Сохранение воспоминаний
Сохранение воспоминаний
http://www.sciam.ru/2005/5/neyrobiology.shtml
Почему одни события в нашей жизни сохраняются в памяти надолго, а другие исчезают без следа? Исследования процессов, которые формируют развивающийся мозг, позволят ответить на этот вопрос.
В триллере под названием "Помни" ("Memento") главный герой, Леонард, помнит все, что происходило с ним до того, как он получил удар по голове. Однако кого бы он ни встречал и что бы ни делал после той страшной ночи - все забывалось. Он потерял способность переводить кратковременную память в долговременную. Леонард пытался найти убийцу своей жены, но происходящие события мгновенно исчезали из его памяти, поэтому он вынужден был покрывать свое тело татуировками, чтобы сохранить хоть какие-то напоминания.
| КАК ПАМЯТЬ СОХРАНЯЕТСЯ В
УСТОЙЧИВЫХ СВЯЗЯХ
|
В основе сюжета фильма лежит реальная история пациента, описанная в медицинской литературе под псевдонимом "HM". В 9 лет HM упал с велосипеда и получил серьезную травму головы, из-за которой у него возникла тяжелейшая эпилепсия. Чтобы облегчить приступы, с которыми не удавалось справиться никакими другими средствами, хирурги удалили у HM часть гиппокампа и прилежащих тканей. Операция показалась успешной, поскольку приступы стали беспокоить меньше, однако она нарушила ту таинственную связь, которая тянется от кратковременной памяти к долговременной. Информация о людях, месте, событиях (декларативная память) должна пройти через гиппокамп перед тем, как она будет записана в коре больших полушарий. В результате давние воспоминания, уже зафиксированные в мозге HM, остались четкими, но все впечатления, связанные с настоящим, быстро исчезали из его памяти. HM ежемесячно приходил на прием к своему лечащему врачу, но каждый раз вел себя так, как будто они первый раз встретились.
Превращение текущих впечатлений в устойчивую память давно интриговало нейробиологов. Когда вас впервые представляют незнакомому человеку, его имя хранится в кратковременной памяти и через несколько минут может забыться. Однако часть информации, например, кличка друга, переходит в долговременную память и может сохраняться там в течение всей вашей жизни. Механизм, который заставляет мозг хранить одни впечатления и позволяет другим исчезнуть, в последнее время стал более понятным для нас.
Долговременная и кратковременная память сохраняются в связях между нейронами, в местах контакта между ними (синапсы), где отросток нейрона, передающий сигнал (аксон), встречается с одним из десятков выростов соседнего нейрона, принимающих сигнал и называемых дендритами (см. врез на стр. 63). Когда возникает кратковременная память, стимуляции синапса оказывается достаточно для того, чтобы временно сенситизировать его, т.е. повысить эффективность прохождения последующих сигналов. При долговременной памяти повышение эффективности синапса становится постоянным. Однако уже в 60?х гг. стало известно: чтобы запустить синтез белков, требуется активация генов в ядре нейрона.
Исследователи памяти ломали головы над тем, каким образом активность генов в ядре клетки может управлять событиями в удаленных синапсах. Откуда ген "знает", когда нужно усилить синаптическую связь, а когда позволить мимолетному мгновению исчезнуть бесследно? И каким образом белки, закодированные в генах, "знают", на какой именно из тысяч синапсов надо воздействовать? Те же самые вопросы возникают при изучении развития мозга у зародыша, когда мозг решает, какие синаптические связи сохранить, а какие ликвидировать. Изучая это явление, наша лаборатория разгадала одну из интригующих загадок мозга. И, подобно сказочной героине Элли, обладавшей волшебными башмачками с самого начала своих приключений и не знавшей, что именно они ей нужны для возвращения домой, мы поняли, что ответ был все время у нас перед глазами.
| КАК ПРОИСХОДИТ ЗАПОМИНАНИЕ |
Запоминание происходит тогда, когда у нервных клеток повышается эффективность связей, называемых синапсами. В случае кратковременной памяти эффект длится всего минуты или часы. При долговременной памяти синаптическая связь усиливается надолго.
