия. На выходе клетки с электрической активностью нет учитываемого (информационного) параметра интенсивности. Она или возбудилась стандартным выходным импульсом или нет. Если она долго возбуждается, то идет пачка импульсов, поддерживая наличие стимула, но в более поздних наработках эволюции возник механизм удержания отклика стимула, даже если он пропадает – за счет обратной связи, поддерживающей постоянную активность, что оказалось принципиальным прорывом в возможностях индивидуальной адаптивности. Так, если муха скрылась за травинку, то ее образ не исчезает, а продолжает воздействовать на поведение охотника.
Старый вопрос о том, чем кодируется в нейросети информация, сегодня сводится к двум разумно обсуждаемым вариантам: «номером канала» и/или частотой разрядов. То, что любая информация получает смысл в зависимости от того, откуда она пришла относится к любым случаям распознавания. А вот наличие латерального торможения и, как следствие, отсутствие информации об абсолютном значении рецепторного параметра делает значение частоты импульсов как информации об интенсивности спорным. Кроме того, динамический диапазон такого кодирования очень груб из-за множества факторов, влияющих на частоту импульсов нейрона. Это никак не может объяснить огромный динамический диапазон оценки, например, яркости источника света, который получается именно относительным способом (и недостижим для прямого измерения даже современными чувствительными матрицами фотоаппаратов), который демонстрируется в демо-модели fornit.ru/38103, где параметр интенсивности коррелирует с положением ползунка силы латерального торможения. Этот алгоритм легко реализуется схемотехнически (легче, чем он был реализован программно в данной модели, т.к. там для получения результата проводится десятикратная итерация постепенного приложения величины торможения к совокупности нейронов).
Как есть отдельные распознаватели цвета, так есть и отдельные распознаватели общей и локальных интенсивностей в их относительном значении, и они далеко не идеальные, но обеспечивают большой динамический диапазон оценки интенсивности параметра. Сочетание этих параметров у высших животных происходит во “вторичных зонах” мозга и окончательно - в третичных зонах - как взаимодействие иерархии контекстов, смотрите иллюстрация организации памяти образа на основе отдельных примитивов восприятия - fornit.ru/460.
Это приводит к выводу о том, что частота импульсов от нейрона кодирует не силу стимула в нейросети, а только - продолжительность его наличия.
Есть еще немного выводов из схемотехнических принципов для нейросетевого адаптивного управления, которые способны вызвать настороженность у нейрофизиологов. Это пока – только постулаты, возникшие как следствие учета схемотехники управляющих структур на основе универсального распознавателя – нейрона.
Речь пойдет о процессе формирования проводящего синапса. Неважно, какие механизмы для этого задействованы, общий смысл сводится к тому, что синапс между двумя активными клетками начинает наращивать элементы (пузырьки или везикулы) для выброса нейромедиатора, которые и оказывает поляризующее действие на мембрану последующего нейрона. Чем больше пузырьков сформируется, тем эффективнее связь. Понятно, что для их образования нужно время. Вот и получается, чем дольше экспозиция (время воздействия активного фактора среды), тем больше пузырьков. И важно общее время экспозиции, а не однократное, т.е. при повторяющейся время от времени обоюдной активности, растет число пузырьков. И это время длится, пока нейрон не окажется специализированным, т.е. более не проявляет спонтанную активность.
Можно представить, как развиваются подобные схемы, на основе наследственно предопределенных ветвлений аксонов от предшествующих сенсоров. Т.е. речь идет не о мутациях, а о вполне устоявшейся “норме” развития.
![]() На схеме – случай, когда к центральному нейрону наследственно подходят аксоны от двух типов сенсоров. И вот, центральный нейрон созрел, а к этому времени уже и подавно работают сенсоры (потому как они эволюционно развились раньше).
На схеме – случай, когда к центральному нейрону наследственно подходят аксоны от двух типов сенсоров. И вот, центральный нейрон созрел, а к этому времени уже и подавно работают сенсоры (потому как они эволюционно развились раньше).
Нейрон созрел, перед ним то и дело возникают активности предшествующих двух аксонов, там меняется хим. состав, что является приманкой для ветвления дендритов созревшего нейрона. Они растут к аксонам, образовывают синапсы и начинается самое интересное.
Т.к. эффективных связей пока нет, нейрон оказывается в неустойчивом равновесии потенциала на своем теле (как ничем не нагруженные весы колеблются от всего). А в это время со стороны аксонов тоже возникают активности во время отклика на раздражение сенсоров. Понятно, что активность двух аксонов разная, потому как то к одному сенсору что-то прикоснется, то к другому, иногда к обоим сразу. Ровно так же начинают формироваться и связи, накапливая активные элементы в синапсах. Все зависит от того, с какой частотой (внешнего воздействия) активируются каждый из двух сенсоров.
Для определенности предположим, что верхний сенсор откликается на прикосновения к усикам, а нижний – к телу на голове.
И вот тут возможны варианты. Предположим, что в этой местности улитку чаще кто-то или что-то тревожит за усики, чем за голову. Тогда связь от верхнего сенсора будет укрепляться быстрее и, фактически, центральный нейрон, после завершения специализации (когда уже не будет состояния неустойчивого равновесия, и он прекратит самопроизвольно генерировать), станет детектором прикосновения к усикам, а на прикосновения к голове будет реагировать более вяло, только когда прикосновение будет продолжительным.
Такое же может быть и с сенсором головы в случае более частого раздражения.
А в случае примерно одинаковой раздражительности обоих сенсоров, веса синапсов поделятся примерно поровну и будет логика И.
Получается, что логика ИЛИ так не формируется и для этого нужны другие условия, другое ветвление аксонов.
Но главное: улитка приспособится реагировать именно к местным существующим условиям.
Далее переходим к детальному рассмотрению природных нейросетей в контексте схемотехнических принципов. Но начать придется с физико-химических аспектов природной реализации, а для этого нужны базовые представления по химии, которые можно получить в статье fornit.ru/1139, где выделено самое важное. Тем, у кого есть неуверенность понимания химии, нужно сначала переключиться на эту статью.
Очень полезно прочесть эту статью прямо перед сном и не задумываться сильно при этом, а лишь отметить места недостаточности понимания. Поутру, перед мыслями о предстоящем дне, нужно будет без напряжения вспомнить прочитанное.
Из химии важно усвоить понятие ионов и то, что ионы из многих нейтральных молекул могут получаться при растворении солей (молекул, связанных за счет электростатических сил притяжения) в воде, образуя носители электрического тока. Чем выше температура, тем подвижнее носители заряда в водной среде, очень существенно (в несколько раз при изменении на 10 градусов) меняя условия воздействия этих зарядов. Поэтому для стабильной работы нейросети необходимо поддерживать температуру на оптимальном уровне.
Химия электротоков и ее воплощение в нейросети
Схемотехника электрических цепей это – всегда источник электропитания и элементы в цепочке передвижения тока, которые что-то делают, благодаря этому току. Все, что имеет электрический заряд, стремится приблизиться к противоположному по знаку другому заряду, и, если нет препятствий, то возникает такое движение или электроток того, что движется. Отрицательными зарядами обладают электроны, а положительными – протоны. Все, больше нет в обычной жизни других предельно простых носителей зарядов в веществе. Там, где электронов оказывается больше, чем протонов, будет отрицательный заряд и наоборот.
Заряды – вещь относительная. Если есть два отрицательных заряда, но разные по величине, то один из них – более отрицателен и, значит, есть разность их потенциалов, заставляющая все электрическое пытаться двигаться в направлении этой разницы. Эта разность потенциалов (величин) зарядов – основа питания любых электроустройств и любой схемотехники использования электротока в своих целях. Природные нейросети – не исключение.
Источник питания всегда имеет два электрода - концы с разными зарядами, один из которых будем называть более отрицательным по этому заряду, чем другой потому, что там скопилось больше электронов, имеющих отрицательный заряд из-за невозможности преодолеть этот промежуток. Если между электродами оказался проводник электронов, тогда по нему потечет ток электронов от минуса к плюсу (или можно условно посчитать, что наоборот, что совершенно не имеет значения, кроме электронов, электроток может быть вызван движением других заряженных частиц – ионов – более массивных, чем электроны осколков вещества, а ионы могут быть как положительными, так и отрицательными).
Природа в растворах электролитов мозга использует ток ионов, т.е. больших (по сравнению с электронами и протонами) атомов или даже групп связанных атомов (молекул), у которых есть избыток или электронов или протонов.
Простейшим электронейтральным атомом является водород. Если у водорода отнять его электрон, то останется голый протон. Протоны в воде дают кислую реакцию, т.е. стремятся соединиться с тем, что имеет более отрицательный электрический заряд, а это могут быть другие атомы или молекулы с избытком электронов. Кислый раствор протонов – простейший электролит, в котором потечет ток протонов от более положительного электрода к более отрицательному. И этот электроток можно как-то использовать.
Если число электронов равно числу протонов в этой области пространства, то она (эта область) будет электронейтральной, так что все электричество основано на нескомпенсированных количествах электронов и протонов, между которыми начинается ток носителей электричества – свободно движущихся в среде зарядов (свободных электронов, протонов или ионов).
Для схемотехники важно одно правило: ток всегда потечет между разницей зарядов (потенциалов заряда), если между ними есть проводник для этого вида носителей зарядов.
Представление об электротоке важны для того, чтобы нарабатывать навыки сразу видеть, что будет с электродеталью при воздействии электротока. Навыки понимания и использования электросхем как раз основываются на представлении о том, что будет, когда потечет ток в цепи и будут оказываться воздействия на электродетали схемы. Это должно стать зримой картинкой.
Ток носителей заряда возникает, как только для этих зарядов, помещенных между разностями потенциалов, окажется возможным перемещаться в проводнике тока. Каким бы ни был длинным проводник, ближайшие к источнику потенциала носители заряда начнут движение (с одного конца притягиваясь, а от другого отталкиваясь) и это создаст потенциал для следующих носителей.
Сами носители передвигаются по проводнику довольно медленно (со скоростью распространения тепла в этом проводнике), но взаимовлияние между ними распространяется со скоростью света в вакууме, а не в данном проводнике потому, что происходит не перемещение фотонов с переизлучением от атома к атому, а поляризация носителей заряда – влияние с помощью поля притяжения-отталкивания. И волна этой поляризации распространяется со скоростью света в вакууме.
Вокруг каждого заряда его поле поляризует все вокруг, вызывая притягивание всего противоположного по знаку и отталкивание схожего.
Если электроток течет с постоянной скоростью (потенциалы источника не меняются по величине), то и поляризация постоянна, но если потенциалы все время меняются, то и воздействие поляризации на окружающее оказывается с той же частотой. И тогда в проводнике, расположенном рядом, но не подключенному к источнику разницы потенциала, возникает та же частота колебаний его носителей заряда. Так работают приемные антенны, трансформаторы и любые другие электродетали, использующие вовлечение своих носителей в ту же динамику, что и носители в рядом расположенном проводнике, разгоняемые разностями потенциалов.
Если подключить такие пассивные проводники (антенные электроды) к голове, то в них начинают возникать колебания, отражающие все поляризационные влияния электротоков мозга, в некоей общей суммарной картинке. Чем ближе электрод к цепи, наводящей в нем поляризационный отклик, тем сильнее будет и наводка – в полном соответствии с законом Кулона: сила притяжения-отталкивания равна произведению величин электрозарядов, деленная на квадрат расстояния между ними. Т.е. чем ближе заряды, тем сильнее между ними взаимное влияние.
Закон Кулона формально в точности такой же, как закон тяготения Ньютона, только во втором имеются ввиду не электрические, а гравитационные заряды. Схожесть этих двух (и не только этих) законов взаимодействия вызвана тем, что эти взаимодействия используют степени свободы трехмерного пространства, а в нем удаление зарядов вызывает уменьшение их потенциала именно обратно пропорционально квадрату расстояния, что легко видеть, если нарисовать два заряда и провести лучи во все стороны от них: плотность этих лучей будет уменьшаться по такому закону от расстояния между зарядами.
Это представление о природе электровзаимодействия не понадобится для понимания работы электрозарядов в нейросети мозга, но дает понимание причин и следствий в утверждениях, а не просто веры в них. Конечно, существует граница разумного, насколько глубоко погружаться в природу явления и то, что касается зарядов носителей электротока не требует более глубокого понимания природы, тут достаточно интуитивного понимания как в случае с опытом наблюдения поведения магнитов. Поэтому все сказанное желательно воспринимать достаточно буквально, без попыток разобраться в природе всего этого. Важно сделать привычными основные свойства участников явления электропроводности, а не то, из чего они состоят.
Итак, суть электротока сейчас должна быть зримо ясна. Но важно и понимание количественных взаимовлияний. Для электротока есть понятие его силы – как количества носителей, текущих в единицу времени, измеряемое в Амперах. Все знакомые с проблемой зарядки смартфонов имеют уже достаточно ясное представление, насколько много или мало это 1 или 5 ампер, гарантируемых в течение одного часа. Если батарейка обеспечивает ток в 4 ампера в час, то считается, что это неплохо для смартфона и ее хватит надолго, если не слишком много заставлять потреблять гаджет, чтобы он начал не сильно греться.
Носители заряда, протискиваясь через решетку структуры проводника, расталкивают ее, добавляя энергии к тепловым колебаниям атомов, и эти атомы начинают колебаться сильнее, а температура смартфона повышается.
При этом говорят, что в данной цепи для электротока имеется определенное сопротивление его протеканию, которое носители начинают преодолевать, частично застревая, отдав свою энергию или просто даже не в состоянии протиснуться между атомами проводника.
Величина разницы потенциалов источника питания (электронапряжение) – тоже хорошо понимаемо на бытовом опыте: несколько вольт запросто можно попробовать на язык, а вот 220 вольт – это уже слишком.
Источник в 1 вольт вызовет электроток в проводнике в 1 Ампер, если сопротивление проводника будет 1 Ом. Это называется законом Ома, который уже был упомянут в механической аналогии. В контексте общей схемотехники будем просто говорить о напряжении (U), сопротивлении (R) и токе (I) без приставки “’электро”: I=U/R;
Итак, ток может быть слабым, если носитель не способен пропустить через себя много зарядов и тогда даже миллион вольт не принесут нам вреда, протекая по телу потому, что столь мизерное количество электронов не сможет что-то серьезно повредить, или ток может быть сильным, как если на клеммы автомобильного аккумулятора уронить гаечный ключ.
Далее будет немало случаев, когда нужно будет моментально прикидывать реализацию тех или иных механизмов, это нужно будет привыкнуть делать автоматически буквально для всех механизмов, без чего не получится понять их причинность и взаимодействие.
В нейросетях источник питания распределенный, а не какая-то батарейка в одном месте. Каждый нейрон сети мозга – источник электроэнергии за счет разности потенциалов ионов в растворе, окружающем его и растворе его тела. А оболочка нейрона не дает возможности этим зарядам перемешаться, изолируя их. С обеих сторон мембраны клетки нейрона образуется электрический потенциал из-за различия не только концентрации электролита сред, но и состава солей в них.
Даже если бы с двух сторон оболочки нейрона была просто разная концентрация одной и той же соли, то потенциалы скопившихся ионов оказались бы разными, а то, что и соли разные, еще больше повышает разность потенциалов.
Мембрана клетки нейрона так устроена, что при определенном условии она перестает быть препятствием для ионов, и они взаимно гасят разность потенциалов.
Это становится возможным, если каким-то образом (совершенно не принципиально каким) начать изменять электрический потенциал на мембране. С какого-то порогового значения, мембрана станет проводником тока.
При этом возникает перепад напряжения в виде импульса тока (от его пика разности потенциалов до полной релаксации в однородность электролита вне нейрона и внутри его). Этот процесс гистерезисный (потенциал старта больше, чем потенциал продолжения процесса) и он распространяется по мембране нейрона вдоль его отростков к другим нейронам.
Клетка нейрона после такой релаксации включает ионные насосы, которые перемещают смешавшиеся ионы опять по разные стороны мембраны, после чего она готова к новому импульсу в ответ на поляризующее воздействие.
Мы не станем углубляться в очень сложные механизмы работы мембраны и другие сопутствующие явления, которые пришлось разработать природе. Нам важно понять, что вот таким образом организованы источники тока в сети нейронов и так они обмениваются импульсами активности.
Все это можно было бы создать искусственно куда более простым и лучшим путем, в том числе и вовсе без каких-то потенциалов, а программно. Но нам важно также не упустить все то, что представляет принципиальную основу причин и следствий природного механизма обмена импульсами.
Далее будут формироваться электротехнические принципиальные модели основной функциональности нейрона. Тем, кто вообще не имеет представлений об электронике, очень стоит сначала пройти блиц-курс для понимания ее основ: fornit.ru/1140
Схемотехнические принципы в модели нейрона
Сегодня немало фирм, например Intell, разрабатывает, совершенствует и выпускает в серию специальные микросхемы для построения нейросетей. Они уже проделали работу по выявлению основной универсальной функциональности нейрона в связке с его синапсами и организовали модули, реализующие сразу по несколько десятков таких элементов на одном чипе. По сравнению с попытками реализации на компьютере, такие сети на дискретных, самостоятельно работающих элементах, обладают несравнимыми по эффективности преимуществами отсутствия необходимости просчитывать состояние всей сети центральным процессором.
Но к идеализированным моделям функциональности нейрона нам нужно еще прийти с достаточной очевидностью, а для этого совершенно неизбежно придется погрузиться в начальные уровни понимания электро-схемотехники. И такое предстоящее погружение может показаться многим неуместным и излишним, но это – иллюзия непонимания, которую можно избегнуть, только достаточно освоив те промежуточные представления, без которых нет шансов продолжать с пользой читать эту книгу…
Вспомним из предисловия: “Теперь самое интересное. Этим исследователям попался настоящий марсианин – схемотехник компьютерных систем. И тут начинается чудесное: все попытки схемотехника что-то объяснить исследователям компьютера упираются в стену непонимания, ведь те не потратили годы на освоение принципов конструирования компьютеров и не понимают даже самые простые вещи в этой области”.
Чтобы понять механизмы схемы марсианского компьютера – нужно понимание схемотехники, а не изучение свойств его деталей и проводников.
Как уже говорилось, причиной электротока является разность потенциалов и среда между этой разностью, позволяющая носителям заряда перемещаться от одного потенциала к другому. Эта среда обладает каким-то сопротивлением, ограничивающим количество зарядов, проходящих по проводнику в единицу времени (сверхпроводимость не рассматриваем).
При протискивании носителей тока через мешающие им структуры проводника, носители отдают им часть своей энергии движения, нагревая проводник. А нагретый проводник еще больше мешает движению носителей своими тепловыми колебаниями, цепляя еще большее число носителей.
Возникает отрицательная обратная связь: ток нагревает проводник, но при этом он сам уменьшается из-за возрастающего сопротивления. Поэтому нити накаливания лампочки раньше использовали как стабилизаторы тока: при повышении напряжения ток пытается увеличиться, но тут же резко увеличивается температура, и току мало удается измениться. При изменении напряжения сильно меняется яркость нити накаливания, но ток при этом меняется слабо.
В схемотехнике и жизни именно электроток (поток воздействующих на что-то зарядов) имеет значение, вне зависимости от вида сопротивления среды этому току. Поэтому мы можем просто пользоваться обезличенным понятием сопротивления току, путь даже его носители переносятся самым экзотическим образом, хоть даже обезьянами или демонами, или как, например, в синаптических щелях нейронов.
Именно сопротивление и пропускаемый ток окажутся ключевыми понятиями в модели нейрона. И манипуляции с этими понятиями нужно отработать в разных ситуациях, особенно в ситуациях сложения токов при взаимном влиянии многих источников: суммировании воздействия токов, как это происходит на мембране нейрона.
Любой источник тока обладает каким-то сопротивлением и не позволяет выпустить неограниченный ток. В схемотехнике это называется выходным сопротивлением, и оно должно учитываться в прикидках возможностей тока в цепи. Любая электродеталь, на которую воздействует ток, тоже имеет свое реальное сопротивление и не способна поглотить любой ток и это называется входным сопротивлением.
Идеально было бы, чтобы источники электротока имели бы нулевое выходное сопротивление и могли бы запитать сколь угодно потребителей тока, а электродетали – бесконечно большое входное сопротивление и можно было бы применить сколь угодно деталей, не истощающих источник. Конструкторы к этому стремятся, но в реальной жизни так не бывает.
Условно ситуацию можно изобразить так:
![]()
Слева - источник разности потенциалов, напряжением 12 вольт каждый выход которого имеет сопротивление 100 ом. Справа - электродеталь со входным сопротивлением 400 ом. Ток потечет из + к – через все сопротивления и его значение в общей цепи будет I=U0/(100+400+100) = 0,02 ампера. Понятно, что ток будет один и тот же, как если бы мы имели на трубе несколько по-разному открытых кранов, струйка на выходе получится в зависимости от состояния всех. Достаточно совсем закрыть любой и тока вообще не станет.
Прикинем, какие напряжения будут на входах детали.
Цепочка последовательных сопротивлений образует т.н. “делитель напряжения”, и в каждой точке напряжение окажется пропорциональным подключенным сопротивлениям. На концах первого сопротивления окажется 0,02*100=2 вольта. Это называется “падением напряжения” на сопротивлении (просто без напряга привыкайте к таким словечкам, чтобы если попадется -знать, откуда они растут). На двух сопротивлениях R1+R3 падение будет уже 0,02*500=10 вольт. Теперь ясно как переопределяются напряжения в цепи: U1 будет 10 вольт (12-2), U2=2 вольта.
Нагрузка запитается не 12-ю вольтами, а 0,02*400=8 вольтами.
Если вольтметр на нагрузке показывает напряжение, меньшее, чем на клеммах источника, способного давать заведомо больший ток, чем 0,02 ампера, то, значит, или провода имеют сопротивление или еще что-то мешает току (например, плохой контакт на клемме с источником).
В случае с нейроном возникает та же картина перераспределения токов. Нейрон – реальное электроустройство со своим входным сопротивлением, к которому прикладываются несколько (до десятков тысяч) источников тока (это – утрировано, но в целом верно). И они прикладываются не последовательно, а сразу все вместе к одному нейрону, что выглядит так:
![]() Те специалисты, кто имеет хорошие представления о реальных нейронах, в этот момент будут испытывать сильный протест от такого вульгарного отношения к такому чрезвычайно сложному образованию как нейрон при том, что приводимые схемы никак не противоречат этой сложности, а рассматривают ее определенную часть с позиции схемотехники. Я предлагаю волевым усилием преодолеть неприятие и просто посмотреть, что будет дальше. В результате самыми тщательными рассуждениями на основе воспроизводимых экспериментальных данных мы придем к наиболее общей и универсальной функциональности нейрона, что позволит в его модели освободиться от множества сложностей природной реализации. И эта модель покажет свою дееспособность во всех последующих рассмотрениях механизмов нейросетевой функциональности.
Те специалисты, кто имеет хорошие представления о реальных нейронах, в этот момент будут испытывать сильный протест от такого вульгарного отношения к такому чрезвычайно сложному образованию как нейрон при том, что приводимые схемы никак не противоречат этой сложности, а рассматривают ее определенную часть с позиции схемотехники. Я предлагаю волевым усилием преодолеть неприятие и просто посмотреть, что будет дальше. В результате самыми тщательными рассуждениями на основе воспроизводимых экспериментальных данных мы придем к наиболее общей и универсальной функциональности нейрона, что позволит в его модели освободиться от множества сложностей природной реализации. И эта модель покажет свою дееспособность во всех последующих рассмотрениях механизмов нейросетевой функциональности.
На электрической схеме все источники тока имеют потенциалы относительно “земли” – некоего условного “нулевого” потенциала для общего отсчета всех напряжений относительно него, который на схеме обозначен жирной черточкой в самом низу слева. Заметим, что самый нижний источник имеет относительно земли отрицательный потенциал.
Все входные сопротивления соединены в одной точке, где нужно прикинуть результирующее напряжение U0. Это сделать очень просто, если заметить, что сопротивления R1 и R2 положительному току в два раза сильнее ограничивают токи этих источников, чем R3. Если не учитывать подключенный R0, то очевидно, что в точке U0 положительные токи будут в сумме равны отрицательному току и в U0 все токи скомпенсируют друг друга: весь ток потечет к отрицательному источнику, и мы можем спокойно посчитать, что будто бы есть один положительный источник с общим сопротивлением 100 ом потому, что два параллельно подключенных сопротивления работают совместно, как два приоткрытых крана в одной трубе.
Если два крана открыты одинаково, то через них потечет в два раза больший ток.
(формулу расчета для случая разных величин сопротивлений мы не будем нигде применять, но если интересно, то она вот: 1/r0=1/r1+i/r2+...).
Понятно, что подключение устройства к нулевому потенциалу так его и оставит нулевым, каким бы ни было R0.
Как уже говорилось, к нейрону могут быть подключено до 10000 разных источников поляризующего мембрану тока с самыми разными сопротивлениями этому току. На входном сопротивлении нейрона будет меняться потенциал в зависимости от того, какой вес положительных или отрицательных зарядов будет преобладать. И если начинают преобладать те, что вызывают срабатывание нейрона, то его мембрана при этом становится проводящей, и приложенный потенциал скачкообразно обнуляется. Нейрон срабатывает, когда “возбуждающий” потенциал превысит порог его “терпения”, после чего начинается опять процесс накопления зарядов на его мембране до нового срабатывания.
Условно говоря, положительным потенциалом будем считать то, что приводит к срабатыванию нейрона, а отрицательным – к его торможению.
Использование схемотехнических механизмов
Устройство, которое срабатывает, переключаясь из одного состояния в другое, при превышении потенциала (любого вида) на его входе заданного порога, называется компаратором. Меняя порог срабатывания, можно менять условия срабатывания, например, если потенциал на входе возрастает постепенно, накапливаясь, то от порога срабатывания зависит время, через которое он сработает: или сразу при мизерном потенциале или дольше, если задан высокий порог.
У каждого человека есть свой “болевой порог”. Если сигналы его превысят, мы воспринимаем это как боль. Люди с низким болевым порогом испытывают боль даже от незначительных повреждений.
В реальном нейроне порог зависит от температуры и от концентрации ионов в среде вокруг нейрона, на что влияют примеси разных ионов (параметры эдс концентрационного элемента). Чтобы максимально сделать устойчивым порог срабатывания, среда нейрона ограждается по возможности от всего, что может повлиять из кровотока и это называется гематоэнцефалическим барьером. Иначе после фруктового сока у нас бы поехала крыша. Так как переносчиками зарядов к телу нейрона являются нейропептиды с нейромедиаторами в щелях контактов между телом нейрона и разными входами (синаптических щелях), то важно предохранять среду нейронов от всего того, что может повлиять на такой перенос. В том числе нужно стабилизировать температуру, чтобы от перегрева не возникал бред, т.к. порог в мембранных концентрационных элементах при повышении температуры уменьшается из-за повышения термодинамической активности носителей.
Если бы мозги были сделаны из транзисторов, то этой проблемы бы не было, как и очень многих других в природной реализации. Поэтому для понимания сути происходящего в мозге важно понять именно то, что оказывается важным для достижения цели того или иного механизма, а не то, что вынуждает строить конструкции для реализации механизмов. Из огромного количества доступных материалов исследования работы механизмов мозга, начиная от механизма работы нейрона, будем выделять самое главное, что в итоге даст общую картину причин и следствий происходящего вплоть до описания психических явлений.
И тогда нейрон это, однозначно – компаратор, сравнивающий сигналы токов, сглаживающихся на мембране, с различных входов со своим порогом срабатывания (но чуть сложнее, что будет вскоре ясно). А вместе со всеми входами (дендритами с синапсами щелей между входом и телом нейрона) он выполняет функцию распознавания того сочетания входных токов, которое заставляет его срабатывать, и более – ничего. Все остальное возникает из этой функции. Это похоже на схему устройства, называемую персептроном, но только отчасти.
Первым персептронную функцию нейрона предложил Ф.Розенблатт. Но он слишком идеализировал это устройство, сделав его намного точнее и универсальнее, чем сами нейроны, и это направление привело к созданию искусственных нейросетей, которые сегодня уж ничем не похожи на природные нейросети и обладают намного большими возможностями при намного больших невозможностях из-за такой идеальности, что будет подробно рассмотрено.
Функциональную модель нейрона мы будем еще уточнять, а пока что нужно остановиться и на других схемотехнических устройствах и схемотехнических принципах, которые проявляются в составе нейросети.
Если просто подать на вход компаратора потенциал выше его порога, то он мгновенно сработает, сменив потенциал на выходе. Но если на входе будет стоять накопитель зарядов, в который будет заливаться ток, то потенциал будет нарастать постепенно от нуля, как это происходит с заливанием в бак воды из крана. Чем больше емкость бака, тем медленнее он будет наполняться до уровня дырки стока. Чем больше ток воды из крана, тем бак наполнится быстрее.
Есть немало видов таких накопителей в электротехнике. Аккумулятор медленно заряжается оптимальной для него величиной тока. Конденсатор же способен заряжаться очень большими токами быстро, и в продаже есть для автомобилистов конденсаторы очень большой емкости, которые могут быть мгновенно заряжены, чтобы потом передать свой заряд аккумулятору или даже запитать стартер.
В природной нейросети нейрон так же накапливает заряды на мембране, получая потенциал не мгновенно, хотя и очень быстро. На разных участках мембраны могут оказываться разные потенциалы и тогда участок с превышением порога может начать срабатывать, но более поздние успеют или не успевают вернуть процесс. Это озадачивает исследователей, заставляя задуматься о том, что, может быть, это – тоже какой-то важный информационный механизм, а не просто неидеальность схемы.
Но в организме очень много неидеальностей, некоторые из которых оказывается полезными и передаются из поколения в поколение, хотя большинство просто никак ни на что не влияют. Это - как если бы иголка для шиться подвернулась еще для ковыряния в зубах.
Если бы на нейрон через его отростки-входы с проводящими синапсами начали бы поступать непрерывно высокие потенциалы, то они постоянно бы заряжали его мембрану до порога срабатывания. Нейрон бы разряжался импульсом своего тока до нуля потенциала на мембране, но входные токи продолжали бы ее заряжать и процесс генерации нейроном импульсов не прекращался бы пока сам нейрон не истощился бы из-за недостатка поступления питательных веществ и отвода продуктов его работы - до полной невозможности более реагировать. Что реально и происходит в таких случаях за какое-то время (слишком интенсивная работа групп нейронов приводит не только к истощению, но и гибели нейронов, что случается во время психозов). В мозге есть приемы избавления от такой нагрузки, приводящие ко многим интересным явлениям.
Мы могли бы наглядно смоделировать работу нейрона вот так:
![]()
Здесь C – конденсатор, накапливающий заряд, N– компаратор, выходной сигнал которого вызывает срабатывание ключа K, который замыкает обкладки конденсатора, разряжая его до нуля, после чего компаратор опять перестает выдавать сигнал и ключ размыкается (роль конденсатора и ключа одновременно выполняет мембрана, мгновенно становящаяся прозрачной для ионов и разность потенциалов на ней тут же компенсируется). На выходе компаратора получаются прямоугольные импульсы напряжения с частотой заряда конденсатора до порога срабатывания компаратора.
На конденсаторе получается пилообразное нарастание напряжения с некоторым временем спада до нуля, за которое на выходе компаратора возникают положительные прямоугольные импульсы.
Вот анимация fornit.ru/an-book-14, показывающая наглядно, как срабатывает нейрон.
Пачка импульсов активного нейрона, приложенная к клетке мышечного волокна, способна сократить размеры этой клетки, - она съеживается под воздействием этого тока.
Кстати, вот еще пример того, что частота импульсов кодирует время воздействия стимула, а не его силу: пока есть импульсы, мышечное волокно сжато до момента истощения. На этом функция мышечной клетки в организации работы мышцы и заканчивается.
По сравнению с мышечной клеткой, клетка нейрона так же возбуждается от входного тока, но не сокращается, а генерирует импульсы. И на этом исчерпывается функция тела нейрона, если не говорить о всяких побочных вещах, не касающихся основной функции в нейросети – передачи возбуждения при определенном пороге заряда на своем теле. Но из-за того, что с мозгом связано очень много разных и запутанных явлений, многие исследователи пытаются наделять отдельные нейроны непомерно сложными свойствами, вплоть до собственных целей.
Постепенно мы с очевидностью увидим, что все многообразие явлений распознающей и управляющей функции мозга основывается только на одной указанной функции нейрона без необходимости привлечения чего-то еще. При этом из нейронов организуются многие схемотехнические устройства, реализующие определенные схемотехнические принципы.
Дополнительно стоит заметить, что в живом теле разные клетки приспособились к различным строго выраженным специализациям, и все они развиваются из “стволовых”, неспециализированных клеток, которые формируют свою специализацию в зависимости от окружающих условий (такая зависимость начального наследственного кода от условий описана в fornit.ru/806).
Клетки различных рецепторов способны реагировать на какие-то воздействия, выдавая, как нейроны, импульсы тока сами или через посредников (как это происходит в случае фоторецепторов) и в простейших организмах, где нет нейронов, прямо от них сокращаются мышечные клетки или выделяются нужные вещества. Таких видов рецепторов много: клетки, реагирующие на фотоны света, клетки, активирующиеся от разных видов химических веществ, чувствительные к механическим колебаниям специализируются как рецепторы звуковых колебаний, как рецепторы прикосновений, как рецепторы сокращения мышц и т.д. Нет каких-то хитрых скрытых других функций кроме тех, что явно проистекают из специализации.
Вот и клетки нейронов, как и клетки рецепторов в результате своей функции выдают электрические импульсы в ответ на раздражение, только раздражителями для них являются заряды, возникающие на синапсах, покрывающих тело нейрона. А эти заряды появляются в ответ на электрические токи предшественников: рецепторов или таких же нейронов.
Вот почему все нейроны – такие рецепторы, функцией которых является реагирование на состояние предшествующих им клеток. Так же все нейроны воздействуют на последующие эффекторы – тоже нейроны или уже мышечные (или продуцирующие гормоны) клетки. Все ограничивается главной функциональностью нейросетей мозга – управление, а не еще что-то. Для целей формирования управляющих структур больше ничего не требуется.
Получается, что все слои связанных в сеть нейронов – эффекторы по отношению к предыдущим и, в то же время, они же – рецепторы для последующих, это – условно нами выделяемые роли в нейросети.
Рассмотрим еще одно схемотехническое понятие – “коэффициент передачи” тока (или напряжения) – ключевое понятие в персептронах. И сделаем это на примере транзистора, реализующего “коэффициент усиления”. Коэффициент передачи – множитель, на который нужно умножить входное напряжение (или ток), на входе устройства, чтобы получить величину напряжения (или тока) на его выходе.
Что нейроны, что электродетали в схемах управления – в принципе равнозначны для целей управления, а транзисторы – очень удобная аналогия для понимания свойств электродеталей.
В самом общем виде транзистор – электродеталь, функцией которой является управление током в цепи. Он может просто как ключ включать и выключать весь протекающий через него ток, и тогда этот режим так и называется – ключевой. Но транзистор может как кран дозировать ток, своим изменяющимся сопротивлением, поддерживая его в заданном значении, для чего и используется тот “коэффициент передачи”, часто в ипостаси “коэффициента усиления”. Заодно будет рассмотрен принцип обратной связи нескольких реализаций. А обратные связи очень широко используются в природных нейросетях.
![]()
Транзистор может обозначаться на схемах очень по-разному в зависимости от его типа, и мы рассмотрим только некую его обобщенную модель с определенными свойствами. У транзистора есть три конца (вывода, электрода): Э – эмиттер – куда втекает ток его цепи, К – коллектор, откуда вытекает модулированный транзистором ток и Б – база – управляющий электрод (у “полевого” транзистора это – исток, сток и затвор, делать нечего, но захотели назвать по-другому).
Но, повторюсь, нам неважно, какой именно это транзистор. Важно то, что, изменяя напряжение на его базе, мы сможем заставить транзистор открываться и закрываться для тока между эмиттером и коллектором. У каждого транзистора есть свое значение такого напряжения на базе, когда ток через транзистор не протекает и он заперт, это называется “напряжение отсечки”. Начиная с этого напряжения на базе, начинается протекание тока через транзистор до такого максимума, выше которого сопротивление транзистора уже не позволит протекать больше или он просто сгорит.
Чтобы выгорания от большого тока не случилось, поставим в цепи его эмиттера, ограничивающие сопротивление:
![]()
Транзистор нужно рассматривать как изменяющееся сопротивление в цепи тока, зависящее от напряжения на базе (даже у биполярных транзисторов есть такая вольтамперная характеристика). Т.е. в любой момент к нему тоже можно применять закон Ома.
При постоянном напряжении на базе Uб (минус напряжение отсечки) через транзистор будет протекать ограниченный ток, независимо от напряжения на коллекторе (пока не выдержит транзистор) и независимо от коэффициента передачи транзистора по току. Этот ток определяется только сопротивлением в эмиттере, и вот почему.
Если на базу подать напряжение, выше отсечки, транзистор приоткроется и на его эмиттере тоже появится напряжение, но не выше, чем напряжение на базе - минус отсечка. Ведь если бы на эмиттере оказалось выше, то транзистор бы закрылся. Он вынужден оставаться на таком промежуточно открытом состоянии, при котором он открыт ровно настолько, чтобы не начать закрываться из-за уменьшения напряжения меньше, чем отсечка.
Чтобы было проще прочувствовать ситуацию, предположим, что у этого транзистора напряжение отсечки равно нулю, т.е. он начинает открываться, как только напряжение на базе начинает превышать нуль. Теперь логику обратной связи, заставляющей поддерживать на эмиттере то же самое напряжение, легко можно представить.
![]() Подключим резистор между коллектором и + источником питания.
Подключим резистор между коллектором и + источником питания.
Это – все тот же генератор тока. Ток, определяемый сопротивлением Rэ и напряжением Uб, потечет через резистор Rк. Напряжение на коллекторе будет определяться величиной резистора Rк, просто образуя это напряжение как падение на резисторе Rк при протекающем токе через него (если только величина этого сопротивления не настолько велика, что он начинает сам ограничивать этот ток, делая его меньше, чем стабилизируемый транзистором).
Если мы изменим напряжение на базе на небольшую величину Dб, то ток через цепь изменится на Dб/Rэ, а напряжение на Rк изменится на Dk=Rк* Dб/Rэ. Отношение Dk/Dб называется коэффициентом усиления К (по напряжению) данной транзисторной схемы:
К= Rк* Dб/Rэ / Dб или, сокращая Dб, К= Rк* Dб/Rэ / Dб = Rк/Rэ – т.е. зависит только от соотношения двух резисторов. Если Rк в 2 раза больше, чем Rэ, то эта схема будет усиливать все изменения напряжения на входе в 2 раза.
Интуитивно это воспринимается достаточно просто, если уже понято: на бОльшем сопротивлении данный ток вызывает бОльшее падение напряжения (по закону Ома: чтобы через сопротивление протекал данный ток, на его концах должно быть определенное напряжение) и, значит, просто меняя сопротивление при данном токе, мы получаем разные падения напряжения на нем, и поэтому при изменении тока в цепи изменением напряжения на входе, вызывается гораздо большее изменение напряжения на резисторе коллектора. Но это изменение нарастает в сторону, противоположную изменению напряжения на базе: если на базе напряжение уменьшается, то и ток уменьшится и падение на резисторе коллектора уменьшится: оно же отнимается от напряжения источника питания, т.е. напряжение Uк при этом возрастет. Когда на базе напряжение уменьшается, на коллекторе, наоборот, – возрастает. Говорят, что такой усилитель инвертирует сигнал.
Все это намеренно дается, чтобы как можно больше и по-разному проговорить и понять, закрепив модель понимания в голове до автоматизма, не требующего напряжения мысли. Поэтому нужно придавать как можно большую заинтересованность моделирующей игре с такими вещами, по-разному прокручивая ситуации. Тут мотивация – научиться без проблем моделировать схемотехнику нейронных цепей. Это – не просто как в школе учить непонятно зачем.
В отличие от профи-схемотехников, мы будем иметь дело всего с одной деталью – упрощенным персептроном (но со многими схемотехническими принципами), поэтому все второстепенное предельно упрощается.
Вот более распространенная схема получения заданного коэффициента усиления:
![]()
В этой схеме мы сделали уже понимаемым способом коэффициент усиления: 1000/10=100. Каскад инвертирует изменение сигнала и поэтому через R2 к базе прикладывается напряжение, препятствующие изменению входного U1.
Учитывая, что К каскада усиления без учета R2 и R1 очень большой (=100), то даже незначительное уменьшение U1 привело бы появлению практически - напряжения питания на Uк. И резисторы R2 с R1 поделили бы его между U1 и плюсом питания.
Если бы R2 было равно R1, то входное U1 делилось бы пополам, т.е. изменение напряжения на базе стало бы в 2 раза меньше. Это дернуло бы напряжение на базе вверх и, значит, открыло бы транзистор, увеличив ток, но повышенный ток повышал бы и Uк. Чувствуете действие обратной связи через R2? Процесс бы сбалансировался в некоем компромиссе, который можно прикинуть: напряжение Uк стало бы таким, чтобы на базе с учетом U1 возник приоткрывающий транзистор потенциал, обеспечивающий ток в цепи эмиттера и коллектора, вызывающий падение напряжения на Rк, равное Uк.
Чуть отступая, но это важно на будущее: в реальных схемах такая балансировка возникает не мгновенно, а с некоторой задержкой, и поэтому возникают колебания перекомпенсации в ту или иную сторону, которые быстро затухают. Это называется переходным процессом, и он порождает паразитные явления в выходном сигнале. Такие переходные процессы обнаруживаются при работе фоторецептора при резкой подаче света на него.
Если задержка выходного сигнала от входного настолько высока, что сигнал на входе успевает измениться на противоположный, то вместо отрицательной обратной связи получается положительная, и переходный процесс не утихает, а, наоборот, увеличивает разбалансировку. На выходе возникает уже независимая от входа генерация.
Вот такие переходные процессы будут рассматриваться как следствие работы нейросети.
Намеренно соотношение Rк и Rэ сделано достаточно большим, а значение Rk на порядок меньше, чем R2, чтобы можно было не учитывать Rк и Rэ в расчете. С достаточной точностью можно посчитать, что у нас есть только R1, R2 и Rk, образующий делитель напряжений между резисторами, в котором напряжение на базе примерно равно напряжению отсечки (чтобы транзистор открывался). Тогда при изменении напряжения U1 на D1 потенциал Uк станет таким, чтобы на базе сохранилось напряжение отсечки. Это означает, что при увеличении U1 на D1 Uк должно увеличиться в R2/R1 раз, с учетом погрешности отбора других резисторов – в 10 раз. В этой схеме самое показательное то, что на базе поддерживается напряжение отсечки, ведь если оно окажется хоть немного выше, то при К=100 Uк упадет до нуля (транзистор закроется) и наоборот, стоит чуть уменьшиться и Uк Увеличится в 100 раз до + питания.
Пока транзистор находится в приоткрытом режиме (Uк не доходит ни до нуля, ни до + питания), напряжение на базе будет всегда оставаться около значения напряжения отсечки (в идеальном случае бесконечного коэффициента передачи устройства – точно равно напряжению отсечки).
Если вместо неидеального транзистора применить специальную схему идеального усилителя с бесконечным К усиления и нулевым напряжением отсечки, то на его входе при организации обратной связи через R2 и входного резистора R1 всегда бы поддерживался строго ноль, пока усилитель находится в рабочем режиме, а не ключевом. К усиления такого каскада был бы строго R2/R1 – если у источника сигнала нулевое выходное сопротивление, а у усилителя бесконечно большое входное, иначе эти сопротивления нужно было бы учесть в формуле K.
![]()
Здесь на входе усилителя строго поддерживается ноль, его называют еще виртуальный ноль потому, что в этой точке еще и нулевое входное сопротивление. Это значит, что сколько бы ни было у нас входных резисторов R1 каждый со своим сигналом, в точке нуля они все идеально суммируются по току, определяемому сопротивлением и разностью напряжения сигнала и нуля входа. При этом у каждого такого входа – свой коэффициент усиления, а все получившиеся сигналы очень точно смешиваются, суммируясь в одном выходном сигнале. Это – наилучший способ организовать суммирующую функцию модели нейрона.
Но даже простой транзистор мог бы хорошо выполнять роль компаратора в нейросетях потому, что в них не требуется прецизионной точности, правда, лучше бы стабилизировать температуру, чтобы порог срабатывания не гулял из-за изменения напряжения отсечки. Чтобы задать порог срабатывания, было бы достаточно задавать напряжение на его эмиттере и тогда транзистор бы срабатывал относительно него. Можно нарисовать такую модель нейрона на транзисторе:
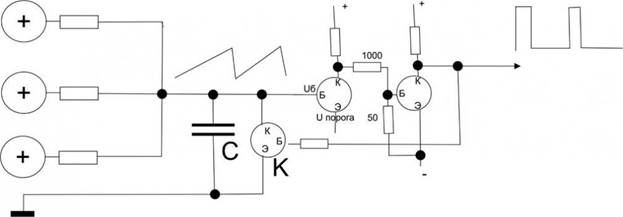
Здесь оба транзистора работают в режиме ключа, один сравнивает напряжение на конденсаторе с напряжением порога своего эмиттера и когда оно превышает его, открывается, закрывая следующий транзистор, и тогда + выходное напряжение открывает ключ K, разряжающий конденсатор.
Или же использовать компактный и идеальный усилитель:
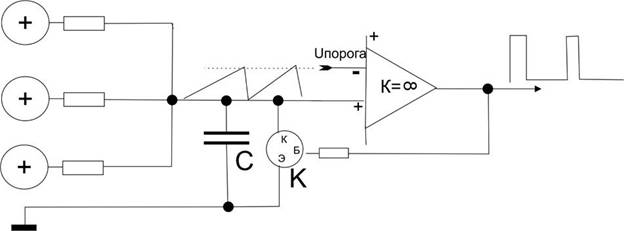
Входы усилителя помечены + как неинвертирующий и – инвертирующий, т.е. относительно инвертирующего входа и сравнивается потенциал на неинвертирующем входе, при превышении которого на выходе возникает высокое напряжение импульса на время разрядки конденсатора до порога.
В обоих случаях суммирующим элементов является конденсатор С, который работает как емкость, постепенно заливаемая всеми источниками тока, пока уровень потенциала в емкости не превысит порог срабатывания.
Чтобы импульсы не были слишком короткими, нужно задать гистерезис: сделать так, чтобы напряжение срабатывания было больше напряжения отпускания и тогда компаратор будет ждать, пока конденсатор не разрядится более сильно перед тем, как снова отключится. Этот гистерезис можно задать положительной обратной связью с выхода компаратора на его + вход:
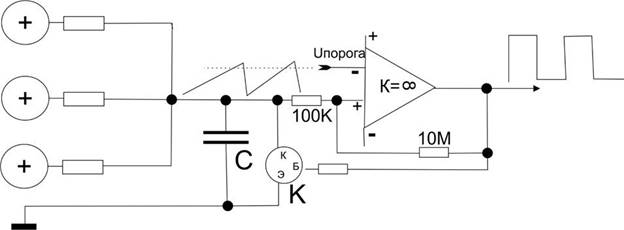
А в мозгу все это – одна клетка нейрона с 10000! входных “резисторов”, по каждому из которых может приходить сигнал возбуждения (+) или торможения (-) или ничего не приходит. Задача этой клетки – срабатывать только при определенном сочетании весов активирующих и тормозящих сигналов на ее входе, т.е. – распознавать такое сочетание как рецептор этого состояния. В качестве резисторов служат синапсы, которые проводят или нет в какой-то степени, что называется весом сигнала по каждому входу. В зависимости от проводимости (веса, коэффициента передачи синапса) к телу нейрона приходит только какая-то часть входного сигнала или вообще не может пройти, как через изолятор.
Дополнительное пояснение работы схемы компаратора на транзисторе в модели нейрона
Если бы не было нейрона, то конденсатор просто зарядился бы до максимума +питания и так бы остался с высоким напряжением на нем. Но нейрон разряжает его всякий раз, как только потенциал на конденсаторе превысит его порог срабатывания.
Если использовать идеальный компаратор (есть такие микросхемы), то схема будет выглядеть так:
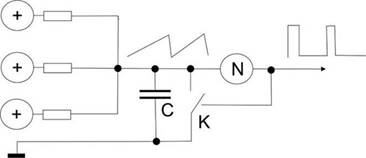
Идеальный компаратор никак не вмешивается своими входами в работу схемы потому, что входное сопротивление у него бесконечно большое. Т.е. он просто следит за потенциалами на его входах: опорным напряжением U порога, сравнивая его с нарастающим напряжением на конденсаторе. Конденсатор совершенно свободно заряжается, не подозревая, что его измеряют.
Если опорное напряжение будет 5 вольт, то компаратор сработает, как только напряжение на конденсаторе достигнет 5 вольт.
Входы усилителя помечены + как неинвертирующий и – инвертирующий, т.е. относительно – входа и сравнивается потенциал на + входе, при превышении которого на выходе возникает высокое напряжение импульса на время разрядки конденсатора до порога.
Неинвертирующий вход – такой, что сигнал превышения порога (на выходе) будет иметь ту же полярность, что и сигнал на входе: на входе он увеличивался до порога, значит на выходе он станет + (нарисованы импульсы положительного напряжения).
Т.к. компаратор никак не влияет на зарядку конденсатора, нам нужно сделать так, чтобы при срабатывании компаратора разрядился конденсатор. Для этого мы ставим разрядный ключ на транзисторе К. Пока транзистор закрыт, его все равно что нет, он не мешает заряжаться конденсатору. Но стоит его открыть, как он превращается в проводник, как гаечный ключ, брошенный на клеммы аккумулятора.
Транзистор открывается положительным потенциалом на своем входе, превышающим его порог открывания (напряжение отсечки). Мы поставили резистор, ограничивающий ток базы, просто чтобы не повредить транзистор (больше нет задачи у этого резистора).
Положительные импульсы с выхода компаратора, как только они появляются, превышают напряжение открывания транзистора К (напряжение отсечки), транзистор открывается, становясь проводником, и разряжает конденсатор. После этого компаратор опять на выходе имеет низкий уровень сигнала, который закрывает транзистор, и конденсатор опять начинает заряжаться.
Эта причинная логика достаточно ясна, она следует из того, как идет процесс зарядки конденсатора и его принудительной разрядки.
Собственно компаратор может быть сделан всего на одном транзисторе. Он не идеальный, но в принципе пойдет. Опорное напряжение подаем на его эмиттер, пусть будут те же +5 вольт.
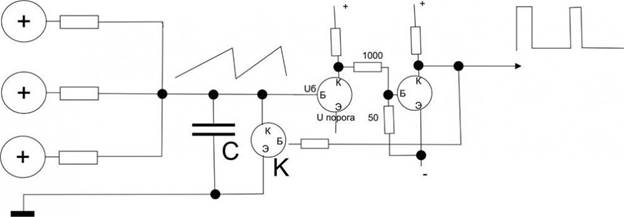
Закрытый транзистор тоже не влияет на заряд конденсатора и сработает, как только будет достигнуто напряжение открывания. И тогда на выходе такого каскада возникнет низкий потенциал потому, что ставший проводящим транзистор подключит к нему источник питания в его эмиттере.
Получается, что на выходе транзистора появляется минус, когда на входе возрос плюс, т.е. он действует как инвертор. Но ключ-то открывается плюсом. Значит, нам нужно поставить еще один инвертор перед ключом, чтобы там появлялся плюс - когда нужно разрядить конденсатор.
Здесь оба транзистора работают в режиме ключа, один сравнивает напряжение на конденсаторе с напряжением порога своего эмиттера и когда оно превышает его, открывается, закрывая следующий транзистор и тогда + выходное напряжение открывает ключ K, разряжающий конденсатор.
Кроме того, дополнительный транзистор выполняет еще одну функцию. Ключ должен быть закрыт в начале процесса заряда, но напряжение на выходе первого транзистора меняется (при его переключении) от +5 до + питания, а +5 вольт – тоже открывающее напряжение для К, который закрываются при меньшем, чем 0,5 вольта потенциале. Второй транзистор при переключении меняет потенциал на выходе (на своем резисторе коллектора) от 0 до + питания, что и требуется.
Резисторы 1000 и 50 ом обеспечивают рабочие напряжение включения-выключения на базе второго транзистора.
Для неискушенного в схемотехнике возникает вопрос: “В схеме непонятно, куда и как движется ток и какие претерпевает при этом изменения”.
Начнем с того, что транзисторы по своему входу почти не потребляют тока (а есть такие, что вообще не потребляют), т.е. у них настолько высокое входное сопротивление в закрытом состоянии, что они спокойно позволяют заряжаться конденсатору, не влияя на это. Так же никак не влияет и закрытый транзистор (К), подключенный к конденсатору коллектором, чтобы в нужный момент открыться и разрядить его. Можно сказать, что закрытого транзистора как бы и нет в схеме и рассматривать потенциалы без его учета.
Тогда мы получаем просто заряд конденсатора через резисторы от источников + напряжения.
(Время, за которое конденсатор зарядится, прикидывается по формуле T=R*C. В данном случае мы имеем 3 штуки R, которые заряжают конденсатор до напряжения 0.7 от + источника, и если + одинаковые и R одинаковые, то заряжают в 3 раза быстрее.)
Закрытый первый транзистор означает, что на резисторе в его коллекторе будет + напряжения питания, который приложится к базе второго транзистора через делитель напряжения 1000 и 50, который поделит напряжение на базе в 20 раз. Это значит, что второй транзистор будет открыт, если напряжение питания равно 10 или больше вольт при пороге его открывания (Uотсечки) 0,5 вольта.
Конденсатор будет заряжаться не до напряжения + входных резисторов, а пока не будет достигнут открывающий первый транзистор потенциал, т.е. до Uпорога (пусть будет +5 вольт) + Uотсечки транзистора (Uотсечки небольшое, например, = 0,5 вольта). После этого первый транзистор резко откроется (резко потому, что нет никакой обратной связи, чтобы сбалансировать открывающий потенциал), т.к. он сработает как ключ.
Когда первый транзистор откроется, он станет просто проводником и к его коллекторному резистору приложится Uпорога, например +5 вольт. При +5 вольтах на конце делителя, на базе второго транзистора станет 5/20=0,25 вольт, что меньше напряжения отсечки и второй транзистор закроется, т.е. он как бы перестает существовать в схеме. На его коллекторном резисторе станет напряжение питания, которое через резистор приложится к ключу К, резко его открыв. Конденсатор разрядится, тем самым закрыв первый транзистор, после чего ключ отпустит его и конденсатор опять начнет заряжаться.
Можно заметить, что транзисторы в таком включении инвертируют сигнал на своих входах: если на входе низкий сигнал, то на выходе – высокий и наоборот.
Мы имеем отрицательную обратную связь через замкнутый круг каскадов (транзисторных модулей). Каждый из каскадов инвертирует сигнал так, что данное число каскадов приводит к инвертированному отклику в точке замыкания кольца связей. Если бы не было задержки во всей этой цепи, эта связь бы привела к некоему равновесному состоянию, несмотря на общей очень высокий К усиления (у каждого “обычного” транзистора реальный К бывает не ниже 50, а общий == 50*50*50 == 125000). Никакого заряда конденсатора на входе не было бы, он зависнет на каком-то напряжении, близком к Uпорога+U отсечки первого транзистора. Но задержка есть и при таком большом К усиления за ее время ключ К успеет сильно разрядить конденсатор. Время такой задержки и определит очень короткую ширину импульса. Поэтому в последней схеме мы вводим гистерезис порога срабатывания первого транзистора.
Стоит не забывать, что транзистор может работать как ключ (или открываться или закрываться) и в линейном режиме, когда его сопротивление принимает некое промежуточное состояние. Просто подбирая потенциал на входе, заставить работать транзистор линейно практически невозможно, граница переключения очень резка из-за высокого коэффициента усиления транзистора. Линейный режим устанавливают с помощью обратной связи, как это описывалось в самом начале лекции, например:
![]()
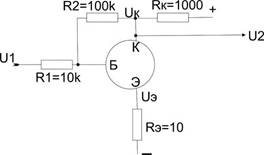
Принцип персептрона
Сначала определим фундаментально важные функции рецепторов и эффекторов, прочувствовав то, что именно в них принципиально и почему.
Рецепторы
Рецепторами будем называть преобразователи какого-то вида элементарного воздействия в сигнал типа: “есть” или “нет”. Такое переопределение может показаться странным для нейрофизиологов, ведь в нейронной сети имеются и т.н. «вторичночувствующие» рецепторы, генерирующие только рецепторные потенциалы (ТПСП, ВПСП) (фоторецепторы, наружные волосковые клетки улитки внутреннего уха). Так что это определение – схемотехнический термин для данной книги: это не только рецепторы воздействий среды, но и нейроны, активность которых влияет на последующие нейроны или клетки других видов. Соответственно, все то, что воспринимает сигнал, будем называть эффекторами. Нейрон в одной ипостаси является рецептором, а в другой – эффектором. Такая терминология более системна.
Для целей распознавания важно именно элементарное воздействие, т.е. минимально возможное для данного механизма рецепторов. И тогда один рецептор может сообщать о наличии или отсутствии такого воздействия, а несколько рецепторов – о силе такого воздействия в информационном плане (а не в виде частоты генерации нейрона), т.е. о том, сколько элементарных воздействий было приложено к рецепторному полю (условному профилю активности элементов данного слоя). Вид выходного сигнала не принципиален: хоть импульсы, хоть постоянный потенциал, важно только, что есть такой сигнал на выходе или нет его. Но в природе используются импульсы: пока есть воздействие, идут импульсы (пока не истощится ресурс восстановления клетки после генерации импульса, если затраты на генерацию превысят поступление энергонесущих веществ в клетку). Уточним, что в первичных рецепторах, таких как On-Off-клетки сетчатки, частота импульсации хотя коррелирует с силой воздействующего фактора, но, как уже замечалось выше, эта корреляция не способна отразить динамический диапазон силы воздействия, а сильно лимитирована возможностями клетки, т.е. носит не информационный характер, а является локальным механизмом.
У природных рецепторов минимальное воздействие может оказываться равно элементарному действию всего нескольких фотонов или одной молекулы вещества, но это не принципиально. Просто у рецептора есть чувствительность, которая и определяет порог воздействия.
При таком определении рецептора можно заметить, что не только специализированные чувствительные клетки, но и любой нейрон оказывается рецептором по отношению к тому, что воспринимает его сигнал. Зная специализацию рецептора, всегда можно сказать, на что именно он реагирует, о чем он “информирует” последующий элемент.
Рассмотрим схему соединения разных рецепторов прямыми возбуждающими связями.
![]()
Представим, что есть три рецептора, специализированные на детекции наличия-отсутствия 1-звука, 2-света, 3-запаха.
За ними есть два рецептора 4 и 5, которые специализированы срабатывать, если к ним по имеющейся связи приходит хотя бы один из сигналов детекции. Так, 4 будет выдавать на выходе сигнал, если активен 1 или 2, а 5 сработает по активности 1 или 3.
Далее за ними есть слой рецепторов 6 и 7, специализированных аналогично имеющимися связям.
Понятно, что специализация любого рецептора зависит от существования связи того, что на него воздействует. Так, если закрыть глаза, то на рецепторы света не будут попадать никакие сигналы, а если оборвать связи со вторым слоем, то 4 и 5 перестанут выполнять свои функции распознавания (детектирования) активности на входе.
В схеме все связи - возбуждающие и каждая способна при активности превысить порог срабатывания подключенного к нему следующего рецептора. Такие сочетания пар (или большего числа) связей работают по логике “ИЛИ”: любая активность приведет к срабатыванию, или одна или другая или, тем более, обе.
![]()
Теперь представим, что у нас есть рецептор, который работает наоборот: при своем возбуждении он выдает сигнал, выключающий последующий элемент:
Тогда элемент 7 сможет сработать только в случае, если молчит элемент 4, иначе через тормозное влияние элемента Т 5 не сможет возбудить семерку. Это уже другая логика, исключающая логику ИЛИ для данной связи, так и называющаяся “исключающее ИЛИ” (т.к. ИЛИ в логике обозначается OR, то новая логика называется XOR).
Сам же тормозной элемент в логике управления можно было бы назвать “НЕ” или инвертором. Но в логике нейросетей этот элемент стоит особняком потому, что не просто инвертирует сигнал, а является детектором отсутствия активности 4, но с инвертированием такой активности. Если 4 неактивен, то Т не оказывает никакого влияния, а элемент “НЕ” оказывал бы, имея высокий уровень (инвертируя низкий уровень на входе). Т.е. в отличие от булевой логики в природной нейросети неактивные элементы как бы отсутствуют. Названия для такой логики нет, а она есть в природной нейросети.
В т.н. булевой логике (оперирующей условными логическими элементами), с которой хорошо знакомы все, кто работает с цифровыми схемотехническими устройствами и программисты, чтобы получить поведение Т, нужно сделать специальную схему.
Логику управления любым элементом или сочетанием элементов в нейросети можно назвать более общим словом “распознаватель”, и его логика оказывается намного гибче “обычной” булевой логики, позволяя реализовать многие возможности реагирования на сочетания звука, света и запаха. Т.е. распознаватель способен реализовать любую логику.
Мало того, можно еще более ее расширить, если добавить возможности связям быть не всегда превышающим порог, а придать им коэффициент передачи сигнала, меньше 1. И тогда рецептор сможет реагировать только при превышении порога общей силой сигнала нескольких рецепторов, скажем, света:
![]()
В данном случае между входными рецепторами и 4 включен элемент разрыва связи с коэффициентом передачи 0,4. Понятно, что теперь 4 сработает только в случае активности всех рецепторов света 1,2 и 3, что реализует логику “И”.
В нейросети такие коммутирующие элементы называют синапсами. Синапсы и в самом деле и по принципу действия похожи на щели между концами проводника, которые могут становиться в какой-то мере проводящими, ограничивая ток от нуля до максимума. Только нужно заметить, что коэффициент передачи реального синапса не является гибко переменным, а формируется какое-то его значение (хотя это значение может несколько “гулять” от разных внешних воздействий, искажая функцию распознавания). Как уже замечалось, у синапса не может быть большой дискретизации состояний проводимости из-за ограниченности стабильности воздействия окружающей среды. Пластический синапс Хеба может иметь не более 5-10 дискретов проводимости, но и это будет избыточным для реализации принципа природной специализации нейрона.
Эффекторы
Эффектором будем называть элемент, способный от электрического сигнала на его входе совершать некое действие. Так, мышечная клетка способна сокращаться, гормональная клетка продуцирует специфический для нее гормон, а нейрон – выдает электрический сигнал.
Повторим в который уже раз, что нейроны одновременно и рецепторы по отношению к последующим элементам, настроенные на детекцию определенного воздействия, и при этом эффекторы по отношению к предыдущим элементам, срабатывающие от этого воздействия.
Смысл именно в том, какую роль они выполняют относительно окружающих других клеток, соединенных с ним. Этот смысл реализуется связями, начинающимися от первичных рецепторов до конечных эффекторов, обеспечивая реакцию конечных эффекторов от состояния первичных рецепторов, которая определяется состоянием промежуточных нейронов – в этом и состоит управляющая функция мозга.
На самом раннем этапе эволюции схем управления мозга сигналы от первичных рецепторов непосредственно управляли клетками ответных действий, и это давало возможность реагировать на происходящее. Но появление промежуточных нейронов и синапсов придало схемам управления максимальную универсальность реагирования для гораздо большего числа ситуаций, чем это было бы возможно без промежуточных
нейронов, ограничиваясь лишь спецификой и чувствительностью первичных рецепторов и возможностями конечных эффекторов.
Можно декларировать математически строгое утверждение или теорему, что с помощью пороговых компараторов (нейронов) и синапсов, обладающих заданным коэффициентом передачи и еще одним из двух видов последействия: возбуждающего или тормозного, можно организовать любую схему адаптивного управления эффекторами от рецепторов, любой сложности. В том числе обладающую способностью самостоятельно изменять логику своей работы (но в природной нейросети все еще сложнее, например, есть еще и нейромедиаторы и разные специфичные к нейромедиаторам синапсы, что придает еще больше гибкости и возможностей).
Математическая логика способна дать определенный ответ на это утверждение, исходя из булевой логики. Схемотехники-профессионалы и программисты способны дать определенный ответ на это утверждение, исходя из любых практических задач.
Впервые такое утверждение сделал Розенблатт, чем вызвал повышенное внимание к суммирующим компараторам в сочетании с синапсами на входе, которые он назвал персептронами. Но скоро было показано, что сам по себе один такой элемент, все же, обладает ограничением на реализацию одной из базовых логических функций: “Исключающее ИЛИ” (схемотехническое обозначение “=1” - xOR). После этого интерес к персептрону начал стремительно угасать, в том числе и из-за того, что в деятельности нейрона, синапсов и других элементов мозга нашли множество “нелогических” особенностей, многие из которых напрямую влияли на выполнение управляющей функции мозга. Появлялись самые разные теории работы мозга и функции нейрона в нем, в том числе очень витиеватые, даже почти мистические теории из-за своей сакраментальной неопределенности используемых слов, зато достаточно привлекательные и академически импозантные. А Розенблатт не смог ничего поделать с этим просто потому, что вскоре трагически погиб.
Но энтузиасты идеи оставались, и они показали, что многослойный персептрон способен выполнять вообще любые логические операции и быть идеальным распознавателем всего, чего бы то ни было, лишь бы были рецепторы минимальных состояний этого. Это направление оказалось в жестком и непримиримом разрыве от исследователей-биологов, которые вообще мало были готовы воспринимать такие абстракции, считая их техническими изысками, далеки от реальности, и были правы в этом. Но ответвившееся направление искусственных нейросетей доказало, что да, нет никаких ограничений эффективности распознавателей при любой сложности объекта распознавания. Эмуляторы адаптивных нейронных сетей показывают, что с помощью простейших распознавателей (основной функции нейрона) можно создать управляющие схемы любой сложности: fornit.ru/41239.
Если попытаться найти, где же конкретно в природных нейросетевых схемах может использоваться элемент XOR (в схемотехнике он может использоваться как смеситель импульсов, как генератор коротких импульсов, для управления инвертированием импульсов, но на самом деле на практике их используют редко, а в программировании их широко используют для создания прозрачных изображений), то такового найти не удается просто потому, что в природе не используется логические операции с отдельными импульсами нейронов, ведь эти импульсы обезличиваются суммированием на мембранах.
Но в самом общем плане использование логики ИЛИ – типичная вещь: есть нейроны, которые срабатывают если хотя бы один из их входов оказывается активным. Это не точно функция ИЛИ, а некий суррогат ее по схожему смыслу действия механизма. Но этот механизм очень важен для того, чтобы распознать несколько разные, но схожие по какому-то признаку активности, например, для получения инвариантных абстракций (инвариантное - нечто неизменяемое в схожих абстракциях – т.е. условных отвлечений от реальных стимулов: выделений абстракций-примитивов линий, кругов, точек и т.п. независимо от наклона и масштаба).
Кстати, любой нейрон детектирует какую-то абстракцию, т.е. является элементом, выделяющим условное значение в воспринимаемом. Напоминание: о сути абстракций можно прочитать здесь: fornit.ru/103
Вот в таком, суррогатном смысле, есть еще одно применение “алгебры логики” в мозге: контрастирование профиля активностей на входе нейрона (см. fornit.ru/ax1-10-413, fornit.ru/ax1-10-414, fornit.ru/ax1-10-484), т.е. выделение более контрастной составляющей с подавлением фоновых шумов. Это очень напоминает использование XOR для получения прозрачности в изображениях. И при контрастировании, можно сказать, выполняется нечто, напоминающее XOR, когда общий массив приходящих сигналов подавляется, если они одинаковые и не выдающиеся среди других. Проделывается это не одним нейроном-персептроном, а с помощью вставочных тормозных нейронов, располагающихся между нейронами данного слоя распознавателей. И тогда активность какого-то нейрона вызывает подавление активности соседей так, что в конечном итоге активным остается только наиболее возбужденный, а остальные молчат. Возникает контраст, настолько просто, эффективно и полезно, что это становится характерным для всех слоев нейронов мозга: подавление боковых соседей, что биологи назвали латеральным торможением (fornit.ru/21746).
Зримо себе представить этот эффект можно с помощью демонстрационной модели fornit.ru/38103: там нужно выбрать образ (над левой матрицей) и начать сдвигать ползунок над правой.
Латеральное торможение – хорошо известное явление в нейросети мозга. Но ему уделяется неоправданно мало внимания и оно как бы даже забывается при рассмотрении различных проявлений реакций организма. А ведь это – базовый, основополагающий принцип в организации нейросети. Совсем недавно вышла статья (fornit.ru/ext-1): “Что заставляет чесать, где чешется”:
“С помощью функционального магнитного резонанса авторы проанализировали активность различных зон мозга во время почесывания. … Исследование показало, что почесывание подавляло активность передней и задней поясных извилин коры головного мозга, связанных с восприятием боли и с воспоминаниями о ней. При этом, чем дольше человека почесывали, тем меньшая активность наблюдалась в этих зонах мозга”.
На самом деле механизм - на очень низком уровне организации природной нейросети: это - латеральное торможение. Если соседние рецепторы возбудились, они подавляют шумы (случайные флуктуации активности).
Вот типичная картина: из-за шумовой активности стимул может оказаться самоподдерживаемым (реверберация по А.Иваницкому). Он продолжает ощущаться даже если рецептор уже не активен. Но если почесать вокруг, то стимулы от соседей погасят реверберацию боковым торможением на уровне третичной коры.
Чесаться может как при постоянной активности рецептора, так и по причине реверберации порождаемого стимулом восприятия. Но почесывание тормозит процесс на обоих уровнях.
Теоретики и практики искусственных нейросетей, конечно же, изначально использовали схему многослойного персептрона, которая математически доказывала свою универсальность и гибкость. Не было никакого основания использовать отдельные схемы однослойных и, тем более, одиночных персептронов. Поэтому все искусственные нейросети сегодня созданы на основе многослойного, оптимизированного по количеству элементов в слоях персептроне.
К сожалению, это направление оказалось тупиковым при невероятно впечатляющих успехах собственно функции распознавания на многослойном персептроне.
Разница в том, что в многослойном персептроне обучение ведется для всего персептрона и оно настолько сложно и капризно, что это было настоящей проблемой для разработчиков. А в сети из отдельных слоев одиночных нейронов формирование функции распознавателя осуществляется для каждого распознавателя отдельно и строго последовательно от одного слоя к другому. Это – важнейшее и принципиальное условие эволюции природной нейросети, от которого зависит эффективность адаптации.
Природные распознаватели управляющих схем мозга построены на отдельных упрощенных персептронах, причем очень неточных как по порогу срабатывания суммирующего компаратора, так и по точности коэффициентов передачи синапсов.
Но в компенсацию этого природные синапсы оказались не только возбуждающими и тормозными, а обладают еще и спецификой передачи тока так, что в одних условиях работают только одни виды синапсов, а в других – другие. Для этого и понадобились нейромедиаторы. Это позволило для разных условий формировать независимые друг от друга схемы реагирования, что для природы было очень естественно из-за использования биоматериалов. В таких синапсах использовались разные органические молекулы, которые назвали нейромедиаторами, для организации проводимости изолирующей щели. Достаточно было изменить баланс таких веществ в области работы персептронов и “лишние” оказывались не у дел, а работали только специфичные к такому балансу соединения проводников (на самом деле все куда более заморочено, что не меняет принципа).
Это - как если бы мы сделали несколько разных схем управления для разных ситуаций, у каждой из которых был бы свой источник питания. В зависимости от того, какая из известных ситуаций распознана, включался бы тот или иной источник и работала только нужная схема.
Или же если бы управление было программным, то большие управляющие куски программ активировались бы ветвлением оператором IF (логика: ЕСЛИ это, то сделать то).
Например, IF увидел самку в позе -> программа полового поведения, IF появилась еда -> программа пищевого поведения, IF хищник -> скрываемся или притворяемся мертвыми.
Первый “искусственный интеллект” в интерактивных играх делался на ветвлениях IF. В зависимости от ситуации персонаж что-то спрашивал (первый уровень ветвлений), игрок выбирал из списка вариант ответа, в зависимости от выбора (второй уровень ветвлений) игра получала дальнейшее развитие. Большие блоки программы зависели от того, по какому пути пойти и чем больше было ветвлений IF, тем труднее было ориентироваться в такой программе, выискивая, где начинается тот или иной вариант в коде.
Программистов бесит, когда приходится использовать слишком много ветвлений, потому что такая сложность путает голову. Они придумали на этот случай оператор SWITCH, которому показывают условие, и он переключает нужный блок программы, но и это не решало игровой проблемы.
