Правило Лагерквиста. Физико-химическое обоснование симметрий и соотношений Румера.
Относится к разделу Молекулярная биология
Данная статья посвящена разбору физико-химических механизмов, которые обеспечивают закономерности распределения вырожденных и невырожденных кодонов, включая симметрические преобразования и соотношения, установленные Юрием Борисовичем Румером.
Дело в том, что симметрические преобразования Румера, и соотношения Румера могут быть однозначно выведены из так называемого правила Лагерквиста [Lagerkvist U. 1978]:
Если первые две позиции кодона формируют сильные уотсон-криковские пары SS (S=G или C), то основы кодонов сильные. Если же первые две позиции формируют слабые пары WW (W=A или U), то основы кодонов слабые. Если основа кодонов представлена нуклеотидами, формирующими и сильную, и слабую пару, то сила основ определяется пурин-пиримидиновым типом второго основания в кодоне - если второе основание представлено пиримидином, то основа сильная, если же пурином, то слабая.
Из данного правила автоматически вытекают симметрические преобразования Румера и соотношения Румера. Поэтому найдя объяснение правилу Лагерквиста, мы автоматически сможем обосновать и факт существования симметрий кода, обнаруженных Юрием Борисовичем Румером.
В предыдущей статье "Молекулярные механизмы обеспечения вырожденности кода" было показано, что вырожденность ГК в большинстве известных организмов обеспечивается взаимодействием модифицированных или не модифицированных первых нуклеотидов антикодона с третьим нуклеотидом кодона на мРНК. Однако в простейших прокариотических системах (например, митохондриях) третий нуклеотидом кодона может вообще специфически не взаимодействовать с первым нуклеотидом антикодона, тем не менее генетическое кодирование осуществляется с приемлемой (хотя и меньшей) точностью (это значит, что возможно ошибочное включение тРНК с другой аминокислотой), причём в случае несколько видоизменённых митохондриальных и других прокариотических геномов правило Лагерквиста, а, значит и симметричные соотношения Румера и правила Румера продолжают выполняться. Дело в том, что наряду с механизмом "качения"("wobbling"), обеспечивающим высокую точность декодирования, существует во всех организмах ещё и другой механизм, функционирования которого достаточно для обеспечения декодирования значительно более простых геномов. Этот факт даёт основание предположить, что описанный ниже механизм является более древним и по-видимому существовал непосредственно в период формирования ГК, а затем был дополнен более совершенным механизмом "качания" третьего основания кодона, обеспечившим большую точность процесса трансляции (синтеза белка). При взаимодействии кодона и антикодона образуется двойная спираль, которая узнаётся декодирующим центром рибосомы, представленным рибосомной РНК малой субъединицы. Декодирующий центр может распознавать только двойную спираль из кодон-антикодоновой пары. Но стабильность этой двойной спирали, оказывается, сильно зависит от того, какие именно основания принимают участие в кодон-антикодоновом взаимодействии.
Причина зависимости стабильности кодон-антикодоновой пары от состава нуклеотидов заключается в том, что комплементарная пара G-C образует три водородных связи и является наиболее энергетически выгодной по сравнению с парой A-U, в которой образуются только две водородных связи (для тех, кто забыл, схемы образования водородных связей в парах G-C и A-U приведены на рис. 2. в статье "Молекулярные механизмы обеспечения вырожденности кода"). Поэтому если первые два основания в кодон-антикодоновом взаимодействии представлены только парами G-C (6 водородных связей), то энергии этих связей ДОСТАТОЧНО для образования стабильной кодон-антикодоновой пары, которая будет распознана декодирующим центром малой субъединицы рибосомы.
Если же первые два основания в кодоне представлены только парами A-U (4 водородных связи), то стабильности этих пар в отсутствие стабильного спаривани третьего нуклеотида кодона с первым антикодона уже НЕДОСТАТОЧНО для формирования двойной спирали, распознаваемой декодирующим центром рибосомы. В этом случае двойная спираль может быть стабилизирована только взаимодействием третьего нуклеотида кодона с первым антикодона.
Промежуточная ситуация наблюдается для случая, если один из двух первых нуклеотидов кодон-аниткодоновой представлен парой A-U, а другая G-C. В этом случае стабильность комплекса "кодон-антикодон" будет занимать промежуточное положение и существенно будет зависеть от того, представлено ли второе основание пурином или пиримидином. Разберёмся, почему стабильность двойной спирали в отсутствие стабильного спаривани третьего нуклеотида кодона с первым антикодона существенно зависит от того, представлено ли второе основание кодона пурином или пиримидином.
Дело в том, что если второй нуклеотид в антикодоне пурин (а в кодоне значит пиримидин), то при взаимодействии кодона с антикодоном этот пурин получает возможность дополнительно взаимодействать с уридином тРНК, расположенного в позиции 33 (о нумерации позиций см. "Транспортные РНК как молекулярные реликты"). Схема этого взаимодействия приведена на рисунках 1 и 2.