Память формируется как следствие прохождения сигналов через синапсы. Сообщения начинают передаваться от одного нейрона (пресинаптической клетки) к другому тогда, когда электрический импульс, известный как потенциал действия (см. ниже), достигнет кончика отростка первого нейрона, называемого аксоном.
ПЕРЕДАЧА СИГНАЛА В СИНАПСЕ
Импульс,
пришедший в окончание аксона (вставка внизу), заставляет синаптические
пузырьки, хранящиеся в пресинаптическом нейроне, высвобождать химические
вещества, называемые нейромедиаторами, в синаптическую щель - узкий зазор
между аксоном и дендритом второго, постсинаптического нейрона.
Нейромедиаторы связываются с рецепторами на дендрите, запуская локальную
деполяризацию мембраны постсинаптического нейрона.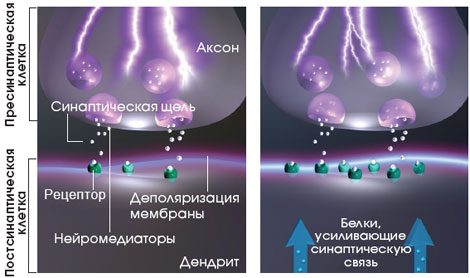
УСИЛЕНИЕ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
Если
синапс работает недолго, но с высокой частотой, то он становится более
эффективным, и в ответ на последующие стимулы в нем будут возникать более
сильные отклонения потенциала. Такое временное усиление синаптической
связи лежит в основе кратковременной памяти. Хотя еще далеко не все
понятно, ученые уже знают, что для долговременного усиления синаптической
связи постсинаптическая клетка должна выработать специальные белки,
усиливающие синаптическую связь (слева). Эти белки могут добавлять новые
рецепторы или как-то иначе изменять постсинаптическую часть синапса, а
также, возможно, влиять на пресинаптическую клетку.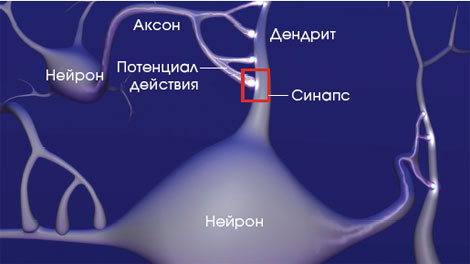
Генетическая память
Молекулярные биологи знали, что гены принимают участие в превращении памяти из кратковременной в долговременную. Эксперименты с животными показали, что их обучение требует синтеза новых белков в мозге в течение нескольких первых минут тренинга, в противном случае информация в памяти будет утеряна. Чтобы произвести новый белок, необходимо участок ДНК, находящийся в клеточном ядре, скопировать на относительно небольшую подвижную молекулу, называемую матричной РНК (мРНК), которая затем выходит в цитоплазму клетки, где специальные клеточные органеллы считывают закодированные в ней инструкции и синтезируют молекулы белка. Исследователи обнаружили, что если заблокировать процесс транскрипции ДНК в мРНК или трансляции мРНК в белок, то образование долговременной памяти нарушится, в то время как кратковременная не пострадает.
Один нейрон способен образовывать десятки тысяч синаптических связей, и поэтому трудно себе представить, чтобы для каждого из синапсов существовал свой собственный ген. Нейробиологи стремились найти объяснение тому, каким образом клеточное ядро управляет эффективностью работы каждого синапса в отдельности. Они предположили, что в синапсе, получившем достаточную стимуляцию, должны вырабатываться молекулы какого-то неизвестного сигнального вещества. После того как эффективность синапса повысилась, он, видимо, может поддерживать память еще в течение некоторого времени, пока сигнальная молекула находится на пути к ядру нервной клетки. Там она могла бы активировать определенные гены, необходимые для синтеза белков, которые усилят синаптическую связь на длительное время. Однако оставалась неразрешенной вторая проблема: каким образом белок, синтезированный в теле нейрона, сможет отыскать среди тысяч синапсов именно тот, который его затребовал.