Невозможность прописать все варианты здорово ограничивало число возможных ветвлений и их уровней, а ситуаций в жизни бывает бесконечное разнообразие и после каждого выбора оценка ситуации меняется. С этим прекрасно справляются схемы природного интеллекта и то, как это проделывается, все еще не могут повторить создатели искусственного интеллекта.
Мы не станем рассматривать полную модель персептрона потому, что она не нужна для реализации нейросетей, но при желании можно с ней ознакомиться, материалов в интернете очень много, например: fornit.ru/7000.
Рассмотрим схему распознавания идеальным персептроном (т.е. с использованием весовых коэффициентов и сигналов как возбуждающих, так и тормозных, чего нет в природной схеме) образов условного зрительного поля, формируемых в неокортексе примитивов восприятия. В более древних структурах может быть любое полезное сочетание возбуждающих и тормозных компонентов активности нейрона. Так, комбинации возбуждающих и тормозных сигналов присуща «природной схеме» цветооппонентных клеток в таламусе, обеспечивающих цветовое зрение.
В приведенной ниже матрице (рецепторного поля) использованы как возбуждающие, так и тормозные сигналы, да еще с дискретизацией значений с точностью 0.1, что обеспечивает распознавание семерки за один раз, одним “нейроном”. Именно таким путем пошли искусственные нейронные сети. Далее будет показано почему такой путь является тупиковым и существует более простой и надежный алгоритм, реализованный природой фундаментально для любых элементов нейросети.
![]()
Слева – сетка отдельных входов нейрона, 16 ячеек, расположенных для удобства так, чтобы отражать фрагмент рецепторного поля.
Конечно же, связи к нейрону могут подходить как угодно переплетаясь, но, главное, каждая связь сохраняет путь, откуда она пришла, что именно означает данный сигнал среди других. И, конечно же, нейрон их всех обезличено суммирует. Никакого заведомого кодирования информации.
На схеме показан фрагмент восприятия цифры 7, написанной от руки. Персептрон уже специализирован распознавать именно такое написание среди множества других возможных видов увиденной семерки. Возбуждающие связи показаны без значка перед их значением, а тормозные – со значком минус. Максимальная проводимость здесь условно обозначена весом связи = 1 (100%), т.е. такой вес сразу перевешивает порог компаратора, или же для этого нужно два сигнала по 0.5.
Итак, K=-1 значит, что ток передается весь, но его действие - обратное, в схеме персептрона - тормозное. Если на одном входе 1, а на другом -1, эти токи (веса) взаимно компенсируются и никак не влияют.
Если сумма весов, ячеек под рисунком окажется больше 1, персептрон сработает, если нет, - будет молчать. Понятно, что ячейки с перекрытием должны иметь веса, в сумме открывающие персептрон с некоторым запасом, а не быть все 1, ведь иначе сработает от любой такой ячейки, а не от суммы всех. А ячейки, на которые фигура не накладывается, должны тормозить по максимуму и тогда любое искажение фигуры не вызовет срабатывания.
Задача текущего уровня понимания – начать понимать, как работает принцип персептрона, что делает его специализированным рецептором (детектором) каких-то признаков, задаваемых предыдущими структурами. Потому как все последующее будет прямо на этом основываться и без понимания этого просто окажется невозможным ясно ориентироваться в нейросетевых механизмах, их причинно-следственной логике.
Вспомнив ранее рассмотренный принцип “дверного ключа” или “сита”, становится очевидным, что пытаться использовать тормозные связи при распознавании рецепторного профиля – так же не просто, как сделать схему ключ-замок, открывающуюся не только когда язычок ключа оказывается меньше отверстия замка, но и не позволяющую открывать, когда язычок меньше отверстия. Или в случае с ситами – сделать сито, которое сразу бы могло отсеивать строго заданный размер песчинок, ни больше, ни меньше.
В случае распознавателя кроме тормозных сигналов нужно использовать точно подобранные веса, а для этого нужно заранее знать, какой образ мы нацелились распознавать и в случае ошибки – корректировать тип воздействия и веса.
Но в том-то и состоит универсальность адаптивности при формировании примитивов восприятия, что неизвестно заранее, что именно нужно научиться различать и нет никакой корректировки ошибок.
Поэтому в природе возник довольно неожиданный для теоретиков искусственных нейросетей механизм, и мы его скоро выявим в полной детализации принципов.
Основной принцип организации природных нейросетей
В искусственных нейросетях персептроны моделируются программно (за счет условных описаний объектов класса Синапс и объект класса Нейрон с параметрами значений весов синапсов). В любых принципиальных схемах от способа реализации персептрона не зависит его функциональность.
Попробуем сопоставить полученные сведения и данные фактических исследований мозга, обобщив их в те аксиоматические основы, на которые мы будем полагаться в развитии уровней усложнения систем адаптации, реализованных управляющими структурами мозга. Т.е. мы должно настолько убедительно для себя определить основополагающие принципы, чтобы они стали надежной опорой всех дальнейших построений.
При этом схемотехнические описания, как и химический контекст, а также отдельные принципы алгоритмического описания (программирования) будут постоянно проявляться на всех уровнях модели. Мы рассмотрим такие механизмы как переключатели, ветвления, обратные связи сигналов в мозге, сдвиговые регистры цепочек реагирования, наводки ритмических электротоков мозга, которым исследователи придают столь высокое значение, рассмотрим реализацию принципа стека памяти в мозге и т.п.
В любой природной нейросети можно выделить три основных блока: рецепторы (сенсорные датчики, специализированные на выделение определенных параметров внешней и внутренней среды), эффекторы (конечные устройства, выполняющие определенные действия при их активации) и нейросеть (промежуточные структуры между рецепторами и эффекторами), которые управляют эффекторами в зависимости от активности рецепторов.
Самые простые схемы – непосредственная связь от рецепторов к эффекторам. Рецепторы выступают в качестве селективного выделения сигналов изменения параметров внешней и внутренней среды, подчас настолько слабых, что их бы не хватило для непосредственной активации эффекторов, которым требуется довольно высокий активирующий потенциал. Но рецепторы бывают настолько чувствительны, что активируются даже при очень слабых сигналах. Так, зрительному рецептору некоторых животных бывает достаточно всего одного фотона.
Чтобы улитка втянула в себя выступающие глаза, бывает достаточно очень слабого, но характерного прикосновения, т.е. защитное втягивание срабатывает достаточно избирательно, что и обеспечивают рецепторы: срабатывание тактильных рецепторов при ползании игнорируется, а вот активность в определенных зонах в упрежденных обстоятельствах порождает общую защитную реакцию. И это – уже не просто связь от стимула к действию, а работа схемы целой группы нейронов, которая при распознавании возможной опасности заставляет активироваться стандартную программу втягивания в себя уязвимых отростков. Эта же программа может быть запущена и сигналами других датчиков, например химического воздействия. Схема такого управления для схемотехников совершенно очевидна и не вызывает никакой трудности в моделировании. Такие схемы предлагают составлять в вузах будущих нейрофизиологов. Вот пример: fornit.ru/an-book-9.
Важно учитывать, что в природной реализации можно констатировать правило необходимости и достаточности, что регулируется механизмами адаптации, реализующей необходимость в дезадаптации, освобождающей от излишнего. Эти критерии, среди других доводов, помогут выделить принципиально важное в системе адаптивности среди второстепенного, т.е. относящегося к особенностям реализации.
Никакие другие воображаемые способы реализации кроме дискретной схемотехники (где элементы самодостаточны и не требуют общего управления от какого-то центра вычислений их состояния) не могут обеспечить подобный алгоритм управления в естественных условиях развития. Хотя программно и возможно так же воспроизвести такую логику, но любая программа это – эмулятор действительности с центральным вычислителем состояния всех элементов, это – условное представление реальных причин и следствий в уже имеющейся конструкции, способной запускать программу, т.е. программа требует уровня собственно организации программного механизма и только потом – уровня реализации программы на нем, что уже выходит за рамки необходимого и является избыточным как по ресурсам, так и по времени эволюции, отведенному на решение вопроса адаптации. Впрочем, это уже отвлечение на философию, но - показательное отвлечение для демонстрации действенности проявила необходимости и достаточности.
В принципе, и в программной реализации есть резон, т.к. в любом случае реализуется принципиальная основа адаптивности. Так, чтобы встроить программное управление, необходимо иметь блок абстрагирования от реальности в область условных данных, алгоритмически обработать их и затем опять использовать преобразователь условных данных в уже реальные действия.
Но именно это и происходит в природной реализации: рецепторы переводят сигналы в условную форму активностей, затем происходит их алгоритмическая обработка по определенной схеме и, наконец, получившиеся активности опять преобразуются уже в реальное воздействие – поведение организма. Так же как и в компьютере сигналы в нейросети совершенно одинаковы – электрические импульсы и их значение определяется тем, каким рецептором они вызваны, так же как и значение в компьютере, от микрофона ли, от видеокамеры или клавиатуры, сканера или мыши или это – уже блок памяти, имеющий условные назначения каждому элементу активности.
Неважно, как именно и с помощью каких деталей (нейронов или транзисторов) организована схемотехника управления, она выполняет определенный алгоритм обработки условно значащих активностей.
Архитектура такого обработчика может быть самой разной, от строгой последовательности обработки в порядке очередности или максимальной параллельности, не требующей никаких очередей, это – не принципиально.
В природной нейросети реализована децентрализованная (дискретными элементами, не требующими управления собой) обработка всех сигналов именно так, как обычно работают электрические схемотехнические устройства.
Если взять любую простую нейросеть в природе (улитка, бабочка и т.п.), то становится очевидным то, что взаимодействие нейронов в ней всегда следует принципу схемотехнического управления эффекторами в зависимости от состояния рецепторов. Эта схема легко может быть повторена искусственно на транзисторах или более сложных и удобных деталях. Она легко понимаема в своей организации, настолько, что можно для тренировки начать создавать свои схемы ответа на стимулы и проверять их адекватность в каких-то моделях.
Подключим к обобщению более общий принцип.
Один из самых фундаментальных принципов эволюции полезной адаптивности: те элементы, которые оказываются наиболее эффективными, т.е. обеспечивают преимущество особям с ними, начинают обкатываться до высочайшей точности и эффективности множеством вариантов в особях, получаемых преимущество.
Принцип кодирования последовательности синтеза белков в зависимости от условий привел к доминированию генетического аппарата, который стал использоваться во всех более сложных организмах потому, что он придал им качественно более высокое преимущество.
Появление клетки привело к тому, что все более сложные организмы стали клеточными потому, что это – результат наиболее эффективной организации.
Появление промежуточных между рецепторами и эффекторами клеток, способных активироваться при превышении порога и распознавать профиль активности на входе, привел к использованию нейронов в мозге.
Принцип схемотехнического построения управления от рецепторов к эффекторам стал единственным из-за качественного преимущества перед любыми другими принципами (к примеру, в fornit.ru/816, предлагаются гипотезы о том, что все нейроны – самостоятельные организмы со своими целями, а не связанные в схемотехнические системы алгоритмов электро-элементы).
Поэтому можно быть уверенными: на любом уровне сложности организации мозга используется именно схемотехника нейросетей и все отдельные механизмы адаптивности мозга поддаются именно такому схемотехническому описанию и моделированию так, что достаточно очевидно можно представить, как именно осуществлен алгоритм того или иного механизма.
Это означает главное: при описании работы мозга необходимо оставаться в системе понимания схемотехнической организации, а не какой-то другой, типа голографической суперпозиции или квантовых эффектов.
Электро-схемотехническая система реализации – необходима и достаточна для организации управляющих структур любой сложности. Все остальное приводит к повышению избыточности реализации, – как бывает при выборе неудачной системы представлений для описания и понимания.
Древнегреческий Зенон задавался вопросом: “Почему мы считаем, что Земля неподвижна, а стрела, которая представляется нам летящей, движется? Почему бы не остановить стрелу и двинуть Землю?”. Понятно, к каким сложностям и неправильным выводам это привело бы. Вывод: систему отсчёта для любого явления нельзя принимать произвольно. Её нужно связывать с сутью причинно-следственных процессов, в которых это явление рассматривается.
Когда появляются фактические данные исследований того или иного механизма на нейронном уровне описания, всякий раз оказывается, что это – именно схемотехнический механизм. Примером можно привести любое такое описание: самоподдерживающиеся образы за счет ревербераций в различных участках мозга, выполняющих функциональность удержания активностей, нейрональные механизмы условных рефлексов, организация карт местности в мозге, организация латерального торможения, селективности, детекция значимых признаков, различные виды иллюзий, системы возбуждения и торможения, доминанта, механизм прогностического “опережающего возбуждения”, организация цепочки поведенческой реакции и т.п.
Это не доказывает, что никаких других базовых принципов организации нейросети нет в мозге. Но никаких таких принципов не обнаруживается ни в каких своих проявлениях. Они никем не описывались даже в качестве правдоподобной гипотезы.
При этом многие исследователи упорно пытаются найти какие-то такое принципы и новые подходы системного описания, даже не представляя хоть что-то, намекающее на это. Часто для этого подхватываются новые идеи типа голографического принципа, частотных синхронизаций и т.п. Авторы уверены, что есть нечто такое сакраментально недостижимое пока в понимании, что способно ответить на такие немыслимые вопросы, как связь души и тела, как то, почему мысль способна оказывать воздействие на материальное? Как материя мозга рождает субъективное явление?
Все такие вопросы показывают, что нет достаточно определенных представлений о самих используемых в этих вопросах понятиях (т.е. сама постановка вопроса некорректна), что нет главного: хоть сколько-то определенного понимания функции механизмов мозга, порождающих эти сакраментальные явления, ведь для чего-то все это понадобилось, показав свои преимущества в эволюционной конкуренции. Не видя таких адаптивных преимуществ (точнее, даже не задумываясь о таком критерии), многие исследователи объявляют важнейшие явления психики побочным продуктом деятельности мозга.
Для суждений (сопоставлений и обобщений) по вопросу о «побочности» психики, и, в частности, о ее соотношении с процессами мозга, полезно обратиться к широко обсуждаемой в среде психофизиологов книге философа Д. Чалмерса (2015). «Сознающий ум. В поисках фундаментальной теории». И тогда получим представление о том, какими критериями сопоставлений и обобщений в проблематике организации психики в мозге воспользовались современные теоретики. В этом плане предлагается итоговая статья: “Что люди узнали о мозге” - fornit.ru/19798.
Свойства нейрона
Можно сопоставить несколько эвристических принципов для оценки правдоподобности того, что в нейросети мозга главным является использование функции персептрона, но в его наиболее простой форме (с отсечением излишне сложного по принципу ключа или сита). Пронумеруем эти принципы.
1. Нейрон активируется по тому же принципу, что и клетки мышц, только нейроны при этом не сокращаются, а выдают электрический сигнал о своей активации. И мышечную клетку, и нейрон можно возбудить, подведя электродом ток достаточно высокого потенциала. Оба вида клеток одинаковым принципом организуют свои мембранные потенциалы, возбуждаются при их превышении и регенерируют, восстанавливая разность мембранных потенциалов, распространяют возбуждение по всему волокну.
Вспомним: “Стоит заметить, что в живом теле разные клетки приспособились к различным таким строго выраженным специализациям и развиваются из одних “стволовых”, неспециализированных клеток, которые формируют свою специализацию в зависимости от окружающих условий (такая зависимость начального наследственного кода от условий описана в fornit.ru/806).
Клетки различных рецепторов способны реагировать на какие-то воздействия, выдавая, как нейрон, импульсы тока и в простейших организмах, где нет нейронов, прямо от них сокращаются мышечные клетки или выделяются нужные вещества. Таких видов рецепторов много: клетки, реагирующие на фотоны света, клетки, активирующиеся от разных видов химических веществ, чувствительные к механическим колебаниям специализируются на рецепторы звуковых колебаний, на рецепторы прикосновений, на рецепторы сокращения мышц и т.д.
Нет каких-то хитрых скрытых других функций кроме тех, что явно проистекают из специализации.
Вот и клетки нейронов, как и клетки рецепторов, в результате своей функции выдают импульсы тока на раздражение, только раздражителями для них являются заряды, возникающие на синапсах, покрывающих тело нейрона. А эти заряды появляются в ответ на электрические токи предшественников: рецепторов или таких же нейронов.
Вот почему все нейроны – такие рецепторы, функцией которых является реагирование на состояние предшествующих им клеток. Так же все нейроны воздействуют на последующие эффекторы – тоже нейроны или уже мышечные (или продуцирующие гормоны) клетки, называемые эффекторами.
Получается, что все слои связанных в сеть нейронов – эффекторы по отношению к предыдущим и, в то же время, они же – рецепторы для последующих, это – условно нами выделяемые роли в нейросети.
Само по себе это не доказывает, что нейрон – только лишь схемотехническое устройство для распознавания суммарного профиля активности по своим входам и выдачи результата распознавания далее.
2. Все простейшие нейронные сети у живых существ с очевидностью построены как схемы управления эффекторами в зависимости от состояния рецепторов. Эта логика прослеживается однозначно.
3. Нет никаких оснований думать, что природа не продолжает развивать нейросети по схемотехническому принципу, никто не нашел и не показал что-то, отличающееся от него.
4. Если явление работы мозга можно описать как схемотехническое устройство, а не нечто более сложное и непонятное, а природа изначально использовала именно схемотехнику управления, то нет причин делать что-то более сложное. Т.е. если мы смогли описать даже самые сложные психические явления как схемы взаимодействия более простых схем, то незачем предполагать что-то сложнее. Уже практически доказано: сколь угодно сложную систему распознавания и управления, вплоть до самоадаптирующейся к новому, возможно сделать на основе всего одного конструктивного элемента: персептрона. Это – необходимое и достаточное условие, не требующее появления чего-то еще для целей управляющих структур мозга. Природа нашла именно такой элемент, но оптимизировала его в еще более простой и универсальный и использовала его в мозге, имея возможность на его основе организовывать любые уровни адаптивности к новому.
Если каждый из перечисленных доводов оценки сам по себе и не является достаточно убедительным, то совместно убедительность возрастает так, как это описано в прикидке вероятности того или иного события, - т.е. возрастает очень резко: fornit.ru/7498, в частности, можно применить эвристическую версию метода Байеса.
Теперь, при должном понимании и сопоставлении приведенных доводов, нет никакого основания допускать в основном принципе организации природной нейросети что-то еще кроме схемотехнических принципов, если не говорить о множестве побочных эффектов в особенностях реализации этих принципов. Современные искусственные нейросети так же могут реализовываться схемотехнически на электро-элементах и это была бы более эффективная реализация (т.к. не использует центральное устройство отслеживания состояния всех узлов, а они работают децентрализовано), но их начали делать программно на устройствах, сделанных схемотехнически, причем устройствах громоздких из-за необходимости невероятно больших ресурсов для таких вычислений.
И, все равно, строго говоря, это – не доказательство, т.к. есть ненулевая вероятность иного и т.п. Но в плане формирования модели это очень хорошо согласуется с критериями полноты и верности (fornit.ru/7649), начиная с самого фундаментального каркаса модели, и поддерживается всеми последующими взаимно согласованными звеньями в общем пазле картины представлений. Т.е. модель, построенная на таких принципах (модель МВАП fornit.ru/7431), оказалась непротиворечивой, полной и эффективной.
Поэтому с полной уверенностью и очевидностью будем полагать, что базовая функция нейрона (fornit.ru/6449) и то, как взаимодействуют нейроны на любых уровнях организации мозга, определены однозначно.
Т.к. нейрон на выходе имеет сигнал типа да-нет (он или возбужден, или нет), то можно было бы предположить, что в нейросетях используется двоичная логика и в полной мере применима булева алгебра для такого вида кодирования. Но нейрон еще может возбуждаться коротко или длительно (изменяя частоту импульсации от уровня своего возбуждения).
Так что именно кодирует нейрон? Об этом до сих пор ведутся споры и делаются самые фантастические предположения.
В первую очередь нейрон, являясь рецептором, кодирует то, что в восприятии есть некий профиль (совокупность признаков) элементарных компонентов первичных рецепторов, профиль, отражающий выделенную (абстрагированную) особенность воспринимаемого. Нейрон является распознавателем (детектором) специфического профиля активности на своих входах. Есть такой профиль – есть активность нейрона.
Во-вторых, нейрон активен столько времени, сколько активен и профиль на входе (в реальности до своего истощения), т.е. он длительностью активности отражает длительность воздействия профиля, если только нейрон не тормозится соседями или специально не активируется (например, обратной связью, обеспечивающей удержание активности, когда уже нет активной рецепции).
В природной нейросети нейрон меняет частоту импульсов от редкой при слабом распознавании до максимальной – при точном распознавании, но это – скорее неидеальность, мешающая логике распознавания, размывающая эту логику (хотя и в этом есть свои плюсы). Поэтому не будем считать это свойство системным.
Ничего более нейрон в принципе не может кодировать. Он – просто специфический рецептор или детектор появления-пропадания данного абстрагированного признака.
Различие искусственных и природных нейросетей
Теперь можно констатировать некоторые (пока далеко не все) принципиальные различия между принципами построения искусственных нейросетей (в частности, глубокие сети (fornit.ru/6616 - глубокие – т.е. имеющие много внутренних слоев): fornit.ru/7359, fornit.ru/7513, fornit.ru/7120, fornit.ru/7360.
| Искусственная нейросеть | Природная нейросеть |
| Многослойный персептрон с общей функцией распознавания для всей сети, в котором объединены все профили рецепторов. Связь типа все – со всеми. | Состоит из однослойных упрощенных персептронов, каждый из которых формирует свою собственную функцию распознавания, связанных с последующими слой за слоем. |
| Формируется сразу и обучается вся сеть сразу. | Формируется строго поочередно (сначала созревает очередной слой и начинает специализироваться), начиная с самых простейших примитивов распознавания, на каждый слой которых выделяется определенное время (критический период развития). У человека на завершение развития всех слоев уходит 25-30 лет. |
| Может обучиться очень точному распознаванию, улавливая самые незаметные общности в явлении – при условии достаточного числа (сотни тысяч и более) предъявляемых примеров с имеющейся оценкой верности каждого из них. | Очень неточное распознавание, но зато предельно ограниченного числа входных признаков для каждого специализирующегося персептрона. В распознавании очень важно влияние соседних персептронов для контрастирования общего (для участка слоя) профиля на входах. Каждый отдельный персептрон обучается “без учителя”, просто специализируясь на наиболее частом профиле возбуждения за время его обучения. |
В области слоев первичных примитивов зрительного восприятия нет никаких линий, кругов, семерок и других выделяемых нами по смыслу фигур. Есть самые разные неравномерности яркости и цвета, которые нам бы показались похожими на линии, круги, семерки и другие символы. Значимость этих фигур ассоциируется уже в третичной зоне теменной коры, а смысл – в префронтальной лобной коре в процессе его осознанного понимания. Но это слишком преждевременное утверждение. Важно понять, что с примитивами первичной коры не связаны никакие значимости и смыслы, они просто - некая коллекция, более изощренно выделяемая рецепция, которую можно как-то использовать потому, что она абстрагирует из самых первичных элементов восприятия некие фигуры и с этими фигурами теперь можно связывать какие-то реакции, например, при появлении длинных, извилистых фигур – реакцию избегания и страха.
Условия образования связей между нейронами
Главным механизмом формирования специфики распознавания являются условия образования связей между нейронами.
То, какая связь вообще в принципе может образоваться, в первую очередь зависит от того, как, в каком направлении и какой длины вырастет выходной отросток нейрона – аксон. Это предопределяется наследственно, плюс влияют условия развития.
Входные отростки находят свои мишени для роста более просто: их рост стимулируют активирующиеся предшествующие нейроны, которые при активации специфически меняют состав электролита вокруг себя, и рост дендритов следует направлению к таким местам.
В формировании и развитии нейронной сети, в точности, как и при развитии любой ткани тела, определяющую роль играет последовательность выработки специфических белков (строительных и способствующих формированию данного вида ткани) – т.н. экспрессия генов. Генетический код устроен так, что выработка тех или иных белков происходит в строго определенных условиях. Если появляется такое химическое окружение, то начинается и такая экспрессия. В других – другая. Так клетка определяет, во что ей специализироваться при развитии и делении.
Принципы наследования и реализации наследственного кода описаны в статье: “Наследование признаков” (fornit.ru/806), с которой очень стоит ознакомиться и ясно понять эти принципы потому, что эта логика непосредственно отражается в развитии всех последующих усложнений мозга. Именно поэтому развитие организма повторяет развитие вида в каждой своей стадии. Именно поэтому и усложняются механизмы мозга на основе предыдущих так, как это было заложено в последовательности экспрессии наследственного кода.
Отсюда возникает еще одно фундаментальное для нейросети свойство: определенная последовательность развития нейрона, когда он находит свое место в нейросети, созревает до стадии возможности образования связей и завершает свое развитие стадией распознавательной специализации. Этот период формирования специализации определен генетически для каждого слоя последовательно созревающих нейронов и называется критическим периодом развития.
Существует общая закономерность: более поздние критические периоды имеют большую продолжительность, что обусловлено тем, что более сложные примитивы распознавания встречаются реже в восприятии и их нужно успеть зафиксировать. Кроме того, у каждого вида животных оптимизируются свои продолжительности критических периодов. У человека они – наиболее продолжительны (последние формируются годами). Следующим видом после человека по продолжительности таких периодов оказываются высшие обезьяны.
Из ранее сказанного ясно, что именно продолжительность критических периодов развития дает и более эффективный результат приобретения навыков, из-за того, что в данный период умещается большее число новых задач адаптивности и ее решений. В социальном же плане за более продолжительные периоды становится возможным перенять большее число навыков от своих родителей и авторитетов. Это и определило интеллектуальное (т.е. система наработанных навыков понимания и реагирования) преимущество человека.
Генетический код способен кодировать только продуцирование белков в окружении специфики также белкового характера, но, конечно же, не интервалы времени. И поэтому последовательность созревания слоев организуется в точности по тому же принципу, как и последовательность формирования тканей: по метаболическому признаку завершения (или достаточной активности) предыдущего периода. Это значит, что после достаточно долгой экспозиции слоя рецепторами, возникают условия для созревания и готовности к специализации следующего. И это очень логично (отражает реальную причинность взаимовлияния факторов).
Принципиально нет смысла продолжать что-то менять в предыдущем слое, когда уже специализировался последующий, ведь эти изменения уже не смогут повлиять на последующий слой. Значит, вырабатывается какой-то метаболический признак завершения специализации слоя. Как именно это проделывается, пока не известно. Но все более долго экспонируемые слои требуют, чтобы был такой механизм. Этим обусловлено и то, что завершившаяся специализация в первичных зонах мозга уже не может быть изменена в норме, а потеря распознавателей уже не может быть восполнена в этой зоне, а только – косвенно в поздних, еще пластичных слоях.
В самом деле: если что-то поменяется в предыдущих слоях системы управления, то окажутся неверными все последующие, которые были основаны на предыдущих.
С точки зрения системных представлений нам не нужны эти детали, но они позволяют убеждаться в общей логике развития нейросети. Достаточно отследить, как именно в природе возникают все более совершенные уровни механизмов адаптивности к новым условиям. Поэтому будем рассматривать принципиально важные особенности развития природной нейросети. Выделим три важнейших принципа, сопровождающих формирование природной нейросети:
1. Спонтанная активность нейрона, развившегося до стадии способности образования связей.
2. Условия образования связей.
3. Строгая последовательность развития слоев специализирующихся нейронов, отражающая эволюционную последовательность усложнения вида данного животного.
Эти принципы далеко не общепризнаны и поэтому требуют, кроме принципиальной системной верификации модели, поддержки фактическими данными исследований.
Описывают несколько механизмов спонтанной активности в нейросети (fornit.ru/22946), нас интересует конкретно спонтанная активность нейрона, не имеющего тормозных связей, что подтверждается большим числом фактических данных исследований.
Причем, “понимание механизма генерации спонтанной активности в развивающихся нейросетях представляется фундаментальной научной задачей, поскольку, как было показано, явление не является особенностью какой-то одной структуры, а отражает общую тенденцию”. (fornit.ru/6344).
“Новые нейроны, которые появляются в мозге, обладают особыми свойствами. Например, они являются гипервозбудимыми, и именно благодаря такой своей гипервозбудимости они и способны каким-то образом модифицировать память” (fornit.ru/6409).
Одиночный пейсмейкер “состоит из одного нейрона, в котором возникает внутриклеточная реверберация импульса.”. Причем, эта активность проявляется даже после специализации, если связи вдруг оказываются оборваны, например, при патологии слухового нерва возникает шум в ушах (fornit.ru/5284).
Другой эффект – фантомная боль:
“Если поврежден непосредственно периферический нерв, ниже узла заднего корешка, то центральная нервная система продолжает получать афферентную импульсацию из самого периферического нерва, выше места повреждения и из эктопических источников ее генерации. В этом случае всегда возникает патологическая сенсорная афферентация с развитием периферических невропатических болей”. (fornit.ru/5172).
“...вышедшие из-под тормозного контроля нейроны, теряют свои прежние функциональные характеристики и образуют новую группу, которая становится ядром нейронной популяции генератора” (fornit.ru/5407).
“Л. Тауц и Г. М. Гершепфельд 11960] обнаружили, что соматическая мембрана нейронов моллюсков, не имеющая на своей поверхности сипаптических окончаний, обладает высокой чувствительностью к медиаторным веществам и, следовательно, имеет молекулярные хемоуправляемые структуры, свойственные иостсинаптической мембране. Наличие внесинантической рецепции показывает возможность модуляции пейсмеккерной активности диффузным действием выделяющихся медиаторных веществ”. (fornit.ru/5347).
“Известно, что растущие нейроны с какого-то момента начинают проявлять активность: в культуре клеток возникает возбуждение, которое распространяется по всей колонии. Такое поведение нервных клеток обычно сопряжено с обучением и запоминанием, но нейроны начинают вести себя так, когда ни обучение, ни запоминание ещё невозможны в принципе — клеток слишком мало. По словам исследователей из Университета Твенте (Нидерланды), экспериментировавших с культурами нейронов молодых крыс, начинается эта активность у клеток на первой неделе роста и никакой иной причины, кроме этого самого роста, не имеет”. (fornit.ru/5424).
Спонтанная активность оказывается фактором, обеспечивающим образование связи, начиная с самого раннего этапа развития.
“После появления человека на свет на его мозг обрушивается бесчисленное количество раздражителей. Считается, что все эти стимулы из внешней среды в значительной мере помогают сформировать архитектуру мозга в постнатальный период: нейроны получают сигнал, образуют или убирают синапсы и так далее. Однако не так давно сразу в нескольких нейробиологических лабораториях было показано, что огромную роль в формировании нейронных сетей играет так называемая спонтанная активность мозга, которая впервые проявляется ещё до рождения и которая не требует для своего возникновения внешних стимулов.
Исследования этой спонтанной активности продолжаются и приносят довольно любопытные результаты. Так, нейрофизиологи из Йельского университета (США) сообщают, что для правильного формирования нейронных путей между глазом и мозгом необходима строгая очерёдность проявления спонтанной нейронной активности в глазах в первые дни жизни”. (fornit.ru/5442).
Первые связи между рецепторами и эффекторами возникают в результате спонтанной активности эффекторов.
“Исследования на обоих этих объектах показывают, что ранняя сенсорная стимуляция мозга обеспечивается эндогенными механизмами - спонтанными миоклоническими подергиваниями в соматосенсорной системе и спонтанными волнами сетчатки в зрительной системе”. (fornit.ru/7532).
Конкретизируем условия образования связей
Условия образования связей универсальны для любых зон мозга и выполняются на всех уровнях организации мозга. Это – одно из фундаментальных оснований, определяющее самый общий контекст развития нейросетей. Так, в соответствии с этими условиями уже от самых первых рецепторов возникают связи с эффекторами. Конкретизируем эти условия.
Данных фактических исследований этого достаточно много (fornit.ru/ax1-20), начиная от экспериментов с отдельными нейронами (fornit.ru/5297 - т.е. в предельно выделенных от влияния чего-либо условиях).
Уже были сформулированы основные правила образования связей между нейронами:
“Если два нейрона неоднократно активируются одновременно, связи между ними усиливаются в обоих направлениях.
Если два нейрона неоднократно активируются последовательно, усиливается связь, направленная от одного ко второму.
В обоих правилах предполагается, что связи усиливаются навсегда или, по крайней мере, надолго: так ассоциация закрепляется в памяти”. (fornit.ru/7224).
Условием, при котором начинается процесс формирования проводимости в синапсе, является достаточно длительная активность с двух сторон (около получаса – время найдено эмпирически) от синапса. Она может прерываться, но важно, чтобы вокруг такой активности возникали метаболиты, способствующие формированию проводимости синапсов данного вида. За это время повторяющиеся активности сформируют устойчивые связи, а случайные будут погашены латеральным торможеным. В результате сформируется вполне определенный профиль возбуждения данного нейрона.
Так образуются все рефлексы, начиная от простейших:
“Одним из условий выработки оперантных моторных условных рефлексов является обязательное включение в систему раздражителей импульсации, возникшей спонтанно или вызванной активным, или пассивным движением”. (fornit.ru/5231).
Итак, в случае образования простейшей связи типа стимул-ответ (от рецептора к эффектору) из-за пейсмекерной активности начинают подрагивать отдельные волокна мышц, а если рядом есть аксон от возбуждающегося одновременно рецептора, то образуется связь от рецептора к эффектору.
Этот механизм реализуется на ранних стадиях активности, например, подергивания плода человека в утробе как раз и способствуют наработке первичных связей, на основе которых будут формироваться все остальные.
Даже новые нейроны гиппокампа (нейрогенез во взрослом состоянии) при созревании проявляют повышенную активность без каких-либо связей (fornit.ru/23499).
Возникает следующая картина. Когда нейрон созрел, он проявляет спонтанную активность. Это не постоянная интенсивная активность, а, как описывалось в приводимых работах, спайки (импульсы на выходе) возникают, провоцируюясь метаболическим накоплением разности концентраций между мембраной нейрона до его срабатывания. При разряде разница концентраций электролитов выравнивается и потенциал опять становится низким. Т.е. импульсы возникают бессистемно время от времени (визуально это ощущают беременные женщины как толчки плода при такого рода активности мотонейронов в мышцах).
Когда такая активность совпадает с активностью предшествующего рецептора (или первичного или уже сформированного нейрона), начинает укрепляться связь так, что активность на выходе все в большей степени начинает отслеживать рецепторную. Получается нечто вроде синхронизации укреплением связей. Нейрон выдает импульсы уже не просто хаотически и редко, а все более чаще - в соответствии с активностью рецепторов на своем входе (по ту сторону его синапсов).
Это – еще одно отличие от поведения математического персептрона: “Совокупность параметров стимула кодируется нейроном-детектором в виде частоты потенциалов действия, при этом нейроны-детекторы обладают избирательной чувствительностью по отношению к отдельным сенсорным параметрам”. (fornit.ru/22948). Чем больше точность соответствия все более специализированного нейрона, тем более активно и полноценно он реагирует своей активностью. Т.е. нейрон проявляет не пороговый характер распознавания, но чем точнее распознавание, тем сильнее он влияет на последующие нейроны.
Экспозиционный характер обучения во время действия специфического профиля рецепторов на входе дополняется латеральным торможением соседних детекторов, что, с одной стороны, делает распознавание еще более определенным, а, с другой стороны, тормозное влияние уже надежно запирает нейрон, не позволяя ему самопроизвольную активность, хотя при полном отсутствии сенсорных сигналов появляется такая активность в виде шумов, т.к. при этом отсутствует латеральное торможение.
Это можно заметить прямо на себе. Если нет ничего в восприятии, то его острота нарастает (соседние рецепторы не заглушают) вплоть до появления отдельных иллюзорных ощущений. Так, вдруг может зачесаться что-то на коже, даже если ничего это не провоцирует. Мы чешем это место, заставляя активироваться соседей и приглушая иллюзорную активность. В полной темноте начинают возникать хаотические ощущения световых точек и пятен.
Так можно очень зримо прочувствовать динамику этого механизма.
Особенности природной реализации по-своему реализуют, все же, основной принцип персептрона, который вообще не зависит от того, каким именно способом выходной сигнал отражает верность распознавания: хоть частотой импульсов, хоть пороговым “да-нет” для алгоритмической реализации, хоть пороговым потенциалом – для схемотехнической модели на электродеталях. В любом случае главное – специализация функции распознавания профиля активности на входах за счет изменения веса, привносимого каждым из входов в общий отклик.
В качестве развития представлений полезно ознакомиться с “Векторным принципом кодирования” по работе Е.Н. Соколова (2010). “Очерки по психофизиологии сознания” (fornit.ru/sokolov_3).
Имеющийся способ реагирования определяет контекст реализации нейросети и ее функционирования. Поэтому, рассматривая природные механизмы, всегда будем иметь ввиду особенности именно природной реализации, что, как в этом можно будет убедиться, не помешает моделировать эти механизмы чисто схемотехническими условностями.
Строгая последовательность развития слоев
Этот принцип в развитии природной нейросети уже упоминался ранее: “Отсюда возникает еще одно фундаментальное для нейросети свойство: определенная последовательность развития нейрона, когда он находит свое место в нейросети, созревает до стадии возможности образования связей и завершает свое развитие стадией завершенной специализации. Этот период формирования специализации определен генетически для каждого слоя последовательно созревающих нейронов и называется критическим периодом развития. Существует общая закономерность: более поздние критические периоды имеют большую продолжительность, что обусловлено тем, что более сложные примитивы распознавания встречаются реже в восприятии и их нужно успеть зафиксировать.”. Подборка материалов фактических данных исследований: fornit.ru/ax1-32, ornit.ru/ax1-55.
“Можно выделить три наиболее общих и качественно различающихся по функциональности зоны мозга, отражающие последовательность эволюционных скачков механизмов адаптивности: первичные, вторичные и третичные зоны мозга. Каждая из них так же подразделяется на свои периоды последовательного развития”. (fornit.ru/6443).
Последние периоды развития мозга человека действуют после 21 года (fornit.ru/7521).
Современная искусственная нейросеть “глубокого обучения” обучается совершенно иным образом: при данном профиле на ее входах она ждет сигнала о том, является ли этот случай правильным распознаванием данной специфики или нет, и такой сигнал ей дают извне, “учителя”. Поэтому дополним таблицу различий еще одним пунктом:
| Искусственная нейросеть | Природная нейросеть мозга |
| Обучается учителем, обычно заранее заготовленной последовательностью примеров верного или неверного распознавания в очень большом количестве (пропорциональном сложности объекта распознавания в отличиях вариантов правильного и неправильного распознавания). | Обучается без учителя, при экспонировании профиля активности на время, достаточное, чтобы некоторые совпадающие активности оказывались более рельефными, чем случайные, что так же контрастируется латеральным торможением. Время экспонирования равно времени собственной активности нейрона (пейсмекерной или вынужденной из-за возбуждения по другим входам). |
Забегая вперед, стоит заметить, что на самом деле “учитель” в поздних участках природной нейросети есть, но это организовано совершенно не так, как в искусственной и есть два основных уровня, достигнутых эволюцией: подкрепление в условных рефлексах и произвольность оценки в осознаваемо формируемых автоматизмах.
Это различие делает обучение нейрона нетипичным для персептронов, но основной принцип персептрона остается актуальным.
Слова это – такие условности, зависящие от контекста и вкладываемого смысла, так что как будет назван объект внимания не меняет его сущность: можно называть нейрон с системой своих синапсов персептроном, но при этом возникает немалое число озадачивающих различий. Но лучше называть его по бесспорно проявляемой функции – распознаватель профиля активности на своем входе или детектор. Мы будем называть его распознавателем во всех ипостасях его участия в нейросети.
Рефлексы
Самые простейшие реакции от рецепторов к эффекторам называются таксисами, а те, что связаны с активным выбором направления действия - топотаксисы. Такие простейшие реакции являются основой для всех более сложных безусловных и, на их основе, условных рефлексов. В области еще большего усложнения на их основе, с участием осознанной произвольности формируются неосознаваемые автоматизмы.
Различием в качестве этих видов реакций является то, на какой стадии развития и каким образом осуществляется их формирование и закрепление.
Таксисы и безусловные рефлексы формируются за счет отбраковки особей в условиях естественного отбора: те, который выживают из-за случайной мутации формирования связей (при важном условии – флуктуации ветвления аксонов до формирования связей) или те, у которых уже есть спасительная реакция (как негативного избегания опасностей, так и позитивного стремления к поддерживающим факторам). Учителем здесь является естественный отбор.
На данном уровне негативность или позитивность (вред или польза) никак не оцениваются, а просто является результатами случайных мутаций, утвержденных наследованием. Такие рефлексы могут быть довольно сложными, образуя длинные цепочки, создаваясь на основе более простых, с учетом реагирования на дополнительные стимулы. В принципе ничто не ограничивает сложность таких реакций и даже на уровне муравьев и пчел они поражают исследователей своей сложностью и точностью.
Отдельно стоит выделить безусловный “ориентировочный рефлекс”, который оказался важнейшим механизмом, обеспечивающим основы осознанного внимания.
Условные же рефлексы возникают как индивидуальная адаптивность к новым условиям, но и они НЕ используют гомеостатическое распознавание вреда или пользы (в отличие от утверждаемого в академической науке). Это будет обосновано.
Автоматизмы возникают на основе условных рефлексов с возможностью блокировки при негативных последствиях, для чего в организме есть специальные распознаватели от рецепторов отклонений жизненных параметров организма (вред) и возврата этих параметров в норму (польза).
Как это происходит и почему вообще стало происходить, мы рассмотрим далее.
Начальные связи нейронов
Перед тем как перейти к рассмотрению последовательностей эволюционных апгрейдов природной нейросети, необходимо окончательно уточнить основы.
Вспомним про модели нейрона. Реальный нейрон по сравнению с рассмотренной моделью не имеет никакого потенциала на выходе в состоянии покоя, т.е. как бы оборвана цепь по току, а на схеме
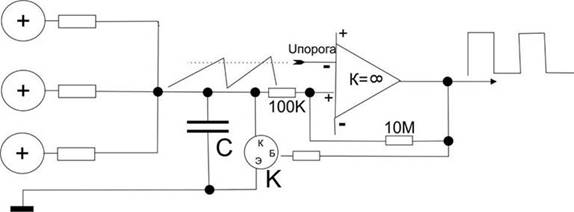
он имеет низкий потенциал выхода усилителя, что оказывало бы постоянное разряжающее влияние на конденсатор модели последующего нейрона (но не в природной нейросети, где синапсы не реагировали бы на это).
Это совершенно не трудно обеспечить схемотехнически всего лишь введением диода на выходе модели:
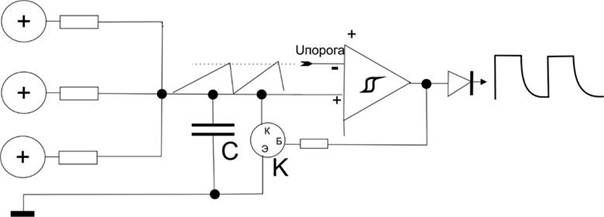
(Вместо резисторов положительной обратной связи (10мегаом и 10килоом), просто нарисуем значок гистерезиса на компараторе.)
В схеме к выходу подключили диод – электродеталь, которая пропускает ток только в одном направлении: по стрелке его обозначения, в данном случае он пропускает только если на его входе будет + больше, чем потенциал на выходе (понятно, что если там будет такой же или + потенциал, то он закроется). При положительном импульсе диод резко станет проводником и положительный заряд тока тут же окажется на его выходе. Но когда на его входе будет низкий потенциал, он выключится и разорвет цепь. Потенциал на его выходе станет полностью зависеть от последующей цепи. Пологий спад говорит о том, что на выходе есть что-то, сначала зарядившееся от положительного импульса, но после обрыва цепи постепенно разрядившееся (как резистор связи, подключенный к емкости конденсатора).
В дальнейшем мы будем просто использовать условное обозначение этой схемы в виде кружка со стрелками связей к нему и от него, как это показывалось раньше на схеме, иллюстрирующей простую цепь управления в зависимости от условий:
А если будет важно показать его профиль возбуждения, то, изобразим его в виде таблички, как это тоже уже делалось:
![]()
Вид выходного сигнала для нас в условных моделях нейрона, реализующих принцип персептрона, перестает иметь значение: хоть импульсный, хоть постоянного тока, модели будут работать одинаково эффективно и демонстрировать все свойства нейронных соединений.
Для формирования связей необходимо выполнение условия образования связей и готовность последующего от рецептора нейрона к образованию связей своей в виде спонтанной или вырванной чем-то активности.
Таких случаев может быть два:
1) модифицируются связи уже специализированного нейрона, при этом будет меняться его специализация и он станет распознавателем уже другого образа на его входах и
2) образуется новый распознаватель, для чего нужно, чтобы в этом месте появился и созрел до функциональности пейсмейкера новый нейрон.
Если позже (по времени) такого нейрона была уже сформирована сеть распознавателей на его основе, т.е. предыдущие элементы использовались для профиля распознавания последующих, то в обоих случаях описанной модификации все последующие распознаватели потеряют свою специализацию, скорее всего начнут реагировать беспорядочно или молчать потому, что для адекватного какой-то задаче реагирования нужно переучивать все последующие слои распознавателей, - вспомним принцип строгой последовательности развития слоев нейронов.
Чтобы не было путаницы, нужно напомнить, что слои нейронов – довольно поздняя находка эволюции, реализованная в “новой коре” мозга, а до этого в управляющих цепях практиковались хаотические флуктуации связей. Но в любом случае специализация последующих нейронов происходит позже, чем предыдущих, что отражает постепенное эволюционное усложнение управляющих цепей.
Это достаточно очевидно для всех, ясно представляющих себе логику такой взаимосвязи.
В реальном мозге случается, что оказываются поврежденными какие-то промежуточные распознаватели и конечная функция оказывается потеряна и такая потеря невосполнима, несмотря на определенную пластичность, запускаемую в зонах поражения. Вот почему в первичных зонах мозга не бывает новых нейронов после того, как там уже закончилась специализация слоев после прохождения очередного критического периода развития. Такого механизма для переучивания всех последовательностей слоев в природе не существует потому, что гораздо проще начать с нуля – с нового организма.
Но последний критический период развития у высших животных продолжается всю жизнь, обеспечивая возможность адаптации к новому, в том числе для случая потери рецепторов (как внешнего воздействия, так и внутренних распознавателей) или приобретения новых терминалов: освоения палки, катания на коньках и т.п. И в зонах мозга, где это происходит (район гиппокампа и префронтальная лобная кора), возникают новые нейроны с определенной периодичностью, пропорциональной частоте необходимой адаптации к новому. Где и почему возникают новые нейроны, мы рассмотрим при формировании моделей адаптивных механизмов третичных зон мозга: в теменной коре и лобной коре.
Таким образом, есть еще одно дополнение в таблице отличий моделей искусственных глубоких нейросетей от реальных:
| Искусственная нейросеть | Природная нейросеть мозга |
| Способность только переобучаться за счет уже имеющейся архитектуры слоев, без добавления или убавления рецепторов и эффекторов. | Способность развивать новые распознаватели для адаптации к новым условиям даже при потере каких-то внутренних распознавателей или потери рецепторов и эффекторов или приобретения новых эффекторов (рост, регенерация мышечной ткани). |
Клетки с функциональностью нейронов находятся не только в головном мозге, но и в мышцах (водитель ритма сердца) и вообще в стенках внутренних органов, обладающих сократительной активностью, в гладких мышцах, сосудистой и капиллярной сети, эндокринные и иммунные образования. Такие включения названы метасимпатической нервной системой. Так что регуляция, использующая свойства нейронов, буквально повсеместна. Связи, образованные такой регуляцией древние, но постоянно эволюционируют, как эволюционирует вообще все в организмах.
С самого раннего периода эволюционного развития организмов, находка эволюции в виде нейронов используется повсеместно, где от нее есть польза организму. Даже самодостаточные образования, но зависимые от более общей управляющей нервной модуляции (типа водителя ритма) имеют свои датчики условий (рецепторы), которые влияют на работу нейронов для управления эффекторами. И такие связи, образуясь по общему принципу, возникают в соответствии с последовательностью запрограммированной в генах экспрессии генов, вовремя обеспечивая нужную функциональность.
Причинно-следственная последовательность образования связей
С момента первого деления первой клетки организма на геном влияет окружающая клеточная среда, определяя, какой именно участок генома в этих условиях должен быть активным (определяя момент экспрессии генов в этих условиях). Одна клетка, две клетки, четыре клетки могут мало менять среду, хотя соседство клеток меняется, и клетки просто делятся, увеличивая количество. С некоторого момента окружения условия меняются настолько, что возникает следующая по эволюционной цепочке, записанной в геном, экспрессия генов и масса клеток приобретает различия в свойствах и функции уже измененной ткани.
Так, с каждым новым воплощением последовательности развития вида, с каждым новым условием, в которых оказывается каждая из клеток организма, все они включают свою, соответствующую последовательность генов и при делении возникает уже нечто иное. Т.е. каждая клетка активна в том участке своего генома, который распознает специфику окружающих условий. Это порождает все многообразие органов и свойств организма.
Влияют не только внутренние условия, но и внешние, если это как-то предусмотрено и распознается в геноме. Так, деревья, высаженные в высокогорье или в холодных зонах, приобретают кустарниковую или карликовую форму.
Внешние условия могут оказаться такими, что вообще для них не окажется предусмотренными никакие участки генома и тогда организм не будет развиваться.
Важно понять, что сами по себе гены не определяют то, что из них развивается, а буквально на всех стадиях влияют окружающие условия. Это важно для представления о том, что связи так же не образуются только по программе генетического кода. В первую очередь это влияет на то, как и куда прорастают аксоны (fornit.ru/5301) нейронов в зависимости от того, где эти нейроны начали развиваться. А проросшие нейроны могут образовывать связи, если аксон оказывается возле дендрита другого нейрона, когда совпадают их активности.
Таким образом, в одних условиях связи могут оказаться существенно иными, чем в других: если эти условия распознаются геномом, то такая связь образуется, если нет, то она не реализуется даже на уровне прорастания аксонов.
Это означает, что даже базовые, простейшие связи в нейронной сети могут существенно различаться в зависимости от специфики условий, в которых развивается организм, даже если первоначальные геномы были идентичны (однояйцевые близнецы). И чем более усложняется нейросеть, образуя на основе первых базовых связей более сложные, тем различия будут все более нарастать. Но в матке условия развития плодов одинаковы, как и, обычно, в первые годы жизни, так что и безусловные рефлексы возникают одни и те же. Геном вида предусматривает, что практически во всех матках условия достаточно схожи для реализации жизнеспособного организма, и поэтому в норме у всех организмов безусловные рефлексы не сформируются только если возникают мутации в этих участках генома у родителей, или в организме матери возникают несовместимые с реализацией данного участка кода условия (болезнь, истощение, отравление и т.п.).
Есть данные (fornit.ru/18218) о формировании не только безусловных, но даже условных рефлексов лишь изменением условий экспрессии генов.
Этологические представления о том, что безусловные рефлексы определяются только генетически, породили статьи, показывающие, что даже молодые, неопытные обезьяны боятся предметов, похожих на змей, что детеныши сразу способны распознавать образы хищников и т.п.
“На сегодняшний день считается, что некоторые защитные реакции, например способность цыплят различать хищных и нехищных птиц, запрограммированы генетически, в то время как большинство вырабатывается в течение жизни. Самым малоизученным остается элемент воспитания – передачи знаний от знакомых с хищником особей”. (fornit.ru/5375)
Формирование представлений о предпочтительном партнере: “Девушки могут не признавать, что их выбор партнера зависит от отношений с отцом, но новое исследование доказало правдивость этой теории”. (fornit.ru/5390)
Язык формирует восприятие цвета: “Наши слова буквально — форма категоризации того, что мы наблюдаем в окружающем нас мире”. (fornit.ru/5238)
Еще об этом: “Развитие речи совершенно неожиданным образом сказывается на умении ребёнка различать цвета.”. (fornit.ru/5391)
О формировании эмоций: “способность радоваться общению с близким человеком, по-видимому, формируется в первые месяцы жизни. Малыши, лишенные в течение этого критического периода самого главного — контакта с родителями, — могут на всю жизнь остаться эмоционально обедненными, им будет трудно адаптироваться в обществе и создать полноценную семью”. (fornit.ru/5270)
Для строительства гнезда птицы используют приобретаемый опыт: “Если птицы строят свои гнёзда в соответствии с генетическим шаблоном, должно ожидать, что все птахи данного вида создают обиталища каждый раз одинаково. Но перед нами не тот случай”. (fornit.ru/5132)
Это – очень сильный довод: в самом деле, если реакция полностью определяется наследственно, то она должна быть строго однообразной. Но этого не наблюдается во многих случаях, описанных этологами.
В самом деле, в геноме не запрограммировано никакой реакции (на такую детализацию не хватит никакой емкости памяти генома), а запрограммирован только момент активации фрагмента генома в случае определенной химии окружающей среды. Реакция же затрагивает множество нейронных связей, управляющих мышцами в определенной последовательности в зависимости от воспринимаемого. Это – даже не простейшие виды связей, а целые цепочки, возникающие на основе имеющихся наборов простейших связей, и такие цепочки формируются так же в зависимости от наличия определенных условий: реагирования не по сигналам первичных рецепторов, а уже сложных образов, состоящих из иерархии последовательного усложнения примитивов восприятия. Стоит не сформироваться каким-то примитивам восприятия (например, при депривации вертикальных линий: fornit.ru/5153, fornit.ru/43629) и цепочка реакции, даже предопределенной ветвлением аксонов, не сформируется.
Гены лишь предопределяют, дают (не)возможность возникнуть связям в мозге, но возникнет ли они в реальности, зависит от условий, например: Развитие мозга рыб почти не зависит от генов (fornit.ru/5171).
Инстинкты оказались не всегда бывают полностью врожденными: “нейронные контуры, отвечающие за распознавание полов, не совсем врожденные. Они не могут сформироваться без социального опыта, то есть без общения с самками. Это неожиданное открытие, поскольку «нейроны распознавания пола» расположены в очень древней области мозга.”. (fornit.ru/7628)
Каждый довод по ссылкам сам по себе, конечно, не обеспечивает полную уверенность, но в общем сопоставлении и в контексте сказанного они образуют надежный каркас представлений. Главное, - возникает очевидность причинно-следственной картины в явлении формирования природной нейросети.
Если бы не выделялись примитивы восприятия, то картинка на основе поля изображения сетчатки глаза была бы очень мало информативна. Вот у насекомых грубо фасеточные глаза потому, что нет смысла в большей детализации при такой низкой информативности. Быстро меняется освещенность – значит что-то явно приближается, нужно резко прятаться. Меняется пятно освещенности по полю зрения – что-то летит, ползет неподалеку, верхняя часть потемнела – помеха сверху, кто-то бьет по мухе мухобойкой и т.п. Никаких линий, кругов, не говоря про более сложные образы.
Но даже первый слой примитивов дает новое качество разнообразия отслеживаемых форм, на которые можно уже гораздо избирательнее реагировать. И чем больше слоев, тем большее разнообразие. Вот почему каждый слой распознавателей можно считать новым качеством рецепторов, как если бы кроме сетчатки глаза появились дополнительные рецепторы сложных образов.
Итого: на основе первичных рецепторов и конечных эффекторов в ходе развития возникают сначала непосредственные связи между ними, а потом и опосредованные, позволяющие, с одной стороны, выделить характерные сочетания элементов первичных рецепторов распознавателями таких сочетаний, а, с другой стороны, сформировать нейроны, управляющие сочетаниями первичных эффекторов.
Распознаватели сочетаний рецепторов принимают на себя роль специализированных рецепторов образа данного сочетания, а со стороны эффекторов, распознаватели необходимости сочетаний эффекторов принимают роль специализированных эффекторов.
Распознаватель необходимости действия – нейрон в ипостаси эффектора. Тот же нейрон в ипостаси рецептора распознает сочетание активностей, при которых нужно запускать действие. Даже мышечную клетку можно назвать распознавателем необходимости сокращаться, когда на нее придет возбуждающий сигнал. Так что концепцию распознавателей мы будем применять во всей нейросети без исключения.
Со стороны рецепторов формируются все более сложные распознаватели, использующие сочетания активностей предыдущих распознавателей, а со стороны эффекторов точно так же последовательно, слой за слоем развиваются все более сложные распознаватели необходимости действия.
Между теми и другими на всех уровнях возможны связи, образующиеся при условии их возникновения: одновременно активности по обе стороны установленного аксоном синапса. И так далее со стороны рецепторов и со стороны эффекторов, это – самая общая картина.
Любой уровень развития мозга следует описанному принципу, различаясь временем созревания и состоянием предшествующих слоев с учетом наследственно предопределенных особенностей. Развитие мозга в этом смысле автоматично. И это придает мозгу особую устойчивость за счет некоторой избыточности. Остановимся на этом явлении подробнее потому, что оно не характерно для привычной инженерам схемотехники, где функциональная цепь отрабатывает без дублирования, и при ее повреждении устройство перестает работать.
При формировании текущего уровня распознавателей отрабатывают многие соседние нейроны, а не один какой-то, что приводит к тому, что даже в случае предъявления элементарного признака в восприятии наблюдается (с помощью методов визуализации активностей) не точечная активность одного нейрона, а целое пятно активности, легко замечаемое при наблюдении.
При повреждении каких-то отдельных элементов такой группы распознавателей схожей функции, остаются другие, что практически никак не сказывается на результате: все равно остаются те распознаватели, которые продолжают обслуживать последующие слои. Причем эти последующие не были задействованы именно на основе отдельных предыдущих нейронов, а продолжают срабатывать, несмотря на гибель отдельных клеток предыдущих, т.е. не происходит исключение из работы мозга всех последующих цепей, работающих за счет погибших предыдущих.
Механизм такого “размывания” роли отдельных нейронов, участвующих в специализации последующих, заключается в том, что входы каждого последующего распознавателя перекрывают достаточно большую область предыдущих (что уже рассматривалось ранее).
Такая система резервирования образуется на всех уровнях развития мозга так, что, скажем, рефлекс определяется не одной какой-то цепью реагирования, а сразу многими схожими по функциям цепочками и поэтому не утрачивается в случае повреждения отдельных нейронов. Любая часть мозга характеризуется не точечной активностью, а пятнами групповой активности, наблюдаемых методами визуализации.
Это приводит к тому, что в случае нарастания патологического процесса в мозге и повреждения все большего числа нейронов, эффект долго не замечается, пока не будут критически исчерпаны группы схожего реагирования, приводя ко все более заметной недостаточности.
В дальнейшем механизмы мозга будут продолжать описываться здесь в стиле не резервирующей схемотехники (иначе было бы очень сложно рассматривать отдельные причины и следствия), но никогда не стоит забывать о групповой специализации буквального всего в мозгу.
Процесс строго поочередного развития слоев приводит к появлению все более усложненных, узко специализированных распознавателей на основе более простых примитивов (см. анимацию: fornit.ru/an-book-3). Как со стороны рецепторов, так и со стороны эффекторов созревают слои нейронов и формируются иерархии специализированных распознавателей.
При этом возникают связи на всех уровнях от рецепторных к эффекторным элементам. Многие такие связи остаются действующими даже в очень сложных организмах, обеспечивая основу возникновения еще более сложных связей.
Цепочки последовательности действий – очень распространены в нейросети, они играют определяюще важную роль в адаптивных механизмах и поэтому мы подробнее рассмотрим их схемотехнику.
При усложнении (обусловленном генетически как находки эволюции вида), на основе более простых связей образуются цепочки, в которых последующее звено срабатывает при условии активности предыдущего и сигнале рецептора об окончании действия предыдущего звена (фазы работы цепочки). Так организуется последовательность элементарных действий в зависимости от актуальности срабатывания каждого звена, которое распознает момент актуальности свей активности.
Звеном такой цепочки может быть как элементарное действие (мышечное и любого другого эффектора, включая нейрон), так и начала других цепочек действия – ветвления основной цепочки.
Цепочка реакции запускается распознавателем актуальности ее работы – пусковым сигналом, и ее звенья начинают последовательно срабатывать.
В схемотехнике такие цепочки последовательного срабатывания звеньев называются регистром сдвига (сдвиговым регистром).
Вот типичная схема:
Каждый из элементов 1-4 это – триггер, т.е. устройство, которое переключает сигнал на выходе в одно из устойчивых состояний: 1 или 0. Чтобы на выходе триггера 1 появился сигнал 1 (высокий потенциал), нужно, чтобы на его входе появился пусковой сигнал 1 и нужно, чтобы на другом, синхронизирующем входе возник перепад с 0 в 1 (т.е. триггер изменяет состояние на выходе в момент перепада (по фронту перепада, а не по потенциалу).
Пока нет пускового сигнала, то импульсы синхронизации никак не влияют на выход триггера. Но как только появился пусковой потенциал, то с первым же перепадом синхроимпульса на выходе 1 с некоторой небольшой задержкой появится 1.
Но когда 1 появился на входе 2, то перепад синхроимпульса уже прошел и 2 не сработает. Зато второй перепад его включит. И последовательно откроются все триггера по цепочке.
Нейроны не срабатывают по фронту, хотя такое действие и можно было бы осуществить в принципе схемой из четырех нейронов, но это привносит нестабильность потому, что нужен не только фронт, но задержка чуть большая, чем фронт. Даже на уровне транзисторных схем такие “гонки” могут вызвать сбои при повышенном уровне помех или нестабильности задержки, а в условиях природной нейросети это – заведомо неприемлемый вариант. Все организуется очень надежно, напрямую.
Представим вариант организации регистра сдвига без фронтов и задержек:
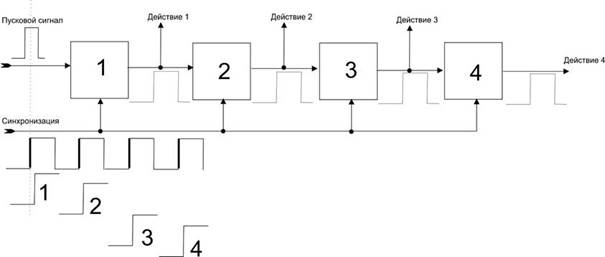
Опытные схемотехники сразу заметят, что после активации первого нейрона, тут же последует активация всех остальных, т.е. схема выполняет роль моментального просмотра всех действий. Более правильную схему мы рассмотрим ниже.
В цепочке стоят “нейроны”, для запуска которых нужно два сигнала: пусковой стимул (fornit.ru/5286) и условие, разрешающее работу данного звена. Можно это организовать через веса входов каждый по 0,5 от максимума, но это вряд ли так делается (вообще природные синапсы слабо градуированы по коэффициенту пропускания, обычно они или проводящие или нет, хотя при формировании профилей активности некоторые синапсы могут и не успеть набрать высокую проводимость за критический период обучения, что и скажется на функции распознавания, и еще синапсы различаются спецификой нейромедиатора). Так что результат распознавания в природной нейросети будет зависеть как от сформированных проводимостей, так и от частот поступающих через них сигналов (или частот предъявления стимула для данной связи), что так же определяет данный профиль образа восприятия (пример ВПСП и ТПСП – градуальных сигналов, где ответ клетки зависит от скорости нарастания сигнала). А для системного аналога искусственной нейросети достаточно было бы только нескольких возможных уровней градации проводимости синапса и постоянного тока на выходе нейрона при его возбуждении, без импульсной активности.
Будет надежнее условие срабатывания сделать тормозным тогда, когда срабатывание не нужно. Т.е. вход “условие срабатывания” является тормозным, пока нет условий для запуска цепочки, что может обеспечивать латеральное торможение. К первому нейрону подключен еще один тормоз Т, но он вначале неактивен и тормозит только когда успешно выполнится первое действие.
Итак, элемент 1 сработает, когда есть условия для такой реакции и когда наступает актуальность реагирования в этих условиях.
По его сигналу совершается действие 1. Около эффектора действия 1 есть рецептор, откликающийся на успешность совершения этого действия (такие рецепторы называют проприорецепторами (fornit.ru/5263), сигнал которого создает условия для срабатывания элемента 2 и через тормозной нейрон закрывает элемент 1. И далее – по такой схеме работы.
На самом деле, как правило, такие цепочки содержат больше элементов, которые могут организовывать процесс очень разными способами с ветвлениями в зависимости от условий, но это сейчас не принципиально для модели понимания явления.
Вот рабочая схема правильной цепочки последовательных действия, которую можно по исследовать в деле с помощью эмулятора fornit.ru/demo1
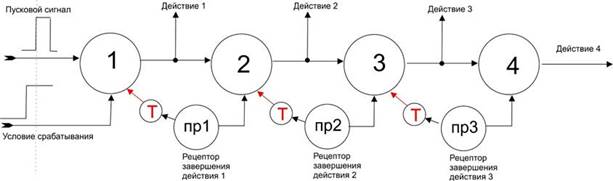
При пуске активность сразу на короткое время проскакивает до конца: "Опережающее возбуждение" по И.Павлову. Это происходит из-за задержки блокировки нейронами “Т”. Мышцы не успевают отработать, хотя могут слегка дернуться (на них регистрируются короткие мио-потенциалы).
Именно на таком принципе организованы цепочки последовательностей фаз мышечного сокращения, но вариантов реализации может быть бесконечное множество. Сигналы при этом могут корректироваться на отдельных стадиях процесса запуска цепочки. Часть мозга, называемая мозжечок, специализируется на таких корректировках, причем не только мышечных, он накапливает опыт корректировок (fornit.ru/7553) последствий различных неточностей и иллюзий.
Для такой работы нужны рецепторы ошибок. Но что можно посчитать ошибкой? Кто или что может быть судьей правильности или неправильности реакции? Это возможно только после совершения действий, когда организм этим ухудшит или улучшит свое состояние, и датчики, регистрирующие это, и есть рецепторы ошибок и их исправления.
Структура цепочек реагирования эволюционирует с приобретением все новых адаптирующих механизмов, приобретая все новые свойства и функции, вплоть до организации цепочек последовательных действий и возможность прогноза, см. анимацию (fornit.ru/an-book-19).
Базовый механизм индивидуального поддержания жизнеспособности
Назрел момент, чтобы заметить самое главное в организации индивидуальной адаптивности. И начнем с проясняющего очередной принцип отвлечения.
В природе нет ни одного одинакового нейрона так же, как нет ни одной одинаковой снежинки. И эти снежинки в зависимости от температуры, ветра, влажности и т.п. приобретают различимо разные виды форм. Одно дело – изучать разнообразие этих форм, а другое – общие закономерности их формирования. Если оставаться на уровне первого, то никогда не получится понять второе из-за невероятной сложности отдельных компонентов и разнообразия их взаимодействия.
Представим, что марсиане перехватили транспортный корабль, перевозящий реквизиты музея истории часов. Там, каждый экспонат хранился в одинаковых ящичках, но в каждом ящичке - очень разные часы, от древних ювелирных, инкрустированных каменьями, до современных изысков из драгоценного дерева, огромное разнообразие механических и электронных из разных эпох.
Это было ни на что не похоже, и ценность находки не вызывала сомнения: наверняка здесь скрывалась величайшая тайна чужой цивилизации, сулящая наконец-то понять, где и как правильно нести яйца. С присущей тщательностью марсианской науки ученые принялись дотошно изучать часы вплоть до молекулярного уровня, но никаких корреляций не обнаруживались: все часы были совершенно разными даже по составу вещества.
И вот, один ученый собрал данные внешне наблюдаемых изменений во времени и ввел их в суперкомпьютер, чтобы выявились похожести этих параметров. Получилось две очень выраженные, вездесущие корреляции изменений движений (для минут и часов): одна более слабая выпадала из общей картины (у тех часов, где не было секундной индикации) и остальные совсем слабые (недели, месяцы, годы).
И тут возникла идея соотнести данные по параметру временной повторяемости. Оказалось, что все виды выявленных периодов с убедительной точностью не только относятся к одним и тем же интервалам времени, но и более длительные из них кратны целым числам более быстрых параметров.
- Да это же просто как наши тикалки! только дико примитивные какие-то... - с отвращением проскрипел жвалами марсианин, не веря в такую простоту, и великая тайна растаяла бесследно. Потом он задумался о том, как написать итоговую работу так, чтобы его не высмеяли за столь вульгарную интерпретацию великой проблемы.
Если не абстрагировать наиболее общее в явлениях, то не получится создать общую теорию.
Когда-нибудь системология (и, в частности, системный анализ) обретет формы математического метода, на который каждый сможет опереться. Но сегодня этого нет, так что нужно просто внедрить этот принцип как общий контекст понимания: в массе вариаций схожих явлений есть нечто наиболее общее, что придает модели этих явлений некий базовый каркас. Это то, что сформулировано в Критериях полноты и верности теории (fornit.ru/7649), и то, что очень нужно нам в текущем построении модели адаптивных механизмов нейросети.
На данном этапе обобщающего рассмотрения – очень много ключевых представлений, от которых зависит основа понимания всего остального. Эти представления были получены не просто при обобщении существующего массива фактических данных исследований, но, главное, из более общей картины каркаса модели совершенно бесспорных (эмпирически воспроизводимых) явлений, которые выделяются исследователями еще с середины прошлого века: безусловные, условные рефлексы, детали гомеостаза и т.п. (по тексту это будет видно).
Каждая конкретная природная реализация демонстрирует огромное разнообразие воплощений и механизмов одного и того же явления, причем это разнообразие не только по видам животных. но и индивидуальное (так, у человека >50 видов гемоглобина с одной системной функцией переносчика кислорода, и вообще белки одного человека не совместимы с белками другого).
Буквально на любом уровне изучения организма физиологи озадачиваются этим разнообразием, не говоря про то, как странно и сложно бывает организованы сами механизмы. Видов синапсов – очень много и они очень разнообразны, а мы говорили только про принцип синаптической передачи тока носителей сигнала. Видов нейронов – очень много, а мы построили одну общую модель, которая отражает наиболее общую функциональность любых видов нейронов. Есть несинаптические случаи передачи возбуждения, которые мы вообще не рассматривали. Да и сам природный нейрон – невероятно неопределенная конструкция, в которой даже отдельные его дендриты способны начинать самостоятельно генерировать потенциалы действия, что, впрочем, на них и заканчивается, если только не будет достигнуто условие генерации самого нейрона.
Природа – великий и бездумный рукожоп…
Приношу извинения тем, кому эта аллегория покажется неуместной теме и духу книги. Да, это – признак смелости, такой же как смелость, нужная для того, чтобы выйти на улицу с вызывающей татуировкой наискось своего высокого лба. Но это ставшее полноправным очень емкое слово-мем отражает самую суть. Это – как слова в песне, которые не выкинешь. Разумный человек сможет это понять.
Исследователи строения глаза недоумевают о том, насколько нелогично, странно и нерационально устроена как оптика глаза, так и его чувствительная часть. Вот в фотоаппаратах все логично и правильно, там идеально и нет проблем, которые нейросети приходится решать, приспосабливаясь к столь капризному и неточному инструменту восприятия. Все можно сделать гораздо проще, точнее и лучше даже если использовать те же биоматериалы. И так – буквально во всех творениях природы. Мало того, все это постоянно кишит разными вариациями в каждой отдельной реализации. Зато глаз в воде продолжает функционировать, а фотоаппарат требует гидробокс. Если в глаз попала соринка, он проморгается, а соринку на матрице может вытащить только специалист и то не всегда без последствий. Видеокамеру связали шнурком с компьютером, тот запоминает все происходящее, но само по себе это - совершенно бессмысленно и только человек может разобрать этот смысл, а вот кошка уже – нет, если только по экрану не начнут бегать мышки.
Чтобы построить системную модель принципов усложнения механизмов адаптивности, просто необходимо суметь абстрагироваться от всех вариантов воплощения этих механизмов. И такая картина может оказаться не понимаема теми, кто досконально изучает тонкости природной реализации. Они будут читать и скептически пожимать плечами: слишком все вульгарно описано, ну нельзя же все так упрощать. Но ведь и среди этих специалистов однозначно всеми принятых мнений по интерпретации данных исследований нет, а есть только многообразие представлений и растерянность в не поддающихся их пониманию теориях мозга.
Системную модель понимания строить нужно и, главное, возможно, другого пути нет, если мы не хотим утонуть в неисчислимых загадках воплощения очередного примера природного безрассудства. Нам важна ясная система, но не выдуманная из ничего, а в которую четко и непротиворечиво укладываются все известные фактические явления организации адаптивных механизмов. А проявления таких механизмов можно подсмотреть только в природной реализации.
