|

Специализированный комплекс отделов мозга, управляющий социальным поведением, сначала был обнаружен у млекопитающих, затем у других наземных позвоночных и даже у рыб. Американские биологи показали, что у разных групп позвоночных сходна не только структура этого комплекса, но и характер работы ключевых генов в нем. Те различия, которые всё же имеются, в большей степени затрагивают синтез сигнальных веществ (нейромедиаторов) и в меньшей — распределение рецепторов, реагирующих на эти вещества. По-видимому, уже у последнего общего предка лучеперых рыб и наземных позвоночных имелась социально-ориентированная нейронная сеть, основные структурные и нейрохимические свойства которой в ходе дальнейшей эволюции менялись весьма медленно.
Исследования последних лет показали, что у костистых рыб, амфибий, рептилий, птиц и млекопитающих социальное поведение (включая агрессию, брачное поведение и заботу о потомстве) контролируется одним и тем же комплексом отделов мозга. Этот комплекс называют «нейронной сетью социального поведения» (social behaviour network). В работе этой сети важную роль играют стероидные половые гормоны (андрогены, эстрогены) и нейропептиды (в том числе окситоцин, вазопрессин и их гомологи, см.: Гены управляют поведением, а поведение — генами, «Элементы», 12.11.2008). Сеть социального поведения тесно связана с системой внутреннего вознаграждения, задача которой — отличать, образно говоря, хорошее от плохого и сообщать о результатах другим отделам мозга при помощи нейромедиатора дофамина (см.: В «системе вознаграждения» найдены нейроны, возбуждающиеся от хороших предчувствий, «Элементы», 10.02.2012). Вместе эти два нейронных контура (социального поведения и вознаграждения) образуют «систему принятия социальных решений» (SDM, social decision-making network). Основные компоненты сети SDM показаны на рис. 1.
Самое удивительное свойство сети SDM — это ее эволюционный консерватизм, то есть крайне медленный темп эволюционных изменений. На первый взгляд, это противоречит тому громадному многообразию форм социального поведения, которое наблюдается у позвоночных. Хотя, с другой стороны, базовые социально-ориентированные задачи у всех позвоночных схожи: привлечь хороших половых партнеров, одолеть конкурентов, повысить свой социальный статус, вырастить побольше здоровых потомков... Это фундаментальное сходство жизненных устремлений, вероятно, и создает предпосылки для развития более или менее универсальных социально-ориентированных нейронных структур в ходе эволюции. Та нейронная сеть, которая сформировалась еще у доисторических рыб — общих предков современных лучеперых рыб и наземных позвоночных, — очевидно, оказалась вполне универсальной. Даже очень небольшие модификации отдельных компонентов сети SDM могут оказаться достаточными для значительного изменения социального поведения (см.: Goodson, 2005. The Vertebrate Social Behavior Network: Evolutionary Themes and Variations). Иногда, конечно, может развиться и нечто принципиально новое. Скажем, у млекопитающих значительную часть социальных функций взяла на себя префронтальная кора, аналогов которой нет у других позвоночных (именно поэтому, кстати, префронтальная кора не рассматривалась в исследовании, о котором пойдет речь).
Лорен О’Коннелл (Lauren A. O’Connell) и Ханс Хофманн (Hans A. Hofmann ) из Техасского университета в Остине решили выяснить, была ли сеть SDM так же консервативна на уровне генной экспрессии (работы генов), как и на уровне структуры. Для этого они проанализировали опубликованные результаты 152 исследований, в которых изучалась активность генов в разных участках сети SDM у разных позвоночных. Авторы сконцентрировались на десяти генных продуктах, работа которых особенно важна для сети SDM. Первые четыре из них связаны с производством лигандов, то есть сигнальных веществ, воздействующих на те или иные рецепторы, остальные шесть — сами рецепторы:
1. TH — тирозингидроксилаза, фермент, необходимый для синтеза дофамина;
2. AVP — аргинин-вазопрессин, который чаще для краткости называют просто «вазопрессином»;
3. OXY — окситоцин;
4. Aromatase — ароматаза, фермент, отвечающий за синтез эстрогенов;
5. D1aR — дофаминовый рецептор первого типа;
6. ER — эстрогеновый рецептор;
7. AR — андрогеновый рецептор;
8. PR — прогестероновый рецептор;
9. V1aR — вазопрессиновый рецептор;
10. OTR — окситоциновый рецептор.
Имеющихся данных пока недостаточно, чтобы сравнивать активность этих генов количественно, поэтому анализ проводился на качественном уровне. Для каждой группы позвоночных и для каждого отдела сети SDM авторы пытались выяснить, в каком состоянии находится каждый из 10 генов: включенном или выключенном («экспрессия есть» или «экспрессия отсутствует»).
Распределение генной экспрессии по отделам сети SDM оказалось весьма сходным у всех изученных групп. Наиболее консервативен рисунок экспрессии рецепторов. Почти у всех исследованных видов позвоночных все шесть рецепторов экспрессируются во всех или почти всех отделах сети SDM (рис. 2).