К середине 90-х гг. исследователи памяти уже знали, что транскрипционный фактор под названием CREB играет ключевую роль в превращении кратковременной памяти в долговременную у столь далеких друг от друга видов, как мухи и мыши (см. врез на стр. 64). Транскрипционные факторы - это управляющие белки, содержащиеся в клеточном ядре, которые отыскивают конкретные последовательности ДНК и связываются с ними. Фактически они являются выключателями, управляющими транскрипцией генов. Поэтому активация CREB в нейроне ведет к активации генов, что приводит к производству таинственных белков, усиливающих синаптическую связь, и превращают кратковременную память в долговременную.
В 1997 г. Юв Фрей (Uwe Frey) из немецкого федеративного Института нейробиологии, генетической регуляции и пластичности и Ричард Моррис (Richard G. M. Morris) из Эдинбургского университета в своих экспериментах показали, что, чем бы ни были "белки памяти", от них не требуется адресации к определенным синапсам. Они могут распространиться по всей клетке, но окажут влияние только на те синапсы, которые претерпели временное повышение своей эффективности, и повысят силу этих связей на длительное время.
Однако ответ на вопрос, что представляет собой сигнальная молекула, путешествующая из синапса в ядро и определяющая, когда следует активировать CREB и сохранить след памяти, так и не был получен. Приблизительно в это же время мы с моими коллегами столкнулись с теми же проблемами, над которыми бились исследователи памяти, но рассмотрели их под другим углом зрения. В лаборатории Национального института детского здоровья и развития человека мы изучаем, каким образом формируются связи в мозге во время внутриутробного развития. Нас интересовало, как гены могут кодировать все те миллионы соединений, которые возникают в развивающемся мозге.
Мы, как и другие нейробиологи, изучающие развитие мозга, уже тогда подозревали, что личный опыт может играть определенную роль при отлаживании схемы связей мозга. Развивающийся мозг может вначале иметь лишь грубую приблизительную схему связей, запрограммированную генами. Затем молодой мозг сохраняет самые эффективные из них и уничтожает непригодные к использованию. Но как он определяет, какие связи следует сохранить?
Строительство мозга
Еще в 1949 г. психолог Дональд Хебб (Donald Hebb) предложил простое правило, описывающее, каким образом пережитый опыт мог бы усиливать отдельные нервные связи. Вдохновленный знаменитыми экспериментами И.П. Павлова на собаках, Хебб предположил, что связи между нейронами, которые разряжаются одновременно, должны усиливаться. Например, когда один нейрон, разряжающийся при звуке колокольчика, расположен поблизости от другого, который реагирует на одновременное предъявление пищи, то они приобретают более тесную связь между собой. В результате формируется клеточная система, запоминающая взаимосвязь двух событий.
Не каждый синаптический вход на нервную клетку обладает эффективностью, чтобы заставить ее разряжаться. Нейрон подобен электронному микропроцессору, поскольку он получает на свои дендриты тысячи сигналов и постоянно интегрирует всю входящую информацию. Однако в отличие от микропроцессора, обладающего множеством выходных контактов, нейрон имеет всего один выход, свой аксон. В результате нейрон может реагировать на входящие сигналы только одним способом: он может либо решить послать сигнал следующему нейрону в цепи, разрядившись импульсом и направив его по своему аксону, либо ничего не сделать.
Когда нейрон получает сигнал, потенциал мембраны его дендрита слегка отклоняется в сторону положительного значения. Когда через синапс проходят высокочастотные залпы импульсов, происходит временное повышение его эффективности, проявляющееся как образование кратковременной памяти. Недолгой работы одиночного синапса обычно бывает недостаточно для того, чтобы заставить нейрон разрядиться импульсом, который правильнее называть потенциалом действия. Однако когда множество синапсов, приходящихся на один нейрон, срабатывают одновременно, их совместные усилия настолько резко изменяют потенциал нейрона, что вынуждают его разрядиться потенциалом действия и передать сигнал следующему в цепи.
Хебб предположил также, что, подобно музыканту оркестра, не попадающему в ритм, синапс, работающий не синхронно с другими входами нейрона, должен быть исключен, в то время как синапсы, разряжающиеся одновременно (если им удается совместно заставить нейрон разрядиться потенциалом действия), должны быть усилены. Таким образом мозг получает возможность устанавливать соединения в соответствии с распространением импульсов в развивающихся нервных цепях, отлаживая и совершенствуя исходную схему связей.