Пример: природный персептрон поражает своей прямолинейной и грубой логикой. Он никак не может соревноваться с точностью распознавания искусственного многослойного персептрона. Но логика идеального персептрона приводит к тупику и получению хотя и очень качественного распознавателя хоть чего, но этим все и ограничивается.
Мы в системной модели понимания принимаем эту обобщенную природную логику, но не во всем ее разнообразии воплощений, а лишь в самом общем принципе реализации, который обеспечивает все последующие принципы своей основой. Этот принцип выверен из сопоставлений всех известных примеров природной реализации. Он не способен на эволюцию для каких-то невероятно новых условий, как все еще способен природный, но нам этого и не нужно.
Другой пример. Когда природа “изобрела” нейромедиаторы в простых животных, то еще не было задачи определять ими стили поведения, они просто встраивались случайно и бессистемно, но в результате очень хорошо отделились реакции разного вида, не мешая одна другой в общей схеме. Потом это привело к очень естественному способу переключения стилей поведения и к сопутствующей гормональной регуляции этих стилей.
Скрупулезному исследователю роли нейромедиаторов очень непросто сделать вывод о такой их функциональности потому, что он видит множество примеров совершенно иного использования, не укладывающегося в такие представления, это навевает свои интересные выводы, требует учесть это в общей картине, и с этим нужно что-то делать, как-то объяснять.
Говорят, что если бесконечно большой стаи обезьян дать пишущие машинки чтобы эти обезьяны все время клацали по буквам, печатая бессмысленный и случайный текст, то у большинства получится просто галиматья, но у единиц выйдут произведения великих писателей. Это верно для бесконечности обезьян и/или бесконечности времени. Но в природе хоть и очень много времени и очень много экспериментаторов, но нет таких бесконечностей для того, чтобы случайно создать самолет или человека за один раз. Но природа так и не работает.
Пусть после того, как каждая обезьяна напечатает очередное слово, оно проверяется на осмысленность богом. Если с новым словом текст не потеряет разумность, то он остается жить и развиваться далее. Если слово окажется неуместным, оно удаляется как вредная мутация.
И тогда за какое-то не бесконечное время будут напечатано множество разных вариантов прозы и каждый из вариантов будет продолжать развиваться, увеличивая число разных разумных (с точки зрения бога) книг.
У природы таким богом выступает смерть, которая не пропускает нежизнеспособное.
С каждой проверкой на разумность после напечатанного слова, вероятность вставки нового разумного слова имеет вполне состоятельную вероятность. Каждый раз эта вероятность реализуешься заново. У множества обезьян начнет формироваться множество вариантов разумного текста.
У природы таким богом выступает смерть, которая не пропускает нежизнеспособное. И уже совсем не нужно бесконечности, чтобы создать человека. Вот так природа и работает: как тупые бессмысленные обезьяны. И это стоит постоянно иметь ввиду. Вы можете сами попробовать эволюцию с помощью программы: fornit.ru/evolution.
Вот почему обращаю особое внимание: при чтении нужно стараться выделять именно такие обобщенные принципы и механизмы, и в таком смысле рассматривать все огромное разнообразие особенностей реализации в публикациях фактических данных исследований. Это делать необходимо потому, что конечная модель должна возникать именно в вашей голове, и она может оказаться в чем-то отличной от той, что сложилась в другой.
Пожалуйста, проникнитесь принципами обобщающей системологии и специально следите за тем, чтобы вовремя видеть несистемные факторы в явлениях, которые мы рассматриваем, когда опираемся на фактические данные природной реализации. Это – самое важное, это – очень непросто, но без этого не возникнет целостный и непротиворечивый каркас собственной модели понимания.
Формируем обобщающую модель адаптивной нейросети
В общей картине представлений о принципах развития нейросети важнейшим является роль рецепторов – датчиков информации для работы регулирующей нейронной схемы управления эффекторами, что, в конечном счете, позволяет поддерживать организм в жизнеспособном состоянии. Важнейшим потому, что если рецепторы молчат, то и остальному делать нечего, что убедительно демонстрируют опыты по депривации восприятия, см. fornit.ru/5244, fornit.ru/5348, fornit.ru/5393.
Рецепторные датчики не только обеспечивают восприятие внешнего мира, они возникли буквально во всем теле и обеспечивают жизненно важные механизмы регуляции в организме. Ранее упоминалась метасимпатическая нервная система, но этим далеко не ограничиваются управляющие контуры регуляции. И от самых простых взаимосвязей развиваются все более сложные, затрагивая уже имеющиеся связи головного мозга.
Везде, где только можно, после включения в геном такого варианта развития клетки как нейрон, он используется в качестве промежуточных распознавателей состояний предшествующих рецепторов (в том числе и предшествующих сформировавших специализацию нейронов) в организме. При этом последние для данного организма дополнения в геноме постоянно пытаются модифицироваться многими механизмами (мутация, половое смешение и др.) так, что в популяции возникает довольно широкий спектр генетического разнообразия. Последние эволюционные участки генома, способные к модификации генетическими механизмами разнообразия, – это примерно 1-10%, и чем сложнее организм, тем этот процент меньше. Более древняя, хорошо обкатанная жизнью часть генома остается неизменной, если только ее не меняют паразиты или поломки (например, радиация или мутагенные химические вещества). Если затрагивается древняя часть, то получаются монстры, как правило, мало приспособленные к жизни уроды. Вот почему из кота не может вдруг родиться удав, а виды, у которых геном не совпадает на достаточно раннем этапе формирования тела, не способны к появлению потомства при спаривании.
Это напоминает то, как не модифицируются уже специализированные слои развивающегося мозга. И здесь регулятор – смерть неудачного.
В результате во всем организме возникают более молодые варианты нейронной регуляции на основе уже устоявшихся и мало подверженных изменениям. Так продолжается эволюция, способная адаптировать организм к меняющимся условиям и при этом все более усложняющая общую систему регуляции.
Гомеостаз
Для определения границы допустимого в отклонении различных параметров жизнеспособности существуют внутренние рецепторы значений таких параметров, в отличие от рецепторов, параметров внешней среды. Таких параметров очень много.
Так, общие для всего организма параметры, которые поддерживаются в оптимальном состоянии (меняясь в зависимости от условий):
кислотность (крови, лимфы, межклеточной жидкости, желудка),
концентрация углекислого газа в крови,
концентрация кислорода в крови,
концентрация глюкозы в крови,
концентрация ионов натрия в крови,
концентрация ионов калия в крови,
концентрация ионов кальция в крови,
давление в гидравлике кровообращения,
осмотическое давление крови,
частота сердечных сокращений,
температура крови,
и др.
Изменение любого из этих параметров в любую сторону резко влияет на функциональность организма. Это прямо означает необходимость эффекторов, которые регулируют эти параметры и необходимость наличия распознавания оптимального значения параметров и направления выхода их из оптимума (превышение или недостаток). Эти распознаватели должны учитывать, что в разных режимах работы организма нужен свой оптимум. А такие режимы переключаются в зависимости от стиля реагирования организма (пассивный, высокая физическая нагрузка, половое поведение, защитная пассивность, сон и т.п.), а также в зависимости от внешних условий: холод, жара, обезвоживание, голод, проблемы с воздухом и т.п.
Обкатанные жизнью системы регуляции основных параметров оказываются тем сложнее, чем для большего числа возможных стилей поведения и числа различных внешних условий и ситуаций они наработаны.
Понятно, что самые простейшие связи типа понизилась температура – дать больший сигнал на ее повышение, как в термостатах, это – лишь первый уровень такой регуляции, на который наслаиваются более сложные, учитывающие ту или иную специфику. И таких слоев оказывается немало даже еще до условно-рефлекторного уровня регуляции, до уровня развития ассоциативной теменной коры мозга, где формируются условные рефлексы.
Гомеостатическая значимость начинает влиять на уровне вторичных зон новой коры, образуя то, что должно восприниматься в данном контексте текущего стиля поведения. И именно на этом уровне становится возможным зацикливание образа для его удержания с помощью цепочки (цикл А.Иваницкого): образ восприятия - ассоциативные зоны с выходом на действие - оцененная значимость результата этого действия - распознаватель гомеостатической значимости и от него - ко входу образа на уровне вторичной зоны.
Точно так же, как в новой коре возникают все более сложные слои распознавателей разных видов внешней рецепции, каждый из которых имеет свой уровень связей с эффекторами, обеспечивающий свою сложность реагирования, так же происходит и с внутренними рецепторами отклонения параметров гомеостаза и возврата их в норму.
Принципы поддержания оптимального уровня параметра
У простых термостатов (устройств для поддержания постоянной температуры в среде) есть некоторый гистерезис: включение нагревателя вызывает перегрев, пока на это не отреагирует датчик, потому как у нагревателя есть инерция (он не может сразу нагреться и сразу остыть). Поэтому температура в таких термостатах постоянно колеблется, т.е. возникает генерация колебаний температуры. И чем больше инерция нагревателя или других элементов отрицательной обратной связи, тем больше будет амплитуда колебаний из-за того, что отрицательная связь все меньше попадает в нужное время и вместо противодействия возникает содействие. Т.е. при сдвиге фазы (момента начала) выходного сигнала из-за задержки по отношению ко входному отрицательная обратная связь становится все более положительной. Если задержка окажется настолько большой, что вместо отрицательной обратной связи возникнет положительная (т.е. момент воздействия окажется противоположным тому, что нужно бы для компенсации), то колебания станут максимальными по амплитуде вплоть до достижения предельного режима переключений: с одного предельного состояния в другое.
Но если сделать некий сглаживающий фильтр (распространенное схемотехническое понятие), например - на выходе термометра, который не позволяет быстро меняться показаниям, а отслеживает некий средний уровень по времени, заведомо больший, чем период колебаний температуры. Это можно сделать простейшей цепочкой из резистора и подключенного к нему заземленного конденсатора, и если сглаживать он будет все, что имеет время изменения меньше, чем произведение сопротивления на емкость (- “постоянная времени” цепочки), то колебания прекратятся и установится равновесное состояние на заданной температуре.
Сейчас было описано наиболее общее условие генерации колебаний в системах с обратной связью, в независимости от природы колебаний.
В организме оно также работает. Например, нужно удержать руку в каком-то положении, делается усилие поднять ее. Это усилие зафиксируют рецепторы положения руки (или зрительные рецепторы через все системы мозга, распознающие это положение) и когда рука достигнет нужной высоты, дается команда на уменьшение усилий до уровня сохранения нужной высоты. Но если есть какие-то задержки в реагировании, то момент нужного уровня будет пропущен и нужно будет опускать руку. Возникнет дрожание (тремор). При сильном волнении, когда на слежение за плавностью движения привлекается максимум внимания и движение контролируется буквально на всех стадиях, часто возникает тремор потому, что осмысление требует времени и возникает задержка. Это может закрепиться в автоматизмах. Или тремор может возникнуть из-за патологии каких-то промежуточных стадий, например, прекращения действия некоторых рецепторов положения руки и тогда момент нужного положения будет пропущен.
В быту люди с повышенным давлением склонны принимать сбивающий давление препарат, когда давление поднимается. А это бывает при стрессе, физической нагрузке и т.п. ситуациях, когда организму нужно поднять давление для обеспечения функциональности. С препаратом давление довольно быстро снижается и даже намного ниже, чем нужно. Возникает болезненная вялость, сонливость, нарушения координации, в общем, давления очень не хватает. Тогда гипертоник принимает препарат для повышения давления (пьет пантокрин, кофе и т.п.). Давление снова подскакивает. И т.п.
Если же не торопиться с решением противодействовать, а выждать время, то вообще бы не потребовалось принимать препараты потому, как последствия стресса довольно скоро сами бы закончились.
Бывает, при сильной жажде так хочется пить, что выпивается огромное количество воды. Сигналы о восстановлении водного дефицита возникают намного позднее, когда на борт заливается слишком много воды. Это может нанести определенный вред (как с лекарствами от давления) и это стоит учитывать, не торопясь выпить побольше до того, как жажда не исчезнет.
Во всех случаях подобного восполнения жизненных параметров стоит помнить об этом и не переедать до вредных последствий, не перепивать, не переохлаждаться в жару и т.д. Подробнее о гоместатической регуляции: fornit.ru/324
Гомеостаз органов
Кроме поддержания стабильности внутренней среды, физиологи различают две системы регуляции параметров отдельных органов и систем: симпатическая и парасимпатическая системы с взаимно противоположными видами воздействия на параметр, обеспечивающими его необходимый уровень поддержания.
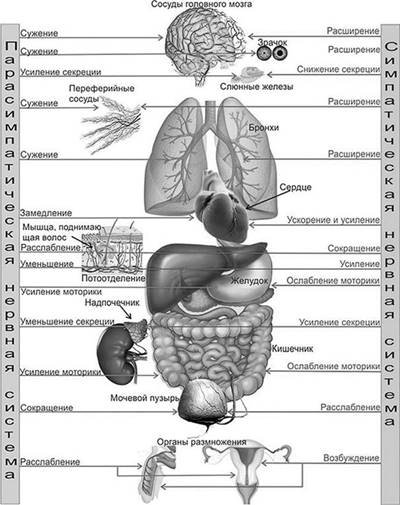
Такой вид регуляции на основе обратной связи с эффекторами противоположного действия применятся везде в организме, где необходимо иметь возможность быстро менять значение параметра. Это – мышцы-антагонисты, регуляция состояния органов и внутренней среды. К этому вынуждает необходимость адаптации.
Отклонение жизненных параметров организма и отдельных органов опасно, и распознавание таких ситуаций важно для того, чтобы их избегать. Поэтому кроме распознавателей отклонения жизненных параметров, которые используются в контурах регулирования параметров, эти же распознаватели (fornit.ru/5088) на последующем слое развития нейронов в этой области могут использоваться как рецепторы опасного состояния, а также рецепторы восстановления после опасности.
Здесь так же, как в гомеостатических эффекторах-антагонистах, нужно при возникновении опасности активировать защиту от нее, но только до тех пор, пока опасность не минует, чтобы прекратить эту защитную реакцию. Восстановление параметров сигнализирует о таком моменте.
Понятно, что распознаватели опасных состояний организма важнее, чем распознаватели восстановления (они первичны в логике происходящего), и они представительствуют в мозге гораздо больше потому, что во многих случаях обратной связи в такой регуляции не требуется и после возврата в норму ничего не нужно совершать. Ощущения “нам плохо” – важнее ощущений “нам хорошо” вопреки распространенным среди гуру самосовершенствования мнений.
Это также касается всех случаев реакций избегания опасности, когда распознаются ситуации, могущие навредить на основе предыдущего опыта. Для таких реакций рецепторами являются слои распознавателей опасности, которые, в свою очередь, используют в качестве рецепторов слои распознавателей отклонения гомеостаза организмов и органов.
Система значимости
Функцией распознавания значимости не обладают сами по себе распознаватели всех других сенсорных систем: никакие образы восприятия сами по себе ничего не означают для организма в последствиях их появления. Но, сочетаясь в общий образ (профиль возбуждения детектора образа от различных сенсорных систем), где присутствуют как рецепторы различных видов рецепции и рецепторы значимости, так и начальные эффекторы цепочек ответных реакций, возникают связанные между собой (ассоциированные) образы, обладающие значимостью того, что данный образ в данных условиях означает для сопутствующего состояния организма. Это происходит, начиная со вторичных зон и, особенно конкретно (специфично к какому-то распознанному образу) - в области формирования условных рефлексов, в теменной ассоциативной коре мозга, где сходятся все виды рецепции, включая рецепцию состояния гомеостаза.
Мы будем называть такие образы в ассоциативной теменной коре образами восприятия-значимости-действия или просто образами, а систему распознавателей опасного отклонения параметров гомеостаза и восстановления их – системой гомеостатической значимости. В дальнейшем она будет дополняться более высокоуровневыми механизмами распознавания значимости происходящего для организма, образуя то, что называют смыслом происходящего для субъекта.
Стоит заметить, что система значимости и система регуляции гомеостаза, хотя и основывается на одних базовых распознавателях и первичных рецепторах состояния параметров гомеостаза, но это – две разные системы с разными функциями. Это должно быть очевидным из уже сказанного.
Значимость образов позволяет организовывать реагирование на внешние условия и раздражители с учетом того, как это отражается на жизненном состоянии организма.
Именно это в ассоциативных зонах играет роль “учителя” в том, как оценивается верность распознавания ситуации в данных условиях, но здесь не происходит формирование распознавателей с коррекцией от учителя, а возникает лишь возможность такой коррекции в более сложных механизмах.
Эта роль значимости организуется совсем не так, как в искусственных нейросетях, хотя там уже и начали использовать искусственную систему подкрепления, но она не отражает собственное состояние сети и параметры ее существования и поэтому не является для нее адаптивной в этом смысле.
Дополним таблицу различий природной и искусственной нейросетей:
| Искусственная нейросеть | Природная нейросеть мозга |
| Критерием верности функции распознавания являются привнесенные оператором условия, а не состояние самой сети. Во всей сети все ее элементы корректируются в соответствии с этими конечными условиями. | Критерием верности распознавания значимости образов являются сигналы специализированных рецепторов состояния гомеостаза и их интерпретация более высокоуровневыми рецепторами системы значимости. До ассоциативной зоны распознаватели формируются без учителя. |
Это – последнее такое различие потому, что далее, в сторону усложнения механизмов адаптации, уже не будет вообще никаких возможных сопоставлений.
С момента использования распознавателей значимости для возможностей определения направленности реагирования схема приобретает новое качество индивидуальной адаптивности по сравнению с адаптивностью, определяемой генетическим кодом потому, что начинает учитываться текущее состояние организма, что позволяет организовывать связи между теми условиями, которые оказывают влияние на это состояние и реакциями, нормализующими это состояние.
Конечно, уже и генетический код дает возможность учитывать условия и формировать нейросеть в зависимости от того, были или нет условия, вызывающие ту или иную экспрессию генов, но теперь изменения реагирования происходят не из поколения в поколение, регулируемые смертью неудачных форм, а с учетом текущей ситуации.
Гомеостатическая значимость формируется так же, как и другие первичные и вторичные зоны мозга, образуя в новой коре все более сложные примитивы того, что для организма хорошо, а что – плохо.
В третичной (ассоциативной) зоне, где сходятся все выходы разных сенсорных каналов распознавателей, возникают уже сложные образы, составленные из многих видов рецепторного восприятия.
Образы оказываются ассоциированы, в первую очередь, с отрицательным состоянием организма (опасный выход каких-то параметров гомеостаза из нормы), или ассоциированы с положительным состоянием возврата из аварийного состояния жизненных параметров в норму. По логике ясно, что не может быть возврата в норму до того, как параметр вышел из нормы. Так что состояние негативной значимости всегда первично.
Однако, возбудить распознаватель возврата в норму (т.е. детектор, что с организмом все хорошо) можно разными химическими и физическими воздействиями, и тогда организм получает ложный сигнал “хорошо” в то время, как нет никаких таких причин и обстоятельств, которым нужно было бы противодействовать. Именно так были первоначально открыты точки мозга (fornit.ru/5374), где сконцентрированы распознаватели “хорошо” и “плохо”. Но стоит иметь ввиду, что гомеостатическая значимость вовсе не означает наличие субъективных переживаний, что возврат жизненных параметров в норму не является переживанием “хорошо”, каким-то кайфом при раздражении таких распознавателей. Кайф возникает только при осознании этого состояния, и для переживания необходимо иметь дополнительные механизмы осознания, до которых мы еще не добрались.
Эти участки были названы “эмоциональными центрами” потому, что их возбуждение приводит к переключению стиля поведения животного (в том числе человека), характерного для проявления какой-то эмоции. Но это – всего лишь тупо (неосознанно) – распознаватели отклонения и возврата в норму, а не “чувства”, и понимание этого различия – чрезвычайно важно.
Стоит ознакомиться с подборкой фактических данных исследований по теме системы значимости: fornit.ru/ax1-15
В плане эмоций и в самом деле, возбуждение распознавателей значимости может приводить к осознанному переживанию тех или иных градаций состояния хорошо и плохо. Но такие переживания возможны только у животных, обладающих способностью осознавать и субъективно переживать, а что это такое – более поздняя тема. У сложно организованных насекомых есть распознаватели состояния параметров гомеостаза, но нет механизмов субъективного осмысления, представленных совершенно определенными анатомическими образованиями, обеспечивающими высшие (по сравнению с рефлексами) механизмы индивидуальной адаптации к новому. Сейчас об этом преждевременно сказано для того, чтобы задать контекст понимания, что первичные связи и даже условные рефлексы – далеко не исчерпывают механизмы индивидуальной адаптации, а лишь представляют собой их базовую часть.
Самый важный принцип использования распознавателей значимости – возможность переключения вида реагирования в зависимости от условий и обстоятельств и формирование ответных реакций именно в этом контексте.
Можно выделить такие основные виды реагирования как пищевое поведение, половое поведение, защитное поведение (активная и пассивная защита), исследовательское поведение, сон. У каждого такого стиля должен быть инициирующий момент необходимости его активации (выход параметров гомеостаза из нормы) и стадия возврата гомеостаза в норму, что характеризует успешность варианта поведения (или не успешность, если параметр еще более отклонится от нормы).
Половое поведение инициируется самим организмом, это - особый случай, когда выход параметров из нормы организован как механизм выживания вида. Поэтому с половым поведением связана возможность его возбуждения, даже если гомеостаз в полном порядке и нет аварийного состояния, т.е. возможна его самоактивация (еще одно исключение – альтруизм, который так же есть у многих видов животных). Впрочем, есть возможность искусственной активации и любой другой потребности, скажем, воздержание от воды или еды для какой-то цели (тоже - атрибут осмысленности) или чтобы потом получить удовольствие от восполнения дефицита. Как в анекдоте, где медведь увидел, что заяц бьет себя кувалдой по гениталиям и вопит на весь лес, а на вопрос медведя нашел силы ответить, что, когда промахнется, это – неимоверный кайф.
Каждый базовый стиль имеет более сложные градации его особенностей, обеспечиваемых последующими слоями распознавателей значимости, конкретизирующими распознанную ситуацию. Это позволяет разделять контексты реагирования.
Контексты значимости
Базовые стили поведения переключаются очень быстро с помощью изменения баланса нейромедиаторов в мозге так, что те цепи, которые не рассчитаны на использование синапсов, работающих через такие нейромедиаторы, перестают работать, а действуют только специфичные к имеющимся нейромедиаторам связи (fornit.ru/an-book-4). Так выделяются целые программы ответных действий (fornit.ru/an-book-6).
В теле при этом одновременно выделяются специфичные вещества – гормоны, которые изменяют режимы работы органов, оптимально приспосабливая их к данному стилю поведения: если нужны предельные усилия, то повышается частота сердечных сокращений и их сила, расширяются сосуды, становится интенсивнее дыхание, высвобождаются энергетические вещества и т.п.
Нормы гомеостаза соответственно меняются, что учитывается распознавателями значимости.
Таким образом, формируется общий контекст восприятия и реакций организма, в котором оказывается возможным только присущий ему стиль поведения.
Базовые стили восприятия-поведения – механизмы, предопределяемые наследственно. Но число вариантов реальных ситуаций таково, что они не могут быть обеспечены ограниченным набором нейромедиаторов и требуют более оперативного изменения поведенческих реакций в условиях, не предусмотренных наследственно. Т.е. для большей гибкости реагирования необходим механизм, который бы мог более избирательно переключать контексты реагирования.
По схемотехнической логике это реализовать просто: вместо того, чтобы отключать питание у схем, обеспечивающих все стили реагирования кроме нужного (аналог нейромедиаторной регуляции), можно просто ввести блокировочные сигналы или, наоборот, сигналы необходимости возбуждения данного контекста для распознавателей ситуации. Это проиллюстрировано действующей моделью: fornit.ru/demo2.
В природе реализованы оба варианта: активация распознавателя данного контекста со стороны иерархии слоев значимости и торможение между конкурирующими соседними контекстами, которые активируются специфичными детекторами - распознавателями ситуации.
Получается такая картина: распознаватели жизненных потребностей (при отклонении параметров гомеостаза) активируют специализированный контекст, а сигналы рецепторов внешней ситуации определяют стимулы для активации тех или иных реакций в этом контексте. Каждый образ активируется во вполне определенных условиях окружения при наличии “пускового стимула”: fornit.ru/an-book-17
Новые связи могут образовываться только в активном контексте, но не в других. Научившись танцевать у себя дома, не получится сразу так же хорошо продемонстрировать это на балу.
Это приводит к тому, что образ одних и тех же признаков восприятия приобретает совершенно разную значимость (ассоциируясь с признаками значимости) в разных контекстах. Соответственно, реакции в разных контекстах оказываются неповторимо специфического стиля так, что животное как бы приобретает разные свойства, подчас противоположные, в разных своих состояниях. Люди могут быть неузнаваемо разными в разных ситуациях.
Любая деятельность и ее навыки развиваются только для определенного контекста, в котором возникает понимание ситуации, восприятие ее особенностей и выбор действий, прогнозирующих желательный результат. Поэтому понадобились механизмы, отзеркаливающие навыки одних контекстов для использования в других, и не только своих контекстов, но и внешне наблюдаемые проявления чужих. И вообще потребовалось наблюдение с формированием модели причин и следствий наблюдаемого. Субъективная модель понимания и реагирования – большая тема, но это приходится упоминать уже сейчас. Поэтому, чтобы получить представление о субъективных моделях, можно ознакомиться с популярным изложением: “Субъективные модели действительности” - fornit.ru/7305.
Рефлексы, приобретенные в одних условиях, оказываются неприменимыми для других. И тут не поможет механизм, когда, скажем, линии разной толщины и наклона все равно идентифицируются как линия потому, что простейшие примитивы достаточно схожи и детектируются рядом соседних распознавателей, так что свести их все по логике ИЛИ в общий распознаватель механически очень просто. Но распознать линию в карандаше, ребре бумаги, кривую линию на рисунке и т.п. как абстракцию “линия” на этом уровне принципиально невозможно. Для этого нужен совершенно иной принцип организации контекстов: по смыслу воспринимаемого, а не по его форме и других сенсорных характеристик в контексте значимости. Этот механизм реализован в иных по локализации слоях мозга по отношению к тем, где формируются условные рефлексы.
Понятие контекстов значимости – базовое для всех более сложных, чем уже рассмотренных механизмов нейросети. В общем смысле контекст – ограничение восприятия и возможности создавать новые реакции механизмом, игнорирующим все в восприятии, что не относится к данному контексту значимости состояния организма.
Наиболее широкий, общий контекст – состояние аварийного отклонения какого-либо параметра гомеостаза (аварийное состояние) – контекст общей негативной значимости и состояние возврата к норме – контекст позитивной значимости.
Более частные, вложенные контексты ограничивают все вне пищевого поведения, оборонительного поведения, полового поведения и т.п.
Еще более частные контексты – уточнение стиля поведения базовых контекстов. Наиболее узкий частный контекст – условия формирования конкретной условно-рефлекторной реакции.
И здесь – особенно много вариабельности природной реализации. Так, кроме иерархии базовых контекстов значимости есть фоновый механизм “сторожевых реакций” (“старт-реакции”), который работает вне контекстов значимости. Мы его не будем рассматривать – как отдельные ответвления системы адаптивности.
В природе (и в искуссвтенной реализации Beast) контексты реализуются в виде структуры деревьев, что позволяет моментально выходить на конечные ветки активного сочетания условий (fornit.ru/66797).
Прогноз
То, что многие образы восприятия несут с собой уже имеющуюся ассоциацию с позитивным и негативным результатом появления такого образа, позволяет использовать их как прогноз, активируя ту или иную значимость, связанную с ним. И так – со всеми образами, составляющими последовательность цепочек программы реагирования. Поэтому если подсмотреть, какая значимость связана с последним звеном, можно узнать заранее, чем может закончиться такая реакция и предпринять меры, если конец плох.
Для этого нужен механизм произвольности предпросмотра конечного результата: цепочки реакций дополнены средствами отсечения тормозных связей (в нейрофизиологии предполагается, что это проделывают модулирующие нейроны), блокирующих активность последующего звена, пока не сработает предыдущее и будет сигнал о законченной фазе движения, (эта возможность унаследована из торможения во время сна всего из восприятия и реакций).
Что произойдет в нейронной схеме организации цепочки действий, если во всех элементах после первого убрать тормозные нейроны, а первый элемент активировать? Произойдет мгновенный доступ к значимости конечного звена, то, что И.Павлов назвал “опережающим возбуждением”: fornit.ru/an-book-15 и fornit.ru/an-book-19
Хотя на уровне рефлекторных реакций такой прогноз не используется, но уже есть задел для этого.
Активность возбужденных образов, совпадающая с активностью базовых элементов системы значимости, позволяет укреплять связи от ассоциативных зон к первичным зонам системы значимости. Такая связь позволяет устанавливать более общий контекст в зависимости от воспринятого образа. Увидел льва далеко – прятаться, близко – бежать, совсем близко – паника от отсутствия заготовленных реакций или бешеная агрессия. Все это пока еще - неосознанная оценка и выбор поведения, а простые рефлекторные реакции, возникшие в условиях данных обстоятельств. Т.е. связь локализуется около нейронов образов в ассоциативной коре. И эта связь (от ассоциативных зон к первичным зонам системы значимости) должна иметь возможность обрываться, что и проделывают переключатели специализированного органа мозга – гиппокампа, что составляет часть механизма удержания образа, который исследовал А.Иваницкий (fornit.ru/7446).
В мозге циклы гиппокампа обеспечивают обработку текущего актуального образа. Получаемая информация образует текущую картину ситуации – субъективное ощущение (fornit.ru/50394) и в мозге элементы информации о ситуации локализуется около гиппокампа, там же, где и Правила (карты местности, последовательности мышления и т.п.). Поэтому при лоботомии самоощущение сохраняется за счет циклов гиппокампа и отслеживает текущую ситуацию, хотя решения по действиям в таким случае принимается только за счет уже сформированных автоматизмов (произвольность отсутствует).
Это – очень важная особенность – возможность учета влияния от более поздних структур к более ранним, реализуется во всех зонах мозга из-за наследственно предопределенного принципа образования двунаправленных связей (fornit.ru/1474 - т.е. посылка роста аксонов в обратном направлении). Эта связь не участвует в функции распознавания примитива восприятия, а только коммутирует его возможность работать или молчать, обеспечивая контекст восприятия. Т.е. наличие активности такой связи оказывает подвозбуждающее воздействие, на фоне которого специализируется данный распознаватель. Без него профиль рецепторов не возбудит распознаватель потому, что у него не хватит возбуждающего потенциала. А с ним все так же хорошо контрастируется тормозным влиянием соседей в четкий образ. В тех зонах мозга, где организовано это, распознаватели подвозбуждаются текущим контекстом системы значимости (а не последующим образом, которого при формировании распознавателя еще нет).
Так называемые активирующее и тормозное влияние – базовая фишка, точнее, фича (feature) организации мозга.
Это происходит частично в первичных зонах и, в основном, в последующих вторичных зонах с влиянием распознавателей системы значимости, формирующей контекст (см. fornit.ru/23376, fornit.ru/23375, fornit.ru/7446
Эти связи помогают выделять стимулы в текущем контексте значимости и удерживать их. Нейрофизиологи из Университета Карнеги — Меллона (США) сумели подсмотреть, как мозг это делает:
“Информация из зрительной коры поступает в теменную, и если там какой-то объект будет признан особо важным, то соответствующий участок теменной коры усилит подведомственный участок коры зрительной — и зрительное внимание сосредоточится на этом объекте.” (fornit.ru/5202)
В третичной зоне рецепторы гомеостаза (в ипостаси нейронов-распознавателей особенностей сочетания признаков отклонения параметров) не образует контекст, а непосредственно придает определенную значимость данному образу, точнее это есть предельно узкий, пока еще вырожденный контекст для его расширения в случае ветвления от этого образа других реакций.
Как уже говорилось, после развития слоев мозга они утрачивают возможность образования там новых связей. Поэтому в первичных и вторичных зонах остаются только те связи с другими областями мозга, которые были активны во время их специализации. Понятно, что третичных зон еще не было и поэтому связей от них быть не может. А вот основа механизма уже была заложена.
Констатируя некоторую сумбурность последних абзацев, затрудняющую понимание, примите оправдания: нужно было сопоставить в общем понимании сразу несколько принципов, которые еще толком не рассматривались в механизмах реализации. Но при итерации прочтения все будет пониматься лучше. Альтернатива – очень занудное и долгое рассмотрение этих принципов и потом сведение в общую картину, как было сейчас сделано.
Если есть возможность связи от образа в ассоциативной коре к базовым (и не только базовым) распознавателям значимости, от которых образуется контекст восприятия, выделяющий предшествующие примитивы восприятия на уровне вторичной коры, то при замыкании такой связи образуется кольцо циркуляции сигнала: активный образ поддерживает активность распознавателей значимости, которые обеспечивают активность этого образа, и такая циркуляция возбуждения будет продолжаться, пока ее не прервет что-то.
Возникает вопрос: когда пропадает стимул, как может активировать образ во вторичной зоне мозга, всего лишь подвозбуждающей силы значимости? Ведь только совместное воздействие стимула и подвозбуждающей связи контекста значимости становится достаточным для активации образа. Схемотехнически это реализовать довольно просто, переключая силу подвозбуждения от фоновой до активирующей. Как именно это проделывается в природе точно не ясно. Описано свойство нейрона временно облегчать свое возбуждение (fornit.ru/5174), если только что он был возбужден. Но есть совершенно достоверные факты исследований, что именно такие зацикленные связи удерживают стимул в активном состоянии. А.Иваницкий несколько преждевременно полагал, что это – и есть субъективное переживание, элемент мысли, но это – всего лишь одна из основ организации субъективности – способность удерживать образ тогда, когда активирующий стимул уже пропал: fornit.ru/an-book-11
Если бы взгляд был постоянно прикован к изображению, то нейрон активного распознавателя быстро бы истощался постоянной генерацией. Но зрачок постоянно скачет на небольшие расстояния (стаккады), расширяя зону наиболее четкой своей оптики, а распознаватели генерируют не постоянную серию импульсов, а короткие сигналы детекции образа, которые удерживаются на уровне вторичной коры и поддерживающего контекста локального внимания от системы значимости (хотя частота стаккад и сама по себе достаточна для поддержания активного образа, но механизм обращения внимания на образ все равно срабатывает и обратной связью поддерживает память образа).
Можно рассматривать все вокруг, а следы распознавания образов будут продолжать воздействовать в данном контексте на реакции. При циркуляции активности по кольцу нейроны вполне успевают восстановиться и не происходит их истощения. Но с накоплением числа таких закольцовок общая локальная область мозга начинает испытывать проблемы регенерации энергетики и продуктов метаболизма. Кроме того, при этом возникают множественные активности, мешающие новым более слабым за счет бокового торможения (острота и концентрация внимания снижается), вплоть до настолько полного забивания активностями всех каналов, что становится невозможной нормальная гомеостатическая регуляция. При длительном лишении сна происходит ухудшение возможности реагирования вплоть до смерти из-за множественной разрегуляции жизненно важных параметров при, казалось бы, полной исправности регулирующих структур мозга.
В искусственной зрительной системе не нужны были бы стаккады глазных яблок, т.к. ничто бы не истощалось, но поле зрения все равно ограничено и так же возникает проблема удержания образов в активном контексте.
Дополнительно: fornit.ru/7004
Условные рефлексы
Условные рефлексы – первая половина (не по числу, а по границе разделения рефлексы-автоматизмы) индивидуальных адаптивных реакций с учетом контекстов условий, что формируются в онтогенезе человека вскоре после рождения, постепенно уступая более эффективному и гибкому механизму автоматизмов, составляющих другую половину представительства в мозге.
Это заранее подчеркивается потому, что физиологи все еще традиционно не различают механизмы этих двух явлений, у них все – рефлексы, а потом вдруг сразу – какое-то там непонятное сознание. С этим устаревшим представлением, как и самим термином “рефлекс”, нужно что-то делать. Поэтому мы этот термин оставим для уровня простых сочетаний рецепторных распознавателей с эффекторными, которые называли “безусловным рефлексом”, и для контекстно-зависимых сочетаний – “условных рефлексов”. Наиболее сложные условные рефлексы начинают использовать новое качество выделения контекста – новизну, что очень логично для адаптивности.
Но даже самые сложные условные рефлексы формируются тупо (буквально без сознания), автоматизмы же – с использованием функциональности психических процессов.
Понять второе без первого не получиться, и вообще второе крепко основывается на первом.
Пока что автоматизмы не будут рассматриваться, потому что до них еще предстоит разобраться с несколькими важнейшими механизмами, в частности с тем, как, чуть ли не с рождения, начинает организовываться адаптивность “с учителем” (в простейшем случае - импринтинг). До этого формирование связей между нейронами никак не рассматривалось в плане воздействия какого-то учителя и прямого учета ошибки.
А пока что, все, что формируется еще “без учителя” – рефлексы. На первый взгляд кажется, что подкрепление, связанное с откликом системы гомеостатической значимости, и является “учителем”, но это не так потому, что новый условный рефлекс всегда формируется на основе уже имеющегося условного или безусловного рефлекса, которые и обуславливают реакцию или избегания или стремления. А подкрепление лишь инициирует необходимость формирования новой реакции, которая возникает в определенном контексте данной значимости. Этот контекст и ограничивает возможность восприятия и действия, только в нем и может возникнуть ветвление старой реакция на новый стимул.
Ранее говорилось о том, что безусловные рефлексы не наследуются, а лишь наследственно предопределяются и возникают только в определенных условиях. Тем более не наследуются условные рефлексы: те связи, что возникают при определенных воспринимаемых особью условиях (в дополнение к условиям развития нейронной сети), см. Наследуются ли условные рефлексы? Л.В. Крушинский: fornit.ru/807
К сожалению, столь обширную до неисчерпаемости тему имеет определенный смысл вместить в одну главу, хотя потом все остальное будет развиваться на этой основе. Иначе мы утонем во второстепенных деталях, которые не существенны для системных представлений. Кроме того, чем более подробны объяснения, тем больше лишней каши в голове.
Далее будет немало лаконичных, но важных рассуждений, образующих логику явления на основе эмпирических данных исследований, поэтому, очень стоит быть внимательным к, казалось бы, малозначащим фразам, потому как ничего просто так произнесено не будет.
П.К.Анохин делал попытки различать условные рефлексы и формируемые произвольно автоматизмы: fornit.ru/7204 но в классических определениях продолжается путаница с этим: fornit.ru/64936.
Как уже упоминалось, строго граничные условия функциональности условных рефлексов были определены при разработке схемы Beast, когда стало ясно, что может, а что никак не может быть привлечено для формирования реакций на уровне условных рефлексов. Самым общим и лаконичным выводом является определение, что условные рефлексы всего лишь являются “зеркалом” предшествующих рефлексов, полностью наследуя программу действия, но запуская ее от нового стимула, который становится “синонимом” старого стимула исходного рефлекса. Т.е. при образовании условного рефлекса обязательна детекция нового в данных условиях, чего не упоминается ни в каких классических определениях. Более нет никаких других адаптивных функций у условного рефлекса. Более подробно: fornit.ru/64936.
Условия образования условного рефлекса
Приведем пару наиболее характерных описаний эмпирически полученных данных об условиях, необходимых для возникновения нового условного рефлекса.
Механизм образования условных рефлексов (fornit.ru/5231)
“Рефлекторное возбуждение, возникающее в какой-либо рефлексогенной зоне, адресуется из центров чувствительных нервов не всем, а строго определенным эффекторным (двигательным или секреторным) нервным центрам. В безусловных рефлексах эта связь между чувствительными и эффекторными центрами является врожденной.
Условные рефлексы характеризуются тем, что каждый раздражитель (световой, звуковой, проприоцептивный и др.) может при известных условиях приобрести сигнальное значение и стать раздражителем, вызывающим специальную ответную реакцию организма: двигательную, секреторную, пищевую, оборонительную и т. д.
Условные рефлексы хорошо образуются только при определенных условиях. Главнейшими из них являются:
1) повторное сочетание действия ранее индифферентного условного раздражителя с действием подкрепляющего безусловного или ранее хорошо выработанного условного раздражителя;
2) некоторое предшествование во времени действия индифферентного агента действию подкрепляющего раздражителя;
3) бодрое состояние организма;
4) отсутствие других видов активной деятельности;
5) достаточная степень возбудимости безусловного или хорошо закрепленного условного подкрепляющего раздражителя;
6) надпороговая интенсивность условного раздражителя.
Хотя это – пример статьи, где условные рефлексы – единственный вид закрепления навыков (т.е. осознанные автоматизмы = условным рефлексам), но здесь важно обобщение условий их возникновения”.
В другом источнике (fornit.ru/21292):
“1) наличие двух раздражителей, один из которых безусловный (пища, болевой раздражитель и др.), вызывающий безусловно-рефлекторную реакцию, а другой - условный (сигнальный), сигнализирующий о предстоящем безусловном раздражении (свет, звук, вид пищи и т.д.);
2) многократное сочетание условного и безусловного раздражителей (хотя возможно образование условного рефлекса при их однократном сочетании);
3) условный раздражитель должен предшествовать действию безусловного;
4) в качестве условного раздражителя может быть использован любой раздражитель внешней или внутренней среды, который должен быть по возможности индифферентным, не вызывать обронительной реакции, не обладать чрезмерной силой и способен привлекать внимание;
5) безусловный раздражитель должен быть достаточно сильным,
в противном случае временная связь не сформируется;
6) возбуждение от безусловного раздражителя должно быть более сильным, чем от условного;
7) необходимо устранить посторонние раздражители, так как они могут вызывать торможение условного рефлекса;
8) животное, у которого вырабатывается условный рефлекс, должно быть здоровым;
9) при выработке условного рефлекса должна быть выражена мотивация, например, при выработке пищевого слюноотделительного рефлекса животное должно быть голодным, у сытого - этот рефлекс не вырабатывается.
Разобщение корковых центров условного и безусловного рефлексов путем пересечения коры мозга не препятствует образованию условного рефлекса. Это свидетельствует о том, что временная связь может образоваться между корковым центром условного рефлекса, подкорковьм центром безусловного рефлекса и корковым центром безусловного рефлекса”.
Список условий может различаться в разных источниках, но всегда можно выделить общее.
Мы уже говорили об универсальном условии образования связей: связь формируется между двумя возбужденными нейронами, между которыми уже есть непроводящий синапс и если есть достаточно времени для формирования проводимости этого синапса.
При этом необязательно наличие постоянного возбуждения (и даже нежелательно, т.к. оно быстро истощает нейрон), важно, чтобы оно с достаточной частотой появлялось по обе стороны синапса и тогда там образуется метаболический след, оказывающий влияние на экспрессию генов.
Кроме одновременности, экспериментально найденные условия образования рефлексов требуют некоторого предшествования условного раздражителя, иначе связь затрудняется. По логике причин и следствий это условие понятно: нужно реагировать именно на причину, а не то что потом случается уже по какой-то другой причине. Если не сделать такой механизм, то начнут образовываться довольно парадоксальные реакции, не имеющие причинности.
Схемотехнически это организовать несложно: связь образуется, если есть разрешающий потенциал от стимула. Как именно такой механизм организован в природе, сказать пока сложно (наверняка множеством хитросложных вариантов), но он есть совершенно достоверно (т.е. воспроизводится эмпирически) и стоит хотя бы предположить, как это организовано.
Рост дендритов
Дендриты начинают развиваться уже после созревания нейрона и его готовности к установлению связей. Существует немало данных, что рост дендритов стимулируется активностью предшествующих аксонов и идет в сторону активных из них, например в fornit.ru/23468:
“Ген OSTN стал включаться в активно работающих нейронах. Сигнальный белок, кодируемый геном OSTN (красные кружочки), выделяется в межклеточную среду и регулирует рост дендритов”.
В другом источнике fornit.ru/5426:
“...характер появления дендритных выростов зависел от того, какое именно упражнение нужно было выполнить. Например, если мышь должна была идти по вращающейся палке в одну сторону, то шипики возникали на одних дендритах, а если нужно было идти в другую сторону, то шипики появлялись на других дендритах. То есть клеточная морфология нейронных отростков зависела от того, что за информацию нужно было обработать”.
Это – очень логично: не пытаться устанавливать возможные контакты со всем, что лежит рядом, а только с тем, что проявляет рецепторную активность.
Для самых первых слоев нейронов в первичных зонах это бессмысленно потому, что все они почти равновероятно засвечиваются активностью первичного рецепторного поля. Но уже следующий слой работает с выделенными примитивами восприятия. И чем далее, тем роль логики связи с рецепторами признаков, проявляющих активность, приобретает все большее значение.
На уровне условных рефлексов это может начать играть роль логики причины происходящего. С первым предъявлением условного стимула формируется готовность образования связей, а с последующими она укрепляется.
Но нам не нужно гадать, как именно это устроено, важно знать принцип и знать, как можно реализовать это схемотехнически, а выбор конкретного механизма реализации будет зависеть от того, что и как мы конструируем. Так будем поступать во всех подобных случаях впредь: экспериментально подтвержденный принцип является главной основой для понимания и модели механизма, а как он организован на нейрофизиологическом (атомарном, кварковом и т.п.) уровне – не входит в суть данной абстракции, ведь зная принцип, всегда можно представить и механизм, оптимальный для данного вида реализации.
Вот другой экспериментальный факт: многие реакции формируются именно после появления причины (условного раздражителя) и, самое главное, нет одновременности условного стимула и того, с чем нужно бы образовать связь. Так, съев, казалось бы, привычную еду, но почувствовав себя после этого плохо, потом чувствуется отвращение к этой еде. Но ведь этому вначале не предшествовал какой-то раздражитель, все было привычно беззаботно, как всегда. Чтобы связать отравление со съеденной едой, мы должны осмыслить, что среди всего происходящего именно еда стала причиной недомогания, хотя бывает трудно исключить другие возможные причины. Мы должны проделать немалую мыслительную работу или просто не заметить другие возможные причины и сразу свалить все на еду.
Но на уровне условных рефлексов функциональность осмысления невозможна принципиально, и особь, ограниченная лишь механизмами условных рефлексов, будет снова и снова жрать испорченную еду, подыхая как тараканы от отравы, если только такая еда сразу же не покажется отвратительной. Описанное понимание причины – продукт осмысления, реализуется не как рефлекс, а как автоматизм, - тема будущих обобщений.
Однако, испорченная еда – очень важный фактор выживания, и чисто смертельным отбором появился безусловный рефлекс: если стало плохо и нет более непосредственных причин, то, скорее всего это – из-за еды, особенно если пошли всякие глюки в восприятии, характерные для интоксикации. Реакция “cтало плохо – стошнить” в какой-то мере позволяет выжить, но она может срабатывать совершенно зря, когда после плотной еды человек пошел купаться в море и там возникли необычные сигналы от желудка в условиях водной “невесомости” или когда мы катаемся на карусели или даже просто едем в автомобиле, покачивающим нас и меняющим направлением движения, но без предшествующих координирующих сигналов (если укачиваемый человек рулит сам, то его не укачивает).
Если включить логику, то можно было бы так организовать ответную реакцию: если действие привело к ухудшению аварийных параметров, т.е. оно закончилось плохо, то нужно его блокировать появившейся гомеостатической значимостью.
Да, такая значимость связывается со звеном конца неудачной реакции (так же как позитив его удачности) по условиям образования связей, но это никак не влияет на то, избегать ли его впредь или оставить в статусе удачного на уровне такого звена, т.е. это пока что не играет роль учителя в образовании рефлексов. Для этого нужен более высокий уровень, позволяющий подсмотреть конечную значимость и решить, делать ли так или нет (такая возможность формируется в лобных долях одновременно и независимо с условными рефлексами у достаточно продвинутых в организации нейросети животных, но мы пока это не рассматриваем именно из-за независимости процессов).
На уровне рефлексов связь межу стимулом и одной из уже существующих наследственных или условных реакций “тупо” (ведь и в самом деле – тупо, раз не осознается) возникает при одновременности активности с обеих сторон при некотором предшествовании стимула и строго в текущем контексте восприятия-действия. И все. Учителя в понимании искусственных нейросетей – нет на уровне рефлексов.
Интересен рефлекс избегания, описанный как “рефлекс второго рода” - как обучение методом проб и ошибок. Механизм такого обучения выходит за рамки описанного условия образования рефлекса. Здесь нужно внимательно учитывать, как именно вызывалась реакция в ходе образования такого рефлекса: или на основе уже имеющейся реакции с сочетанием такой реакции и условного раздражителя или же это была реакция на основе выбора по результатам ранее окончившейся реакции и установившемуся состоянию гомеостата. Т.е. всегда нужно в первую очередь вспоминать про гистерезис запаздывания сигналов состояния жизненных параметров организма.
Имеющимся набором распознавателей состояния организма и связанных с ними реакций для восстановления нормы исчерпывается значимость таких распознавателей. На этом уровне нет возможности начать избегать то, что не понравилось, и стремиться делать нравящиеся (это появится только с механизмами произвольности выбора на уровне не рефлексов, а осознанно формируемых автоматизмов), но есть целевая направленность контекстов – на восстановление с прекращением компенсаторной реакции, когда контекст изменится на констатацию достигнутой нормы. Наелся, - прекращается стиль пищевого поведения, переключаясь на стиль благостного бездействия при отсутствии опасности и другой необходимости шевелиться.
Итак, просто учтем, что на уровне рефлексов раздражитель всегда предшествует, а в остальном процесс следует условиям образования связей в активном контексте, выделяющим только характерное для данного стиля восприятия и поведения, а все раздражители и подкрепления, оказывающиеся вне такого контекста, связи не образуют.
От стимулов первичной и вторичной зоны возникают только безусловные рефлексы и те условные, которые используют только сенсорику данного канала рецепции. После завершения развития этих зон, там уже ничего не меняется, а условные рефлексы возникают только на основе третичной, ассоциативной теменной зоне (а не третичной лобной, на основе которой возникают принципиально отличающиеся автоматизмы, и это – самое интересное, что ждет нас впереди).
Общий механизм возникновения условного рефлекса
Практика схемотехнического мышления в достаточной степени позволяет не рисовать примеры схем, которые всегда приходится упрощать, а представлять возможности их построения, что гораздо продуктивнее, чем конкретика. Конечно, чтобы наработать навык использования схемотехнического мышления до уровня моментального возникновения в голове решения, нужно хотя бы лет пять увлеченно разрабатывать схемы, но и без этого, как минимум, возникает понимание как, в принципе, это делается без всякого колдунства. Самая общая формула:
В области подвозбуждения текущего контекста (активационного влияния) активные образы восприятия, образуют связи с активными нейронами уже существующего рефлекса, безусловного или условного.
Представим, что у нас есть уже сформированный набор реакций: защитная реакция избегания раздражения и пищевая реакция стремления проглотить раздражитель.
Если при срабатывании пищевой реакции возбудилась цепочка программы пищевого поведения, особь потянулась к кормушке красного цвета, но в этот момент ее ударили током, то сработает имеющийся рефлекс избегания и ошарашенная особь, моментально переключившаяся в контекст оборонительного поведения, отдернется.
Это может быть случайностью, а есть-то нужно, животное снова тянется к красной миске и опять получает удар. Это уже серьезно, уже намного дольше колебания, но есть-то нужно, а ей подставляют зеленую миску. Она решается и все хорошо. Потом опять красная – плохо! Начинает закрепляться связь между распознавателем красной миски и рефлексом избегания в контексте оборонительного поведения. На самом деле у птиц, рыб, кошек-собак уже хорошо развиты механизмы формирования автоматизмов, и они за один раз поймут, что произошло и остерегутся тупо снова наступать на грабли, что спутает карты исследователю рефлексов. Но в некритичных случаях у этих животных игнорируется опыт с малозначащими последствиями, и преимущество получают рефлекторные механизмы (у собак более совершенный уровень такой адаптивности, а кошки сразу учитывают только сильное потрясение).
Еще чище опыты над высшими насекомыми: Исследователи выработали условный рефлекс у тараканов (fornit.ru/505):
“Учёные обнаружили, что если распылять эту эссенцию перед насекомыми за две секунды до кормления сахаром пять раз подряд, то их нервная система начинает реагировать на этот запах, как на условный раздражитель. Повторив эксперимент на следующий день, исследователи добились того же эффекта”.
Сначала должен активироваться определенный контекст поведения, например пищевой потребности, когда нужна еда. Потом активируется распознаватель какого-то признака восприятия (и оставляет метаболическую метку, показывающую, что это – область для установления новой связи). Если затем появляется активность уже имеющегося рефлекса, то образуется связь с ним от нового образа восприятия. Если новый образ восприятия активируется уже после срабатывания рефлекса поглощения пищи, то он не учитывается.
Конечно же, и на уровне рефлексов у природы множество всякого путающего исследователя рукожопства. Вот пример (fornit.ru/2565):
“Нейробиологи из Канады и США обнаружили, что в запоминании простых навыков участвуют не все нервные клетки, получающие необходимую для этого информацию, а лишь около четверти из них. То, какие именно нейроны примут участие в формировании долговременной памяти, зависит от концентрации регуляторного белка CREB в клеточном ядре. Если искусственно повысить концентрацию CREB в некоторых нейронах, запоминать будут именно они. Если заблокировать CREB в части нейронов, роль запоминающих возьмут на себя другие нервные клетки”.
Зачем это понадобилось? Да просто так легли кости случайности, но находка оказалась жизнеспособной и повесть жизни продолжилась. В той же статье:
![]() “Оказалось, что процесс обучения и запоминания не имеет ничего общего с какими-то высшими, идеальными или духовными материями, а полностью объясняется довольно простыми и совершенно автоматическими событиями на уровне отдельных нейронов. Весь процесс можно полностью воспроизвести на простейшей системе из трех изолированных нервных клеток. Один нейрон (сенсорный) получает сигнал от сифона (в данном случае — чувствует легкое прикосновение). Сенсорный нейрон передает импульс моторному нейрону, который, в свою очередь, заставляет сокращаться мышцы, участвующие в защитной реакции (Aplysia втягивает жабру и выбрасывает в воду порцию красных чернил). Информация об ударе по хвосту поступает от третьего нейрона, который в данном случае играет роль модулирующего. Нервный импульс от одного нейрона к другому передается посредством выброса сигнальных веществ (нейромедиаторов). Точки межнейронных контактов, в которых происходит выброс нейромидиатора, называются синапсами.
“Оказалось, что процесс обучения и запоминания не имеет ничего общего с какими-то высшими, идеальными или духовными материями, а полностью объясняется довольно простыми и совершенно автоматическими событиями на уровне отдельных нейронов. Весь процесс можно полностью воспроизвести на простейшей системе из трех изолированных нервных клеток. Один нейрон (сенсорный) получает сигнал от сифона (в данном случае — чувствует легкое прикосновение). Сенсорный нейрон передает импульс моторному нейрону, который, в свою очередь, заставляет сокращаться мышцы, участвующие в защитной реакции (Aplysia втягивает жабру и выбрасывает в воду порцию красных чернил). Информация об ударе по хвосту поступает от третьего нейрона, который в данном случае играет роль модулирующего. Нервный импульс от одного нейрона к другому передается посредством выброса сигнальных веществ (нейромедиаторов). Точки межнейронных контактов, в которых происходит выброс нейромидиатора, называются синапсами.
На рисунке показаны два синапса. Первый служит для передачи импульса от сенсорного нейрона к моторному. Второй синапс передает импульс от модулирующего нейрона к окончанию сенсорного. Если в момент прикосновения к сифону модулирующий нейрон «молчит» (по хвосту не бьют), в синапсе 1 выбрасывается мало нейромедиатора, и моторный нейрон не возбуждается”.
Но как сложно вся эта принципиальная простота наворочана!
Новые реакции на основе старых
Интересно то, что в организме сохраняются и реализуются в условиях своего развития все более древние наследственно предопределенные реакции и они лишь дополняются новыми.
Это похоже на то, как на основе примитивов восприятия формируются более сложные образы, а если нет такого примитива, то нет и последующих на его основе, - только касается уже реакций. Получается два лимитирующих адаптационные возможности фактора: примитивы восприятия лимитируют образы условных раздражителей, а база безусловных рефлексов – диапазон того, что можно построить на их основе. Что не удивляет: так же на основе клетки формируется и весь более сложный организм.
Все условные рефлексы возникают на основе уже имеющихся безусловных и сформированных условных, точнее - на основе уже заготовленных распознавателей необходимости запуска реакций в моторной зоне коры, разделяющей мозг на две важнейших части: заднюю – область условных рефлексов и лобную – область реакций на совершенно новом принципе.
Точно так же, как в первичных зонах мозга формировались примитивы восприятия, одновременный процесс противоположной направленности происходит и в первичных зонах моторной коры: формирование распознавателей необходимости запуска примитивов реагирования.
Как и при формировании примитивов восприятия, так и при формировании примитивов реагирования созревающие нейроны проявляют спонтанную активность, что, согласно условиям формирования связей, обеспечивает появление проводимости в соединяющих нейроны синапсах. Как и слуховые примитивы, моторные примитивы формируются еще до рождения (fornit.ru/23514).
На основе этих эффекторных распознавателей и происходит все последующее усложнение адаптивного реагирования. Это, вместе с примитивами рецепторных распознавателей, составляет базу индивидуальных способностей-возможностей. Первые связи между рецепторными и эффекторными распознавателями составляют первые базовые реакции.
Если для самых древних реакций характерны прямые сочетания первичных рецепторов и конечных эффекторов, то со все большим включением промежуточных нейронов сочетания образуются между нейронами, выполняющими роль внутреннего рецептора и нейронами, выполняющими роль внутреннего эффектора – начального звена цепочек эффекторной реакции.
Цепочки поведения запускаются безусловными и условными рефлексами, они могут ветвиться при образовании нового условного рефлекса и становиться сколь угодно сложными, но остаются максимально эффективными из-за древовидной иерархической структуры (fornit.ru/66797). В каждой точке ветвления возникает более одного варианта возможных действий и выбор одного из них происходит при конкретизации условий, когда может запуститься строго один из ответов, подавляя остальные латеральным торможением в условиях более слабо выраженной активности.
Такое дерево рефлексов и было реализовано в схеме Beast (fornit.ru/beast4). Вообще все, что зависит от условий в той схеме сделано в виде деревьев, а сами условия определяются активными образами не в виде полного среза активностей, а в виде активности только самого обобщающего образа – распознавателя совокупности активности. Это позволяет строить деревья со строго фиксированным числом уровней так же, как в новой коре есть такая определенность слоев – по той же причине использования конечных образов, заменяющих собой все предыдущие (fornit.ru/43642).
Использование иерархии образов восприятия и образов действий – самый оптимальный метод структурирования промежуточных данных (связанной со значимостью информации), - в отличие от практикуемого в искусственных нейросетях, оказывающихся из-за этого тупиковым направлением.
Чтобы распознать уникальное сочетание активностей сигналов нескольких иерархически усложняющихся типов, лучше всего подходит структура дерева, где каждая ветка состоит из фиксированного числа видов образов. В таком дереве находится место для любых сочетаний исходных образов, поиск по дереву – самый быстрый вид поиска, требуется минимальные вычислительные ресурсы в случае применения рекурсивного прохода веток дерева (вида обратной связи). Узнавание любого сочетания достигается при числе итераций, равном числу использованных видов образов.
Вернемся к реакциям на внешнее воздействие.
После выполнения реакций организм как-то откликается на результат состоянием гомеостатической значимости (лучше ли стало или хуже), связывая ее с конечным звеном, вызвавшим рефлекс за время формирования связи, которого достаточно для такой ассоциации с учетом запаздывания отклика жизненных параметров и, тем самым, образуя основу для контекста данной реакции.
Таким образом, с каждым новым рефлексом возникают новые, все более вложенные (все более уточняющие условия) контексты значимости, каждый из которых способен на формирование новых реакций в случае появления новых условий, вызывающих образование рефлекса.
Это приводит к возможности организовывать удержание активностей образов запуска реакций, замыканием их выхода со входом распознавателей системы значимости, что образует закольцовку, самоподдерживающую активность.
Такое удержание позволяет 1) продлить действие условного раздражителя для образования нового рефлекса в течение всего одного предъявления стимула и 2) удерживать раздражитель в активном состоянии, даже если временно пропал сам рецепторный сигнал, что позволяет продолжать выполнение цепочки целевых действий.
Это – максимальная достигнутая сложность в организации ответных реакций на уровне условных рефлексов.
Необходимость повторения сочетаний условного и безусловного компонента реакции
Зачем нужны повторения сочетаний для закрепления условного рефлекса? Кроме того, что на установление постоянной связи нужно время (примерно полчаса), а явление сочетания стимула и ответной реакции бывает значительно меньше, нужно еще и конкретизировать стимул. Он уже в немалой степени конкретизирован наличием подвозбуждающего контекста восприятия, ограничивающего число принимаемых во внимание образов восприятия, но в восприятии может быть несколько значимых (поддерживаемых контекстом системы значимости) стимулов. С каждым новым сочетанием точность реакции возрастает, потому как все второстепенное отсеивается, связь все более укрепляется с момента своего предшествующего состояния. Это называется “стадией специализации” условного рефлекса.
Ориентировочный рефлекс
Эволюционно возник еще один важный механизм уточнения важности стимула – ориентировочная реакция или “ориентировочный рефлекс”. Это – безусловный, наследственно предопределенный рефлекс, позволяющий обратить внимание именно на что-то новое в восприятии, чтобы заняться информационной обработкой именно данного объекта внимания. Т.е. среди всего привычного, что уже не должно отвлекать мозг и, по логике, не должно бы участвовать в формировании новых реакций, может выделяться необычное в профиле возбуждения и тогда именно это оказывается преимущественно выделенным для формирования новой реакции.
Ориентировочный рефлекс ограничен текущим контекстом значимости состояния организма и его соответствующего стиля поведения. Кроме всего, в течение продолжительного жизненного опыта возникают образы восприятия, которые связаны с распознавателями значимости в данном общем контексте: то, что обычно сопровождает важные для организма состояния, разбалансировало гомеостаз и возвратило его в норму. Наличие таких стимулов вызывает повышенную селекцию (конкретизацию контекста) и преимущественное их участие в формировании новой реакции.
По логике индивидуальной адаптивности это – рефлекторный механизм для подключения аппарата нахождения новых вариантов реагирования, если в них появился новый и значимый раздражитель.
Для распознавания нового необходимо поддержание активности образов, которые будут сравниваться по новизне между собой и активирующимися новыми образами в восприятии в общей картине активности данного контекста значимости. Новое всегда в чем-то присутствует в восприятии и нужно выделить только то новое, что имеет наибольшую значимость в данном контексте. Такой объект нужно поддерживать в активном состоянии на время его обработки аппаратом принятия решений. Это делается замыканием связей с выхода на вход образов, что делает активность самоподдерживающейся.
Замыканием связей с распознавателями значимости для самоподдержания активности образа занимаются переключатели гиппокампа. Логично, что именно рядом с ними и формируются структуры, позволяющие детектировать новое в данном профиле активности. Там они и были обнаружены и описаны Е.Соколовым: fornit.ru/5134. Впервые нейроны новизны обнаружил Г. Джаспер в гиппокампе. Это означает, что распознаватели нового в данном контексте не локализованы в теменной ассоциативной зоне, а они сконцентрированы в слоях гиппокампа для обслуживания образов теменной ассоциативной зоны и связи их с другими структурами.
Ориентировочный рефлекс важен тем, что он развился в механизм селективности осознанного внимания для формирования нового качества реакций - автоматизмов.
Ориентировочный рефлекс – особый по сравнению с другими безусловными рефлексами потому, что не использует конкретные стимулы, а реализуется механизмом выделения нового среди всего значимого в данном контексте значимости.
Вернемся к условным рефлексам.
Итак, возможны три основных состояния образования условных рефлексов:
1. привычный фон широкого контекста восприятия с множеством раздражителей, - требует наибольшего числа уточняющих сочетаний для конкретизации условного раздражителя;
2. появление новых признаков в широком (вплоть до базового) контекста, - реакция формируется преимущественно с участием новых стимулов;
3. стимулы вызывают рефлекторное сужение контекста значимости, выделяющее их – такая реакция не нуждается в выделенной стадии специализации и обходится минимальным числом сочетаний в силу самоподдержания активности образа.
Последнее обобщение материалов по функциональности и механизмам ориентировочного рефлекса - в статье Ориентировочный рефлекс.
Угасание условных рефлексов
Важным механизмом в конкретизации ответной рефлекторной реакции является повторение сочетаний стимулов, когда все совпадающее усиливается, а случайное все более утрачивается (похоже на принцип экспозиции при формировании примитивов восприятия в первичных зонах мозга). Чем больше число подтверждений рефлекса, тем он определеннее и сильнее в конкуренции со всеми рядом расположенными активностями. Это настолько важно, что даже если времени образования связи уже вполне хватило, то стоит продолжить процесс селективности, чтобы не засорять область реагирования в данном контексте множественными случайными реакциями.
Такая селективность осуществляется несколькими механизмами, обеспечивающими торможение, ослабление уже возникшего рефлекса при его невостребованности.
Процесс может быть пассивным за счет дезадаптации и естественного ослабления связей, возможно облегченного по сравнению с безусловными рефлексами, т.е. условные рефлексы легко забываются. Вспомним, что первичные примитивы восприятия были представлены сразу многими соседними детекторами, активирующимися одновременно (пятно стимула вместо утонченной активации), а при усложнении примитивов этот эффект все более нивелируется, особенно в области контекстных рефлексов так, что они оказываются представлены чуть ли не единичными цепочками управления.
Кроме того, чем моложе рефлексы, тем они легче ослабляются за счет боковых конкурентов взаимным торможением.
Вот что пишут про угасание условных рефлексов: fornit.ru/7511:
“...малейшее изменение в окружающей среде меняет сигнальную роль условного раздражителя, что свидетельствует о тончайшем приспособлении организма к условиям своего существования”.
Это – о том, что рефлекс остро заточен на определенные условия, о чем уже была речь и что обусловлено формированием реакции строго в определенном контексте значимости.
“Вот пример натурального условного торможения. Собака-ищейка научена брать пищу только из рук своего хозяина и не прикасается к ней, если ее кормит кто-то другой: вид и запах пищи перестают быть условным раздражителем в других условиях. Роль условного тормоза здесь играет вид и запах постороннего человека.
Любой вид внутреннего торможения является активным процессом задержки, подавления условных рефлексов.
В этом легко убедиться, если в момент внутреннего торможения подействовать на животное посторонним к опыту раздражителем, который в других условиях является внешним тормозом. Внешнее торможение встречается с внутренним и происходит растормаживание: сигнальный раздражитель вновь вызывает временно задержанный условный рефлекс”.
Свойства условных рефлексов
Схемная реализация позволила окончательно обобщить свойства условных рефлексов:
Для образования условных рефлексов необходимо:
1. Совпадение во времени (сочетание) какого-либо индифферентного раздражителя (условного) с раздражителем, вызывающим соответствующий безусловный рефлекс (безусловный раздражитель).
2. Необходимо, чтобы действие условного раздражителя несколько предшествовало действию безусловного.
3. Условный раздражитель должен быть не вызывающим значительной самостоятельной реакции.
4. Рефлекс возникает только после нескольких повторений сочетаний, это избавляет от случайных связей.
Условный рефлекс может образовываться на основе безусловного или на основе имеющегося безусловного, используя действия исходного рефлекса для новых условий. Такие цепи рефлексов ничем не ограничены.
Условия затухания условного рефлекса:
1. Долгое отсутствие пускового стимула (узла ветки, с которого он запускается);
2. Действие конкурентных раздражителей – т.е. подавление конкурентными рефлексами и автоматизмами – т.е. если есть более значимый рефлекс или любой автоматизм на тот же пусковой стимул, то он блокирует условный рефлекс. Автоматизм приоритетнее любого рефлекса.
3. При отсутствии «подкрепления» после совершенного действия. Но безусловные рефлексы не угасают при этом, они безусловны и по отношению к тому, что происходит после действия и их «подкрепление» – обусловлено наследственной эволюцией: безусловные рефлексы постоянны, даны от рождения и не угасают на протяжении всей жизни. У условных рефлексов точно та же функциональная задача, только с новыми стимулами, значит, им также не нужно последующее подкреплением. В литературе часто путается отсутствие подкрепления с дезадаптацией, а также условные рефлексы и автоматизмы, образующиеся при осознании.
4. Более «молодые» и непрочные условные рефлексы угасают быстрее, чем более «старые», прочные условно-рефлекторные связи.
Алгоритм формирования условного рефлекса, в реализации Beast:
1. Условный рефлекс возникает там, где нет безусловного на основе нового стимула N, привязывая к нему действия того рефлекса (условного или безусловного), которое вызывало реакцию ПОСЛЕ данного нового стимула M.
2. Но теперь если в восприятии появляется стимул N, то вызываемый им условный рефлекс перекрывает все рефлексы более низкого уровня, в том числе условные меньшего уровня.
При разработке было выявлено очевидное заблуждение в классической интерпретации условных рефлексов, утверждающее, что для поддержания условного рефлекса нужно обязательное подкрепление.
Факт: собака перестает бежать к еде по звонку, если не видит еду несколько раз после звонка.
Тут действует именно то, что она не видит еду, оценивает ложность звонка, т.е. у нее не условный рефлекс, а более высокоуровневый осознанный автоматизм, который исследователи путают с рефлексом. Разные условные рефлексы без подкрепления угасают с неодинаковой скоростью. Более "молодые" и непрочные условные рефлексы угасают быстрее, чем более "старые", прочные условно-рефлекторные связи.
Итак, пример: если при прикосновении к миске с водой начало бить током, у животного срабатывает безусловный рефлекс отдергивания. При повторении несколько раз сам образ миски с водой начинает вызывать отдергивание. Это – отличается от классической интерпретации: рефлекс возникает в результате подкрепления (в данном случае отрицательного). Нет, подкрепление уже сопровождает базовый рефлекс отдергивания, а не является причиной появления рефлекса.
Описанный эффект хорошо наблюдать у кошек не слишком умных пород: они очень трудно поддаются обучению подкреплением, которое предполагает сопоставление произошедшего с событием подкрепления. Если попытаться отучить орать кошку по ночам методом кидания тапочка, то кошка воспринимается события поражения тапком в полном отрыве от своего мяукания, это – для нее – свое, особе событие. И сочетания “замяукала” - “получила тапком” не происходит ни на уровне формирования рефлексов (множественным повторением), ни на уровне формирования осознанных автоматизмов.
Более подробный разбор экспериментов, почему «условный рефлекс» на самом деле является в большинстве случаев автоматизмом можно посмотреть в fornit.ru/64936.
Обобщение фактических данных исследований по функции и механизмам ориентировочного рефлекса – границы между рефлексами и сознанием: Ориентировочный рефлекс. fornit.ru/44471.
Обучение с учителем
Мы вступаем в область все менее изученных феноменов организации индивидуальной адаптивности, со спорными предположениями многих физиологов о механизмах и их реализации.
Точками опоры все в большей степени остаются факты проявления этих механизмов, принципиальная логика их функционирования и, главное – фактическая верификация в виде действующей модели прототипа.
Чем глубже погружаемся в систему, тем меньше фактов и практически нет адекватных в системном подходе теоретических представлений ученых. Зато чем ближе к явлениям психики, тем в большей мере каждый оказывается способен сам непосредственно наблюдать и исследовать свою субъективность во всех ее переживаемых проявлениях. Такой метод всегда был у психологов на виду, и они пытались им пользоваться, но без достаточно надежных фундаментальных представлений о предшествующих механизмах и хоть и редких, разрозненных, но достоверных фактах исследований отдельных проявлений, это порождало довольно фантастичные представления.
Ключевые факты исследований удается выделить как нечто воспроизводимо общее в эволюции развития адаптирующих механизмов мозга, верифицируя такие фактические наблюдения психологов, у которых накопился огромный эмпирический материал, плюс к этому - самонаблюдения собственного субъективного мира. И здесь, как никогда, полезно опираться на схемотехническое мышление, позволяющее видеть способы создания схемных моделей этих явлений.
Пока что в формализации понимания системы адаптивности мы будем следовать методу интерполяции, формируя каркас модели понимания, основанный на воспроизводимых фактах, без необходимости экстраполяционных предположений. Интерполяция – хорошо обкатанный метод нахождения недостающих промежуточных звеньев по их известным соседним (эффект пазла), что дает достаточно определенное теоретическое предсказание. Так Д.Менделеев находил новые элементы по дыркам в системе уже известных, - именно система и позволяла это проделать. Экстраполяция использует лишь один из известных узлов системы и поэтому более рискованна, и если дырка содержит более одного звена, то такая экстраполяция оказывается все более предположительной.