Если центральный нуклеотид антикодона является пурином, то при формировании кодон-антикодоновой двойной спирали (U33)02'H (эта запись означает "атом водорода в положении 2' (см. рис.2 слева) уридина тРНК в положении 33") взаимодействует с (A35)N7 (эта запись означает "седьмым атомом азота (см. рис. 2 слева) аденина в положении 35 тРНК). Если же центральный нуклеотид в антикодоне является гуанин, то он также взаимодействует с (U33)02'H, но не только с помощью (G35)N7, но и также (G35)O6, образуя в водородной связи разветвление (бифуркацию), как это хорошо видно на рис.2 слева. В обоих случаях (взаимодействия аденина и гуанина) образуемая водородная связь сравнима по прочности с обычной уотсон-криковской водородной связью и придаёт кодон-антикодоновому комплексу дополнительную стабильность. Именно поэтому если второй нуклеотид в антикодоне пурин, а, значит в кодоне - пиримидин, то кодон-антикодоновая пара только с одной GC-связью может стабилизировать двойную цепь даже в отсутствие взаимодействия третьего нуклеотида в случае наиболее примитивных прокариотических геномов.
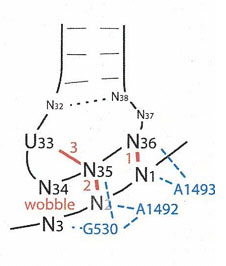
Рис. 1. Взаимодействие кодона мРНК с антикодоном тРНК. Красным цветом показано взаимодействие нуклеотидов кодона, антикодона и тРНК. Голубым цветом показано взаимодействие с нуклеотидами РНК декодирующего центра малой субъединицы рибосомы (номера нуклеотидов соответствуют стандартным общепринятым). Рисунок взят из [Lehmann J., Libchaber A. 2008]
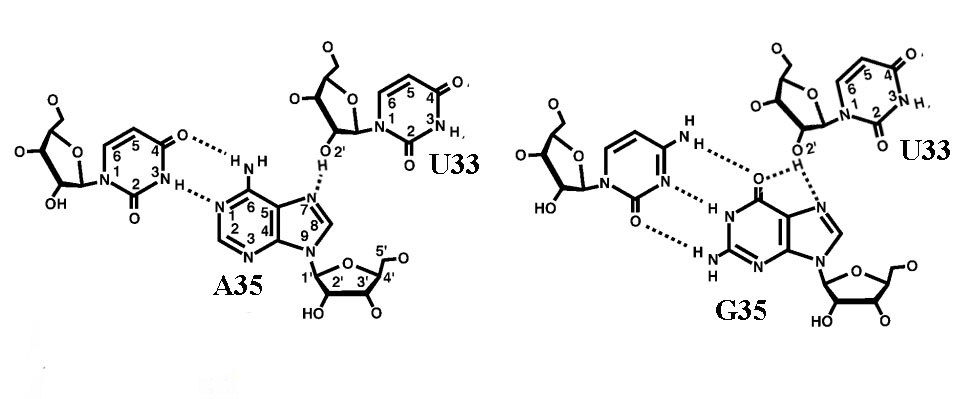
Рис. 2. Взаимодействие второго аденина (слева) и второго гуанина (справа) антикодона с U33 тРНК с образованием стабильной водородной связи.
Если же второй нуклеотид атникодона является пиримидином, то дополнительной стабильной водородной связи с U33 не образуется.
Декодирующий центр малой субъединицы рибосомы может узнавать только двойную спираль, образуемую кодон-антикодоновой парой. Три нуклеотидных остатка рибосомы (G530, A1492 и A1493) взаимодействую с малой бороздокой двойной спирали, образованной кодон-антикодоновой парой. A1493 взаимодействует с малой бороздкой первой пары оснований, тогда как G530 и A1492 взаимодействует с малой бороздкой от второй пары оснований (на рисунке 1 эти взаимодействия показаны голубым цветом).
Таким образом, характерное для большинства современных организмов специфическое взаимодействие первого нуклеотида антикодона с третьим нуклеотидом кодона в некоторых наиболее примитивных прокариотических геномах, ряде экспериментальных бесклеточных системах синтеза белка (трансляции in vitro) и предположительно в древних системах синтеза белка не обязательно происходит для сильных основ кодонов, в которых взаимодействия первых двух нуклеотидов кодона было достаточным для образования двойной кодон-антикодоновой спирали, распознаваемой декодирующим центром рибосомы.
Это правило распространялось на любые альтернативные варианты ГК, поэтому правило Лагерквиста и румеровские симметрии и соотношения оказались справедливыми как для канонического ГК, так и для его различных диалектов.
Литература.
1. Lagerkvist U. "Two out of three": An alternative method for codon reading. // PNAS USA. 1978. V.75. P.1759-1762.
2. Lehmann J., Libchaber A. Degeneracy of the genetic code and stability of the base pair at the second position of the anticodon. // RNA 2008. V.14. P.1264-1269.
Статьи, связанные с темой:
Молекулярные механизмы обеспечения вырожденности кода
Транспортные РНК как молекулярные реликты
Список публикаций >>
Тест: А не зомбируют ли меня? Тест: Определение веса ненаучности
Создан синаптический коммутатор с автономной памятью и низким потреблением
Ученые Северо-Западного университета, Бостонского колледжа и Массачусетского технологического института создали новый синаптический транзистор, который имитирует работу синапсов в человеческом мозге.
Тематическая статья: Как образумить Beast?
Рецензия: Статья П.К.Анохина ФИЛОСОФСКИЙ СМЫСЛ ПРОБЛЕМЫ ЕСТЕСТВЕННОГО И ИСКУССТВЕННОГО ИНТЕЛЛЕКТА