Поскольку информация в нервной системе кодируется паттерном импульсов нейронной активности в мозге, я предположил, что определенные гены в нервных клетках должны включаться и выключаться в зависимости от характера импульсного разряда. Для того чтобы проверить эту гипотезу, мы с Коиши Ито (Kouichi Itoh) начали брать нейроны из зародышей мыши и выращивать их в культуре ткани. Подводя электроды прямо к чашке Петри, мы могли стимулировать нейроны, заставляя их разряжаться потенциалами действия с различными паттернами следования импульсов, а после этого измеряли количество мРНК известных генов, участвующих в процессах формирования нервных цепей или в адаптации к внешней среде. Было обнаружено, что наше предсказание оказалось правильным. Мы могли включать или выключать конкретные гены, всего лишь устанавливая соответствующую им частоту стимулов на нашем электрофизиологическом стимуляторе.
| КАК ГЕНЫ ЗАСТАВЛЯЮТ ПАМЯТЬ СОХРАНЯТЬСЯ |
)
Временное кодирование
Когда мы обнаружили, что гены нейронов могут регулироваться в соответствии с паттерном импульсов, генерируемых клеткой, мы решили выяснить, каким образом характер изменений электрического потенциала, происходящих на поверхности клетки, может управлять генами, расположенными в ядре нейрона. Для этого нам необходимо было исследовать цитоплазму клетки и узнать, какие преобразования претерпевает та информация, которая распространяется от поверхности к ядру.
Как и в хитросплетении дорог, здесь также существует множество пересекающихся биохимических путей, передающих сигнал через многочисленные перекрестки от клеточной мембраны вглубь клетки. Каким-то образом электрические сигналы, идущие с различной частотой по клеточной мембране, находят свою дорогу через цитоплазму к конечной станции - ядру.
Информация об электрическом состоянии нейрональной мембраны подается в эту систему химических реакций, происходящих в цитоплазме, через управление входом ионов кальция сквозь потенциал-чувствительные каналы клеточной мембраны. Можно сказать, что нейроны окружены морем ионов кальция, однако внутри нейронов их концентрация поддерживается на чрезвычайно низком уровне - в 20 тыс. раз ниже, чем снаружи. Когда потенциал на клеточной мембране достигает критического уровня, клетка разряжается потенциалом действия, заставляя кальциевые каналы открываться на короткое время. Потоки ионов кальция, втекающие в нейрон с каждым нервным импульсом, переводят электрический код в химический, который понятен биохимической машине, работающей внутри нейрона.
Подобно тому, как падающие костяшки домино толкают друг друга, входящие в цитоплазму ионы кальция активируют ферменты, называемые протеинкиназами. Последние запускают другие ферменты путем химической реакции, называемой фосфорилированием, при которой к белкам присоединяется фосфатная метка. Подобно бегунам, передающим эстафетную палочку, ферменты, снабженные такой меткой, выходят из состояния покоя и стимулируют активность транскрипционных факторов. CREB, например, активируется кальций-зависимыми ферментами, которые его фосфорилируют, и инактивируется ферментами, снимающими с него фосфатную метку. Однако в клетке содержатся сотни различных транскрипционных факторов и протеинкиназ. Мы хотели узнать, как конкретная частота следования потенциалов действия может управлять потоком кальция таким образом, чтобы воздействовать на нужные протеинкиназы и далее на нужные транскрипционные факторы, управляя, в конечном счете, правильным геном.
Наполняя нейроны красителем, который при повышении концентрации кальция в цитоплазме начинает флуоресцировать зеленым цветом, мы смогли проследить, каким образом различные паттерны разряда потенциала действия переводились на язык динамически меняющейся концентрации кальция. Одна возможность заключалась в том, что транскрипция генов может регулироваться степенью прироста концентрации кальция в нейроне и что различные гены по-разному реагируют на различные уровни кальция. Однако результат превзошел наши ожидания: степень повышения концентрации кальция в нейроне имела гораздо меньшее значение для регуляции работы конкретных генов, чем временные паттерны кальциевых вспышек, отражающих временной код породивших их нервных импульсов.
Фелик Эшет (Feleke Eshete) проследил кальциевые сигналы вплоть до ферментов, активировавшихся ими, и до транскрипционных факторов, регулировавшихся этими ферментами. В результате стало ясно, каким образом различные паттерны нервных импульсов могут быть переданы по различным путям внутриклеточной сигнализации. Важнейшим фактором оказалось время.