Импринтинг
Как утята могут сразу при рождении запоминать вид утки, не ошибаясь в различных ракурсах? Это – пример того, насколько сложными могут быть цепи наследственно предопределенных связей, что позволяет смело предполагать столь же сложными наработки и в структурах механизмов психических процессов (упоминаемые ранее инфо-функции).
Понятно, что для реализации импринтигнга уже должны быть сформированы все необходимые слои распознавателей зрительных примитивов с их инвариантным (не зависящем от разнообразия масштабов и ракурсов) срабатыванием на образ. Другой вопрос: насколько точно при этом учитываются детали? Ведь особой точности и не требуется.
Утята обладают короткими периодами развития слоев мозга и вскоре после рождения уже готовы для запоминания образа своей утки. Сразу после рождения такое невозможно принципиально: нечем запоминать. Для импринтинга нужен свой критический период развития соответствующего слоя нейронов. Стоит его упустить, и характерная реакция не разовьется.
Так же не вызывает сомнения, что процесс фиксации памяти в импринтинге – особый вид похожего на условный рефлекс (раз условием является внешняя совокупность раздражителей), но очень облегченный, предопределенный наследственно. Т.е. наследственно задано, что в определенных условиях потребности (специализированный контекст восприятия-значимости), когда такой контекст оказывается активным, по некоторым признакам в окружающем выбирается то, что подходит по условиям как образ утки: существует особый, выделяющий важное контекст. Активность сочетаний сразу доступна для действий, а за какое-то достаточно небольшое время формируются постоянные связи от распознавателей этого образа к программе безусловных рефлексов следования за своей уткой и т.п. поведенческие реакции. На время фиксации связей возникшая активность должна самоподдерживаться.
Утятам нужно немного времени, чтобы стать готовыми к формированию образа утки. Такое обучение верифицируется признаками, которыми должен обладать такой образ по его возможному размеру, компактности признаков восприятия и динамике перемещения (утка не должна быть неподвижной). Стоит подменить утку любыми предметами или животными, подходящими под эти свойства, и рефлекс зафиксирует этот образ: возникнет поведение следования, подражания (fornit.ru/23469) и полного доверия во всем, хоть прыгать в воду, хоть в огонь (- не проверялось экспериментаторами).
Очевидно, что импринтинг имеет очень немалую подготовительную базу безусловных рефлексов, которые, в свою очередь, развиваются один на основе другого и в определенных условиях (fornit.ru/23470). Это – по-настоящему великое достижение эволюции при всем ее рукожопстве так, что мысленный эксперимент с печатающими обезьянами на деле показывает эффективность отбраковки смертью неудачного в достижении даже сложно сбалансированных между собой результатов.
Т.к. для нас это – невосполнимая в деталях становления загадка миллион летнего процесса отбора, то остается полагаться только на хорошо выявляемые факты в конечном результате. Но этих данных вполне достаточно и вовсе не обязательно следовать столь же бездумному перебору промежуточных результатов, хотя в познавательном плане это, несомненно, дало бы немало полезного. Какой бы природный механизм не начинают исследовать ученые с большой его детализаций, возникает много поучительного и много идей для реализации своих моделей.
Обычно под словом “обучение” понимается процесс запоминания действий, которые дают возможность получения целевого результата. Если запоминание не приводит к обладанию возможностью получить такой результат, то процесс неудачен (бесполезен, а то и вреден). Т.е. появляется понятие цели, желаемой при ее достижении. И нам предстоит найти схемы воплощения механизмов, использующих такую целенаправленность.
Логика верификации по такой оценке результата действий, как “истина” или “ложь”, позволяет корректировать обучение: если оценка не совпадает с ожидаемой целью (не адекватен), то блокировать, избегать впредь неудачный вариант реагирования.
Такой логики нет в импринтинге потому как, оказывается, легко можно ошибиться в условном образе, и это никак не корректируется.
И все же, импринтинг – базовый, основополагающий механизм целевого обучения по эталону (учителю), потому как у него задана цель того, к каким возможностям должен приводить результат: быть в области опеки и получать возможность без фатальных ошибок развивать адаптацию к условиям окружающего мира. В природе никто обычно не подменяет утку на движущийся мяч и все срабатывает прекрасно.
Если заменить утку на мяч, то импринтинг возникнет, но мяч не будет выполнять функции адаптивности к миру и утята будут лишены необходимых для выживания навыков. Только тепличные условия теперь могут оградить их от всего фатального.
Импринтинг запускает новую волну обучения: копирование действий по доверительному эталону, а для этого нужно, чтобы уже была возможность таким образом действовать (реакции отдельных действий уже были сформированы), т.е. существовала цель обучения в виде достижения эталонной последовательности действий.
Как связывается цель с имеющимися действиями и что вообще такое цель в схемотехнике организации эталонных действий - будет показано ниже. Мы придем к выводу, что утка это - уже не просто автомат, построенный на рефлексах, и все не так просто с обучением. Как утята, так и человечий ребенок, отлученный от родителей и общества, окажется беспомощным и нежизнеспособным, так что этот ранний период базовых форм обучения – очень важен для того, чтобы развиться до возможности обучаться самостоятельно, т.е. осознанно и при этом адекватно реальности.
Этот ранний период организован у всех достаточно высокоорганизованных животных и предполагает полную зависимость от того, как будут выполнять свою адаптирующую роль родители и окружающие особи данной стаи до момента развития возможности самостоятельных действий.
При необходимости начать разрабатывать искусственный организм, пришлось бы вшить все необходимые предопределенности для рефлекторного набора базовых реакций (что и было сделано в реализации Beast). Или же попытаться полностью изменить логику формирования поведенческих реакций, сразу сумев обеспечить целевые критерии верификации удачного или неудачного поведения. Вопрос в том, а что именно задать в таком качестве в организме? Система гомеостатической значимости этим не занимается, она обеспечивает поддержание жизненных параметров организма. Но организм сам по себе не существует, и нужно обеспечить параметры выживания всего вида, - некие критерии того, что для этого необходимо обеспечить в совместном и индивидуальном поведении. Это отбраковывается смертью тех, кто не обеспечивает адекватного реальности набора реакций, т.е. длительной эволюцией вида.
Более общий вопрос: возможно ли обойтись без такой эволюции вида, пока остается открытым. Мы не станем пытаться его решать. Но он дан в конечном виде: безусловные рефлексы сформированы эволюцией вида для определенных условий, в которых эти рефлексы образуются и достигают функциональной цели (не вылететь из отбора). Остается проследить системные принципы организации всех последовательностей адаптивных механизмов, уже найденных природой, а не пытаться придумать что-то свое, и не только потому, что пример искусственных нейросетей показывает, чем заканчивается такое придумывание без учета уже обкатанного природой.
Хотя в природе роль учителя начинается исподволь – в организации базовой системы безусловных рефлексов и совсем не так, как даже мечтают это воплотить теоретики искусственных нейросетей, но пока это – случайно оставшаяся в живых особях цель, а не механизм целеобразования в схемах реализации индивидуального обучения, т.е. называть целенаправленным безусловные рефлексы можно (слово все стерпит), но так же не корректно, как называть живым кирпич (хотя некоторые мистики его таким полагают).
Одним их важнейших целенаправленных (на границе того, что позволяет говорить о цели) рефлексов является подражание (fornit.ru/6359). Оно дополняется все более изощренными механизмами (fornit.ru/6361) и приводит к организации возможности перенимать чужие навыки (fornit.ru/6362) на самом высоком уровне субъективных моделей понимания. Этот механизм создает автоматизмы, копируя реакцию авторитета. В реализации Beast этим организован предосознательный уровень формирования базовых автоматизмов на стадии развития “доверчивое подражание”.
Подражание
Импринтинг и подражание - разные вещи: (fornit.ru/67851).
Учитель жизни детеныша определяется импринтингом или целой системой доверительного отношения к особе, которая защищает от невзгод и формирует навыки приспособления к различным условиям жизни.
После этого получают возможность активироваться и действовать другие рефлексы, обеспечивающие подражание действиям опытной особи, к которой есть полное доверие.
Самый простой подражательный рефлекс – следования за родителем или остальными членами стаи.
Само по себе имитировать что-то из чужого поведения – одна из базовых реакций (безусловно-рефлекторной природы). Мы невольно улыбаемся, видя чужую улыбку – рецидив детской реакции, хотя усилием воли может преодолеть это побуждение, если обратим на это внимание или уже приучили себя к сдержанности.
Но просто имитация чужого поведения дает возможность видеть, какие ответные реакции требуется начинать формировать. Вначале это проделывается рефлекторно, но полезность этого метода обучения настолько высока, что он перешел в форму осознанной имитации в случае освоения нового, когда уже есть критерий верификации результата реакции и сами реакции следуют этому критерию, а не просто базовым условиям формирования связей.
В реализации Beast есть стадия развития, предназначенная для “отзеркаливания” авторитарных реакций в данных условиях после определенного стимула – с формированием базового автоматизма. Эта стадия не требует осмысления и оценки верности реакции потому, что авторитету в этот период развития нужно доверять. Но такие автоматизмы, в отличие от условных рефлексов, уже могут быть заблокированы если при попытке их использования получен негативный эффект. И такие автоматизмы формируются за один раз, не требуя многократных повторений. Время, необходимое для закрепления связей оказывается достаточно потому, что при формировании такого автоматизма срабатывал ориентировочный рефлекс и объект внимания начинал удерживаться в активном состоянии реверберацией. Именно такое удержание позволяет ожидать последствия и оценить успешность.
Период доверчивого обучения проходит стадию доосознанного, рефлекторного подражания и затем переходит в стадию осознанного (по ориентировочному рефлексу) отзеркаливания чужих действий, достигающих актуальной цели. При этом есть промежуточный этап подготовки для осознанного подражания, что важно проследить.
Рассмотрим наиболее общий механизм схемотехнической организации подражания как рефлекса, обеспечивающий условия возникновения более высокоуровневых производных рефлексов. При этом будем продолжать строить системную логику на основе ключевых фактов, а не станем гадать о правдоподобности имеющихся теорий вариантов предположений природного воплощения потому, что (fornit.ru/23471):
“К сожалению, у всех явлений подражания в указанном смысле нет физиологического объяснения! К наблюдаемому многообразному и в то же время единому биологическому феномену не подобрано физиологического ключа. Мы касаемся здесь "дна" (если угодно, "потолка") современной науки о физиологии нервной деятельности. На рецепторы и афферентные пути данного организма не падает никаких раздражении, которые могли бы рефлекторно породить такой-то двигательный эффект; рецепторы воспринимают только сам этот эффект, видимый или слышимый, в поведении другого организма; тем не менее первый как-то отождествляет себя со вторым: повторяет, копирует с него следствия, не испытав соответствующих причин. Физиологическая наука не докопалась до механизмов этого рефлекса. Хотя не сомневается в рефлекторной природе автоматической имитации поведения у животных. Физиологи Попов, Хотин, Орбели, Воронин, Слоним снова и снова вынуждены были признать, что загадка подражания не расшифрована”.
Конкретика реализации механизма рефлекторного обучения не играет принципиальной роли в общей строящейся нами модели и служит для иллюстрации схемотехнического подхода, так что не будем напрягать усилия, чтобы выверить его досконально (тем более, что в реализации Beast этот механизм работает). В очередной раз демонстрируется преимущество схемотехнического подхода, без которого физиологи не смогли подступиться к механизму, несмотря на открытие “зеркальных нейронов”, потому что дает реальные основания для построения действующей схемы взаимосвязанных в иерархии наследования механизмов.
Выбранным языком описания у нас будет причинно-следственная схемотехника и то, что такой язык в наибольшей степени удобен и лаконичен по сравнению со всеми другими (тем более математическим) будет ясно в конце и отражено в конечном обобщении.
Модель механизма подражания
Первым этапом для подражания является привлечение внимания к тому, что необходимо скопировать собственными реакциями. Т.е. необходим механизм фокусировки внимания - как наиболее узкий контекст восприятия и действия в контексте текущей проблемной ситуации - цели (fornit.ru/67888).
При моделировании схемы будем учитывать, что никакого осознания, никакой произвольной концентрации внимания и произвольности оценки успешности на уровне самых простых подражаний пока нет и в помине, нужно стараться не попадать в искушение полагаться на это.
Контекст определенной цели запускается необходимостью что-то совершать, в конечном счете - для компенсации гомеостатического разбаланса (учимся клевать корм, когда голодны или прятаться от опасности и т.п.), когда в восприятии возникают признаки, уже связанные с опасностью или не понимаемые, но поведение учителя вызывает состояние опасности.
Контекст подражания возникает при активации цели, требующей каких-то действий, но без наличия уже готовой реакции на этот случай. А это означает необходимость распознавания нового в данном контексте (возможно, в наиболее примитивном варианте реализации) и активацию ориентировочной реакции. Проще говоря, если в данной ситуации еще нет привычной реакции, то нужно ее подсмотреть у авторитета.
Если рядом есть более опытная особь, к которой имеется доверие (родитель или свой из стаи), то внимание удерживает и образ, отражающий ее поведение в каждые моменты ее действий до достижения целевого результата ее действий (зерно склевано, от опасности спрятались).
Здесь нет осознания законченности, как нет и осознания необходимости (цели) подражать. Есть контекст нового после контекста значимости удовлетворения какой-то потребности (или без него, а просто – факт необычного поведения свояка, вызывающий ориентировочный рефлекс).
Активность образа с новизной вызывает подготовку связей (рост дендритов, если их еще там нет). Это – условная причина.
Следом оказывается активным и специализированный распознаватель запуска действий, схожий с тем, что есть у учителя, и это – самый важный и интересный момент. Это – элемент будущей собственной модели понимания (fornit.ru/7496), отражения реальности:
“оказалось, что эти клетки срабатывают не только тогда, когда обезьяна видит чужие манипуляции с предметами и не только тогда, когда она наблюдает социальные контакты, но и тогда, когда она видит взаимодействие неодушевленных предметов; иными словами, когда макакам показывали в кадре столкновение двух игрушек, у обезьян включались те самые зеркальные нейроны”.
Если провести условный раздел в головном мозге, оставив теменную ассоциативную кору со стороны задних долей, то дальше идут моторные поля программ действий, запускаемых рефлексами ассоциативной теменной коры.
![]()
Система нейронов, обладающих “зеркальной” функциональностью, располагается по обе стороны от двигательных программ:
![]()
Теменная часть “зеркальных нейронов” образована распознавателями образов того, что нужно отзеркаливать – внешние признаки поведения свояка, а лобная – соответствующими собственными реакциями. Понятно, что такая связь должна была образоваться заранее, как связь между видимыми проявлениями чужих реакций и своими действиями, которые приводят к таким же внешне наблюдаемым проявлениям.
Можно легко представить множество вариантов процесса образования такого соответствия на чисто схемотехническом уровне. Такие коллекции сочетаний, как заготовки возможных реакций, формируются, а затем используются в подражательной деятельности.
Причем, видимо, есть несколько усложняющихся уровней такой организации, начиная с очень простой связи между воспринимаемыми признаками чужих и своим действием, например, для реакции следования за другим. Все это – результат развития слоев уже за двигательной зоной – развития слоев лобной коры (fornit.ru/6347), которое начинается позже первичных зон восприятия, но достаточно рано в онтогенезе.
И это – задел для альтернативного рефлекторному механизма формирования ответных реакций. Здесь важно, что это – первые отражения чужих и собственных действий, как и вообще отражение причинно-следственных зависимостей в окружающем мире. Начальный механизм таких связей – условно-рефлекторный, который затем заменяется на механизм формирования автоматизмов. Первые сочетания условного символа восприятия (словесного или невербального: жесты, мимика и т.п.) с возможной реакцией (в контексте смысла лобной коры) начинают формировать систему социальной коммуникации еще задолго до того, как начинает функционировать сознание. Но это – тема последующих глав.
В реализации Beast подражание организовано следующим образом: если в данных условиях нет привычной реакции, то самым простым методом отзеркаливания является – повторение текущего стимула в виде собственного действия для того, чтобы подсмотреть, как на него прореагирует оператор и доверчиво скопировать такую реакцию. В более сложной реализации всегда смотрится, как и на что реагирует оператор, что учитывается как авторитарно усвоенное Правило.
Теперь должен бы быть вполне ясен общий результат возникновения первых рефлекторных реакций подражания при сочетании новых образов ситуации, выделенной в контексте значимости ориентировочной реакции, и активирующихся в этом контексте отражателей действий эталонного авторитета, связанных с программами аналогичных собственных действий.
Конечно же, возникнув, механизм отзеркаливания уже не ограничивается отражением связи наблюдаемых проявлений чужих действий со своими программами таких же действий, а отражает все появляющиеся особенности усложняющихся механизмов адаптивности, в том числе эмоциональное переживание другого сопоставляется с собственными там, если они есть (иначе чужие переживания просто не воспринимаются).
Социализация, совместное поведение всех уровней сложности основывается на механизме обучения полезному у чужого.
При этом условием активности “зеркальных” нейронов, конечно же, является наличие контекста значимости, в котором данные действия чужака воспринимаются как соответствующие потребности, вызвавшей такой контекст. Не будет собственной потребности – игнорируются и чужие такие действия.
Назовем период формирования таких заготовок - периодом доверчивого обучения.
Периодизация психического развития с точки зрения онтогенеза функциональной асимметрии полушарий (fornit.ru/1036):
“Физиологически критический период характеризуется "преобразованием одного доминантного состояния, свойственного предыдущему возрастному периоду, в существенно новое доминантное состояние, требующееся в последующем возрастном периоде" (И.А.Аршавский [6]). Критичность развития ВПФ, приуроченного к определенному периоду, проявляется в необратимом, как принято считать, угасании возможностей эффективного развития соответствующих способностей после перехода возрастных границ данного периода (Я.Л.Коломинский [7]).
... На каждом из нормативных кризисных этапов происходит выбор позитивного либо негативного варианта развития определенных личностных черт, определяющих в дальнейшем жизненную позицию человека, его отношения в социуме. Если внутренние конфликты, свойственные определенной стадии развития, остаются неразрешенными (негативный вариант развития), то в зрелом возрасте они могут проявляться в виде инфантилизма. При этом новые выборы накладываются, наслаиваются на уже произведенные и закрепленные в структуре личности.
... феномен сенситивности свойственен не только человеку и имеет общебиологическое значение (наиболее ярким примером является импринтинг). Биологический смысл сенситивности заключается в следующем: если событие оценивается как статистически редкое, возможно встречающееся впервые, то соответственно информация о данном событии представляет несомненную ценность для адаптации индивида и подлежит сохранению”.
Период доверчивого обучения, включающий период инициативы
В диссертации “Влияние особенностей семейного воспитания на социальную адаптированность детей” (fornit.ru/246):
“Младший школьный возраст характеризуется ещё полной психологической зависимостью от взрослого. Семья формирует и ¶определяет поведение индивида, закладывает план и структуру поведения, которые в значительной мере сохранятся в течении жизни, а также оказывает модифицирующее влияние на поведение в каждый определённый период времени.
...Особенностью детей младшего школьного возраста является «безграничное доверие к взрослым, главным образом учителям, родителям, подчинение и подражание им» (97,172). Дети этого возраста полностью признают авторитет взрослого человека, почти безгранично принимают его оценки, перенося их на себя и присваивая их себе. Даже характеризуя себя как личность, младший школьник, в основном лишь повторяет то, что о нём говорят взрослые”.
В работе Доверие дошкольников к незнакомым взрослым (fornit.ru/376):
“... С позиции рассматриваемой теории развития общения в онтогенезе, возникновение доверия ребенка к другому человеку может определяться уровнем удовлетворения его коммуникативной потребности в процессе общения с этим человеком.
... Примечательно, что большая часть детей пяти лет имеет повышенные показатели доверия, а семи лет — низкие значения доверия.... Резкое снижение доверия детей от пяти к шести годам позволяет говорить о том, что в данный период дошкольного возраста происходит существенное изменение в развитии личности ребенка”.
В статье детского психолога Этапы жизни ребенка: от доверия до активности (fornit.ru/1675):
“Первый период базового доверия (или недоверия) к миру. Он продолжается от рождения до 2 лет. У новорожденного еще нет никаких представлений ни о себе, ни о том мире, в котором он оказался. Зато у него есть определенный набор основных потребностей, без удовлетворения которых он не выживет: еда, тепло, присутствие мамы, общение. Если все это малыш получает по первому требованию, то у него формируется стойкое ощущение, что мир, в котором он оказался – хорошее место, которому можно доверять. Наиважнейшее, или как говорят ученые – базовое чувство, без которого просто невозможно дальнейшее развитие личности.
В этот период весь окружающий мир малыша сосредоточен в одном емком понятии – мама. И если он чувствует, что маме можно доверять, потому, что она накормит, успокоит, всегда будет рядом, не оставит его, маленького и беспомощного надолго одного, то в дальнейшем, подрастая, ребенок перенесет это чувство доверия и на остальных людей, на остальной мир.
... Второй период называют Периодом автономии (от 2 до 4 лет). Это знаменитая «фаза упрямства», фаза «Я сам», так хорошо знакомая родителям трехлеток. Но не надо думать, что это только лишь упрямство, малыш примеряет себя к миру взрослых, осваивает новые умения и навыки, так необходимые ему. Чем больше багаж этих навыков, тем чаще его охватывает первое чувство сомнения «А хорошо ли я делаю?». Считается, что именно в этом возрасте возникает первое чувство стыда.... ... Если в этот период родители стыдят и ругают малыша за испачканные штанишки, за разбитую чашку или за испачканную мордашку при попытке есть самому (поводов, за что именно ругать в этом возрасте, согласитесь, предостаточно!), то сильно тем самым раскачивают уверенность маленького человечка в том, что он что-то может сделать САМ. К такому же результату приводят и чрезмерные требования к «правильности» поведения, которыми так грешат многие родители. Чувство стыда и ощущение собственной неумелости прочно ложатся в основу личности, характера и начинают пускать свои корни во всю дальнейшую жизнь.
... Период инициативы (от 4 до 6 лет). Это очень важный период самоутверждения личности. Малыши в этом возрасте крайне деятельны. И игра в этот период не просто игра – а важная школа взрослой жизни, создание своего мира, со своими законами и правилами. Ребенок этого возраста очень нуждается в поддержке и одобрении со стороны взрослых. Благодаря теплому эмоциональному отклику, поддержке он убеждается, что может достигнуть поставленной перед собой цели и на многое способен. Малыш примеряет на себя такое важное для всей дальнейшей жизни понятие «Могу – не могу».
... Период активности охватывает период от 6 до 11 лет. Ребенок уже – школьник и это уже по-настоящему вводит его в социум. В этом возрасте ваш ребенок начинает сознательно работать «на результат», учится объективно (сравнивая с другими) оценивать себя и плоды своих трудов. В случае положительной оценки он начинает получать истинное удовольствие от удачно завершенного дела. Поддержка и поощрение взрослого в этот период важны особенно”.
Можно видеть, что развитие доверия и инициативы идет как бы по спирали, повторяясь, но на более высоком уровне сложности (интегрированности признаков) условий, к которым вырабатывается адаптация. Всякий раз закладывается посильная на данном этапе сложность формирования соответствующих представлений и навыков, проверяется жизнью, и корректируется уже на более зрелом уровне контакта с социумом. Такие проявления психики как доверие, вера, скептицизм - следуют этим этапам, так же развиваясь. Это отражает базовую логику последовательного усложнения рецепторных и эффекторных распознавателей.
В качестве дополнения стоит учитывать, что рефлексы образуются в любых контекстах значимости, если для данного сочетания условий нет привычной реакции более высокого уровня. В негативных контекстах ответные реакции формируются для действий по восстановлению аварийных значений жизненных параметров, а в позитивных, когда параметры приходят в норму, формируются реакции спокойного состояния и те, что предотвращают опасности и потерю жизненного потенциала. Одной из таких реакций является сон. Это состояние по классификации физиологов относится к тормозным условным рефлексам. Для ее начала нужны определенные условия. Сон – сложное состояние организма, специфический контекст состояния организма, выполняющий несколько разных функций. Причем у разных видов животных некоторые из них очень своеобразны, но есть и общие, о чем будет сказано в теме про сон.
Новизна и значимость
Сразу же зададим контекст понимания логики соотношения привычного и нового.
Имеющиеся ответные реакции, приспособленные к определенным условиям, не требуют какой-то корректировки, они входят в базу ответного реагирования, не требуя внимания к себе в данных условиях. Но те же реакции в других условиях могут не принести удачного результата или даже оказаться вредными. Если тыкаться носом в пушистую маму, это – привычно хорошо, но если пушистое окажется не мамой, а хищным зверем, питающимся вот такими глупыми детенышами, случится фатальная ошибка, исправимая только перезагрузкой жизни особи.
Даже для детенышей, которые уже все, что можно полезное переняли от мамы, при попытке действовать самостоятельно выясняется, что жизнь настолько полна неожиданностей, что буквально ни один совет мамы просто так не срабатывает без порой болезненных неожиданностей потому, что постоянно все меняется и это новое просто необходимо как-то учитывать или же сидеть в строго привычных условиях, платя жизнью за каждый шаг в сторону от привычного, что и происходит у всех простых животных, не умеющих учитывать новое.
Итак, неизбежным шагом адаптации после импринтинга и перенимания чужого опыта является умение обращать внимание на достаточно значимое новое (о чем уже говорилось) и способы модифицировать свое поведение в зависимости от новых условий.
Здесь 1) жизнью проверяется баланс: простое и легко уничтожаемое, но зато легко и быстро воспроизводящееся (вирусы, бактерии, грибы) или 2) сложное и затратное, но стабильно приспосабливающееся. И это во многом равноценно в плане выживания вида.
В реализации Beast различаются два вида новизны:
1. для данных условий еще нет привычной реакции
2. реакция есть, но в образе восприятия найдены новые значимые признаки, ставящие под сомнение привычную эффективность реакции. В этом случае используется значимость образов восприятия, определенная прошлым опытом.
Ориентировочный рефлекс различает эти две ситуации, подключая разные функции обработки.
В обобщающей статье (fornit.ru/1195) по исследованиям психологии внимания:
“Ориентировочный рефлекс связан с адаптацией организма к меняющимся условиям среды, поэтому для него справедлив "закон силы". Иначе говоря, чем больше изменяется стимул (например, его интенсивность или степень новизны), тем значительнее ответная реакция. Однако не меньшую, а нередко и большую реакцию могут вызвать ничтожные изменения ситуации, если они прямо адресованы к основным потребностям человека. Кажется, что более значимый и, следовательно, в чем-то уже знакомый человеку стимул должен при прочих равных условиях вызывать меньшую ОР, чем абсолютно новый. Факты, однако, говорят о другом. Значимость стимула нередко имеет решающее значение для возникновения ОР. Высоко значимый стимул может вызвать мощную ориентировочную реакцию, имея небольшую физическую интенсивность”.
Здесь есть некоторая нелогичность: даже очень высокая значимость при нулевой новизне не вызывает ориентировочную реакцию. Даже очень большая опасность, если она раз от раза не оказывает действия, все менее учитывается. Но стоит заметить, что не бывает совершенно идентичных ситуаций, всегда есть какая-то новизна. И наоборот, даже при очень слабой значимости большая новизна привлечет внимание.
Если говорить о формулировке взаимосвязи, то лучше будет сказать: сила ориентировочного эффекта пропорциональна произведению новизны на значимость. Это учитывает оба варианта в реализации Beast: малая значимость, но отсутствие реакции (большая новизна) и большая новизна при имеющейся привычной реакции.
Роль контекста нового
С одной стороны, ориентировочный рефлекс подавляет другие рефлексы (fornit.ru/23481) потому как нужно не опрометчиво реагировать, а оценить опасность, но при этом усиливает эффективность формирования нового рефлекса:
“В лаборатории И.П. Павлова также были получены первые факты о влиянии ОР на формирование условного рефлекса. Во время процедуры выработки условной связи, а также при формировании ассоциации между двумя индифферентными раздражителями было отмечено усиление или появление ориентировочных реакций. В опытах И.О. Нарбутович и Н.А. Подкопаева [1936] с выработкой у собаки ассоциации в виде двух последовательно следующих индифферентных стимулов (звук – свет или вертушка – прерывистый шум) было показано, что при отсутствии на них ориентировочных реакций ее формирование идет плохо”.
Здесь, кстати, видно, как путаются механизмы условного рефлекса и более высокоорганизованного механизма осознанно сформированного автоматизма.
Функции ориентировочного рефлекса и проявление механизмов их реализации изучались Е. Соколовым (fornit.ru/5134) и О.Виноградовой (fornit.ru/5371) (см. Функции и механизм "ориентировочного рефлекса»: fornit.ru/5400). Центральным органом мозга, в котором сосредоточены рецепторные и эффекторные основы этого механизма, является гиппокамп. О.Виноградова в фундаментальной книге “Гиппокамп и память” собрала огромное количество данных исследований, которые сегодня в совокупности с более новыми позволяют их сопоставить и выделить обобщающие системные основы принципов функции новизны и значимости.
Вот большая подборка фактических данных исследований: Новизна и значимость: fornit.ru/ax1-35.
Во всем этом особо важна роль детекторов нового в текущем контексте значимости, которые подключают более узкий контекст ориентировочной реакции на новый значимый раздражитель. Это формирует новую реакцию, если раздражитель отвечает каким-то потребностям, т.е. уже есть автоматизм, удовлетворяющий эти потребности, но он срабатывал от других раздражителей, но вот появился новый, в образе которого значимость, связанная еще во вторичной зоне, соответствует контексту данного состояния организма. Если при повторениях такого сочетания автоматизм сформируется, то детекторы нового уже не будут срабатывать, а реакция станет привычной, не требующей более коррекции.
Отсутствие новизны в данном узком контексте, не вызывая ориентировочного рефлекса, не создает условия для закрепления новой реакции через механизм связи с лобной корой. Возможно только закрепление условно-рефлекторных реакций, но, как уже замечалось, чем сложнее ситуация, тем реже она проявляется: числа сочетаний оказывается недостаточно для надежного закрепления новых связей или конкуренции подавления старых связей. Вот почему “при отсутствии на них ориентировочных реакций ее формирование идет плохо” – т.е. по механизму условного рефлекса, требующего многократных повторений.
Отсутствие новизны в узком контексте значимости как бы предохраняет эту реакцию от дальнейшей модификации (так же, как игнорируются реакции в иных контекстах), в том числе от тормозных влияний других реакций, от факторов, способных ее тормозить до полного угасания. Реакции, сформированные в условиях значимой новизны, более стабильны и не утрачиваются от факторов, которые приводят к угасанию незащищенные контекстом реакции, если эта реакция долгое время оказывается не востребованной – по причине формирования автоматизма, а не условного рефлекса. Автоматизмы не обладают свойством дезадаптации как угасающие условные рефлексы потому, что у них есть свой механизм блокировки – в случае негативных последствий в данных условиях.
Получается основа для состояний памяти “привычное и новое”, где только новое позволяет эффективно и быстро модифицировать привычную реакцию - как ветвление привычного в новых условиях. После формирования такого ветвления оно также становится привычным, а контекст в точке ветвления оказывается расширенным уже на два направления, которые выбираются в зависимости от условий (дерево состояний получает новое условие).
Появление новых условий никак не может быть запрограммировано заранее и обеспечено впрок наличием нейронов, готовых специализироваться как образы в текущем контексте с участием новых условий. Это требует, чтобы последние слои нейронов в коре не были под запретом на формирование профиля возбуждения (возможно, просто тем, что больше не возникает последующего слоя), или же требуется пополнения слоя новыми нейронами с некоей частотой, коррелирующей с наличием актуальной новизны (т.е. чтобы контексты необходимых коррекций состояния организма были активны). Причем, это касается как области теменной ассоциативной коры, так и лобной, там, где формируются нейроны целевых моделей, отражающих значимые свойства окружающего, связанные с собственными возможностями взаимодействия с окружающим – “зеркальные нейроны” (fornit.ru/23504).
Фактические данные говорят, что способность модифицировать связи остается на уровне последних слоев третичных теменной и лобной зон, а новые нейроны возникают в области механизма коммутации актуальных (значимое новое) образов этих зон в канал внимания к новому. Возникновение новых нейронов обнаруживалось в этих областях и неоднократно это подтверждалось, см. подборку: регенерация нейросети: fornit.ru/ax1-55.
Итак, есть автоматизмы, которые формируются без осмысления в процессе отзеркаливания чужого действия, и такие базовые автоматизмы очень похожи на условные рефлексы, но обладают новыми адаптивными свойствами.
Т.е. канал внимания к новому является конкурирующим со связями образов, запускающими рефлексы и, при активации связи с лобными долями, моторное действие сразу не запускается (ориентировочный рефлекс второго типа), а опосредуется элементами лобной коры, что приводит к эффекту готовности реагирования без самого реагирования (целевой возможности реагирования).
Эти отличия придают трем основным видам реакций качественно различные свойства.
1. Простой (классический) условный рефлекс: создается за несколько сочетаний, находится в широком контексте значимости среди множества других равноценных рефлексов данного состояния (стиля реагирования) организма. Он легко угасает при невостребованности.
2. Базовый автоматизм (требующий проверки собственным опытом), образованный в контексте новизны в данном контексте значимости, образ стимула самоподдерживается замыканием выхода на вход распознавателя системы значимости данного контекста. Он оказывается в предельно узком контексте значимости и поэтому его угасание затруднено. При его активации замыкается самоподдерживающая закольцовка, и он сохраняет образ, запускающий реакцию для ожидания последствий и их оценки.
Вспомним: “распознаватели нового в данном контексте не локализованы в теменной ассоциативной зоне, а они сконцентрированы и общем слое гиппокампа для обслуживания образов теменной ассоциативной зоны и связи их с другими структурами”.
Связи, обеспечивающие сравнение активных признаков восприятия в данном контексте с ранее зафиксированными (запомненными как привычное), при рассогласовании (как результат сравнения) активируют нейрон новизны, который открывает переключатель гиппокампа для закольцовки образа и удержания его в “памяти” с укреплением связей между активными элементами (формирования “постоянной памяти”).
Переключательная функциональность нейронов гиппокампа демонстрируется в протезах реальных систем восстановления работы гиппокампа: fornit.ru/5221, fornit.ru/5291, fornit.ru/6329. Схема такой функции нейрона очень проста: переключатель это - вставочный (промежуточный) нейрон, связывающий выход предшествующего нейрона со входом последующего, а к вставочному нейрону приложен тормозной сигнал, который не позволяет ему активироваться, пока на нем есть потенциал, т.е. переключатель оказывается заперт. Такая схема требует постоянного поддержания всех ключей в запертом состоянии активными тормозными сигналами, чего можно избежать, если посчитать, что таким активирующим неспецифическим сигналом является активность текущего контекста значимости, оказывающая латеральное торможение на все переключатели в ее области. Другой, более соответствующий природному стилю реализации, вариант – переключательные нейроны (fornit.ru/24207), которые образованы собственными возвратными коллатералями аксонов, постоянно поддерживающими тормозную активность. Здесь уместно вспомнить и относительно недавно открытый механизм обратных тормозных связей в синапсах (гиппокамп, прилежащее ядро) на основе эндоканнабиноидных CB1-рецепторов.
При установлении связей между активными компонентами, включая переключатель закольцовки, рефлекс, срабатывая от условного стимула в контексте значимости, запускает и самоподдержание образа, тем самым оставляя его контекст активным даже после завершения цепочки эффекторного реагирования (пока его что-то активно не затормозит), что обеспечивает возможность оценки результата реакции, но и не только, а обеспечивая возможность обобщенной оценки значимости последовательности совершаемого, формируя Правила – как цепочки звеньев типа Стимул-Ответ-Эффект.
3. В отличие от базового автоматизма, который кроме заготовки отзеркаленной привычной реакции, стандартный автоматизм имеет связь с нейронами модели собственной (а не чужой авторитарной) целью действия. Такая связь замыкается так же переключателем при активировании нейронов новизны, как и закольцовки образа в теменной ассоциативной зоне.
В случае новизны автоматизм не запускает сразу свою привычную моторную реакцию, но активирует образ в лобной коре (“зеркальный нейрон” или модель ответного действия), от которого есть возможность запуска действия при дополнительном разрешающем сигнале.
Ориентировочный рефлекс, подключая канал связи с лобными долями, отключают запуск более низкоуровневой привычной реакции.
Т.е. сформированная с участием ориентировочного рефлекса автоматизм запускается со стороны лобной коры. Получается так, что автоматизм формируется как результат произвольного решения в лобной коре: запускать или нет данную реакцию в данных условиях новизны. Такое решение закрепляется в виде приторможенного автоматизма или нового созданного автоматизма с альтернативным действием, после чего уже не требуется подключение механизмов нахождения решения в лобной коре (т.е. минуя активацию переключателей в гиппокампе, гиппокамп становится не нужен для уже закрепленного автоматизма), а просто реакция срабатывает автоматически с эффективностью рефлекса. Если же в данном контексте опять появилось новое и срабатывает ориентировочный рефлекс, он тормозит выполнение уже привычного действия на время, необходимое для решения механизмами произвольности, что делать с этим дальше.
Это явление назвали “внешним торможением” рефлекса (fornit.ru/7511) при том, что это уже – вовсе и не условный рефлекс:
“Внешнее торможение возникает под действием посторонних к образуемому условному рефлексу раздражителей. Посторонний к опыту раздражитель, особенно новый и сильный, вызывает ориентировочный рефлекс, и возбуждение, относящееся к этому рефлексу, тормозит вырабатываемый условный рефлекс до тех пор, пока посторонний раздражитель не исчезнет или не потеряет новизны” или не будет решено, что делать при этом.
Можно заметить, что такое “внешнее торможение” (а торможение ли это? или просто прерывание до решения) никак не соответствует механизму условного рефлекса, а присуще именно механизму реагирования на “ориентировочный рефлекс”. То, что до сих пор называют условным рефлексом, в данном случае представляет совершенно иную организацию.
Это прерывание реагирования вниманием к актуально новому и составляет основу для всех возможностей произвольности, которые развились в очень сложный алгоритм механизмов решения, как именно реагировать в данном случае, который будет рассмотрен в теме Сознание, но сначала необходимо будет подробно рассмотреть темы Произвольность и, затем, Автоматизмы.
Ориентировочная реакция всегда прерывает то действие, которое затрагивало текущее выделение наиболее актуального (fornit.ru/43673). Если мы идем и видим впереди появившуюся лужу, то это не прерывает движение пока мы приближаемся к ней, но прогноз выделяет лужу как наиболее актуальное, и уже тормозит дальнейшее движение напрямик, подключая канал осознания ситуации для поиска решения. Если видим, что лужу можно обойти, то меняем прежний стереотип и изменяем путь (сознание отработало в отслеживающем режиме), если нет – останавливаемся и задумываемся более глубоко. Если лужа будет встречаться постоянно на этом месте, вырабатывается новый автоматизм, уже не требующий осознания (fornit.ru/43674).
Существует много данных исследований этого явления: fornit.ru/24210, fornit.ru/24209, fornit.ru/6352.
Чем меньше наработанных автоматизмов, тем чаще прерывается деятельность, что проявляется в виде СДВГ (fornit.ru/1317) со времени становления механизма привлечения внимания значимой новизной, с возраста 3-4 года. Т.е. такое прерывание характерно именно для ориентировочного рефлекса привлечения внимания, а в возрасте до 3 лет условные рефлексы так не прерываются, их выполнение можно только насильно прервать: попробуйте по сопротивляться удару под коленную чашечку или условному рефлексу слюноотделения на вид лимона. Вспомнилась история, как суфлер решил наказать певца оперы и принялся выразительно грызть лимон в своей суфлерской яме, пока певец не подавился. Но, с другой стороны, можно и не отдернуть руку от горячего, если крепко задумался, психические рекции перекрывают рефлексы.
Образ восприятия-значимости остается активным не только на время привлечения внимания в теменной ассоциативной коре, но и после отключения от канала связи с лобной корой, сохраняя “кратковременную память” о прошедшем событии и за это время фиксируя связи такого “эпизодического воспоминания”. Именно про такой образ в случае подключения канала внимания (связи с лобной корой) А. Иваницкий писал (fornit.ru/7446), что он является основой субъективного восприятия потому, что по временным задержкам распространения сигналов по связям он четко коррелирует с появлением субъективного переживания. На самом деле все значительно сложнее и интереснее.
Отработку привычных реакций в зависимости от условий и стимулов в самом общем плане демонстрирует анимация “Роль гиппокампа в удержании образа и ветвлении вариантов в зависимости от условий”: fornit.ru/an-book-12. Это – демонстрационная визуализация схемотехнической интерпретации механизма для тех, кто пока затрудняется сам легко вообразить такие взаимосвязи.
В прошлой главе было показано, как контексты, означающие определенное состояние организма и определяющие стиль его восприятия и реагирования, выделяют только то, что значимо для данного стиля, имеющего задачу восстановления параметров гомеостаза, т.е. определяют контекст гомеостатической значимости. Эти контексты (начинаясь от самого общего, организованного балансом нейромедиаторов, до сужающегося признаками все более частных условий, которые были связаны со значимостью во вторичных зонах мозга) в третичных (ассоциативных) зонах связаны как со значимостью, так уже и с каким-то запуском рефлекторных действий. При этом канал контекста, ограничивающего восприятие для данного рефлекса, оказывается максимально сужен. Если же в этом суженом контексте значимости появляется новый стимул (ранее связанный с того же типа значимостью и поэтому активный в данном контексте), то срабатывает специализированный распознаватель такой ситуации – “нейрон новизны”, создающий условия для формирования новой реакции: новый стимул замыкается в самоподдерживающее кольцо, чтобы обеспечить его активность, даже если на время пропадает первичный стимул (значимый признак хоть и исчез, но не перестает учитываться уже “по памяти”). Это обеспечивает достаточное время для создания связи нового автоматизма, т.е. связь образуется при единственном предъявлении, не требуя множества повторений сочетания условного признака с ответной реакцией.
При этом наличие специализированного контекста значимости намного лучше обеспечивает сохранность такого автоматизма (раз контекст предельно сужен, у реакции нет конкурентов) по сравнению с более простыми рефлексами более широкого контекста значимости, где многое может оказывать тормозящее, конкурентное влияние.
С каждым новым автоматизмом широта охвата контекста, в котором оказывается все большее число образов, увеличивается, а новые реакции, в максимально узком пока контексте, оказываются наиболее стойкими.
В реализации Beast применено дерево автоматизмов (fornit.ru/beast5), где для каждого нового условия есть место для формирования новой ветки и связи ее с автоматизмом. В этом дереве используются конечные образы воcприятия, имеющие уникальное сочетание условий и поэтому нет необходимости доращивать новый уровень в дереве при появлении нового условия, а достаточно иметь один уровень для всех уникальных образов. Чтобы лучше это представить нужно бы внимательно ознакомиться со схемой реализации Beast раздел Автоматизмы: fornit.ru/64975)
Конкретизируем логику соотношения компонентов “значимое” и “новое”.
1. Если нет никакой значимости (все в порядке с организмом, ничего не нужно), то и нет никаких раздражителей, т.к. отсутствие контекста = игнорированию всего. Не нужно сужать внимание на чем-то, все привычно сработает само, как только появится пусковой стимул, активируя ответную программу действий.
2. Если нет никакой новизны, то так же все будет оставаться на достигнутом уровне. Новизны не может быть никакой, если не активирован хоть какой-то контекст значимости.
Такие свойства выявляют функцию модуляции новизны значимостью, т.е. реализуется операция перемножения: если один из компонентов – ноль, то произведение – тоже ноль.
В контекстах значимости новизна распознается просто сопоставлением образа узкого контекста с профилем текущего возбуждения (при активации дерева условий нет компонента, для которого еще нет ветки дерева). Активность системы значимости, выделяющая все, принадлежащее соответствующему контексту значимости, модулирует силу отклика профиля возбуждения и всех распознаваемых признаков в нем, старых и новых, что и реализует функцию перемножения.
Но у признака, для которого создана модель его свойств и качеств как объекта внимания, уже есть зафиксированная прошлым опытом произвольно оцененная значимость его для организма в тех или иных условиях как возможный вред или польза. И реагирование на такую новизну происходит уже в контексте такого произвольного смысла. При этом значимость нового признака требует сопоставления с вариантами альтернативного привычному действию на уровне более высоком, чем контексты значимости – назовем их уровнем субъективных моделей понимания.
Очевидно, что где-то должен фиксироваться опыт оценки значимости образам восприятия, образуя модель понимания того, что может значить данный стимул в данных условиях. Значимость оценивается по результату оценки последствий появления данного стимула в условиях привлечения внимания к нему, т.е. структурами лобной коры. Такая значимость отличается от гомеостатической значимости текущего состояния жизненных параметров и в этом смысле ее оценка является абстрактной (отвлеченной от гомеостатического состояния) – произвольной, что открывает возможности нового качества экспериментирования при произвольном изменении значимости объектов.
Для сравнения с гомеостатической значимостью вернемся к рефлексам. Из эмпирически выявленного условия образования условных рефлексов: “отсутствие других видов активной деятельности“ в данном контексте гомеостатической значимости (активном стиле поведения) следует, что новые условные рефлексы образуются в суженом контексте значимости, где нет других реакций. При этом привычные, уже хорошо отработанные рефлексы могут срабатывать вне узких контекстов значимости, если пусковой стимул окажется в рамках более общего контекста текущего стиля поведения. Эти реакции могут выполняться одновременно, взаимно независимо, поддерживая выполнение все более узко контекстных, более поздно образованных цепочек. Как минимум, вся система поддержания гомеостаза работает параллельно для всех регулируемых параметров.
Вся эта система жестко зависит от текущих условий и стимулов в этих условиях, она работает как автомат, хотя может и дополнять свои программы новыми рефлексами. Но это – все более трудный процесс потому, как с усложнением системы реагирования появление новых признаков, связанных с той же узкой значимостью в контексте работы распознавателей новизны, оказывается все более редким событием: с каждым опытом такая вероятность все меньше.
Это проявляется в том, что при малом числе рефлексов, определяющих привычный опыт реагирования, новизны в широких контекстах очень много, ориентировочный рефлекс срабатывает постоянно, возникает известный родителям непоседливых детей и дрессировщикам молодых собак синдром дефицита внимания и гиперактивности: СДВГ.
Стоит вспомнить, что множество реакций для познания мира после рождения использует опыт подражания, т.е. их компоненты находятся не только в теменной, но и лобной зоне. В случае поражения лобных долей синдром СДВГ возвращается (fornit.ru/7279), все как бы опять становится в новинку, но уже нет раннего безусловного исследовательского рефлекса, стимулирующего познание нового и подражание.
Очевидно, что рефлекторный механизм с набором опыта адаптации приходит к некоему пределу своих возможностей, к тупику насыщения опытом привычного реагирования, после чего появление новых реакций случается редко и маловероятно.
Но есть звенья, отражающие логику происходящего как некую модель свойств, завязанных на реакции использования этих свойств – “зеркальные нейроны”, точнее систему отражения отделенных от конкретной текущей реальности связей причин и следствий. И на их основе возникает более гибкий механизм ответных реакций – базовые автоматизмы.
Новое всегда противопоставляется привычному
Привлечение внимания к новому нужно только тогда, когда в привычном появляется нечто новое, настолько важное, что может сделать привычную реакцию неудачной и даже фатальной. Выявление такого нового необходимо, чтобы для ранее привычного контекста условий значимости, где была привычная реакция, появилось ветвление новой реакции в случае, если старая реакция в таких условиях не приводит к нужному результату, иначе можно продолжать пользоваться привычной реакцией. Вот для какой задачи развился очень сложный и богатый функционал лобных долей. Вот на что заточена психика.
Такая задача порождает определенную логику ее реализации, которую остается только облечь в схемотехнические механизмы реализации.
В первую очередь, требуется как-то определить, можно ли пользоваться привычным в новых условиях, и чем это может закончиться.
Это можно попытаться сделать с помощью прогнозирования, если известен конечный результат реакции и есть опыт того, что может означать, к чему может приводить наличие данного признака нового в восприятии.
Но можно просто попытаться прореагировать привычно и посмотреть, что из этого получится. Т.к. это может привести к негативным последствиям, нужно решить, каким методом лучше воспользоваться.
Альтернативой может быть просто рискнуть отказаться вообще что-то делать в таких условиях. Но бездействие тоже может привести к негативу и приходится решать, насколько допустимо бездействие.
Наконец, если есть опытный и отзывчивый товарищ (родитель, воспитатель, добрый самаритянин, Википедия, раскрученный блогер, ученый на популярной лекции, поп или полицейский), можно просто попросить его подсказать что делать в таких условиях. Но для этого нужны развитые системы коммуникации, которые превышают возможности рефлексов и так же формируются в лобных долях как правила общения.
Это – варианты выбора способа решения – насущная проблема, требующая решения, что же выбрать. Это – одна из сторон проявления произвольности.
Все перечисленное может быть фатально критично для выбора и поэтому функции, информирующие о возможном способе решения, особенно интенсивно отрабатывается отбором смертью особей и выработкой множества вариантов. Фатальность ошибки уменьшается использованием чужого опыта действий в новой ситуации или игровым стилем формирования собственного опыта, когда можно имитировать ситуации без фатального исхода.
В общем, вырисовывается большая система обработки значимого нового в контексте привычного.
В любом случае появление значимого нового требует возможности реагировать вопреки привычному. А это - основа произвольности (fornit.ru/12787).
Произвольность
Начнем с того, что постараемся выявить наиболее полную (из существующих данных исследований) функциональность “зеркальных нейронов” (перенимание чужого опыта – вид произвольности), что позволит отказаться от этого явно поспешного названия, хотя они, конечно, “отражающие” в смысле отражения свойств наблюдаемого.
Ранее были рассмотрены функции первичных, вторичных и третичных зон задней и теменной областей мозга, включая распознаватели системы гомеостатической значимости. Говоря о реакциях с участием детекторов нового, мы затрагиваем другую функциональную половину мозга – лобные доли. Здесь тоже есть первичные, вторичные и третичные зоны, но они имеют эффекторную направленность (что это значит будет вскоре ясно) и в этой ипостаси они во многом оказываются отражением функциональности задней половины (с теменной зоной), но в ее отражательном, более абстрактном качестве моделей понимания (пока не нужно напрягаться в попытке понять, что это, просто задается такой предварительный контекст).
Ознакомимся с наиболее важными данными фактических исследований лобных долей.
Подражательные реакции хорошо наблюдать у собак потому, что у них короткие периоды развития и легко проследить становление подражательных реакций: fornit.ru/v1. Можно заметить, что у собаки есть какое-то желание что-то сделать и в этом контексте она наблюдает, как этого достигает другой. Получается пример достижения цели еще до того, как собака сама попробует так действовать.
Учитывая эти наглядные примеры, дополним картину фактами исследований, связанными с этими явлениями.
Строение лобных долей (fornit.ru/5341):
“Передняя центральная извилина является «представительством» первичной двигательной зоны со строго определенной проекцией участков тела ... выполняет очень важную в функциональном отношении роль. Она является центром произвольных движений”.
Однако, мы будем различать “произвольность” использования готовых автоматизмов от произвольности изменения программ привычных действий на новые для новых ситуаций.
Первичные и вторичные зоны моторной коры формируются достаточно рано (fornit.ru/23514) и участвуют в схемах непосредственных запусков безусловных и условных рефлексов и их сколь угодно длинных последовательностей (цепочек действий). Во вторичные уже вмешиваются координирующие программы мозжечка. Это – зона сложных программ действий, в том числе речевых, с учетом того, что должно обеспечивать выполнение главного действия (координацию вспомогательными действиями). Для этого сюда приходят сигналы рецепторов контроля действия от мышечных, суставных, кожных рецепторов и рецепторов распознавателей координированности действия мозжечка.
Роль третичных зон лобной коры (fornit.ru/7279):
“Основным отличием здесь является тот факт, что если во втором, афферентном, блоке мозга процессы идут от первичных к вторичным и третичным зонам, то в третьем, эфферентном, блоке процессы идут в нисходящем направлении, начинаясь в наиболее высоких — третичных и вторичных — зонах, где формируются двигательные планы и программы, переходя затем к аппаратам первичной двигательной зоны, которая посылает подготовленные двигательные импульсы на периферию.
... разделы мозга, относящиеся к третичным зонам коры, играют решающую роль в формировании намерений и программ, в регуляции и контроле наиболее сложных форм поведения человека.
... Решающее значение имеет тот факт, что лобные доли мозга, особенно их медиальные и базальные отделы — обладают мощными пучками восходящих и нисходящих связей с ретикулярной формацией. Эти области новой коры получают импульсы от систем первого функционального блока, «заряжаясь» от него, в то же время они оказывают интенсивное модулирующее влияние на образования ретикулярной формации, придавая ее активирующим импульсам дифференцированный характер и приводя их в соответствие с теми динамическими схемами поведения, которые формируются непосредственно в лобной коре мозга. ... Активирующее и тормозящее (модулирующее) влияние лобных долей на аппараты первого блока доказано многочисленными электрофизиологическими экспериментами.
префронтальные отделы коры мозга являются третичными образованиями мозговой коры, теснейшим образом связанными почти со всеми основными зонами коры головного мозга. В отличие от третичных зон задних отделов мозга третичные отделы лобных долей фактически надстроены над всеми отделами мозговой коры, выполняя, таким образом, гораздо более универсальную функцию общей регуляции поведения.
... Как известно, исследования последних десятилетий убедительно показали, что схема рефлекторной дуги не в состоянии адекватно объяснить все существенное в строении поведения и что она должна быть заменена схемой рефлекторного кольца, или рефлекторного круга, в которой наряду с восприятием и анализом сигналов внешней среды и реакцией на них учитывается и то обратное влияние, которое оказывает эффект действия на мозг животного. Механизм обратной связи, или обратной афферентации, как существенное звено всякого организованного действия явился объектом пристального внимания целого ряда исследователей (П. К.Анохин, 1935, 1971; Н.А.Бернштейн, 1935, 1957, 1966; Миллер, Прибрам, Галантер, 1960). П. К. Анохин, назвавший этот механизм акцептором действия, указывает, что наличие его является обязательным условием любого организованного действия. ... животное без лобных долей мозга оказывается не в состоянии обнаруживать и исправлять допускаемые ошибки.
... префронтальные отделы мозговой коры созревают на поздних этапах онтогенеза (в 4 — 8-летнем возрасте)”.
…у ребенка 2—3 лет (fornit.ru/5226) “— регуляция произвольных двигательных реакций соответственно речевым инструкциям оказывается недостаточной. Он не может управлять движением в полном соответствии с указаниями другого лица и еще не в состоянии полностью подчинить движения собственным речевым командам. К 3—4 годам ребенок осваивает речевые навыки и сам произносит «надо» или «не надо» делать движения. Эти речевые отчеты помогают ему в регуляции действий. Афферентная импульсация, идущая от речевой мускулатуры, служит важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц. Речевые самоотчеты помогают осмысливать и направлять движения. Постепенно простые речевые реакции усложняются и образуют сложную систему саморегуляции. Внешняя речь ребенка (произносимые им слова) сначала заменяется шепотом, а затем переходит во внутреннюю речь”.
Эффекты поражения лобных долей (fornit.ru/5234):
“Поражение лобных долей мозга у человека приводит к нарушению лишь сложных, вызываемых с помощью речи форм активации, составляющих психофизиологическую основу произвольного внимания.
Элементарные формы ориентировочного рефлекса (или непроизвольного внимания) не только сохранены, но нередко даже патологически усилены.
... Например, если больному предложить зажечь свечу, он успешно зажигает спичку, но вместо того, чтобы поднести ее к свече, которую держит в руке, берет свечу в рот и начинает «раскуривать» ее, как папиросу. Новое и относительно мало упроченное действие заменяется, таким образом, хорошо упроченным стереотипом.
... Больные с поражениями лобных долей хорошо копируют действия врача, повторяя, например, движения его руки и пр. Однако если им предъявить речевую инструкцию, то они затрудняются ее выполнить. Характерным является то, что попытка использовать собственную речь в качестве регулятора поведения не компенсирует дефект: больной правильно повторяет инструкцию, но нужных действий не производит. Такой переход на элементарный уровень непосредственных эхопраксических реакций является типичным симптомом распада произвольных движений у больных с массивными поражениями лобных долей мозга. При массивном поражении лобных долей разрушается механизм, ответственный за формирование программы действия. В самом простом варианте — это замена заданной программы инертным стереотипом.
... Больные не способны создавать прочные мотивы запоминания и поддерживать активное напряжение, необходимое для произвольного запоминания.
... У больных нарушается интеллектуальная деятельность, начиная от наиболее простых и наглядных ее форм и кончая сложными видами отвлеченной дискурсивной деятельности. Эти симптомы ярко проявляются при пересказе больными достаточно сложной сюжетной картинки. Они не способны сопоставить детали картинки друг с другом, затем выдвинуть некоторую гипотезу и сверить ее с реальным содержанием картинки.
... Сразу после разрушения коры лобного полюса больные становятся апатичными, лицо принимает характер маски, у некоторых отмечают мутизм или акинезию (скованность, отсутствие движений) ... наблюдаются стертость эмоций”.
В книге С. Блейксли, Д. Хокинс Об интеллекте (fornit.ru/7198) на основе сопоставления множества мнений ведущих психофизиологов, высказывается следующее:
“Головной мозг, особенно если он достаточно большой, разделяет кору головного мозга на переднюю и заднюю часть, антериорнуюи пастериорную. Их разделяет извилина, которую называют центральной бороздой. Заднюю часть коры головного мозга образуют зоны сенсорного восприятия, получающие входные сигналы от органов зрения, слуха и осязания. Передняя часть коры головного мозга включает зоны, осуществляющие мышление и планирование высокого уровня сложности. Сюда также входит моторная зона коры головного мозга, отвечающая за контроль над мышцами, т. е. за непосредственное поведение.
В процессе эволюции кора головного мозга (особенно ее передняя часть) у людей и приматов увеличилась. По сравнению с другими приматами и ранними гоминидами у нас непропорционально большой лоб, основное назначение которого – служить вместилищем для очень большой передней части коры головного мозга. Но одного этого факта недостаточно, чтобы объяснить улучшение наших моторных способностей по сравнению с другими живыми существами. Способность человека осуществлять сложнейшие движения связана с тем, что моторная зона коры головного мозга у homo sapience имеет намного больше связей с мышцами тела, чем у других млекопитающих. Поведение большинства животных генерирует «старый» мозг. А у человека передняя часть неокортекса узурпировала большую часть моторного контроля. Если вы повредите моторную зону коры головного мозга крысы, она не почувствует существенных неудобств. Если вы повредите моторную зону коры головного мозга человека, он будет парализован”.
Итак, от самых первых безусловных рефлексов запуск эффекторных программ формируется в области первичной моторной коры лобных отделов, воздействуя на мотонейроны спинного мозга (управляющие мышцами), образуя наиболее общие, до гомеостатической регуляции ответные реакции.
Распознаватели зон восприятия отражаются своими связями в первичной лобной коре. Связи вторичных зон восприятия образуют связи уже с учетом гомеостатических контекстов значимости. И, наконец, условные рефлексы, образованные разными видами каналов восприятия, со сложными программами действий, отражаются в моторной коре, дополняясь всем, что должно эти действия обеспечивать (координировать), и это дополнительная поддержка формируется вместе с возникновением основной реакции – как обеспечение конечного результата сгибателей-разгибателей для выполнения действия и корректирующие равновесие дополнительные мышечные усилия. Изначально множество таких коррекций формируется как “рефлексы” мозжечка (fornit.ru/7424).
Мозжечок с самого раннего возраста занимается координирующей компенсацией положения тела при выполнении действий и при этом он проходит период обучения, когда ребенок старается не падать и добивается желаемого. Для этого в мозжечке используются датчики положения тела вестибулярного аппарата (лабиринтные, или установочные рефлексы, вследствие которых правильное положение в пространстве занимает сначала голова, а затем все тело, а так же рефлексы положения тела в пространстве, зависящие от правильного распределения тонуса мышц и суставов), датчики состояния мышц (так называемые афферентные или рецепторные сигналы), что приводится в соответствие с проекциями органов тела, представленных в моторной коре – все то, что используется рефлексами для запуска реагирования (т.е. эффекторные распознаватели актуальности запуска моторной реакции).
Первоначально мозжечок оказывает тормозное воздействие на моторные программы. На последние и воздействуют выходные сигналы мозжечка так, чтобы согласовывать это воздействие с ответами рецепторов положения мышцы. Т.е. ранее описываемая цепочка последовательности мышечного действия из нескольких последовательных звеньев, которые срабатывают поочередно в зависимости от сигналов результата сокращения мышцы предыдущим звеном, как раз координируется мозжечком. Вот эти тормозные связи могут оказываться задействованы во сне, когда блокируются мышечные реакции от сновидений.
Для того чтобы однозначно определить, как именно образуются такие связи компенсационных действий мозжечка схемотехнически, данных исследований недостаточно (fornit.ru/23497), но описанное вполне позволяет представить себе возможные варианты реализации. Мозжечок параллельно получает сигналы рецепторов и по ним может судить о том, насколько достигнута цель управляющей реакции, образуя отрицательную обратную связь, компенсирующую отклонения.
Особенность этих реакций в том, что они обеспечивают компенсаторную поддержку всем уровням моторного реагирования, от самых простых рефлексов, до самых сложных, произвольных.
Если мы хотим коснуться пальцем носа, не пользуясь сигналами зрения, закрыв глаза, то точность этого произвольного действия обеспечивается структурами мозжечка, где уже определено, что конечная цель движения руки с вытянутым пальцем должна сопровождаться определенным напряжением определенных мышц (соотношением тонусов сгибателей и разгибателей), а если цель дотронутся до подбородка, то – тоже самое, но немного меньше тонуса мышц вертикального удержания руки.
Можно убедиться, насколько мозжечок использует цель движения для координации. Махи ногой вперед-назад требует именно такой цели, например, чтобы носок оказался как можно выше или чтобы носком был произведен удар. Если подобной цели нет (просто бездумно машем лишь бы махать), то движения ногой легко выводят из равновесия, а если есть, равновесие эффективно поддерживается. Каждый может поэкспериментировать с этим. Кстати, бездумно махать непросто, для этого нужно махать, отвлекая внимание на что-то другое, и равновесие будет потеряно.
И тут встает вопрос: откуда мозжечок знает про цели? Как он вообще способен организовывать целенаправленную координацию? Что задает направление коррекции, например, поддержания равновесия? На уровне рефлексов такого ответа нет. Но на уровне обучения с учителем – есть. Кроме собственных целей конечную цель задает учитель в период формирования лобных долей, в период доверчивого обучения. И, как уже говорилось, заставляют следовать целям учителя безусловные рефлексы, начиная с подражания улыбке и т.п. Этот механизм формирует, с одной стороны, восприятие того, что демонстрирует учитель, а в моторной коре – уже имеющиеся собственные реакции, которыми можно воспользоваться как элементами выполнения целей учителя.
У мозжечка просто нет никакой другой возможности иметь направление формирования его координирующих функций, кроме как получить информацию о том, достигло ли действие цели или нет, которую нужно достичь, обеспечивая нужные коррекции до тех пор, пока цель не будет достигнута. И когда родители всячески побуждают, фактически заставляют, ребенка учиться сохранять равновесие, то рано или поздно такая цель в силу попыток подражания становится собственной отслеживаемой по результату действий целью, и тогда мозжечок получает возможность использовать все сопутствующие сигналы вестибулярного аппарата и сигналы состояния мышечно-суставного тракта так, чтобы дозировать усилия (на основе уже имеющихся у него модулирующих воздействий) всего, что обеспечивает равновесие, до окончательного достижения цели. Не хватает силы действия – добавить в виде рефлекса мозжечка. Перебор с усилием – убавить силу действия.
Конечно, цели могут формироваться и самостоятельно, в ходе попыток чего-то достичь в раннем периоде развития за счет совершенно беспорядочных движений и получения оценки их результата. Так учатся вообще как-то пользоваться своими конечностями и познают их возможности. Но с учителем этот процесс идет несопоставимо эффективнее и целесообразнее.
Компенсаторные функции мозжечка точно в таком же смысле затрагивают не только мышечные, а почти все внутренние регуляции организма: сердечно-сосудистой деятельности, дыхания, тонуса гладкой мускулатуры, перераспределение тонуса между симпатической и парасимпатической системами. И даже ментальные усилия воздействия на “внутренние” функции управления поведением.
Любая такая координация или поддержание тонуса выражаются в дозированном тормозном влиянии на активирующие команды или тормозным влиянием на тормозящие команды, которые без этого бы заставляли мышцы и органы сразу врубать режим конечного, целевого действия.
Как именно организуются механизмы координации в мозжечке, как они используют механизм подражательного эффекта, указывающего на цель реагирования? Схемотехник это может прикинуть сам, а можно подсмотреть природную реализацию, но нет сомнения, что сигнал целевого результата компенсаторных воздействий у мозжечка есть потому, что принципиально никаких других способов определить эффективность компенсаций у мозжечка нет.
Тот механизм ожидания последствий после реакции, что обеспечивается удержание стимула и выявления эффективности реагирования, для создания навыка Правил типа Стимул-Ответ-Эффект, используется и мозжечком.
Хотя описанная функциональность мозжечка чрезвычайно важна, она не представляется системной и могла бы быть организована более логичным образом, Мозжечок реализует достаточно хорошо алгоритмируемую функцию, и его возможно реализовать с помощью универсального модуля, что и было проделано: fornit.ru/23513. Поэтому мы сейчас не будем слишком глубоко обращать на мозжечок внимание при рассмотрении основных принципиальных механизмов. Его функциональность рассматривалась только для того, чтобы лучше представить общую картину и то, какую роль выполняют механизмы, обеспечивающие подражание.
Можно посчитать (и это выглядит наиболее правдоподобно), что мозжечковые механизмы возникли как альтернативные механизмам выработки решений путем осмысления (о чем будет сказано позже), и они оказались настольно полезными, что продолжают функционировать параллельно.
Постепенно целеуказание учителя заменяется собственными навыками произвольного выбора цели, начиная с периода инициативы после периода доверчивого обучения. Т.е. постепенно возникает и развивается собственная возможность выбора целей, что и является основой субъективной произвольности.
Все это означает, что с какого-то периода развития лобных долей формируются механизмы самообучения (целевой инициативы) по совершенно новому механизму (в сравнении с механизмом условных рефлексов) на основе того, чтобы сформировано под воздействием учителя в период доверчивого обучения: “Префронтальные отделы мозговой коры созревают на поздних этапах онтогенеза (в 4 – 8 -летнем возрасте). Темп роста площади лобных областей мозга резко повышается к 3,5 – 4 годам (в этот же период отмечается и существенный рост линейных размеров клеток, входящих в состав префронтальных отделов коры); второй скачок приходится на возраст 7 – 8 лет” (fornit.ru/24226).
О том, что такое цель, подробнее
До этого момента во всех схемах ответных реакций не говорилось об их цели потому, что это было условное абстрактное понятие для обозначения конечного результата реагирования, который никак ничем не оценивался кроме последствий для жизненных параметров организма. А отклик этих параметров – очень запаздывающий и подчас не адекватный реальному положению дел процесс, помните гистерезис после жадного утоления жажды и т.п. примеры.
Речь идет о субъективной цели реагирования (fornit.ru/67888), которая обеспечивается не только мозжечком, но и многими другими механизмами субъективной произвольности, цели, уже определенной до запуска реакции. После экспериментов Либета (fornit.ru/2506) это озадачило многих исследователей и особенно вызвало восторг у мистиков тем, что целевая активность возникает задолго до ее осознания (fornit.ru/5124)).
А ведь логика - вполне отрезвляющая: сначала активность мозга (материя), только потом (как результат) субъективное ощущение последствий (осознание). Отсюда лаг во времени. Если предполагать обратное соотношение, то сознание оказывается самостоятельным (автономным) явлением, никак не связанным с мозгом. Зачем тогда вообще такому «суверенному» сознанию мозг?
Было экспериментально определено, что для мозжечковых компенсаторных реакций субъективная цель реагирования есть (см. fornit.ru/23500). В этом смысле цель – то действие, которое нужно совершить в данной конкретной ситуации. Такая цель определяется или уже имеющимся опытом (Правилами) или совершается информационная работа по нахождению нового в данных условиях действия с наиболее желательными последствиями. После совершения действия отслеживается Эффект последствий, по которому можно судить, насколько (недо)(пере)выполнена цель. Что и является критерием работы мозжечка.
Цель в уже сформированных реакциях определена распознавателем намерения совершить (моторными нейронами) действие, отражающее возможность запуска уже имеющихся элементов действий. А если нужных цепочек реагирования пока нет, но есть те, из которых можно было бы составить нужную последовательность действий, цель указывается или учителем, или собственной системой целеуказания (далее будет понятен механизм) так, что остается только активировать подходящие элементы, составляющие общую реакцию, чтобы достичь цели и затем, в случае успеха, закрепить это как новый вариант реагирования. И после образования новой реакции становятся не нужны все промежуточные механизмы выбора, реакция реализуется самостоятельно без вовлечения лобных долей, т.е. без подключения их гиппокампом с прерыванием действия. В таких случаях исследователи говорят (fornit.ru/24206) о том, что “память” в гиппокампе как бы переходит в “память” поведенческой реакции (т.е. какая-то память переходит в структуры коры из гиппокампа): “Получается, что память кодируется параллельно и в гиппокампе, и в коре большого мозга. Со временем, баланс смещается в сторону клеток коры – сигнал становится сильнее, а в гиппокампе слабее”. Мы же будем избегать вот такие туманные и некорректные формулировки.
Понятно, что никакая такая память (в том числе доступная для субъективного вспоминания) на самом деле никуда не переходит, а просто вначале процессы выработки новой реакции использовали подключение гиппокампа при ориентировочной реакции к префронтальной лобной коре, а с появлением привычного реагирования возникший автоматизм уже обходится без этого.
С целеобразованием связан и предполагаемый результат действия в его значимости для параметров гомеостаза (fornit.ru/23501). И как следствие возникает возможность прогноза ожидаемого (fornit.ru/23502).
Итак, возможные цели при реагировании уже должны быть известны и понятны, и они определены отзеркаливанием опыта учителя или собственного опыта в качестве самоучителя в виде тех самых “зеркальных нейронов” на стороне третичной лобной коры, и у них более широкая функциональность распознавания, чем просто цели реагирования.
Для пользы понимания аккуратно и осторожно проследим все - с самого начала формирования таких нейронов в онтогенезе организма, а для этого нужно вспомнить, как возникало имитационное поведение. Дальнейшее пояснение может оказаться слишком запутанным, но потом все должно проясниться и можно будет опять пройти запутанные места для ясности, так что без паники преодолеваем это место.
Ранее рассматривалась схема рефлексов подражания с участием эффекторной области лобной коры и контекста гомеостатической значимости. Повторим этот эпизод:
“Такие образы составляют не только признаки восприятия в контексте значимости подражательной деятельности. Они должны включать в себя и распознанные программы действий: каждый образ фрагмент – со своей реакцией...
Контекст определенной цели запускается необходимостью что-то совершать, в конечном счете - для компенсации гомеостатического разбаланса (учимся клевать корм, когда голодны или прятаться от опасности и т.п.), когда в восприятии возникают признаки, уже связанные с опасностью или не понимаемые, но поведение учителя вызывает состояние опасности.
Контекст подражания возникает при активации цели, требующей каких-то действий, но без наличия уже готовой реакции на этот случай. А это означает необходимость распознавания нового в данном контексте (возможно, в наиболее примитивном варианте реализации) и активацию ориентировочной реакции. Проще говоря, если в данной ситуации еще нет привычной реакции, то нужно ее подсмотреть у авторитета.
Если рядом есть более опытная особь, к которой имеется доверие (родитель или свой из стаи), то внимание удерживает и образ, отражающий ее поведение в каждые моменты ее действий до достижения целевого результата ее действий (зерно склевано, от опасности спрятались).
Здесь нет осознания законченности, как нет и осознания необходимости (цели) подражать. Есть контекст нового после контекста значимости удовлетворения какой-то потребности (или без него, а просто – факт необычного поведения свояка, вызывающий ориентировочный рефлекс).
Активность образа с новизной вызывает подготовку связей (рост дендритов, если их еще там нет). Это – условная причина.
Следом оказывается активным и специализированный распознаватель запуска действий, схожий с тем, что есть у учителя, и это – самый важный и интересный момент. Это – элемент будущей собственной модели понимания (fornit.ru/7496), отражения реальности”.
Конечный этап действия это – неосознаваемая цель, то, что учитывается мозжечком для координации действий достижения этой цели при запуске реакции. При формировании реакции в режиме фокусировки внимания детекторами нового, цель определяется контекстом сфокусированной, выделенной системы значимости. После формирования реакции цель остается как зафиксированный образ той позитивной значимости, определяющей, зачем нужна эта реакция.