Экспрессия генов, связанных с производством лигандов, более изменчива. Самые значительные изменения произошли при отделении предков лучеперых рыб от предков лопастеперых и тетрапод (наземных позвоночных), а также при расхождении линий птиц и рептилий (рептилии были представлены змеями и ящерицами; по крокодилам и черепахам данные, к сожалению, пока отсутствуют).
|
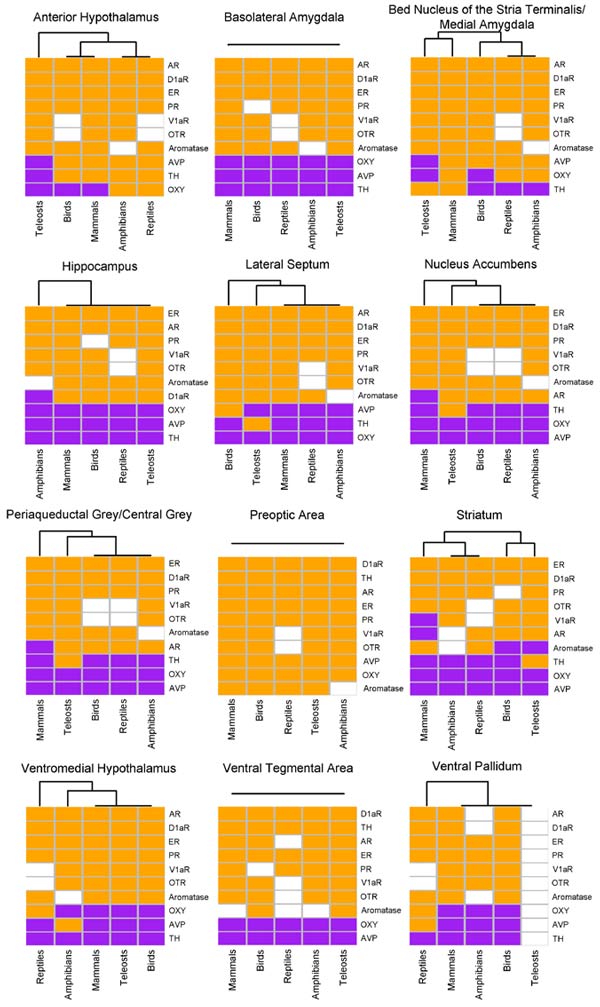
Рыбы отличаются от других позвоночных тем, что синтез TH (тирозингидроксилазы) происходит у них в большем числе отделов. В центральном сером веществе, латеральной перегородке, прилежащем ядре и стриатуме экспрессия TH зарегистрирована только у рыб. Из-за отсутствия внешней группы (то есть данных по более «базальным» позвоночным, таким как хрящевые рыбы или миноги) пока нельзя сказать, каким был рисунок экспрессии TH у общего предка рыб и тетрапод и в какой из двух линий произошло изменение.
У чешуйчатых рептилий (ящериц и змей) окситоцин производится в вентральном паллидуме и вентромедиальном гипоталамусе, где у других позвоночных ген окситоцина выключен. Отличительными особенностями млекопитающих являются отсутствие экспрессии вазопрессинового рецептора в стриатуме, андрогенового рецептора — в стриатуме, центральном сером веществе и прилежащем ядре.
Самый консервативный профиль экспрессии характерен для преоптической области (в ней все 10 генов включены у всех групп), а также для вентральной области покрышки (в ней у всех позвоночных гены окситоцина и вазопрессина выключены, остальные 8 — включены). Наибольшая эволюционная пластичность характерна для стриатума (striatum), или полосатого тела: в этом отделе мозга нейрохимический профиль менялся в ходе эволюции сильнее всего.
Главный вывод исследования состоит в том, что «сеть принятия социальных решений» у позвоночных оказалась крайне консервативной не только на структурном, но и на нейрохимическом уровне. Все эволюционные изменения социального поведения у позвоночных осуществлялись, по-видимому, за счет относительно небольших изменений соответствующих нейронных сетей. При этом распределение сигнальных веществ — лигандов — менялось сильнее, чем распределение рецепторов, реагирующих на эти лиганды. Нечто подобное наблюдалось и в эволюции индивидуального развития (онтогенеза) животных: распределение лигандов (морфогенов) по телу эмбриона менялось быстрее, чем распределение соответствующих рецепторов.
Очевидно, нейрологические основы разнообразия форм социального поведения позвоночных скрываются в более тонких деталях строения и работы сети SDM, чем те, что рассматривались в данной работе. Следующим шагом к пониманию этих основ, по идее, должно стать количественное, а не только качественное, сравнение уровней экспрессии генов в разных отделах мозга у позвоночных.
Источник: Lauren A. O’Connell, Hans A. Hofmann. Evolution of a Vertebrate Social Decision-Making Network // Science. 2012. V. 366. P. 1154–1157.
Рис. 1. Отделы мозга, ответственные за принятие социально-ориентированных решений у позвоночных (сверху вниз: рептилии, птицы, млекопитающие, амфибии, костистые рыбы). Для каждой группы показана серия схематических поперечных срезов мозга (точнее, одной половинки мозга) на разных уровнях; слева направо увеличивается расстояние от переднего конца мозга. Компоненты нейронной сети социального поведения показаны желтым цветом, системы внутреннего вознаграждения — голубым. Отделы, ответственные за связь этих двух систем, показаны зеленым цветом. AH — передний гипоталамус, blAMY — базолатеральная миндалина, BNST — ядро ложа терминальной полоски, HIP — гиппокамп, LS — латеральная перегородка, meAMY — медиальная миндалина, NAcc — прилежащее ядро, PAG — центральное серое вещество, POA — преоптическая область гипоталамуса, Str — стриатум, VMH — вентромедиальный гипоталамус, VP — вентральный паллидум (бледный шар), VTA — вентральная область покрышки среднего мозга. Изображение из обсуждаемой статьи в Science