Мы обнаружили, что нельзя представить путь от клеточной мембраны до ДНК в виде одной простой последовательности химических реакций. На каждом этапе, начиная со входа кальция через мембрану, реакции разветвлялись по сети различных, хотя и переплетавшихся путей, каждый из которых имел свои собственные временные параметры, определяющие, насколько хорошо данный путь будет реагировать на прерывистые сигналы той или иной частоты. От этого и зависело, какой именно сигнальный путь достигнет ядра при любой конкретной частоте следования потенциалов действия.
Одни сигнальные пути отвечали быстро и тут же возвращались в исходное состояние. Таким образом, они реагировали на высокочастотные залпы потенциалов действия, но не могли поддерживать постоянную активацию, если вспышки импульсов прерывались длительными паузами. Другие пути медленно передавали сигналы и не могли ответить на быстрые залпы импульсов. Тем не менее, будучи активированными, они очень медленно выключались, что означает, что они способны сохранять активность между залпами импульсов, разделенными длительными промежутками покоя. Активация генов по такому пути будет ответом на нечасто, но регулярно поступающие стимулы.
Другими словами, мы наблюдали, что сигналы с разными временными паттернами распространялись по разным путям, которые были настроены именно на них, и, в конечном счете, управляли различными транскрипционными факторами и генами. Наши измерения показали, что CREB быстро активировался потенциалами действия, однако медленно инактивировался после того, как стимуляция нейрона прекращалась. Очевидно, что CREB способен сохранять активацию между повторяющимися залпами импульсов, разделенными интервалами в 30 и более минут, такими же промежутками бывают разделены повторы, необходимые для обучения новым навыкам или запоминания новых фактов.
Но могут ли сигнальные механизмы, изученные нами ради познания развития мозга, иметь отношение и к механизмам памяти?
Память в чашке Петри
Если ту часть мозга, которая была удалена у пациента HM (гиппокамп), изъять из мозга крысы и поддерживать ее жизнедеятельность в растворе солей, то с помощью микроэлектродов и электронных усилителей можно зарегистрировать электрические импульсы от отдельных синаптических связей нейрона. Подавая залп электрических импульсов на синапс и заставляя его разряжаться согласно заданному паттерну, можно усилить данную синаптическую связь. Проще говоря, в ответ на последующие стимулы этот синапс будет давать удвоенное отклонение потенциала по сравнению с тем, которое возникало до высокочастотной стимуляции.
Увеличение эффективности синапса, называемое длительной потенциацией (ДП), вопреки своему названию сохраняется не так уж и долго. Если после высокочастотной стимуляции изредка подавать тестирующие импульсы, то потенциал, возникающий в синапсе, будет постепенно уменьшаться и спустя несколько часов вернется к своему исходному значению. Это временное повышение эффективности синапса, известное под названием ранняя ДП, может служить клеточной моделью кратковременной памяти.
Удивительно, но если ту же самую высокочастотную стимуляцию подавать повторно, то возникает устойчивое повышение эффективности синапса, и это состояние называют поздней ДП. Но стимулы нельзя подавать сразу друг за другом, т.к. залпы должны быть разделены достаточно длительными периодами покоя. А добавление в солевой раствор, омывающий срез, химических веществ, блокирующих синтез мРНК или белка, приводит к падению эффективности синапса до его исходного значение в течение двух-трех часов. На клеточной модели обнаруживается, что кратковременная память не связана с ядром, в то время как долговременная зависит от него.
С помощью данной методики Фрей и Моррис показали, что белки, повышающие эффективность синапса, влияют на любой временно усиленный синапс. Сначала они кратковременно стимулировали один синапс, чтобы вызвать в нем раннюю ДП. Затем они заставляли работать второй синапс на том же самом нейроне в том режиме, чтобы вызывать позднюю ДП (три залпа с интервалом в 10 минут). В результате эффективность обоих синапсов возросла на длительное время. Более сильный стимул посылал в ядро сигнал, требующий производства белков памяти, и затем эти белки "находили" любой синапс, подготовленный к их воздействию.