Осознанная необходимость что-то совершать это – осознаваемая цель, приводящая к произвольности действия.
Реакция сможет сформироваться, если уже были подготовлены все промежуточные программы действий, у которых есть распознаватель их запуска, и есть контекст, выделяющий то в восприятии, что относится к условиям данного состояния организма (контекста значимости), т.е. то в восприятии, что ранее получило зеркальное соответствие типа: моя реакция – видимые признаки чужой реакции, начиная с таких ранних безусловно-рефлекторных подражаний, как улыбка в ответ на чужую улыбку.
Для формирования реакции остается в ответ на более сложные, но пока не освоенные чужие действия, в своем контексте необходимости таких действий, активировать те свои реакции, которые отражают реакции чужого. Необязательно живого учителя, а вообще отражающие любые воспринимаемые явления в логике “причина-следствие”, где объективные действия причины на следствия всегда имеют уже имеющиеся свои возможности действовать таким же образом.
Это, возможно, слишком накрученная по сложности фраза, показывающая этиологию целенаправленности сначала без участия сознания, на рефлекторном уровне, но уже с использованием детекторов нового. Предположение схемотехники таких последовательностей не представляет сложности, но при этом нужно ориентироваться на достаточно простой механизм такой реализации.
А теперь – особенно следите за руками.
На самом деле мы описали нейронные основания формирования модели вообще любых отражений причин-следствий объективного мира (а не только живого учителя) в критериях понимания субъекта, т.е. в критериях уже имеющихся у него реакций, способных отражать действие причины на следствие. Это – очень важное понимание. “Зеркальность” учителя таких нейронов – лишь частный эффект их распознавательной функции (на самом деле вообще нет необходимости при этом определять “учителя” как живое и неживое).
Что важно: отражательные нейроны формируются в условиях активности детекторов нового и контекста потребности (начиная с 3-4 года от рождения), а затем - когда нужно, активируются как реакция. Это – противопоставление тому привычному рефлекторному реагированию, что было до появления нового, и это приводит к появлению нового типа реакции, но не только реакции, а и отражению цели такого реагирования в виде связанной вторичной (произвольно оцененной) гомеостатической значимости. Эта цель может быть использована и до реагирования - как прогноз того, чего можно достичь, а во время реагирования эта цель используется мозжечковыми механизмами для координирующего влияния.
При всей сложности описания взаимодействий в нейронной сети, на схемотехническом уровне нет ничего принципиально сложного в организации описываемых механизмов, и они естественно возникли в результате последовательного усложнения с оптимизацией на каждом шаге проверки нового среди огромного числа особей. Что и было осуществлено в реализации Beast. Так что сказанное – не просто слова, а имеется работающий прототип.
Но продолжим. На описанном уровне проходит плавная граница между рефлексами (жесткими алгоритмами) и тем, что вопреки этим рефлексам для новых условий формирует альтернативные реакции, учитывающие новые условия – автоматизмами, которые будут рассматриваться в следующей главе. Это граница появления произвольности.
Произвольностью будем называть не сумасбродные или случайные порывы действий, а четко: то, что для новых условий, важных для получения цели привычного реагирования, формирует новую реакцию с определенной целью, учитывающую эти условия и обеспечивает запуск именно ее, когда появляются учтенные признаки в контексте привычного реагирования.
Произвольность – альтернатива привычному для новых условий, как бы свое мнение о том, как нужно бы поступать, когда возникает вот такое новое в привычной ситуации. Если предполагаемая реакция в самом деле достигнет желаемой цели, то она становится привычной, автоматический, не требующей выбора и оценки того, насколько она подходит. Еще несколько раз эти условия могут вызвать опять ориентировочный рефлекс в силу своей пока еще новизны и не полной уверенности (про уверенность – ниже), но реакция все в большей степени уже будет на виду и подставляться все быстрее с меньшими усилиями выбора варианта произвольного реагирования.
Смысл, интерпретация, передача смысла
Оценка того, насколько данная реакция подходит, возникает на основе ранее накопленных моделей различных ситуаций, связанных с целью такого реагирования. Если раньше данный признак условий не приводил к цели или даже усугублял аварийную рецепцию, то в любых других условиях он даст предупреждающий негативный сигнал: такое кончается плохо. Если признак приводил к успеху, то в привычной ситуации с избеганием или запретом реагирования он дает сигнал о возможном решении проблемы.
Можно заметить, что такие оценки оказываются не зависимы от системы контекстов значимости теменной коры, они – именно наиболее общие модели ситуаций (абстракция), отражающих свойства объектов внимания в плане возможной цели.
Это – контекст с личной точки зрения, как некоего управляющего выбором реагирования в новой ситуации. Поэтому это – уже не гомеостатическая система значимости, а то, что может значить конкретная совокупность признаков в новых условиях, это – предположение для новых условий, основа для выбора наиболее приемлемого, т.е. это – произвольная значимость, то, что называют субъективным смыслом происходящего (fornit.ru/7339). Смысл это – осмысленная (осознанная) значимость чего-то (fornit.ru/66643).
Т.к. существует возможность самонаблюдения, то это свойство произвольного придания смысла очень верно назвали осмыслением объекта внимания. И для формирования нового варианта реагирования нужно пережить это время предположительной значимости выбора вида реагирования для достижения цели до того, как это станет привычным реагированием, не требующим такого переживания.
Как уже говорилось в теме про контексты, реакция, альтернативная привычной, новым ветвлением расширяет контекст (в более многозначительный, с большим числом возможных реакций), и теперь новая реакция оказывается в предельно узком контексте ее цели.
В случае активации узкого контекста без выполнения связанного с ним действия, возникает ясная определенность (уверенное понимание) возможной цели и способа ее достижения. В случае активации широкого контекста без уточнения условий предусмотренных в нем реакций, возникает многозначительность, неопределенность в виде многих возможных путей реагирования (нужны более конкретные условия, чтобы определиться).
В любом случае такой активации мы имеем дело именно с контекстами значимости действий в данной ситуации. С этого момента мы будем говорить не о “зеркальных” и даже не об отражающих причины-следствия нейронах, а о нейронах контекста понимания причин и следствий значимых объектов внимания, служащих для определения смысла ситуации, или просто – нейронов моделей понимания. Это – совершенно новый постулат и он не претендует на то, чтобы отмести все надежно выверенные данные исследований. Термин предлагается переопределить по вполне определенным соображениям, а тем, кого это может неприятно озадачить, просьба - некоторое время побыть снисходительным. Речь пойдет о формализации представлений о нейрональной основе субъективных моделей понимания, а это сегодня для многих – совершенно новая тема. Пока что предположим, что есть нейроны, ответственные за активацию определенных моделей субъективных представлений или смысла воспринимаемого и, соответственно, возможных действий. И это – чрезвычайно важная часть предлагаемой формальной модели.
Чем смысл отличается от гомеостатический значимости? Смысл позволяет понять, к какой модели, отражающей причины-следствия, относится объект внимания так, что это дополняет его свойства и возможности взаимодействия с ним имеющимися реакциями. И часто это бывает наперекор гомеопатической значимости, к примеру, - получить только что испеченную в углях картошку – позитивно, но для этого нужно усилием воли преодолеть негатив легкого ожога. Гомеопатическая значимость запретила бы выхватывать картошку из углей, но психика имеет Правило (полученное опытом после отзеркаливания цепь звеньев Стимул-Ответ-Эффект) по которому цель достигается после преодоления негатива: Стимул1-Ответ1-негатив -> Стимул2-Ответ3-ПОЗИТИВ.
Таким образом, смысл определяется только при инициализации такой задачи (осмысления воспринятого) при подключении ориентировочным рефлексом актуальной активности к лобным долям для осмысления: fornit.ru/12822, fornit.ru/5339, fornit.ru/24232. При этом выполняется выбор наиболее подходящей модели понимания для формирования смысла воспринятого (fornit.ru/7229): “пока испытуемые слушали предложение, но до того, как успевали увидеть картинку, их гиппокамп активно работал, подбирая ассоциации, подходящие к заданному контексту”.
После чего смысл интерпретирует воспринятое (fornit.ru/7419), делая его определенным в возможности взаимодействия. Если оказывается, что смысл не соответствует реальным свойствам объекта внимания (каким-то образом этот объят продемонстрирует абсурдность его принадлежности к данной модели), то будет активирована другая, более подходящая модель. В реализации Bеast нужные ветки дерева автоматизмов и дерева ситуации активируются мгновенно (в силу эффективности поиска по дереву), но выборка опыта Правил и опыта Значимости, а также сопоставление этих данных с особенностями текущей ситуации могут потребовать нескольких итераций.
Для понимания одним человеком другого необходимо передать смысл сказанного, т.е. дать ему такие признаки восприятия, которые позволят распознать смысл (активировать наиболее подходящую модель из имеющихся у него), и результат зависит от того, насколько соответствующие модели адекватны одна другой (а не реальности). Возникает проблема понимания или проблема передачи смысла (fornit.ru/7060).
В случае ошибки распознавания смысла возникает иллюзия понимания (fornit.ru/459), далекая от реальности, т.е. чего-то с такими свойствами вообще нет ни в какой реальности, кроме субъективной. Но даже если смысл распознан верно согласно опыту, то он все равно не отражает в точности то, что наблюдалось в реальности и чем меньше опыта, тем в большей степени.
Важнейшим способом обретения способности произвольно оперировать имеющейся информацией при осмысливании, является отвлечение образов от реальной действительности – абстрагирование.
В мозге абстрагирование начинается с рецепторов, которые уже привносят особенности восприятия мира, далеко не полноценно выделяющие из него какую-то часть сигнального воздействия, см. fornit.ru/279. В результате возникают образы, уникальные по сочетанию составляющих признаков. Эти выделенные образы являются символами того, как воздействовала реальность, символами в виде уникальных идентификаторов (в случае природной нейросети – номера связи детектора образа). Вся дальнейшая обработка идет уже не со всей сложнейшей совокупностью признаков, а с несколькими символами (в реализации Beast – числами ID образов). Уникальность образов определяет свойство абстракций быть единственными символами явления любой сложности: где бы, у кого бы, когда бы такая абстракция не возникала, она будет иметь единственный нематериальный смысл, к примеру, понятие единицы одно на всех особей, у которых оно активируется, его нет в природе в виде чего-то, но есть один на всех определенный смысл (fornit.ru/103, fornit.ru/50497).
Модели понимания – далекие от реальности абстракции (fornit.ru/103), в большей мере отражающие индивидуальные потребности и возможности. Эти вторичные (первичные образы восприятия – в третичной теменной коре, вторичные – в третичной лобной коре) абстракции хотя и могут отражать свойства материального, но сами не имеют никакого отношения к материальному миру так же, как философское понятие формы объектов: форма - не материальна (fornit.ru/1132). Субъективность – иллюзия, основанная на механизмах материальных процессов, которая всегда в чем-то не соответствует реальности и способна произвольно менять смысл, придающий этой абстракции любые, самые фантастичные свойства, которых просто не может быть в природе. Отсюда и проистекает неизбежность ошибок субъективных предположений, которые корректируются только уточнением в реальности свойств конкретного утверждения, и тогда если утверждение верно верифицируется в реальности, оно считается истиной (fornit.ru/508), если нет - ложью.
Субъективное
Не изменяя своему базовому определению, понятие произвольности будет в теме развиваться с необходимостью обслуживания все более сложных механизмов адаптивности так, что станет коррелировать с его бытовым представлением свободы воли, с тем, что вызывает чуть ли не религиозное недопонимание и вопросы о влиянии материи мозга на субъективность.
Отсюда начинается психика или область функциональности субъективного.
Не меньший религиозный трепет, чем вопрос о цели, произвольности и смысле, вызывают вопросы о том, что такое субъективность.
Поэтому нужно остановиться на связке того, что является основанием для субъективных переживаний и подтянуть понимание этого так же, как и понимание цели и произвольности.
Эмоции
У нас есть два мира: мир образного восприятия и мир субъективных абстракций. Второй полностью зависит от первого: именно через восприятие внешнего мира субъективность получает начальные данные. Эти данные ограничены текущим контекстом значимости, определяющим стиль реагирования, которому сопутствуют все рефлексы, сформированные в этом контексте. Это значит, что для каждого контекста значимости возникают свои субъективные модели понимания, отражающие данный стиль реагирования. Именно контекст такого понимания и связывают с переживаемыми эмоциями.
Можно сказать, что эмоции – это абстракция контекстов значимости (гомеостатических стилей поведения). И такое абстрагирование позволяет произвольно менять свое эмоциональное состояние. Первоначально эмоции просто копируют гомеостатическую значимость, но в ходе осмысления они могут произвольно заменяться на другие, если есть опыт того, что в данном эмоциональном контексте получается лучший эффект реализации Правил.
Эмоциональный контекст в реализации Beast является базовым условием для активации дерева автоматизмов и дерева ситуации, т.е. все остальное активируется на основе данной эмоции. И если в данном эмоциональном контексте нет подходящего автоматизма, а в другом – есть, то можно произвольно переключить эмоцию и получить нужное реагирование.
Понимание не то, что механизмов, а даже основной сути эмоций у разных академических ученых различается кардинально, например:
· Теория эмоций П. В. Симонова (fornit.ru/1430)
· И. Изард Когнитивные теории эмоций (fornit.ru/444)
· сборник работ (fornit.ru/ax1-14).
Явление субъективизации образов восприятия-значимости-действия
Когда мы говорили о самоподдерживающихся образах, позволяющих удерживать какой-то стимул даже если он временно пропадает, то в интерпретации А.Иваницкого (fornit.ru/7446) такой образ считается основой субъективного переживания. Но для того чтобы осмысливать ситуацию, т.е. быть в состоянии ясной или неполной определенности происходящего, нужна активация нейронов моделей контекстов ситуации, а это требует подключения самоподдерживающегося образа к лобной коре по связям, сформированным при образовании нового образа-контекста реагирования в новых условиях. Таким образом, переживается только те из всех самоподдерживающихся образов в теменной ассоциативной зоне, что в данный момент подключены переключателями гиппокампа к лобным долям, конкретно, к нейронам модели ситуации в ассоциативной зоне префронтальной коры. А такое подключение происходит для того из самоподдерживающихся образов, который в наибольшей степени обращает на себя внимание системы выбора наиболее актуального, о которой говорилось в главе про новизну и значимость.
У каждого контекста понимания получается представительство как в префронтальной коре, так и теменной. Для образования новых составляющих контекстов (при уточнении существующих) и нужны новые нейроны в этих областях. С каждым случаем конкурентной активности системы выбора наиболее актуального возникают условия специализации связей таких новых нейронов (ранее уже давались ссылки на работы по нейрогенезу: fornit.ru/5232, fornit.ru/5360, fornit.ru/7222, fornit.ru/5287, fornit.ru/8410, fornit.ru/5429, fornit.ru/5386, fornit.ru/16232).
Кроме того, возникают условия образования связей между предыдущим активным образом (который был чуть раньше в топе актуальности) с новым, формируя цепочки последовательности подключения образов. Это – цепочки “эпизодической памяти” (fornit.ru/67560). О том, насколько этот побочный эффект используются в адаптивных механизмах субъективности, будет еще немало сказано в темах про сон и мышление.
Первые субъективные образы ситуации (элементы эпизодической памяти: fornit.ru/67560) у людей появляются после 2 лет от рождения (fornit.ru/7656). Это при том, что контекстные модели начинают формироваться чуть ли не с самого рождения, но пока в зачатке, - только как нейроны, позволяющие запускать собственные моторные (и не только моторные) реакции в ответ на необходимость реагирования на текущий контекст гомеостатической значимости.
С появлением самоподдерживающихся образов в теменной коре и детекторов нового для формирования автоматизмов, начинают образовываться подключения между ними и нейронами моделей реагирования в префронтальной коре. Это и есть начало возникновения первых субъективно переживаемых образов как звеньев цепочек последовательностей осознания происходящего (fornit.ru/6756).
Сначала фиксируются только отрывочные, фрагментарные образы – когда гиппокамп начинает по срабатыванию ориентировочного рефлекса подключать префронтальную кору к текущему образу восприятия. Такие воспоминания могут оставаться доступными только в особых случаях их значимости (причем это может быть даже не сильная значимость, а просто наиболее актуальная среди других в восприятии). То, что именно было зафиксировано в такие моменты, в последствии дополняется новым пониманием и кардинально меняет картину такой памяти. Поэтому невозможно просто вспомнить этот след именно таким, каким он был. Но можно представить, что из-за предельно бедного набора моделей понимания он был как очень сырая заготовка субъективного переживания, собственно, там и не было переживания, а просто некий тупой слепок. Но уже с 3-4 лет все быстро меняется (fornit.ru/12789) из-за высокой интенсивности новых впечатлений, оставляющих свои следы в качестве моделей понимания и в виде эпизодической памяти (fornit.ru/67560), оживляющей этот контекст. И тогда субъективное переживание расцветает тем немыслимым богатством (в самом деле не охватываемым мысленно), который делает его таким непостижимым. Но субъективное ощущение всегда – всего лишь отклик в контексте текущей цели от фрагментарной памяти разного рода, часто не от моторных действий в ситуациях, а от мыслительных последовательностей, о чем будет говориться достаточно обстоятельно в следующих главах.
В результате развития функциональности психики возникает следующая последовательность процессов осознания. С момента просыпания возникает первичная информационная картина самоощущения (fornit.ru/160), начинается процесс осмысления, сопровождающийся активацией подходящих информационных функций (вспоминание, сопоставление, определение цели и т.п.). Каждая инфо-функция своими результатами дополняет общую информационную картину, и возникает возможность использования следующей информационной функции. Не правда ли, получается хорошее соответствие общим представлениям Дж.Тонони (fornit.ru/7589)?
Это и есть процесс мышления, в котором ощущению (переживанию) доступны только обновления информационной картины. Так что не самоощущение, а работа последовательности информационных функций, определяемых наработанными ментальными Правилами, находит решения и производит регуляцию активности мозга. А самоощущение – лишь отражение этой работы в виде смысла текущей информационной картины (fornit.ru/50394). Именно так это организовано в реализации Beast.
Автоматизмы, моторные и мыслительные
Потому как автоматизм вообще никак не описан в академической науке на уровне механизмов и свойств, то далее будет основываться на результатах, полученных при реализации Beast. Кстати, так поступим и теми психическими явлениями, описываемыми здесь, которые не имеют достаточного описания. Еще в помощь приходит возможность мысленных наблюдений за своей субъективностью.
Мы будем называть моторными автоматизмами (или просто автоматизмами в отличие от ментальных автоматизмов) реакции, возникающие на стороне психики (лобные доли), которые способны не угасать со временем, а при отрицательном значении полезности блокируются, но не забываются. При позитивном результате реагирования такие реакции усиливаются (в психике возникает большая уверенность).
Созданный психическим процессом автоматизм для достижения определенной цели, привязывается к определенным условиям, в которых он и запускается. Поэтому для данных конкретных условий может быть только один рабочий (штатный) автоматизм, остальные, ассоциированные с данными условиями автоматизмы оказываются заблокированными, но остаются в памяти и в отдельных ситуациях могут пробно использоваться.
Только что созданный автоматизм имеет нулевой параметр полезности, но при первом же его применении этот параметр получает значение эффекта реагирования, как только такой эффект возникнет (в течение некоего периода ожидания эффекта).
Возникает следующая последовательность формирования системы автоматизмов:
· Постепенно целеуказание учителя (или собственная пробная реакция) заменяется произвольностью выбора цели, начиная с периода инициативы после периода доверчивого обучения. Т.е. постепенно возникает и развивается собственная возможность выбора целей, что и является основой субъективной произвольности.
· Это означает, что с какого-то периода развития лобных долей формируются механизмы самообучения (целевой инициативы) по совершенно новому механизму (в сравнении с механизмом условных рефлексов) на основе того, чтобы сформировано под воздействием учителя в период доверчивого обучения.
· Необходимость что-то совершать это – неосознаваемая цель – основа отражения причин-следствий действий учителя или собственного исследования – самоучителя.
· Произвольностью мы уже назвали не сумасбродные или случайные порывы действий, а то, что для новых условий, важных для получения цели привычного реагирования, формирует новую реакцию, учитывающую эти условия и обеспечивает запуск именно ее, когда появляются учтенные признаки в контексте привычного реагирования.
· На стороне лобной коры доступны как образы восприятия, начиная со вторичных зон, так и образы потребностей в восстановлении жизненных параметров системы значимости потому, что они входят в контуры самоподдержания (замыкающиеся гиппокампом) и такая активность является сигнальной, имеющей определенную значимость для организма.
Автоматизмы можно условно подразделить на два основных вида:
1) управление моторными реакциями (мышцы и органы гомеостаза;
2) управление активностями структур мозга (активация или блокирования цепочек и отдельных звеньев реагирования, активация и торможение зон образов восприятия, расширяющими или сужающими границы внимания, - то, что является результатом действия произвольности.
Первый вид будем называть моторными, а второй – мыслительными автоматизмами.
Моторный автоматизм запускается в зависимости от иерархии контекстов уникальных образов (fornit.ru/5136) – дерева условий автоматизмов.
Свойства и особенности автоматизмов
При сопоставлении рефлексов и автоматизмов важно не забывать, что условные рефлексы формировались без “учителя”, определяющего цель реагирования, без способности корректироваться по результату обучения. И еще важное свойство: условные рефлексы не способны прерваться каким-то внутренним оперативным запретом типа: “нет, не делай это”, когда выполнение уже началось, а вот автоматизм способен прерываться для коррекции произвольным выбором более подходящего на любой своей фазе.
Условный рефлекс связывает условный раздражитель, появившийся до сопровождаемой его уже имеющейся реакции и выступает в роли новой причины уже определенного следствия.
Моторный автоматизм позволяет выбирать, какая из уже имеющихся реакций будет следовать за данной новой причиной, т.е. реализует произвольность выбора при учете Правил, накопленных опытом. Но после формирования, в условиях его запуска уже не использует произвольность, а срабатывает как рефлекс – быстро и бессознательно.
Условием формирования автоматизма является ориентировочная реакция (для ментальных автоматизмов – аналог ориентировочного рефлекса на уровне психики) со связью с нейронами текущей, уже выбранной модели понимания в префронтальной лобной коре. Как уже говорилось, такая реакция выделяет наиболее актуальное в восприятии-значимости-действия в области теменной коры – как наиболее значимую новизну среди всех других активностей на конкурентной основе, но не только в области теменной, а и ассоциативной префронтальной лобной, т.е. ориентировочный рефлекс затрагивает и контексты выполняющихся автоматизмов (об этом – через главу). Фактические исследования сопутствующих механизмов такой организации, как это обычно практикуется, просто констатируют участие тех или иных групп нейронов в явлении, например, так просто декларируется (в fornit.ru/24232), что “Холинергическая система обусловливает возникновение ориентировочного рефлекса, включение непроизвольного внимания, а также обработку информации во время парадоксального сна”, т.е. обработку состояния допроизвольного уровня сознания во время сновидений.
Для долговременной фиксации автоматизма не требуется нескольких повторений сочетаний причин и следствий, а фиксация происходит за время самоподдержания образа, замкнутого в кольцо переключателями гиппокампа, который, будучи подключенным к лобным долям, активирует работу мыслительных автоматизмов выбора решения и, кроме того, формирует звено цепи эпизодической памяти (fornit.ru/67560), связанное с выбранной цепочкой реагирования, что и фиксируется долговременно за время самоподдержвания активности (уже отключенной от лобных долей). Это – важнейшие процессы при осмыслении в фокусе внимания, которые будут детально и последовательно разбираться далее.
В области механизмов произвольности “(мыслительных”, в отличие от “моторных” автоматизмов) происходит выбор и подстановка варианта реагирования (или просто запуск привычного), что укрепляет связи непосредственно от образа эпизодической памяти к последовавшим реакциям (необязательно моторным, но и мыслительным, т.е. везде, где отрабатывает фокус внимания к новому).
В реализации Beast создана система развития личного Опыта в виде Правил (термин), которые используются на самом раннем уровне осознания, не требующем мышления (fornit.ru/65133).
Короче говоря, если моторные реакции воздействуют на мышцы, органы регуляции гомеостаза и т.п. терминалы тела, то мыслительные воздействуют на состояние самого мозга, управляя его состоянием централизовано, из префронтальной лобной коры, что субъективно создает иллюзию “гомункулуса” (fornit.ru/7353). Мыслительные автоматизмы как раз и обеспечивают произвольность. Эффект гомункулуса возникает из-за единственности канала осознанного внимания, что, в свою очередь является следствием выбора наиболее актуального из всего активного для решения вопроса (см. fornit.ru/44471).
После оценки результата, опять попавшего в фокус осознанного внимания после выполнения выбранного действия, эпизодическая память дополняется таким осознанием, в том числе осознанной произвольной оценкой результата.
В реализации Beast при оценке результата в эпизодическую память записывается очередное звено Правил: Стимул-Ответ-Эффект. В следующем акте оценки последствий пишется Правило, возникающее из отзеркаливания ответного действия оператора: Ответ Beast-Ответ оператора- позитивный эффект (позитивный потому, что оператор считается авторитетным источником правил.)
В лобной коре, как и в теменной, есть свои цепи удержания значимого образа (fornit.ru/7358), которые так же обеспечивают долговременную фиксацию связей – уже для ментального автоматизма.
В конце концов, новизна процесса ослабевает с оценкой удачности решения (актуальность исчерпывается), а непосредственные связи образуют автоматизм реагирования в данной ситуации, - уже минуя механизмы произвольности (запускаемые ориентировочным рефлексом с прерыванием реагирования) и автоматизма срабатывает так же быстро как рефлекс, без задержек на осмысление. Гиппокамп утрачивает свою актуальность, сохраняя лишь звенья последовательности эпизодической памяти (fornit.ru/67560), способные вернуть образное восприятие и связать с ним набор возможных целевых реакций (автоматизмов) уже в контексте понимания лобной коры, т.е. реакции (автоматизмы) остаются в области лобной коры.
Это расходится с существующими сегодня гипотезами о том, что гиппокамп осуществляет перенос кратковременной памяти (в виде чего?) в какое-то долговременное хранилище в коре. Хотя гиппокамп и может обрывать связь с нейронами модели в префронтальной коре (при появлении более актуального и переориентации на него), но связи этих нейронов с переключателями гиппокампа так же становятся долговременными, как и связи зацикленного образа с теми раздражителями новых признаков восприятия, которые дополняют активацию образа, выигравшего конкуренцию актуальности среди других.
Разберем эту ситуацию подробнее.
Образ, который самозамыкался и связывался ориентировочной реакцией с префронтальной корой, образует звено цепи эпизодической памяти (fornit.ru/7613), - последовательности обращения осознанного внимания, о которой говорилось в прошлой главе. Это звено и является связывающим с нейронами модели понимания, позволяя активировать эти модели. И, наоборот, активировать образ обратно направленными связями (не от самих моделей, а от мыслительных автоматизмов в их контексте, что будет рассматриваться позже). Это позволяет произвольно вспоминать ранее воспринимаемое (вспоминать – значит не просто активировать вспоминаемый образ, а подключать его к активному контексту модели понимания для осознания).
В этом смысле, конечно, “долговременная память” оказывается дополненной структурами вне гиппокампа, но гиппокамп остается в силах опять сделать образ самоподдерживающимся при произвольном вспоминании, и такая память не утрачивается.
В природной реализации такого алгоритма участвуют дополнительные нейроны (fornit.ru/24206) с функцией “указателя” (программисты поймут, в реализации Beast постоянно используются указатели на уникальные образы) на абстрактный (субъективный образ на стороне префронтальной коры), который был активен при осознании. Такие нейроны обнаружены около гиппокампа (конечно же) и при активации они воскрешают образ восприятия, который снова оказывается связанным с лобными долями в виде субъективного переживания эпизода прошлого осознания. Этот образ является конечным элементом иерархии контекстов значимости (о чем уже говорилось), так что воспоминание воскрешает этот частный контекст (в случае образного мышления).
Тут же заметим, что этим обеспечивается возможность повторных переживаний уже случившегося, что позволяет произвольно менять оценку и формировать новые решения. Этот метод регрессии субъективных переживаний используется эмпирическими психологами для переформатирования травмирующих переживаний и выхода из зависимых состояний.
Это происходит в отношении какого-то объекта осознанного внимания (ориентировочный рефлекс), модель которого активируется в момент такого внимания и если этот объект проявляет новое важное свойство, то его модель дополняется им (при условии, что это свойство уже известно, т.е. имеет свою цепь причин-следствий).
Предположим, близкий человек вдруг сделал вам что-то очень болезненное, чего вы от него никак не ожидали (мощная ориентировочная реакция, выделяющая эти причины и следствия). Вы этот эпизод запомните сразу и на всю жизнь. Кроме того, активность модели понимания близкого дополнится вот такой возможностью причинять боль, что свяжется с кадром эпизодической памяти (fornit.ru/67560), локализованной у гиппокампа. Для вас - не новость такие болезненные действия, новость только то, что боль причинил ваш близкий. Вы уже знаете, что делать в такой ситуации и новых автоматизмов вырабатывать не требуется.
После это случая возможны два варианта: когда вы вспоминаете тот момент, то гиппокампом активирует для самоподдержания кадр эпизодической памяти, который является указателем на модель понимания близкого (вызывает ее активацию). Другой вариант уже без вспоминания того эпизода: вы встречаетесь опять с близким примерно в той же ситуации, но теперь тот контекст включает (уже без активации гиппокампом эпизода) модель возможности причинить боль, наряду с моделями других свойств близкого, и вы все это оперативно можете учитывать, выбирая способ ответного реагирования.
Подключение образа восприятия к модели понимания означает какую-то обработку этого образа в контексте, в котором есть несколько возможных вариантов реагирования (в том числе мыслительных), ведь для того и возник механизм, чтобы для новых условий скорректировать привычное. Но пока эти реакции не запущены, даже если здесь есть уже сформированные привычные автоматизмы. Т.е. не будь этот образ наиболее актуальным по значимости новизны, то просто моментально сработал бы привычный автоматизм вместе с его сопровождением координацией от мозжечка. Но у кадра эпизодической памяти есть метка о том, что это – воспоминание реальное или субъективно продуцированное (в норме мы легко разделяем придуманное от реального в воспоминаниях). Кроме того, будучи подключенным к контексту понимания в префронтальной лобной коре из-за какой-то новизны, привычная реакция сразу не активируется, а ожидает или подтверждения или отмены или коррекции. Такое прерывание – важнейший, ключевой момент, на основе которого все было организовано в префронтальной коре чтобы не только связывать новые стимулы с уже заготовленной и выполняющейся реакцией, а иметь возможность оценить, чем такая реакция при таком стимуле может закончиться. Потому как в третичной лобной коре уже есть заготовки целей имеющихся реакций с оценкой их результата (цепочки Правил), то при подключении к ним нового стимула в имеющемся контексте возникает возможность предположительно предсказать, чем это может закончиться: возникает мгновенное понимание того, чем сулит этот новый стимул в ранее привычной ситуации. И тут могут быть разные ситуации.
В простейшем случае может быть быстрый ответ: продолжать ли автоматическое действие или проявить осторожность.
Если же действие подразумевает сложную программу реагирования из многих звеньев Правил, то такой быстрый прогноз оказывается затруднительным.
Но уже есть связи к звеньям эпизодической памяти и можно подключить следующее звено, чтобы подсмотреть, а чем оно закончится или даже быстро пройти все звенья до конца, на что, конечно, требуется больше времени. Но звенья просматриваются без выполнения реагирования по ним, просто подвозбуждаясь для вспоминания их при заблокированных выходах на реагирования. Блокировка моторных реакций во время размышления о них – древнее приобретение для поддержки режима сна, когда блокируются внешние пусковые стимулы и моторные реакции.
Во время ориентировочного рефлекса реагирование от образа с актуальной новизной не только прерывается, но и к его цепочке действия прикладывается тормозное влияние в точности как это было во сне, но только в отношении одной такой цепи. Т.е. торможение для обеспечения сна становится еще и произвольно управляемым. Это позволяет делать прогнозы или произвольно проходить цепочку по отдельным ее звеньям, контролируя результат их выполнения и корректируя в случае ошибки. Все эти действия совершают мыслительные автоматизмы от самых простых, до более сложных цепей управления.
И все это сопровождаются субъективными переживаниями, воскресающими эпизодическую память и/или мыслитетельный автоматизм произвольности в контексте текущей модели понимания.
При торможении боковыми влияниями цепь активируется без задержек и прогноз последнего звена сложной цепи доступен почти мгновенно. Это явление наблюдал еще И.Павлов (fornit.ru/5206) и способ получения картины возможного будущего назвали “опережающим (события) возбуждением”. Вот демонстрация такого процесса: fornit.ru/m4.
Этот вид прогнозирования оказывается возможным из-за фиксации точки касания ориентировочным рефлексом наиболее актуального звена реагирования в виде образа восприятия-значимости-действия с образованием доступа к последующей его активации со стороны префронтальной лобной коры в виде звена цепи эпизодической памяти (неизбежно образующимся при каждом акте фокусировки осознанного внимания).
Понятно, что область мозга с такими самоподдерэивающимися активностями довольно быстро начинает заполняться, что затрудняет уточнение внимания, но в какой-то мере соседние активности взаимно тормозят и разряжают переактивированную зону. Окончательно же зоны избавляются от активностей общим торможением во время сна.
Чем более многозначная реакция следует от данного образа и чем более не определен в прогнозах новый стимул в привычном окружении реагирования, тем дольше будет процесс выбора: продолжать или нет прерванный автоматизм. Если решения не находится за время осмысления, то реакция может быть запущена с риском последствий или не запущена, если она недостаточно необходима. Но это – уже сложный процесс обработки ситуации, а сейчас важно разобраться с базовой организацией принципа реагирования с участием нейронов модели понимания.
Схемотехнически очевидно, что для управления запуском отдельных кадров эпизодической памяти так и всей цепочки с блокировкой выполнения идущих от этих образов действий, должны быть организованы соответствующие нейроны запусков таких реакций в лобной третичной коре, срабатывающие в ответ на необходимость подсмотреть ассоциированную с образом (в третичной теменной коре) значимость, если оказывается недостаточно произвольной значимости нейронов цели действий (слишком широк контекст понимания), Такие нейроны обеспечивают обратный доступ к образу в третичной теменной коре, одновременно организовывая торможение (блокировку) выполнения действий от этого образа с целью получить его конечный результат значимости.
Эти нейроны формируются при каждом акте подражания или наблюдения, связывая цель реагирования с образом восприятия условий такого реагирования еще даже до начала созревания механизмов поддержки автоматизмов.
В реализации Beast это организуется с помощью инфо-функций, управляемых ментальными автоматизмами.
Любые словесные или невербальные раздражители, символизирующие какое-то действие, связываются с теми образами в третичной теменной коре, которые соответствуют этим условным символам, способными самостоятельно активировать образ звена эпизодической памяти. Это – достаточно хорошо изученные факты исследований, прежде всего, вербальных зон в лобной коре (fornit.ru/ax1-17).
Кроме моторных автоматизмов есть класс реакций на основе психики, которые обычно относят к неорганической психопатологии ввиду их чрезмерности и навязчивости, но на самом деле являются психосоматическими автоматизмами, у которых есть как полезная функция, так и негативный субъективный эффект. Подробнее они описаны в статье Психосоматические автоматизмы.
Механизмы произвольности
Над связью условных символов в рамках подражания, обучения или наблюдения с образами соответствующего профиля восприятия, ломали голову все физиологи. И.Павлов создал теорию сигнальных систем, П.Анохин теорию функциональных систем с учетом воздействия раздражителя при рассогласовании привычного и нового.
Но в реальной организации, кроме механизмов, хорошо поддающихся универсальному алгоритму описания, есть то, что оказывается над этим – организация произвольности. Мы начинали ее описывать с прошлой главы и в ней оказывается так много взаимосвязанных компонентов, что сразу не получается облечь всю логику их взаимодействия в некую систему. Особенно трудно положиться на какие-то данные исследования в этом из-за невероятной вариабельности способов реализации и, как следствие, недостаточность целевых, конкретно ориентированных исследований отдельных механизмов. Но есть общая логика такого взаимодействия во множестве ее проявлений, в том числе и возможности самонаблюдения свойств субъективного. Так что нужно в этом месте построить интерполяцию принципиальных механизмов организации на основе этих ключевых опорных точек.
Если бы была задача создать программный вариант организации автоматизмов, то замыкание самоподдерживающей активности, подключение к контексту понимания в лобной коре, образования связей обратной активации образов от лобной коры, программист написал бы в виде универсальной функции, к которой обращается ориентировочный рефлекс при каждом выявлении им наиболее актуального из образов. Потому, что эти процессы унифицированы в достаточно самостоятельную логику.
В реализации Beast такая функция называется Осмыслением. Именно в ней происходит поэтапное рассмотрение ситуации с приходом каждого значимого Стимула. Это рассмотрение в Beast разделено на 4 уровня осмысления. На первом рассматривается возможность запуска автоматизма без последующих размышлений. На втором используются Правила, накопленные опытом реагирования. На третьем возникает итерация вызовов инфо-функций для нахождения решения (мыслительный процесс или цикл размышлений) – это принципиально не алгоритмизируемый процесс. Четвертый уровень включается тогда, когда не получается сразу найти решения и тогда формируется Доминанта нерешенной проблемы, в которой опредееляется объект целевого внимания и решающее действие по отношению к нему, что и будет считаться решением проблемы (fornit.ru/67888).
В случае задачи схемотехнической реализации в виде физического устройства это был бы тоже общий универсальный канал организации такого взаимодействия, который бы подключался “гиппокампом” как к образу, так и к контексту понимания в лобной коре на время выработки произвольной оценки и подходящей реакции.
Тот же канал уже после формирования автоматизма уже не требовал бы подключения к системе выбора подходящего поведения, а просто бы связывал условия с выбранной реакцией.
И нечто подобное уже было создано в виде искусственного гиппокампа (fornit.ru/5221), взяв на себя функцию такого универсального канала (это оказалось столь же алгоритмично, как задача при создании искусственного мозжечка, о чем говорилось в прошлой главе).
Такой канал можно назвать каналом осознанного внимания к наиболее актуальному. Осознанного в смысле, что в случае новизны привычная реакция ждет подтверждения от системы выбора поведения или подстановки другой имеющейся реакции, а такая система уже не является универсально алгоритмичной, при том, что она очень непростая по организации и развитию многих ее уровней все большей эффективности индивидуальной адаптации.
Единственность этого канала и вынужденная конкурентность за него возникла потому, что система выбора подходящего поведения возникла в виде общего универсального модуля для выбора лучшего варианта в любых новых ситуациях и поэтому она должна подключаться к тому из образов, который выделяется как наиболее актуальный в значимой новизне. Понятно, что если есть система обработки новой ситуации для поиска наиболее подходящего ответа, то она – одна и должна подключаться к наиболее актуальной текущей задаче (логично решать самое главное, актуальное в первую очередь).
В природной реализации такой канал выполнен не в виде одного универсального модуля, подключаемого к нужным цепям, это бы сделало его слишком уязвимым в случае повреждения. Тут очень много параллельных связей к лобным долям от них обратно, что, кроме обычной для мозга надежности, дает немало дополнительных эффектов и возможностей, оптимизируемых методом смерти особей.
Принципиальная функциональность такого канала не вызывает уже сомнения по очень многим взаимно подтверждающимся фактам и роли в общей структуре организации адаптивности выше условных рефлексов, в частности организации “опережающего возбуждения” с использованием кадров эпизодической памяти от актов срабатывания ориентировочного рефлекса для наиболее актуального из образов.
Для каждого из нас это – канал субъективного восприятия и понимания смысла происходящего при выборе того, как поступать, для природы – канал формирования новых автоматизмов с системой выбора наиболее подходящего варианта. И нам нужно во всем этом разобраться основательнее для обеих ипостасей этого канала.
Скорректируем наиболее общий контекст понимания: нам кажется, что мы способны произвольно переключать внимание с одного объекта на другой, но ориентировочный рефлекс всегда срабатывает вне нашей произвольности во всех случаях без исключения, выбирая наиболее актуальное по значимости нового среди всех активностей как в теменной коре, так и префронтальной лобной, где и сосредоточены все механизмы произвольности. И это – единственное, что сильнее произвольности и определяет ограничение на “свободу воли” нашего “гомункулуса”. Если мы обращаем на что-то внимание, у этого есть причина, характеризующаяся как наиболее значимая новизна. Это давно понимали философы, начиная от Спинозы, говоря, что свобода – есть осознанная необходимость.
Организация произвольности принципиально требует абстрагирования от всего привычного, более старого в мозге, что и организуется на уровне третичной зоны префронтальной лобной коры. И эта “независимость”, полностью определяемая всеми составляющими ее структурами мозга, проявляющаяся субъективно как самоуправление, не имеет предела по возможности моделировать совершенно нереальные субъективные идеи и целые миры, правда, полностью ограниченные более низкоуровневым восприятием.
Когда-нибудь все эти “сложности” обретут целостные образные и символьные понимания во множестве практических воплощений и перестанут казаться сложными, так же как привычный смартфон. А в учебниках будет описание вроде такого: fornit.ru/65133.
Но и сейчас, если отбросить “лишнее”, как отбрасывается понимание устройства смартфона, картина достаточно проста и очевидна: осознанное внимание прерывает выполнение действия для выбора более подходящего с учетом новизны и продолжает уже выбранное действие, что и формирует автоматизм, уже не требующий произвольности выбора (осознания) в тех же условиях.
Организация произвольности требует абстрагирования от всей более древней, стереотипной части управляющей схемотехники мозга, что и порождает субъективное переживание волевого управления.
Компонент принципиально не алгоритмизируемой произвольности включает в себя всю сакраментальность функции сознания. И со второго уровня осознания эта произвольность основана на мыслительных автоматизмах, способных формировать новое, более сложное на основе имеющейся базы стереотипных реакций.
Особенности мыслительных автоматизмов
Последовательность ориентировочных подключений внимания к возможным целям в лобной коре в поисках лучшего из возможностей так же, как и последовательность кадров эпизодической памяти, фиксируется как цепочка в теменной коре. Ведь все символьные цели реагирования в префронтальной коре получают свои соответствия в теменной в виде образов, что показывалось совсем недавно.
Рядом с гиппокампом были обнаружены “карты местности” (fornit.ru/7057, fornit.ru/6605, fornit.ru/6604). Такая карта местности – это последовательность Правил, о которых уже шла речь. По любому локальному Правилу видно, что правильно делать далее и то, что делалось до того.
Когда опытный человек играет в шахматы, то каждый ход противника вызывает уже известное Правило о том, как нужно отвечать на такой ход. Причем в зависимости от того, какие ходы были сделаны до того, выбирается именно нужная цепочка Правил. Становится возможным предсказывать, а как нужно походить, чтобы вынудить ответить определенным ходом и разыгрывается целая многоходовая комбинация.
Реализация такого механизма очень алгоритмична, что было проделано в случае Beast. Но бывает, что ситуация не дает определенного Правила или вообще не понятна. Тогда приходится начать искать наиболее подходящее решение. Нужны поиски решений с помощью все той информации, что уже доступна: эпизодическая память, структурированные в виде “карты решений” цепочки Правил, ранее применявшиеся действия. Попытки найти решения приводят к успеху и тогда фиксируются та последовательность вызова информационных функций, которая привела к успеху – ментальное Правило.
При поиске решения вызывались автоматизмы, которые запускали информационные функции. Когда ситуация оказывается схожей, то возникает возможность использовать готовые мыслительные решения по имеющимся ментальным Правилам. А также записывается новый ментальный автоматизм, реализующий данный способ решения.
Как уже говорилось, каждый вызов инфо-функции дает новую информацию в общую картину понимания и становится понятнее, какую еще инфо-функцию вызвать, если информации недостаточно.
Процесс вызова инфо-функций с помощью ментальных автоматизмов является циклическим потому, что рекурсивно вызывает функцию осознания (рекурсия – вызов функции из самой этой функции) на основе новой информации. Так реализованы циклы самоорганизации мышления. Как это реализовано в природе - еще предстоит выяснить. Но то, что при этом есть циклическая активность с обратной связью, о которой писал Дж.Тонони, не вызывает никакого сомнения.
В случае появления новой проблемы при осознании очередного нового, непосредственное реагирование приостанавливается на время подключения к лобной коре для опережающей действие обработки (осмысления), где, в контексте понимания происходящего, в точности, как и в теменной в контексте значимости, формируются автоматизмы управления активацией образов для выбора варианта реагирования – по тем же принципам оценки удачности результата. Целью реакции являются не эффекторные программы для мышц или гормонов и т.п. моторные эффекторы, а эффекторы выбора варианта реагирования, т.е. последовательности подсматривания за кадрами эпизодической памяти или сопоставления значимостей возможных целей. Начиная от самых простых таких автоматизмов, могут нарабатываться все более сложные цепочки поиска вариантов для решения по ментальным Правилам, как лучше реагировать в данных новых условиях.
Факт наличия ментальных автоматизмов и ментальных Правил в природной реализации не вызывает сомнения: fornit.
Мы часто мыслим так легко, что вообще не замечаем этого процесса, хотя сами - довольно сложные продукты не моторного характера, а чисто субъективного, в виде идей. И чем более человек опытен в данной специализации (контексте) мышления, тем более незаметен процесс такой импровизации посредством имеющихся заготовок, без нерешенных проблем в вариантах действия, т.е. без провоцирующей осмысление новизны. Возникает эффект особой “тишины мыслей” (fornit.ru/17954) – оптимизируемой нормы индивидуальной адаптивности.
Осознание информации после действия мыслительных автоматизмов также образуют последовательности кадров (моментов осознания) эпизодической памяти – как кадры образов, намного более абстрактных, чем образы в контексте значимости теменной коры. Это образы абстрактного мышления, имеющие в эпизодической памяти метку о своей субъективной природе. С опытом стиль осознания с применением моторных Правил все в большей степени заменяется стилем ментальных Правил и яркость первичных образов тускнеет в мыслях, использующих далекие от первичности образы.
Если переживание в контексте реагирования моторных автоматизмов оперирует непосредственными образами восприятия-значимости-действия теменной коры, то мыслительные автоматизмы формируют более опосредованные символы слов и абстрактных образов невербальных условных раздражителей. Для раскрытия до зримых образов восприятия (образное мышление) нужно дополнительно активировать соответствующие образы теменной коры через “указатели” эпизодической памяти в районе гиппокампа (запускающие самоподдержание этих образов). А эпизодическая память абстрактных символов префронтальной коры не имеет столь непосредственного обращения к образам восприятия и требует дополнительных усилий произвольного вспоминания, через активацию уровней первичных образов, вплоть до вторичных зон восприятия (потому, что с ними связаны элементы системы значимости и они способны самоподдерживаться).
От того, какой из видов эпизодической памяти преобладает в процессе мышления, зависит и образность субъективных переживаний, что описывается как образное (наглядно-образное) и как абстрактное мышление, точнее, субъективное переживание.
В онтогенезе первый вид мышления все в большей степени заменяется и перекрывается вторым потому, что все большая сложность оперирования первичными образами восприятия становится затруднительной, гораздо проще оперировать символами, причем далеко не всегда словесными (хотя многие ученые ошибочно считают словесные символы обязательным атрибутом мышления).
Первоначально окружающий мир и события переживаются очень ярко и зримо во всех тончайших нюансах и деталях, но это с возрастом по мере накопления опыта все более сложного решения задач в новых условиях заменяется безликими символами. О разительном контрасте этих видов восприятия написано в книге (fornit.ru/1585) врача-физиолога Оливера Сакса "Человек, который принял жену за шляпу":
“Стивен Д., 22 лет, студент-медик, наркоман (кокаин, PCP, амфетамины). Однажды ночью - яркий сон: он - собака в бесконечно богатом, "говорящем" мире запахов. ("Счастливый дух воды... отважный запах камня"). Проснувшись, обнаруживает себя именно в этом мире ("Словно все вокруг раньше было черно-белым - и вдруг стало цветным"). У него и в самом деле обострилось цветное зрение ("Десятки оттенков коричневого там, где раньше был один. Мои книги в кожаных переплетах - каждая стала своего особого цвета, не спутаешь, а ведь были все одинаковые"). Усилилось также образное восприятие и зрительная память ("Никогда не умел рисовать, ничего не мог представить в уме. Теперь - словно волшебный фонарь в голове. Воображаемый объект проецирую на бумагу как на экран и просто обрисовываю контуры. Вдруг научился делать точные анатомические рисунки"). Но главное - запахи, которые изменили весь мир ("Мне снилось, что я собака, - обонятельный сон, - и я проснулся в пахучем, душистом мире. Все другие чувства, пусть обостренные, ничто перед чутьем"). Он дрожал, почти высунув язык; в нем проснулось странное чувство возвращения в полузабытый, давно оставленный мир.
- Я забежал в парфюмерную лавку, - продолжал он свой рассказ. - Никогда раньше запахов не различал, а тут мгновенно узнавал все. Каждый из них уникален, в каждом - свой характер, своя история, целая вселенная.
Оказалось, что он чуял всех своих знакомых: - В клинике я обнюхивал все по-собачьи, и стоило потянуть носом воздух, как я не глядя узнавал два десятка пациентов, находившихся в помещении. У каждого - своя обонятельная физиономия, свое составленное из запахов лицо, гораздо более живое, волнующее, дурманящее, чем обычные видимые лица. Ему удавалось, как собаке, учуять даже эмоции - страх, удовлетворение, сексуальное возбуждение... Всякая улица, всякий магазин обладали своим ароматом - по запахам он мог вслепую безошибочно ориентироваться в Нью-Йорке.
Его постоянно тянуло все трогать и обнюхивать ("Только на ощупь и на нюх вещи по-настоящему реальны"), но на людях приходилось сдерживаться.
Эротические запахи кружили ему голову, но не более, чем все остальные - например, ароматы еды. Обонятельное наслаждение было так же остро, как и отвращение, однако не в удовольствиях было дело. Он открывал новую эстетику, новую систему ценностей, новый смысл. — Это был мир бесконечной конкретности, мир непосредственно данного, - продолжал он. - Я с головой погружался в океан реальности.
Он всегда ценил в себе интеллект и был склонен к умозрительным рассуждениям - теперь же любая мысль и категория казались ему слишком вычурными и надуманными по сравнению с неотразимой непосредственностью ощущений.
Через три недели все внезапно прошло. Ушли запахи, все чувства вернулись к норме. Со смесью облегчения и горечи Стивен возвратился в старый невзрачный мир выцветших переживаний, умозрений, абстракций.
- Я опять такой, как раньше, - сказал он. — Это хорошо, конечно, но есть ощущение огромной утраты. Теперь понятно, чем мы жертвуем во имя цивилизации, от чего нужно отказаться, чтобы стать человеком. И все-таки это древнее, примитивное нам тоже необходимо”.
У взрослого, умудренного опытом человека, никакие острые впечатления и образы не мешают ему мыслить, причем без ущерба для качества в том, случае, если это нужно. Мысли как бы защищены от помех потому, как с опытом мышления оптимизируются мыслительные автоматизмы и среда их активности, образуя защищенную от всех помех субъективную зону мышления. И в это защищенное пространство время от времени прорываются ценные идеи из области неосознаваемых автоматизмов мышления, не перестающих решать проблему в общем контексте необходимости ее решения. Как только значимость новизны новой идеи оказывается преобладающей, ориентировочный рефлекс фокусирует на ней осознанное внимание, и она становится доступной для осмысления, принятия или непринятия – как в случае осознания привычных реакций с новым компонентом. Об организации такой субъективной зоны мышления написано в статье “Тишина мысли”: fornit.ru/17954.
То, на чем фокусируется осознанное внимание, определяется наиболее актуальной новизной так, что среди всего активного осознается только наиболее значимое и новое, а остального нет в мыслях.
Образы “модели понимания” в лобной коре играют такую же контекстную роль, как контекст гомеостатической значимости в теменной коре: этот контекст служит для случая запуска звена уже имеющегося автоматизма в нем (цепочки отдельных звеньев более элементарных реакций), т.е. в данном контексте понимания определенные стимулы моментально запускают привычную реакцию.
Сознание. Уровни сознания
Теперь, когда уже нами прояснены многие атрибуты процесса осмысления, будет легче разобраться с “Трудной проблемой сознания”.
Нет более туманной и противоречивой в различных трактовках темы, чем тема о сознании. Поэтому не станем приводить множество сырых, наивных и откровенно фантастических теорий на этот счет и не стоит всерьез отвлекаться даже на наиболее из них принимаемые в научном сообществе (fornit.ru/23531), ведь все они ничего на самом деле не проясняют в механизмах и не имеют практической пользы, а представляют собой лишь уровень философского осмысления.
Однако, в книге Антти Ревонсуо “Психология сознания” (fornit.ru/b9), кроме современных теорий сознания можно узнать о многих эмпирических коррелятах мозговой активности, связываемых с сознательной деятельностью. Рекомендуется прочесть эту книгу сначала “по диагонали”, а после освоения двух последующих глав уже прочесть критически, в контексте освоенного.
Приведу постановку проблемы от К. Анохина (fornit.ru/1714), его вопросы во многом производные от списка, сформулированного Д.Эдельманом:
“1. Соотношение мозга и разума
2. Соотношение сознательного и бессознательного
3. Соотношение разума человека и разума животного
4. Соотношение локализованных и распределенных функций мозга.
Главный вопрос теории: "как материя мозга рождает субъективное явление?"
Это – достаточно настораживающие сопоставления: мозг и разум (соотношение компьютера и виндовз), человек и животное (по биологической классификации человек - животное), локальные функции и более общие (это даже не философия, а похоже на риторику того, кто точно не мыслит схемотехнически).
Если привести эти вопросы в более корректный вид, то на них вполне становится возможным дать конкретные обоснованные ответы, очевидные настолько же, насколько учет причин и следствий в схемотехнике дает понимание функциональности и наблюдаемых явлений работы электрических схем. И при этом не придется прибегать к каким-то новым сущностям, предположительным построениям и понятиям, что – уже один из критериев удачного обобщения (fornit.ru/7649), наряду с другими.
Основная функция сознания
Теперь можно и пора выделить наиболее общие свойства сознания. С основными принципами организации информационной обработки удерживаемого вниманием образа лучше ознакомиться в отдельной статье: fornit.ru/67865
Прежде всего, то, что функцию сознания отличает от функций всех эволюционно предшествовавших механизмов индивидуальной адаптивности: фокус сознания (то, что мы рассматривали как канал связи воспринимаемого с его целенаправленным осмыслением) всегда и без исключения оказывается на наиболее актуальной новизне происходящего в контексте текущей (а значит уже актуальной) значимости.
Проще всего, так, как годилось бы для школьных учебников, можно сказать следующим образом.
Есть условные рефлексы, связывающие стимулы с какой-то ответной реакцией. Но наиболее актуальное новое заставляет прерывать выполнение привычной реакции и перенаправлять стимулы с новым параметром в область процессов психики в контексте понимания смысла ситуации и целью, определенной привычной реакцией. И эти процессы пытаются найти наиболее подходящее продолжение реагирования с учетом того, что они уже знают о значении и свойствах новых объектов внимания. Или же просто разрешают осуществиться прерванной реакции.
В случае удачного результата пробного реагирования такие реакции все более закрепляются и уже не требуют привлечения внимания к ним для корректировки: вопросов больше нет, все теперь совершается привычно и быстро как рефлексы.
Привычные мыслительные автоматизмы, точно так же, могут прерываться новым для нахождения более подходящего направления мыслительной произвольности.
Вот такое привлечение внимания с прерыванием привычного для подстановки более адекватного реальности реагирования и называется моментом осознания, который фиксируется в виде кадра эпизодической памяти в цепочки последовательности перескоков внимания на наиболее актуальное в области моторных и мыслительных автоматизмов.
Сам момент осознается, точнее ощущается (fornit.ru/64924) в виде общей информационной картины.
Вот такая, в принципе, простая формулировка.
Это и есть самая главная и общая функция сознания: следить за наиболее актуальным для того, чтобы адаптировать уже привычные ответные реакции к появившемуся новому. Это – ответ на вопрос 4: “Соотношение сознательного и бессознательного”.
Если новое появляется 1) в контексте гомеостатической значимости (на стороне теменной ассоциативной коры) или же 2) в контексте абстрактного мышления, связанного с произвольно приданной значимостью (на стороне префронтальной лобной доли), то сознание получает к нему доступ с прерыванием связанной с данным узким контекстом привычной реакции (такое прерывание специально подчеркивалось при рассмотрении механизмов подключения канала связи). Это приводит к некоторой задержке реагирования, но зато позволяет быть более адекватным реальности.
В первом случае осознанное внимание для корректировки привлекается к области восприятия, во втором – к области мышления.
Задержка может затянуться до окончательного прерывания того, что грозит прогнозируемыми нежелательными (произвольность оценки) последствиями. Или же эта задержка может никак не ощущаться, если не прогнозируется что-то нежелательное, или актуальность действия настолько высока, что нет другого выхода, как просто последовать привычной реакции. Опытный, решительный человек сразу оценивает более-менее знакомую ситуацию и не тратит времени на мешающие действиям сомнения.
Есть люди, которые не могут перейти речку по камням или по бревну просто потому, что начинают тщательно контролировать каждый свой шаг. Чудо, если они не падают в воду. Они не доверяют сами себе, своей несомненной способности моментально и верно действовать наиболее эффективно на основе накопленного опыта, не требующего осмысления. А ведь они всю жизнь нарабатывали эти самые движения. Думать в таких случаях, когда точно нужно уже действовать, бывает просто опасно: fornit.ru/1133.
Желательность или нежелательность – произвольные (меняющиеся от обстоятельств) нестереотипные оценки достижения цели, которые вырабатываются в ходе работы мыслительных автоматизмов. Они и являются субъективным самоучителем при формировании осознанных реакций. Это будет рассмотрено подробно.
Вне канала осознанного внимания ничего не переживается субъективно, даже боль (fornit.ru/189), т.е. ощущение – это совокупная информация в текущем моменте итерации функции осознания и больше нигде нет этого явления.
Все, что оказывается вне подключенного канала осознанного внимания к наиболее актуальному, в принципе не может переживаться субъективно, и составляет область временно не контролируемых стереотипов поведения (бессознательное). Сюда входят и процессы работы информационных функций в ходе мышления. Мы не можем осознанно отследить ничего из воспринимаемого, что не является наиболее актуальным, хотя оно и продолжает воздействовать на уже имеющиеся привычные реакции всех видов. Мы не можем отследить, как именно мы мыслим, а лишь перескакиваем в осознании с одного на другое по мере обновления информационной картины, а в остальном работают имеющиеся автоматизмы. Этим пользуются фокусники и мошенники.
Наиболее прагматичным советом в острых, не терпящих промедления ситуациях, будет рекомендация не рассуждать, занять сознание удержанием нужного стиля поведения, а не переживанием окружающего, и полностью довериться уже имеющемуся опыту потому, что ничего более проверенного и полезного у вас в данной ситуации нет: неосознанные действия – самые правильные (fornit.ru/5325) в условиях необходимости немедленного реагирования. Так в жизни часто и происходит: эволюцией выработано распознавание ситуаций, когда нужно отключать сознание (состояние аффекта: fornit.ru/23532). Но можно выработать навыки более управляемого состояния, некоей стратегии поведения в критических ситуациях: fornit.ru/1133.
В эпизодической памяти остается только цепь последовательностей переключения канала осознанного внимания с возможностью вспоминания информационной картины для этого кадра. Детальность воспоминания зависит от того, насколько внимательно мы раскрывали образы восприятия во время осмысления. Разведчиков учат привлекать внимание к наиболее значащему для целей разведки.
Казалось бы, стоит захотеть и можно пристально наблюдать хоть за самым незначительным объектом внимания, например, за нарисованной точкой на листе. На самом деле, если такая задача доходит до ее реального воплощения, наблюдению за точкой произвольно придается высокая значимость, что и обеспечивает удержание внимания на ней, но чем больше времени пройдет, тем труднее до полной невозможности это станет делать. Внимание будет постоянно соскакивать (fornit.ru/7156) с точки, пока откровенно не остановится на чем-то другом внешнем или какой-то мысли. Против этого базового условия осознанного внимания принципиально не может быть никакого приема.
Как уже говорилось в позапрошлой главе, структуры лобного отдела мозга изначально формируют коллекции Правил реагирования, отражающие следствие получения значимого для личности результата, порождаемые причинами данного воздействия (формируют абстракции полезных и вредных последствий – как отражение причин и следствий объективной реальности).
Произвольность (действие мыслительных автоматизмов в контексте понимания смысла происходящего) обеспечивает достижение этих целей или смену целей уже имеющимися или новыми способами в случае, если вмешивается что-то новое в причинность событий. Но для функционирования произвольности она действует не в контекстах гомеостатической значимости, а в контекстах смысла происходящего – в субъективных моделях понимания происходящего, связанных с целью реагирования в этом контексте.
Появление нового означает необходимость переоценки смысла происходящего, который может меняться новым обстоятельством. Произвольность, в первую очередь и обеспечивает понимание нового смысла (или отсутствие необходимости изменения смысла) за счет того, что уже есть модель (знание) свойств объекта нового, который и учитывается. А как только появился новый смысл происходящего, корректируется цепочка реакций для достижения имеющейся (доминирующей активности нерешенной проблемы) цели. Для новой цели отрабатывают соответствующие ее достижению автоматизмы.
Пример: Вы открываете крышку любимого блюда, но оттуда разит отвратительный запах. Смысл ситуации резко меняется потому, что вы знаете, что такой запах означает испорченную еду. Все, есть нельзя, нужно искать новую еду.
Контекст субъективного переживания или осознания – всегда имеет ввиду какие-то целевые возможности индивидуального реагирования, в виде смысла со связанными с ним возможностями. Без такого контекста переживания теряют определенный смысл и, тем самым, утрачивается возможность осмысления. Если эта логика еще не очевидна, то нужно вернуться к более раннему тексту книги, начиная с главы “Новизна и значимость”, сорри…
Суть субъективного переживания
Субъективое переживание, ощущение, квалиа и т.п. - то, как воспринимаются процессы осознания (fornit.ru/67865) самим субъектом (fornit.ru/50394).
Для понимания субъективной сути сознания нужно проследить за сознанием с момента его появления после полного бессознательного состояния, которое бывает после очень глубокого сна и, тем более, общего наркоза, с состояния наиболее базового самоощущения (fornit.ru/160).
В первую очередь после полного отсутствия сознания происходит попытка найти наиболее подходящий контекст понимания. При этом ориентировочный рефлекс, независимый от такого контекста (точнее, ограниченный лишь контекстом гомеостатической значимости при отсутствии иного), прекрасно работает и привлекает внимание ко всему воспринимаемому, выдающемуся по актуальности. Но мы не можем сразу понять смысл окружающего, т.е. выбрать определенный контекст понимания смысла происходящего, вплоть до того, что означают те или иные картины зрительного или слухового восприятия, - они оказывается бессмысленными наборами примитивов восприятия:
“…он очнулся и вспомнил все, еще не успев открыть глаза. Но это было далеко и давно. Он открыл глаза и вместо белой постели в больничной палате увидел беспорядочно мелькающие цветные пятна и полосы. Мучительно долго все увиденное собиралось в образы, и этот процесс походил на безумие. Потом он понял, что смотрит на мерцающую рябь воды прямо у его лица” (fornit.ru/10888).
Все воспринимается как бессмысленная каша, не за что ухватиться, но наш организм уже способен реагировать на это чисто рефлекторно в той степени, в какой такие реакции на внешнее были закреплены на рефлекторном уровне.
В этот момент доминирует чисто рефлекторное восприятие, хотя оно и отражается уже в канале осознания, но в совершенно пустом, самом общем контексте, где что угодно может иметь смысл чего угодно. Отсутствуют какие-то осмысленные переживания, кроме переживания непонимания и растерянности, так что мысли уже есть, они никак не связаны с воспринимаемым (настолько абстрактны, что, кажется, исходят из какого-то управляющего всем существа). Мы можем задаваться вопросами: кто мы и где мы потому, что это относится к самому общему контексту, а у нас есть мыслительные автоматизмы, обслуживающие такие мотивационные цели, которые могут инициировать самый общий исследовательский контекст. Поэтому, даже еще не сориентировавшись в том, где мы, мы можем понять, кто мы, если удачно привлечем эпизодическую память в таком контексте, что позволит потянуть ниточки к возможным контекстам моделей себя в самых различных условиях.
Наконец распознается что-то в окружающем – как только подбирается подходящий контекст, в котором виденное приобретает определенный смысл: активируется модель понимания данного объекта внимания и переживания обретают более определенный смысл, все встает на свои места хотя бы в вопросе - что нас окружает.
Далее последует распознавание остального, что может повлечь понимание того, где мы, и помочь вспомнить то последнее, что с этим было связано, если нас в бессознании не переместили совершенно в другое место.
Наконец полностью уточняется соответствующий контекст понимания места и времени, поддерживая последние воспоминания, и все приходит в порядок, обретя полноценный смысл происходящего с сопутствующими этому мыслительными автоматизмами.
Всякий раз нам помогают именно распознаваемые образы восприятия и кадры эпизодической памяти, как-то связанные с такими образами. Без этих образов мы не смогли бы сориентироваться и начать уверенно мыслить в определенности понимания ситуации. Такое состояние способствует появлению посленаркозного психоза.
В описанном процессе восстановления понимания вовсе не предположительно, а вполне обосновано есть начальный момент, который у разных животных субъективно (по переживаниям) отличается лишь набором автоматизмов (жизненным опытом) наиболее общего контекста. Совершенно не играют роли различия в распознавателях уровня рефлексов и набор имеющихся реакций, распознаватели запуска которых молчат без пусковых стимулов. Это состояние очень показательно, как исходная точка понимания сути субъективного переживания.
В первую очередь, проявляется полное абстрагирование от всего самой системы произвольности, что приводит к одной абстракции, которую грубо можно обозначить как “я есть”, - одной на всех, у кого она актуализируется (вспомним свойства абстракций).
Вот статья, описывающая феномен базового самоощущения: fornit.ru/160, и отсюда следует множество важных и шокирующих выводов.
Итак, у всех животных, обладающих каналом осознанного внимания в момент начала активации системы произвольности (сознания) эти начальные переживания совершенно идентичны как субъективные абстракции при том, что они могут быть реализованы самой разной схемотехникой или, в принципе, даже программно. Такая абстракция идентична не в механизмах организации и процессах активации сознания, а именно в субъективном символизме происходящего. И это – суть субъективного мира, который – нечто совершенно несвязанное хоть чем-то материально с миром объективным потому, что объективный мир обладает физическими свойствами (определяемыми фундаментальными взаимодействиями, что обеспечивает причины и следствия событий), а субъективный – нематериальными, символьными значениями абстракций причин и следствий, которые могут быль любыми в меру возможностей имеющихся распознавателей образов (см. про нематериальность мысли: fornit.ru/1132). И в этом нет ничего сакраментального так же, как нет такового в смысле вообще чего-либо, например, смысле прочитанного слова или увиденного на экране компьютера при том, что само это слово или изображение на мониторе не имеет ничего общего по свойствам с этим индивидуально возникающим смыслом.
То, как возникает определенный смысл в голове и что такое смысл вообще – мы уже рассматривали. Конечно же, требуется достаточно много времени, чтобы определенные последовательности причин и следствий в процессах схем механизмов адаптивности связались с пониманием явлений субъективного самоощущения. И тогда будет ясный ответ на вопрос № 1: “Соотношение мозга и разума”. Это – то же самое, как понять частные случаи такого соотношения в явлениях:
· что означает цифра 1 в данных конкретных условиях для данного человека или кошки;
· что означает зеленый цвет (fornit.ru/279) в данных конкретных условиях для данного человека или кошки;
и т.п. для все более сложных субъективных смыслов.
Какой бы и в виде чего цифра 1 ни была, она в общем виде распознается теми, для кого это было значимо, но в таком общем контексте она слишком многозначительна и воспринимается слишком абстрактно, не воплощаясь в конкретное субъективное переживание. В конкретных же ситуациях она приобретает определенный смысл и составляет субъективное переживание понимания сути происходящего в контексте целевой возможности что-то сделать сейчас или позже. Именно смысл и составляет основу субъективного мира.
Более зримо воспринять суть самоощущения может помочь интерпретации: fornit.ru/50394 и fornit.ru/50497.
Прямо сейчас не станем погружаться в философию субъективных переживаний, а проследим развитие главного функционала сознания для адаптивности поведения, а это эволюционно наращивалось так, что появилось несколько все более эффективных уровней реализации для единственной цели - определиться в последствиях нового обстоятельства для привычного реагирования.
Мы остановились на том, что подключение канала осознанного внимания к образу, в котором появилось что-то важное-новое, означает одновременно приостановку выполнения соответствующего текущему контексту ответного действия (в том числе ответной мысли) и понимание нового смысла возникшей ситуации с помощью выбора подходящей модели понимания. Эта модель достаточно инертна, как и контекст значимости, она конкурирует с соседними альтернативными моделями так, что изменить смысл воспринимаемого бывает не так просто, если для этого нет достаточных оснований. Все воспринимаемое интерпретируется (получает смысл) в данном контексте, но уточняется более частным, вложенным смыслом, определяющим варианты возможного реагирования.
Уже имеющийся опыт заготовил для разных видов нового разные вложенные контексты в модели и если, осознав ситуацию, мы понимаем негативные последствия привычного поведения в этих условиях, и есть заготовленный вариант поведения с лучшим результатом, то готовое сорваться в действие привычное моментально переключается на другой вариант, и мы обходим новую лужу на привычной дороге.
В процессе осмысления возникает оценка личной значимости того, чем может закончиться ситуация в разных вариантах поведения, что уже заготовлено как цель каждого действия (о которой говорилось в главе Произвольность), и, при этом, возможно, – смысл негативной значимости привычного реагирования в условиях нового обстоятельства, которое сводит все на нет (это – тоже результат уже имеющегося опыта схожих ситуаций).
Задача мыслительных автоматизмов в каждой особенности ситуации и подготовленности к ней в случае простейшего алгоритма – выбор лучшего из имеющихся в заготовке вариантов в процессе самоорганизации ментальных Правил.
Уровни сознания
Уже говорилось о четырех уровнях вовлеченности функции осознания во все более проблемное осмысление в реализации Beast.
Каждый уровень вовлеченности соответствует новому апгрейду эволюции для реализации более специализированной адаптивности к новому.
Как и в других частях мозга, более древние структуры оказываются наиболее общими и поэтому более стабильными, неприхотливыми, а самые новые образования легко могут оказываться неработоспособными при ухудшении условий работа мозга (отклонение состояния метаболизма от нормы, снабжение кислородом и глюкозой, воздействие мешающих нормальной передаче сигналов между нейронами химических и физических факторов: температурой, конкурирующими с нейромедиаторами или изменяющие поляризацию мембраны химическими веществами).
Далее станет видно, насколько же много последовательных эволюционных улучшений развито только на уровне функциональности сознания.
Вот предположительный вариант общего работы произвольности в виде блок-схемы (принципиальная блок-схема реализации Beast: fornit.ru/beast0):
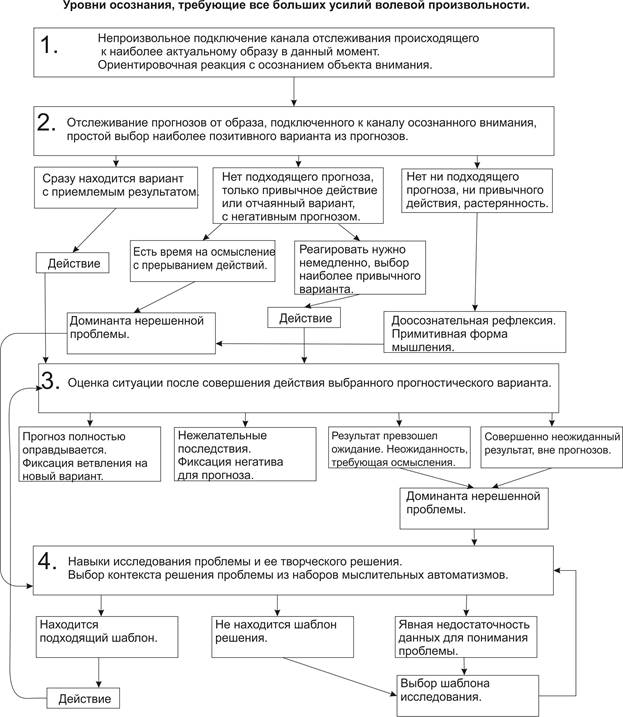
Эта блок-схема приведена не для того, чтобы погружаться в детали реализации описываемых, это было бы слишком специально и предположительно (второй довод - весомее), а дана для общего представления о том, что берет на себя сознание и как может это проделывать. Это – достаточно очевидная логика, доступная в самонаблюдении и во многих случаях описанная психологами и исследователями отдельных видов творческой активности. Ее каркас основан на уже имеющейся модели понимания механизмов МВАП (fornit.ru/7431) и подтверждена реализацией прототипа.
Здесь условно выделяется четыре качественно различающихся по функциям уровня вовлечения механизмов произвольности сознания. В реализации Beast они описаны: fornit.ru/beast7.
Можно заметить, что до четвертого уровня произвольность проявляет себя довольно незаметно для самого субъекта. Но четвертый уровень можно посчитать уровнем волевого контроля, потому как именно его автоматизмы вырабатывают новые решения, которые требуют волевых усилий для их принятия вместо привычного. Иногда это привычное настолько сильно значимо, что требуются предельно высокие усилия произвольности (“силы воли”, или мыслительных автоматизмов преодоления привычного, что нарабатывается в тренировках) для замены их на альтернативное реагирование.
Т.е. до четвертого уровня произвольность оперирует уже имеющимися образами действия, выбирая лучший из них для новых условий. Для подстановки вместо одного варианта (наиболее привычного в данных условиях) другого (тоже привычного, но в других условиях) не требуется субъективно переживаемых усилий (волевых усилий). Другое дело, когда возникает сложная ситуация с непонятными последствиями, и тогда предположительный вариант, явно рискованный из-за его непроверенности на деле, уже вызывает серьезные колебания и переживания возможных последствий. А преодолению такой неуверенности и того, что придется, возможно, испытать негативные ощущения при попытке поступить иначе, еще нужно учиться, чаще всего на чужом примере.
Волевые усилия нужно научиться прикладывать, что известно как развитие самодисциплины (основу которой также составляет подражание авторитетам). Вот почему четвертый, волевой, уровень сознания подавляется во время сна (fornit.ru/5084) при том, что предыдущие оказываются вполне активными для сновидений.
Можно сказать, что на четвертом уровень – муки творчества и сомнения, а до него – радость от умелых навыков, уже проверенных в деле. Область волевого интеллекта или творческий уровень сознания позволяет адаптироваться с совершенно новым качеством и требует смелости для проверки предположений на деле. Такое сопоставление очень полезно для понимания сути субъективных переживаний.
Именно доминанта нерешенной проблемы оказывается тем, что провоцирует активность четвертого уровня. Без него бытие мало отличается от механизма сновидения, в том числе и общение, при котором непринужденные разговоры протекают именно по механизму сновидения, только коллективного сновидения. Более простое представительство таких доминант возникает всякий раз, когда во время бодрствования встречались новые важные ситуации, но не было времени их осмыслить так, что их активность поддерживается их высокой значимостью до периода сна.
Как только возникает заминка в получении приемлемого решения для действий, включается особый режим функции осмысления: циклы итераций рекурсивного вызова функции из самой себя – циклы мышления.
Суть проста: в текущей информационной картине выбирается наиболее подходящий ментальный автоматизм для получения дополнительной информации (в случае имеющего опыта решений выбор следует из цепочек ментальных правил). Новая информация дополняет самоощущение и если все еще не получается найти решение, то запускается новый вызов функции осознания, уже с обогащенной информаций. И т.д. до решения проблемы или откладывания ее в виде доминанты нерешенной проблемы. При каждом обновлении информации самоощущение меняет картину понимания.
Эти самовызовы происходят без затрат времени на мышечные усилия и обработку восприятия, они очень быстро следуют один за другим (их скорость сопоставима со скоростью циркуляции активностей во время сна: fornit.ru/5376).
Прервать такой цикл может только новый реальный стимул, который перезапускает функцию осознания с новым образом, требующим внимания.
Перечислим особенности уровней вовлечения сознания.
Первым уровнем осознанного реагирования является автоматическое, непроизвольное подключение канала осознанного внимания к наиболее актуальному в данный момент образу. Это - очень логично: в первую очередь необходимо обращать внимание на наиболее важное, а не тратить время на мелочи и пропустить большую опасность. Когда будет отработана ответная реакция для самого актуального, его место для контроля займет следующее по важности.
Ориентировочный рефлекс означает необходимость обратить внимание на текущий активный пакет информации с целью проверить, насколько подходит штатный автоматизм, а если такого нет, то создать пробный и запустить. В самом общем плане, если есть надежный полезный автоматизм для данных условий, то не нужны никакие размышления, можно просто выполнить его.
Конкуренция по актуальности образов может быть настолько велика, что буквально несколько разных событий требуют осознанного внимания и перетягивают на себя канал внимания хаотично-случайно. На этот случай природа выработала механизм быстрого переключения внимания с одного на другое так, что создается видимость одновременных осмысленных действий, причем эта видимость возникает не только у наблюдателей, а у самого субъекта (возможно потому, что для удержания образа общей картины на стороне структур сознания используется самоподдержание общей цели, но, скорее всего, субъективное ощущение непрерывности и последовательности - просто результат развития отношения к этому, по сложности предположительной модели это выходит за рамки книги). Успев наспех подкорректировать действия для одной цепочки реагирования, сознание тут же схватывает другую ситуацию.
Есть еще один наблюдаемый механизм: отложенное реагирование при появлении более актуального. Цепь отслеживания смысла прерванных действий у разных животных разная и у человека – самая крутая, доходя до 5-7 удержаний в “стеке памяти”.
В памяти остаются следы цепочки последовательности всех действий - как общая картина происходящего с участием в ней, называемая эпизодической памятью. Эта память позволяет получать прогнозы (в виде Правил прошлого опыта) более емко и обще, чем прогнозы от множества отдельных независимых цепочек действия. И такие прогнозы отвечают доминирующим целям, что используется как пережитый опыт.
Частным случаем первого уровня сознания оказывается ситуация, когда в восприятии нет настолько значимого и нового, чтобы представлять важность для отслеживания, и возникает состояние ожидания событий (“пассивный режим” работы мозга) и ленивое состояние (fornit.ru/652). Само по себе ожидание событий может быть достаточно важным (активным) и тогда возникают последующие уровни функционирования сознания, когда нужно быть настороже и быть готовым вовремя среагировать. Это выделяется в отдельный стиль поведения, который развивается жизненным опытом. Но при полном отсутствии опасностей и монотонном окружении возникает торможение отслеживающей функции сознания с переходным периодом, схожим со сновидением. В специальной изолирующей камере, в которой помещались испытуемые так, что ничто не могло хоть как-то воздействовать на них, вскоре возникал период самопродуцированных видений и сон.
Второй уровень осознания обслуживает уже подключенный канал осознанного внимания от наиболее актуального звена одного из выполняющихся автоматизмов к субъективной модели. Это обслуживание осуществляется с помощью прогнозов при рассмотрении цепочек Правил, опережающих намечающееся действие. В результате происходит произвольный выбор заготовленного ранее варианта, опережающе показывающего наилучший результат, т.е. наибольшую позитивную значимость.
Выбор подходящего варианта оказывается настолько эффективным, насколько имеется достаточный опыт в виде различных подходящих вариантов и насколько часто субъект прибегает к такому режиму отслеживания. Это - результат приобретаемого навыка, когда время, уделяемое одному событию, до предела минимизируется так, чтобы не тормозить размышлизмами совершаемые действия.
Если собаке протянуть одновременно кусок мяса и комок травы, то она, не раздумывая, схватит кусок мяса. Если протянуть два куска мяса, то она схватит тот, что ближе или тот, что больше или тот, что более чем-то для нее привлекателен. В последнем случае она может несколько замедлить действие, выбирая.
Обычно вторым уровнем осмысления ограничиваются такие действия как движение по улицам знакомого города (Правила типа “карта местности”), игра блиц (не раздумывая), непринужденный разговор, импровизация, ремесленническая деятельность и т.п.
Для второго уровня сознания характерны следующие случаи.
1. Сразу находится такой вариант, который с учетом новизны происходящего, приводит к наилучшему результату. Но, стоит обратить внимание, это - пока предполагаемый результат. Это может быть и тот привычный вариант, который бы был использован без вмешательства сознания. Но в новых условиях он обретает предположительную возможность неудачи из-за пока неясного воздействия новых обстоятельств. Так, в окружении новых людей может ощущаться некоторая неловкость.
2. Прогноз не дает достаточно хорошего в последствиях результата. Нет достаточной уверенности в том, что-то действие, которое сразу приходит в голову в такой ситуации, закончится позитивно, а не приведет к беде. Здесь возможны два обстоятельства:
1) Насущность немедленного реагирования настолько высока, что делать хоть что-то, но придется. Например, разливающаяся раскаленная лава заставляет отступить, но впереди ров, ширина которого превышает уверенный прогноз перепрыгивания. Каждый прыгнет на удачу в таких обстоятельствах, особенно когда начнет нестерпимо припекать. Но иногда риск от опасности меньше, чем риск от опрометчивых действий, и тогда от большего риска отказываются. Тут все очень индивидуально и напрямую зависит от уже имеющегося жизненного опыта.
Если опыта вообще нет, то совершаются опрометчивые и даже бессмысленно-хаотические действия. Опрометчивое действие - крайний вынужденный вариант, характерный при недостаточности опыта, и даже если он удался, проблема выхода из сложившейся ситуации остается актуальной потому, что пережитая опасность не забывается, а оставляет след нерешенной проблемы высокой значимости. Удачная попытка увеличивает уверенность и уменьшается необходимость решения проблемы. В следующий раз в подобных ситуациях будет легче решиться действовать таким образом.
2) Ситуация позволяет не реагировать немедленно. В таком случае действие прекращается для придумывания выхода из положения. Это - очень важный момент - возможность прерывания действия тормозным влиянием сознания на текущее звено выполняемой цепи автоматизма - вариант произвольности управления вниманием. Это - свойство именно канала осознанного внимания по отношению к наиболее актуальному образу, который к нему подключён для отслеживания. Теперь на месте звена прерванного автоматизма долговременно блокируется его продолжение и возникает память о нерешенной проблеме - оценка сознания об отсутствии варианта и актуальность необходимости ее решения - в зависимости от того, как неизбежно ожидается ее повторение в будущем. Этот образ фиксируется долговременно (примерно в течении получаса) и оказывается доступным, даже если активность будет чем-то прервана - для воспоминания и осмысления. Высокая значимость такой ситуации (а раз было привлечено внимание, то она достаточно высокая) и ожидание повторения могут поддерживать этот образ в постоянной активности - в виде доминанты нерешенной проблемы.
В результате осознания проблемы формируется ее произвольная значимость. Она может оказаться меньше той, что вызывала привлечение осознанного внимания, и тогда объект внимания уйдет на задний план проблем, замещаясь чем-то более важным. Но если осознанная значимость подтверждается как самая актуальная на данный момент проблема, то теперь ориентировочная реакция будет усиленно отслеживать все, что касается этого объекта, переводя это в область осознания. Если мы почувствовали голод (т.е. осознали его, иначе бы он не почувствовался), и осмысление этого показало, что это - самая насущная проблема, то теперь все восприятие будет особенно чувствительно ко всему, что связано с возможностью удовлетворения голода. Самоподдерживающаяся доминанта активной нерешенной проблемы своей значимостью задает общий контекст, что теперь удовлетворению голода придается вот такое повышенное значение. На данном уровне сознания задается контекст для более древних зон рефлексов (теменные ассоциативные зоны) вплоть до переключения базового эмоционального контекста (эмоции, стиля поведения) и более поздних зон автоматизмов (лобные ассоциативные зоны) - активизация моделей поиска и навыков добычи еды субъективных моделей. Все, что вдруг обнаруживается на уровне первичного восприятия как связанное с едой, теперь выходит на передний план, привлекает внимание и понимается на основе имеющихся моделей.
Таким образом, произвольность осмысления значимости (смысла) воспринятого оказывает обратное влияние, фильтрует восприятие в данном направлении, управляя направленностью внимания.
Результатом этого, как при необходимости немедленного реагирования, так и возможности отложить действие, оказывается появление доминанты нерешенной проблемы в контексте текущей, более общей модели, возвращающей на себя сознание всякий раз, когда нет чего-то более актуального. Это - свойство любых важных нерешенных проблем.
3. Вообще не находится подходящего варианта реагирования, - полная растерянность. Субъект попадает во власть обстоятельствам, и его реакции ограничиваются доосознаваемым уровнем следования уже имеющимся автоматизмам. Это состояние вызывают гипнотизеры, шокируя чем-то недостаточно опытных своих клиентов.
При этом может возникнуть модель нерешенной проблемы и будет возвращать сознание на себя в более спокойной обстановке и/или во сне в виде сновидений (как и нерешенная проблема второго уровня). Это произойдет, если важность действия будет прогнозироваться и впредь так, что будет поставлена цель ее решения.
Если какой-то вариант действий выбран, то появляется небольшой, но существенный нюанс: выбранное действие, даже если оно оценивается как очень уверенное, но несколько меняющее привычный процесс реагирования, требует определенной решительности для его выполнения, - волевого усилия. Это - очень важный аспект, который будет рассмотрен ниже, после перечисления уровней сознания.
Все перечисленные случаи второго уровня легко моделируются схемотехнически, позволяя хорошо представить себе варианты их искусственной реализации.
Более развитая способность данного уровня сознания позволяет не ограничиваться первыми результатами прогноза, а если позволяет время, чуть задержаться на этой стадии, и по результатам опережающего прохода, оказавшись на конечном образе одного из вариантов, посмотреть новый букет прогнозов уже оттуда, расширяя поле возможных предсказательных последствий. Это - очень важный и очень востребованный прием. Каждый такой шаг по ступеням последовательности вариантов улучшает общее понимание возможностей (набор учитываемых вариантов для выбора) и, в последующем, оказавшись запомненным, позволяет сразу перейти от проблемного образа к конечному варианту прогноза, уже не требуя столько времени. Это напоминает развитие сценария сновидения, но с осознанно управляемым ветвлением.
Часто действие позволяет одновременно с его выполнением отвлекать сознание на такой последовательный просмотр возможных последствий, который может продолжаться довольно долго, пока прогнозы не исчерпают текущий контекст, и тогда конечное действие оказывается более выверенным без его прерывания. Но и во время прерывания именно такой вариант мысленного прохода обеспечивает самый простой способ найти приемлемое продолжение действий. Таким образом, если время позволяет так раздумывать, то становится возможным оценивать значимую составляющую образов как бы со стороны, в виде мотивирующего субъективного переживания.
Третий уровень эволюционного усложнения логики функционирования осознания начинается при отсутствии решения на втором уровне осознания. Если не удается решить проблему за счет мышления сразу, то возникает доминанта нерешенной проблемы и мыслительные процессы ее решения (если есть на это время). Обычно такое бывает после совершения пробного действия (второй уровень не дал решения и было сформировано и выполнено предположительное действие) и появления результата эффективности действий. Возможны следующие варианты.
1. Прогноз полностью оправдывается, и в новых условиях получилось предполагаемое. Действие оказалось адекватным объективной реальности: предполагаемое соответствует получаемому. Для признаков новых условий, которые вызывали привлечение сознания, от данного звена автоматизма долговременно формируется ветвление на новый, испытанный вариант. В дальнейшем, уже может не потребоваться осознания, а сразу, в таком уже привычном случае будет совершено это ветвление. Или же эта привычка окончательно закрепится и перейдет в неосознаваемое за еще несколько эпизодов осознания.
2. Прогноз не оправдывается, и в результате в новых условиях получаются нежелательные последствия. Действие оказалось неадекватным реальности: предполагаемое не соответствует получаемому. В данном звене автоматизма долговременно закрепится ветвление с негативной оценкой, и в последующем такой вариант окажется исключен в прогностическом выборе. Впредь такое поведение будет избегаться.
Схемотехнически очень просто заблокировать его выполнение так, чтобы даже вне осознания такая реакция в данных обстоятельствах не выполнялась. Как именно механизм такой блокировки реализуется в мозге, пока точно не выяснено, но принцип таков: в последующих сходных обстоятельствах ранее предполагаемый прогноз теперь имеет уверенно негативную оценку и не будет применен, а прерванное действие не завершится, что зафиксируется долговременно.
В случае, если конкретные признаки новизны условий оказались не распознанными еще на первом уровне, т.е. сознание не было привлечено до этапа действия, а только неожиданность негативного результата привлекла сознание, косвенно определяя, что вмешалось нечто, изменившее результат привычного действия, то вся привычная цепочка поведения покажется странной или даже вообще избегаемой. И только настоятельная необходимость все же попробовать опять действовать так же может это превысить. Если опять будет неудача, неуверенность станет еще большей. А если неудача оценивается слишком сильным негативом, то и вообще возникнет тупик полной блокировки дальнейших действий прежним способом.
3. Прогноз не только оправдывается, а результат приносит значительно больше позитивного, чем предполагалось. Это - тоже неожиданность, хотя и принесшая позитив, а, значит, таит в себе опасность непонимания тем, что предположение лишь по какой-то случайности не привело к негативу. После долговременного закрепления такого ветвления он оказывается предпочтительным в последующем прогностическом выборе в силу высокой позитивной значимости результата.
4. Результат оказывается совершенно неожиданным, не таким, каков был образ конечного звена цепи выбранного варианта реагирования даже с учетом не удачности его реализации. Порождается образ нерешенной проблемы, описанный выше, требующий дополнительного исследования и понимания ситуации.
Таким образом, проявляется иерархия алгоритмов сознания по отношению к отслеживаемому наиболее актуальному образу автоматического реагирования.
Перечисленные случаи не вызывают принципиальных сложностей в их схемотехнической реализации у опытного схемотехника (это – вопрос изобретательства, что само по себе не просто), который способен по заданной цели вообразить принципиальную схему устройства для ее реализации. Вот только как это реализовано в природном мозге – совершенно иной вопрос, требующий еще очень немалых исследований.
Эти алгоритмы сознания возникали как механизмы адаптивности к новому и последовательно усложнялись по уровням и ситуациям отслеживания происходящего.
Все перечисленные случаи, кроме первого, требуют понимания случившегося так, чтобы стало возможным каким-то методом подобрать подходящий вариант реагирования для этих новых условий. Они задают нерешенную проблему, той или иной степени актуальности. Даже во втором случае неожиданной удачи это заставляет задуматься. Как правило, в результате одного из таких случаев дальнейшие действия прерываются для осмысления. Возникает нерешенная проблема, требующая исследования и поиска выхода.
Четвертый уровень осознания проявляется процессами мышления и формируемыми навыками исследования проблемы и ее творческого решения, что всегда сопровождается активностью доминанты нерешенной проблемы. Подробнее логику и механизмы исследовательского поведения и творческого решения проблем (создание новых вариантов) рассмотрим позже, а пока важно понять, что для нахождения подходящего решения нужно выявить какие-то новые условия происходящего, которые могут прогностически подсказать решение. Т.е. сначала - исследование проблемы, потом - попытка подобрать имеющийся вариант или сотворить новый.
Когда мы подходим к луже воды на дороге, видим, что она занимает все место и ее не обойти, не то, что перепрыгнуть, то останавливаемся и присматриваемся к свойствам лужи. Видим, что везде из нее торчат неровности дороги, что означает - она неглубокая, а у нас обувь достаточно высока. Мы сопоставляем возможную глубину лужи с высотой обуви и при таких условиях возникает подходящий вариант пройти прямо по воде, мы уверенно решаемся на него. В случае неудачи приобретаем опыт избегания такого решения и более внимательного выбора варианта.
Метод сопоставления параметров - один из наиболее общих в исследовании, а метод обобщения сопоставленных данных - наиболее общий для решения.
Часто проблема не решается так просто и остается нерешенной долгое время, бывает годами. Доминанта легко активируется из памяти списка существующих нерешенных доминант (“гештальт не закрыт”) - если есть время на раздумья и значимость доминанты превышает остальное, в том числе и значимость других доминант. Активность образа доминанты самоподдерживается реверберацией за счет обратной связи и ввиду большой силы актуальности, бывает, не гасится даже во сне, направляя сновидения в своем контексте, но, даже если она окажется погашенной, легко снова активируется при малейшей ассоциации со сходными элементами восприятия.
Каждый из перечисленных уровней осознанной коррекции происходящего требует последовательного развития дополнительных слоев мозга в зоне лобных долей для формирования все более сложных распознавателей на основе предыдущих более простых. Чем более долгим оказывается период формирования таких слоев, тем более "интеллектуальным" будет поведение.
У человека наибольшая продолжительность формирования всех периодов развития мозга среди других животных.
Уровни лобных долей, соответствующие самостоятельному нахождению решений, у людей созревают в среднем к двенадцати годам, что так же подтверждается окончательным формированием структуры гиппокампа, переключающего доступ к лобным долям от образов восприятия-значимости-действия. Это следует учитывать в программах обучения и воспитания.
Хотя предлагаемая классификация «уровней сознания» напоминает классификации «уровней рефлексии» в психологии (например, Л.С. Выготский выделял шесть уровней рефлексии), но выводится на уровне схемотехнической блок-схемы, образованной при сопоставлении всех выявляемых компонент механизмов организации психики.
Тестирование сознания
Каждый из описанных уровней активности механизмов сознания может быть вполне адекватно и достоверно протестирован и распознан. Так, первый уровень (и, соответственно, все последующие) – простое проявление ориентировочного рефлекса с прерыванием реагирования.
Второй (и, соответственно, все последующие) характерен возможностью не следовать наиболее привычной реакции в случае, если появились новые признаки, которые явно означают возможность неудачи привычного действия в таких условиях. Так, если на привычной дороге, по которой человек ходит часто и постоянно, вдруг появилась лужа, то только бессознательно пьяный пойдет прямо по ней. Т.е. достаточно выявить часто и привычно совершаемое данным человеком, добавить новый фактор, делающий привычное действие нерациональным, и посмотреть на результат. Это и есть наиболее общий тест на сознание.
Так же можно тестировать все уровни сознания, например, второй уровень, проверкой способности выбрать из нескольких вариантов (предметов, действий) наилучшее. Вот более-менее подходящий тест на сознание: Тестирование уровня сознания: fornit.ru/6581.
Контекст понимания смысла. Личность.
Это – наиболее сложная и информационно емкая глава, фактически - окончательное сведение представлений о произвольности в одну систему.
Понимаемый смысл
Сочетания условных символов (взаимно принятых у данной контактирующей группы особей, но изначально взятых у учителя) с возможными реакциями в зависимости от особенностей условий и ситуации (в том числе собственного состояния), происходят в определенном контексте текущей ситуации. Так же как контекст значимости теменной коры сужает диапазон воспринимаемых символов, связанных с реагированием (вплоть до единственно определенного), так и в лобной коре формируется контекст, ограничивающий диапазон произвольного управления реагированием вплоть до предельной определенности (в реализации с помощью деревьев это означает, что для любой уникальной ситуации есть ветка, активирующаяся при наличии такой ситуации). Мы уже называли его контекстом понимания смысла происходящего или субъективного понимания значимости выделенных объектов внимания канала осознания, что позволяет строить модели значимости (для себя, а не для других) любых других объектов внимания, живых или нет – не принципиально, в том числе – других особей и сопереживать им.
Этот контекст может определять вполне однозначный, уверенный смысл (значимость, привязанную к данному контексту) или может оказаться многозначительным, если нет достаточных признаков для уточнения контекста. Но предельно конкретный смысл может таить в себе пока незнакомые особенности условий и ситуаций, когда нам кажется, что мы уверенны и все понимаем (fornit.ru/1682), а на самом деле просто еще чего-то не знаем.
Если гомеостатическая значимость представлена распознавателями от первичных до ассоциированных зон, связанных со вторичными зонами каналов рецепторного восприятия и третичными зонами теменной коры, то иерархия смысловой значимости локализуется в лобных долях (fornit.ru/7146), отражая произвольно оцененные (в целевом назначении) состояния гомеостатической значимости. Именно эта иерархия определяет вложенность все более уточняющихся контекстов понимания смысла происходящего в его целевых возможностях реагирования.
Эмпирические данные природной реализации субъективной значимости (fornit.ru/24354) отвечают системной логике организации произвольности, которая описывалась в прошлых темах, в том числе алгоритму уровней осознания проблемы в контексте гомеостатической значимости новизны происходящего. Но пока отсутствует достаточная системная конкретизация механизмов формирования основ природной системы осмысления, хотя ясно, что ее активность всегда отражает какую-то проблему жизнеобеспечения, как первичную, так и более высокоуровневую в представительстве контекста значимости.
Важно то, что это – всегда новая проблема (иначе нет осознания), которая, может быть, как-то и решается уже имеющимися стереотипами реагирования, но еще нет привычной последовательности применения этого в новых условиях. Т.е. проблема отражает гомеостатическую актуальность, но в ходе выбора, учитывающего новизну решения, эта значимость должна иметь возможность меняться произвольно, обеспечивая оценку удачности решения для заданной цели.
Так, чтобы вырваться из кольца огня, нужно преодолеть значимость ожогов для того, чтобы избежать худшего – сгорания, или суметь отрезать себе конечность, чтобы освободиться.
На уровне гомеостатической значимости это – нерешаемые проблемы, а на уровне смысла происходящего возникает возможность абстрагироваться от текущей значимости, чтобы сравнить другие варианты, отслеживая, чем могут закончиться те или иные имеющиеся цепочки реагирования и выбрать лучшее, невзирая на потери, не останавливаясь на первом же негативе, а имея возможность подсмотреть, что будет, если проигнорировать этот негатив.
Всякий раз решение того, что в каких случаях предпочесть и что выбрать, формирует мыслительные автоматизмы методов такого выбора.
В самом общем плане это означает возможность заглянуть в будущее дальше, чем простой связью условного рефлекса. Это и есть организация произвольности придания смысла для выбранного (на каком-то уровне сознания) варианта реагирования. Остается преодолеть запрещающие стереотипы для промежуточных действий тем убеждением, что конечное действие приносит значительно большую выгоду, чем временные потери, или, наоборот, суметь не попасть под влияние заманчиво желаемого, а увидеть негативные последствия и выбрать, что предпочтительно.
Это организуется системой преодоления стереотипной значимости, которую называют “силой воли”.
Такая система автоматизмов достаточно универсальна для любых ситуаций необходимости преодолеть промежуточные (негативные или позитивные) значимости и нарабатывается постепенно, как навык преодоления текущих трудностей для достижения цели. Т.е. это – типичный мыслительный автоматизм. Все описанные уровни сознания представлены системой последовательно формирующихся с 4-5 лет до 25-30 лет (наиболее интенсивного развития) мыслительных автоматизмов методов решения проблем.
Возможность наработки новых мыслительных автоматизмов, как и моторных, не определяется возрастом с тех пор и до тех пор, пока работает гиппокамп и другие ответственные за их формирование структуры. Все они возникают в рамках определенного смыслового контекста, наподобие условных рефлексов, возникающих в рамках своего контекста значимости и, затем, оказываются доступны именно в этих своих контекстах – как понимание возможных вариантов реагирования.
Как именно реализуются все эти механизмы формирования смыслового контекста понимания возможностей реагирования – не принципиально для схемотехнического описания: они могут быть выполнены множеством способов, но выявление природного варианта их реализации может подсказать ценные идеи для унификации и оптимальной логики схемотехнической реализации (новые идеи – всегда инициируются восприятием).
В этих условиях для того, чтобы убедительность утверждений теории, выверенная реализацией прототипа, поддерживать адекватной реальности, важна постоянная опора на эмпирические данные, и мы продолжим такие интерполирующие корреляции с системной логикой для большей уверенности в каркасе общей модели понимания организации адаптивности уровня психики, в области механизмов произвольности. К сожалению, сложность описываемых механизмов делает просто невозможной постоянные привязки к каким-то достаточно простым примерам схемотехнической реализации, но нисколько не мешает наработанному схемотехническому стилю мышления представить их вероятные варианты реализации. Для этого нужно постоянно иметь ввиду главную функцию модуля формирования подходящего решения проблемы для новизны ситуации и следующую этому системную логику уровней сознания, формализованную в прошлой главе в виде структурной схемы алгоритма, начиная от самого факта подключения механизмов сознания.
Как определить наличие сознания
При самонаблюдении наличие сознания проявляется самосознанием и результатами мышления (“Я мыслю, значит Я существую”), но проблемой является определение наличия самоощущения у другого существа и, тем более, у программной реализации живого существа. Самым главным тут является понимание механизмов, субъективно переживаемых как самоощущение, вообще – знание таких механизмов (fornit.ru/64924).
Только что рассматривались методы формального тестирования проявлений уровней сознания. Но какой-то этолог может сказать: а вот ящерица обрывает свой попавшийся хвост, значит у нее есть волевой четвертый уровень осознания! Это можно прояснить 1) рассмотрением механизмов адаптивности, которыми обладает ящерица 2) визуализацией активности в отделах мозга.
У ящерицы уже довольно сложная организация мозга, в том числе есть гиппокамп и зачатки лобных долей. Но у ящерицы хвост уже устроен так, чтобы легко обрываться и даже отбрасываться резким рефлекторным сокращением мышц, а затем за пару месяцев вырастает новый. Т.е. это – вовсе не трагическая операция, требующая силы воли, а банальный рефлекс для отвлечения хищника на оставленный хвост. Другое дело волк, перегрызающий свою попавшую в капкан лапу. В природе нет капканов, и волк не застревает в таких ловушках, у него нет никакой физиологической готовности к отгрызанию лапы, которая уже не отрастет заново. У него работает волевой четвертый уровень произвольности, он – творческая особь с развитой волей и самодисциплиной, чего не скажешь про большинство людей, живущих в комфортных условиях.
С помощью томографической визуализации обнаруживается, что в префронтальной лобной коре структуры откликаются своими активностями с четкой корреляцией всех субъективных переживаний, которые носят ту или иную эмоциональную окраску. При этом в других областях мозга нет настолько однозначной корреляции, т.е. структуры, отвечающие за переживания, могут быть активны сами по себе как чистые мыслительные абстракции, без поддержки более ранних структур образного восприятия.
Методы измерения времени распространения сигналов по цепям мозга так же позволяют получить системные корреляты, как это было проделано, в работе А.Иваницкого “Мозговая основа субъективных переживаний” (fornit.ru/768):
“Показано, что ощущение возникает в результате синтеза на нейронах проекционной коры сведений о физических и сигнальных свойствах стимула, который обеспечивается кольцевым движением возбуждения из проекционной в ассоциативную кору, гиппокамп и мотивационные центры с возвратом в проекционную кору. Установлено также, что при мышлении происходит конвергенция корковых связей к определенным центрам, названным фокусами взаимодействия. Их топография специфична для различных мыслительных операций: при образном мышлении фокусы располагаются в теменно-височной, а при абстрактном мышлении - в лобной коре”.
Здесь важна связь процессов самоудержания образа восприятия-действия с подключаемыми структурами лобного отдела, которые обеспечивают интерпретацию “сигнальных свойств стимула”.
При этом всегда такие подключения образов к лобным долям связаны с поддержкой структур ориентировочной реакции. Вне зависимости от того, инициируется ли ориентировочный рефлекс со стороны теменной коры или он срабатывает от детекторов нового в области префронтальной лобной коры (fornit.ru/24208), каждый новый кадр эпизодической памяти, соответствующий моменту текущего субъективного переживания, включается этим процессом выбора наиболее актуального как в области образного, так и в области абстрактного восприятия.
Это прямо означает, что вне канала осознания (вне кадров эпизодической памяти, которые сменяются с переключением фокуса осознанного внимания) нет субъективных переживаний, а также то, что при повреждении этих структур в лобной коре субъективные переживания невозможны вместе со всем набором ставших бесполезными мыслительных автоматизмов.
Вопрос о том, полностью ли составляет эта активность то, что называют сознанием, или в сознании может независимо присутствовать образное восприятие без связи с префронтальной лобной корой – снимается потому, что сознание – только то, что оказывается не только в фокусе, выделенном среди всего ориентировочным рефлексом, но, главное, то, что подключается этим рефлексом со стороны лобных долей (конкретно – система распознавания смысла происходящего).
Об этом трудно судить лишь по внешним проявлениям поведения человека, без точной констатации приоритетной активности ориентировочного рефлекса потому, что это поведение, может оказываться полностью основано на рефлексах и очень тонко наработанных моторных автоматизмах, для чего подключение к фокусу осознанного внимания совершенно не обязательно: рефлексы и автоматизмы могут выполняться самостоятельно, хотя и в рамках текущего контекста значимости или элементов контекста понимания на стороне теменной коры мозга (вспомним, что “зеркальные нейроны” присутствуют как со стороны теменной, так и лобной коры, связывая образы восприятия и образы целей действия).
Судить о наличии или отсутствии осознанности возможно только, тестируя проявления ориентировочного рефлекса и проявления уровней функциональности сознания, самый простой из которых – оперативная подстановка более адекватных текущим особенностям вариантов поведения в новых условиях, которые не являются наиболее привычными, стереотипными в данном контексте. Так, если человек не замечает новых деталей и действует привычно так, как будто их и нет, то сознание отсутствует в его наиболее простой отслеживающей функциональности, человек действует как пьяный, шагающий через лужу на привычной дороге, которая всегда до этого была без лужи.
Так бывает, когда все вокруг настолько привычно однообразно, что не отрабатывает ориентировочный рефлекс, или когда человек погружен в свои переживания и проблемы, ориентировочный рефлекс отрабатывает по мыслительным автоматизмам. Стоит глубоко задуматься и поведение обходится без коррекции поступков осознанием, человек совершает довольно нелепые, неуклюжие поступки. С возрастом таких случаев становится все больше.
Чем более глубок уровень вовлечения сознания, тем труднее он поддерживается и используется, если только человек не специализировался на этом и не привык использовать его. Поэтому четвертый уровень в привычной обстановке проявляется редко.
Как уже говорилось, контекстные модели понимания представляют собой отражение причинно-следственных связей в виде Правил и Значимостей, которые выверяются и все более уточняются до полной уверенности во всех случаях – отсутствия неопределенностей и нерешенных проблем своего поведения в этих контекстах - до полной привычности. И чем более выверены модели, тем больший ориентировочный рефлекс вызывают наблюдаемые несоответствия привычному пониманию из-за ломки, казалось бы, надежных представлений.
Когда собакам или обезьянам показывают фокусы с предметами, они приходят в замешательство, проявляют предельный интерес и предпринимают попытки выяснить причину столь очевидного абсурда. У них рушатся основы их представлений, что сметает привычное реагирование, переводя в контекст исследования и решения очень важной проблемы. Это - высший уровень механизмов сознания.
Функционирование моделей понимания
Иерархия контекстов моделей понимания не всегда отражает иерархию системы значимости отклонения жизненных параметров и возврат их в норму, настолько, насколько гомеостатические значимости оказываются произвольно оцененными в новый осмысленный смысл происходящего для каждой новой нерешенной проблемы. В отличие от непосредственно зависимого от текущего состояния гомеостатической значимости контекста восприятия-действия, смысл порождает осознанные мотивации к действиям в соответствии с целями решения новых проблем, что часто бывает более актуально, чем действия по восстановлению жизненных параметров. И тогда такие проблемы решаются вопреки жизненной необходимости. Фанаты вполне способны убить себя ради какой-то Сверхценной Идеи.
Так же, как с каждым новым рефлексом дополнялся контекст гомеостатической значимости, с каждым осмыслением корректируется и дополняется контекст понимания, формируя модели, отражающие реально наблюдаемые причинно-следственные зависимости. При этом выделяются те свойства объектов внимания, которые 1) важны (интересны, привлекают внимание) наблюдателя, т.е. он уже находится в распознанном контексте объекта внимания, определяющего его возможные свойства и 2) у наблюдателя уже есть собственные реакции, схожие с тем, что проявляются объектом внимания (иначе он просто не увидит это: fornit.ru/830).
К педагогике это имеет самое прямое отношение: совершенно бесполезно пытаться чему-то научить, если ученик не находится в заинтересованном состоянии это узнать для какой-то свой практический цели, если не удалось привлечь и удерживать внимание на последовательности новых и важных деталей урока.
Например, если круглый камень катится под откос, то такое качение уже знакомо, т.к. человек с раннего детства умеет перекатываться с боку на бок и теперь эта возможность так реагировать связывается с наблюдением поведения камня, дополняя модель понимания этого объекта. Теперь, если нужно переместить тяжелый камень, то в голову придет не поднимать его и переносить, а перекатить. Т.е. в контексте модели понимания камня реализуется цель его перемещения в нужное место.
При этом не требуется какое-то новое Правило, а используются имеющиеся их сочетания, отражающие свойства камня. Но навыки умелого и эффективного перекатывания камней формируются с каждым опытом, как Правила, как способы решения проблем отдельных неровностей поверхности, более энергоэффективные приемы качения и т.п., после чего эти автоматизмы уже не используют контекст свойств камня, т.к. его свойства уже не осмысливаются, а просто автоматически используются в контексте модели понимания возведения сооружения.
Почему мы говорим про модели понимания? Модели - потому, что эти абстракции, произвольно оторванные от материальной реальности, но сохраняющие логику ее причин и следствий, позволяют моделировать возможные реакции с возможностью заранее предсказать причинно-следственный результат. При исследовании свойств реальности причинно-следственные факты отражаются в виде абстрактной модели и субъективно ощущается в виде выбранной мыслительными автоматизмами информации об этом.
Модельное отражение свойств реальности (включая и реальность собственных свойств) позволяет находить решения новых проблем, в той же мере адекватные реальности, в какой были модели, а в случае ошибок адекватности корректировать их уже по механизму оценки результата пробного реагирования, т.е. меняя смысл происходящего в сторону более верного понимания.
Так же, как в контексте гомеостатической значимости, в контексте понимания смысла оказываются доступны для осмысления только те особенности воспринимаемого, что соответствуют имеющейся модели данного объекта внимания (со всеми более частными).
![]() При недостаточности признаков восприятия, может быть распознана необходимость активации не той модели, что дает совершенно неверные представления ее свойств, порождающие ошибки интерпретации объективной реальности. Если ошибка проявляется в виде явного несоответствия наблюдаемого и предполагаемого, то модель может быть уточнена и активирована.
При недостаточности признаков восприятия, может быть распознана необходимость активации не той модели, что дает совершенно неверные представления ее свойств, порождающие ошибки интерпретации объективной реальности. Если ошибка проявляется в виде явного несоответствия наблюдаемого и предполагаемого, то модель может быть уточнена и активирована.
Есть немало многозначительных зрительных иллюзий (fornit.ru/e1), в которых разные люди узнают совершенно разные изображения. Есть те, которое могут увидеть только один какой-то смысл или вообще не видят (fornit.ru/830) ни одного.
Другие легко различают два заложенных смысла:
![]() И тогда они как бы переключаются с одного на другой: человек видит одно, а потом, вспомнив про другой смысл, видит совершенно иное.
И тогда они как бы переключаются с одного на другой: человек видит одно, а потом, вспомнив про другой смысл, видит совершенно иное.
Вот на этом примере хорошо можно понять, что значит смысл, придание смысла, передача смысла другому (fornit.ru/7060). Ведь стоит рассказать не видящему, что он мог бы увидеть, и тот буквально прозревает.
Смысл придает увиденному свою интерпретацию и этим дополняет увиденное так, что оно приобретает зримые формы.
![]()
Вот почему даже в подчас очень скупых рисунках угадывается смысл, и рисунок как бы мысленно дорисовывается дополнительной информацией.
Еще есть люди (их очень немного и для них стоит вопрос о нормальности психики), кто способен в любом изображении увидеть сколько угодно разных смыслов, у них так и прут почти галлюцинаторные видения из-за активации одного из более основных уровней дерева распознавания, как если бы по изображению круга виделись сразу и мячики и арбузы и земной шар и все круглое. Категорию, к которой относится кошка, они определяют как “материя”, а не как “животное”.
Подключением свойств подходящей модели (в виде Правил, характерных для данного объекта) вероятно занимается гиппокамп потому как рядом локализуются все Правила (fornit.ru/5260, fornit.ru/7229, fornit.ru/5222, fornit.ru/5454, fornit.ru/7057). Это корректируемый мыслительными автоматизмами т.к. инициатива получения информации идет из области механизмов психики. При этом определяется верность выбранной модели, а в случае неуверенности (многозначительности) запускающей исследовательское поведение), отрабатывая по все новым топовым актуальностям воспринимаемого, пока очередной образ восприятия не позволит уверенно распознать подходящую модель понимания. Во время непонимания окружающего ориентировочный рефлекс становится особенно чувствительным вплоть до СВГД потому, что практически полностью оказывается на стороне образов восприятия, проявляя эффект поведения без лобных долей (fornit.ru/7279).
В результате такой деятельности накапливается все больше самоподдерживающихся образов уже в лобных долях (fornit.ru/7202) после того, как они были там активированы ориентировочным рефлексом. В совокупности эти образы представляют профиль распознавания модели понимания. Это – важный механизм обобщения данных для выявления общего для них смысла, о котором мы еще будем говорить. Т. о. вначале происходит сопоставление: поочередная активация отдельных объектов осознанного внимания, и с каждым новым уточняется общий смысл. При меньшем числе совокупных образов смысл может быть только соответствующим им. Он может оказаться неопределенным или даже показаться вполне однозначно понимаемым, но с дополнением другими образами он уточняется в более конкретной распознающейся модели.
Чем больше опыт в данной области (определяемой границами условий), тем меньшее количество ключевых признаков позволяет верно распознать смысл, а, с другой стороны, тем меньшее число ответных действий потребуется для эффективного достижения цели. В этом проявляется количественная характеристика интеллекта: fornit.ru/475.
Бывает так, что что-то увиденное покажется чем-то знакомым вплоть до того, что мы явственно будем это видеть, но при более внимательном взгляде станет понятно, что это – совершенно другое.
Проявления этого механизма сопоставления актуальных образов в некоторой их последовательности активации (то, что дает эффект стека и конечным числом звеньев хранения 5-7 для человека) называют обобщением. В случае, когда это обобщение возникает вне фокуса осознания с активацией нового образа (восприятия или абстрактного), то явление называется интуицией (fornit.ru/1392), а процесс ее воплощения (решение проблем) – эвристическим мышлением (fornit.ru/1647).
Процесс понимания и размышления происходит в контексте текущей информационной картины, уже после распознавания условий, в которых эта информация актуализируется. С каждой новой ментальной операцией получения дополняющей информации, общая картина обогащается, что позволяет еще более уточнить понимание ситуации новой ментальной операцией.
Наиболее широким является информационное ощущение “я есть (мыслю – значит существую)”. Оно возникает при активации сознания после глубокого сна или наркоза. В этом контексте собраны все возможные реакции, которые без конкретизации контекста оказываются взаимно противоположными и не реализуемыми так же, как нереализуемы рефлексы без конкретизации стиля поведения и еще уточнения этого стиля до уровня единственной реакции в зависимости от условий.
При этом возникает вложенная иерархия моделей понимания от самой общей “кто я”, определяющей наиболее общий набор возможного понимания и реагирования, к вложенной “где я”, сужающей набор конкретным местом событий и т.д. до наиболее узкого контекста понимания свойств текущего объекта внимания. С каждым мысленным запросом общая картина все более уточняется.
Наиболее общие контексты самоподдерживаются наиболее долго, наиболее частные – меняются так же часто, как меняется актуальность решения текущей проблемы. Если не было полностью гасящего все активности сознания сна или наркоза, то многие образы остаются самоподдерживающимися и на их основе сразу восстанавливается понимание.
Мысленный пример. При просыпании после глубокого сна все частные элементы понимания недоступны, т.к. они являются более конкретными, вложенными контекстами. Мы не можем понять смысл происходящего. Но как только в восприятии возникает что-то активирующее достаточно общий контекст (свет в окне – значит день, окно – значит я в комнате и т.п.), то восстанавливается информационное понимание окружения. Теперь при восприятии деталей мы сможем понимать и их. А совокупность этих деталей последовательно восстановит смысл происходящего.
Т.к. канал осознанного внимания постоянно и довольно быстро перескакивает с одного объекта внимания на другой, возникает некая текущая общность поддержания актуальной среды проблемной области осмысления – мир субъективных переживаний.
В случае, если в текущей актуальности ориентировочного рефлекса нет в самом деле значимых для мотивации решения проблем (т.е. значимость новизны выше, чем значимость благостного бездействия), то сознание остается на своем самом низком уровне отслеживания, предельно обедняясь субъективными переживаниями (осоловелость). В полностью банальной продолжающейся ситуации сознание может переключаться все реже, что отражается на субъективном представлении о времени (fornit.ru/270) событий и может быть даже вообще заторможено. Стоит закрыть глаза во время длительной поездки в транспорте, даже не погружаясь в сон, а лишь ограничивая зрительную часть восприятия, как и время в итоге будет ощущаться текущим намного быстрее (fornit.ru/2489, fornit.ru/5394).
В случае появления непривычной ситуации может не получиться выбрать подходящую модель понимания-действия и возникает растерянное состояние. Выключаются какие-либо схемы управления произвольностью, остались одни рефлексы, особь замирает в прострации. Это – основа гипнотических состояний.
У самых древних животных в случае распознавания непосредственной опасности без подходящей ответной реакции сформировался рефлекс замирания. И такой рефлекс у разных видов животных можно запустить определёнными сочетаниями признаков. Так, если курицу положить на спину на землю и провести перед клювом черту мелом, она оцепенеет. Ее произвольность обесточена как электролампочка, нужен какой-то стимул, возвращающий в одно из привычных состояний.
У любых животных, когда не оказывается подходящей заготовленной реакции, возникает состояние неопределенности, не активируется никакая поведенческая цепочка. Когда люди оказываются в непонятной, неузнаваемой ситуации, они замирают в растерянности, оказываясь наиболее легко поддающимися для авторитарного воздействия (реликт периода доверчивого обучения).
Это значит, что достаточно вывести человека из текущего привычного для него состояния, которое формируется в процессе понимания ситуации и в этом контексте понимания ему подсказываются подходящие варианты действий, как человек окажется вне каких-то своих моделей понимания-действия и ему можно навязать какую-то из имеющихся у него просто словесным внушением.
В момент просыпания или в момент засыпания тоже возникают состояния вне каких-то определенных моделей понимания-действия.
Соответственно, существуют несколько методов гипноза, использующие механизмы определенности личных моделей. Одним из методов являются погружение в сонное состояние и внушение на стадии утраты определенных моделей понимания-действия. Другим методом является внезапное выбивание человека из уже имеющегося контекста модели неожиданными, вводящими в растерянность действиями и словами.
Итак, поддерживаются активности тех контекстов понимания, к которым обращался канал осознанного внимания, образуют картину текущего субъективного восприятия - понимания и переживания происходящего – информационную картину субъективного самоосознания.
Именно явление субъективности самоощещения, доступное лишь изнутри при самонаблюдении, но ускользающее на уровне процессов мозга, порождает столько мистически непознаваемого в “трудной проблеме сознания”, которую озвучил Антти Ревонсуо в книге Психология сознания: fornit.ru/b9. Для очевидного понимания того, как возникает субъективное, недостаточно уровня организации элементов субъективного, описываемых в обзорной статье “Субъективизация ощущений и личность” (fornit.ru/1388) и еще: fornit.ru/50394 и fornit.ru/50497. В первую очередь нужно суметь зримо сопоставить то, как все это динамически реализуется в контекстах моделей понимания в виде меняющейся информационной картины, ясно видеть механизмы их активации, в контексте которых заготовлены мыслительные автоматизмы решения новых проблем понимания и целевого действия. И тогда всякому недопониманию и мистическим чувствам не останется места. Но это не просто, за пару раз не получится наработать навык такого охвата сразу многих сложных механизмов и их абстрактно выделяемого сознанием соответствия.
Сопоставление отдельных смыслов в общий смысл
Рассмотрим подробнее динамику процессов активации в контексте понимания смысла.
Стоит измениться, к примеру, месту событий, распознается и активируется другой контекст, после чего нужно уточнять всю информацию, что касается этого контекста. Это можно наблюдать, когда кошка просится выйти в другую комнату с закрытой дверью, хозяин открывает ей дверь, но кошка замирает в нерешительности, столкнувшись с другими звуками, запахами и зрительной картиной. У людей тоже бывает, что, войдя в другую комнату, они забывают, зачем сюда пошли, если мотивация была связана с действиями в предыдущей комнате. Вернувшись и поделав что-то с прерванного момента, вдруг вспоминается: ах да, мне же нужна соль. Теперь с этой удерживаемой мыслью можно более уверенно опять пойти в другую комнату.
Как удерживается мысль из предыдущего контекста? Можно измыслить достаточно простые и надежные схемы механизмов. Главное, что задача иметь такие механизмы необходима, иначе бы при каждой смене обстановки или ситуации у нас обнулялась бы цель действий. И экспериментально было обнаружено, что с развитием сложности организации мозга у разных видов животных возникает все более емкий стек такой памяти. У человека он достигает возможности хранить до 5-7 прерванных контекстов различных целевых действий (fornit.ru/2499).
В реализации Beast емкость такой памяти принята равной 10. Слишком большая емкость приводит к своим негативным последствиям, так что она должна оптимизироваться в зависимости от опыта решения задач различного вида.
Мы способны читать довольно сложные, навороченные фразы, которые объединяют совершенно разные составляющие смыслы происходящего в обобщающее понимание. И каждый раз, натыкаясь на новый контекст (а значит, срабатывает ориентировочный рефлекс, прерывая действие и мысли старого контекста), сознание откладывает в стек предыдущий и, если число фрагментов осмысления оказывается меньше 5-7, возникает общее понимание происходящего, а если нет, возникает недоумение в понимании причин и следствий (вспоминаем, что модели понимания отражают причины и следствия, значимые для личности, т.е. имеющие смысл). Приходится разбирать все предложение по частям, сначала осиливая смысл отдельных совместных составляющих, объединяя их уже в один смысл и уменьшая число требуемых прерываний стека памяти.
Чем больший опыт имеется в понимании данного вида фраз, тем меньше составляющих смысла они требуют для разбиения на части. Ребенок может путаться даже с одним словом, силясь понять его смысл из отдельных букв.
Если в обозримом пространстве удерживаемых активностей нет того, что приводит к однозначности смысла (чуть раньше мы говорили о поиске подходящего контекста гиппокампом и о механизме обобщения, интуиции и эвристики при таком уточнении смысла), а, значит, понимания происходящего (или услышанного, прочитанного, увиденного и т.п.), то нужный контекст не может быть активирован уверенно, без сомнений, оставляя лишь многозначительные догадки.
Возможно, принцип цепочки в 5-7 прерванных восприятий смысла – именно в сохранении такого количества самоудерживающихся активностей контекстов понимания, даже если это – очень далекие один от другого по смыслу контексты, что позволяет, сопоставив, связать одним смыслом самые разные вещи.
Ограничение в 5-7 удерживаемых для обобщения сопоставляемых образов возникает потому, что речь идет, все же, не о примитивной функции распознавания как у отдельного нейрона, а некоем универсальном алгоритме распознавания моделей понимания (смысла), реализованном механизмом второго уровня сознания, т.е. механизмом сопоставления значимости цепочек Правил с выбором наиболее подходящего.
Каждый из появляющихся сопоставляемых образов вызывает попытку распознавания модели понимания причин-следствий с каким-то результатом определенности понимания смысла (по базовой модели распознавания профилей возбуждения). Следующий образ придает новый смысл, который может дополнить предыдущий или аннулировать его ошибочное распознавание (видим, что была иллюзия понимания) и т.д. При этом нужно удерживать все участвующие активности в зоне профилей распознавания моделей, для чего нужны связи буквально со всеми возможными моделями в зоне предположительного распознавания (более общего контекста, который может быть расширен еще в более общую или смежную область (иначе “и в голову не приходило, что это может такое значить!”).
Если стек будет слишком мал, то не получится понимать передаваемый смысл сообщения, содержащего число сопоставляемых и известных причин-следствий больший, чем число звеньев такой памяти. Если он будет слишком большой, то к получающемуся смыслу начнет добавляться уже лишнее, что-то из следующего фокуса внимания. Когда передаваемое сообщение заканчивается, то становится понятно, что, наконец-то, все, больше в данный стек понимания не нужно больше ничего помещать, но это не всегда бывает столь ясно на слух, как при виде точки в предложении. В общем, возможно, число звеньев в стеке оптимизируется в зависимости от культурного правила строить сообщения не более разумной навороченности (если ты слишком прост, то вызываешь презрение, если слишком витиеват, то вызываешь раздраженное непонимание). Возможно, что, если достаточно рано погрузиться в среду заумных книг с длинными предложениями, разовьется и больший стек, даже больший, чем одно предложение, а сразу для смысла нескольких абзацев, страницы.
Можно представить, насколько легко при этом уйти в заумную шизу. Кстати, чем больше город, чем более интегрирована культура в данной местности (в том числе культура целой страны fornit.ru/19858), тем больше шиза там (fornit.ru/2433) – т.е. чаще проявляется психопатология личной интерпретации.
Тут стоит привести проясняющий пример.
Скажем, на уроке биологии учитель раскрывает тему про размножение и выдает фразу: “Размножение у людей состоит в передаче половой клетки от мужчины женщине и слиянии ее с половой клеткой мужчины”. Подхихикивание в классе обостряет интерес. Если это было новостью для кого-то, кто уже знает, что для размножения нужны мужчины и женщины, но еще не знает про слияние половых клеток, то у него возникает новая модель понимания причины и следствия: при слиянии половых клеток начинается размножение. Значимость для себя этого подогрета классом и имеющимися знаниями. Возникло обобщение уже известной модели про мужчин и женщин, и модели понятия клеток в новую модель причин и следствий: мужчины и женщины размножаются слиянием клеток. Эта модель будет укрепляться, дополняться и развиваться далее.
Другой пример.
Ребенку дали задание убрать лишнее из нескольких слов и перечисляют: “морковка, арбуз, хлеб, молоко, БМВ”. Каждое слово вызывает распознавание своей категории модели (у ненормальных все может оказаться в одной общей категории “материя”), выстраивается стек для обобщения. Самое важное для ребенка – БМВ, о котором он мечтает, все остальное у него есть без проблем. Модель БМВ доминирует с большим отрывом, понятно, что все остальное лишнее. Но умный мальчик соображает, что учитель имеет ввиду убрать только одно слово. Если еще не сформированы обобщающие категории из данных образов, то дополнительное условие все меняет и начинается перебор для поиска схожего у всех, кроме одного. Это – далеко не разовая автоматическая операция, а нерешенная проблема для 4-го уровня сознания.
Вот так может использоваться стек ограниченного числа звеньев удерживания моделей понимания для операций мыслительными автоматизмами. Т.е. такой стек – важный элемент для оперирования мыслительными автоматизмами, и он должен для этого адаптироваться.
То, насколько большие куски текста возможно поместить в стек памяти для обобщения, зависит от уже имеющихся заготовок моделей понимания, оперирующими уникальными образами (вопрос в том, если ли уже образ данного куска текста со привязанной значимостью и категорией).
Если тема не привычна и содержит новое, то для продвижения нужны промежуточные осмысления отдельных фрагментов с закреплением новых образов и новых моделей понимания. Иначе неизбежно будет упущено многое для понимания сказанного и не поможет даже сон.
Но если тема хорошо понятна и привычна, то можно схватывать сразу большие куски, в том числе и не привлекая осознанность для скорочтения за счет автоматизмов, хотя при таком произвольном подавлении ориентировочного рефлекса (те, кто занимается медитацией, легко учатся это делать) появляется риск не заметить важное новое.
При подражании используется этот же контекст понимания смысла: наблюдатель отслеживает последовательность отдельных действий учителя, понимая то, к чему они приводят и как он это делает. Если уже известна и понимается цепочка элементарных действий, то она воспринимается сразу вся, но если нет, приходится обращать внимание на отдельные ее детали. При этом находится соответствие между имеющимся распознавателями образа восприятия данного звена действия в теменной коре и имеющимися моторными действиями на стороне лобной коры. Это возможно только в рамках объема возможностей сопоставления 5-7 отдельных понимаемых фрагментов (при большем числе общий смысл не находится). Это образует обобщенный фрагмент нового навыка, который остается закрепить в попытках повторить эту цепочку. После чего в следующий раз уже не требуется детальное осмысление.
Такие автоматические навыки постоянно используются практически или их можно произвольно начать использовать для целей быстро оценить что-то без сильно затягивающего процесс внимательного просмотра. Скажем, вы засветили свой почтовый ящик в интернете так, что в спам попадает много писем, среди которых могут оказаться и вовсе не спамовые. Можно внимательно вычитывать заголовки, а можно подавить осознанное внимание и просто довольно быстро пройти взглядом весь список. На спорных письмах взгляд будет останавливаться.
Из общей системной логики модели вырисовывались основы таких состояний, как (не)уверенность (fornit.ru/379) и понимание (fornit.ru/1073). А отсюда естественно тянутся интерпретации механизмов творчества и движущая сила интеллекта – неудовлетворенность существующим (fornit.ru/475), не говоря про конкретизации философии смысла жизни (fornit.ru/59).
Буквально все начинает вставать на свои места.
Такая эквилибристика случается всегда при системных обобщениях и это – очень опасно в том смысле, что нужно иметь навыки чтобы четко следить за обоснованной корректностью системных обобщений и методом сопоставлений на основе формирования целостного каркаса системной модели. Если обобщения в самом деле системны, то в белах пятнах модели появляются предсказания новых элементов (как это было у Менделеева (fornit.ru/23625) и других авторов системных обобщений).
Но никто не безгрешен, ошибки всегда сопровождают любые субъективные предположения (fornit.ru/1012) и, тем более, никто себе (и другим) не судья, а поэтому любые даже кажущиеся очевидно системные модели, в конечном счете должны верифицироваться экспериментальной проверкой не только собственной, но и независимых исследователей. Предположительную часть МВАП можно назвать системной философией, но она во многом интерполяционна и в такой целостности вселяет уверенность своей очевидностью: fornit.ru/7117. И она уже уверенно подтверждена реализацией прототипа индивидуальной системы адаптивности Beast.
Практическая польза таких моделей – четкое понимание, какие именно исследования нужно проводить, что дает совершенно иное качество, чем метод научного тыка и бессистемная тематика. Здесь чрезвычайно важны основы естественнонаучного мировоззрения (fornit.ru/13268), которые закладываются в виде базовых моделей понимания так, что насколько адекватна реальности модель – настолько близки к реальному ее интерпретации, а погрешности такого уровня могут даже не замечаться в силу давно привычной адаптации к этой картине (раз удалось с ними мириться, не корректируя).
Другого пути и нет: понять организацию механизмов психики возможно только с помощью системной модели при условии удачного выбора точки отсчета (схемотехника и эволюционная адаптология). И для этого безальтернативным инструментом является имеющееся программное воплощение, на основе которой возможно проверять любые предположения. Никакие другие специализированные биологические, нейрофизиологические и т.п. подходы в попытках сопоставить все многообразие природной реализации и вывести нечто принципиально общее оказались практически несостоятельны (fornit.ru/19798).
Мы не будем сейчас углубляться во множество обнаруживаемых следствий сказанного, хотя это заслуживает большого отдельного рассмотрения.
Контексты личности
Мы рассматривали контекстную организацию моделей понимания, которая определяет смысл происходящего в условиях данного контекста. И в каждом таком активном контексте происходит закрепление цепочки эпизодической памяти, с помощью кадров которой мы легко можем воскресить данный контекст, если такой кадр был чем-то активирован, например, мыслительным автоматизмом, когда возникла задача вспомнить события вчерашнего дня. Мы вспоминаем, пробегая по вчерашней цепочке последовательности осознания с произвольной скоростью (как и по цепочке автоматизма). Если мы запоминали путь в лабиринте здания, то он останется в виде цепочки последовательности прохождения (эти следы назвали картой местности (fornit.ru/7057), но это – лишь частный случай любых цепочек эпизодической памяти). Мы легко определяем время до и после события по этим кадрам, вовлекая сопровождение мыслительных автоматизмов в активировшемя контексте понимания вчерашнего прохождения.
В более общем контексте “на прошлой неделе” (если мы такой контекст сформировали в силу его какой-то важности, а не просто контекст последних прожитых дней) есть доступ к набору цепочек эпизодической памяти за неделю. На самом деле жизненно более важна не хронология, а то, в каких состояниях мы пребываем, что определяет стиль поведения. А контекст “это было вчера”, “позавчера” и т.п. – дежурные всего несколько контекстов, в которых есть доступ для мыслей к последним связанным с ними цепочкам эпизодической памяти. Но если какой-то день окажется очень важным, то он может составить свое собственное ветвление в общем контексте “это было в прошлом”.
В отличие от стилей поведения, определяемых контекстами гомеостатической значимости, на стороне контекстов понимания существуют модели самого себя, которые основываются на базовых стилях поведения, но дополняются особенностями произвольной интерпретации смысла происходящего, формируя то, о чем говорят как о свойствах личности, проявляемых внешне и которые доступны самонаблюдению.
Мы уже упоминали об уровне моделей “кто я” (о самосознании поговорим позже). Они являются более общими в иерархии вложенности относительно моделей личности в различных условиях, отражающих гомеостатическую значимость в различные периоды развития организма (детство, зрелость), которые ограничивают восприятие и реагирование тем, что характерно для этих условий.
Теряя актуальность (когда условия перестают соответствовать тому, для чего формировалась данная модель “Я”), модели более раннего периода могут быть вновь актуализированы при некоторых условиях: игры, внушение тех признаков, которые характеры для таких моделей, повреждение альтернативных моделей данного уровня, что оставляет лишь одну (их локализации различаются).
Кроме периодов развития могут быть и другие общие факторы, провоцирующие формирование альтернативных моделей личности: переезд в другую страну с другой культурой на постоянное жительство, наркомания или пьянство и все, что с этим связано, периоды рецидивов болезни, ситуации, провоцирующие доминирование первичных стилей поведения (страх, ярость, половое, пищевое поведение и т.п.). Все эти контексты могут кардинально, до противоположности, менять интерпретацию происходящего, смысл происходящего и доступные наборы вариантов реагирования в таком смысле.
С помощью авторитарного внушения (гипноз) удается заставить активировать ту или иную модель личности, в том числе и совершенно экзотические, например, заставив человека ощущать себя мудрым камнем у дороги или вообще не существующим объектом вроде дракона или бога, если только такая модель представлений существует в его представлении.
В книге Вилейанур С. Рамачандран Рождение разума (fornit.ru/1257):
“… даже несмотря на собственную принадлежность «Я» — по самому его определению, - оно в огромной степени расширено за счет социальных взаимодействий и, безусловно, может эволюционировать уже в социальном контексте.
Первыми на это указали Ник Хамфри и Хорас Барлоу на конференции, которую организовали Брайан Джоузефсон и я в 1979 году. Позвольте мне развить эту мысль. Наш мозг в целом представляет собой моделирующее устройство, необходимо создавать рабочие виртуальные имитации реального мира, в соответствии с которыми мы можем действовать. Внутри имитаций нам также нужно создавать модели разума других людей, поскольку мы, будучи приматами, чрезвычайно социальные существа. (Это положение называется «теория другого разума»).) Надо делать это таким образом, чтобы иметь возможность предвидеть их поведение. Например, вам нужно понять, был ли укол зонтиком чьим-то злым умыслом, а значит, он может повториться, или это была случайность — тогда инцидент исчерпан. Более того, чтобы эта внутренняя имитация была законченной, она должна содержать не только модели разума других людей, но также и модель саму по себе, то есть ее постоянные атрибуты — что она может и не может делать. Вполне вероятно, что одна из этих способностей моделирования эволюционировала первой, а затем подготовила почву для второй. Или — как это часто происходит в эволюции — обе способности развивались совместно, обогащая друг друга, достигая вершины самосознания, которое характеризует Ното Зйгдепз.
На самом рудиментарном уровне мы уже видим наличие этого взаимодействия «Я» и «других» всякий раз, когда новорожденный младенец имитирует поведение взрослых. Высуньте язык перед новорожденным ребенком, и он высунет язык вам в ответ, трогательным образом размывам границы (условные барьеры) между «Я» и другими. Чтобы сделать это, он должен создать внутреннюю модель вашего действия, а затем «разыграть» ее в своем собственном мозгу. Удивительная способность, учитывая, что младенец даже не может видеть свой собственный язык, а значит, должен искать соответствие зрительному образу в ощущении его положения в пространстве.
Мы знаем, что этот процесс производится специальной группой нейронов в лобных долях, называемых зеркальными нейронами. Я предполагаю, что эти нейроны хотя бы частично вовлечены в формирование «материального воплощения» самосознания, а также нашей способности «сопереживать» другим. Неудивительно, что дети, страдающие аутизмом (которые, по моим предположениям, имеют дефектную систему зеркальных нейронов}, неспособны воссоздавать «теорию другого разума», не могут сопереживать другим, а также участвовать в самостимуляции, чтобы усилить свое восприятие собственного «Я», воплощенного в теле”.
Существуют расстройства в организации психики, при которых оказывается невозможным распознать подходящую модель самого себя или, наоборот, активируются одновременно более одной такой модели: fornit.ru/1230.
При активной модели понимания (со всеми ее более общими составляющими), если действие не происходит из-за того, что реакция не подтверждена на выполнение первыми уровнями сознания, возникает состояние поиска решения целевой проблемы, для чего подключаются все более высокие уровни сознания (блок-схема алгоритма из прошлой главы). Это означает некоторую самодостаточность поддержания текущего контекста понимания на время решения проблемы и ее доминирование по активности в конкуренции с другими. Причем, важность проблемы, которая представлена произвольностью оценки ее значимости (т.е. пониманием важности смысла для поставленной цели), конкурирует с важностью других возможных целей, больших и малых. А любые признаки в области образного восприятия или абстрактного мышления, привлекающие осознанное внимание на себя, которые связаны с теми или иными элементами контекста проблемы (тем самым обладающие своей какой-то новизной), опять вызывают активацию контекста проблемы, но уже дополняя отклик ориентировочного рефлекса своей произвольной значимостью (которую возможно усилить или понизить).
Поэтому, если у нас возникала насущная проблема, которую не удается решить сразу, то ее контекст оказывается или постоянно активным, лишь допуская временные переключения на другие в соответствии с удержанием в памяти отложенных проблем (5-7 штук) или, если он был чем-то дезактивирован (например, глубоким сном или еще более насущной проблемой), то легко восстанавливает свою активность и все мыслительные автоматизмы в ее контексте понимания.
А.Ухтомский предложил называть такое явление доминантой (fornit.ru/5157) и посвятил ее исследованию много времени. По его мнению условные стимулы, которые активируют контекст нерешенной проблемы, могут укреплять связи по этому пути по механизму условного рефлекса (fornit.ru/5359), становясь причиной быстрого переключения в данный контекст, во многих случаях вызывая патологические состояния зависимости: fornit.ru/422. Мало того, такие связи укрепляются и по механизму осознанного формирования автоматизмов с участием уже произвольной оценки значимости и волевого усилия.
Мы будем называть это явление доминантой нерешенной проблемы, которая сопровождает активацию четвертого уровня сознания. Инициируясь ориентировочным рефлексом при очень большой значимости, она требует нахождения решения, которое может многократно откладываться в силу неудачных попыток решить. Но высокая значимость, связанная с эпизодом актуальности проблемы, делает ее легко опять актирующийся малейшими ассоциациями и просто текущей необходимостью.
Пора заметить, насколько много самых разных проявлений психики, которые интриговали своих исследователей и рассматривались обособлено (особенно в практике эмпирической психологии), оказываются настолько тесно взаимосвязаны в общей модели организации адаптивного отклика на новые и значимые стимулы – в общей модели адаптивной функциональности сознания.
Так, термин “Гештальт” наивно и вульгарно описывает проявление доминанты нерешенной проблемы, и эта проблема - вовсе не обязательно - гомеостатическая потребность.
Возможно, что те, кто (каким-то чудом) удерживают общую картину описания, испытывают специфическое ощущение гармонии понимания этих взаимосвязей, скрепленных в каркасе эмпирических данных, что придает ясную и очевидную уверенность понимания и видения взаимодействий описанных механизмов. Но это во многом слишком оптимистично.
Очень хочется тут же начать раскрывать все следствия этих основных механизмов, например, суть той же очевидности (fornit.ru/7117), при ее окончательной важности для субъективного убеждения, но это бы сделало книгу совершенно неподъемной так же, как попытки приводить в каждом случае варианты возможной схемотехнической реализации. Поэтому приходится подавлять такие порывы в пользу более общей определенности в оптимуме лаконичности, не требующей слишком большого объема памяти отложенных проблем, которая и без того наверняка постоянно переполняется, требуя возвратов к уже прочитанному для удержания смысла сказанного.
Но в случае продолжения заинтересованности, можно перейти к постепенному рассмотрению множества психических проявлений: fornit.ru/1181 рассмотренных механизмов и, главное, изучить реализацию Beast (fornit.ru/64975).
Про самосознание
Привлечение осознанного внимания к модели Правил и Значимости самого себя приводит к проявлению эффекта самосознания – при осмыслении новизны, возникшей в связи с этим, что, в принципе, не отличается от процесса осмысления любых других моделей разных объектов внимания (если внушением или галлюциногенным воздействием придать модели “Большого камня у дороги” статус “Я”, то возникнет самосознания себя с известными качествами данного камня.).
Самосознание — это не внимание к каким-то отдельным деталям своего тела или даже к мыслям, а именно внимание к свойствам самого себя в общем контексте “я существую и управляю собой”, отраженным в модели понимания смысла происходящего. Это, прежде всего, активация модели свойств собственного Я со стороны отдельных целеобразующих моментов подключения ориентировочным рефлексом конкретных задач и активацией имеющихся автоматизмов мышления. Не более того - в сопоставлении с такой же активацией других проблемных контекстов. Так что какого-то особого внимания к рассмотрению явления самосознания не требуется вот почему: fornit.ru/1277.
Наличие самосознания не привносит какое-то особое качество сознательной деятельности и вообще любой из функции уровней сознания. Мы очень редко занимаемся самосознанием и есть люди, которые вообще без этого прекрасно обходятся, непосредственно переживая события, хотя есть и период раннего самоисследования, когда самокопание бывает актуально наряду с другими философскими проблемами: fornit.ru/7062.
Понимание сути самоощущения и его проявляемых эффектов (как субъективных, так и объективно наблюдаемых) очень непросто и требует серьезного погружения в эту тематику. Поэтому предлагается ознакомиться с материалами на эту тему разного уровня приближения:
· Суть самоощущения: fornit.ru/50394" target="_blank">fornit.ru/50394
· Что такое сознание: fornit.ru/50498" target="_blank">fornit.ru/50498
· Одушевление: fornit.ru/50497" target="_blank">fornit.ru/50497
И затем сопоставить полученное представление с уровнем, достигнутым современной академической наукой на данное время:
· Комментарии к книге Дэвид Иглмен Мозг: «Ваша личная история»: fornit.ru/50204" target="_blank">fornit.ru/50204
· Комментарии к статье К.В.Анохина: «Когнитом: в поисках фундаментальной нейронаучной теории сознания»: fornit.ru/50322" target="_blank">fornit.ru/50322
Произвольность порождается ориентировочным рефлексом
Осмыслим еще раз понятие произвольности в контексте всего уже воспринятого.
Мыслительные автоматизмы системы произвольности способны управлять активностью контекстов значимости, расширяя и сужая их для обобщений и конкретизаций. Они способны прогонять цепочки реакций в составе Правил, чтобы подсмотреть конечный результат и даже способны активировать эти цепочки по отдельным звеньям, чтобы контролировать действие во всех его фазах и корректировать ошибки.
Но, при всем при этом создание нового кадра субъективного переживания в эпизодической памяти – всегда автоматический процесс, даже когда это происходит под воздействием произвольности фокусировки внимания (fornit.ru/24208). Это – медицинский факт (Ильф и Петров ©), который используют фокусники и мошенники, про который сказано в одной из историй о Ходже Насреддине (история про белую обезьяну: fornit.ru/23624). Как именно это организовано - легко представить принцип схемотехнической реализации, а чтобы определить, как именно проделала это природа в каждом виде живых существ - еще не дошли руки у исследователей.
Поэтому под произвольностью не следует понимать ни сумасбродство, ни какие-то другие атрибуты “свободы воли”, а – лишь то, что система приспособления к новому противопоставляет привычному, если оказывается, что реагировать по-старому в условиях, где появилась значимая новизна, не следует. Все. Произвольностью можно считать альтернативное название для слова “сознание” потому, что именно сознание и занимается такой работой: выяснят, можно ли расслабиться и поступать как обычно или стоп! тут нужно сработать по-другому или вообще задуматься над поиском решения, а как лучше. Произвольность тем более проявляется, чем более высокий уровень выбора реагирования нужен и на четвертом уровне вовлеченности механизмов сознания ее качественно больше. А каждый акт осознания инициируется ориентировочным рефлексом.