Мы предположили, что когда синапс разряжается достаточно сильно или синхронно с другими синапсами, заставляя нейрон посылать потенциалы действия по своему аксону, кальций должен входить в нейрон прямо через потенциал-чувствительные кальциевые каналы на теле нейрона и активировать те пути, которые мы уже изучили, приводя в конечном счете к активации CREB в ядре.
Чтобы проверить эту гипотезу, Серена Дьюдек (Serena Dudek) и я применили химическое вещество, которое блокирует работу синапсов в срезе мозга. Затем мы непосредственно стимулировали клеточные тела нейронов и аксоны с помощью введенного электрода. В результате нейроны разряжались потенциалами действия, но синаптические входы на эти нейроны функционировать не могли. Если молекула, несущая сигнал из синапса в ядро, действительно необходима для индукции поздней ДП, нашей модели формирования долговременной памяти, то такая процедура не должна подействовать, поскольку синапсы были заглушены химическим веществом. С другой стороны, если сигналы, поступающие в ядро, возникают как следствие потенциалов действия, что было показано в наших исследованиях развития мозга, то блокирование синапсов не предотвратит активацию в ядре генов, кодирующих белки памяти.
После стимуляции мы обработали мозговые ткани таким образом, чтобы определить, был ли активирован транскрипционный фактор CREB. Оказалось, что в небольшом участке среза мозга, в котором под воздействием стимуляции возникали потенциалы действия при полном отсутствии синаптической активности, ко всем молекулам CREB были прикреплены фосфатные группы, т.е. CREB был переведен в активное состояние.
Далее мы проверили активность гена zif268, связанного с возникновением ДП и памяти. Было обнаружено, что он также оказался активированным разрядом гиппокампальных нейронов при отключенных синапсах. Однако когда мы провели такую же стимуляцию в присутствии другого вещества, которое блокирует потенциал-чувствительные кальциевые каналы - которые, как мы подозревали, и были истинным источником сигнала от мембраны в ядро, - то обнаружили, что в ответ на разряд нейронов в них уже не происходило ни фосфорилирования CREB, ни активации zif268 и другого белка, связанного с поздней ДП, называемого MAPK.
Полученные нами результаты ясно показали, что нет никакой необходимости в посреднике между синапсом и ядром. Как и в наших исследованиях развития мозга, деполяризация мембраны потенциалами действия открывала кальциевые каналы, расположенные в нейрональной мембране, запускала сигнальные пути, ведущие в ядро и включающие определенные гены. Такой принцип работы памяти представляется мне очень разумным. Вместо того чтобы вынуждать каждый синапс посылать свое собственное сообщение в ядро, транскрибирующие механизмы ядра просто прослушивают выходной сигнал нейрона и на его основе решают, синтезировать ли белки памяти или нет.
Молекулярное "помни"
Возможно, существуют пока неизвестные сигнальные молекулы, распространяющиеся из синапса в ядро и действительно участвующие в процессах памяти, однако эксперименты показали, что в них нет необходимости. Как предсказывают правила обучения Хебба, разряд нейрона, происходящий вследствие возбуждения синаптических входов клетки, является тем фактором, который необходим для упрочения памяти.
Подобно Леонарду в фильме "Помни", мы не всегда заранее знаем, какие события следует сохранить в памяти надолго. Оперативная память, необходимая для действий в настоящем, обеспечивается кратковременными изменениями силы отдельных синапсов. Но если событие достаточно важное или повторяется многократно, то синапсы заставляют нейрон, в свою очередь, выдавать нервные импульсы интенсивно и многократно, заявляя тем самым: "это событие следует запомнить". Включаются соответствующие гены, белки памяти отыскивают те синапсы, в которых удерживается кратковременная память, и, можно сказать, помечают их клеймом.
ОБ АВТОРЕ:
Дуглас Филдз (R. Douglas Fields) - заведующий
отделом развития и пластичности нервной системы Национального института
детского здоровья и развития человека, а также адъюнкт-профессор программы
по нейронаукам и когнитивным наукам Мэрилендского университета. Его
предыдущая статья "Другая часть мозга" ("В мире науки", №7, 2004 г.)
рассказывала о том, какую роль играют глиальные клетки в мышлении и
обучении.