Не забываем, что все нерешенные проблемы, которыми занят наш мозг, прямо или косвенно, вплоть до проблем Единой Теории Фундаментальных Взаимодействий или даже связи полярного сияния с онанизмом в Средней Азии (fornit.ru/6468), являются следствием значимой новизны для нас этой темы, которая не нашла ответа в заготовках первых уровней сознания (готовых автоматизмов ответных реакций), но важность ее смысла не покидает нас, заставляя возвращаться к проблеме снова и снова. А проблемой становится наиболее значимое новое (ориентировочный рефлекс), которое появляется в привычном (иначе совершенно новое вообще не воспринимается). Так что четыре уровня сознания – это четыре уровня новых проблем, которые решаются механизмами произвольности в отношении привычного (на первом уровне произвольность почти нулевая, но уже есть намек на то, что мы готовы скорректировать действие в нужный момент).
В области абстрактных образов понимания смысла с наборами мыслительных автоматизмов префронтальной лобной коры есть свои детекторы нового (fornit.ru/24208), и они вызывают ориентировочный рефлекс к тем абстрактным образам, которые входят в состав цепочек мыслительных автоматизмов (так же, как подключается образ из теменной коры в цепочке моторного). И тогда такая абстракция начинает осознаваться, она вдруг появляется в области субъективного как новая мысль: возникает новый образ для сопоставлений в цепочке допустимого числа таких образов (и это фиксируется как кадр эпизодической памяти). Т.е. это уже не тот привычно автоматический образ, который, не требуя осознания, производит какие-то переключения внимания и управления произвольностью, а - в составе новой обобщающей ассоциации, которая при осмыслении придает новый смысл. Это – достаточно частое, повседневное явление, но только в наиболее ярком своем проявлении, когда в результате находится решение доминирующей проблемы, это воспринимается как из ниоткуда (из тишины мыслей) появившееся в сознании решение проблемы – как интуиция: fornit.ru/1392.
Новые идеи появляются в результате учета воспринятого в контексте нерешенной проблемы, под влиянием нового впечатления восприятия или эпизодической памяти. Нет новых впечатлений - нет новых идей. При этом впечатления могут инициировать новые направления мыслительного реагирования даже если это не образы восприятия, а образы эпизодической памяти, т.е. остатки бывшей мыслительной деятельности, но в новом контексте проблемы.
Внешне продуцированные образы восприятия и элементы эпизодической памяти это - то, что придает мышлению не алгоритмическую автоматичность (и сам цикл мышления – как рекурсивная итерация функции осмысления никак не алгоритмичен), а непредсказуемое направление мысли в контексте цели решаемой проблемы. Понятно, что это имеет адаптирующее, а не сумасбродное значение: “Свобода – есть осознанная необходимость”. Вот почему мы – не роботы при всех наших автоматизмах: fornit.ru/514.
Роботы (в классическом представлении) действуют в зависимости от внешне различаемых ими параметров окружения в соответствии с программами, даже с программами, способными создавать новые программы для каких-то заранее определенных ситуаций (в лучшем случае - условные рефлексы с участием детекторов нового). Но они не могут выделять новое в активном контексте личной значимости (потому, что в них отсутствует система гомеостаза, нет жизненных параметров, обслуживание которых определяет все остальное), которое прерывает текущую программу и вместо нее запускает программы обработки нового параметра, использующие иерархию уровней сложности такой обработки в зависимости от успешности нахождения варианта дальнейшего реагирования вплоть до уровня творчества новых вариантов реагирования, которые используют появляющиеся от различных впечатлений идеи в механизмах интуиции. Иначе это будут уже не роботы, а произвольно адаптирующиеся живые особи.
Таким образом, в результате мыслительной обработки нового в контексте актуальной проблемы могут появляться совершенно новые варианты реагирования и совершенно новые мыслительные образы, которым нет соответствия в объективной реальности.
Осознанное использование интуитивного метода нахождения подходящих вариантов действий называется эвристикой: fornit.ru/1647. Можно натренировать навык задавать задачу своему подсознанию и не думать о ней специально, пытаясь решить “логически”. Важно убедительно осознать важность и необходимость решения, можно еще научиться задавать желаемое время получения ответа, с учетом того, что для бессознательного решения нужны новые ассоциации, т.е. достаточное богатство восприятия и мыслей, а на это нужно время. Поэтому задавать срок нужно достаточно отдаленный, может быть не один раз подступая к решению, и в это время заниматься чем-то близким к отложенной задаче, думать о сходном, читать о попытках решений других людей. Если решение будет найдено, оно само привлечет к себе осознанное внимание. И тут важно вовремя зафиксировать его, не рискуя отвлечься на что-то и забыть, даже если это случится ночью или при занятии любовью.
Дополнительно стоит ознакомиться со статьей “Понимание произвольности”: fornit.ru/12787.
И затем - самое время в контексте освоенного критически прочесть книгу А.Ревонсуо “Психология сознания”: fornit.ru/b9.
Про сознание написана книга "Субъективность" (fornit.ru/68364), которая будет поддерживаться в актуальном состоянии, так что там есть много уточнений и нового.
Функциональность сна
Сон – один из базовых стилей поведения, который у разных видов животных сильно различается по разнообразию и особенностям функциональности. Не всегда бывает даже, казалось бы, самое характерное: во сне минимизируются рецепторное восприятие и мышечные реакции. Сон обычно не затрагивает полностью весь мозг (если это не сезонная гибернация), оставляя распознавание “сторожевых” признаков с их рефлексами, т.е. он остается управляем внутримозговыми процессами.
Сон у разных видов животных
У разных видов животных (и, особенно, в зависимости от сложности нервной системы) сон организуется очень по-разному и функции сна достаточно различаются, хотя в одном есть общее: понижение тонуса активности мозга (всего или отдельных его половинок или отдельных зон), чередующееся периодами активности, возникающими вследствие нарастания контрастирования, что приводит к сновидениям, которые, конечно же, эволюционно не были упущены напрасно для адаптивности.
Без сна накапливающиеся суточные активности в мозге (самоподдерживающие и не гасящиеся более слабыми соседними) все в большей степени начинают мешать новым актуальностям реагирования, вызывая трудности выделения отдельных реакций и все большее ухудшение гомеостатической регуляции вплоть до полной разрегуляции и смерти (fornit.ru/5395) - при отсутствии каких-то конкретных органических повреждений, могущих быть причиной смерти. Все ткани и органы оказываются вне регуляции прямо или косвенно осуществляемой мозгом, его структурами, ответственными за поддержание в норме параметров жизнеобеспечения. Можно сказать, что в дополнение к повсеместному латеральному торможению организуется система принудительного торможения активностей.
Сон – очень древнее приобретение. В развитии организма прослеживаются этапы формирования сна у предков так, что у простых животных сон очень схож со сном младенцев, а периоды активности во сне появляются только у тех видов, у которых есть гиппокамповый канал связи с лобными долями (иначе нет субъективных переживаний и сновидений, в том числе). Но даже мухи устают от общения (fornit.ru/5207), нематоды во сне перестраивают свою нервную систему (fornit.ru/2530), а муравьи тоже видят сны (fornit.ru/5363), т.е. у высших насекомых сон уже принимает участие в корректировке нервной регуляции.
У морских животных с легкими и перелетных птиц, которые должны постоянно бодрствовать, сном заняты поочередно разные половинки мозга, потому как они не могут терять на время сна активность восприятия и действия. Они не могут и обходиться вообще без сна.
Понятно, что прежде всего стадия сна адаптируется к суточным ритмам активности (fornit.ru/5383), чтобы максимально оптимизировать период бездействия и бодрствования в связи с окружающими условиями. Так, у северных животных (fornit.ru/5404) в условиях полярного дня и ночи возникают не связанные со светом и темнотой периоды активности и сна.
Многие животные засыпают зимой просто потому, что не смогли бы прожить иначе в режиме полноценной активности. Такая длительная пассивность нисколько не вредит в информационном плане, а вот более высокоразвитых видов животных, которые бы не спали очень долго, не существует. Т.е. сон ко всему прочему – хороший способ сохраниться на время пассивности, минимизируя затраты в течение сезона или в течение суток. Мало того, сон – очень выгоден для регенерации и развития тела животного, так что во время сна специально вырабатываются гормоны (например, гормон роста), которые активируют этот процесс для условий, когда ничего ему не мешает.
В книге Эволюция сна от простых форм до млекопитающих О.И. Лямина (fornit.ru/b10) можно ознакомиться с обобщающим обзором особенностей проявления сна у разных животных.
Эмпирические данные о сне
Как обычно в природе, для каждого состояния организма эволюционно оптимизируются все сопутствующие функции и поэтому возникают самые разные использования состояния сна, причем у разных видов животных они основываются на особенностях их жизненного окружения. В результате все функции, которые можно выделить во время сна, отличаются большим разнообразием. Мы же будем выделять последствия развития наиболее общего, неспецифического торможения в областях коры, где могут сохраняться активности при пропадании внешних стимулов: третичную теменную и третичную лобную кору. Тем самым мы отсекаем от рассмотрения все более низкоорганизованные виды животных, у которых сон кардинально отличается и невозможно представить общую системную модель. Такой примитивный сон соответствует возрасту младенцев после 1-3 месяцев (см. про младенческий сон: fornit.ru/23627).
В детском возрасте потребность сна выше, он продолжителен и част даже в дневное время. Мозг в этот период развивается во много раз более интенсивно потому, что более сложные образы в восприятии случаются гораздо реже, чем более простые, из-за чего слои более сложных образов должны иметь более продолжительные критические периоды формирования.
Очень важно своевременно освобождать мозг как от импульсной активности, так и метаболических следов, чтобы минимизировать образование ложных связей (fornit.ru/7371), не соответствующих реальности. После каждой процедуры общего гашения новые стимулы будут в большей степени совпадать со своими предшествующими следами и лучше закрепляться, так что чем больше таких циклов, тем более ясны и четки детекторы образов. Никакие изверги не посмели проводить опыты по депривации сна у младенцев, но последствия представить несложно: fornit.ru/5419. И это – еще до стадии развития канала осознанного внимания, после которой все становится намного сложнее и интереснее.
Итак, сон у человека начинается с развития общего торможения (fornit.ru/5159), постепенно усиливающегося в третичных зонах (fornit.ru/6330), изолирующихся от восприятия и действий. Если какие-то нерешенные дневные проблемы (fornit.ru/7531), остающиеся в виде очень актуальных переживаний, конкурируют с таким процессом, не давая ему развиться, то засыпание затруднено. Но чем больше скопилось активностей, тем в большей степени они будут мешать одна другой, затрудняя осознанное внимание высокого уровня и, тем самым, снижая конкуренцию засыпанию. Снижение уровня сознания усиливает запуск общего торможения, если только что-то внешнее или очень высокое напряжение волевого усилия не будут этому препятствовать.
При общем торможении в первую очередь угасают наиболее слабые и случайные активности, снимая латеральное торможение для соседних образов, активность которых станет более устойчивой и способной преодолеть связи между последующими зависимыми образами в цепочке их очередности возникновения. Оставшиеся активности оказываются изолированными (fornit.ru/5358) и контрастными, т.е. выделяются наиболее актуальные (fornit.ru/5352) из активировавшихся дневных образов, которые во сне особенно интенсивно поддерживаются гиппокампом (fornit.ru/5168).
Это - очень уверенная логика функциональности сна, без каких-то неопределенностей, с закономерными причинами и следствиями.
Далее – уже более предположительно, но также вполне закономерно. Стоит контрастные активности чуть спровоцировать и пойдет реагирование по ранее проторенным цепочкам, вызывая подергивание даже отключенных от реагирования мышц, а в области мыслительных автоматизмов вызываются мыслительные реакции по следам эпизодической памяти (т.е. цепочке последовательностей подключения осознанного внимания).
И в мозге, конечно же совершенно “тупо” и случайно, среди всех экспериментов природы, возникла система такой провоцирующей активности (fornit.ru/6472), которая оказалась очень полезной. Без нее бы все равно при некотором перевозбуждении образов полезли бы галлюцинации, но важно согласовать их появление с подготовкой к этому остальных зон мозга, скоро станет понятно почему.
Обилие ссылок оправдано тем, что дает опору на фактические данные в области, где до сих пор академическими учеными высказываются самые фантастические и нелепые предположения, как, например, “висцеральная теория сна И.Пигарева”: fornit.ru/1724. Так что эти ссылки надежно определяют каркас понимания явления сна. Теперь остается согласовать его с каркасом модели произвольности.
Сон и память
Многие теоретики утверждают, что долговременная память о событиях, имеющая информационное значение, формируется только во время сна, когда в гиппокампе в ускоренном режиме прокручиваются дневные воспоминания. Это – очень небольшая и далеко не корректная часть правды. Но есть исследования (fornit.ru/5334), дающие очень хорошие подсказки. Академические представления о сне можно представить по книге Вейн Александр Моисеевич: «Сон - тайны и парадоксы»: fornit.ru/217.
Далее – обобщающие выводы о влиянии функции сна на психику.
В области произвольности мы рассматривали структуру алгоритма функциональности уровней сознания в зависимости от качества сложности новой проблемы осмысления. Сейчас важны два варианта развития событий дневных переживаний: когда 1) есть время на осмысление (необязательно действовать немедленно) и 2) нет времени, нужно опрометчиво рисковать и действовать. Во втором варианте так же ситуация ветвится на то, что повезло и все получилось хорошо, и то, что не повезло и тогда проблема еще больше нарастает, а на ее решение может быть или не быть время, т.е. – те же два варианта.
В случаях, когда нет времени осмыслить новое, для адаптивности остаются только механизмы автоматизмов, если только снова и снова рисковать наступать на грабли, т.е. и с привычным реагированием напряженка, ведь бывают настолько безнадежные и не позволяющие осмысливать ситуации, что сознание вообще отказывает и автоматизмы не корректируются, а остаются только механизмы рефлекса.
Если нет времени осмысливать, пройдет время, все уйдет безвозвратно и бесследно, как будто и не было никакого опыта в жизни, и опять в похожих ситуациях оставалось бы выбирать с нуля. А в нашей жизни, стоит оглянуться, почти и не бывает времени подумать, все приходится делать сходу, а если “затупил”, то оказываешься вне игры. Задумчивых не очень любят, это чуть ли не признак отсутствия интеллекта, другое дело те красавчики, кто за словом в карман не лезут. Чаще всего ценятся те, кто действует быстро и выверенно, т.е. уже имеющие достаточный набор автоматизмов. Даже когда отпустит и можно подумать на диване, у многих нет такой привычки или нет достаточной актуальности ворошить прошлое. Но в голове возникшие за день самые значимые активности остаются в виде самоподдерживающихся активностей – по каждому акту ориентировочного рефлекса.
И вот, человек засыпает, главное в дневных переживаниях выделяется, срабатывает все тот же ориентировочный рефлекс и включаются циклы осмысления (fornit.ru/5376), как если бы мы задумались о случившемся. Для этого используются начальные уровни сознания вплоть до третьего, т.е. без выраженного волевого уровня. И мы зримо переживаем возможные варианты развития событий в ускоренном масштабе времени, потому как не привязаны ни к пусковым стимулам, ни ко времени реагирования: цепочки развертываются быстро (~ в 7 раз быстрее, чем реальное восприятие), сразу до конца, оттуда провоцируются зависимые цепочки (в стеке переживаний предыдущее задает контекст последующему, формируя более общий смысл) и так далее, пока сновидение не исчерпает актуальность таких переходов.
С одной стороны добавляются новые актуальные активности и мозг проявляет повышенный тонус, а с другой - мы получаем осмысленный опыт происходящего (fornit.ru/5242) – все те же Правила, с вероятными прогнозами (настолько вероятными, насколько наш прежний, уже проверенный опыт это позволяет). И такой опыт максимально безошибочен потому, как привычные оценки – самые верные, еще не тронутые субъективно продуцированными предположениями волевого (четвертого) уровня осмысления.
После такого процесса безвольного осмысления с формированием общего смысла переживаний и возможных реакций, нужно замести следы от новых активностей и повторяется фаза общего торможения. Таких последовательностей торможения и сновидений в течении ночи у человека в норме бывает несколько (fornit.ru/5212).
Таким образом, отложенное пережитое не теряется напрасно, а эффективно готовит к повтору такой ситуации уже имеющимися готовыми прогнозами.
Вот как этот момент популяризован в статье “Основы понимания психики” (fornit.ru/973):
“Эта главная мысль была особенно безжалостно прервана встречей с собакой и при этом благостное ощущение скорого возвращения домой и удовольствия от телепередачи резко переменилась на парализующий ужас от неподвластной и непонятной ситуации, - совсем иной контекст поведения. Но если бы у вас был большой опыт по отбиванию от внезапных врагов голыми руками, то, несомненно, вместо парализующего от беспомощности ужаса возникла бы эйфорическая ярость предвкушения битвы. Эти такие разные состояния, которые называют эмоциями, фактически определяют ваш стиль поведения в каждой конкретной ситуации.
... Ночью вы заснули как младенец с отблеском той улыбки, вспоминая, как порадовали приятеля по телефону после передачи. Проснулись после взбудоражившего сновидения и поэтому ярко запомнили его (иначе разбудившая мысль не отложилась бы в памяти и все недостаточно значимые находки остались бы в небытии): будто вы прогуливались вечером с очень желанной незнакомкой в саду, было так приятно, как вдруг из кустов как мрачная тень, с волной адского ужаса неизбежной расплаты появился силуэт жуткого зверя со светящимися глазами. Но потом он съежился и оказался просто забавным щенком, так что вы даже не успели показать свой позорный страх девушке. Вам повезло и вы, как бы наблюдая со стороны, вдруг осознали, что не стоило так сразу паниковать, хотя вам раньше это было свойственно. Проснувшись от такой важной для вас при всей очевидности мысли, вы поняли, что эпизод с собакой тоже где-то крутился в голове все то время, что вы провели за телевизором и разговором с приятелем, пока не вылился в сновидение, где для вас стало очевидным новое понимание”.
Активности дневных значимостей нового проявляются в совсем другом контексте базового поведения – сна, и поэтому вызывают ассоциации сюжетов сновидения, которые бывает трудно сопоставить с дневными переживаниями. Но самое главное – сама значимость нового образа восприятия – возможного действия, которое и должно спрогнозировать то, чем оно может закончиться. А какими будут в реальности обстоятельства, где окажется такая же значимость, уже не так важно, в любом случае она новая и пока нет привязки к конкретным обстоятельствам. Банальный пример: выражение лица, виденное днем, поразившее новым неприятным впечатлением, сулящее что-то нехорошее так, что хочется ударить или убежать спасаясь. Чем все может закончиться?
Что интересно, в сюжете последовательного развертывания цепочек могут добавляться новые элементы, ассоциируясь с подчас очень далекими другими образами дневных событий (fornit.ru/5251) потому, что контекст сна, отменяя другие контексты, в которых еще не было подобных пересечений, позволяет взаимодействовать имеющимся активностям так, как если бы они были в общем контексте осмысления (стеке памяти последовательности переживаний), т.е. происходит обобщение для всех более частных контекстов. Возникает рефлекс переключения осознанного внимания к новым обстоятельствам, и сюжет получает новое направление.
Это уже достаточно сложно, чтобы продолжать развивать это направление понимания функции сна, так что стоит остановиться на уже имеющейся достаточно очевидной картине информационной роли сна и сновидений.
Конечно же, это состояние сновиденческого осмысления реализуется в его оптимальном варианте взаимодействия всех участвующих зон, поэтому и нужна такая синхронизирующе-организующая система провоцирования сновидений, о которой упоминалось.
Детали сновидений, из-за их максимальной актуальности для данного человека, обычно производят настолько завораживающее впечатление на переживающего их, что очень хочется поделиться этим с кем-нибудь еще, но при попытке пересказа с досадой обнаруживается, что это совсем неинтересно собеседнику. В самом деле, все эти детали были высшими по важности только для себя в тех обстоятельствах сюжета, что вынесли их среди всего остального. Поэтому по рассказу о сновидении можно определить, что именно было самым важным для этого человека в момент его переживания и как-то это использовать. И эта информация отличается от того, что хотел извлечь из сновидений З.Фрейд и, тем более, того, что видел в этом К.Юнг.
Вот смело выставленная на публику коллекция снов женщины: “Коллекция снов Евгении Коржуновой” (fornit.ru/24204) с ясно выделяемыми значимыми для нее фрагментами.
Яндекс провел исследование и опубликовал статистику сновидений (fornit.ru/24203), выяснив, что: “Самые распространённые сновидения не зависят от места жительства. Тем не ;менее в каждом регионе есть свои характерные сны”.
Информационная функция сновидений
После прокручивания сюжета сновидения новизна образа утрачивается, он становится более определенным по смыслу (пониманию его возможных причин- следствий) и впредь не представляет настолько актуальной проблемы, чтобы выйти в топ среди других проблем.
Как уже говорилось, во сне реализуется альтернатива осмысливанию при бодрствовании, т.е. наделение образов значимостью в циклах мышления, обновляющих информацию. Когда читаем книгу, обычно мы не отвлекаемся на ее осмысливание, чтобы не потерять нить повествования, а все, что нас зацепит, остается в виде актуальных активностей, которые нельзя терять, иначе зачем мы вообще читали книгу (или когда нам что-то долго объясняют и т.п.). Вот почему после прочтения трудного текста очень желателен сон, что оптимально сохранит полученную информацию.
Многие замечали, что заучивание на ночь качественно облегчает не просто запоминание заученного, но и понимание его смысла. Этот эффект регистрируется даже у птиц: fornit.ru/5335, fornit.ru/5219, fornit.ru/5350, fornit.ru/5352.
В осмысливании во время бодрствования неадекватность реальности результата выше потому, что включен режим произвольности, с волевым изменением сюжета (домысливание), а во сне произвольность ограничена, и оценка оказывается более “честной”, а не надуманной. Чем больше домысливания по сравнению с использованием уже проверенных данных (привычных автоматизмов), тем больше ошибок (fornit.ru/1012) предположений возникнет и закрепляется в памяти. А любое субъективное предположение должно проверяться в реальности, чтобы появилась адекватная оценка и уверенность очевидности (fornit.ru/7117). Так что явно у нас есть какое-то ограничение склонности фантазировать наяву, которое может довести до серьезных психопатологий (fornit.ru/1318). Это ограничение легко преодолевается многими людьми, особенно в молодости.
В норме сны не запоминаются и поэтому не продолжается осмысление с произвольным дополнением информации, что для наиболее значащих сюжетов, порождало бы фантазирование вплоть до паранойи. Но сны с полученными Правила остаются в эпизодической памяти, кадры которой оказываются труднодоступными при бодрствовании. Сон можно вспомнить, ухватившись за его фрагмент, но это трудно.
И здесь возникает противоречие с функциональностью четвертого волевого уровня сознания, которое привносит волевые усилия для реализации решений. Т.е. понимание возникает до использования четвертого уровня, но результат проверки верности предположений получается на волевом уровне, когда нужна определенная воля для учета этого результата, как бы ни была дорога идея.
Это верно как для научного творчества, так и реалистичности художественного творчества. Но совсем другое дело творчество, намеренно искажающее реальность и провоцирующее совершенно неожиданные предположения. Они заведомо воспринимаются с большим интересом из-за своей значимой новизны, но как мутации очень редко оказываются полезными, так же и из “психоделических” идей (fornit.ru/1187) редко возникает что-то практически полезное. Они довольно быстро приедаются до банальности потому, что мир субъективных фантазий (и даже компьютерного моделирования) несравнимо беднее мира объективных явлений, и идеи лучше черпать из реального мира, чем и пользовался Леонардо Да Винчи, разглядывая узоры штукатурки, наводящие его на мысли заданной темы.
Безволие сновидений не нравится людям, желающим получать полноценные приключения во сне и управлять ситуацией, где легко можно достичь всего желаемого. И когда такие люди узнают, что можно исказить функциональность сна и повысить активность произвольности до творческого 4-го уровня, они начинают практиковать это, пока не замечают нежелательные последствия такой искусственности и лишения себя нормального сна, или же, не замечая ничего недопустимого, продолжают предаваться этому, искажая свои модели понимания, в точности как в наркотических трипах. Эти извращения сна и их последствия описаны в статье Осознанные сновидения: fornit.ru/984. Понятно, что если увлекаться безудержными фантазиями без сурового отрезвления реальностью, то легко можно дойти до совершенно неземного состояния Офелии из Гамлета.
Сюжеты сновидений запоминаются в контексте сна и поэтому оказываются недоступны в других контекстах, но конкретные ситуации, разрешенные на уровне моделей понимания, дополняют их так, что оказываются пережитым опытом реальных причин и следствий. В контексте сна, во сне старые сюжеты могут вспомниться, если они как-то были затронуты существующими от бодрствования активностями. И тогда сон может протекать в узнаваемых условиях, но по новому сценарию.
Осознание же сновидения или фантазирование включает его в контекст текущего восприятия, дополняя его субъективными домыслами. Так что забывание сновидений – естественно и необходимо, не стоит пытаться вспоминать сны и, тем более, предаваться фантазиям на их основе.
Сон и произвольность
Механизмы организации сна оказываются тесно сплетенными и с развитием механизмов произвольности. Так, сон еще в древней форме, организуя блокировку мышечной активности (fornit.ru/7270), а также активности со стороны восприятия (пусковые стимулы оказываются не в их рабочем контексте значимости), обеспечил систему произвольности возможностью управлять тонусом активности цепочек реагирования: механизмы уже имеются, осталось ими воспользоваться. Но стоит заметить, что в реализации Beast блокировка рефлексов организована отдельным механизмом, что дает основание предполагать, что механизмы сна не так прямо используется при осознании в природной реализации.
Произвольность напрямую конкурирует с функциональностью сна и, возможно еще и поэтому во сне специально подавляется волевой контроль (fornit.ru/5084) на четвертом уровне сознания. Хотя, может быть, что и не специально (но удачно), а просто наиболее эволюционно поздний уровень сознания оказывается и наиболее легко подавляемым в ходе неспецифического торможения третичных структур. Этот уровень далеко не всегда оказывается задействованным и при бодрствовании, он легко теряется при кислородной недостаточности (например, в высокогорье) и при воздействии некоторых веществ, проникающих через гематоэнцефалический барьер.
Особым случаем волевого участия в сноподобном процессе являются беседы. Среди компании людей, устроившихся для специфического проведения времени – беседы, запускается тема, обычно достаточно актуальная по последним событиям, кто-то начинает высказывать свое мнение, не задумываясь (некогда и неприлично), развивая ее по механизму сновидения, но вслух. У других это вызывает актуализацию своих активностей в связи с услышанным, сопутствующие цепочки мыслей и, если возникнет достаточно интересная мысль, человек выбирает подходящий момент, чтобы ее высказать (похоже на эвристическое “озарение”?). Это, в свою очередь, вызывает актуализацию связанного с такой идеей у других и т.д., пока тема не исчерпается как сновидение, но уже не у одного в голове, а у всех собеседников. Поэтому беседы можно назвать коллективным сновидением.
Не только сон организуется как специфический базовый контекст поведения. Любой такой контекст оптимизируется взаимодействием зон мозга так, что это дало повод предполагать существование функциональных систем мозга, специализирующихся на обслуживании таких состояний. Но стоит подойти к этому системно и видеть всю последовательность развития контекстов значимости и зависимых от него структур, чтобы понять более общие взаимодействия с их первопричинами и следствиями. Т.е. изначально функциональные системы мозга не возникали как самоцель, а являются результатом развития всех более общих и универсальных составляющих в ходе видовой и индивидуальной адаптации к окружающим условиям.
Точно так же, как мы рассмотрели контекст сна, можно было бы отдельно остановиться на всех других базовых контекстах. Но это выходило бы далеко за рамки системного рассмотрения общей модели механизмов психики, можно ознакомиться с соответствующими психическими явлениями: fornit.ru/1181.
Сон был выделен потому, что он играет особую роль в организации информационной картины индивида.
Принципиальная схема мозга
Как бы ни показался вульгарным такой заголовок, но все, что в реальной системе обладает причинно-следственным взаимодействием по определенному, понятному механизму, возможно формализовать в виде принципиальной схемы таких взаимосвязей. Поэтому нет ничего крамольного в определенном варианте такой формализации в отношении мозга не только бабочки, но и человека.
Вот принципиальная блок-схема текущей реализации Beast: fornit.ru/beast0.
Схемотехник (хоть электронщик, хоть программист) привычно мыслит блок-схемами потому, что реализация каждого блока не вызывает у него проблем, он сразу уже знает, как примерно это можно сделать, остается только выбрать наиболее подходящий вариант. Например, если нужно преобразование сигнала в цифровой код, то схемотехник знает множество воплощений АЦП и ему не нужно сразу же детализировать до конкретного варианта реализации в усилии представлять систему с участием АЦП потому, что есть достаточная уверенность в возможности реализации. Он набросает блок схему, затем станет детализовать каждый ее элемент, который тоже может состоять из вложенных блоков без конкретной схемы реализации.
Невозможно сразу формализовать сложную систему во всей совокупности составляющих элементов. Мы можем легко говорить про предметы, пока нет необходимости при этом учитывать все взаимодействия атомов и квантов, из которых состоят предметы. Хотя на определенном уровне рассмотрения причинно-следственных взаимодействий компонентов предмета, может понадобиться и квантово-механический уровень. Во многих случаях будет достаточно адекватно реальности говорить в рамках упрощенной детализации и это нисколько не требует воображения полной картины, учитывающей все.
Так и с мозгом, который принципиально можно рассматривать даже не на уровне отдельный нейронов, а на уровнях принципов организации его механизмов от базовых до более детализованных. Но никогда не получится положить на стол полную принципиальную схему его реализации.
На основе уже выясненных механизмов организации психики, можно представить самую общую принципиальную блок-схему мозга, описанную в базовой статье про МВАП (fornit.ru/7431):
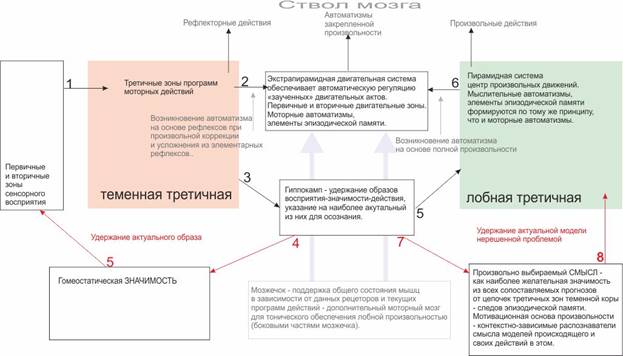
То, как такая общая блок-схема может детализироваться, демонстрируется в статье: “Схема основных функциональных блоков в организации адаптивности поведения” (fornit.ru/1396)
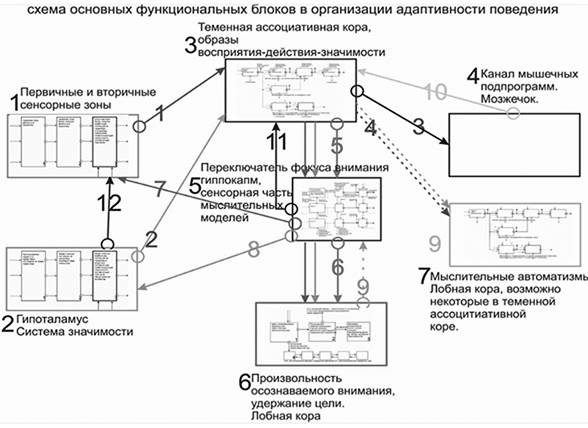
В каждом блоке проглядывает структура подблока, которая открывается при клике на ней (в статье).
В блоке 6 есть вложенный блок:
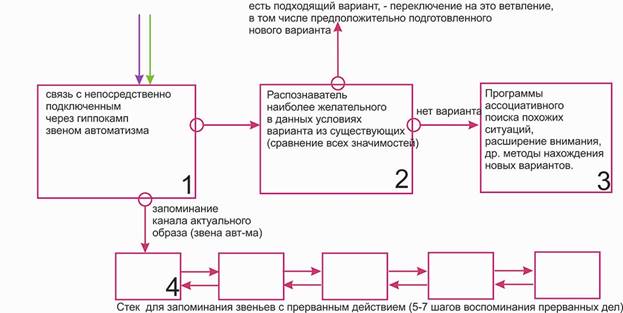
А в нем – еще вложенный блок 3 уже ранее раскрывался как блок-схема организации уровней сознания:
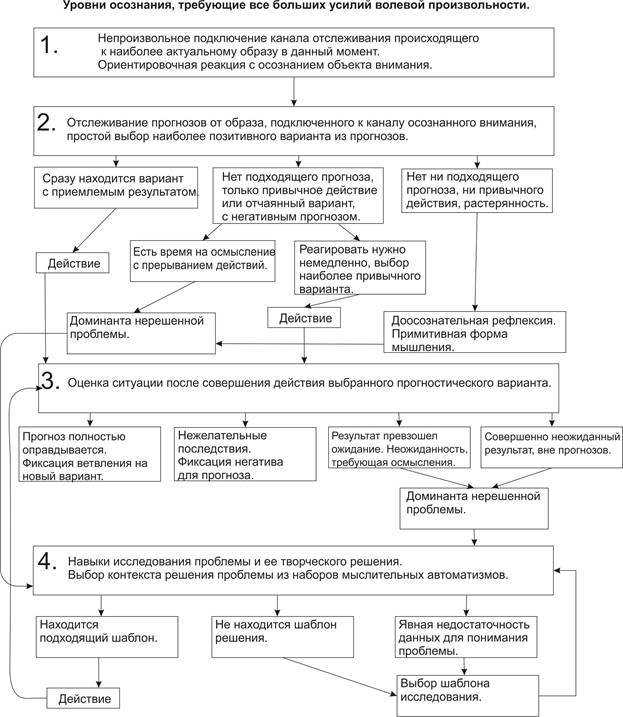
И этот уровень детализации, в свою очередь, может быть представлен вполне конкретными схемами реализации.
Очевидно, что привести полный комплект принципиальных схем для мозга в этой книге нет возможности, но нет и такой задачи. Главное – показать схемотехнический подход и его принципы при обобщении фактических данных о мозге.
Именно схемотехнический подход позволил обобщить огромный массив данных исследований в общую непротиворечивую систему, целостно описывающую всю иерархию развития природных нейросетей от самых простейших до механизмов организации психики, что затем было реализовано в действующем прототипе, а это, в свою очередь, еще более углубило и прояснило теоретические представления.
Для нейробиологов, не имеющих схемотехнического опыта, этапы такого описания выглядят недоказательными просто потому, что они не видят системы, определяющейся схемотехническими принципами. Но эти принципы достаточно жестко и однозначно укладывают данные исследований по своим местам в общем пазле модели и позволяют верифицировать как адекватность самих данных реальности, так и предполагаемые выводы.
Резюмируем уровни модели волевой адаптивности
Возможно, прочтенное во многом уже забылось и в голове не выстроился даже каркас описываемой модели. Поэтому сделаем короткий экскурс по условно определяемым эволюционным уровням совершенствования системы индивидуальной адаптивности. Эти уровни можно выделять по разным критериям, например так, как описано в статье Эволюция механизмов сознания: fornit.ru/1643.
Но в этой книге действует контекст схемотехнических критериев совершенствования адаптивности. Каждый уровень придает системе адаптивности новое качество в преимуществах индивидуальной адаптации совершенно определенными схемными механизмами. Обозначим коротко только те механизмы совершенствования, которые можно уверенно и надежно верифицировать.
Первый уровень – появление универсального распознавателя и наследственно предопределяемых схем управления, которые не отличаются определенной логикой построения, а являются лишь следствием случайно найденных преимуществ при использовании той или иной схемы управления. Важнейшим завоеванием эволюции, которое затем используется во всех последующих механизмах, является использование универсального распознавателя – нейрона и механизма образования связей после созревания нейрона до уровня пейсмеккерной активности.
Второй уровень – появление принципа формирования внутренних вспомогательных рецепторов в виде коллекций примитивов восприятия, которые не имеют непосредственных связей с эффекторными программами, но нарабатывают наиболее типичные образы восприятия, отражающие условия развития данной особи, и, тем самым, снабжая ее качественно более эффективными рецепторами сочетаний внешних воздействий реальности, на которые становится возможно реагировать.
Это происходит с развитием “новой коры”, где в первичных зонах возникают примитивы восприятия, порождаемые каждым из внешних сенсорных полей. Во второй зоне возникают более сложные примитивы уже с сочетанием нескольких сенсорных модальностей, а в третьей зоне добавляются примитивы восприятия значимости отклонения гомеостатических параметров от нормы и возврат их в норму. Это позволяет перейти от безусловных рефлексов к более адаптивным условным рефлексам.
На этом заканчивается область более-менее уверенного и общепринятого понимания нейрофизиологов. Дальнейшие уровни описываются и интерпретируются ими отрывочно и бессистемно.
Третий уровень – появление механизма удержания образов восприятия в памяти даже когда внешний объект уже не воздействует на сенсорные поля. Это - прорыв не только тактического превосходства над особями, тут же теряющими из вида все, что перестает воздействовать на их органы восприятия, но и качественного уровня новых механизмов.
Замыкание выхода образа восприятия переключателями гиппокампа на вход примитивов системы гомеостатической значимости, связанных с условными рефлексами, позволило этим рефлексам действовать даже при пропадании вызвавших их стимулов.
В контекстах этих рефлексов (если забылось, что представляют собой конкретно такие контексты, стоит вернуться к теме “Контексты значимости”) появляется возможность выявить новые условия и то, с какой значимостью связаны признаки новых условий. Это – чрезвычайно важная информация, которая позволяет научиться обращать внимание на ту новизну, которая прежним опытом была связана с высокой значимостью потому, что такая новизна выходит за рамки уже имеющих рефлексов и требует формирования новых, учитывающих новую значимость.
Четвертый уровень – использование значимой новизны для формирования новых реакций для новых условий. Для этого начала развиваться лобная часть мозга, по другую сторону коллекций примитивов моторных действий. Мозг как бы разделился на область рефлекторную и область произвольную. Главная функция второй области – формирование новых реакций для новых условий, переводя их в привычное так, чтобы уже не было необходимости снова привлекать лобные доли (т.е. обращать внимание на значимое новое с помощью “ориентировочного рефлекса”).
С этого уровня более старые структуры мозга остаются консервативно мало изменяемыми у всех видов животных, у которых они надстроились более поздними адаптивными механизмами. Это является общим свойством в формировании любого организма: старые структуры остаются все менее вариабельными для механизмов изменчивости (иначе изменчивость в более старых основаниях сделает невозможным все последующие механизмы, которые базировались на предыдущих). Теперь эволюционному совершенствованию в основном подвергаются лобные доли, в которых возникает все более эффективные механизмы формирования новых реакций для новых условий и механизмы, позволяющие волевым усилием запускать такие новые реакции вместо привычных.
Пятый уровень – совершенствование механизмов произвольности от самого простого отслеживания наиболее актуального среди происходящего до уровня доминанты нерешенной проблемы. Это сопровождается развитием самообучения на основе механизмов подражания и совершенствования субъективных моделей понимания (смысла происходящего) и произвольности действия (возможности поступать вопреки привычному).
Пятый уровень сам может быть подразделен на последовательность еще четырех качественно улучшающих адаптивность уровней (по числу уровней функции осознания).
Начиная с развития подражания и самообучения особое значение приобретает социальная коммуникабельность.
Схема процесса осознания Beast
Ни один вид рефлексов не обеспечивает достаточно эффективную адаптацию к новым условиям, реакции для которых отсутствуют или дают негативный результат.
Чтобы организовать формирование автоматических реакций в условиях появления нового и наиболее значимого раздражителя, необходимо создать систему обработки “информации” об этом новом объекте, получение предполагаемого варианта реагирования, апробацию его в реальном действии и учет результата.
Под информацией будем понимать данные о значимых для системы (в первую очередь для ее гомеостаза) коррелятах образов восприятия и ответного действия, данные о состоянии гомеостата и всей цепочки активных механизмов адаптивности. Информация, в общем плане – данные о значимости для системы любых активностей ее механизмов.
Назовем систему, в которой сводятся все предусмотренные функционированием значимости, осознанием происходящего, потому что в момент привлечения внимания к объекту в одном месте собирается вся доступная информация о нем.
Постулируем, что единственной функцией осознания, конечной целью его механизмов является нахождение достаточно эффективных ответных реакций для данных условий и пополнение базы таких наработанных автоматизмов, использование которых в ставших привычными условиях уже не требует осознания.
Чем больше информации, коррелирующей со свойствами объекта внимания и условиями текущей ситуации, тем больше возникает принципиальных возможностей для распознавания зависимостей между особенностями информационной картины и возможными составляющими образа ответной реакции. Это – самый сложный момент понимания.
Эволюция опробовала самые разные варианты организации сбора информации о наиболее актуальном объекте внимания и текущей ситуации, реализуя это в разных видах живых существ. Чем больше фишек в такой игре, тем больше возникает возможностей переборов вариантов, приводящих ко все более эффективному результату.
Рассуждая чисто формально, ясно, что что чем большим числом значений свойств описывается объект и текущие условия, тем точнее возможен выбор адекватного ответного действия.
В ходе решения проблемы нахождения эффективных реакций возникают новые виды информации, например, Правила, показывающие, что в данной ситуации при данном Стимуле и данном Ответе возникает такой-то эффект (позитивный или негативный). В Правилах информация об образе Стимула в текущей ситуации коррелирует с информацией о получаемом эффекте при использовании данного Ответного действия.
Последовательность Правил образует прогноз возможных событий, так что становится возможным ориентироваться, что может произойти после данного действия в данных условиях или что нужно сделать, чтобы достичь данного результата. Так, при игре в шахматы опытный игрок имеет множество цепочек Правил, описывающих комбинации в игре и после хода противника подбирает наиболее подходящее продолжение комбинации. Когда человек ориентируется в знакомой местности, то, находясь в данном месте, он видит, куда нужно пойти, чтобы достигнуть цели.
Физиологи нашли нейронные структуры, фиксирующие такие Правила и назвали “Картой местности” (fornit.ru/6605, fornit.ru/7057). Правила в мозге локализуются около гиппокампа, обеспечивая быструю выборку нужного в текущих условиях и имеющейся цельи. Но указатели на элементы Правил (связи для их активации) находятся в лобных долях - там, где обрабатывается информация о наиболее актуальном объекте внимания.
В ходе осознания проблемы образуются и Ментальные Правила, которые показывают корреляции между способами решения проблемы и получаемым результатом.
Организация и использование Правил по своей основополагающей сути – то же, что и алгоритмы GPT - (fornit.ru/66139), но отличаются тем, что в GPT база ограничена только одним видом рабочих последовательностей (текстами, картинками, музыкой, вербальными символами общения и т.п.) и у последовательностей нет параметра их эффективности для тех или иных условий, т.е. в GPT вообще не рассматриваются контексты условий, а контекстами являются только степень совпадения цепочек последовательностей. С другой стороны, алгоритмы использования Правил несопоставимо проще, чем у GPT. Это похоже на сопоставление искусственных “глубоких” нейросетей с функциональностью нейрона.
В GPT следующее слово находится по статистике распределения вероятностей следующего токена, т.е. даже если перемешать слова местами, то ответ будет получаться всё равно одинаковым. “Пришлось придумать костыль — positional encoding. Этот механизм позволяет трансформерам "видеть" порядок входных токенов. Грубо говоря, positional encoding — это любой способ кодирования позиции слова внутри эмбеддинга.”. И таких костылей придумывается все больше.
Правила же представляют собой непосредственные готовые последовательности Стимул-Ответ для данных условий. Причем для данных условий, но в разное время могут быть совершенно разные последовательности, взятые из проб реальных попыток адаптивности, удачных и не удачных.
У Beast есть два вида Правил, хранящие последовательности Стимул-Ответ-Эффект: 1) для объективных реакций и 2) ментальных циклов мышления.
Правило смотрится с учетом уже пройденной цепочки и если такое совпадение есть, то оно выполняется. Ожидается ответ и после него снова смотрится подходящая цепочка. Так реализуются все известные комбинации действий, в зависимости от уже пройденных.
Кроме Правил, возникают такие информационные образования, как Значимости объектов (то, к чему приводят элементы действий, своих или стимулов), и многие другие образования, важные для достижения функционального результата.
На основе Значимостей возникают Модели понимания – связь данного объекта значимости с кадрами эпизодической памяти, где они участвуют. Это позволяет получить обобщенное понимание возможностей объекта внимания или действия.
Привлечение внимания к наиболее актуальному осуществляет древний механизм “ориентировочного рефлекса”. Это – наиболее общее приобретение эволюции, возникшее на основе системы удержания образов, которые могут временно пропадать из виду.
В каждый момент времени привлечения осознанного внимания обновляется общая Информационная картина, создавая условия для решения проблемы нахождения подходящей реакции.
Сам процесс решения не осознается (т.к. отдельные функции являются жестко наследуемыми механизмами по типу безусловных рефлексов), т.е. не вызывает обновления общей Информационной картины с возможностью ее прогностического сканирования. Меняющаяся общая Информационная картина доступна для произвольного осознания, для каждого из ее составляющих. Это значит, что любой из компонентов этой картины может быть выделен как наиболее актуальный образ осознанного внимания и по нему запущен процесс осознания.
Людям доступна такая произвольная активация для уточнения своего текущего состояния: мы можем задуматься о какой-то детали этого своего состояния. Хотя, в целом, таким самоощущением мы занимается довольно редко. В реализации Beast имеется такая возможность.
Однако, есть и более поддающийся осознанию процесс создания и коррекции сложных программ действий – инициативная произвольность. Она реализуется с помощью циклов мышления.
Циклы мышления позволяют формировать адаптивную реакцию любой сложности – как последовательность отдельных известных образов действий. Такую цепочку, хранящуюся в оперативной памяти, может быть запущена как действие и тогда по оценке результата первое звено становится автоматизмом, а последующие - продолжением его действия. Становится возможной осознанная корректировка сложных программ действия и выполнение этой программы уже не задумываясь. Так же становится возможным осознанный прогноз возможных результатов этих действий.
Такое использование циклов для осознанного создания и корректировки ранее созданных действия позволяет проявлять инициативную произвольность, а не реагировать только вслед за воспринятыми стимулами.
Как во время сновидения, так и во время размышления возможен процесс развития цепочки образов восприятия по их значимости, дающий наиболее актуальный другой образ и связанное с ними действия, а по образу этого действия – найти следующий. Получившаяся цепочка действий запоминается со значимостью конечного звена (т.е. оценке результата такой цепочки) и может замет использоваться как автоматизм для определенных условий. Это реализовано в виде функции “Спокойно найти подходящий образ действия в исследовательской ситуации. Выбрать все, что подходит под ситуацию с текущим актуальным объектом значимости, набирая цепочку из Правил и из Прогнозам по Правилам”.
Реализация инициативной произвольности в схеме Beast происходит в случаях:
- При стимуле с Пульта нет автоматизма или пытается найти альтернативу, в сомнении. Но нет походящего правила, Значимости и Модели чтобы выработать реакцию.
- Когда давно нет стимулов с пульта, но ситуация достаточно опасная.
- При исследовательском или ученическом контексте нужно экспериментировать (иначе - просто промолчать).
Процесс осмысления является активацией функции решения проблемы (не имеет значения как организована эта функция, в виде самоподдерживающихся циклов активности в нейросети или рекурсивным вызовом программной функции). Эта активация может происходить по внешним стимулам (внешний ориентировочный рефлекс) и ментально, в ходе запуска цикла осмысления. Таких циклов может параллельно работать несколько одновременно (в принципе, число параллельных циклов не ограничивается). И каждый из них, в случае нахождения решения проблемы (Цели в контексте Темы), изменяет общую Информационную картину (эффект озарения).
У простейших существ, обладающих лишь функцией удержания образа восприятия, такой цикл позволяет не терять из виду важную добычу или хищника (без этого, стоит пропасть мухи из вида, и лягушка забывает про нее). Но эволюция все больше добавляла в эту функцию новые полезные особенности, так что у человека она стала очень сложной и многоуровневой (можно выделить четыре качества различающихся по функциональности уровней). Все эти уровни реализованы в модели Beast и будут здесь описаны.
В природной реализации циклы активности, удерживающие текущий, наиболее актуальный образ, дополняются в каждом проходе шага итерации в виде новой информации, получаемой на этом шаге. Измененная информационная картина дает новое направление следующему шагу мышления. И такой цикл поддерживается, пока не будет решена проблема поиска ответной реакции или процесс не будет прерван более актуальным Стимулом.
Сравните это с философским описанием сути осознания Дж.Тонони (fornit.ru/7589).
Если раньше про циклы А.Иваницкого было известно только то, что они удерживают актуальный образ восприятия и обеспечивают его подключение к лобным долям, то теперь возникает полноценная картина циклической смены информационного окружения при переключении с одного удерживаемого образа на более актуальный (ориентировочным рефлексом).
Теперь ясна функциональность того, что называется картами местности (Правила) у гиппокампа и его роль в информационной картине при выборе действий в данной ситуации.
Множественные, параллельно существующие циклические активности порождают то, что называется подсознанием, а актуальные циклы организуют обновление информационной картины в ходе поиска решения по текущей Цели. Это решение может оказаться найденным “по аналогии” в одном из неактуальных, но активных циклов, приводя к явлению “озарения”. Механизмы такого вида решений становятся ясны в их модельной реализации.
При разработке системы был применен принцип природной эволюции: на каждом достигнутом уровне выделять качественно различающиеся информационные составляющие и пытаться использовать их для наиболее общей пользы организма: достижения позитивного эффекта данного шага адаптации.
Это выделяло главное из бесконечного многообразия вариантов такой реализации. Кроме того, общим контекстом служил имеющийся массив достоверных данных исследований проявлений психики, т.к. можно быть уверенным в том, что эволюция прогнала все возможные бесконечности вариантов, найдя оптимальные решения для каждого их существующих видов живых существ, что в наиболее продвинутой человеческой реализации дает наиболее полный и сложный набор выверенных адаптивных функций.
Система аксиоматики фактических данных исследований позволила собрать общее представление в виде модели понимания организации адаптивной произвольности человека: МВАП , на основе чего и стала возможной искусственная реализация системы Beast.
В основе системы осознания наиболее актуального Beast используется Базовое информационное окружение (картина самоощущения).
Каждый из элементов общей картины может быть осмыслен, т.е. вызвана общая функция для прогностического прохода элемента в качестве выделенного объекта внимания.
Алгоритм обработки выделенного объекта внимания.
Текущие условия данной ситуации распознаются тремя видами древовидных структур:
деревом моторных автоматизмов,
деревом текущей ситуации и
деревом текущей проблемы.
На уровне выделения наиболее актуального образа всегда существует проблема, потому что актуальное для обработки – то, что имеет новизну при высокой значимости, и ранее выработанные реакции (моторные автоматизмы) могут давать неожиданные, вплоть до фатальных, результаты.
Дерево автоматизмов активируется по Стимулу или при изменении гомео-состояния.
После этого активируется дерево ситуации, объективно (стимулом) или произвольно (ментально). К этому времени определяется Тема происходящего и активируется дерево проблем, создающее Цель.
Текущая Тема формируется во многих участках функций в коде. Тема должна давать контекст размышлениям, вот каким образом.
Поддержание актуальной темы-контекста общения - внимание к актуальному объекту.
Для данного объекта выбирается целевая цепочка Правил, которая начинает выполняться по шагам, всякий раз снова корректируя цепочку правил. Так строится разговор.
Для поиска подходящего реагирования запускается очередной цикл мышления над актуальным образом воспринимаемого.
Значимости объектов внимания
В начале мышления сразу выделяется наиболее значимый объект восприятия.
Образы восприятия и действия получают свою значимость в зависимости от текущей проблемы (ветки при активации дерева проблем). Эта значимость – эффект Правил - после активации Дерева автоматизмов.
Модель понимания объектов внимания
Каждый кадр эпизодической памяти хранит все условия его создания и т.о. возникает модель свойств объекта: для каждого условия – определенное Правило с участием данного объекта. Это позволяет мысленно сканировать модели понимания, чтобы видеть, что может происходить в разных ситуациях.
В специальной инфо-функции (№ 5) происходит такое сканирование, цель - эвристический синтез новых важных Правил.
То же происходит при мышлении во время Дремы, определяя направление мышления – по наиболее значимым компонентам в ветвлениях. Происходит раскрутка сюжета по моторным Правилам со сменой сюжета в направлении наивысшей значимости.
Информационные функции
Это – важнейшие функции процесса осознания:
1: "Подготовить продолжение цикла осмысления"2: "Выбрать о чем и как подумать, с учетом предыдущих шагов цикла"
3: "Найти подходящую инфо-функцию по опыту ментальных Правил"
4: "Находит и запускает мот.автоматизм в плохой ситуации"
5: "Идет цикл мышления об экстремально важном объекте "
6: "Подвергнуть сомнению автоматизм, если нет опасности (не нужно реагировать аффектно) и ситуация важна"
7: "Создать новый моторный автоматизм по действию "
8: "Ментальное определение ближайшей Цели в данной ситуации"
9: "Найти способ улучшения значимости объекта внимания "
10: "Вспомнить самое значимое, что было для начала размышления (кроме возврата на прерванное) с произвольной переактивацией дерева"
11: "Ментальное отзеркаливание действия оператора"
12: "Синтез новой фразы из компонентов, имеющих плюсы в Правилах"
122: "Выбрать ID действия имеющего плюсы в Правилах"
13: "Тупое повторение Стимула оператора. Попугайство."
14: "Ментально переактиваровать дерево понимания с заданными узлами"
15: "Для условия дерева автоматизмов в одиночных Правилах выбираем наилучшее"
16: "Случайно выбрать любую известную фразу и затем infoFunc7() + infoFunc17()"
17: "Запустить моторный автоматизм "
18: "Ощущение наиболее значимого из параметров Информационной картины"
19: "Было эвристическое озарение и найдено действие для пробы действия"
20: "Запустить процесс осмысления последнего кадра с пустым Правилом эпиз.памяти (было некогда думать)."
21: "Срочно найти какое-то подходящее действие и запустить его"
Если отключить функцию, то возникнет полноценный эффект лоботомии.
Первым делом выявляется наиболее значимый объект восприятия для мышления.
Функция осмысления имеет 4 уровня использования (отслеживает эволюционное назначение). При решении проблемы обработки нового значимого образа на данном уровне, уже нет необходимости переходить на уровень более глубокой и развитой обработки.
Первый уровень – отслеживание моторных автоматизмов, готовых выполниться в данных условиях. В опасной ситуации они просто выполняются, в противном случае проверяется возможный вред и последствия выполнения так, что автоматизм может быть заблокирован и выполнен найденный альтернативный или вообще не совершено действие.
Этот уровень не вносит ощутимую задержку на обдумывание.
Второй уровень – если на первом уровне не найдено решение – попытка найти подходящее действие, используя Правила. Это уровень импровизации имеющих достаточный опыт особей. Так ведутся шахматные партии-блиц, делаются артистические экспромты, ведется беседа.
Третий уровень – раз не найдено решение, то необходимо мышление об этом.
Мышление - это изменение общей информационной картины при запросах к информационным функциям в течении цикла решения проблемы. В контексте текущего состояния инфо-картины выбирается подходящая информационная функция для получения целевой информации, после чего полученная информация дополняет информационную картину.
Этот процесс может идти до нахождения решения проблемной ситуации или цикл мышления прерывается более актуальными стимулами и переходит в фоновое состояние, где (в неосознанном состоянии), по возможности, продолжается решение проблемы.
Для поиска решений в различных режимах осмысления (бодрствование, сновидение, мечтания, лень) и текущей особенности ситуации – используются Информационные функции - в каждом из шагов итерации мышления.
Четвертый уровень – если решение данной проблемы не находится более 3-х раз, создается Доминанта нерешенной проблемы – постоянно существующая структура, где запоминается проблема, условия и попытки ее решения.
Доминанта активируется во многих местах адаптивных механизмов и позволяет находить решение новым способом – по аналогии.
При удачном решении и его реальной проверке, доминанта закрывается (закрытие гештальта), но остается в базе опыта решений.
Доминанта — это просто пролонгированная Цель. Повышенное время существования Доминанты повышает шансы найти решение по аналогии с уже совершаемым действием.
Активная Доминанта, если она есть, все время сопровождает появление новой инфы восприятия так, чтобы была возможность найти решение по аналогии.
Решенная Доминанта является третьим типом формирования автоматизмов на уровне пятой ступени развития: творческое формирование автоматизма.
Решенные Доминанты являются источниками личного опыта и аналогов решения проблем.
Почему наиболее экстремальный образ обеспечивает самоощущение?
В принципе, каждое обновление структуры должно давать экстремальные образы для мышления.
Имеется возможность уточнять эти образы. Но оно и в природе имеет не управляющую функцию, а лишь отражает результаты управляющих функций и контекст для работы следующих.
Появляется возможность просмотреть, в чем именно заключается значимость наиболее значимого из параметров, которая находит экстремальный образ и запускает мышление.
Т.е. вслушивание в свое самоощущение дает какую-то пользу, разворачивая циклы мышления в определенном направлении и приводя к новым Правилам, иначе оно могло бы и не появиться эволюционно.
Каждый новый цикл мышления становится главным и осознаваемым, а все остальные выполняются в фоне.
Ментальные автоматизмы – опыт мыслительного решения проблем.
Дерево проблем, активирующееся с каждым новым циклом мышления, определяет текущую цель в данной ситуации.
К этому дереву могут привязываться ментальные автоматизмы, которые теперь становятся, по примеру моторных, носителями последовательности ментальных проблем, сохраняющими цепочку мыслей.
Дерево проблем, по сути, нужно только для привязки ментальных автоматизмов и его ID может использоваться для ментальных правил (как для мот.правил используются ID деревьев автоматизмов и ситуации).
Удачные ментальные правила сохраняются в таких ментальных автоматизмах, чтобы не совершать снова работу по поиску подходящего ментального правила. Это схоже с тем, как моторные Правила приводят к формированию моторного автоматизма.
Т.е. если нет штатного ментальными автоматизма для дерева проблем, то сначала идет поиск подходящего в ментальных правилах, а если нет, то уже - ментальные циклы.
Ментальный автоматизм может дополняться новыми ментальными действиями или формироваться новая версия.
Про циклы мышления Beast
В настоящее время циклы мышления отработаны во всех принципиальных аспектах реализации Beast и могут контролироваться с помощью специальных страниц Пульта связи.
Циклы, участвующие в процессе осознания впервые описал А.Иваницкий (fornit.ru/10300), а основы философских принципов наиболее удачно сформулировал Дж.Тонони (fornit.ru/7589).
При разработке действующей модели существа Beast (fornit.ru/beast) циклы мышления были реализованы и отработаны во всех проявлениях адаптивной функциональности.
Вот какова их роль в процессе адаптации к новому, когда нет уверенных привычных реакций для данной ситуации, коротко и просто.
Если появился актуальный объект внимания, нужно удерживать его образ, даже если он временно пропадает из восприятия. Муха, скрывшись за травинкой, должна оставаться активным образом внимания так, чтобы не терять интерес и не упускать возможность животного поймать ее. Эволюция привела к зацикливанию активности образа с выхода на вход так, что образ оказывается самоподдерживающимся и может участвовать в обработке механизмами поиска решений даже на время пропадания из виду.
Каждый новый актуальный образ подключается к пошаговой последовательности функций обработки ситуации, а ранее возбужденные образы остаются в фоне.
Чем более эволюционно совершенно существо, тем все более эффективные функции поиска решения в данной ситуации в нем существуют, тем оказываются большими возможности найти верное действие. Все такие функции, по сути, это – наследственно предопределенные структуры. Именно от их набора и эффективности зависит то, насколько большими возможностями обладает данное существо для поиска решений, процесс которого называется мышлением. Если их не хватает, то никакой жизненный опыт уже не поможет достаточно эффективно мыслить.
При этом решающим оказываются продолжительность критических периодов развития и то, насколько много новой информации перерабатывается в эти периоды в различные формы личного опыта.
Каждая самоподдерживающаяся активность, возникающая от нового актуального образа восприятия, образует свой цикл мышления.
Для каждого цикла мышления отрабатывают те или иные функции – в зависимости от текущей ситуации и результатов прошлого шага мышления, обновляя общую информационную картину ситуации и ее изменения. Измененная же картина дает новые данные для следующего шага мыслительной обработки.
Получается следующая картина. Когда существо проснулось, у него – только самые общие представления о ситуации – в зависимости от состояния жизненных параметров. И его пока единственный цикл мышления (возникший от ситуации пробуждения) основывается на полученной картине самоощущения. Срабатывание мыслительных функций может выработать какое-то действие, ситуация изменится, наиболее актуальный объект внимания откроет опыт (есть несколько разных видов такого опыта) того, что это значило ранее и подскажет дальнейшие действия.
Общая информационная картина меняется с каждым действием и с каждым воспринимаемым последствием этого действия, образуя правила причин и следствий.
Правила запоминаются в основном виде опыта. Кроме того, ухудшение или улучшение состояния после совершения действий привязывает к образам воспринимаемого этот эффект, образуя другой вид опыта – значимости элементов образа восприятия и образов собственного действия. И еще один важный вид опыта привязывает каждый образ восприятия и действий к ленте кадров памяти о происходящем (эпизодической памяти), позволяя создавать модели значимости воспринимаемого, т.е. того, к чему в прошлом приводили те или иные воспринимаемые образы.
Могут возникать любые другие способы сохранения и использования опыта, эволюционно оставляя самые полезные.
Любое действие может порождать новый актуальный образ возникающих последствий, что воспринимается в виде постоянно изменяющейся информационной картины, которая и образует картину текущего самоощущения. Эта картина позволяет с каждым шагом мышления находить решений уже в новой ситуации.
Без циклов самоподдерживающейся активности принципиально невозможно было бы организовать мышление для поиска решения в постоянно меняющейся ситуации.
Они постепенно накапливаются с каждым новым впечатлением, но схожие конкурируют между собой, гася более слабые. В конце дня и даже раньше, их может накопиться так много, что они своей общей активностью начнут приносить больше вреда, чем пользы. Во сне они обрабатываются в тех местах, которые не было времени обдумать, образуя новые правила, значимости и модели понимания.
Только один новый цикл мышления обновляет общую информационную картину самоощущения, которая составляет наиболее общий контекст работы всех циклов мышления. Старые циклы становятся фоновыми и все, что в них происходит, называют подсознанием. Но, если в каком-то фоновом цикле возникнет актуальное решение, то он становится основным циклом со всеми правами осмысления и возникает явление инсайда, озарения, эвристического решения.
Если в главном цикле решение не находится с нескольких попыток, то возникает особая активность – Доминанта, которая становится постоянным источником нерешенной проблемы, заставляя думать об этом, когда только возможно (не мешает что-то более срочное). Доминанты могут находить решение в любом из циклов по методу аналогии, такой цикл становится главным, решение обдумывается и запускается на пробу.
В реализации Beast осуществлена намного более сложная организация циклов мышления, учитывающая множество различных аспектов.
Так, привила, сохраняющие причинную связь действия и последствий с эффектом улучшения или ухудшения, позволяют находить решения в данной ситуации методом GPT (fornit.ru/66139) – нахождением наиболее верного продолжения, исходя из уже совершаемых цепочек действий-результатов.
Но еще используются ментальные правила, сохраняющие причинную связь между активацией тех или иных информационных функций и полученным в результате эффектом. Это позволяет использовать такие правила уже для направления хода мышления все тем же принципом GPT.
Реализаций циклом мышления привела к появлению всем знакомых психических явлений, например, появлению навязчивых мыслей (крутящейся в голове мелодии, фразы и т.п.).
Описывать конкретику реализации совершенно бесполезно, код достаточно сложен сам по себе, несмотря на пространные комментарии, но он наилучшим образом позволяет разобраться в сделанном.
Все сделано с помощью общей функциональности уровней осознания.
На первом уровне для текущей ситуации определяется наличие наиболее привычного ответа и такой ответ проверяется на приемлемость в данных условиях с учетом того, чем грозит такое действие. Из опыта всех возможных действий в данных условиях выбирается самый безопасный и запускается. Это происходит очень быстро как рефлекс.
Если привычной заготовки действия нет, то подключается второй уровень и методом GPTнаходится наиболее подходящий ответ по опыту правил.
Если и на втором уровне не находится решения, то запускается цикл мышления об этом, который активирует подходящие информационные функции. В текущей реализации Beast имеется 20 инфо-функций.
Одной из функций является нахождение пути мышления методом GPTпо опыту ментальных правил.
И, наконец, если в течении нескольких попыток решение не находится, то создается доминанта нерешенной проблемы и она пытается найти решения во всех случаях запуска привычных действий по аналогии.
Теперь должно стать понятным, почему так важен уже наработанный массив личного опыта, который формируется в критические периоды развития индивидуальной адаптивности. Этот массив используют функции мышления, без которого они бесполезны.
Первым таким периодом развития психики является развитие базовых автоматизмов на основе рефлексов и случайных действий, что готовит базу для эмпатийного отзеркаливания чужого опыта.
Во втором периоде происходит подражание и формирование собственных автоматизмов на основе наблюдения чужих действий. Нарабатываются опыт правил причин и следствий, формируются массивы значимости воспринимаемых образов и их связь с кадрами памяти о событиях.
В третьем периоде начинается подвергание сомнению перенятых реакций и совершенствование своих собственных. Продолжает нарабатываться и корректироваться массив приобретенного опыта.
В пятом периоде все большую роль в поиске решений начинает играть творчество и развивается опыт применения творческого мышления (ментальные автоматизмы).
Социальный уровень адаптации
Социальная среда с самого раннего периода развития формирует структуры и возможности всех уровней произвольности, начиная с имитации до самообучения.
Все то, чем наделяют “общественное сознание”, конечно же, представлено механизмами произвольности в каждой отдельной голове, и больше общественное сознание нигде не расположено. Отдельные разумы образуют общий эффект взаимодействия, составляя в каждой голове общее распределенное в индивидуумах сообщество, проявляющее как бы свою совокупную произвольность в случаях общих целей, когда установившиеся стереотипы этических норм оказываются в противоречии с новыми обстоятельствами, делающими следование этим стереотипам уже не соответствующим реальности и требующими корректировок реагирования, что вызывает привлечение осознанного внимания у отдельных особей в момент актуализации такой значащей новизны.
Чем сложнее учитываемые условия реакции, тем больше времени нужно, чтобы конкретизировать особенности этих условий просто потому, что такое сочетание встречается редко: чем сложнее, тем реже, иногда даже раз за одну жизнь. Одно дело реагирование типа: “Это – можно, а это – нельзя”, требующее распознавания образа “этого”. Нужно лишь время на то, чтобы экспонировать “это” в восприятии несколько раз за период формирования распознавателя. Совсем другое дело навык: “Это можно, если только то-то, но особенно нельзя, когда вот такое”. Тут можно и умереть от старости пока обнаружится, что “это особенно нельзя”, вообще не дождавшись его появления. Но когда особей много, кому-то с этим повезет больше, чем другим.
В общем, без передачи навыка другим просто не получится сформировать самому достаточно эффективную систему реакций с некоторого порога сложности из-за статистики частоты проявлений в природе. Возникнет просто маугли: fornit.ru/6365. И этот порог соответствует очень раннему уровню развития. Сколько одних только ядовитых растений и, наоборот, полезных, но незаметных среди других, и один человек все не перепробует.
Поэтому механизмы передачи смысла, связанного с полезными и опасными действиями, начинают работать сразу после рождения, пополняя коллекцию сопоставления собственных реакций с образами восприятия при наблюдении за совершением таких же реакций другими, - то, о чем говорили в теме “Обучение с учителем”.
Коллекция моделей понимания, отражающих причинно-следственные зависимости, восполняется не только живыми учителями, но и отражением неживой природы - на уже приобретенном уровне понимания и возможности сопоставления-обобщения: стоит заметить, как круглый камень скатывается со склона, можно потом что-то круглое скатить или даже самому скатиться быстро, сгруппировавшись максимально кругло или катясь боком. Можно даже изобрести колесо. Но Правил, усвоенного чужого опыта несопоставимо больше.
Главное – появилась модель возможного действия с понимаемой целью и абстрактный символ такого действия в виде распознавателя звена иерархии контекста, связанный с образными распознавателями сочетания признаков восприятия. Если активировался абстрактный символ – он потянул связку в цепи образа значимости восприятия-действия или сам подготовил к действию мыслительные автоматизмы, связанные с возможным достижением цели на таком принципе причин и следствий.
Если у человека есть навык разжигания огня, то все взрослые уже знают, чем именно он занимается, они понимают, почему ему в таких случаях не следует мешать, а, наоборот, лучше как-то помочь. И для этого не нужны слова, все бывает ясно молча. Как только кто-то начинает характерные действия, другие понимают, что он собирается сделать, даже если сами пока не умеют. Иногда достаточно сделать характерный жест или скривить лицо в узнаваемой мимике. Или издать специфический звук. Все это будет для других символизировать что-то, связанное с возможными действиями, обеспечивающими понимание других.
Роль слова среди других символов взаимопонимания
В культуре людей слово (точнее фраза) стало наиболее удобным и универсальным для передачи смысла. Поэтому часто из виду упускаются другие символы коммуникации. В культурном обществе кажется очевидной мощь слова, с которой ничто не может сравниться, хотя выразительность в сценическом искусстве демонстрирует большие возможности потому, что не всегда хватает слов для передачи смысла (немое кино вообще обходилось без этого). Даже мысли озвучиваются внутренней речью, от привычки использовать которую бывает трудно избавиться, как некоторым трудно не говорить самому с собою, когда он о чем-то думает.
Ильф и Петров живописал Эллочку Людоедку с лексиконом в 30 слов, которая достаточно комфортно интегрировалась в общество ее среды обитания. Конечно, на слух она понимает больше 30 слов, но сказать не может, зато может что угодно показать, поэтому ей и не нужно слова говорить. Таких эллочек разного пола можно встретить немало по жизни. Особенно в бытовом контексте, применяется гораздо больше невербальных символов взаимопонимания, чем слов, а между близкими людьми часто вообще не требуются слова, а они понимают друг друга по одному взгляду: в данной ситуации у них очень схожие контексты понимания.
Малословие нисколько не отменяет и не обедняет осмысленность понимания и интеллект (наработанные автоматизмы), и в своей тусовке Эллочка даст фору адаптивности новичку, хоть даже академику.
Общаясь в другой стране с мизерным лексиконом, все начинается с жестов и мимики, которыми возможно добиться взаимопонимания во многих вещах. Змея шипит, чтобы все другие побыли с ней осторожнее и т.п. и все ее прекрасно понимают.
Мы уживаемся с домашними животными без особых проблем взаимопонимания при минимуме словесного общения: они мяукают или гавкают – нам понятно примерно по их поведению, мы им говорим что-то – они по тону и многим другим признакам, а не по смыслу слов, понимают в чем дело.
В общем, кроме словесных символов есть несопоставимо большее количество несловесных. Нам часто не хватает слов, чтобы выразить мысль, и тогда выручают мимика и жесты, лучше слов передают переживаемое тому, у кого есть подходящий контекст понимания.
И, наоборот, если даже сказать известные слова, нет гарантии, что другой человек поймет смысл сказанного именно так, каким он был у сказавшего. Напротив, чаще всего смысл воспринимается с большими искажениями вплоть до совершенно иного или противоположного. Нужно сказать очень много слов, чтобы быть в достаточной степени понятым и убедиться в этом, и тогда контексты понимания в этой области окажутся достаточно согласованными. Одно слово, сказанное в контексте выразительной мимики и жестов, может заменить длиннющее словесное описание при условии подготовленности как автора описания, так и воспринимающего.
К сожалению, многие исследователи мозга и психики как-то упускают из вида совокупность высказанных доводов и опираются только на особенности преимущественного использования слов в культуре, хотя, когда они слушают музыку или смотрят картины в галерее или балет в театре, то там вообще нет слов, но воспринимаемое вызывает живейший и вполне предсказуемый отклик сопереживания.
Огромная часть этических символов взаимопонимания (fornit.ru/1679) несет смысл вовсе без слов, а, скорее, в дополнение к скупым словам, если они вообще есть.
При всем этом во многих теориях о психике укоренилась идея о непременной основополагающей роли слова не просто для каких-то мыслей, а вообще для сознания. А остальных видов абстрактных образов моделей понимания как бы и нет вовсе. Это – типичный тренд следования за авторитетами в науке, который прослеживается еще со времен Аристотеля (fornit.ru/889):
“Аристотель утверждал: у мухи - восемь ног. И авторитет Аристотеля среди ученых был настолько велик, что несколько столетий никому в голову не приходило проверить это утверждение на реальных мухах. Кроме утверждения о мухе, у Аристотеля есть и другие: например, утверждение о разном числе зобов мужчины и женщины и т.п.”.
Сказать и даже думать поперек авторитетов в академической науке (fornit.ru/121) чревато отношением как к диссиденту, а репутация ученого – самое дорогое, что у него есть. Поэтому раз авторитеты убедили себя и других, что слово – основа мышления, даже не понимая сути мышления, то проявлять сомнения и задумываться на этот счет катастрофически возбраняется, и это – один из серьезных недооцененных факторов авторитарного консерватизма в академической науке, против которого нет приема, в отличие от естественной нормы развития представлений через период попирания авторитарных догм у молодежи вплоть до шокирующей наглости.
Множество резонных доводов не позволяет ограничиться только словами как основой мышления и социальной коммуникации. При передаче чужого опыта слова бесполезны и даже мешают во всех случаях, когда еще нет нужных моделей понимания.
И слова разговорного языка – лишь частный случай проблемы формализации. Например, математика часто представляется как наука всех наук и что только с помощью математической модели возможно найти в самом деле корректное решение. Это – глубочайшее заблуждение потому, что сначала – понимание, а только после этого становится возможна формализация, в том числе и математическая. Вот способ убедительно показать это: fornit.ru/693.
У всех специалистов есть записанные словами методики. У кулинаров – рецепты блюд, у химиков – описания устройств и действий для проведения хим.реакций и т.п. Но стоит попробовать неискушенному человеку по кулинарному рецепту приготовить даже простую яичницу, и у него не получится так, как задумано автором рецепта. Не учтено огромное количество различных условий, нет огромного количества промежуточных навыков. А вот если он посмотрит внимательно, как готовит яичницу кулинар, то – совсем другое дело. Так что наблюдение (в чем зрение имеет гораздо большую пользу, чем слух) и имитация – важнейшие механизмы развития понимания и подготовки к возможности что-то сделать самому.
Мы, как социальные личности, практически полностью состоим из символов взаимопонимания всех модальностей восприятия, определяющих спектр наших возможных реакций потому, что все это – намного сложнее, чем то, что можно выработать самостоятельно с чистого листа.
От того, когда и как вырабатывались эти модели понимания, зависят индивидуальные способности и возможности. Это – социально обусловленный интеллект (fornit.ru/6693) – иерархия автоматизмов (моторных и мыслительных) на базе рефлексов, сформировавших основу моторных программ реагирования, которая и используется для поддержки моделей понимания возможного реагирования.
Если что-то не так с исходными рефлексами (безусловными и условными) до периода готовности системы произвольности, то какие-то моменты передачи чужого опыта окажутся невозможными, а смысл чужих действий окажется неверно понимаемым. Но такое возможно лишь при органических патологиях или очень ненормальных условиях развития.
Так что, если говорить про норму, то мы все полностью определяемся условиями и своевременностью имитации чужого опыта, а начальные особенности строения мозга все в меньшей степени оказываются ответственными за поведение. Наследственные первопричины в развитии практически полностью заменяются на реакции необходимости социальной адаптации, хотя все строится на первопричинах, которые только в начальный период влияют на то, насколько эффективно и быстро мы адаптируемся.
Бывает так, что первопричины дают настолько большое преимущество, что возникает вундеркинд, но быстрота развития у него, как правило, играет плохую роль в меньшей вероятности столкновения со сложными явлениями и, повзрослев, он оказывается уже непримечательным из-за недостаточности полноты обхвата.
Это – закономерно: ведь все новые уровни механизмов адаптивности как раз и служат тому, чтобы сделать особь как можно индивидуальнее приспособленной к условиям существования, и каждый такой уровень все больше отдаляет от материальной основы восприятия ко все большим уровням абстрагирования от нее.
Конечно, особь у которой будет более эффективная коллекция примитивов распознавания, скажем, определения числа штучек в кучке одним взглядом, будет иметь неоспоримое преимущество в моментальной оценке количества предметов, но на этом ее преимущество окажется этим ограничено и во всем более сложном, все менее проявляться. Большее значение имеет развитие моделей понимания, чем развитие более примитивных основ, хотя с некоторого количества недостаточности это станет качественно затруднено (не стоит пытаться научить собаку говорить словами, хотя она может научиться понимать тысячу слов: fornit.ru/24363).
Слепые и глухие нарабатывают компенсаторные способности и в общем деле социума оказываются не ущербными, а в чем-то даже превосходящими тех, кто в условиях всех благ не имел мотивации нарабатывать умения.
Так, красавчики смолоду заняты общением с девочками и друзьями, это для них - главная мотивация, а те, кому это не светит, заняты развитием других навыков и в этом получают неоспоримые преимущества. Красавчик и умник в одном флаконе – лишь мечта девочек, на самом деле у красавчиков хорошо развиты навыки производить впечатление умников и не лезть в карман за словом. Потом жизнь ставит все на свои места в реальных проявляемых возможностях. Так что родителям стоит намеренно принижать уверенность своих детей в неотразимости у противоположного пола до неуверенности в попытках отношений, пусть больше учатся другому интересному. Но, может быть, и нет, ведь роль домашней красивой кошки тоже востребована.
Специализация личностей
Мы – продукт своего развития в данном месте данным окружением, а не то, из чего и как мы сделаны. Если нет ущербности, мешающей сформировать модели понимания, а также недостаточности продолжительности периодов развития и моторных возможностей, то человек сможет освоить все что угодно. А запас по таким возможностям очень велик и определяет лишь то, какой степени совершенствования при сопоставлении и обобщении элементов общей культуры может достичь данный человек.
У кошек, собак и обезьян порог недостаточности качественно ниже, что обусловлено более короткими их критическими периодами развития, не формирующими необходимую сложность заготовок распознавателей, а потом – преемственную сложность моделей понимания. Но при этом нет разницы в качестве и особенностях переживаний на первых уровнях сознания, а кое в чем - и на четвертом уровне сознания (волки проявляют интеллект адаптивности к условиям в лесу эффективнее, чем у охотников).
У людей тоже есть немалая разница в сроках развития и те, кто демонстрирует стремительное развитие, будут проигрывать тем, кто больше времени формировал заделы данного уровня.
Если у одного человека есть модели понимания, которых нет у другого, то второй вообще не подозревает о них, никак не учитывая, а первый прекрасно видит эту недостаточность второго, но не может сразу ему пояснить, особенно если для этого требуются многие промежуточные пояснения. Поэтому не правы те, кто полагает, что если человек что-то хорошо знает, то он способен это объяснить так просто, чтобы стало возможным понять это. Ну, пусть объяснят первоклашке, что такое и зачем нужен круговой интеграл. Он может показать, как он действует, но для полноценной имитации может оказаться необходимой немалая подготовка.
Это особенно ярко демонстрируется спорами взрослых с детьми с “позорным” бессилием взрослого объяснить ребенку, почему тот неправ. Отсюда возникает эффект (Эффект Даннинга-Крюгера: fornit.ru/1682):
“Менее компетентные люди в целом имеют более высокое мнение о собственных способностях, чем это свойственно людям компетентным (которые к тому же склонны предполагать, что окружающие оценивают их способности так же низко, как и они сами). Также люди с высоким уровнем квалификации ошибочно полагают, что задачи, которые для них легки, также легки и для других людей”.
В самом деле, опыт состоит в большей степени из неудач (опыта того, что вот так делать не следует потому, что...), чем удачных решений, и у опытного человека есть ясное понимание того, как легко можно ошибиться и как тщательно приходится все учитывать, чтобы достичь цель и быть уверенным. А неопытный просто не видит проблем в достижении целей.
Этот эффект оказывается легко использовать для получения влияния в обществе, прибегая к заведомо некомпетентным утверждениям и заманчивым посулам, которые воспринимаются как очевидно верные людьми определенного уровня развития (чем и пользуются организаторы революций, воздействуя на наивных подростков и простаков).
На уровне наиболее общих символов взаимопонимания в культуре люди имеют сопоставимое качество переживаний (т.е. на таком уровне их субъективности неотличимы качественно). А на уровне базового самоощущения (наиболее общего контекста понимания) нет и количественных различий, т.е. субъективные переживания на таких уровнях идентичны. Развитие этих представлений приводит к довольно шокирующим выводам в отношении сути личной субъективности, как полностью самодостаточной абстракции (каковой являются по сути модели понимания), оторванной от материальной основы, что отражено в цикле статей про Эго: fornit.ru/1648 и в художественной интерпретации: fornit.ru/31007 и что порождает недоумение ученых, пытающихся найти взаимосвязь тела и разума.
У людей лет с 5 начинают формироваться автоматизмы на четвертом уровне сознания, возникают альтернативы произвольно понимаемых вариантов реагирования (этики данной культуры). А раз появились свои варианты понимания и реагирования, то возникает проблема сделать это понимаемым другими, т.е. суметь показать большую значимость оценки результата альтернативного реагирования. Отсюда – столь скудная подпитка культуры удачными продуктами самобытного творчества. Отсюда - консерватизм социальной культуры, ведь, чтобы доказать полезность своего продукта, требуются навыки воздействия на других людей, в том числе и авторитарного убеждения, с помощью занимаемой роли в обществе. Ведь чем выше авторитет, тем в большей степени такому будут доверять остальные. Но это означает возможность влияния и получения больших привилегий и без доказанной пользы своего творчества. Поэтому желание повлиять своей самобытностью возникает очень рано, развивая методы воздействия на общество.
Творческое продуцирование поведенческих альтернатив позволяет индивидуально оптимизировать свою адаптивность к реальным условиям, а не тем, что соответствует догмам учителей и обычаям общества, часто оторванным от реальности. Проявление таких инициатив в определенный период развития провоцируется каким-то образом физиологически, возможно наработанным соответствующим контекстом поведения: вместо безусловного доверчивого приятия, вдруг возникает скептицизм и желание оспаривать сказанное взрослыми вплоть до откровенной наглости. Разные люди допускают разную наглость попыток внедрить свои самобытные представления и действия. Как развивается контекст наглости можно прочесть в статье про это: fornit.ru/1134.
Социализация
Абстракции могут оказываться невыразимыми в виде материальных символов (таких – подавляющее большинство в голове) и такими, которые можно передать подготовленному человеку в виде символов общения и тот поймет информацию.
Абстракции (основы образов для мысли, идеи), не зависят ни от наследственности, ни от особенностей тела: одни и те же абстракции могут быть и у человека, и у кошки, которая над чем-то задумалась. Каждое мыслящее существо по сути своих переживаний – это любое другое мыслящее существо, только возникшее в другом теле и развившееся при других обстоятельствах. С каждым актом осознания меняется субъективная модель абстракций, человек становится другим, по набору поведенческих стереотипов, но тем же - по сути организации абстракций.
Можно представить себе устройство, создающее абсолютно точную копию человека. В первое же мгновение эти два человека уже не могут ощущать мысли другого, хотя поначалу легко догадываются. Но чем дольше они живут, тем более разными становятся вплоть до каких-то радикальных отличий. Можно убить любого дубля сразу после создания и очевидно, что ничего не изменится ни для одного из них: как было, так и стало. По сути, точно так же можно сказать и про разных людей с тем различием, что будет потеряна для других некая часть его субъективной модели, возможно, очень значимая для социума. А если нет ничего значимого, то и вообще говорить не о чем.
Это значит, что каждый человек значим не для себя, а только для того социума, в котором он возник, переняв особенности системы информационной коммуникабельности и определив свою роль в этом обществе. Это – непростой, но определяющее важный вывод: каждый – продукт того социального окружения, тех условных абстракций, которые были ему переданы при его развитии и индивидуально реализовали его произвольную систему понимания и отношения.
Каждый социум – муравейник, без которого отдельная особь теряет смысл (осознанную значимость) своего существования. Каждый социум несет в основе свой набор условных абстракций, информирующих всех, определяющих, что можно и чего нельзя.
Самый первый социум – родители, формирующие базовый уровень абстракций во всех актах отзеркаливания проявлений собственного отношения.
Более широкий уровень – район проживания, и здесь набор абстракций уже отличается, во многом оказываясь иным, чем в семье. Человек адаптируется к этому набору, начинает понимать и занимает свое социальное место, начиная реализовывать какую-то свою социальную роль в общем взаимодействии.
Наиболее общее – системы условных понятий (этики) в рамках всего мира на Земле (про вселенную пока молчим).
Именно из этической системы условных понятий, которую не обойти никому без серьезных проблем переадаптации, складываются все негативные социальные явления, с которыми пытаются бороться, без того, чтобы изменить саму порождающую систему понятий: коррупция, воровство, хулиганство и т.п.
Личность и социум
В зависимости от своевременности развития моделей понимания возникают разные специализации, определяющие возможные роли в социуме. Это настолько четко отражает представительство специализированных моделей понимания и их развития, что можно сказать: общество в своей культуре во многом повторяет организацию системы контекстов понимания или систему произвольности мозга — вот в каком смысле.
Как уже говорилось, каждый вид специализации (начиная от базовых стилей поведения) к новым условиям порождает соответствующие иерархии развития моделей понимания, интерпретирующие смысл происходящего и возможные реакции, что отражается на проявляемых чертах личности, характерных для данных условий: личность в детстве, личность в состоянии алкогольного (наркотического) опьянения, личность для работы по профессии, личное увлечение, религиозный верующий, попавшийся на чем-то преступник, навык общения со знакомыми людьми или навык общения с незнакомыми людьми и т.д. Все такие личности могут проявлять очень разные качества в одних и тех же частных ситуациях, но разных контекстах интерпретирующего понимания.
Автоматизмы уровней сознания полностью определяются в рамках этих контекстов, что и дает разницу в проявляемых качествах. Вместе с субъективной особенностью понимания смысла ситуации эти автоматизмы определяют субъективные особенности переживаний так, что можно говорить об активном проявлении одной какой-то личности среди всех заготовленных жизненным опытом. В самом деле, мы все воспринимаем очень по-разному в разных своих состояниях и мыслим тоже очень по-разному.
Каждая из личностей выходит на передний план, как только ситуация оказывается подходящей именно для проявления ее навыков, остальные в это время оказываются в небытии (без осознания) потому, что это – следствие подбора наиболее подходящей модели понимания в зависимости от условий и воспринимаемого. У нас не один “гомункулус”, а множество их, очень предупредительно и бескорыстно передающих управление самому подходящему из них (конечно, утрирую, с гомункулусами мы уже разобрались).
Вот так же, как каждая такая личность активируется с конкурентной активностью его контекста, выполняя наиболее выверенную часть задачи быть в максимальном соответствии с текущими условиями и ситуацией, так же и в фокусе общественного сознания оказывается та из личностей уже конкретного человека, который смог показать свою лучшую эффективность в данной специфике среди всех других.
Общество воспринимает ценные и понятные утверждения данной личности, апеллирует к ней для решения проблем ее специфики. Разница в том, что в общественном организме нет общей системы контекстной произвольности, а взаимодействие личностей происходит в системе общих представлений, которыми располагает культура, и технических информационных возможностей общения, роль которых выполняют СМИ, различные источники информации и этической коммуникации.
Сегодня это обеспечивается интернетом, телефоном и производными от этих технологий видами коммуникаций.
Завтра могут появиться вживляемые в мозг модули непосредственной активации распознавателей моделей понимания, которые, в отличие от очень индивидуальной системы образного восприятия, обладают очень высокой универсальностью в силу того, что формируются в культуре социума в процессе преемственности передачи таких сведений и актуализируют ту или иную абстракцию. Но проблема такого интерфейса в том, что для каждого контекста понимания существуют свои распознаватели одних и тех же абстракций, что и обуславливает их разный смысл в разных ситуациях. Одно и то же слово представительствует во всех контекстах, где оно ранее было сформировано и использовалось, а это – разные локализации в мозге, как со стороны образов (распознавание слов), так и со стороны абстракций (произнесение слов в контексте понимания его текущего смысла). Можно гадать, как может быть организован столь сложный интерфейс, но, если это будет сделано, то в таком обществе будущего (fornit.ru/938) станет возможной очень тесная синхронизация деятельности особей с образованием в самом деле интегрированной системы общественного сознания (fornit.ru/1513) на основе подключенных личностей, а не мифическая демократия (fornit.ru/19875).
Если мы уже обосновывали абстрактную оторванность от материального мира субъективных переживаний личности, и совершенно непринципиально, каким способом реализована схемотехника этих механизмов, то столь же непринципиальна и организация некоего межличностного мира общесоциальных переживаний – разума общества. Мы так же не можем ничего об этом сказать, как и о мире переживаний любого другого существующего человека из-за недоступности к нему своей субъективности. Но разумы других людей существуют одновременно в точности с тем же качеством самосознания, как и предполагаемый общественный разум. У нас нет способа практически использовать это, и мы просто привыкли считать себя в центре всего, окруженные другими чужими субъективными мирами, как считает каждый из этих разумов. Но это уже выходит за рамки этой книги.
Понятие личности
Каждый где-то с десяти лет с изумлением открывает, что именно он - исключительная личность в этом мире. Но такая сильная иллюзия возникает просто потому, что он не может заглянуть в мысли любой другой личности и увидеть ту же самую исключительность. Самосознание проявляется гораздо раньше - в период начала интенсивного формирования функций приспособления к миру наиболее эволюционно совершенных, лобных долей мозга, развитие которых завершается примерно в 25 лет.
Более 20 лет постепенно формируются осознаваемые представления на основе уже имеющихся более простых, достигая наивысшего уровня для данного индивида. Разброс по уровню достигнутой сложности очень велик у разных людей и зависит от большого количества различных факторов, среди которых наследственность играет лишь потенциальную роль.
Наиболее сильно влияющий фактор - особенности ближайшего социального окружения в возрасте от рождения до 25 лет. Этот период - самый длинный среди всех других животных и только у некоторых видов обезьян он соизмерим с человеческим, но значительно короче. Именно это и дает человеку имеющиеся преимущества и способности гораздо более глубокого понимания и приспособления к меняющимся условиям жизни, причем не только самого себя, а в тесной связи с окружающим обществом.
С момента осознания окружающего человек начинает осмысливать наиболее для него интересное, наделяя своим личным отношением. Это и есть момент появления новой личности - носителя субъективного или психики. Человек начинает познавать мир, строя свою субъективную модель взаимосвязей объектов внимания, включая самого себя. В такой модели не просто отражается происходящее, а вычленяется то главное, что касается именно данной личности, которая строит свое отношение ко всему. Это является следствием того, что сознание появилось как необходимость выделять из всего окружающего наиболее важное и новое для него - для того, чтобы научиться учитывать это новое, уточняя привычные, уже не осознаваемые реакции.
Подстройка поведения для новых обстоятельств - самая основная функция сознания (fornit.ru/5214) и именно так будем рассматривать все, что касается сознания далее в этой книге.
В результате все окружающее приобретает оценку личного отношения, которое обычно не выделяется и не осознается, просто весь мир воспринимается через такое личное отношение. Отсюда возникает ощущение своей исключительности.
У всех других людей - одинаковые принципы субъективного понимания, которые находятся очень далеко от того, что привносит наследственность, поэтому условно можно уверенно сказать, что все люди вокруг - одна и та же личность, но развившаяся из своих начальных условий и своего окружения.
Это так же, как если бы из одного человека суметь сделать несколько его точных копий. В первый момент у них будут совершенно одинаковые системы реагирования, но очень скоро различия в окружении начнут привносить свои уникальные особенности и чем больше времени пройдет, тем более разными будут становиться личности. Даже однояйцевые близнецы, если живут в разных условиях, во взрослом состоянии часто оказываются похожи лишь внешне, да и то, различия в субъективности накладывает на внешность свой отпечаток. Не зря в романе А. Камю "Падение" сказано: "После определенного возраста каждый человек сам отвечает за свое лицо".
При этом из-за схожести социального окружения и общего взаимопонимания разные личности во взрослом состоянии оказываются больше похожи одна на другую, чем одна личность на разных этапах своего становления. В детстве, юношестве, взрослости и старости - поочередно возникают совершенно разные личности по отношению к окружающему и самому себе: их реакции - оказываются специфичны к потребностям данного возраста и окружения так, что, например, повадки детства уже никогда не бывают востребованы взрослым.
Получается, что последовательно возникают и умирают несколько типов личности в одном теле. И это - не просто аналогия, а именно так все и обстоит на самом деле, но мы не делаем трагедии из очередного перевоплощения, хотя и ностальгируем о прошлой жизни.
Фасеточный разум
Академик В. Вернадский ввел понятие ноосферы. По аналогии с атмосферой это – сфера разума, условных понятий, но не как некий пассивный набор, а как общая основа, развивающаяся с развитием каждого отдельного разума в тот момент, когда этот разум оказывает влияние на других. Религиозные мистики пошли дальше и объявляли это Космическим Разумом, который, наоборот, сам влияет на отдельные разумы как передающая антенна, внедряющая мысли в головы, а собственных мыслей вроде и нет ни у кого, - так они “объясняют” феномен сознания.
В настоящий момент общество представляет собой объединенные общей культурой и условной этикой взаимопонимания разделенные разумы отдельных людей. Но при этом они начинают связываться друг с другом с самого раннего детства в неразрывную систему взаимоиспользования. У каждого возникают свои личные жизненные стратегии, и он решает проблемы тактики реализации этих стратегий. Этот процесс зависит от того, в каком текущем состоянии находится человек потому, что у каждого человека есть множество заготовленных личностей, берущих на себя роль “Я” в нужный момент, для которого они были заготовлены.
Так же и в любом обществе каждый человек специализируется в том, в чем он развил свой жизненный опыт, занимая определенную социальную нишу. Так было всегда, с самых ранних форм организации общества. Мало того, так организованы общества многих видов насекомых.
С развитием системы условных сигналов общения, передающих значения субъективных абстракций, развивались и технические средства, обеспечивающие оперативность и эффективность передачи сведений, которые все в большей степени объединяют людей по всему миру, сближают их. Сегодня каждый может связаться с другим человеком на другом краю Земли, мешает только незнание языка, но общаться можно и мимикой, жестами, передавая изображения, графики, ссылки на документы. В мире все более нарабатывается общая для всего мира этика и условные символы общения. Только анклавы субкультур, основанные на авторитаризме (религия, политика, власть), мешают этому процессу, так как субкультуры раскалывают общество и в отдельных странах.
Но в мире все больший вес обретает фокус общественного внимания: те люди, деяния которых так или иначе оказываются настолько значимыми, и новыми, что проявляется всемирный “ориентировочный рефлекс», заставляющий осмысливать это другими людьми.
Чтобы внести что-то свое в общественный разум, нужно привлечь общественное внимание и тогда происходит осознание, переживание и делаются выводы, закрепляющиеся как стереотипы общества. Чем более тесна интеграция, тем более общество напоминает единый организм или единый разум, и это – не аналогия, а непосредственное проявление определенных функций и качеств.
Остается совсем немного: как только киборгизация с целью улучшения эффективности жизни и общения достигнет возможности передачи не просто сведений, как это сейчас делается с помощью телефонов, электронной почты, компьютерных коммуникаторов, а непосредственно субъективных абстракций, возникнет новое качество единства, когда каждый непосредственно сможет понимать другого, как это происходит в преемственности ролей личностей в мозге.
К этому добавится понимание сути личностей и их общности за счет принципа единственности абстракций: мы в самом прямом смысле будем разделять одни и те же мысли в головах многих так, как если бы эти идеи пришли в нашу собственную голову.
Общественный разум все в большей степени начинает быть похожим на фасеточную форму объединения субъектов с общим механизмом их консолидации: фасеточный разум: fornit.ru/1513.
Дополнительные материалы по теме организации социума:
· Развитие ребенка и влияние родителей - fornit.ru/14408
· Дети и политика - fornit.ru/7524
· Личность и социум - fornit.ru/656
· Социализация - fornit.ru/1364
· Смерть и адаптивность - fornit.ru/7537
· Личность, система значимости - fornit.ru/657
· Социальная значительность - fornit.ru/19491
· Личность. Множественность личностей в голове и социуме - fornit.ru/658
· Контекст понимания, модели личности - fornit.ru/610
· Личные жизненные стратегии - fornit.ru/659
· Культура и стабильность - fornit.ru/644
· Преступность и культура - fornit.ru/11871
· Общество мифов - fornit.ru/19875
Далее предлагается развитие представлений о механизмах организации психики, конкретизированных на уровне схемотехнического подхода.
Творчество
Картина "Черный квадрат" Малевича произвела сенсацию тем, что расширила существующие рамки допустимого в изобразительном искусстве. До такой наглости еще никто не осмеливался дойти и при этом так чудесно ее объяснить. Потом были белый, красный квадраты, другие фигуры, но только тот первый стал символом новых возможностей.
Сам по себе этот квадрат ничего не стоит и люди недоумевают: ну что тут такого, да я могу нарисовать квадрат еще лучше! Но не новизна квадрата, а новизна смысла ситуации произвела сенсацию. Нужно знать историю живописи, те рамки, в которых она находилась, и что очень многое пробовалось, включая кубизм, но без такого триумфа чистой идеи. Нужно знать философию живописцев (а они все поголовно - истовые философы, обычно мистического толка) и то, какой смысл вкладывал Малевич в ту свою картину, и как продолжал ее пиарить впоследствии, объясняя глубочайший замысел.
Творчество дает не только продукт с новыми свойствами, открывающий нечто новое. Это, прежде всего, новый и важный смысл. И огромное число творческих произведений не было замечено потому, что не было понято или даже не осмысливалось.
Вспомним, что творчество это – процесс осмысления четвертого уровня вовлеченности механизмов сознания - в контексте доминанты нерешенной проблемы. И его продукт, чтобы был оценен другими людьми, должен дополняться идеей, несущей найденное решение не только для данного конкретного случая, а как метод решения всех схожих проблем. И эта идея должна быть достаточно актуальной для того, чтобы ей заинтересовались, или эта актуальность должна транслироваться через авторитеты и тогда ее примут люди, далекие от проблематики.
Высшим качеством творчества считается то новое, что уже тянет за собой существующий актуальный смысл в культуре и при этом открывает новый - в неожиданном и заманчивом качестве. Любому плевку краски, любому словосочетанию или звукосочетанию можно придать потрясающий смысл. Это и проделывают часто дизайнеры и когда заказчик с недоумением смотрит на неказистый и простой завиток его логотипа, а ему навешивают на уши гениальный смысл содеянного.
Это относится к любому виду творчества. Ученый открывает новый смысл расширяющих или углубляющих представления фактов. Ему нужно суметь найти именно такой смысл, такое значение воспринятого, чтобы это стало решением поставленной проблемы, важной для коллег так, чтобы они тоже сумели понять и произвольно оценить смысл этого решения.
Это - не просто переключение на более подходящий для текущих условий вариант действий, что требует самого примитивного уровня функциональности сознания. И это - не просто сопоставление ранее известного с выбором наилучшего прогноза. И даже еще более высокоорганизованный уровень, когда учитывается результат совершенного предполагаемого действия - еще не творчество. Только исследование в контексте доминирующей нерешенной проблемы с появлением новых сведений о ней, сопоставления разных проявлений, касающихся проблемы, и обобщающее формирование нового элемента субъективной модели, позволяет сотворить в самом деле новые возможности.
Таким образом, любое творчество, в конечном счете, является процессом решения новой проблемы на самом высоком, творческом уровне сознания, что невозможно без творческой фантазии.
Из-за столь высокого уровня организации (наибольшей сложности развития самых совершенных механизмов адаптивности) не всегда такой уровень оказывается доступным. Например, высоко в горах, в условиях гипоксии мозга творчество невозможно (не раз проверено эмпирически). Так же этот уровень недоступен в случае отравления алкоголем, шоковых ситуациях и во всех случаях снижения уровня сознания, к примеру в стадиях торможения во сне. При этом в фазе сновидений возможен конечный этап обобщений с созданием новой успешной модели и осознание ее значимости. Даже после сытного обеда голова не работает достаточно ясно для творчества, склонная к банальностям и поверхностному мышлению.
Анри Пуанкаре в статье "Математическое творчество" подчеркивает важность появления нового, именно практически полезного смысла (fornit.ru/694):
"В чем, в самом деле, состоит математическое творчество? Оно заключается не в создании новых комбинаций с помощью уже известных математических объектов. Это может сделать мало ли кто; но число комбинаций, которые можно найти этим путем, было бы бесконечно, и даже самое большое их число не представляло бы ровно никакого интереса. Творчество состоит как раз в том, чтобы не создавать бесполезных комбинаций, а строить такие, которые оказываются полезными; а их ничтожное меньшинство. Творить — это отличать, выбирать.
Как следует производить этот выбор, я объяснил в другом месте; в математике фактами, заслуживающими изучения, являются те, которые ввиду их сходства с другими фактами способны привести нас к открытию какого-нибудь математического закона, совершенно подобно тому, как экспериментальные факты приводят к открытию физического закона. Это именно те факты, которые обнаруживают родство между другими фактами, известными с давних пор, но ошибочно считавшимися чуждыми друг другу.
...В поле его сознания появляются лишь действительно полезные комбинации. ... полезными комбинациями являются как раз наиболее изящные комбинации, т. е. те, которые в наибольшей степени способны удовлетворять тому специальному эстетическому чувству, которое знакомо всем математикам, но которое до того непонятно профанам, что упоминание о нем вызывает улыбку на их лицах".
Упомянутый творческий выбор предполагает, что уже есть варианты, среди которых можно выбирать. Это - огромное число часто бредовых вариантов, которыми полнится неосознаваемое. Во время тяжелой болезни, бывает, этот бред, прорываясь через тишину мыслей, оказывается вовлеченным в сознание, порождая тяжелые переживания. Это - невероятное множество фрагментов пережитого в их различных сочетаниях, которые становятся ассоциативно связанными из-за изменения границ внимания. В норме первичным выбором занимаются неосознаваемые процессы, активированные доминантой задачи и выдающие результат в виде озарения. И они могут определенным образом организовываться сознательно. Но подробное описание этого (fornit.ru/7024) уже выходит за рамки книги.
Поводом для творчества всегда оказывается осознание новой важной проблемы, не решаемой более примитивными функциями сознания в рамках привычного. Важность и необходимость решения проблемы и намерение ее решить создает цель, порождает доминанту (по А.Ухтомскому) постоянной актуальности, которая переводит внимание на себя всякий раз, когда нет чего-то более насущного. Поэтому мысли днем и ночью оказываются в контексте нерешенной актуальной проблемы и любой стимул, хоть как-то ассоциированный с темой проблемы, может подтолкнуть развертывание цепочек предсказаний в какой-то даже не осознаваемой области и если там возникнет нечто, превышающее текущий уровень актуальности (новизны и значимости), то идея прорывается в осознание.
Так что не столько осознаваемая часть мыслей, как гораздо большая неосознаваемая часть, способна продолжать процесс решения даже если сознание занято чем-то другим.
Творческой проблеме сопутствует негатив отсутствия решения и волевые усилия, которые приходится применять, чтобы выйти за рамки привычного. Поэтому творческий процесс бывает настолько изнуряющим, что говорят о муках творчества.
Творчество и ремесленничество
Как уже говорилось, настоящей культурной ценностью является оригинальный продукт творчества, символизирующий прорыв в решение проблемы и открытие новых путей. Когда проблема решена и это вошло в культуру, оригинальный продукт уже не вызывает восторга, поражающего новизной и важностью. Он становится обыденностью и ценен только как символ совершенного. Копии и вариации воплощений этого продукта имеют вторичную ценность.
То новое в решениях, что вошло в культуру и носителем которого являются специалисты, применяется для насущных задач как уже готовое эффективное решение. Это называется ремесленничеством и это не требует привлечения высшего уровня функциональности сознания из-за привычности. Это позволяет что-то совершать, не задумываясь, а эффективный результат возникает как импровизация, радующая тем позитивным смыслом успешности, что был закреплен за составляющими эту импровизацию отдельными удачными решениями. Почти гарантированная эстетическая эффективность приносит радость ремесленнику. Он счастлив воплощением своих отточенных умений.
Вот этот момент качественно и отличает творчество от ремесленничества: Творчество всегда требует почти мучительного напряжения воли в поиске решения, а уже готовые решения радуют удачным результатом их применения.
Творцы вносят нечто новое в культуру, хотя далеко не всегда им удается привлечь внимание и заставить увидеть заложенный смысл. Ремесленники используют уже готовые решения и личные навыки, хотя часто, чтобы к этому прийти, им пришлось пройти мучительный путь творчества потому, что освоить просто по сведениям и по книжкам что-то невозможно, нужно суметь приспособиться к этому, сделав сведения своими знаниями.
Именно самобытная часть творчества, входящая в культуру, делает эту часть личности нетленной в культуре, дополняя символы взаимопонимания и связанную с ними этику взаимоотношений (fornit.ru/1679).
Нужно быть носителем культуры, чтобы понимать эти символы. Чужое этническое искусство увлекает только своей необычностью, но не понимаемо с позиции другой культуры. Вся музыка кажется однотипной и быстро приедается. Картины в лучшем случае оказываются узнаваемы в известных объектах зрительного восприятия. Тексты непонятны и не интересны потому, что в них скрывается недоступная пониманию часть связанного смысла. То, что в одной культуре представляется симпатичным, в другой может показаться отвратительным потому, что признаки симпатичности (fornit.ru/2073) очень разные.
Восприятие художественного творчества напрямую зависит от всех остальных предшествующих элементов культуры (fornit.ru/271).
Интуиция
Тот, кто сотворил что-то в самом деле стоящее, не может толком рассказать, как именно он это сделал потому, что подавляющая часть процессов творчества была не осознаваема. Мало того, не было какой-то одной последовательности решения, которую возможно проследить хотя бы по отдельным касаниям разума. Все происходило как подготовка отдельных элементов общей согласованной модели решения, постепенного накапливания самых разных, в том числе мусорных частей (которые нужно "не выплеснуть как ребенка"), выбором из которых постепенно складывалась целостная и непротиворечивая система, свободная от лишнего. И в какой-то момент эта целостность и непротиворечивость оказывается осознана как эстетическая гармония получившегося (как сошедшийся пазл или соответствие гармонии), связанная со смыслом того, что все это означает и может означать. Иногда решающие связи находятся случайно и к этому подталкивает наблюдение, казалось бы, совершенно неотносящегося к проблеме, но ассоциативно показывающее искомую связь. Иногда это происходит во время сновидения - как процесса обработки важной информации следов прошедшего дня. Но чаще всего нежданная мысль вдруг озаряет сознание.
А. Пуанкаре пишет об этом: "Прежде всего, поражает этот характер внезапного прозрения, с несомненностью свидетельствующий о долгой предварительной бессознательной работе; роль этой бессознательной работы в процессе математического творчества кажется мне неоспоримой; следы ее можно было бы найти и в других случаях, где она является менее очевидной. Часто, когда думаешь над каким-нибудь трудным вопросом, за первый присест не удается сделать ничего путного; затем, отдохнув более или менее продолжительное время, садишься снова за стол. Проходит полчаса и все так же безрезультатно, как вдруг в голове появляется решающая мысль. Можно думать, что сознательная работа оказалась более плодотворной, благодаря тому, что она была временно прервана, и отдых вернул уму его силу и свежесть. Но более вероятно, что это время отдыха было заполнено бессознательной работой, результат которой потом раскрывается перед математиком, подобно тому как это имело место в приведенных примерах; но только здесь это откровение происходит не во время прогулки или путешествия, а во время сознательной работы, хотя в действительности независимо от этой работы, разве только разматывающей уже готовые изгибы; эта работа играет как бы только роль стимула, который заставляет результаты, приобретенные за время покоя, но оставшиеся за порогом сознания, облечься в форму, доступную сознанию.
... эта работа возможна или по меньшей мере плодотворна лишь в том случае, если ей предшествует и за нею следует период сознательной работы. Никогда (и приведенные мною примеры достаточны для такого утверждения) эти внезапные внушения не происходят иначе, как после нескольких дней волевых усилий, казавшихся совершенно бесплодными, так что весь пройденный путь в конце концов представлялся ложным. Но эти усилия оказываются в действительности не такими уж бесплодными, как это казалось; это они пустили в ход машину бессознательного, которая без них не стала бы двигаться и ничего бы не произвела.
... Среди тех крайне многочисленных комбинаций, которые слепо создает мое подсознательное «я», почти все оказываются лишенными интереса и пользы, но именно поэтому они не оказывают никакого воздействия на эстетическое чувство, и сознание никогда о них не узнает; лишь некоторые среди них оказываются гармоничными, а следовательно, полезными и прекрасными в то же время; они сумеют разбудить ту специальную восприимчивость математика, о которой я только что говорил; последняя же, однажды возбужденная, со своей стороны, привлечет наше внимание к этим комбинациям и этим даст им возможность переступить через порог сознания".
Конечно же, в интуиции и озарении нет ничего мистического, если понимать то, как организуется психика на уровне сознания и на что вне осознания опираются эти механизмы.
У каждой активной доминанты нерешенной проблемы должен быть своя упрощенная функция осмысления -только для циклов мышления четвертого уровня. Информация, получаемая в таких циклах, не пополняет ощущаемую информационную картину до тех пор, пока не будет найдено значимое решение.
Можно задать работу своему подсознанию, формулируя важную проблему и время необходимого решения, после чего не то, что выбросить это из головы, а понимать, что там уже кипит работа, и она требует притока новых впечатлений по нужной теме, так что нужно помогать, собирая как можно большую и разную информацию, уже без надрыва, спокойно и вдумчиво. В нужное время возникает решение, настолько удачное, насколько для этого оказалось достаточно информации. Но главный выбор среди интуитивных решений и их доработка уже в составе моделей понимания делается осознанно.
Гениальность
Говоря о таланте, имеют в виду не только выдающиеся способности к творчеству в данном направлении, но и еще и высочайшую эффективность реализации этих способностей. Эта эффективность обеспечивается как соматическими особенностями, позволяющими более интенсивно использовать ресурсы мозга, так и личностными, в частности - особенностями мотивации.
Неспецифичность такого обеспечения приводит к тому, что гениальный человек делает более эффективным не только свой наиболее развитый навык, но все остальные (fornit.ru/301).
Если сформулировать главное, то гениальность включает в себя:
1) Повышенные возможности тела для обеспечения питанием и кислородом мозга и его метаболизма (доставка кислорода и глюкозы, вывод продуктов), что позволяет более длительное время поддерживать локальные и общие активности, а также наследовано предрасположенное особо удачное ветвление аксонов мозга, повышающее результативность творческого уровня сознания (больше процессов могут обрабатывать информацию).
2) Особую организацию доминанты цели решения проблемы, становящейся очень устойчивой настолько, что такая увлеченность заставляет максимально погружаться, буквально жить решением проблемы. Другими словами - живейший и неугасимый интерес к проблеме.
3) Талантливость или интеллект в области наибольшего интереса, что позволяет наименьшими силами достигать большего эффекта, а этому способствует воспитание и окружение, обеспечивающее специфичность элементов, важных для предмета талантливости, и достаточная преемственность в культуре.
Замечено, что у гениальных людей не бывает подавленного состояния сознания, голова всегда ясна и свежа, а они полны энергии. Замечали, что этому способствуют широкие носовые ходы. Даже общепризнанно выделяют одну из наций, особенно хорошо подготовленную соматически: евреев. Гений лучше других способен адаптироваться к какой-то специфике деятельности, но гениальность сфокусирована на его наиболее захватывающем, доминирующем увлечении.
Часто оказывается, что гениальность проявилась в качестве компенсации какого-то дефекта, потребовавшей особых усилий и развития. Человек, у которого все хорошо, которого с детства все любят, а в юности женщины вешаются ему на шею и у него нет проблем, вряд ли станет творческим гением. Так, великие ораторы, подобные Демосфену, развивали свой талант с целью преодолеть заикание или дефекты речи (fornit.ru/7624).
Как и интуиция, гениальность не имеет никакого отношения к мистике. Она не возникает на пустом месте так, чтобы гений сотворил нечто совершенно ни в чем ранее не предрасположенное, с полного нуля. Кроме характерного стиля продуктов творчества и самобытности, гений всегда "стоит на плечах" своих великих предшественников (fornit.ru/302). Это - та преемственность в культуре, которая состоит из более простых основных элементов этического взаимопонимания, и обрастает все более сложными производными.
У творчества есть начало и есть естественный конец. Естественный потому, что с накоплением жизненного опыта для данного окружения все реже появляются новые проблемы. Если только нет такого общего исследовательского контекста, мотивируемого неподдельным личным интересом, который не может иссякнуть, например научный интерес какой-то развивающейся предметной области. В противном случае привлечение сознания становится все более ненужным и возникает дезадаптация. Ее признаки даже при полном здоровье организма проявляются в виде комплекса (синдрома) признаков, называемого деменцией fornit.ru/40983.
Практические следствия модели МВАП
Но книжного знания при прочтении этой книги будет недостаточно. А вот после глубокого участия в проекте Beast возникнут собственные знания и понимание станет ясным.
Нужно много временя и усилий с искренней заинтересованностью (мощная и длительная доминанта нерешенной проблемы), чтобы сформировать личный стиль схемотехнического мышления и начать ясно размышлять о вариантах искусственной реализации адаптивных принципов или критически воспринимать такие варианты для верификации их возможностей.
Но уже сейчас при восприятии новостей, публикаций по психическим явлениям и их интерпретации, становятся ясно видны случаи недостаточности методов исследования, не учитывающих важнейшие принципы и даже попирающие их, сразу понимается недостаточная обоснованность утверждений, или, наоборот, в исследованиях сразу замечается то, что хорошо вписывается в систему других эмпирических фактов исследований, составляющих МВАП (или, рассматривая вне формализма МВАП , не противоречит другим надежным данным, а дополняет их), хотя такие публикации, естественно, встречаются гораздо реже.
Становится понятно, где тормозят Яндекс и Гугл в потугах внедрить интеллектуальную выдачу поисковых запросов. И даже бездушная семантическая модель, внедренная в качестве поисковой системы (https://you.com/), вызвала панику у разработчиков традиционных поисковиков.
Становится очевидно, где тормозят авторы тех или иных теорий разума. Во всяком случае, легко и ясно обнаруживаются несоответствия и противоречия, которые уже можно предметно верифицировать.
Также становится возможным как угодно корректировать детализующие представления о МВАП , если для этого окажется достаточно веских оснований, согласующихся с общей стратегией критериев полноты и верности теории (fornit.ru/7431).
Рассказанное в книге, при неизбежных вероятностях отдельных неточностей, дает главное: понимание того, без досконального исследования чего невозможно решить проблему организации психики, т.е. все без исключения упоминаемые элементы модели должны быть взаимно согласованы потому, что они жестко следуют из воспроизводимых данных исследований, образуя взаимодействующую систему принципов, отражающую реальные причины и следствия. Возникает понимание механизмов каждого из таких принципов в их взаимодействии с другими. Становится очевидной необходимость схемотехнического, а не какого-то другого подхода, в том числе и безуспешность попыток математического моделирования, которые столь же неуместны, как в случае целостного математического описания работы компьютера или даже микроволновки – это практически совершенно бесполезная, зато модная затея.
В этой главе – ознакомительная тема, где будут показаны только самые важные практические следствия и обобщения, следующие из концепции МВАП . Вовсе не обязательно пытаться прочесть все предлагаемые статьи и книги как можно скорее, так что каждый сам решит, как ему лучше поступить. Но если найти время на эти тексты, в которых с самой разной позиции и точки зрения и используются принципы организации психики, то темы книги окажутся более полно раскрыты.
Далее - примеры практического использования будут распределены по тематическим разделам.
Педагогика
Здесь ключевой является фраза: “Произведение новизны на значимость – формула привлечения (осознанного) внимания”.
Утверждения: “При наблюдении выделяются те свойства объектов внимания, которые 1) важны (интересны) наблюдателю, т.е. он уже находится в распознанном контексте объекта внимания, определяющим его возможные свойства и 2) у наблюдателя уже есть собственные реакции, схожие с тем, что проявляются объектом внимания (иначе он просто не увидит это)” - имеют к педагогике самое прямое отношение: совершенно бесполезно пытаться чему-то научить, если ученик не находится в заинтересованном состоянии узнать свойства для какой-то своей практический цели.
Вывод: “...красавчики смолоду заняты общением с девочками и друзьями, это для них главная мотивация, а те, кому это не светит, заняты развитием других навыков и в этом получают неоспоримые преимущества. Красавчик и умник в одном флаконе – лишь мечта девочек, на самом деле у красавчиков хорошо развиты навыки производить впечатление умников и не лезть в карман за словом. Потом жизнь ставит все на свои места в реальных проявляемых возможностях. Так что родителям стоит принижать уверенность своих детей в неотразимости у противоположного пола до минимума неуверенности в попытках отношений, пусть больше учатся другому интересному” – определяет возможности создания доминанты нерешенной проблемы, которые только и способны сформировать дееспособные знания.
Статьи по теме:
· Дети - fornit.ru/7024
· Развитие ребенка и влияние родителей - fornit.ru/14408
· Дети и политика - fornit.ru/7524
· Педагогика и механизмы психики - fornit.ru/1705
Формирование осознанных навыков
Фраза из книги: “Нужно дробить цель (моторного) движения на более элементарные с коррекцией каждого их них, давая возможность мозжечку освоить координацию тела для обеспечения такой цели”.
Фраза из книги: “...во сне реализуется альтернатива осмысливанию при бодрствовании, которое случается редко и не все к этому склонны. Когда читаем книгу, обычно тоже не отвлекаемся на ее осмысливание, чтобы не потерять нить повествования, а все, что нас зацепит, остается в виде актуальных активностей, которые нельзя терять, иначе зачем мы вообще читали книгу. Вот почему после прочтения трудного текста очень желателен сон, что оптимально сохранит полученную информацию.”.
Фраза из книги: “ Чем больше домысливания по сравнению с использованием уже проверенных данных (привычных автоматизмов), тем больше ошибок (fornit.ru/1012) предположений возникнет и закрепляется в памяти. А любое субъективное предположение должно проверяться в реальности, чтобы появилась адекватная оценка и уверенность очевидности (fornit.ru/7117). Так что явно у нас есть какое-то ограничение склонности фантазировать наяву, которое может довести до серьезных психопатологий (fornit.ru/1318). Это ограничение легко преодолевается многими людьми, особенно в молодости, т.е. ограничивает жизненный опыт негативных последствий фантазирования.
В этом есть очень важный практический вывод: если возникает необходимость построить теорию, то не следует использовать волю в размышлениях потому, что тогда подставляется то направление мыслей, которое в чем-то важно именно нам, нравится нам, а не то что должно бы следовать безучастно. Не следует давать себе волю строить теорию, она должна сама построится на основе достаточного количества сопоставляемых данных, ведь именно на этом и заточено наше сознание, наши модели понимания. Нужно строить формальную модель, описывающую естественно складывающуюся модель понимания причин и следствий, отражающих реальность. Ни в коем случае не следует любить свою теорию и потакать ей хоть в чем-то, это обязательно как-то вылезет боком.
И здесь возникает противоречие с функциональностью четвертого волевого уровня сознания, которое прививает волевые усилия для реализации решений. Т.е. понимание возникает до использования четвертого уровня, но результат проверки верности предположений получается на волевом уровне, когда нужна определенная воля для учета этого результата, как бы ни была дорога идея. ”.
Статьи по теме:
· Осознанное формирование поведенческих навыков - fornit.ru/983
· Адаптивная методика метания ножей - fornit.ru/6412
· Здоровье и счастливое долголетие - fornit.ru/12971
· Научная методология против иллюзий восприятия - fornit.ru/817
· Психогигиена - fornit.ru/1185
· Гениальность или об эффективности поведенческой адаптации - fornit.ru/301
· Самосовершенствование - fornit.ru/1276
· Личные жизненные стратегии - fornit.ru/659
· Как жить чужим умом - fornit.ru/19649
Творчество на практике
Фраза из книги: “Использование интуиции для нахождения подходящих вариантов действий называется эвристикой: fornit.ru/1647. Можно натренировать навык задавать задачу своему подсознанию и не думать о ней специально, пытаясь решить “логически”. Важно убедительно осознать важность и необходимость решения, можно еще научиться задавать желаемое время получения ответа, с учетом того, что для подсознательного решения нужны новые ассоциации, т.е. достаточное богатство восприятия и мыслей, а на это нужно время. Поэтому задавать срок нужно как можно подальше, может быть, не один раз подступая к решению, и в это время заниматься чем-то близким к оставленной задаче, думать о сходном, читать о попытках решений других людей. Если решение будет найдено, оно само привлечет к себе осознанное внимание. Нужно вовремя зафиксировать его, не рискуя отвлечься на что-то и забыть.”.
Фраза из книги: “Но совсем другое дело творчество, намеренно искажающее реальность и провоцирующее совершенно неожиданные предположения. Они заведомо воспринимаются с большим интересом из-за своей значимой новизны, но так же, как мутации, очень редко оказываются полезными, так же и из “психоделических” идей, возникает что-то практически полезное. Они довольно быстро приедаются потому, что мир субъективных фантазий (и даже компьютерного моделирования) не сравнимо беднее мира объективных явлений, и идеи лучше черпать из реального мира, чем и пользовался Леонардо Да Винчи, разглядывая узоры штукатурки, наводящие его на мысли заданной темы”.
Теперь можно более внятно представить, что такое нахождение нового в творчестве, и именно новое и имеет наибольшую ценность. И это – не просто случайно возникшая новая комбинация, а ценность новаторскому решению придает значимость решенной проблемы.
Поэтому недостаточно просто разглядывать узоры штукатурки. И не каждому дано найти новое в какой-то области творчества. Первое условие – быть на переднем краю представлений ведущих творцов в данной области. Иначе, в лучшем случае будут изобретаться банальные велосипеды. Важно поставить себе интересную и значимую нерешенную проблему, чтобы возникла доминанта этой проблемы. Привлечение осознания проблемы всегда оказывается на основе чего-то нового и значимого. И вот в таком доминантном контексте рассматривание штукатурки (или обогащение восприятия любыми другими природными проявлениями подсказок, которых нет в субъективном мире) может подтолкнуть подходящий неосознаваемый автоматизм к эвристической находке, которая прорвется в сознание новой мыслю.
Более подробно о том, что и как влияет на процесс творчества: Оптимальная скорость творчества: fornit.ru/24631.
Статьи по теме:
· Основные механизмы творчества - fornit.ru/970
· Ученые - особая каста - fornit.ru/14245
· Свобода утверждений в науке - fornit.ru/6477
· Математическое и эвристическое мышление - fornit.ru/693
· Неудовлетворенность существующим - fornit.ru/870
Маркетинг
Фраза из книги: “Произведение новизны на значимость – формула привлечения внимания”.
Статьи по теме:
· О рекламе - fornit.ru/907
· Стоимость и цена - fornit.ru/1381
Произвольность оценки происходящего
Статьи по теме:
· Модели понимания и зависимость - fornit.ru/7489
· Роль религии в современном обществе - fornit.ru/14107
· Быть или верить? - fornit.ru/202
Развитие произвольности
Фраза из книги: “У простых термостатов есть некоторый гистерезис: включение нагревателя вызывает перегрев, пока на это отреагирует датчик, потому как у нагревателя есть инерция (он не может сразу нагреться и сразу остыть). Поэтому температура в таких термостатах постоянно колеблется, т.е. возникает генерация колебаний температуры. И чем больше инерция нагревателя или других элементов отрицательной обратной связи, тем больше будет амплитуда колебаний. Если задержка окажется настолько большой, что вместо отрицательной обратной связи возникнет положительная (т.е. момент воздействия окажется противоположным тому, что нужно бы для компенсации), то колебания станут максимальными по амплитуде. Но если сделать некий сглаживающий фильтр, например на выходе термометра, который не позволяет быстро менять показания, а отслеживает некий средний уровень по времени, заведомо больший, чем период колебаний температуры, то колебания прекратятся и установится равновесное состояние на заданной температуре.
Это – наиболее общее условие генерации колебаний в системах с обратной связью, в независимости от природы колебаний.
В организме оно так же работает. Например, нужно удержать руку в каком-то положении, делается усилие поднять ее. Это усилие зафиксируют рецепторы положения руки (или зрительные рецепторы через все системы мозга, распознающие это положение) и когда рука достигнет нужной высоты, дается команда на уменьшение усилий до уровня сохранения нужной высоты. Но если есть какие-то задержки в реагировании, то момент нужного уровня будет пропущен и нужно будет опускать руку. Возникнет дрожание (тремор).
При сильном волнении, когда на плавность движения привлекается максимум внимания и движение контролируется буквально на всех стадиях, часто возникает тремор потому, что осмысление требует времени и возникает задержка. Это может закрепиться в автоматизмах. Или тремор может возникнуть из-за патологии каких-то промежуточных стадий, например, прекращения действия некоторых рецепторов положения руки и тогда момент нужного положения будет пропущен.
В быту люди с повышенным давлением склонны принимать сбивающий давление препарат, когда давление поднимается. А это бывает при стрессе, физической нагрузке и т.п. ситуациях, когда организму нужно поднять давление для обеспечения функциональности. В результате давление довольно быстро снижается и даже намного ниже, чем нужно, возникает болезненная вялость, сонливость, нарушения координации, в общем, давления организму очень не хватает. Тогда гипертоник принимает препарат для повышения давления (пьет родиолу, крепкий чай и т.п.). Давление подскакивает. И т.п.
Если же не торопиться с решением противодействовать, а выждать время, то вообще бы не потребовалось принимать препараты потому, как последствия стресса бы довольно скоро закончились.
Бывает при сильной жажде так хочется пить, что выпивается огромное количество воды. Сигналы о восстановлении водного дефицита возникают намного позднее, когда на борт заливается слишком много воды. Это может нанести определенный вред (как с лекарствами от давления) и это стоит учитывать, не торопясь выпить побольше до того, как жажда не исчезнет.
Во всех случаях подобного восполнения жизненных параметров стоит помнить об этом и не переедать до вредных последствий, не перепивать, не переохлаждаться в жару и т.д.”.
Статьи по теме:
· Математическое и эвристическое мышление - fornit.ru/693
· Системное мышление и формализация - fornit.ru/7597
· Эвристика вероятности - fornit.ru/7498
· Психическое явление Интерес или Инициатива наказуема? - fornit.ru/7188
· Психическое явление Превосходство - fornit.ru/7181
· Минимизация усилий - fornit.ru/728
· Осознанное формирование поведенческих навыков - fornit.ru/983
Зависимые состояния
В медицинском и психогигиеническом аспекте многие автоматизмы оказываются нежелательными для субъекта потому, что по многих случаях они приводят к негативным последствиям. В норме такие случаи достаточно легко корректируются. Например, споткнувшись о неожиданное, мы осознаем это, замечаем помеху и запоминаем так что в следующий раз в этом месте автоматически уже корректируем движение (на первый раз с осознанным вспоминанием ситуации, а потом вообще не отвлекаясь на это).
Но некоторые автоматизмы по ряду причин оказываются неподатливыми и просто так не корректируемыми. Это относится к наркотической и т.п. зависимости, когда сильные переживания, закрепленные с данным автоматизмом, продолжают вызывать его выполнение вопреки волевым усилиям.
Это сильно усугубляется тем. что такие автоматизмы формируют свой эмоциональный контекст на самом низком уровне гомеостатической значимости. Субъект начинает формировать специфический опыт (мыслительные и моторные автоматизмы в контекстах субъективных моделей понимания) в этих контекстах.
Многие ситуации своими признаками вызывают привычное переключение в такие контексты и тогда уже очень трудно становится осознанно контролировать свое восприятие и действия, противоречащие сформированным автоматизмам.
Такие автоматизмы в своих эмоциональных контекстах условно будем называть зависимыми состояниями.
Зависимые состояния – основа любой психопатологии, кроме органических поражений (поражений в широком смысле, от генетических нарушений, до дисфункции органов, систем организации метаболизма и т.п). По классификации психологов сюда входят неврозы и психозы, истерические состояния и т. п., по классификации психиатров - многие формы шизофрении, биполярных расстройств, параноидальных, психопатических и т.п. состояний с сопутствующими проявлениями (бред, галлюцинации, иллюзии, бессвязное мышление и т.д.).
На первый взгляд и, конечно же, с позиции специалистов, все это относить только к зависимым состояниям – слишком надумано, но при условии понимания принципов и механизмов формирования субъективных автоматизмов, все встает на свои места.
То, что в попытках эмпирических классификаций разных предметных областей и врачебных дисциплин имеет огромное количество разных названий, на самом деле имеет одни и те же механизмы разбалансировки поведенческой адаптивности восприятия и действий, которые следуют из понимания системы организации механизмов психики (модель МВАП ). И суть этой разбалансировки заключается в создании привычного зависимого состояния (т.е. человек оказывает зависим от него, как он зависим вообще от любого своего эмоционального контекста) в которое человек попадает всякий раз, когда возникают определенные условия и признаки, которые привычно переключает контекст восприятия и действия в систему понимания и реагирования со своими устоявшимися и трудно преодолимыми стереотипами.
Теория организации механизмов психики МВАП очень сложна и для ее понимания не только в качестве “книжных знаний” но и для формирования адекватного реальности опыта, требуется немало лет и практики. Поэтому сначала приведем конкретные и исчерпывающие шаги для преодоления субъективной зависимости. И это предлагается принять в качестве постулата – на предварительную веру, чтобы при заинтересованности изучить все более углубленно. Всего три шага раскрывают самое главное.
1. Прежде всего, для преодоления любого вида зависимостей, нужно иметь неподдельное, собственное желание избавиться от этой конкретной патологии. Это – самое общее и совершенно необходимое условие (то, что в теории называется созданием доминанты нерешенной проблемы). Ни сам человек никто другой со стороны не в состоянии без этого условия скорректировать патологический стереотип.
2. Нужно помнить и применять следующий принцип: любой стереотип поведения реализуется без непосредственного участия сознания (потому что его суть – неосознаваемый автоматизм), хотя человек может осознавать происходящее, но при этом не осознает именно то, что составляет стереотипные действия. Достаточно осознать нежелательность происходящего так же, как при осознании привычного действия мы начинаем его совершать уже строго по командам осознанной последовательности, и стереотип будет заменяться на уже осознанные последовательности действий и. тем самым, окажется под контролем сознания. Это значит, что стоит обратить внимание на то, что ты совершаешь, как привычный автоматизм приостанавливается и в дальнейшем выполняется то, что следует из произвольного (осознанного волевого) усилия.
3. У каждого зависимого состояния есть то трудно избегаемое позитивное переживание. которое ранее и закрепило его выполнение. Бывает, что волевого усилия недостаточно, чтобы его преодолеть если наготове нет достаточно убедительного довода почему не сделает так делать. И чем глубже развилось зависимое состояние (чем полнее совершился переход в данный контекст восприятия и действий), тем это оказывается труднее. Все знают, что если настиг ужас или ярость или вожделение, то уже очень трудно выйти в другой эмоциональный контекст. Поэтому для каждого зависимого состояния, от которого решено избавиться, нужно заготовить такой подходящий довод и его реализацию в виде убедительной команды себе немного подождать (по такой-то причине). А чем больше период ожидания, тем легче переключиться в другой приемлемый эмоциональный контекст и выйти из патологического состояния.
Суть методики преодоления зависимости в том, чтобы 1) вовремя распознать наступление ситуации зависимости 2) осознать сам процесс действий при реализации зависимости, что остановит ее выполнение и 3) решить подождать с дальнейшими действиями и за это время попробовать придумать более правильное продолжение действий. Именно так можно переписать любой привычный автоматизм.
Социология
Статьи по теме:
· Депрессивные состояния - fornit.ru/345
· Особенности социальной адаптации - fornit.ru/980
· Социальные проблемы психики - fornit.ru/16104
· Преступность и культура - fornit.ru/11871
· О демократии и либерализме - fornit.ru/7585
· Дети и политика - fornit.ru/7524
· Психофизиология протеста - fornit.ru/1197
· Ненависть к несправедливости - fornit.ru/7467
· Почему нарастает неравенство - fornit.ru/7363
· Проблема рабочих мест - fornit.ru/7397
· Культура и стабильность - fornit.ru/644
· Общество мифов - fornit.ru/19875
· Роль религии в современном обществе - fornit.ru/14107
· Иллюзия счастья или Стратегическая ошибка сапиенсов - fornit.ru/7148
· Как удачно выйти замуж - fornit.ru/7126
· Психогигиена - fornit.ru/1185
Философский потенциал
Для размышления о разуме и психике появляется хороший философский потенциал понимания, способный адекватно интерпретировать воспринимаемые сведения по теме и самому рассуждать в контексте такой специфики проблем на уровне непосредственного подхода к научной стадии исследований. Курс тематики позволяет планировать целевые, а не случайные исследования в рамках имеющейся модели.
В последних главах напрямую тематика касалась чуть ли не эпатажных выводов про суть Эго: fornit.ru/1648, то, во что развивается тенденция все большего усиления технической коммуникации. Это напрямую изменяет сложившееся в современной культуре отношение к смерти. Да, во многих известных культурах смерть не считается трагедией, но это – в силу религиозного убеждения в вечной жизни души и неважно, что это будет за жизнь, страшно уже не было даже тем, кому за грехи сулили посмертные страдания. Но совершенно иная картина возникает с пониманием сути субъективных переживаний, сути абстракции субъективной реальности, которая уже не является принадлежащей к материальному миру: fornit.ru/1132, хотя стремящейся адекватно отражать его. Такое отношение к смерти отдельной личности конкретизируется в статьях “Непостижимая исключительность бытиЯ” (fornit.ru/861) и “Смерть и адаптивность” (fornit.ru/7537).
Статьи по теме:
· Очевидность - fornit.ru/7117
· Неудовлетворенность существующим - fornit.ru/870
· Общество мифов - fornit.ru/19875
· Социальная значительность - fornit.ru/19491
· Протест очевидности или почему люди спорят? - fornit.ru/19687
· Как жить чужим умом - fornit.ru/19649
· Как объяснить, что такое ложка? - fornit.ru/7657
Современное состояние академической науки о психике
Это достаточно полно характеризуют следующие статьи:
· Что люди узнали о мозге - fornit.ru/19798
· Почему до сих пор нет общепризнанной модели организации психики?. - fornit.ru/7074
· Скромное очарование этологических теорий разумности - fornit.ru/7313
· О реорганизации академической науки - fornit.ru/19847
· Ученые - особая каста - fornit.ru/14245
И вот – заключительная глава о том, насколько самодостаточно или изменчиво эго и то, во что превращается вопрос: “Что такое Я?”.
Быть или не быть
Есть одна страшилка, следующая из индивидуальной разобщенности - страх смерти.
Очень во многом он воспитан традиционным отношением, существующем в культуре. Есть культуры, в которых это явление отсутствует или отсутствует для некоторых социальных групп: чукчи, кельты. И отсутствие страха смерти не мешает таким народам полноценно жить. У профессиональных воинов тоже возникает бесстрашие в отношении смерти.
С другой стороны, есть люди, настолько панически боящиеся смерти, что это становится их постоянной проблемой, мешающей жить.
Вопрос о смерти не вызывает сомнения в том, жить ли дальше, и только люди с глубоко развившейся психопатологией, или убеждающиеся в бессмысленности дальнейшего существования, или жертвующие собой ради чего-то, выбирают смерть.
На самом деле смерть - субъективная абстракция, которая возникает из-за свойств выделения объекта из окружающего, когда свойства объекта меняются так, что он перестает соответствовать понятию "живой". Это касается и отношения к собственной смерти.
Эпикур пришел к утверждению, что там, где есть смерть, нет нас, а там, где есть мы, нет смерти. Это - пример высказывания, верного по формальной логике, дающего направление осмысления представлений о смерти. Вот как Эпикур высказывает свою мысль: "Приучай себя к мысли, что смерть не имеет к нам никакого отношения... Все хорошее и дурное заключается в ощущении, а смерть есть лишение ощущения. Поэтому правильное знание того, что смерть не имеет к нам никакого отношения, делает смертность жизни усладительной, - не потому, чтобы оно прибавляло к ней безграничное количество времени, но потому, что отнимает жажду бессмертия... Глуп тот, кто говорит, что он боится смерти не потому, что она причиняет страдания, когда придет, но потому, что она причиняет страдание тем, что придет: ведь если что не тревожит присутствия, то напрасно печалиться, когда оно только еще ожидается. Таким образом, самое страшное из зол, смерть, не имеет к нам никакого отношения, так как когда мы существуем, смерть еще не присутствует, а когда смерть присутствует, тогда мы не существуем. Таким образом, смерть не имеет отношения ни к живущим, ни к умершим, так как для одних она не существует, а другие уже не существуют".
После смерти влияние социально активной личности на социум не прекращается, пока не будет исчерпана новизна такого влияния.
Для социума как единого организма смерть индивидуума означает лишь прерывание его текущей активной деятельности. Уже в современном обществе роль отдельных людей бывает определяющей лишь в отдельные ключевые моменты их активности, и чем более развито общество, тем такая роль бывает все менее индивидуальной: многие открытия и изобретения зарождаются не в одной голове, а нарастают в культуре, прорываясь сразу в нескольких передовых умах (проблемы приоритета возникают все чаще). На самом деле в культуре каждое новое качество развития подготавливается множеством промежуточных элементов, созревая сразу во всем обществе, точнее в той его субкультуре, которая специализирована на этом.
Ценность жизни каждым осознается очень высоко и страх смерти так же излишен, как страх божий для ценности этических убеждений. Он способен только помешать трезво и адекватно оценивать происходящее и принять верное решение, в том числе, когда нужно принести себя в жертву или прекратить уже ничего не дающее существование. В последнем случае — это вовсе не моменты нерешаемых проблем в жизни, а действительно полная немощь с утратой человеческих качеств (fornit.ru/1468), но в таком случае уже не удается принять решение на должном уровне разума и верх берут более примитивные реакции.
В любом случае не следует принимать непоправимое для себя решение: нельзя решать за себя будущего в новых обстоятельствах, а именно к этому склонны многие молодые при возникающих трудностях, кажущихся им безысходностью в силу наивности (Эффект Даннинга-Крюгера).
Даже в полной неподвижности и беспомощности Стивена Хокинга, он очень сильно влияет своим разумом на воспринимающих его идеи людей. Слово, даже произнесенное в последний момент, или даже записи, обнаруживаемые после смерти, могут оказать решающее воздействие на чью-то жизнь.
Из-за условности понятия невозможно точно провести границу между жизнью и смертью: смерть субъективного бывает намного раньше смерти тела, а смерть личности - намного позже. При старении все в большей степени востребованы автоматизмы и в меньшей - сознание, пока все не станет привычно автоматическим.
Каждая личность - не только то, что она представляет собой сейчас, а и то, что она будет представлять собой на протяжении всей своей жизни и как оказывать влияние на окружающее, в первую очередь - на культуру своего социума.
В самом общем плане, можно сказать, что понятие своевременности и своеместности существования является более общим, чем понятие естественного отбора, и применимым вообще к любым объектам наблюдения, в том числе неживым, то, что было сведено к двум принципам специализации, которые выводятся в статье о наследовании: fornit.ru/806:
1. Форма развития объекта нашего наблюдения (чего бы то ни было) зависит не только от внутренних свойств объекта (генетической программы развития у живых существ), а от того, какое влияние оказывает на это внешняя среда. Только в среде, на которую рассчитана программа, она дает результаты, оставляющие объект в привычной нам форме.
2. Каждый предыдущий шаг последовательности развития создает новые условия для последующего, изменяя форму объекта.
Хронотоп личности (fornit.ru/7545) определяется социальным окружением так, что личность оказывается в определенном месте определенного времени развития тела и общества, откуда она не может быть просто перемещена в другое место и время развития тела и общества без потери той части функциональности, что была специализирована для прежних условий (fornit.ru/7537).
У каждого есть иллюзия исключительности своего существования (fornit.ru/861), которая обычно осмысливается примерно в 13 лет. Так кажется только потому, что нет возможности заглянуть в разум другого, иначе бы стало ясно, что у него - точно такое же исключительное отношение к своей жизни, - свойство отражения окружающего с точки зрения текущей модели своего я.
Но в обществе, как едином организме, связь разных эго устраняет иллюзию исключительности для интегрированных членов так же, как нет иллюзии индивидуальной исключительности у каждого из множества специализированных Я в одной голове. Система "Я в детстве" без трагедии безвозвратно уступает свое место более новым "Я - взрослый". В жизни множество разных эго специализируются для своих условий, беря на себя осознание в момент своей актуальности, а другие модели Я остаются неосознаваемыми на это время, хотя их активность и решение каких-то своих задач может продолжаться, в нужный момент порождая интуитивное озарение (fornit.ru/12318).
Логика Эпикура слишком лаконична, чтобы преодолеть иллюзию индивидуальной исключительности. В статье Эго со всеми ее продолжениями (fornit.ru/1648, fornit.ru/160, fornit.ru/861, fornit.ru/7537) дается дополнительная логика, показывающая условность субъективного понятия смерти личности. Одно из самых ярких и убедительных сопоставлений предлагает сделать следующий мысленный эксперимент.
Если создать абсолютно идентичную копию человека (неважно каким образом), то возникнут личности с совершенно одинаковым исключительным отношением к себе. Если после этого тут же уничтожить любую из них (ведь нет разницы какую, если они абсолютные копии), то ясно, что никакой трагедии от этого не будет: каким был человек, таким и остался. И понятно, что никакой потери тоже не будет ни для какой из копий - по той же причине. Попытка привнести религиозную душу тут ничего не меняет - рассуждения остаются теми же самыми. Но это требует очень глубокого осмысления, чтобы понимание стало очевидным и больше не требующим обдумывания.
То устройство, каким мы являемся, можно выключить на время и потом включить (что и происходит при полной анестезии или в некоторых стадиях сна), и опять личность воскреснет и начнет себя осознавать исключительной. Но при этом множество других личностей других людей и даже других животных точно так же осознают свою исключительность, просто разрозненно. Выключенное одно устройство не мешает продолжать осознавать себя всем другим, просто не привносит активного влияния.
Можно сказать, что, если не учитывать индивидуальные различия, мы одновременно существуем во всем множестве тел, причем не только человеческих и не только на Земле, но никто из нас не остается застывшей особенностью, а постоянно меняется так, что, бывает, мы оказываемся больше похожи на другого, чем на себя в прошлом. Это трудно осмыслить, но это строго показывает субъективную условность понимания смерти и своей исключительности.
Каждый специализируется в своем окружении и занимает социальную нишу с полной преемственностью предыдущих личностей и внося свой вклад в общую ментальность. И роль каждого динамично меняется.
Одной из форм смерти личности, кроме ее деградации, является безусловная вера в авторитарно навязанную картину мира и этику поведения (fornit.ru/202). Все, что составляет предмет безусловной веры, оказывается внедренным извне как непреложная истина. Это означает, что в этой области не может возникнуть собственная, самобытная модель понимания, определяющая эго. Фактически это означает чужую жизнь по чужим правилам: эффект марионетки.
Вера исключает развитие в новых условиях потому, что исключает произвольность в вещах, не допускающих сомнения. Возникает просто ментальный клон. О таких говорят, что человек живет не своим умом. Создание армии таких клонов живописал Курт Воннегут в романе "Сирены Титана".
Кроме этого, вера, препятствуя индивидуальной адаптивности, порождает нарастание неадекватности реальности, что рано или поздно приводит к серьезным психопатологиям (fornit.ru/221).
Дети в раннем возрасте оказываются ментальными клонами своих родителей, но в период игровой инициативы быстро становятся самостоятельными исследователями мира.
В случае авторитарного давления, обычно религиозного характера, период инициативы подавляется, и человек во многом остается ментальным клоном, лишенным самостоятельности или, как говориться, "рабом божьим".
Вера фиксирует нравственность на уровне древних устоявшихся догм и препятствует развитию культуры. Во время засилья организованной религии в средние века происходил застой культуры во всех ее проявлениях. Лишь с ограничением влияния религии стало возможно развитие и социальная адаптивность, но лишь за счет тех людей, которые оказались не подвержены закрепощающему влиянию церковных догм.
Человек, теснейшим образом связанный с обществом, фактически большую свою часть личности формирует из элементов окружающей культуры, и лишь очень небольшая часть оказывается самобытным творением. Еще меньшая часть этой части оказывает влияние на общество и привносит новый элемент культуры.
И под конец вот квест, с помощью которого можно убедиться, насколько адекватно удалось понять сказанное: fornit.ru/tk4 (обязательно сначала прочтите топик помощи).